

Abstract
Background
An important functional trait of organisms is their trophic mode. It determines their position within food webs, as well as their function within an ecosystem. For the better part of the 20th century, aquatic protist communities were thought to consist mainly of producers (phytoplankton) and consumers (protozooplankton). Phytoplankton cover their energy requirements through photosynthesis (phototrophy), while protozooplankton graze on prey and organic particles (phagotrophy). However, over the past decades, it was shown that another trophic group (mixoplankton) comprise a notable part of aquatic protist communities. Mixoplankton employ a third trophic mode by combining phototrophy and phagotrophy (mixotrophy). Due to the historical dichotomy, it is not straightforward to gain adequate and correct information on the trophic mode of aquatic protists. Long hours of literature research or expert knowledge are needed to correctly assign trophic modes. Additionally, aquatic protists also have a long history of undergoing taxonomic changes which make it difficult to compare past and present literature. While WoRMS, the World Register of Marine Species, keeps track of the taxonomic changes and assigns each species a unique AphiaID that can be linked to its various historic and present taxonomic hierarchy, there is currently no machine-readable database to query aquatic protists for their trophic modes.
New information
This paper describes a dataset that was submitted to WoRMS and links aquatic protist taxa, with a focus on marine taxa, to their AphiaID and their trophic mode. The bulk of the data used for this dataset stems from (routine) monitoring stations in the North Sea and the Baltic Sea. The data were augmented and checked against state-of-the-art knowledge on mixoplankton taxa by consulting literature and experts. Thus, this dataset provides a first attempt to make the trophic mode of aquatic protists easily accessible in both a human- and machine-readable format.
Keywords: aquatic protists, phytoplankton, protozooplankton, mixoplankton, trophic mode, functional traits, functional biodiversity
Introduction
Protists (i.e. unicellular eukaryotes) form the base of aquatic ecosystems by providing food for higher trophic levels. Even though protist communities are so important for the trophic functioning of aquatic ecosystems, the trophic diversity within those protist communities is not always clear. For the better part of the 20th century, aquatic protist communities were divided into producers, the phytoplankton and grazers, the (proto)zooplankton (Flynn et al. 2013). Over the past decade, there has been an effort to reshape the traditional dichotomy of aquatic protist communities by taking mixotrophic protists into account (Mitra et al. 2016). Mixotrophic protists can function both as producers and grazers and, recently, the term mixoplankton has been suggested to describe this group (Flynn et al. 2019).
However, taking the correct trophic mode into account is still a challenge. Due to the historical bias, most aquatic protists are still by default categorised as either phytoplankton or protozooplankton. Intensive experimental work is required to determine mixotrophy in protists. While, in the past years, quite a few papers were published that contained lists of currently-proven marine mixoplankton (Faure et al. 2019, Leles et al. 2017, Leles et al. 2019), there is no database available which allows the trophic mode of aquatic protists to be queried. This makes it very time-consuming to take the trophic mode into account for large data-driven approaches on aquatic protist communities. A further complication is added through the frequent taxonomic changes within the protist community which make it difficult to compare literature references.
This dataset provides information on the trophic mode of aquatic protists and links them to the WoRMS database that keeps track of taxonomic name changes by using a unique species identifier, the AphiaID. By combining information on trophic modes with an already existing and widely-used database such as WoRMS, the authors hope to make data on trophic modes of aquatic protists more accessible in a machine-readable fashion. Thus, the dataset can help facilitate a better understanding of trophic dynamics and the functional role of protist groups within aquatic ecosystems. The trophic mode of the taxa included in this dataset can be accessed via the attributes of the WoRMS taxa search tool (see, for example, http://www.marinespecies.org/aphia.php?p=taxdetails&id=232772#attributes).
However, this dataset is only a start. The authors hope that, as more information on mixoplankton becomes available, this dataset will actively be expanded through community effort. New data can easily be submitted to WoRMS using the instructions available on https://www.marinespecies.org/contribute.php.
General description
Purpose
The purpose of this project was to establish a dataset on trophic modes of aquatic protists. As correct classification of trophic modes is especially important within the context of analysing routine monitoring data, the idea arose to make this work more accessible to the broad aquatic science and management community. This dataset was assembled in the scope of the H2020 Marie-Curie ITN "MixITiN".
Project description
Title
A dataset on trophic modes of aquatic protists
Personnel
Lisa K. Schneider (management, data collection, literature research, tidy data implementation, data concatenation, manuscript preparation), Konstantinos Anestis (data collection, literature research, manuscript contribution), Joost Mansour (literature research, manuscript contribution), Anna A. Anschütz (literature research, manuscript revision), Nathalie Gypens (data collection, expert knowledge, manuscript revision), Per J. Hansen (data collection, expert knowledge, manuscript revision), Uwe John (data collection, expert knowledge, manuscript revision), Kerstin Klemm (data collection, manuscript revision), Jon Lapeya Martin (data collection, manuscript revision), Nikola Medic (literature research, manuscript revision), Fabrice Not (expert knowledge, manuscript revision), Willem Stolte (management, concept development, expert knowledge, manuscript preparation).
Design description
To gather, analyse and disseminate the trophic mode of aquatic protists, a dataset was submitted to the World Register of Marine Species, WoRMS at http://www.marinespecies.org. WoRMS provides "an authoritative and comprehensive list of names of marine organisms, including information on synonymy" (WoRMS Editorial Board 2017) and this list of marine organisms can be augmented with metadata, such as traits, for example, trophic modes. Each organism is labelled with a unique AphiaID with which it is possible to keep track of taxonomic name changes (Vandepitte et al. 2015). This approach of keeping track of taxonomic name changes allows the database to be accessed and used in different ways, for example, by searching for single organisms or matching a list of taxa.
In this dataset, the trophic mode is defined by assigning one of the three different aquatic protist categories (sensu Flynn et al. (2019)): phytoplankton, protozooplankton or mixoplankton. Phytoplankton are defined as protists that obtain their nourishment via photo(auto)trophy and osmo(hetero)trophy. Irrespective of seasonality or environmental conditions, phytoplankton are always incapable of phago(hetero)trophy. Protozooplankton are defined as protists that obtain their nourishment via phago(hetero)trophy and osmo(hetero)trophy. Irrespective of seasonality or environmental conditions, protozooplankton are always incapable of photo(auto)trophy. Mixoplankton are defined as protists that obtain their nourishment by combining photo(auto)trophy, osmo(hetero)trophy and phago(hetero)trophy, so called mixotrophy.
Furthermore, for each mixoplankton, the type of mixotrophy is assigned as a trait. In the dataset, the type of mixotrophy is defined by assigning CM, GNCM, pSNCM or eSNCM to the mixoplankton, according to the types identified in Mitra et al. (2016). Constitutive Mixoplankton (CM) have the innate ability to perform photosynthesis, while Non-Constitutive Mixoplankton need to acquire chloroplasts from their prey. These Non-Constitutive Mixoplankton are divided into the General Non-Constitutive Mixoplankton (GNCM), the plastid Specialists Non-Constitutive Mixoplankton (pSNCM) and the endosymbiotic Specialist Non-Constitutive Mixoplankton (eSNCM). The GNCMs can use chloroplasts from multiple phototrophic prey, while the pSNCMs and eSNCMs only use chloroplasts from specific preys or endosymbionts.
Funding
This project has received funding from the European Union's Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement No 766327 and EMODnet Biology (EC Service contract – EASME/EMFF/2016/1.3.1.2/Lot 5/SI2.750022).
Sampling methods
Study extent
This dataset focuses on the trophic modes of aquatic protists. It combines data from five different sources: routine monitoring (HELCOM 2019, Rijkswaterstaat 2019), scientific cruises (John 2020, Martin 2020), scientific papers (primary literature), review papers and a book chapter (secondary literature). Sampling for routine monitoring and on scientific cruises was performed using Niskin bottles, followed by inspection of the samples using light microscopy. In the case of the scientific cruises, metabarcoding was employed for further validation of the microscopic data. Suppl. material 1 lists all sources and their complete references.
Sampling description
This dataset (Suppl. material 2) covers 1296 taxa. Fig. 1 shows that the bulk of the taxa stem from routine monitoring (72%) and are mainly labelled as phytoplankton (89%). Secondary literature (reviews and book chapter) contributes 10% and 1.9% of the total taxa, respectively, which are all labelled as mixoplankton. Primary literature (scientific papers) contributes 11% of the total taxa and are divided evenly between protozooplankton and mixoplankton. Recent scientific cruises in the North Sea contribute 3% of the total taxa and display the most even distribution of trophic modes.
Figure 1.

Depiction of a) percentage of the data origin to the complete dataset and b) percentage of trophic mode per data origin.
In total, 21% of the taxa are classified as mixoplankton, 66% as phytoplankton and 13% as protozooplankton (Fig. 2). However, as 72% of the taxa originate from routine monitoring, the percentage of mixoplankton is most likely under-represented. Most routine monitoring undersample mixoplankton due to the employed sampling techniques (Flynn et al. 2019). An example is the routine monitoring data of the Dutch Southern North Sea in which ciliates, as well as nanoflagellates, are often not identified and counted. Both of these groups are known to contain mixoplankton (Haraguchi et al. 2018, Stoecker and Lavrentyev 2018). Furthermore, mixotrophy must be proven in phyto- and protozooplankton by observing either feeding or utilisation of chloroplasts from prey or symbionts. It must be assumed that taxa remain labelled according to the historical dichotomy until proven otherwise. This remains an issue also for this dataset, of which the user should be aware. It can only be remedied by continually updating the dataset as new mixoplankton taxa are empirically determined.
Figure 2.

Contribution of trophic modes to total dataset.
Quality control
In order to ensure consistent taxonomy over the various data sources, each data source was matched against the WoRMS taxonomic database using the WoRMS "Match taxa" tool. This ensured that each taxon was given the currently-accepted scientific name and referenced with an AphiaID. Data sources were tidied (Wickham 2014) and joined into one dataset. If the various sources disagreed with each other on the trophic mode of the taxon, two decision pathways were possible. Firstly, mixoplankton data sources were always given precedence over other sources. Secondly, if the data sources disagreed on non-mixoplankton, then expert knowledge and literature was used to assign the trophic mode of that organism. Lastly, the list was checked by expert witnesses to ensure correct trophic classifications. This described workflow is visualised in Fig. 3.
Figure 3.
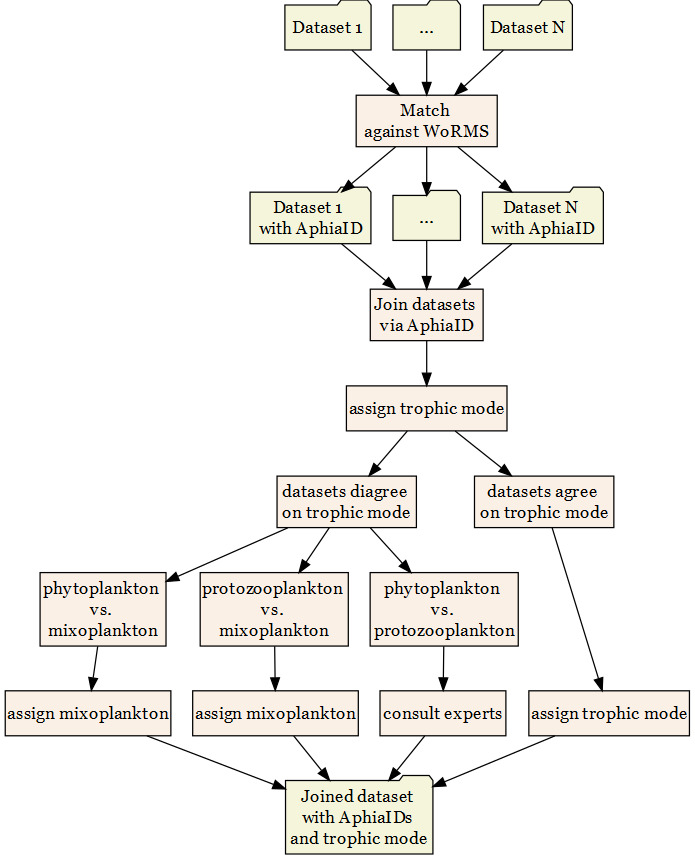
Workflow depiction beginning with single data sources and ending with the complete dataset.
Geographic coverage
Description
As this consortium is based in the EU, data stemming from routine monitoring is biased towards European waters. Data derived from literature extends beyond the EU. We hope that this dataset will be built upon with data contributions from other scientists, to establish a more encompassing collaborative resource that will promote research on trophic modes. New data can easily be submitted to WoRMS using the instructions available on https://www.marinespecies.org/contribute.php.
Taxonomic coverage
Description
Fig. 4 visualises the contribution of each class to the total dataset (Fig. 4a), as well as the contribution of trophic modes across those classes (Fig. 4b). It should be noted that only those classes are displayed that make up 90% of the total dataset. The largest class, represented in the dataset, is Bacillariophyceae, followed by Dinophyceae and Cyanophyceae. In terms of trophy, the classes which contain the most phytoplankton are Bacillariophyceae, Cyanophyceae, Chlorophyceae and Trebouxiophyceae. The classes which contain the most mixoplankton are Dinophyceae, Polycystina, Oligotrichea, Globothalamea, Prymnesiophyceae, Acantharia and Cryptophyceae. Protozooplankton are represented in the classes Dinophyceae, Polycystina, Oligotrichea, Globothalamea and Acantharia, in which they contribute between 5% and 30%.
Figure 4.

Depiction of a) percentage of each class to the complete dataset and b) percentage of trophic mode per class.
Traits coverage
This dataset focuses on the trophic mode of aquatic protists. As mentioned, aquatic protists can be divided into phytoplankton, protozooplankton and mixoplankton. This next section will give more detail on these different functional groups and their impact on aquatic ecosystems.
Phytoplankton are defined as those protists that perform photosynthesis and are incapable of phagotrophy. The most prominent examples of phytoplankton groups are cyanobacteria, diatoms and green algae. Attributing to their need of light for photosynthesis, phytoplankton are found in the euphotic zone, where light is available. It is estimated that aquatic photosynthesis by phytoplankton totals about half of the total primary production on Earth (Falkowski 1994, Field 1998). Phytoplankton in marine ecosystems play an important role in major biogeochemical cycles. For example, cyanobacteria species are known for their capacity to fix atmospheric nitrogen (25-50% of global natural fixation), a unique feature for this phytoplankton group (Canfield et al. 2010). Furthermore, diatoms contribute considerably to the global carbon cycling as they are responsible for 30-40% of global primary productivity (Sarthou et al. 2005). The diatom cell wall is composed of silica and thereby diatoms are considered the world's largest contributors to the silica cycle (Tréguer and De La Rocha 2013). Moreover, sinking of phytoplankton contributes to the carbon export to the deep oceans (Falkowski 1994).
Protozooplankton are defined as those protists that gain their nutrition through capture and ingestion of prey (or organic particles). Protozooplankton do not have the capability for photosynthesis, nor other means of producing their own organic carbon. Examples of protozooplankton are heterotrophic ciliates, heterotrophic dinoflagellates and heterotrophic (nano)flagellates. The grazing of heterotrophic protists on phytoplankton plays an important role in controlling the growth and population of phytoplankton taxa. Heterotrophic protists are the connecting link for energy transfer towards higher trophic levels and, in some cases, can be responsible for the removal of the largest part of primary production (Calbet and Landry 2004, Lawerence and Menden-Deuer 2012). Apart from the consumption of phytoplankton, heterotrophic protists also ingest prokaryotes indicating their further involvement in planktic food web energy transfer (Cho et al. 2000, Pernice et al. 2014, Šimek et al. 2019).
Mixoplankton are defined as those protists that can combine phototrophy and phagotrophy (sensu Flynn et al. 2019). Mixoplankton are often associated with mature ecosystems (Mitra et al. 2014, Moorthi et al. 2017) and many harmful algal bloom species are known to be mixotrophic. Due to their ability to combine phototrophy and phagotrophy, they can simultaneously fulfil many of the functions (Selosse et al. 2016) described for both phytoplankton and protozooplankton. Mixoplankton can thus also contribute significantly to primary productivity and functionality of ecosystems (Ghyoot et al. 2017). The additional energy acquired by the consumption of prey can increase the gross growth efficiency of mixoplankton (Schoener and McManus 2012) and subsequently, have major effects on trophic transfer in the food web (Ward and Follows 2016). CMs (e.g. Prymnesium parvum, Karlodinium veneficum) that have the innate ability to perform photosynthesis, can express bacterivory (Unrein et al. 2007) or ingest other protists to supplement their nutritional needs (Stoecker et al. 2017). NCMs, such as the kleptoplastidic ciliate Strombidium basimorphum, can be voracious grazers, achieving grazing rates comparable with pure heterotrophic species (Maselli et al. 2020). Furthermore, NCMs also contribute significantly to primary production through either their ability to use prey chloroplasts after ingestion (Nielson et al. 2012) or others (eSNCMs, like many Acantharia and Foraminafera) through chloroplast containing endosymbionts (Caron et al. 1995).
Usage rights
Use license
Creative Commons Public Domain Waiver (CC-Zero)
Data resources
Data package title
Trophic modes of aquatic protists
Number of data sets
2
Data set 1.
Data set name
List of trophic citations
Number of columns
2
Description
List of trophic mode references giving the short citation form (used in the dataset) and the full reference.
Data set 1.
| Column label | Column description |
|---|---|
| source | Short citation of the reference source used in the dataset |
| Full citation | Full APA citation of the reference source |
Data set 2.
Data set name
Trophic modes of aquatic protists
Number of columns
13
Description
Dataset listing the trophic modes of aquatic protists (with reference and data source) along with their accepted scientific name, AphiaID and taxonomic hierarchy.
Data set 2.
| Column label | Column description |
|---|---|
| ScientificName | Accepted scientific name retrieved from WoRMS. |
| AphiaID | Accepted AphiaID (unique identifier) retrieved from WoRMS. |
| Trophy | Gives the trophic mode of taxa as either "phytoplankton", "protozooplankton" or "mixoplankon". |
| typeMX | Gives the type of mixotrophy as either "CM" (Consitutive Mixoplankton), "GNCM" (General Non-Constitutive Mixoplankton), "eSNCM" (endosymbiotic Specialist Non-Constitutive Mixoplankton) or "pSNCM" (plastid Specialist Non-Constitutive Mixoplankton). If the type of mixotrophy does not apply (because the organism is labelled as phytoplankton or protozooplankton), the type of mixotrophy is labelled with "NA". |
| source | Gives the reference for the assigned trophic modes. Refers to primary literature or secondary literature (book, review papers or published datasets from routine monitoring). |
| dataType | Denotes the origin of the data point. Can be either "book", "paper", "review", "routine monitoring" or "scientific cruise". |
| Kingdom | Kingdom of the taxa within the taxonomic hierarchy |
| Phylum | Phylum of the taxa within the taxonomic hierarchy |
| Class | Class of the taxa within the taxonomic hierarchy |
| Order | Order of the taxa within the taxonomic hierarchy |
| Family | Family of the taxa within the taxonomic hierarchy |
| Genus | Genus of the taxa within the taxonomic hierarchy |
| Species | Species of the taxa within the taxonomic hierarchy |
Additional information
A github repository is available which contains the code to match the species list against WoRMS, as well as to create the figures: https://github.com/lkschn/trophyProtists.
Supplementary Material
List of trophic citations
Lisa K. Schneider, Konstantinos Anestis, Joost Mansour, Anna A. Anschütz, Nathalie Gypens, Per J. Hansen, Uwe John, Kerstin Klemm, Jon Lapeya Martin, Nikola Medic, Fabrice Not, Willem Stolte
Data type
Short and full citations
Brief description
This list gives all trophic citations in their short form (used in the dataset), as well as their full reference.
File: oo_444242.txt
Dataset on trophic modes of aquatic protist
Lisa K. Schneider, Konstantinos Anestis, Joost Mansour, Anna A. Anschütz, Nathalie Gypens, Per J. Hansen, Uwe John, Kerstin Klemm, Jon Lapeya Martin, Nikola Medic, Fabrice Not, Willem Stolte
Data type
Trophic mode and taxonomy of aquatic protists
Brief description
This dataset lists the trophic modes of aquatic protists (with reference and data origin) along with their accepted scientific name, AphiaID and taxonomic hierarchy.
File: oo_444243.csv
Acknowledgements
The authors want to thank Salvador Fernández for reviewing the manuscript and the staff at WoRMS for their quick and efficient handling of the dataset submission.
This project has received funding from the European Union's Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement No 766327 and EMODnet Biology (EC Service contract – EASME/EMFF/2016/1.3.1.2/Lot 5/SI2.750022).
Funding Statement
This project has received funding from the European Union's Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement No 766327.
References
- Calbet Albert, Landry Michael R. Phytoplankton growth, microzooplankton grazing, and carbon cycling in marine systems. Limnology and Oceanography. 2004;49(1):51–57. doi: 10.4319/lo.2004.49.1.0051. [DOI] [Google Scholar]
- Canfield Donald E., Glazer Alexander N., Falkowski Paul G. The evolution and future of Earth’s nitrogen cycle. Science. 2010;330(6001):192–196. doi: 10.1126/science.1186120. [DOI] [PubMed] [Google Scholar]
- Caron David A, Michaels Anthony F, Swanberg Neil R, Howse Frances A. Primary productivity by symbiont-bearing planktonic sarcodines (Acantharia, Radiolaria, Foraminifera) in surface waters near Bermuda. Journal of Plankton Research. 1995;17(1):103–129. doi: 10.1093/plankt/17.1.103. [DOI] [Google Scholar]
- Cho BC, Na SC, Choi DH. Active ingestion of fluorescently labeled bacteria by mesopelagic heterotrophic nanoflagellates in the East Sea, Korea. Marine Ecology Progress Series. 2000;206:23–32. doi: 10.3354/meps206023. [DOI] [Google Scholar]
- Falkowski P. G. The role of phytoplankton photosynthesis in global biogeochemical cycles. Photosynthesis Research, 1994;39(3):235–258. doi: 10.1007/BF00014586. [DOI] [PubMed] [Google Scholar]
- Faure Emile, Not Fabrice, Benoiston Anne-Sophie, Labadie Karine, Bittner Lucie, Ayata Sakina-Dorothée. Mixotrophic protists display contrasted biogeographies in the global ocean. The ISME Journal. 2019;13(4):1072–1083. doi: 10.1038/s41396-018-0340-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Field C. B. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science. 1998;281(5374):237–240. doi: 10.1126/science.281.5374.237. [DOI] [PubMed] [Google Scholar]
- Flynn Kevin J., Stoecker Diane K., Mitra Aditee, Raven John A., Glibert Patricia M., Hansen Per Juel, Granéli Edna, Burkholder Joann M. Misuse of the phytoplankton–zooplankton dichotomy: the need to assign organisms as mixotrophs within plankton functional types. Journal of Plankton Research. 2013;35(1):3–11. doi: 10.1093/plankt/fbs062. [DOI] [Google Scholar]
- Flynn Kevin J, Mitra Aditee, Anestis Konstantinos, Anschütz Anna A, Calbet Albert, Ferreira Guilherme Duarte, Gypens Nathalie, Hansen Per J, John Uwe, Martin Jon Lapeyra, Mansour Joost S, Maselli Maira, Medić Nikola, Norlin Andreas, Not Fabrice, Pitta Paraskevi, Romano Filomena, Saiz Enric, Schneider Lisa K, Stolte Willem, Traboni Claudia. Mixotrophic protists and a new paradigm for marine ecology: where does plankton research go now? Journal of Plankton Research. 2019;41(4):375–391. doi: 10.1093/plankt/fbz026. [DOI] [Google Scholar]
- Ghyoot Caroline, Lancelot Christiane, Flynn Kevin J., Mitra Aditee, Gypens Nathalie. Introducing mixotrophy into a biogeochemical model describing an eutrophied coastal ecosystem: The Southern North Sea. Progress in Oceanography. 2017;157:1–11. doi: 10.1016/j.pocean.2017.08.002. [DOI] [Google Scholar]
- Haraguchi Lumi, Jakobsen Hans H., Lundholm Nina, Carstensen Jacob. Phytoplankton community dynamic: A driver for ciliate trophic strategies. Frontiers in Marine Science. 2018;5:272. doi: 10.3389/fmars.2018.00272. [DOI] [Google Scholar]
- HELCOM HELCOM Phytoplankton biovolumes. [2019-07-06T00:00:00+03:00];http://www.helcom.fi/helcom-at-work/projects/phytoplankton 2019
- John Uwe. Data of HE513 Pangea. [2020-07-17T00:00:00+03:00];https://doi.pangaea.de/10.1594/PANGAEA.897217 2020
- Lawerence C, Menden-Deuer S. Drivers of protistan grazing pressure: seasonal signals of plankton community composition and environmental conditions. Marine Ecology Progress Series. 2012;459:39–52. doi: 10.3354/meps09771. [DOI] [Google Scholar]
- Leles S. G., Mitra A., Flynn K. J., Stoecker D. K., Hansen P. J., Calbet A., McManus G. B., Sanders R. W., Caron D. A., Not F., Hallegraeff G. M., Pitta P., Raven J. A., Johnson M. D., Glibert P. M., Våge S. Oceanic protists with different forms of acquired phototrophy display contrasting biogeographies and abundance. Proceedings of the Royal Society B: Biological Sciences. 2017;284(1860) doi: 10.1098/rspb.2017.0664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leles Suzana Gonçalves, Mitra Aditee, Flynn Kevin John, Tillmann Urban, Stoecker Diane, Jeong Hae Jin, Burkholder JoAnn, Hansen Per Juel, Caron David A., Glibert Patricia M., Hallegraeff Gustaaf, Raven John A., Sanders Robert W., Zubkov Mikhail. Sampling bias misrepresents the biogeographical significance of constitutive mixotrophs across global oceans. Global Ecology and Biogeography. 2019;28(4):418–428. doi: 10.1111/geb.12853. [DOI] [Google Scholar]
- Martin Jon Lapeyra. Belgian coastal zone sampling with RV Simon Stevin 2020
- Maselli Maira, Altenburger Andreas, Stoecker Diane K., Hansen Per J. Ecophysiological traits of mixotrophic Strombidium spp. Journal of Plankton Research. 2020;43(5) doi: 10.1093/plankt/fbaa041. [DOI] [Google Scholar]
- Mitra A., Flynn K. J., Burkholder J. M., Berge T., Calbet A., Raven J. A., Granéli E., Glibert P. M., Hansen P. J., Stoecker D. K., Thingstad F., Tillmann U., Våge S., Wilken S., Zubkov M. V. The role of mixotrophic protists in the biological carbon pump. Biogeosciences. 2014;11(4):995–1005. doi: 10.5194/bg-11-995-2014. [DOI] [Google Scholar]
- Mitra Aditee, Flynn Kevin J., Tillmann Urban, Raven John A., Caron David, Stoecker Diane K., Not Fabrice, Hansen Per J., Hallegraeff Gustaaf, Sanders Robert, Wilken Susanne, McManus George, Johnson Mathew, Pitta Paraskevi, Våge Selina, Berge Terje, Calbet Albert, Thingstad Frede, Jeong Hae Jin, Burkholder JoAnn, Glibert Patricia M., Granéli Edna, Lundgren Veronica. Defining planktonic protist functional groups on mechanisms for energy and nutrient acquisition: Incorporation of diverse mixotrophic strategies. Protist. 2016;167(2):106–120. doi: 10.1016/j.protis.2016.01.003. [DOI] [PubMed] [Google Scholar]
- Moorthi S. D., Ptacnik Robert, Sanders R. W., Fischer Robert, Busch Michaela, Hillebrand Helmut. The functional role of planktonic mixotrophs in altering seston stoichiometry. Aquatic Microbial Ecology. 2017;79:235–245. doi: 10.3354/ame01832. [DOI] [Google Scholar]
- Nielson L., Krock B., Hansen P. J. Effects of light and food availibility on toxin production, growth and photosynthesis in Dinophysis acuminata. Marine Ecology Progress Series. 2012;471:37–50. doi: 10.3354/meps10027. [DOI] [Google Scholar]
- Pernice Massimo C, Forn Irene, Gomes Ana, Lara Elena, Alonso-Sáez Laura, Arrieta Jesus M, del Carmen Garcia Francisca, Hernando-Morales Victor, MacKenzie Roy, Mestre Mireia, Sintes Eva, Teira Eva, Valencia Joaquin, Varela Marta M, Vaqué Dolors, Duarte Carlos M, Gasol Josep M, Massana Ramon. Global abundance of planktonic heterotrophic protists in the deep ocean. The ISME Journal. 2014;9(3):782–792. doi: 10.1038/ismej.2014.168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rijkswaterstaat Servicedesk data. [2020-07-17T00:00:00+03:00];www.rws.nl. 2019
- Sarthou G, Timmermans K. R., Blain S., Tréguer P. Growth physiology and fate of diatoms in the ocean. Journal of Sea Research. 2005;53(1-2):25–42. doi: 10.1016/j.seares.2004.01.007. [DOI] [Google Scholar]
- Schoener DM, McManus GB. Plastid retention, use, and replacement in a kleptoplastidic ciliate. Aquatic Microbial Ecology. 2012;67(3):177–187. doi: 10.3354/ame01601. [DOI] [Google Scholar]
- Selosse Marc‐André, Charpin Marie, Not Fabrice. Mixotrophy everywhere on land and in water: the grand écart hypothesis. Ecology Letters. 2016;20(2):246–263. doi: 10.1111/ele.12714. [DOI] [PubMed] [Google Scholar]
- Šimek Karel, Grujčić Vesna, Nedoma Jiří, Jezberová Jitka, Šorf Michal, Matoušů Anna, Pechar Libor, Posch Thomas, Bruni Estelle P., Vrba Jaroslav. Microbial food webs in hypertrophic fishponds: Omnivorous ciliate taxa are major protistan bacterivores. Limnology and Oceanography. 2019;64(5):2295–2309. doi: 10.1002/lno.11260. [DOI] [Google Scholar]
- Stoecker Diane K., Hansen Per Juel, Caron David A., Mitra Aditee. Mixotrophy in the marine plankton. Annual Review of Marine Science. 2017;9(1):311–335. doi: 10.1146/annurev-marine-010816-060617. [DOI] [PubMed] [Google Scholar]
- Stoecker Diane K., Lavrentyev Peter J. Mixotrophic plankton in the Polar Seas: A Pan-Arctic review. Frontiers in Marine Science. 2018;5 doi: 10.3389/fmars.2018.00292. [DOI] [Google Scholar]
- Tréguer Paul J, De La Rocha Christina L. The world ocean silica cycle. Annual review of marine science. 2013;5:477–501. doi: 10.1146/annurev-marine-121211-172346. [DOI] [PubMed] [Google Scholar]
- Unrein Fernando, Massana Ramon, Alonso-Sáez Laura, Gasol Josep M. Significant year‐round effect of small mixotrophic flagellates on bacterioplankton in an oligotrophic coastal system. Limnology&Oceanography. 2007;52(1):456–469. doi: 10.4319/lo.2007.52.1.0456. [DOI] [Google Scholar]
- Vandepitte L, Vanhoorne B, Decock W., Dekeyzer S., Trias Verbeeck A., Bovit L., Hernandez F., Mees J. How Aphia - the platform behind several online and taxonomically oriented databases - can serve both the taxonomic community and the field of biodiversity informatics. Journal of Marine Science and Engineering. 2015;3(4) doi: 10.3390/jmse3041448. [DOI] [Google Scholar]
- Ward Ben A., Follows Michael J. Marine mixotrophy increases trophic transfer efficiency, mean organism size, and vertical carbon flux. Proceedings of the National Academy of Sciences. 2016;113(11):2958–2963. doi: 10.1073/pnas.1517118113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wickham Hadley. Tidy Data. Journal of Statistical Software. 2014;59(10) doi: 10.18637/jss.v059.i10. [DOI] [Google Scholar]
- Board WoRMS Editorial. World Register of Marine Species. Available from http://www.marinespecies.org at VLIZ. Accessed 2019-07-03. VLIZ. 2017 doi: 10.14284/170. [DOI]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
List of trophic citations
Lisa K. Schneider, Konstantinos Anestis, Joost Mansour, Anna A. Anschütz, Nathalie Gypens, Per J. Hansen, Uwe John, Kerstin Klemm, Jon Lapeya Martin, Nikola Medic, Fabrice Not, Willem Stolte
Data type
Short and full citations
Brief description
This list gives all trophic citations in their short form (used in the dataset), as well as their full reference.
File: oo_444242.txt
Dataset on trophic modes of aquatic protist
Lisa K. Schneider, Konstantinos Anestis, Joost Mansour, Anna A. Anschütz, Nathalie Gypens, Per J. Hansen, Uwe John, Kerstin Klemm, Jon Lapeya Martin, Nikola Medic, Fabrice Not, Willem Stolte
Data type
Trophic mode and taxonomy of aquatic protists
Brief description
This dataset lists the trophic modes of aquatic protists (with reference and data origin) along with their accepted scientific name, AphiaID and taxonomic hierarchy.
File: oo_444243.csv