

Abstract
The determination of mycotoxins content in food is not sufficient for the prediction of their potential in vivo cytotoxicity because it does not reflect their bioavailability and mutual interactions within complex matrices, which may significantly alter the toxic effects. Moreover, many mycotoxins undergo biotransformation and metabolization during the intestinal absorption process. Biotransformation is predominantly the conversion of mycotoxins meditated by cytochrome P450 and other enzymes. This should transform the toxins to nontoxic metabolites but it may possibly result in unexpectedly high toxicity. Therefore, the verification of biotransformation and bioavailability provides valuable information to correctly interpret occurrence data and biomonitoring results. Among all of the methods available, the in vitro models using monolayer formed by epithelial cells from the human colon (Caco-2 cell) have been extensively used for evaluating the permeability, bioavailability, intestinal transport, and metabolism of toxic and biologically active compounds. Here, the strengths and limitations of both in vivo and in vitro techniques used to determine bioavailability are reviewed, along with current detailed data about biotransformation of mycotoxins. Furthermore, the molecular mechanism of mycotoxin effects is also discussed regarding the disorder of intestinal barrier integrity induced by mycotoxins.
Keywords: permeability, bioavailability, intestinal transport, metabolism, mycotoxins, biotransformation, cytochrome
1. Introduction
Mycotoxins are toxic secondary metabolites secreted by fungi and frequently occurring in food and feed worldwide [1,2,3]. The major fungal genera producing foodborne mycotoxins are Fusarium, Aspergillus, Penicillium, and Alternaria [4,5]. Fusarium is one of the most important producers of toxins falling into the three major classes of mycotoxins, such as fumonisins (FBs), zearalenone (ZEA), trichothecenes (deoxynivalenol (DON), nivalenol (NIV), and T-2 and HT-2 toxins, and also emerging mycotoxins involving beauvericin (BEA) and enniatins (ENNs) [6,7]. Ochratoxin A (OTA), the major mycotoxin of the ochratoxins, is produced by various species of the Aspergillus and Penicillium genus [8]. In addition, Penicillium species are known to produce mycophenolic acid (MPA) [9] and patulin (PAT) [10,11]. Aflatoxins, including aflatoxin B1 (AFB1), aflatoxin B2 (AFB2), aflatoxin M1 (AFM1), aflatoxin M2 (AFM2), aflatoxin G1 (AFG1), and aflatoxin G2 (AFG2), are the most studied group of mycotoxins produced by Aspergillus flavus [12,13]. Alternaria fungi contaminate a wide variety of food items, such as cereals, fruits, wheat, barley, and sorghum, producing several toxins, with alternariol (AOH), alternariol monomethyl ether (AME), tentoxin (TEN), tenuazonic acid (TeA), altenuene (ALT), and altertoxins (ATXs) being the most relevant ones [14,15]. The effects of mycotoxins on cell functions are listed in Table 1.
Table 1.
Toxic effects of mycotoxins.
| Mycotoxins | Effects | LD50 (mg/kg) | References |
|---|---|---|---|
| T-2 and HT-2 | Inhibition of DNA, RNA and protein synthesis. Induction of mutations and apoptosis. |
T-2 Rodents: 5–10 Pig: 5 Chicken: 2–6 Shrimp: 30 Mice: 2–5 HT-2 Rodents: 5–10 |
[16,17,18,19,20,21] |
| DON | Inhibition of DNA, RNA and protein synthesis. Decrease of the cell proliferation. |
Mice: 46–78 Duck: 27 Chicken: 140 |
[22,23,24,25,26] |
| ZEA | Activation of the estrogen receptor. Inhibition of DNA and protein synthesis. Triggering lipid peroxidation and cell death. |
Mice: 2000–20,000 Rat: 4000–10,000 Pig: 5000 |
[27,28,29] |
| BEA | Increase of the biological membrane permeability. Loss of ionic homeostasis. Induction of lipid peroxidation. |
Mice: 100 | [30,31,32] |
| ENNs | Increase of the membrane permeability for cations. | No acute in vivo toxicity data | [32,33] |
| FB1 | Inhibition the activity of ceramide synthase. | >1000 | [34] |
| AOH and AME | Single and double strand DNA breaks. Decrease of the cell proliferation. |
Mice: 400 for AOH and AME | [35,36,37] |
| ATXs | DNA strand breaks. | Mice: 0.2 | [37,38] |
| TeA | Inhibition of protein synthesis. Inhibition of photosynthetic activity. |
Mice: 81(female), 186–225 (male) Rat: 168 (female), 180 (male) |
[39,40,41,42] |
| AFB1 | Damage of DNA Inhibition of protein synthesis through interfering with RNA transcription and translation. Induction of oxidative stress. |
Swine: 0.62 Duck: 0.37 Turkey: 0.5–1 Chicken: 6.5–12.5 Quail: 19.5 |
[22,43] |
| MPA | Inhibition of inosine 5′-monophosphate dehydrogenase. Blocking of the DNA synthesis and proliferation of both T and B lymphocytes. |
Rat: 450 Mice: 1900 |
[40,44] |
| OTA | Inhibition the activity of many enzymes which use phenylalanine as a substrate. Disruption of phenylalanine metabolism. Production of reactive oxygen species Lipid peroxidation, cell membranes and DNA damage |
Dog: 0.2 Pig: 1 Chicken: 3.3 Rat and mouse: 20–50 |
[34,45] |
T-2 toxin (T-2), HT-2 toxin (HT-2), deoxynivalenol (DON), zearalenone (ZEA), beauvericin (BEA), enniatins (ENNs), fumonisin B1 (FB1), alternariol (AOH), alternariol monomethyl ether (AME), altertoxins (ATXs), tenuazonic acid (TeA), aflatoxin B1 (AFB1), mycophenolic acid (MPA), and ochratoxin A (OTA). LD50: Median lethal dose.
The naturally ubiquitous occurrence of mycotoxins in food has been widely documented (Table 2). Thus, mycotoxins can contaminate a variety of foodstuffs, such as grain-based products (wheat, oats, barley, maize, and rye), nuts, dried fruits, spices, cocoa, coffee, beer, wine, fruits, meat, and animal products (eggs, milk, and cheese) [14,46,47,48,49,50]. Some mycotoxins are thermostable, allowing them to endure most food processes [51]. DON is stable up to 120–180 °C and was degraded after 40 min at 210 °C [52]. For ZEA, heat treatment at temperatures up to 160 °C had no significant effect and 85% reduction of the toxin concentration in barley flours was achieved after 60 min at 220 °C [53]. DON content in whole and white breads decreased by 49% and 39%, respectively, compared to the original flours [51]. According to Generotti et al. [54], increasing pH and baking time in an acceptable technological range can reduce DON concentration in the final product [54]. T-2 and HT-2 are relatively stable during the thermal process up to 170 °C and about 45% of T-2 and 20% HT-2 were thermally degraded during biscuit-making [55]. Similarly, the loss of OTA, ZEA, AFB1, BEA, and ENNs in the products showed that thermal processing effectively decreased the content of these mycotoxins [52,56,57]. The 54% AOH reduction was reported for treatment at 110 °C for 20 min, whereas no significant effect was found for AME at the same temperature [58]. Based on Stadler et al. [59], the parent mycotoxin can be structurally modified during food processing, including isomerization, decarboxylation, rearrangements, and the reaction with other small molecules [59]. In summary, food processing such as brewing, cooking, baking, frying, canning flaking, nixtamalization, and extrusion, in general, reduced concentrations of some mycotoxins but did not eliminate them completely [52,59,60].
Table 2.
Occurrence of mycotoxins in food commodities.
| Mycotoxins | Commodity | Concentration Range (µg/kg) | Country | References |
|---|---|---|---|---|
| T-2 and HT-2 | Barley grain | 26–787 | Italy | [61,62] |
| Maize | 146 | Hungary | ||
| Cereal-based products | <LOD-209 | Tunisia | ||
| Wheat | 6.7–15.2 | Spain | ||
| DON | Cereal and corn | 96–1790 | Portugal | [63,64] |
| Wheat-based product | 333–1821 | Portugal | ||
| Maize grain | ND-700 | Ethiopia | ||
| Sorghum grain | 40–112 | Ethiopia | ||
| ZEA | Corn | 59–505 | Philippines | [63,64,65] |
| Cereal and corn | 5–930 | Portugal | ||
| Sorghum grain | 7.2–382 | Ethiopia | ||
| BEA | Rice | 3800–26,300 | Morocco | [46] |
| Cereal | 0.1–10,600 | Morocco | ||
| ENN A | Rice | 8400–119,500 | Morocco | [46] |
| ENN A1 | Rice | 56,200–448,700 | Morocco | |
| ENN B | Rice | 4400–26,200 | Morocco | |
| ENN B1 | Rice | 3600–23,700 | Morocco | |
| FB1 | Maize | ND-1106 | Zimbabwe | [3,63] |
| Industrial processed food | 43–836 | Nigeria | ||
| Dried sweet potato chips | 29.34–628.78 | Tanzania | ||
| Corn | 113–1162 | Portugal | ||
| Corn products | 183–2026 | Portugal | ||
| AOH | Tomato sauce | 1.2–20.8 | Europe | [46,64,66] |
| Sunflower oil | 0.7–2.9 | Europe | ||
| Sorghum grain | 75–1090 | Ethiopia | ||
| Cereal | 0.75–832 | Germany | ||
| Fruit juices | 15–100 | Germany | ||
| AME | Tomato sauce | <LOQ-4.7 | Europe | [46,64,66] |
| Sunflower oil | <LOQ-7.1 | Europe | ||
| Sorghum grain | 13–257 | Ethiopia | ||
| Cereal | 0.3–905 | Germany | ||
| Fruit juices | 0.13–4.9 | Germany | ||
| ALT | Tomato products | 6.1–62 | Belgium | [46] |
| Fruit juices | 1.18–18.4 | Germany | ||
| ATXs | Tomato sauce | 0.5–3.7 | Europe | [66] |
| Sunflower oil | 2–4.7 | Europe | ||
| TeA | Tomato sauce | <LOQ-691 | Europe | [46,66] |
| Sunflower oil | 24–458 | Europe | ||
| Fruit juices | 1.1–250 | Germany | ||
| Infant food | 0.8–1200 | Germany | ||
| TEN | Tomato sauce | 0.2–1.2 | Europe | [46,66] |
| Sunflower oil | <LOQ-21.8 | Europe | ||
| Fruit juices | 0.5–10.7 | Germany | ||
| AFB1 | Polished rice | 1–2546 | Philippines | [64,65] |
| Sorghum grain | <7.5–359 | Ethiopia | ||
| PAT | Apples | 3.2–1500 | Portugal | [63] |
| Quince jam | 9.7–28.7 | Portugal | ||
| OTA | Cereals | 0.27–7.97 | Portugal | [63,64,65] |
| Coffee beans | 8-36,561 | Philippines | ||
| Sorghum grain | 3.7–163 | Ethiopia |
LOQ: Limit of quantitation; ND: Not detected.
Ingestion of contaminated food is considered as a major route for exposure to many mycotoxins [51]. Upon ingestion, mycotoxins may induce local toxicity or cross the intestinal barrier to enter the bloodstream and reach target organs [2]. Nevertheless, to achieve any effect in a specific tissue or organ, the mycotoxins must be available in effective concentration at certain location, which refers to the compound’s tendency to be extracted from the food matrix, and they must then be absorbed from the gut via the intestinal cells [67]. The term bioaccessibility refers to the fraction of a mycotoxin liberated from a food matrix that passes unmodified through complex biochemical reactions related to the gastrointestinal digestion and thus becomes available for absorption in the small intestine [68,69]. Bioaccessibility can be considered as an indicator for the maximal absorption of the toxin, which can be used for realistic worst-case risk assessment of the toxin in a consumer product [70]. In fact, foodborne mycotoxins can be degraded or modified by metabolic processes of the human body, and only a fraction of the initial content can pass the intestinal membrane to enter the bloodstream [71]. In this sense, bioavailability is defined as the portion of ingested contaminant in food that reaches the systemic circulation [72].
To determine the bioavailability of mycotoxins, different in vitro models or in vivo experiments have been carried out. In vivo experiments would be the best way to evaluate the efficacy of binding capacities [73]. However, to avoid the ethically questionable use of animals in the in vivo experiments, the in vitro models have been used instead. The bioavailability studies carried out in animals are complex, expensive, and lengthy, while the in vitro experiments can be simple, rapid, and cost-effective [72]. The advantages and disadvantages of each procedure are summarized in Table 3. Most of the in vitro studies of the gut were done with human colon tumorigenic cell lines Caco-2, T84, TC7, and HT-29 [74]. The brief description of the expression of transporters, enzymes, and other relevant proteins of available cell lines used for the in vitro biotransformation and bioavailability of drugs and xenobiotics is stated in Table 4. Among commercially available cell lines, Caco-2 cells have been widely used to study absorption, metabolism, and bioavailability of drugs and xenobiotics [2,74]. This model is generally suitable for screening drug and nutrient compounds due to a good in vitro–in vivo correlation [75].
Table 3.
Advantages and disadvantages of in vivo and in vitro models in the evaluation of bioavailability.
| Models | Advantages | Disadvantages |
|---|---|---|
| In vitro models | ||
| Simulation of gastrointestinal transformation | Similar to the physiological processes in the human body Suitable for high-throughput format Ability of testing a specific mechanisms of action Focus on small number of components Validation with reference material |
No hormonal and nervous control Lack of feedback mechanisms Absence of mucosal cell activity Deficiency of complexity of peristaltic movements, and involvement of the local immune system Homeostatic mechanisms are not present Difficult to achieve the anaerobic assay conditions |
| Caco-2 cells | Reproducibility of results Provides information about efficiency of digestion, absorption Ability of studying transport mechanisms Phenotypically similar to absorptive epithelial cells Suitable for high-throughput format |
Human colonic adenocarcinoma origin Higher TEER value than human intestine Lack of mucin, microflora, biofilms, and epithelial cell types Variation of efflux transporters expression levels Incapability of simulating the changes of pH |
| In vivo models | In vivo condition Well-known biology Selection of specific subjects Better-understanding kinetic of mycotoxins |
High-throughput limitation Extremely complex functional systems Influence of different factors-phenotypic variation Lack of certified reference standards Ethical issues and high cost Time consuming and labor intensive |
TEER: Transepithelial electrical resistance.
Table 4.
Available human and animal cell lines used for in vitro biotransformation and bioavailability of drugs and xenobiotics.
| Cell line | Origin | Transporters, Enzymes and Other Relevant Proteins | References |
|---|---|---|---|
| Caco-2 | Human colon adenocarcinoma | CYP1A1, 1A2 GST, UGT, SULT, NAT P-gp, MRP-2, BCRP |
[76,77,78] |
| HT-29 | Human colon adenocarcinoma | CYP2C8, CYP2J2, CYP3A4 GST, UGT MRP1, MRP2, p-gp, BCRP |
[79,80,81] |
| TC-7 | Caco-2 subclones | Similar to Caco-2 | [82] |
| T84 | Human colonic carcinoma | P-gp, MRP2, MRP3 | [83,84] |
| H4 | Human small foetal intestine | CYP3A4 | [85] |
| IPEC-J2 | Neonatal pig small intestine | CYP1A1, 1A2, 3A29 P-gp, MRP1, BCRP |
[86,87] |
P-glycoprotein (P-gp), multidrug resistance protein (MRP), breast cancer resistance protein (BCRP), uridinediphosphoglucuronosyl transferase (UGT), sulfotransferase (SULT), N-acetyltransferase (NAT), glutathione–S–transferase (GST), and cytochrome P (CYP).
This review mainly focuses on the biotransformation of mycotoxins via the expression regulation of some critical enzymes and the currently available data regarding the in vitro study of the bioavailability of mycotoxins using the Caco-2 monolayer. Furthermore, the usefulness and limitations of this model are also discussed.
2. Biotransformation of Mycotoxins
Mycotoxins biotransformation is defined as all the complex modifications which alter the structure of mycotoxins by chemical reactions within the body [88]. Biotransformation is often referred to detoxification, but biotransformation enzymes can also convert certain chemicals into highly toxic metabolites (Figure 1) in a process known as bioactivation [89]. Biotransformation of mycotoxins involves two distinct stages, namely phase I and phase II. The biotransformation process allows metabolites created during phase I to enter conjugation processes (phase II), but in some cases, the substances may be eliminated directly after phase I [90].
Figure 1.

Major biotransformation and adverse cellular effects of mycotoxins. CYP450: Cytochrome P450; UGT: Uridine 5′-diphospho-glucuronosyltransferase; GST: Glutathione S-transferase; ROS: Reactive oxygen species.
In phase I, the mycotoxin could be oxidized, reduced, or hydrolyzed based on their chemical structure [90]. The enzymes involved in detoxification belong to the cytochrome P (CYP) superfamily. The CYP superfamily contains the enzymes involved in oxidative metabolism, such as monooxygenases, prostaglandin synthases, amine oxidases and alcohol dehydrogenases; and reductive metabolism mainly governed by epoxide hydrolases, and aldehyde or ketone reductases [91]. CYP450 enzymes play an important role in the oxidative and reductive metabolism of many endogenous or exogenous chemical compounds [34], including most mycotoxins (Table 5). In mammals, CYPs are present in the endoplasmic reticulum and mitochondria of most cells [89]. Among CYPs, CYP3A with an average content from 50–70% of total enteric CYPs is the major subfamily expressed in the human small intestine [92].
Table 5.
CYP450 isoforms induced by mycotoxins and their phase I and II metabolites.
| Mycotoxins | Induced CYP450 | Phase I Biotransformation | Phase II Biotransformation | References |
|---|---|---|---|---|
| T-2 and HT-2 | CYP3A46, 3A29 and 3A22 in pig CYP1A5, 3A37 in chicken CYP1A1 in human |
NEO, 3′-OH-T-2, 3′-OH-HT-2, T-2 triol, T-2 tetraol, and some C12,13-deepoxy products | T-2 glucuronides HT-2 glucuronides |
[34,93,94,95,96,97,98,99,100,101] |
| DON | CYP2B1 and 2B2 | DOM-1 | DON-3-gluccoside, DON-, DOM- and DON -3-Glucoside-sulfonates, DON-3-, DON-7-, DON-8- and DON-15- glucuronides | [34,102] |
| ZEA | CYP1A1, 1A2, 2B6, 2C9, 3A4 and 3A5 in human CYP2C7, 2E1, 3A1 and 3A2 in rat |
α-ZEA and β-ZEA | ZEA, α-ZEA and β-ZEA-glucuronides ZEA-14-Glucoside, α-ZEA-14-Glucoside, β-ZEA-14-Glucoside, ZEA-14-Sulfate and ZEA-16-Glucoside |
[34,103,104,105,106] |
| BEA | CYP3A4/5 and CYP2C19 in human CYP3A1/2 in rat |
No metabolites detected | No metabolites detected | [107,108] |
| ENNs | CYP3A4, 2C9, 1A2 in human CYP3A and 1A in rat and dog |
M1–M12 with rat, dog and human liver microsomes M1–M5, M9–M13 in chicken |
No sulfated or glucuronidated of ENN B and B1 detected | [109,110,111,112,113,114,115] |
| FB1 | CYP 1A1 and 4A1 in rat CYP1B1 in human |
HFB1 and pHFB1 | Unknown | [34,116] |
| AOH and AME | CYP1A1 | OH-AOH and OH-AME | AOH-3-glucoside, AOH-9-glucoside and AME-3-glucoside | [117,118,119,120] |
| ATXs | CYP1A1 | ATX I | No metabolites detected | [121] |
| ALT | Unknown | OH-ALT | ALT-glucuronide | [121] |
| TeA | Unknown | No metabolites detected | No metabolites detected | [121] |
| TEN | CYP3A4 | Monooxidized, mono-methylated and di-methylated metabolites | Unknown | [121] |
| AFB1 | CYP1A1, 1A2, 2B6, 2C9, 3A4 and 3A5 in human liver | AFBO, AFM1, AFL, AFQ1 and AFP1 | AFB1-glutathiones, glucuronides and sulfates | [34,122,123] |
| OTA | CYP1A1, 1A2, 2B6, 2C9, 3A4 and 3A5 in human liver | Lactone-open OTA, OTα, OTB, 4-OH-OTA and 10-OH-OTA | OTA-glutathiones, OTA-hexose/pentose, OTA-sulfates | [34,124,125,126,127,128,129] |
| PAT | CYP1A1, 1A2, 2B6, 2C9, 3A4 and 3A5 in human hepatocytes | E-ascladiol, Z-ascladiol, hydroascladiol and deosypatulinic acid | PAT-glutathiones | [34,130] |
T-2 toxin (T-2), HT-2 toxin (HT-2), neosolaniol (NEO), deoxynivalenol (DON), zearalenone (ZEA), beauvericin (BEA), enniatins (ENNs), fumonisin B1 (FB1), alternariol (AOH), alternariol monomethyl ether (AME), altenuene (ALT), altertoxins (ATXs), tenuazonic acid (TeA), Tentoxin (TEN), aflatoxin B1 (AFB1), mycophenolic acid (MPA), ochratoxin A (OTA), patulin (PAT), 3′-hydroxy-T-2 (3′-OH-T-2), 3′-hydroxy-HT-2 (3′-OH-HT-2), hydroxy-alternariol (OH-AOH), hydroxy-alternariol monomethyl ether (OH-AME), deepoxy-deoxynivalenol (DOM-1), α-zearalenone (α-ZEA), β-zearalenone (β-ZEA), hydroxy-altenuene (OH-ALT), AFB1–8,9-epoxide (AFBO), aflatoxin M1 (AFM1), aflatoxicol (AFL), aflatoxin Q1 (AFQ1), aflatoxin P1 (AFP1), ochratoxin α (OTα), ochratoxin B (OTB), 4-hydroxy-OTA (4-OH-OTA) and 10-hydroxy-OTA (10-OH-OTA), cytochrome P (CYP).
Phase II reactions are known as conjugation reactions, which usually refer to covalent binding of endogenous hydrophilic substances such as glucuronic acid and sulfate. The reactions provide more hydrophilic compounds, which are quickly eliminated. In general, phase II reactions decrease the toxicity [89]. Uridine 5′-diphospho-glucuronosyltransferase (UDP-glucuronosyltransferase–UGT) and glutathione S-transferase (GST) enzymes play an important role in the phase II metabolism [89,91].
Although the liver is the main detoxification organ, extrahepatic tissues in the gastrointestinal tract (GI tract), kidney, and bladder also show metabolic activity. The GI tract is a first physical barrier for mycotoxins but it also influences the biotransformation process and bioavailability of mycotoxins in other ways. Microorganisms from guts have been reported to exhibit the capacity for degrading mycotoxins [131,132,133,134]. Additionally, P-glycoprotein (P-gp) and multidrug resistance protein (MRP), members of the ATP–binding cassette (ABC) superfamily of transport proteins, are able to pump mycotoxins out of the intestinal cells, leading to limit bioavailability of the substrates [71,135]. Both CYP450 and P-gp in the gut play a crucial role in defense mechanisms against mycotoxins that reach the intestinal mucosa [92].
Previous biotransformation studies mainly focused on AFB1, OTA, trichothecenes (T-2 and DON), ZEA, and FBs. Recently, emerging Fusarium and Alternaria mycotoxins have gained more interest [46], although in vivo metabolization data are still limited. The biotransformation products of mycotoxins are summarized in Table 3. These studies revealed that mycotoxins can induce the expression of CYP450 enzymes in animal and human cell lines.
2.1. Biotransformation of Aflatoxins
Native AFB1 itself is not toxic, but the bioactivation by cytochrome CYP450 leads to AFB1-8,9-epoxide (AFBO), which is acutely toxic, mutagenic, and carcinogenic (Figure 2) [34]. Additionally, the metabolic pathway of AFB1 can also give rise to moderately toxic aflatoxicol (AFL) by ketoreduction, mildly toxic AFM1 and relatively nontoxic aflatoxin Q1 (AFQ1) by hydroxylation, and relatively nontoxic aflatoxin P1 (AFP1) by demethylation [123,136]. Their formation is thus considered as a detoxification pathway [137]. CYP enzymes, particularly CYP1A2 and CYP3A4, are predominant in the metabolic activation of AFB1 [122]. The detoxification of AFBO and AFM1 is realized by conjugation with glutathione catalyzed by GST. Otherwise, the unconjugated AFBO is alternatively hydrolyzed to AFB1-dihydrodiol, which is reversibly converted to AFB1-dialdehyde [34,90,138]. AFB1-dialdehyde is metabolized by the enzymes of aldo-keto reductase subfamily 7 (AKR7) and microsomal epoxide hydrolase (mEH) to form the nontoxic AFB1-dialcohol metabolite in humans, rats, mice, and pigs [123,139,140].
Figure 2.

The major metabolic pathways of aflatoxin B1 (AFB1): (A) Aflatoxin M1 (AFM1) and (B) aflatoxin Q1 (AFQ1) by hydroxylation; (C) Aflatoxin P1 (AFP1) by demethylation; (D) AFB1–8,9-epoxide (AFBO) by epoxidation; (E) Aflatoxicol (AFL) by ketoreduction; (F) AFB1-8,9-dihydrodiol by microsomal epoxide hydrolase (mEH); (G) AFB1-dialcohol by aflatoxin-aldehyde reductase (AFAR); and (H) AFBO-glutathione (AFBO-GSH) by conjugation with glutathione. CYP: Cytochrome P; GSTs: Glutathione S-transferases [123].
2.2. Biotransformation of Ochratoxin A
In animals and humans, OTA can be metabolized by both phase I and phase II enzymes to many different products in the liver, kidney, and intestine (Figure 3). Poor biotransformation and slow elimination of metabolites contribute to the toxicity, carcinogenicity, and organ specificity of OTA [139,141]. In the gut, ochratoxin α (OTα), a major metabolite and is formed by carboxypeptidases, which cleave the peptide bond in OTA [34]. Other types of major metabolites of OTA are 4-hydroxy-ochratoxin A (4-OH-OTA) and 10-hydroxyochratoxin A (10-OH-OTA) have been identified from the urine of rats and are also produced by human, pigs, goat, chicken, rat, and rabbit liver microsomes or human bronchial epithelial cells in vitro [142,143,144]. Most of the metabolites of OTA, such as OTα, OTB, 4-OH-OTA, and 10-OH-OTA, are less toxic than the original compound [129,139]. However, opening the lactone ring under alkaline conditions (called the lactone-opened OTA), found in rodents, leads to more toxic metabolites than OTA itself [126]. These phase I-type reactions probably relate to the action of the CYP450 enzyme family, including CYP1A1, CYP1A2, CYP3A1, CYP3A2, CYP3A4, CYP3A5, CYP2B6, and CYP2C9 [124,125,126]. Phase II biotransformation mainly occurs in the liver with conjugation of OTA with sulfate, glucuronide, hexose/pentose, and glutathione [127,128,129].
Figure 3.

The biotransformation of ochratoxin A (OTA): (A) OTα by cleavage of the peptide bond of OTA; (B) lactone-opened OTA by lactone hydrolysis; (C) OTA-quinone by oxidation; (D) 4-hydroxy-ochratoxin A (4-OH-OTA) and (E) 10-hydroxyochratoxin A (10-OH-OTA) by hydroxylation; (F) OTB by dechlorination; (G) OTA-glutathione, OTA-glucuronide and OTA-sulfate by conjugation with glutathione (GSH), glucuronic acid, and sulfate; (H) Hexose/pentose-OTA by conjugation with hexose/pentose, (I) OTA-glutathione by conjugation with glutathione. CYP450: Cytochrome P450; GSTs: Glutathione S-transferases; UGTs: Uridine 5′-diphospho-glucuronosyltransferases [129].
2.3. Biotransformation of Deoxynivalenol
DON is not substrate of phase I metabolism [145]. Major metabolites of DON include the glucuronide and sulphate conjugates of DON (Figure 4) and deepoxy-deoxynivalenol (DOM-1) [146]. DOM-1 showed lower cytotoxicity in pigs [147]. DON conjugates with glycosides or sulfonates to form DON-3-gluccoside (D3G); DON-, DOM- and D3G-sulfonates; and DON-3-, DON-7-, DON-8-, and DON-15- glucuronides identified in porcine, rat, chicken, bovine, and human [34,102,148,149,150]. Other DON-biotransformation products, including DON-glutathione conjugates and the products of glutathione degradation, such as DON-S-cysteinyl-glycine and DON-S-cysteine, have been reported in cereals. Thanks to intestinal microflora, DON could be metabolized in animals and humans but not deposited in the tissues [151,152].
Figure 4.

Phase II biotransformation of deoxynivalenol (DON): (A) Deepoxy-deoxynivalenol (DOM-1) by deepoxidation; (B) DON-3-sulfate, (D) DON-10-sulfate, (G) DON-3-glucoside sulfonate and (H) DOM-1-10-sulfonate by sulfation; (C) DON-glutathiones by conjugation with glutathione; (E) DON-3-glucuronide, DON-7-glucuronide, DON-8-glucuronide, and DON-15-glucuronide by conjugation with glucuronic acid; and (F) DON-3-glucoside by conjugation with glucose. GSTs: Glutathione S-transferases; UGTs: Uridine 5′-diphospho-glucuronosyltransferases [102].
2.4. Biotransformation of T-2 and HT-2
The major metabolic pathways of T-2 include hydroxylation, hydrolysis, deepoxidation, and conjugation (Figure 5) [153]. The typical metabolites of T-2 in human and animals are HT-2 toxin (HT-2), neosolaniol (NEO), 3′-OH-T-2, 3′-OH-HT-2, T-2 triol, T-2 tetraol, and some C12,13-deepoxy products [99,154]. The contributions of the CYP450 enzymes to T-2 metabolism follow the descending order of CYP3A4, CYP2E1, CYP1A2, CYP2C9, and CYP2B6 or CYP2D6 or CYP2C19, in which CYP3A4 contributes the most [93]. In addition, CYP1A1 in human [34]; CYP3A46, CYP3A29, and CYP3A22 in pig [94,95,96]; and CYP1A5 and CYP3A37 in chicken [97,98] mainly convert T-2 to 3′-OH-T-2 and HT-2 to 3′-OH-HT-2 [145]. The carboxylesterase is also an important phase I enzyme, contributing to the rapid metabolism of T-2 to HT-2 [100]. A recent study revealed that cholic acid supplementation promotes the T-2 metabolism through activation of the farnesoid X receptor, which was found to have significantly increased the expression of CYP3A37 [99]. In phase II, glucuronidation of T-2 toxin, HT-2 toxin, and further phase I metabolites essentially contribute to the metabolism and excretion. The transformation of T-2 to T-2-3-glucuronide and HT-2 to HT-2-3-glucuronide and HT-2-4-glucuronide occurs in liver microsomes of rats, mice, pigs and humans [155]. The activities of GSTs and sulfotransferases can be also attributed to the conjugation reaction as a response to T-2 exposure [100,101].
Figure 5.

Metabolic pathway of T-2 toxin (T-2): (A) HT-2 toxin (HT-2), (B) Neosolaniol (NEO), (C) 4-deacetyl-NEO, (D) 15-deacetyl-NEO, (E) T-2 triol and (F) T-2 tetraol by hydrolysis; (G) 3′-hydroxy-T-2, (H) 3′-hydroxy-HT-2 and (I) 3′-hydroxy-T-2 triol by hydroxylation; (J) Deepoxy 3′-hydroxy-T-2 triol, (K) Deepoxy-3′-hydroxy-HT-2; (L) Deeopoxy-T-2 Tetraol by deepoxiadtion; and (M) T-2-3-glucuronide, (N) HT-2-3-glucuronide, and (O) HT-2-4-glucuronide by conjugation with glucuronic acid. UGTs: Uridine 5′-diphospho-glucuronosyltransferases [154].
2.5. Biotransformation of Fumonisins
After oral ingestion, FB1 are excreted primarily in the feces, either in the intact form or converted into aminopentol (HFB1) and partially hydrolyzed FB1 (pHFB1) by the intestinal microbiota (Figure 6) [116]. The supplementation with fumonisin carboxylesterase FumD results in the gastrointestinal degradation of FB1 and is considered as an effective strategy to detoxify FB1 in the digestive tract of turkeys and pigs [156]. The findings of Daud et al. [157] provided evidence that human fecal microbiota are capable of FB1 degradation, and LC-MS/MS fragmentation patterns indicated microbial biotransformation to hydrolyzed and partially hydrolyzed FB1 [157]. FB1 is not metabolized by CYPs. Moreover, it is a selective inhibitor of CYP2C11 and CYP1A2, while the activities of CYP2A1:2A2, CYP2B1:2B2, CYP3A1:3A2, and CYP4A are not significantly affected. The significant inhibition of CYP2C11 might be related to suppressed protein kinase activity as a result of the inhibition of sphingolipid biosynthesis caused by FB1 [158,159,160]. FB1, HFB1, and pHFB1 can be acetylated to form N-acetylated fumonisins with fatty acid of various lengths, and N-acyl forms proved to be more toxic than the parent FB1 [161,162,163].
Figure 6.

Metabolic pathway of fumonisin B1 (FB1): (A) Aminopentol (HFB1) and (B) partially hydrolyzed FB1 (pHFB1) by hydrolysis; (C) N-acyl-HFB1 and (D) N-acyl-FB1 by N-acylation [161,162,163].
2.6. Biotransformation of Zearalenone
ZEA is mainly biotransformed into α-zearalenol (α-ZEA), which shows the highest binding affinity to human and porcine estrogen receptors, whereas in broilers and rats, β-zearalenol (β-ZEA) with the low affinity to the receptor is predominantly produced [103,104,105]. ZEA upregulates mainly mRNA levels of CYP2B6, CYP3A4, CYP1A2 and CYP1A1, followed by CYP3A5 and CYP2C9, together with activation of their transcriptional regulators—aryl the hydrocarbon receptor (AhR), constitutive androstane receptor (CAR), and pregnane X receptor (PXR) [106]. It is well known that ZEA, α-ZEA, and β-ZEA are substrates of UGT, the enzyme responsible for the glucuronidation (Figure 7) [78,105,164,165]. However, the UGT was not only saturated but also inhibited by high concentration of ZEA [166]. Although zearalenone-14-glucoside (ZEA14Glc) has lower toxicity than ZEA due to inability to interact with estrogen receptors, the possible systemic hydrolysis and further activating metabolism of ZEA14Glc leads to ZEA-mediated toxicity [167]. Due to the adverse effect of ZEA on human and animal health, microorganisms have gained great interest in the modulation of ZEA adsorption and transformation [168,169]. Eukaryotic cells were able to biotransform ZEA to α-ZEA and β-ZEA, while prokaryotic cells only absorbed ZEA without any metabolization of this mycotoxin and sequestered ZEA by binding to the cell wall [170,171].
Figure 7.

Metabolic pathway of zearalenone (ZEA): (A) α- zearalenol (B) (α-ZEA) and β- zearalenol (β-ZEA) by hydroxylation; (C) Zearalenone-glucuronide, (D) α-zearalenol-glucuronide and (E) β-zearalenol-glucuronide by glucuronidation; (F) Zearalenone-14-glucoside (ZEA14Glc), (G) Zearalenone-16-glucoside (ZEA16Glc), (H) α- zearalenol-14-glycoside and (I) β-zearalenol-14-glucoside by glycosidation; and (J) Zearalenone-14-sulfate by sulfation. UGTs: Uridine 5′-diphospho-glucuronosyltransferases [170].
2.7. Biotransformation of Enniatins
For ENNs, the most information is currently available for ENN B and B1. In vitro and in vivo studies demonstrated that CYP3A4, CYP2C19, and CYP1A2 play the major role for ENN B metabolism in human microsomes and CYP3A and CYP1A are also included in this process in rats and dogs [109]. The 12 biotransformation products were characterized after the incubation of ENN B with rat, dog, and human liver microsomes (Figure 8): M1–M5 were monohydroxylated and M6 and M7 were N-demethylated, whereas M8–M12 were the result of multiple oxidations [110]. However, only eight metabolites could be detected in the case of chicken liver microsomes, particularly five hydroxylated (M1–M5) and three carboxylated (M9, M11 and M12) metabolites. Moreover, M4 and M13 were major metabolites in egg samples, while M11 and M13 were found in liver and serum samples collected after broilers and hens were given contaminated feed containing ENN B [111]. Similarly, ENN B1 is mainly metabolized by CYP3A4 [112]. In vitro incubation with minipig and slaughter swine liver microsomes resulted in the detection of ten ENN B1 metabolites (M2–M11) and M1 occurred only in the minipig assays, while six metabolites (M5–M8) were detected also in vivo [113]. Rumen microbiota also proved to be able to degrade ENN B up to 72% after 48 h of incubation [114]. Any sulfated or glucuronidated phase II metabolites of ENN B or ENN B1 were detected (Figure 9) [115].
Figure 8.

Molecular structures of ENN B, and B1, and proposed structures of their metabolites [110,111].
Figure 9.

2.8. Biotransformation of Beauvericin
Very few studies have been carried out on BEA (Figure 10) in this regard. No BEA metabolites were detected in the mice feed with BEA in the study of Rodríguez-Carrasco et al. [107], suggesting a higher metabolic stability for BEA [107]. Mei et al. [108] reported that BEA is a potent inhibitor of diverse CYP450 enzymes, including CYP3A4/5 and CYP2C19 in human liver microsomes and CYP3A1/2 in rat liver microsomes [108].
Figure 10.
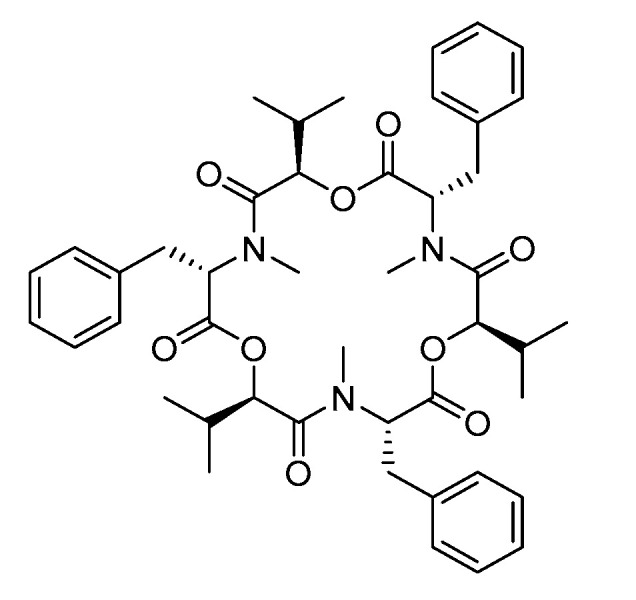
Molecular structure of BEA.
2.9. Biotransformation of Alternaria Mycotoxins
AOH and AME form the metabolites hydroxylated at C-2, C-4, and C-8 by activation of the CYP1A1 enzyme (Figure 11) [172,173]. AOH and AME activate the AhR pathway, which induces CYP1A1 expression [117,118]. AOH is known for its genotoxicity [118]. However, the phase I metabolites, 4-OH-AOH and 4-OH-AME, had minor effect compared to AOH or AME in topoisomerase inhibition and DNA strand-breaking effects [174]. Phase II metabolism includes conjugation with glucuronic acid and sulfate [119]. AME and AOH were enzymatically glycosylated using whole-cell biotransformation system, producing highly effective rates of 58% AOH-3-glucoside, 5% AOH-9-glucoside, and 24% AME-3-glucoside [120]. However, human gut microbiota was not capable of metabolizing AOH, AME, and ALT [175]. The conversion of ATX-II, significantly more genotoxic than AOH, to ATX-I by de-epoxidation in Caco-2 cells did not showed an adequate detoxification but an attenuation of genotoxicity [176]. The metabolic pathway of AOH, AME and other Alternaria mycotoxins, such as TEN, TeA, ALT and ATXs, are summarized in Figure 11.
Figure 11.

Biotransformation pathway of Alternaria mycotoxins: Alternariol (AOH), alternariol monomethyl ether (AME), hydroxy-alternariol (OH-AOH), hydroxy-alternariol monomethyl ether (OH-AME), tenuazonic acid (TeA), altertoxins (ATXs), Tentoxin (TEN), altenuene (ALT), hydroxyl-altenuene (OH-ALT). (A): Demethylation; (B,C,H): Hydroxylation; (D,I): Methylation; (E,F): Sulfation, glycosylation, and glucuronidation; (G): Epoxide reduction. CYP: Cytochrome P; and UGTs: Uridine 5′-diphospho-glucuronosyltransferase [71,117,121,172,173,174,177,178,179,180,181].
2.10. Biotransformation of Patulin
PAT induces the upregulation of PXR and AhR accompanied by the enhancement of CYP1A1, CYP1A2, CYP2B6, CYP2C9, CYP 3A4, and CYP3A5 expression [130]. Moreover, PAT reacts with intracellular glutathione in gastrointestinal mucosa cells [182,183]. The extracellular enzymes of Lactobacillus casei YZU01 induced by PAT mainly degrades PAT, and the cell wall of this bacteria can also absorb a small amount of PAT [184]. Similarly, the degradation of PAT was observed by Saccharomyces cerevisiae during cider fermentation into E-ascladiol and Z-ascladiol (Figure 12), which are not toxic to human [185]. The biotransformation of PAT in humans and animals is not well understood and remains to be established.
Figure 12.

Biotransformation pathway of patulin (PAT): E-ascladiol, Z-ascladiol, hydroascladiol, and desoxypatulinic acid by microorganism, and PAT-glutathiones by reaction with glutathione. GSTs: Glutathione S-transferase [130,182,183,185,186].
3. Assessment of Bioavailability of Mycotoxins Using Caco-2 Cell Monolayer
The Caco-2 cell line is the most common and extensively used in vitro model to study the intestinal absorption of mycotoxins via the intestinal membrane enterocytes [2,10,187,188]. It was originally derived from a heterogeneous human epithelial colorectal adenocarcinoma cells established by Fogh and coworkers in 1977 [189]. The Caco-2 cells have the ability to spontaneously differentiate into a monolayer of cells, expressing many properties typical of absorptive enterocytes with a brush border layer, tight junctions, and efflux and uptake transporters as found in the small intestine [190,191,192]. Moreover, several phenolic compounds (e.g., kaempferol) are able to regulate the MAPK pathway, which is beneficial to the barrier functions [193]. Kaempferol treatment showed significant an increase in claudin 3, claudin 4, and occluden [194]. On the other hand, several mycotoxins—deoxynivalenol, zearalenone, fumonisin B1, T-2 toxin, aflatoxin M1, and ochratoxin A—have a deleterious effect on tight junctions of claudin 3, claudin 4, claudin 7, and occluden [195,196,197,198].
The Caco-2 cells have been shown to be a suitable model for biotransformation study because they express various phase-I hydroxylation and phase-II conjugation enzymes, and transport proteins of the ATP-Binding Cassette (ABC) superfamily [166]. Furthermore, a good correlation has been found for data on oral absorption in humans and the results in the Caco-2 model [199].
To closer mimic the intestinal barrier in vivo, Caco-2 cells were seeded on permeable membranes to form a confluent monolayer with a well-defined tight junction for approximately 21 days post-seeding [78]. The integrity of the Caco-2 monolayer was monitored by measuring the transepithelial electrical resistance (TEER), or by examining the permeability of paracellular markers, such as mannitol, inulin, Dextran, PEG 4000, Lucifer yellow, and phenol red [191,200]. Studies that have investigated the bioavailability of mycotoxins by Caco-2 cells are listed in Table 6. The results of these studies show that mycotoxins are transported through Caco-2 monolayer in different efficiencies.
Table 6.
Studies investigating bioavailability of mycotoxins by Caco-2 cells.
| Mycotoxins | Concentration (µM) | Incubation Time (h) | Major Findings | References |
|---|---|---|---|---|
| AOH and AME | 20 | 1–3 | 22.7–25.8% and 3–7.1% applied AOH and AME reached the basolateral compartment (including their metabolites). | [119] |
| ATXs | 10 | 0.5 | 6% and 0.3% applied ATX I and ATX II found in basolateral compartment. ATX I were not metabolized. 13 and 4% metabolites of ATX II found in apical and basolateral compartments. |
[201] |
| AFB1 | 1–25 | 24–48 | CYP1A2 and 3A4 were the main CYP450 isoforms for AFB1 activation into the genotoxic metabolite aflatoxin-exo-7-8-epoxyde. | [9] |
| AFB1, FB1, OTA and T-2 | 100 | 24 | AFB1, FB1, T2 and OTA disrupted the intestinal barrier permeability. | [198] |
| BEA | 1.5–3 | 4 | Bioavailability was from 50.1–54.3 for BEA | |
| DON | 5-30 | 24 | DON transcellular passage was either by passive/facilitated diffusion or by active transport. DON was a substrate for both P-gp and MRP2. |
[202] |
| ENNs | 1.5–3 | 4 48 |
Duodenal bioavailability: 57.7–76.8% for ENN A, 68.8–70.2% for ENN A1, 65.0–67.0% for ENN B, and 62.2–65.1% for ENN B1. Colonic bioavailability: 17.3–33.3% for ENN A, 40.8–50.0% for ENN A1, 47.7–55.0% for ENN B, and 52.4–57.4% for ENN B1 |
[67] |
| MPA | 0–780 | - | Decrease in the barrier function of Caco-2 cell monolayer. | [9] |
| NIV | 5 | 6 | Bioavailability: 32.6% NIV would not be metabolized in Caco-2 cells. NIV was a substrate for P-gp and MRP2. |
[203] |
| OTA | 1–100 5–45 |
1 3–24 |
OTA was a substrate for MRP2 and BCRP Metabolites were OTB, OTA methyl ester, OTA ethyl ester and the OTA glutathione conjugate. |
[204] [205] |
| ZEA | 25 | 4 | ZEA was substrates for ABCC1, ABCC2 and metabolites into α- and β-zearalenol and glucuronides. | [206] |
Alternariol (AOH), alternariol monomethyl ether (AME), altertoxin (ATXs), aflatoxin B1 (AFB1), fumonisin B1 (FB1), ochratoxin A (OTA), T-2 toxin (T-2), beauverincin (BEA), deoxynivalenol (DON), enniatins (ENNs), mycophenolic acid (MPA), nivalenol (NIV), zearalenone (ZEA), cytochrome P (CYP), P-glycoprotein (P-gp), multidrug resistance protein (MRP), breast cancer resistance protein (BCRP), ATP-Binding Cassette (ABC).
DON, NIV, ZEA ENNs, and BEA cross easily the cell barrier. DON is efficiently transported through the intestinal barrier possibly either by passive/facilitated diffusion [202] or by paracellular passage through intercellular tight junctions [207]. All of the apparent permeability (Papp) values greater than 1 × 10−6 cm/s suggest that these mycotoxins were absorbed efficiently [208]. Papp values for DON have been reported by many researchers. Sergent et al. [207] reported an average Papp value of 5.02 × 10−6 cm/s for absorption (apical (AP)–basolateral (BL) compartment) and excretion (BL–AP direction) [207]. In other study, absorption and excretion Papp values ranged 1.23–2.06 × 10−6 and 2.68–2.8 × 10−6 cm/s, respectively [202]. Finally, Papp value of 3.3 × 10−6 and 2.8 × 10−6 cm/s for absorption and excretion, respectively, were determined in study of Kodota et al. [209]. A faster bidirectional transport of DON in the mixture comparing to pure DON was observed, suggesting that the presence of other mycotoxins including AFB1, FB1, and OTA may promote intestinal transport of DON [210]. For NIV, transcellular transport probably occurred by passive diffusion in the absorptive direction, and Papp values were also higher than 10−6 cm/s [203]. The Papp values obtained with a concentration of 20 µM ZEA in the apical compartment and an incubation time of 1 h were 10.47 ± 4.7 × 10−6 cm/s [211]. About 30% of initial ZEA crossed the cell monolayer after 3 h of exposure, and 40% of ZEA was absorbed by the intestinal after 8 h [78]. ZEA presented higher bioavailability than its metabolites, α-ZEA, ranging from 10% to 36% (0–4 h; 30 µM) [72]. Unlike DON-3-glucoside (neither absorbed or cleaved by Caco-2 cells), ZEA-14Glc and ZEA-16Glc could cross the cell barrier and be absorbed by Caco-2 cells, resulting in further cleavage and the subsequent release of their parent deglycosylated forms [212]. BEA bioavailability was variable from 50% to 54% [213]. Higher duodenal bioavailability compared to colonic bioavailability of ENNs was observed. Particularly, the duodenal bioavailability of ENNs ranged from 58% to 77% for ENN A, from 69% to 70% for ENN A1, from 65% to 67% for ENN B, and from 62% to 65% for ENN B1. Colonic bioavailability ranged from 17% to 33% for ENN A, from 41% to 50% for ENN A1, from 48% to 55% for ENN B, and from 52% to 57% for ENN B1 [67]. In contrast, FB1 was not absorbed by Caco-2 cells [214].
Berger et al. [215] showed that OTA was absorbed by the human intestinal mucosa by passive diffusion of the undissociated form of OTA and it was not appreciably metabolized by Caco-2 cells [215]. DON and NIV were not significantly metabolized or accumulated in Caco-2 cells as well [71,202,203,207,216,217]. Therefore, upon ingestion, these mycotoxins can be absorbed from the gut via intestine cells, then entered into the systemic circulation and thus transported to the whole body. Nevertheless, the intestinal absorption of OTA would be limited thanks to the presence of the MRP2 [215] and breast cancer resistance protein (BCRP) [204]. An efflux of AFB1 was also associated with BCRP [218], and DON was a substrate for both P-gp and MRP2 [202]. P-gp has been shown to be involved in the efflux of FB1 [214], and NIV interacted with P-gp and MRP2 [203]. Several studies showed that DON transport was unaffected by the transporter [207,209]. However, DON uptake and efflux are carrier-mediated processes, and P-gp and organic anion-transporting peptides may be the major efflux/uptake transporters for DON in Caco-2 cells, respectively [219]. The stepwise c-Jun-N-terminal kinase–Akt–nuclear factor kappa-light-chain-enhancer of activated B cells (JNK-Akt-NF-κB) pathway elaborates upon P-gp induction following DON exposure in mammalian cells and provides a self-protection mechanism to resist exogenous toxic compounds such as DON and T-2 [220]. These dissimilarities may be consequences of differences in exposure conditions to the toxin. Particularly, transport experiments were performed in pH gradient, and the acidification of the apical compartment may increase the fraction of the uncharged molecules facilitating diffusion across the cell membrane and intracellular accumulation [221]. Furthermore, differences in the culture medium, passage number, and time in culture before splitting may lead to significant differences in ABC transporter expression and functionality [222].
Intestinal absorption of AOH was more extensive and faster than AME. About 23–26% of the apically applied AOH reached the basolateral compartment, while only about 3–7% of the initial amount of AME in the apical chamber reached the basolateral side. In basolateral medium, several metabolites were also detected: Three AOH metabolites (3-O-sulfate, 3-, and 9-O-glucuronide) and AME-3-O-glucuronide [119]. Several authors have already shown the ability of Caco-2 cells to metabolize ZEA into α- or β-ZEA, as well as into its glucuronidated and sulphated forms [78,166,211]. Videmann et al. [206] established that facilitated or active transport was involved in the transportation of ZEA and its metabolites. Particularly, they were substrates for ABCC1–3 transporters. ZEA and α-ZEA were mostly extruded by ABCC2 at the AP side and ABCC3 was able to transport β-ZEA at the BL side [206].
Treatment of Caco-2 cells with mycotoxins at reasonable concentrations must have no significant effect on cell viability, cell damage, and barrier integrity. FB1 at a concentration of up to 138 µM did not induce variation on cell viability and differentiation [214]. Similarly, ZEA concentration of up to 200 µM had no significant effect on cell viability and cell damage [78,206], and the integrity of the cell monolayers was preserved throughout the incubation with ZEA at a concentration of up to 40 µM, indicating that ZEA does not have detrimental effects on epithelial integrity in vitro [212]. Moreover, Caco-2 cells exposure to 5 µM of NIV showed neither a significant increase in the sucrose flux nor a significant decrease in TEER values [203]. DON also had no significant effect on Caco-2 cell viability at a concentration of up to 33 µM [202,209].
However, other studies reported that mycotoxins such as ZEA, DON, FB1, T-2, PAT, AFB1, and OTA decreased the TEER of intestinal epithelial cell lines in porcine as well as in human epithelium [10,195,196,198,223,224,225,226,227]. A reduction in TEER can cause an increase in the paracellular permeability, changes in transcellular flux through altered plasma channels or pumps, and uncontrolled cell death within the monolayer [228]. Pfeiffer et al. [211] showed that 20 μM of ZEN was able to affect the apparent permeability coefficients of Caco-2 cells, leading to their quick absorption from the intestinal lumen into the portal blood [211]. Moreover, the important indicators of intestinal permeability are tight junction proteins, which are comprised of several multiprotein complexes, including transmembrane proteins (claudin, occludin, and junctional adhesion molecule) and cytoplasmic scaffolding protein and signaling proteins, including zonula occludens [229]. DON, ZEA, FB1, T-2, AFM1, and OTA have a deleterious effect on tight junctions of claudin 3, claudin 4, claudin 7, and occluden [195,196,197,198].
Tight junction structure and function can be regulated by signaling molecules involved in the mitogen-activated protein kinase-dependent (MAPK) pathways [230]. Therefore, the rapid activation of MAPK, ZEA, and DON decreased the expression of tight junction proteins, resulting in intestinal barrier impairments [134,197]. DON and other trichothecenes are known for their binding of the ribosomal peptidyltransferase, inhibition of protein synthesis, and rapid activation of MAPK via inducing two signal transduction pathways of a process named the ribotoxic stress response [227,231,232,233]. The first pathway consists of the double-stranded RNA-activated protein kinase, leading to stimulation of JNK and p38 [25]. The second pathway involves hematopoietic cell kinase belonging to the Src tyrosine kinase family, which are upstream transducers of activation of MAPK. Among the primary MAPK subfamilies, such as p44/42 extracellular signal-regulated protein kinase (ERK), p38, and JNK [234], p44/42 ERK can be involved in intestinal epithelial cell morphology and in the structure of tight junctions. It was reported that the DON-induced activation of the p44/42 ERK signaling pathway inhibits the expression of claudin-4, which leads to reduces the barrier function of the intestine evaluated by TEER, paracellular permeability [197,227]. Treatment with 10 µM of DON also increased ERK, P38, JNK, and c-Jun phosphorylation levels by 2-fold, 30-fold, 61-fold, and 5-fold, respectively, and altered the gene expression levels of occludin, claudin-3, and the composition of tight junction proteins (Figure 13) [235]. The activation of p44/42 MAPK was partially involved in the detrimental effects of the integrity of tight junction caused by AFM1 and OTA [224].
Figure 13.

The effects of DON and other trichothecenes on the tight junction through activation of the MAPK pathway. MAPK: Mitogen-activated protein kinase-dependent, ERK: Extracellular signal regulated protein kinase, JNK: C-Jun-N-terminal kinase. The colored curves represent junction proteins.
In addition to the tight junction, the maintenance of intestinal barrier-related paracellular secretions, such as cytokines and chemokines, are important as well. ZEA metabolites, α- and β-ZEA, can be beneficial to the intestine by decreasing the expression of both interleukin-8 (IL-8) and interleukin-10 in a dose-dependent manner. Its metabolites have a rather anti-inflammatory effect on the epithelial intestinal cells [225]. However, cytokines are related to the impairment of intestinal integrity when exposed to ZEA and FB1 [225,226,236]. Moreover, the correlation between permeability and IL-8 secretion induced by DON in the intestine was investigated by the authors of [209]. IL-8 was examined as a factor affecting intestinal barrier function, and the increased IL-8 secretion may be involved in the TEER decrease [237]. Similar results were reported by the authors of [238]. Consequently, exposure to certain mycotoxins, particularly DON, may cause damage to the intestinal integrity and lead to various chronic intestinal inflammatory diseases, such as inflammatory bowel disease [195]. In addition, the synergic effects of OTA and AFM1 that might exacerbate intestinal inflammation were also reported [239].
Although the Caco-2 cells model offers several advantages, such as the reproducibility of results, controlled environment, and in-depth mechanistic insight [240], some limits of Caco-2 for assessing the bioavailability were also reported [241]. The main disadvantages of these models are the lack of the regulatory processes of the complex mucosal barrier and inability to accurately calculate the fractional transport and flux rate through the static transport conditions [242]. Moreover, it has been shown that significant variation of the expression level of efflux transporters, such as BCRP, MRP2, and MDR1 in the Caco-2 cell monolayer in human small and large intestines, affect the results as well [243,244]. The Caco-2 cell monolayer is somewhat unsuccessful in simulating in vivo intestinal environment due to lack of expression of CYP3A4, which is responsible for the biotransformation of many compounds in the human epithelial cell [245]. Further drawbacks of these models include the incapability of simulating the changes of intestinal pH system, since it is performed at constant pH conditions. In addition, variations in TEER and permeability were also reported to be related to the source of Caco-2 cell and interlaboratory differences in protocol design [192].
To reduce the heterogeneity of the Caco-2 parental cell line and to improve the performance and the stability of this cellular model, some clonal derivative of Caco-2 cells have been established. The Caco-2/TC7 cell line, which was isolated from a late passage of the parental Caco-2 line, is suitable for intestinal absorption model due to a less heterogenic cellular population, resulting in better reproducibility of results [246]. The human intestinal HT-29 cell line is another cell line from colorectal origin with epithelial morphology and has a large proportion of mature goblet cells that can produce mucins. Therefore, the co-culture of Caco-2 and HT-29 with a ratio of 9:1 was used to provide a better representation of the intestinal tract [247]. In addition, the human colon carcinoma (HCT-116) and human colon adenocarcinoma (SW480) cells used in unraveling cancer-related mechanisms and the human duodenum adenocarcinoma (HuTu-80) cell line simulating duodenal cells are less popular [191]. More recently, a combination of in vitro digestion and Caco-2 absorption was used to simulate the physiological settings in the gastrointestinal tract and determine the bioaccessibility and bioavailability of the ZEA reaction products [72].
4. Conclusions
Scientific insights in the production of mycotoxins, their toxicities, biotransformation, and metabolism in different organisms have greatly contributed to a more detailed understanding of the chemical hazards in food. Mycotoxins can notably biotransform and detoxify in the liver, as well as in the digestive tract. The results obtained with Caco-2 monolayer are useful in the prediction of mycotoxins’ intestinal permeability, transport mechanism, and gene regulation of transporters and enzymes in humans, and may help interpret properly data of mycotoxins’ absorption for better comprehension of their possible adverse effects. Furthermore, the combined usage of in vitro digestion models with in vitro intestinal absorption models using Caco-2 cells may offer more complete picture during digestion in the intestinal tract. However, the correlation between in vitro Caco-2 data and in vivo situation necessitates further investigation.
Abbreviations
| 3′-OH-HT-2 | 3′-hydroxy-HT-2 |
| 3′-OH-T-2 | 3′-hydroxy-T-2 |
| 4-OH-OTA | 4-hydroxy-ochratoxin A |
| 10-OH-OTA | 10-hydroxy-ochratoxin A |
| α-ZEA | α-zearalenone |
| β-ZEA | β-zearalenone |
| ABC | ATP–binding cassette |
| AFB1 | aflatoxin B1 |
| AFB2 | aflatoxin B2 |
| AFBO | Aflatoxin B1–8,9-epoxide |
| AFBO-GSH | Aflatoxin B1–8,9-epoxide-glutathiones |
| AFG1 | aflatoxin G1 |
| AFG2 | aflatoxin G2 |
| AFL | aflatoxicol |
| AFM1 | aflatoxin M1 |
| AFM2 | aflatoxin M2 |
| AFP1 | aflatoxin P1 |
| AFQ1 | aflatoxin Q1 |
| AhR | aryl the hydrocarbon receptor |
| AKR7 | aldo-keto reductase subfamily 7 |
| ALT | altenuene |
| AME | alternariol monomethyl ether |
| AOH | alternariol |
| AP | apical compartment |
| ATX I | altertoxin I |
| ATX II | altertoxin II |
| ATXs | altertoxins |
| BCRP | breast cancer resistance protein |
| BEA | beauvericin |
| BL | basolateral compartment |
| Caco-2 | caucasian colon adenocarcinoma |
| Caco-2/TC7 | TC7 clone was isolated from a late passage of the parental Caco-2 line |
| CAR | constitutive androstane receptor |
| CYP | cytochrome P |
| D3G | deoxynivalenol-3-glucoside |
| DNA | Deoxyribonucleic acid |
| DOM-1 | deepoxy-deoxynivalenol |
| DON | deoxynivalenol |
| ENN A | enniatin A |
| ENN A1 | enniatin A1 |
| ENN B | enniatin B |
| ENN B1 | enniatin B1 |
| ENNs | enniatins |
| ERK | extracellular signal regulated protein kinase |
| FB1 | fumonisin B1 |
| FBs | fumonisins |
| GI | gastrointestinal |
| GSH | glutathione |
| GST | glutathione S-transferase |
| HCT-16 | human colon carcinoma |
| HFB1 | aminopentol |
| HT-2 | HT-2 toxin |
| HT-29 | human colorectal adenocarcinoma |
| IL-8 | Interleukin-8 |
| JNK | c-Jun-N-terminal kinase |
| LD50 | median lethal dose |
| LOQ | limit of quantitation |
| MAPK | mitogen-activated protein kinase-dependent |
| mEH | microsomal epoxide hydrolase |
| MPA | mycophenolic acid |
| MRP | multidrug resistance protein |
| NAT | N-acetyltransferaseND (not detected) |
| NEO | neosolaniol |
| NF-κB | nuclear factor kappa–light–chain–enhancer of activated B cells |
| NIV | nivalenol |
| OH-ALT | hydroxy-altenuene |
| OH-AME | hydroxy-alternariol monomethyl ether |
| OH-AOH | hydroxyl-alternariol |
| OTα | ochratoxin α |
| OTA | ochratoxin A |
| OTB | ochratoxin B |
| Papp | apparent permeability |
| P-gp | P-glycoprotein |
| PAT | patulin |
| pHFB1 | partially hydrolyzed fumonisin B1 |
| PXR | pregnane X receptor |
| RNA | Ribonucleic acid |
| ROS | reactive oxygen species |
| SULT | sulfotransferase |
| SW480 | human colon adenocarcinoma |
| T-2 | T-2 toxin |
| TeA | tenuazonic acid |
| TEER | transepithelial electrical resistance |
| TEN | tentoxin |
| UGT: | uridine 5′-diphospho-glucuronosyltransferase |
| ZEA | zearalenone |
| ZEA14Glc | zearalenone-14-glucoside |
| ZEA16Glc | zearalenone-16-glucoside |
Funding
The financial support of the mobility project from the Czech Ministry of Education, Youth and Sports INTER-COST LTC20015 (COST Action CA18111) is gratefully acknowledged. The work was also supported by the Czech National Program of Sustainability NPU I (LO1601, MSMT-43760/2015).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Key Contribution
The paper overviews the biotransformation of mycotoxins in order to transform the toxins to harmless metabolites. However, it may possibly generate products exerting unexpectedly high toxicity.
References
- 1.De Boevre M., Graniczkowska K., Saeger S. De Metabolism of modified mycotoxins studied through in vitro and in vivo models: An overview. Toxicol. Lett. 2015;233:24–28. doi: 10.1016/j.toxlet.2014.12.011. [DOI] [PubMed] [Google Scholar]
- 2.Smith M.C., Gheux A., Coton M., Madec S., Hymery N., Coton E. In vitro co-culture models to evaluate acute cytotoxicity of individual and combined mycotoxin exposures on Caco-2, THP-1 and HepaRG human cell lines. Chem. Biol. Interact. 2018;281:51–59. doi: 10.1016/j.cbi.2017.12.004. [DOI] [PubMed] [Google Scholar]
- 3.Kebede H., Liu X., Jin J., Xing F. Current status of major mycotoxins contamination in food and feed in Africa. Food Control. 2020;110:106975. doi: 10.1016/j.foodcont.2019.106975. [DOI] [Google Scholar]
- 4.Fernández-Blanco C., Elmo L., Waldner T., Ruiz M.J. Cytotoxic effects induced by patulin, deoxynivalenol and toxin T2 individually and in combination in hepatic cells (HepG2) Food Chem. Toxicol. 2018;120:12–23. doi: 10.1016/j.fct.2018.06.019. [DOI] [PubMed] [Google Scholar]
- 5.Pankaj S.K., Shi H., Keener K.M. A review of novel physical and chemical decontamination technologies for aflatoxin in food. Trends Food Sci. Technol. 2018;71:73–83. doi: 10.1016/j.tifs.2017.11.007. [DOI] [Google Scholar]
- 6.Seyed Toutounchi N., Hogenkamp A., Varasteh S., van’t Land B., Garssen J., Kraneveld A.D., Folkerts G., Braber S. Fusarium Mycotoxins Disrupt the Barrier and Induce IL-6 Release in a Human Placental Epithelium Cell Line. Toxins. 2019;11:665. doi: 10.3390/toxins11110665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Stanciu O., Loghin F., Filip L., Cozma A., Miere D., Mañes J., Banc R. Occurence of Fusarium Mycotoxins in Wheat from Europe—A Review. Acta Univ. Cibiniensis. Ser. E Food Technol. 2015;19:35–60. doi: 10.1515/aucft-2015-0005. [DOI] [Google Scholar]
- 8.Raghubeer S., Nagiah S., Chuturgoon A. Ochratoxin A upregulates biomarkers associated with hypoxia and transformation in human kidney cells. Toxicol. In Vitro. 2019;57:211–216. doi: 10.1016/j.tiv.2019.03.016. [DOI] [PubMed] [Google Scholar]
- 9.Hymery N., Mounier J., Coton E. Effect of Penicillium roqueforti mycotoxins on Caco-2 cells: Acute and chronic exposure. Toxicol. In Vitro. 2018;48:188–194. doi: 10.1016/j.tiv.2018.01.017. [DOI] [PubMed] [Google Scholar]
- 10.Assunção R., Ferreira M., Martins C., Diaz I., Padilla B., Dupont D., Bragança M., Alvito P. Applicability of in vitro methods to study patulin bioaccessibility and its effects on intestinal membrane integrity. J. Toxicol. Environ. Heal. Part A Curr. Issues. 2014;77:983–992. doi: 10.1080/15287394.2014.911138. [DOI] [PubMed] [Google Scholar]
- 11.Hussain S., Asi M.R., Iqbal M., Khalid N., Wajih-ul-Hassan S., Ariño A. Patulin Mycotoxin in Mango and Orange Fruits, Juices, Pulps, and Jams Marketed in Pakistan. Toxins. 2020;12:52. doi: 10.3390/toxins12010052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Moss M.O. Risk assessment for aflatoxins in foodstuffs. Int. Biodeterior. Biodegrad. 2002;50:137–142. doi: 10.1016/S0964-8305(02)00078-1. [DOI] [Google Scholar]
- 13.Rushing B.R., Selim M.I. Aflatoxin B1: A review on metabolism, toxicity, occurrence in food, occupational exposure, and detoxification methods. Food Chem. Toxicol. 2019;124:81–100. doi: 10.1016/j.fct.2018.11.047. [DOI] [PubMed] [Google Scholar]
- 14.Rodríguez-Carrasco Y., Mañes J., Berrada H., Juan C. Development and validation of a LC-ESI-MS/MS method for the determination of alternaria toxins alternariol, alternariol methyl-ether and tentoxin in tomato and tomato-based products. Toxins. 2016;8:328. doi: 10.3390/toxins8110328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gotthardt M., Kanawati B., Schmidt F., Asam S., Hammerl R., Frank O., Hofmann T., Schmitt-Kopplin P., Rychlik M. Comprehensive Analysis of the Alternaria Mycobolome Using Mass Spectrometry Based Metabolomics. Mol. Nutr. Food Res. 2020;64:1900558. doi: 10.1002/mnfr.201900558. [DOI] [PubMed] [Google Scholar]
- 16.De Angelis E., Monaci L., Mackie A., Salt L., Visconti A. Reprint of “bioaccessibility of T-2 and HT-2 toxins in mycotoxin contaminated bread models submitted to in vitro human digestion”. Innov. Food Sci. Emerg. Technol. 2013;25:88–96. doi: 10.1016/j.ifset.2014.07.009. [DOI] [Google Scholar]
- 17.Ling A., Sun L., Guo W., Sun S., Yang J., Zhao Z. Individual and combined cytotoxic effects of T-2 toxin and its four metabolites on porcine Leydig cells. Food Chem. Toxicol. 2020;139:111277. doi: 10.1016/j.fct.2020.111277. [DOI] [PubMed] [Google Scholar]
- 18.Huang Z., Wang Y., Qiu M., Sun L., Liao J., Wang R., Sun X., Bi S., Gooneratne R. Effect of T-2 toxin-injected shrimp muscle extracts on mouse macrophage cells (RAW264.7) Drug Chem. Toxicol. 2018;41:16–21. doi: 10.1080/01480545.2016.1278227. [DOI] [PubMed] [Google Scholar]
- 19.Kang R., Perveen A., Li C. Effects of maternal T-2 toxin exposure on the hepatic glycolipid metabolism in young mice. Ecotoxicol. Environ. Saf. 2020;196:110530. doi: 10.1016/j.ecoenv.2020.110530. [DOI] [PubMed] [Google Scholar]
- 20.Kasimir M., Behrens M., Schulz M., Kuchenbuch H., Focke C., Humpf H.-U. Intestinal Metabolism of α- and β-Glucosylated Modified Mycotoxins T-2 and HT-2 Toxin in the Pig Cecum Model. J. Agric. Food Chem. 2020 doi: 10.1021/acs.jafc.0c00576. [DOI] [PubMed] [Google Scholar]
- 21.Schuhmacher-Wolz U., Heine K., Schneider K. Report on toxicity data on trichothecene mycotoxins HT-2 and T-2 toxins. EFSA Support. Publ. 2010;7 doi: 10.2903/sp.efsa.2010.EN-65. [DOI] [Google Scholar]
- 22.Zhou H., George S., Hay C., Lee J., Qian H., Sun X. Individual and combined effects of Aflatoxin B1, Deoxynivalenol and Zearalenone on HepG2 and RAW 264.7 cell lines. Food Chem. Toxicol. 2017;103:18–27. doi: 10.1016/j.fct.2017.02.017. [DOI] [PubMed] [Google Scholar]
- 23.Fernández-Blanco C., Font G., Ruiz M.J. Interaction effects of enniatin B, deoxinivalenol and alternariol in Caco-2 cells. Toxicol. Lett. 2016;241:38–48. doi: 10.1016/j.toxlet.2015.11.005. [DOI] [PubMed] [Google Scholar]
- 24.García G.R., Payros D., Pinton P., Dogi C.A., Laffitte J., Neves M., González Pereyra M.L., Cavaglieri L.R., Oswald I.P. Intestinal toxicity of deoxynivalenol is limited by Lactobacillus rhamnosus RC007 in pig jejunum explants. Arch. Toxicol. 2018;92:983–993. doi: 10.1007/s00204-017-2083-x. [DOI] [PubMed] [Google Scholar]
- 25.Pestka J.J. Mechanisms of deoxynivalenol-induced gene expression and apoptosis. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2008;25:1128–1140. doi: 10.1080/02652030802056626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Pestka J.J., Smolinski A.T. Deoxynivalenol: Toxicology and potential effects on humans. J. Toxicol. Environ. Heal. Part B Crit. Rev. 2005;8:39–69. doi: 10.1080/10937400590889458. [DOI] [PubMed] [Google Scholar]
- 27.Wang H.W., Wang J.Q., Zheng B.Q., Li S.L., Zhang Y.D., Li F.D., Zheng N. Cytotoxicity induced by ochratoxin A, zearalenone, and α-zearalenol: Effects of individual and combined treatment. Food Chem. Toxicol. 2014;71:217–224. doi: 10.1016/j.fct.2014.05.032. [DOI] [PubMed] [Google Scholar]
- 28.Yang D., Jiang X., Sun J., Li X., Li X., Jiao R., Peng Z., Li Y., Bai W. Toxic effects of zearalenone on gametogenesis and embryonic development: A molecular point of review. Food Chem. Toxicol. 2018;119:24–30. doi: 10.1016/j.fct.2018.06.003. [DOI] [PubMed] [Google Scholar]
- 29.Zinedine A., Soriano J.M., Moltó J.C., Mañes J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007;45:1–18. doi: 10.1016/j.fct.2006.07.030. [DOI] [PubMed] [Google Scholar]
- 30.Ruiz M.J., Macáková P., Juan-García A., Font G. Cytotoxic effects of mycotoxin combinations in mammalian kidney cells. Food Chem. Toxicol. 2011;49:2718–2724. doi: 10.1016/j.fct.2011.07.021. [DOI] [PubMed] [Google Scholar]
- 31.Mallebrera B., Prosperini A., Font G., Ruiz M.J. In vitro mechanisms of Beauvericin toxicity: A review. Food Chem. Toxicol. 2018;111:537–545. doi: 10.1016/j.fct.2017.11.019. [DOI] [PubMed] [Google Scholar]
- 32.Maranghi F., Tassinari R., Narciso L., Tait S., Rocca C.L., Felice G.D., Butteroni C., Corinti S., Barletta B., Cordelli E., et al. In vivo toxicity and genotoxicity of beauvericin and enniatins. Combined approach to study in vivo toxicity and genotoxicity of mycotoxins beauvericin (BEA) and enniatin B (ENNB) EFSA Support. Publ. 2018;15:1406E. doi: 10.2903/sp.efsa.2018.EN-1406. [DOI] [Google Scholar]
- 33.Prosperini A., Berrada H., Ruiz M.J., Caloni F., Coccini T., Spicer L.J., Perego M.C., Lafranconi A. A Review of the Mycotoxin Enniatin B. Front. Public Health. 2017;5:1–11. doi: 10.3389/fpubh.2017.00304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Wen J., Mu P., Deng Y. Mycotoxins: Cytotoxicity and biotransformation in animal cells. Toxicol. Res. 2016;5:377–387. doi: 10.1039/C5TX00293A. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Fernández-Blanco C., Juan-García A., Juan C., Font G., Ruiz M.J. Alternariol induce toxicity via cell death and mitochondrial damage on Caco-2 cells. Food Chem. Toxicol. 2016;88:32–39. doi: 10.1016/j.fct.2015.11.022. [DOI] [PubMed] [Google Scholar]
- 36.Solhaug A., Eriksen G.S., Holme J.A. Mechanisms of Action and Toxicity of the Mycotoxin Alternariol: A Review. Basic Clin. Pharmacol. Toxicol. 2016;119:533–539. doi: 10.1111/bcpt.12635. [DOI] [PubMed] [Google Scholar]
- 37.Nawaz S., Scudamore K.A., Rainbird S.C. Mycotoxins in ingredients of animal feeding stuffs: I. determination of Alternaria mycotoxins in oilseed rape meal and sunflower seed meal. Food Addit. Contam. 1997;14:249–262. doi: 10.1080/02652039709374522. [DOI] [PubMed] [Google Scholar]
- 38.Aichinger G., Puntscher H., Beisl J., Kütt M.L., Warth B., Marko D. Delphinidin protects colon carcinoma cells against the genotoxic effects of the mycotoxin altertoxin II. Toxicol. Lett. 2018;284:136–142. doi: 10.1016/j.toxlet.2017.12.002. [DOI] [PubMed] [Google Scholar]
- 39.da Motta S., Valente Soares L.M. Survey of Brazilian tomato products for alternariol, alternariol monomethyl ether, tenuazonic acid and cyclopiazonic acid. Food Addit. Contam. 2001;18:630–634. doi: 10.1080/02652030117707. [DOI] [PubMed] [Google Scholar]
- 40.Gruber-Dorninger C., Novak B., Nagl V., Berthiller F. Emerging Mycotoxins: Beyond Traditionally Determined Food Contaminants. J. Agric. Food Chem. 2017;65:7052–7070. doi: 10.1021/acs.jafc.6b03413. [DOI] [PubMed] [Google Scholar]
- 41.Ge N., Xu J., Peng B., Pan S. Adsorption mechanism of tenuazonic acid using inactivated lactic acid bacteria. Food Control. 2017;82:274–282. doi: 10.1016/j.foodcont.2017.07.009. [DOI] [Google Scholar]
- 42.Kumari A., Tirkey N.N. Recent Trends in Human and Animal Mycology. Springer; Singapore: 2019. Tenuazonic Acid: A Potent Mycotoxin; pp. 203–211. [Google Scholar]
- 43.Di Gregorio M.C., Bordin K., de Castro Souto P.C.M., Corassin C.H., Oliveira C.A.F. Comparative biotransformation of aflatoxin B 1 in swine, domestic fowls, and humans. Toxin Rev. 2015;34:142–150. doi: 10.3109/15569543.2015.1091979. [DOI] [Google Scholar]
- 44.Wu T.Y., Fridley B.L., Jenkins G.D., Batzler A., Wang L., Weinshilboum R.M. Mycophenolic acid response biomarkers: A cell line model system-based genome-wide screen. Int. Immunopharmacol. 2011;11:1057–1064. doi: 10.1016/j.intimp.2011.02.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Marin S., Ramos A.J., Cano-Sancho G., Sanchis V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013;60:218–237. doi: 10.1016/j.fct.2013.07.047. [DOI] [PubMed] [Google Scholar]
- 46.Fraeyman S., Croubels S., Devreese M., Antonissen G. Emerging fusarium and alternaria mycotoxins: Occurrence, toxicity and toxicokinetics. Toxins. 2017;9:228. doi: 10.3390/toxins9070228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.González-Arias C.A., Crespo-Sempere A., Marín S., Sanchis V., Ramos A.J. Modulation of the xenobiotic transformation system and inflammatory response by ochratoxin A exposure using a co-culture system of Caco-2 and HepG2 cells. Food Chem. Toxicol. 2015;86:245–252. doi: 10.1016/j.fct.2015.10.007. [DOI] [PubMed] [Google Scholar]
- 48.Bellí N., Marín S., Sanchis V., Ramos A.J. Ochratoxin A (OTA) in Wines, Musts and Grape Juices: Occurrence, Regulations and Methods of Analysis. Food Sci. Technol. Int. 2002;8:325–335. doi: 10.1177/108201302129174892. [DOI] [Google Scholar]
- 49.Fernández-Cruz M.L., Mansilla M.L., Tadeo J.L. Mycotoxins in fruits and their processed products: Analysis, occurrence and health implications. J. Adv. Res. 2010;1:113–122. doi: 10.1016/j.jare.2010.03.002. [DOI] [Google Scholar]
- 50.Orlando B., Grignon G., Vitry C., Kashefifard K., Valade R. Fusarium species and enniatin mycotoxins in wheat, durum wheat, triticale and barley harvested in France. Mycotoxin Res. 2019;35:369–380. doi: 10.1007/s12550-019-00363-x. [DOI] [PubMed] [Google Scholar]
- 51.Tibola C.S., de Miranda M.Z., Paiva F.F., Fernandes J.M.C., Guarienti E.M., Nicolau M. Effect of breadmaking process on mycotoxin content in white and whole wheat breads. Cereal Chem. 2018;95:660–665. doi: 10.1002/cche.10079. [DOI] [Google Scholar]
- 52.Mousavi Khaneghah A., Fakhri Y., Sant’Ana A.S. Impact of unit operations during processing of cereal-based products on the levels of deoxynivalenol, total aflatoxin, ochratoxin A, and zearalenone: A systematic review and meta-analysis. Food Chem. 2018;268:611–624. doi: 10.1016/j.foodchem.2018.06.072. [DOI] [PubMed] [Google Scholar]
- 53.Pascari X., Maul R., Kemmlein S., Marin S., Sanchis V. The fate of several trichothecenes and zearalenone during roasting and enzymatic treatment of cereal flour applied in cereal-based infant food production. Food Control. 2020;114:107245. doi: 10.1016/j.foodcont.2020.107245. [DOI] [Google Scholar]
- 54.Generotti S., Cirlini M., Šarkanj B., Sulyok M., Berthiller F., Dall’Asta C., Suman M. Formulation and processing factors affecting trichothecene mycotoxins within industrial biscuit-making. Food Chem. 2017;229:597–603. doi: 10.1016/j.foodchem.2017.02.115. [DOI] [PubMed] [Google Scholar]
- 55.Kuchenbuch H.S., Becker S., Schulz M., Cramer B., Humpf H.U. Thermal stability of t-2 and ht-2 toxins during biscuit-and crunchy muesli-making and roasting. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2018;35:2158–2167. doi: 10.1080/19440049.2018.1530456. [DOI] [PubMed] [Google Scholar]
- 56.Serrano A.B., Font G., Mañes J., Ferrer E. Effects of technological processes on enniatin levels in pasta. J. Sci. Food Agric. 2016;96:1756–1763. doi: 10.1002/jsfa.7282. [DOI] [PubMed] [Google Scholar]
- 57.Tolosa J., Font G., Mañes J., Ferrer E. Mitigation of enniatins in edible fish tissues by thermal processes and identification of degradation products. Food Chem. Toxicol. 2017;101:67–74. doi: 10.1016/j.fct.2016.12.039. [DOI] [PubMed] [Google Scholar]
- 58.Estiarte N., Crespo-Sempere A., Marín S., Ramos A.J., Worobo R.W. Stability of alternariol and alternariol monomethyl ether during food processing of tomato products. Food Chem. 2018;245:951–957. doi: 10.1016/j.foodchem.2017.11.078. [DOI] [PubMed] [Google Scholar]
- 59.Stadler D., Berthiller F., Suman M., Schuhmacher R., Krska R. Novel analytical methods to study the fate of mycotoxins during thermal food processing. Anal. Bioanal. Chem. 2020;412:9–16. doi: 10.1007/s00216-019-02101-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Schaarschmidt S., Fauhl-Hassek C. Mycotoxins during the processes of nixtamalization and tortilla production. Toxins. 2019;11:227. doi: 10.3390/toxins11040227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Morcia C., Tumino G., Ghizzoni R., Badeck F.W., Lattanzio V.M.T., Pascale M., Terzi V. Occurrence of Fusarium langsethiae and T-2 and HT-2 toxins in Italian malting barley. Toxins. 2016;8:247. doi: 10.3390/toxins8080247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Agriopoulou S., Stamatelopoulou E., Varzakas T. Control Strategies: Prevention and Detoxification in Foods. Foods. 2020;9:137. doi: 10.3390/foods9020137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Abrunhosa L., Morales H., Soares C., Calado T., Vila-Chã A.S., Pereira M., Venâncio A. A Review of Mycotoxins in Food and Feed Products in Portugal and Estimation of Probable Daily Intakes. Crit. Rev. Food Sci. Nutr. 2016;56:249–265. doi: 10.1080/10408398.2012.720619. [DOI] [PubMed] [Google Scholar]
- 64.Ayelign A., De Saeger S. Mycotoxins in Ethiopia: Current status, implications to food safety and mitigation strategies. Food Control. 2020;113:107163. doi: 10.1016/j.foodcont.2020.107163. [DOI] [Google Scholar]
- 65.Balendres M.A.O., Karlovsky P., Cumagun C.J.R. Mycotoxigenic fungi and mycotoxins in agricultural crop commodities in the Philippines: A review. Foods. 2019;8:249. doi: 10.3390/foods8070249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Puntscher H., Cobankovic I., Marko D., Warth B. Quantitation of free and modified Alternaria mycotoxins in European food products by LC-MS/MS. Food Control. 2019;102:157–165. doi: 10.1016/j.foodcont.2019.03.019. [DOI] [Google Scholar]
- 67.Meca G., Mañes J., Font G., Ruiz M.J. Study of the potential toxicity of enniatins A, A 1, B, B 1 by evaluation of duodenal and colonic bioavailability applying an invitro method by Caco-2 cells. Toxicon. 2012;59:1–11. doi: 10.1016/j.toxicon.2011.10.004. [DOI] [PubMed] [Google Scholar]
- 68.Fernández-García E., Carvajal-Lérida I., Pérez-Gálvez A. In vitro bioaccessibility assessment as a prediction tool of nutritional efficiency. Nutr. Res. 2009;29:751–760. doi: 10.1016/j.nutres.2009.09.016. [DOI] [PubMed] [Google Scholar]
- 69.González-Arias C.A., Marín S., Sanchis V., Ramos A.J. Mycotoxin bioaccessibility/absorption assessment using in vitro digestion models: A review. World Mycotoxin J. 2013;6:167–184. doi: 10.3920/WMJ2012.1521. [DOI] [Google Scholar]
- 70.Brandon E.F.A., Oomen A.G., Rompelberg C.J.M., Versantvoort C.H.M., Van Engelen J.G.M., Sips A.J.A.M. Consumer product in vitro digestion model: Bioaccessibility of contaminants and its application in risk assessment. Regul. Toxicol. Pharmacol. 2006;44:161–171. doi: 10.1016/j.yrtph.2005.10.002. [DOI] [PubMed] [Google Scholar]
- 71.Tran V.N., Viktorova J., Augustynkova K., Jelenova N., Dobiasova S., Rehorova K., Fenclova M., Stranska-Zachariasova M., Vitek L., Hajslova J., et al. In silico and in vitro studies of mycotoxins and their cocktails; Their toxicity and its mitigation by silibinin pre-treatment. Toxins. 2020;12:148. doi: 10.3390/toxins12030148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Bordin K., Saladino F., Fernández-Blanco C., Ruiz M.J., Mañes J., Fernández-Franzón M., Meca G., Luciano F.B. Reaction of zearalenone and α-zearalenol with allyl isothiocyanate, characterization of reaction products, their bioaccessibility and bioavailability in vitro. Food Chem. 2017;217:648–654. doi: 10.1016/j.foodchem.2016.09.044. [DOI] [PubMed] [Google Scholar]
- 73.Kabak B., Brandon E.F.A., Var I., Blokland M., Sips A.J.A.M. Effects of probiotic bacteria on the bioaccessibility of aflatoxin B1 and ochratoxin A using an in vitro digestion model under fed conditions. J. Environ. Sci. Health Part B Pestic. Food Contam. Agric. Wastes. 2009;44:472–480. doi: 10.1080/03601230902935154. [DOI] [PubMed] [Google Scholar]
- 74.CenciČ A., Langerholc T. Functional cell models of the gut and their applications in food microbiology—A review. Int. J. Food Microbiol. 2010;141:S4–S14. doi: 10.1016/j.ijfoodmicro.2010.03.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Artursson P., Karlsson J. Correlation between oral drug absorption in humans and apparent drug permeability coefficients in human intestinal epithelial (Caco-2) cells. Biochem. Biophys. Res. Commun. 1991;175:880–885. doi: 10.1016/0006-291X(91)91647-U. [DOI] [PubMed] [Google Scholar]
- 76.Brandon E.F.A., Bosch T.M., Deenen M.J., Levink R., van der Wal E., van Meerveld J.B.M., Bijl M., Beijnen J.H., Schellens J.H.M., Meijerman I. Validation of in vitro cell models used in drug metabolism and transport studies; genotyping of cytochrome P450, phase II enzymes and drug transporter polymorphisms in the human hepatoma (HepG2), ovarian carcinoma (IGROV-1) and colon carcinoma (CaCo-2, LS180) cell lines. Toxicol. Appl. Pharmacol. 2006;211:1–10. doi: 10.1016/j.taap.2005.05.004. [DOI] [PubMed] [Google Scholar]
- 77.Lněničková K., Šadibolová M., Matoušková P., Szotáková B., Skálová L., Boušová I. The Modulation of Phase II Drug-Metabolizing Enzymes in Proliferating and Differentiated CaCo-2 Cells by Hop-Derived Prenylflavonoids. Nutrients. 2020;12:2138. doi: 10.3390/nu12072138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Schaut A., De Saeger S., Sergent T., Schneider Y.-J., Larondelle Y., Pussemier L., Van Peteghem C. Study of the gastrointestinal biotransformation of zearalenone in a Caco-2 cell culture system with liquid chromatographic methods. J. Appl. Toxicol. 2008;28:966–973. doi: 10.1002/jat.1362. [DOI] [PubMed] [Google Scholar]
- 79.Cizkova K. Expression of cytochrome P450 epoxygenases and soluble epoxide hydrolase is regulated by hypolipidemic drugs in dose-dependent manner. Toxicol. Appl. Pharmacol. 2018;355:156–163. doi: 10.1016/j.taap.2018.06.025. [DOI] [PubMed] [Google Scholar]
- 80.Šemeláková M., Jendželovský R., Fedoročko P. Drug membrane transporters and CYP3A4 are affected by hypericin, hyperforin or aristoforin in colon adenocarcinoma cells. Biomed. Pharmacother. 2016;81:38–47. doi: 10.1016/j.biopha.2016.03.045. [DOI] [PubMed] [Google Scholar]
- 81.Odenthal J., van Heumen B.W.H., Roelofs H.M.J., te Morsche R.H.M., Marian B., Nagengast F.M., Peters W.H.M. The Influence of Curcumin, Quercetin, and Eicosapentaenoic Acid on the Expression of Phase II Detoxification Enzymes in the Intestinal Cell Lines HT-29, Caco-2, HuTu 80, and LT97. Nutr. Cancer. 2012;64:856–863. doi: 10.1080/01635581.2012.700994. [DOI] [PubMed] [Google Scholar]
- 82.Langerholc T., Maragkoudakis P.A., Wollgast J., Gradisnik L., Cencic A. Novel and established intestinal cell line models—An indispensable tool in food science and nutrition. Trends Food Sci. Technol. 2011;22:S11–S20. doi: 10.1016/j.tifs.2011.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Haslam I.S., Jones K., Coleman T., Simmons N.L. Induction of P-glycoprotein expression and function in human intestinal epithelial cells (T84) Biochem. Pharmacol. 2008;76:850–861. doi: 10.1016/j.bcp.2008.07.020. [DOI] [PubMed] [Google Scholar]
- 84.Naruhashi K., Kurahashi Y., Fujita Y., Kawakita E., Yamasaki Y., Hattori K., Nishimura A., Shibata N. Comparison of the Expression and Function of ATP Binding Cassette Transporters in Caco-2 and T84 cells on Stimulation by Selected Endogenous Compounds and Xenobiotics. Drug Metab. Pharmacokinet. 2011;26:145–153. doi: 10.2133/dmpk.DMPK-10-RG-075. [DOI] [PubMed] [Google Scholar]
- 85.Theodoropoulos C., Demers C., Delvin E., Menard D., Gascon-Barre M. Calcitriol regulates the expression of the genes encoding the three key vitamin D3 hydroxylases and the drug-metabolizing enzyme CYP3A4 in the human fetal intestine. Clin. Endocrinol. 2003;58:489–499. doi: 10.1046/j.1365-2265.2003.01743.x. [DOI] [PubMed] [Google Scholar]
- 86.Saaby L., Helms H.C.C., Brodin B. IPEC-J2 MDR1, a Novel High-Resistance Cell Line with Functional Expression of Human P-glycoprotein (ABCB1) for Drug Screening Studies. Mol. Pharm. 2016;13:640–652. doi: 10.1021/acs.molpharmaceut.5b00874. [DOI] [PubMed] [Google Scholar]
- 87.Palócz O., Szita G., Csikó G. Alteration in Inflammatory Responses and Cytochrome P450 Expression of Porcine Jejunal Cells by Drinking Water Supplements. Mediators Inflamm. 2019;2019 doi: 10.1155/2019/5420381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Loi M., Fanelli F., Liuzzi V.C., Logrieco A.F., Mulè G. Mycotoxin biotransformation by native and commercial enzymes: Present and future perspectives. Toxins. 2017;9:111. doi: 10.3390/toxins9040111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Beyerle J., Frei E., Stiborova M., Habermann N., Ulrich C.M. Biotransformation of xenobiotics in the human colon and rectum and its association with colorectal cancer. Drug Metab. Rev. 2015;47:199–221. doi: 10.3109/03602532.2014.996649. [DOI] [PubMed] [Google Scholar]
- 90.Gajecka M., Jakimiuk E., Zielonka L., Obremski K., Gajecka M. The Biotransformation of Chosen Mycotoxins. Pol. J. Vet Sci. 2009;12:293–303. [PubMed] [Google Scholar]
- 91.Galtier P. Biotransformation and Fate of Mycotoxins. J. Toxicol. Toxin Rev. 1999;18:295–312. [Google Scholar]
- 92.Sergent T., Ribonnet L., Kolosova A., Garsou S., Schaut A., De Saeger S., Van Peteghem C., Larondelle Y., Pussemier L., Schneider Y.J. Molecular and cellular effects of food contaminants and secondary plant components and their plausible interactions at the intestinal level. Food Chem. Toxicol. 2008;46:813–841. doi: 10.1016/j.fct.2007.12.006. [DOI] [PubMed] [Google Scholar]
- 93.Lin N.N., Chen J., Xu B., Wei X., Guo L., Xie J.W. The roles of carboxylesterase and CYP isozymes on the in vitro metabolism of T-2 toxin. Mil. Med. Res. 2015;2:13. doi: 10.1186/s40779-015-0041-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Wang J., Jiang J., Zhang H., Wang J., Cai H., Li C., Li K., Liu J., Guo X., Zou G., et al. Integrated transcriptional and proteomic analysis with in vitro biochemical assay reveal the important role of CYP3A46 in T-2 toxin hydroxylation in porcine primary hepatocytes. Mol. Cell. Proteom. 2011;10 doi: 10.1074/mcp.M111.008748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Wu Q., Huang L., Liu Z., Yao M., Wang Y., Dai M., Yuan Z. A comparison of hepatic in vitro metabolism of T-2 toxin in rats, pigs, chickens, and carp. Xenobiotica. 2011;41:863–873. doi: 10.3109/00498254.2011.593206. [DOI] [PubMed] [Google Scholar]
- 96.Ge X., Wang J., Liu J., Jiang J., Lin H., Wu J., Ouyang M., Tang X., Zheng M., Liao M., et al. The catalytic activity of cytochrome P450 3A22 is critical for the metabolism of T-2 toxin in porcine reservoirs. Catal. Commun. 2010;12:71–75. doi: 10.1016/j.catcom.2010.08.003. [DOI] [Google Scholar]
- 97.Yuan Y., Zhou X., Yang J., Li M., Qiu X. T-2 toxin is hydroxylated by chicken CYP3A37. Food Chem. Toxicol. 2013;62:622–627. doi: 10.1016/j.fct.2013.09.031. [DOI] [PubMed] [Google Scholar]
- 98.Shang S., Jiang J., Deng Y. Chicken cytochrome P450 1A5 is the key enzyme for metabolizing T-2 toxin to 3′OH-T-2. Int. J. Mol. Sci. 2013;14:10809–10818. doi: 10.3390/ijms140610809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Dai D., Pan Y., Zeng C.P., Liu S., Yan Y., Wu X., Xu Z., Zhang L. Activated FXR promotes xenobiotic metabolism of T-2 toxin and attenuates oxidative stress in broiler chicken liver. Chem. Biol. Interact. 2020;316:108912. doi: 10.1016/j.cbi.2019.108912. [DOI] [PubMed] [Google Scholar]
- 100.Deng Y., Wang Y., Sun L., Lu P., Wang R., Ye L., Xu D., Ye R., Liu Y., Bi S., et al. Biotransformation enzyme activities and phase I metabolites analysis in Litopenaeus vannamei following intramuscular administration of T-2 toxin. Drug Chem. Toxicol. 2018;41:113–122. doi: 10.1080/01480545.2017.1320407. [DOI] [PubMed] [Google Scholar]
- 101.Wang Y., Wang G., Dai Y., Wang Y., Lee Y.W., Shi J., Xu J. Biodegradation of Deoxynivalenol by a Novel Microbial Consortium. Front. Microbiol. 2020;10:2964. doi: 10.3389/fmicb.2019.02964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Wu Q.H., Wang X., Yang W., Nüssler A.K., Xiong L.Y., Kuča K., Dohnal V., Zhang X.J., Yuan Z.H. Oxidative stress-mediated cytotoxicity and metabolism of T-2 toxin and deoxynivalenol in animals and humans: An update. Arch. Toxicol. 2014;88:1309–1326. doi: 10.1007/s00204-014-1280-0. [DOI] [PubMed] [Google Scholar]
- 103.Catteuw A., Broekaert N., De Baere S., Lauwers M., Gasthuys E., Huybrechts B., Callebaut A., Ivanova L., Uhlig S., De Boevre M., et al. Insights into in Vivo Absolute Oral Bioavailability, Biotransformation, and Toxicokinetics of Zearalenone, α-Zearalenol, β-Zearalenol, Zearalenone-14-glucoside, and Zearalenone-14-sulfate in Pigs. J. Agric. Food Chem. 2019;67:3448–3458. doi: 10.1021/acs.jafc.8b05838. [DOI] [PubMed] [Google Scholar]
- 104.Malekinejad H., Maas-Bakker R., Fink-Gremmels J. Species differences in the hepatic biotransformation of zearalenone. Vet. J. 2006;172:96–102. doi: 10.1016/j.tvjl.2005.03.004. [DOI] [PubMed] [Google Scholar]
- 105.Malekinejad H., Maas-Bakker R.F., Fink-Gremmels J. Bioactivation of zearalenone by porcine hepatic biotransformation. Vet. Res. 2005;36:799–810. doi: 10.1051/vetres:2005034. [DOI] [PubMed] [Google Scholar]
- 106.Ayed-Boussema I., Pascussi J.M., Maurel P., Bacha H., Hassen W. Zearalenone activates pregnane X receptor, constitutive androstane receptor and aryl hydrocarbon receptor and corresponding phase I target genes mRNA in primary cultures of human hepatocytes. Environ. Toxicol. Pharmacol. 2011;31:79–87. doi: 10.1016/j.etap.2010.09.008. [DOI] [PubMed] [Google Scholar]
- 107.Rodríguez-Carrasco Y., Heilos D., Richter L., Süssmuth R.D., Heffeter P., Sulyok M., Kenner L., Berger W., Dornetshuber-Fleiss R. Mouse tissue distribution and persistence of the food-born fusariotoxins Enniatin B and Beauvericin. Toxicol. Lett. 2016;247:35–44. doi: 10.1016/j.toxlet.2016.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Mei L., Zhang L., Dai R. An inhibition study of beauvericin on human and rat cytochrome P450 enzymes and its pharmacokinetics in rats. J. Enzym. Inhib. Med. Chem. 2009;24:753–762. doi: 10.1080/14756360802362041. [DOI] [PubMed] [Google Scholar]
- 109.Fæste C.K., Ivanova L., Uhlig S. In vitro metabolism of the mycotoxin enniatin B in different species and cytochrome P450 enzyme phenotyping by chemical inhibitors. Drug Metab. Dispos. 2011;39:1768–1776. doi: 10.1124/dmd.111.039529. [DOI] [PubMed] [Google Scholar]
- 110.Ivanova L., Fæste C.K., Uhlig S. In vitro phase i metabolism of the depsipeptide enniatin B. Anal. Bioanal. Chem. 2011;400:2889–2901. doi: 10.1007/s00216-011-4964-9. [DOI] [PubMed] [Google Scholar]
- 111.Ivanova L., Fæste C.K., Van Pamel E., Daeseleire E., Callebaut A., Uhlig S. Presence of enniatin B and its hepatic metabolites in plasma and liver samples from broilers and eggs from laying hens. World Mycotoxin J. 2014;7:167–175. doi: 10.3920/WMJ2013.1609. [DOI] [Google Scholar]
- 112.Ivanova L., Denisov I.G., Grinkova Y.V., Sligar S.G., Fæste C.K. Biotransformation of the Mycotoxin Enniatin B1 by CYP P450 3A4 and Potential for Drug-Drug Interactions. Metabolites. 2019;9:158. doi: 10.3390/metabo9080158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Ivanova L., Uhlig S., Devreese M., Croubels S., Fæste C.K. Biotransformation of the mycotoxin enniatin B1 in pigs: A comparative in vitro and in vivo approach. Food Chem. Toxicol. 2017;105:506–517. doi: 10.1016/j.fct.2017.04.041. [DOI] [PubMed] [Google Scholar]
- 114.Debevere S., Cools A., De Baere S., Haesaert G., Rychlik M., Croubels S., Fievez V. In Vitro Rumen Simulations Show a Reduced Disappearance of Deoxynivalenol, Nivalenol and Enniatin B at Conditions of Rumen Acidosis and Lower Microbial Activity. Toxins. 2020;12:101. doi: 10.3390/toxins12020101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Fraeyman S., Devreese M., Antonissen G., De Baere S., Rychlik M., Croubels S. Comparative Oral Bioavailability, Toxicokinetics, and Biotransformation of Enniatin B1 and Enniatin B in Broiler Chickens. J. Agric. Food Chem. 2016;64:7259–7264. doi: 10.1021/acs.jafc.6b02913. [DOI] [PubMed] [Google Scholar]
- 116.Müller S., Dekant W., Mally A. Fumonisin B 1 and the kidney: Modes of action for renal tumor formation by fumonisin B 1 in rodents. Food Chem. Toxicol. 2012;50:3833–3846. doi: 10.1016/j.fct.2012.06.053. [DOI] [PubMed] [Google Scholar]
- 117.Schreck I., Deigendesch U., Burkhardt B., Marko D., Weiss C. The Alternaria mycotoxins alternariol and alternariol methyl ether induce cytochrome P450 1A1 and apoptosis in murine hepatoma cells dependent on the aryl hydrocarbon receptor. Arch. Toxicol. 2012;86:625–632. doi: 10.1007/s00204-011-0781-3. [DOI] [PubMed] [Google Scholar]
- 118.Aichinger G., Krüger F., Puntscher H., Preindl K., Warth B., Marko D. Naturally occurring mixtures of Alternaria toxins: Anti-estrogenic and genotoxic effects in vitro. Arch. Toxicol. 2019;93:3021–3031. doi: 10.1007/s00204-019-02545-z. [DOI] [PubMed] [Google Scholar]
- 119.Burkhardt B., Pfeiffer E., Metzler M. Absorption and metabolism of the mycotoxins alternariol and alternariol-9-methyl ether in Caco-2 cells in vitro. Mycotoxin Res. 2009;25:149–157. doi: 10.1007/s12550-009-0022-2. [DOI] [PubMed] [Google Scholar]
- 120.Scheibenzuber S., Hoffmann T., Effenberger I., Schwab W., Asam S., Rychlik M. Enzymatic synthesis of modified alternaria mycotoxins using a whole-cell biotransformation system. Toxins. 2020;12:264. doi: 10.3390/toxins12040264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Puntscher H., Hankele S., Tillmann K., Attakpah E., Braun D., Kütt M.L., Del Favero G., Aichinger G., Pahlke G., Höger H., et al. First insights into Alternaria multi-toxin in vivo metabolism. Toxicol. Lett. 2019;301:168–178. doi: 10.1016/j.toxlet.2018.10.006. [DOI] [PubMed] [Google Scholar]
- 122.Yang X.J., Lu H.Y., Li Z.Y., Bian Q., Qiu L.L., Li Z., Liu Q., Li J., Wang X., Wang S.L. Cytochrome P450 2A13 mediates aflatoxin B1-induced cytotoxicity and apoptosis in human bronchial epithelial cells. Toxicology. 2012;300:138–148. doi: 10.1016/j.tox.2012.06.010. [DOI] [PubMed] [Google Scholar]
- 123.Deng J., Zhao L., Zhang N.Y., Karrow N.A., Krumm C.S., Qi D.S., Sun L.H. Aflatoxin B 1 metabolism: Regulation by phase I and II metabolizing enzymes and chemoprotective agents. Mutat. Res. Rev. Mutat. Res. 2018;778:79–89. doi: 10.1016/j.mrrev.2018.10.002. [DOI] [PubMed] [Google Scholar]
- 124.Ayed-Boussema I., Pascussi J.M., Zaied C., Maurel P., Bacha H., Hassen W. Ochratoxin A induces CYP3A4, 2B6, 3A5, 2C9, 1A1, and CYP1A2 gene expression in primary cultured human hepatocytes: A possible activation of nuclear receptors. Drug Chem. Toxicol. 2012;35:71–80. doi: 10.3109/01480545.2011.589438. [DOI] [PubMed] [Google Scholar]
- 125.Shin H.S., Lee H.J., Pyo M.C., Ryu D., Lee K.W. Ochratoxin a-induced hepatotoxicity through phase i and phase ii reactions regulated by ahr in liver cells. Toxins. 2019;11:377. doi: 10.3390/toxins11070377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Tao Y., Xie S., Xu F., Liu A., Wang Y., Chen D., Pan Y., Huang L., Peng D., Wang X., et al. Ochratoxin A: Toxicity, oxidative stress and metabolism. Food Chem. Toxicol. 2018;112:320–331. doi: 10.1016/j.fct.2018.01.002. [DOI] [PubMed] [Google Scholar]
- 127.Gross-Steinmeyer K., Weymann J., Hege H.G., Metzler M. Metabolism and lack of DNA reactivity of the mycotoxin ochratoxin A in cultured rat and human primary hepatocytes. J. Agric. Food Chem. 2002;50:938–945. doi: 10.1021/jf0111817. [DOI] [PubMed] [Google Scholar]
- 128.Tozlovanu M., Canadas D., Pfohl-Leszkowicz A., Frenette C., Paugh R.J., Manderville R.A. Glutathione conjugates of ochratoxin a as biomarkers of exposure. Arh. Hig. Rada Toksikol. 2012;63:417–425. doi: 10.2478/10004-1254-63-2012-2202. [DOI] [PubMed] [Google Scholar]
- 129.Kőszegi T., Poór M. Ochratoxin a: Molecular interactions, mechanisms of toxicity and prevention at the molecular level. Toxins. 2016;8:111. doi: 10.3390/toxins8040111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Ayed-Boussema I., Pascussi J.M., Rjiba K., Maurel P., Bacha H., Hassen W. The mycotoxin, patulin, increases the expression of PXR and AhR and their target cytochrome P450s in primary cultured human hepatocytes. Drug Chem. Toxicol. 2012;35:241–250. doi: 10.3109/01480545.2011.592194. [DOI] [PubMed] [Google Scholar]
- 131.Ji C., Fan Y., Zhao L. Review on biological degradation of mycotoxins. Anim. Nutr. 2016;2:127–133. doi: 10.1016/j.aninu.2016.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Young J.C., Zhou T., Yu H., Zhu H., Gong J. Degradation of trichothecene mycotoxins by chicken intestinal microbes. Food Chem. Toxicol. 2007;45:136–143. doi: 10.1016/j.fct.2006.07.028. [DOI] [PubMed] [Google Scholar]
- 133.Pierron A., Mimoun S., Murate L.S., Loiseau N., Lippi Y., Bracarense A.P.F.L., Schatzmayr G., He J.W., Zhou T., Moll W.D., et al. Microbial biotransformation of DON: Molecular basis for reduced toxicity. Sci. Rep. 2016;6:29105. doi: 10.1038/srep29105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Du K., Wang C., Liu P., Li Y., Ma X. Effects of Dietary Mycotoxins on Gut Microbiome. Protein Pept. Lett. 2017;24:397–405. doi: 10.2174/0929866524666170223095207. [DOI] [PubMed] [Google Scholar]
- 135.Antonissen G., Devreese M., De Baere S., Martel A., Van Immerseel F., Croubels S. Impact of Fusarium mycotoxins on hepatic and intestinal mRNA expression of cytochrome P450 enzymes and drug transporters, and on the pharmacokinetics of oral enrofloxacin in broiler chickens. Food Chem. Toxicol. 2017;101:75–83. doi: 10.1016/j.fct.2017.01.006. [DOI] [PubMed] [Google Scholar]
- 136.Murcia H.W., Diaz G.J. In vitro hepatic aflatoxicol production is related to a higher resistance to aflatoxin B1 in poultry. Sci. Rep. 2020;10:5508. doi: 10.1038/s41598-020-62415-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Gross-Steinmeyer K., Eaton D.L. Dietary modulation of the biotransformation and genotoxicity of aflatoxin B1. Toxicology. 2012;299:69–79. doi: 10.1016/j.tox.2012.05.016. [DOI] [PubMed] [Google Scholar]
- 138.Peles F., Sipos P., Győri Z., Pfliegler W.P., Giacometti F., Serraino A., Pagliuca G., Gazzotti T., Pócsi I. Adverse Effects, Transformation and Channeling of Aflatoxins Into Food Raw Materials in Livestock. Front. Microbiol. 2019;10:1–26. doi: 10.3389/fmicb.2019.02861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Wu J., Xu W., Zhang C., Chang Q., Tang X., Li K., Deng Y. Trp266 determines the binding specificity of a porcine aflatoxin B 1 aldehyde reductase for aflatoxin B1-dialdehyde. Biochem. Pharmacol. 2013;86:1357–1365. doi: 10.1016/j.bcp.2013.08.014. [DOI] [PubMed] [Google Scholar]
- 140.Lyagin I., Efremenko E. Enzymes for Detoxification of Various Mycotoxins: Origins and Mechanisms of Catalytic Action. Molecules. 2019;24:2362. doi: 10.3390/molecules24132362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Schrenk D., Bodin L., Chipman J.K., del Mazo J., Grasl-Kraupp B., Hogstrand C., Hoogenboom L. (Ron), Leblanc J., Nebbia C.S., Nielsen E., et al. Risk assessment of ochratoxin A in food. EFSA J. 2020;18 doi: 10.2903/j.efsa.2020.6113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Li P., Su R., Yin R., Lai D., Wang M., Liu Y., Zhou L. Detoxification of mycotoxins through biotransformation. Toxins. 2020;12:121. doi: 10.3390/toxins12020121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Ringot D., Chango A., Schneider Y.-J., Larondelle Y. Toxicokinetics and toxicodynamics of ochratoxin A, an update. Chem. Biol. Interact. 2006;159:18–46. doi: 10.1016/j.cbi.2005.10.106. [DOI] [PubMed] [Google Scholar]
- 144.Wu Q., Dohnal V., Huang L., Kuca K., Wang X., Chen G., Yuan Z. Metabolic Pathways of Ochratoxin A. Curr. Drug Metab. 2011;12:1–10. doi: 10.2174/138920011794520026. [DOI] [PubMed] [Google Scholar]
- 145.Schelstraete W., Devreese M., Croubels S. Impact of subacute exposure to T-2 toxin and zearalenone on the pharmacokinetics of midazolam as CYP3A probe drug in a porcine animal model: A pilot study. Front. Pharmacol. 2019;10:399. doi: 10.3389/fphar.2019.00399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Payros D., Alassane-Kpembi I., Pierron A., Loiseau N., Pinton P., Oswald I.P. Toxicology of deoxynivalenol and its acetylated and modified forms. Arch. Toxicol. 2016;90:2931–2957. doi: 10.1007/s00204-016-1826-4. [DOI] [PubMed] [Google Scholar]
- 147.Springler A., Hessenberger S., Reisinger N., Kern C., Nagl V., Schatzmayr G., Mayer E. Deoxynivalenol and its metabolite deepoxy-deoxynivalenol: Multi-parameter analysis for the evaluation of cytotoxicity and cellular effects. Mycotoxin Res. 2017;33:25–37. doi: 10.1007/s12550-016-0260-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Nagl V., Woechtl B., Schwartz-Zimmermann H.E., Hennig-Pauka I., Moll W.D., Adam G., Berthiller F. Metabolism of the masked mycotoxin deoxynivalenol-3-glucoside in pigs. Toxicol. Lett. 2014;229:190–197. doi: 10.1016/j.toxlet.2014.06.032. [DOI] [PubMed] [Google Scholar]
- 149.Schwartz H.E., Hametner C., Slavik V., Greitbauer O., Bichl G., Kunz-Vekiru E., Schatzmayr D., Berthiller F. Characterization of three deoxynivalenol sulfonates formed by reaction of deoxynivalenol with sulfur reagents. J. Agric. Food Chem. 2013;61:8941–8948. doi: 10.1021/jf403438b. [DOI] [PubMed] [Google Scholar]
- 150.Gerding J., Cramer B., Humpf H.U. Determination of mycotoxin exposure in Germany using an LC-MS/MS multibiomarker approach. Mol. Nutr. Food Res. 2014;58:2358–2368. doi: 10.1002/mnfr.201400406. [DOI] [PubMed] [Google Scholar]
- 151.Pestka J.J. Deoxynivalenol: Toxicity, mechanisms and animal health risks. Anim. Feed Sci. Technol. 2007;137:283–298. doi: 10.1016/j.anifeedsci.2007.06.006. [DOI] [Google Scholar]
- 152.Gao X., Mu P., Zhu X., Chen X., Tang S., Wu Y., Miao X., Wang X., Wen J., Deng Y. Dual function of a novel bacterium, slackia sp. D-G6: Detoxifying deoxynivalenol and producing the Natural Estrogen Analogue, Equol. Toxins. 2020;12:85. doi: 10.3390/toxins12020085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Mackei M., Orbán K., Molnár A., Pál L., Dublecz K., Husvéth F., Neogrády Z., Mátis G. Cellular Effects of T-2 Toxin on Primary Hepatic Cell Culture Models of Chickens. Toxins. 2020;12:46. doi: 10.3390/toxins12010046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Li Y., Wang Z., Beier R.C., Shen J., De Smet D., De Saeger S., Zhang S. T-2 toxin, a trichothecene mycotoxin: Review of toxicity, metabolism, and analytical methods. J. Agric. Food Chem. 2011;59:3441–3453. doi: 10.1021/jf200767q. [DOI] [PubMed] [Google Scholar]
- 155.Welsch T., Humpf H.U. HT-2 toxin 4-glucuronide as new T-2 toxin metabolite: Enzymatic synthesis, analysis, and species specific formation of T-2 and HT-2 toxin glucuronides by rat, mouse, pig, and human liver microsomes. J. Agric. Food Chem. 2012;60:10170–10178. doi: 10.1021/jf302571y. [DOI] [PubMed] [Google Scholar]
- 156.Masching S., Naehrer K., Schwartz-Zimmermann H.E., Sărăndan M., Schaumberger S., Dohnal I., Nagl V., Schatzmayr D. Gastrointestinal degradation of fumonisin B1 by carboxylesterase FumD prevents fumonisin induced alteration of sphingolipid metabolism in Turkey and swine. Toxins. 2016;8:84. doi: 10.3390/toxins8030084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Daud N., Currie V., Duncan G., Busman M., Gratz S.W. Intestinal hydrolysis and microbial biotransformation of diacetoxyscirpenol-α-glucoside, HT-2-β-glucoside and N-(1-deoxy-d-fructos-1-yl) fumonisin B1 by human gut microbiota in vitro. Int. J. Food Sci. Nutr. 2019;71:540–548. doi: 10.1080/09637486.2019.1698015. [DOI] [PubMed] [Google Scholar]
- 158.Merrill A.H., Morgan E.T., Nikolova-Karakashian M., Stewart J. Sphingomyelin hydrolysis and regulation of the expression of the gene for cytochrome P450. Biochem. Soc. Trans. 1999;27:383–387. doi: 10.1042/bst0270383. [DOI] [PubMed] [Google Scholar]
- 159.Spotti M., Maas R.F.M., De Nijs C.M., Fink-Gremmels J. Effect of fumonisin B1 on rat hepatic P450 system. Environ. Toxicol. Pharmacol. 2000;8:197–204. doi: 10.1016/S1382-6689(00)00040-5. [DOI] [PubMed] [Google Scholar]
- 160.Riley R.T., Merrill A.H. Ceramide synthase inhibition by fumonisins: A perfect storm of perturbed sphingolipid metabolism, signaling, and disease. J. Lipid Res. 2019;60:1183–1189. doi: 10.1194/jlr.S093815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Harrer H., Laviad E.L., Humpf H.U., Futerman A.H. Identification of N-acyl-fumonisin B1 as new cytotoxic metabolites of fumonisin mycotoxins. Mol. Nutr. Food Res. 2013;57:516–522. doi: 10.1002/mnfr.201200465. [DOI] [PubMed] [Google Scholar]
- 162.Dellafiora L., Galaverna G., Dall’Asta C. Mechanisms of Fumonisin B1 Toxicity: A Computational Perspective beyond the Ceramide Synthases Inhibition. Chem. Res. Toxicol. 2018;31:1203–1212. doi: 10.1021/acs.chemrestox.8b00188. [DOI] [PubMed] [Google Scholar]
- 163.Knutsen H.K., Alexander J., Barregård L., Bignami M., Brüschweiler B., Ceccatelli S., Cottrill B., Dinovi M., Edler L., Grasl-Kraupp B., et al. Risks for animal health related to the presence of fumonisins, their modified forms and hidden forms in feed. EFSA J. 2018;16:e05242. doi: 10.2903/j.efsa.2018.5242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Ieko T., Inoue S., Inomata Y., Inoue H., Fujiki J., Iwano H. Glucuronidation as a metabolic barrier against zearalenone in rat everted intestine. J. Vet. Med. Sci. 2020;82:153–161. doi: 10.1292/jvms.19-0570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Binder S.B., Schwartz-Zimmermann H.E., Varga E., Bichl G., Michlmayr H., Adam G., Berthiller F. Metabolism of zearalenone and its major modified forms in pigs. Toxins. 2017;9:56. doi: 10.3390/toxins9020056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Videmann B., Mazallon M., Tep J., Lecoeur S. Metabolism and transfer of the mycotoxin zearalenone in human intestinal Caco-2 cells. Food Chem. Toxicol. 2008;46:3279–3286. doi: 10.1016/j.fct.2008.07.011. [DOI] [PubMed] [Google Scholar]
- 167.Dellafiora L., Galaverna G., Righi F., Cozzini P., Dall’Asta C. Assessing the hydrolytic fate of the masked mycotoxin zearalenone-14-glucoside—A warning light for the need to look at the “maskedome”. Food Chem. Toxicol. 2017;99:9–16. doi: 10.1016/j.fct.2016.11.013. [DOI] [PubMed] [Google Scholar]
- 168.Keller L., Abrunhosa L., Keller K., Rosa C., Cavaglieri L., Venâncio A. Zearalenone and Its Derivatives α-Zearalenol and β-Zearalenol Decontamination by Saccharomyces cerevisiae Strains Isolated from Bovine Forage. Toxins. 2015;7:3297–3308. doi: 10.3390/toxins7083297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Rogowska A., Pomastowski P., Walczak J., Railean-Plugaru V., Rudnicka J., Buszewski B. Investigation of zearalenone adsorption and biotransformation by microorganisms cultured under cellular stress conditions. Toxins. 2019;11:463. doi: 10.3390/toxins11080463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Rogowska A., Pomastowski P., Rafińska K., Railean-Plugaru V., Złoch M., Walczak J., Buszewski B. A study of zearalenone biosorption and metabolisation by prokaryotic and eukaryotic cells. Toxicon. 2019;169:81–90. doi: 10.1016/j.toxicon.2019.09.008. [DOI] [PubMed] [Google Scholar]
- 171.Chlebicz A., Śliżewska K. In Vitro Detoxification of Aflatoxin B1, Deoxynivalenol, Fumonisins, T-2 Toxin and Zearalenone by Probiotic Bacteria from Genus Lactobacillus and Saccharomyces cerevisiae Yeast. Probiotics Antimicrob. Proteins. 2020;12:289–301. doi: 10.1007/s12602-018-9512-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Pfeiffer E., Schebb N.H., Podlech J., Metzler M. Novel oxidative in vitro metabolites of the mycotoxins alternariol and alternariol methyl ether. Mol. Nutr. Food Res. 2007;51:307–316. doi: 10.1002/mnfr.200600237. [DOI] [PubMed] [Google Scholar]
- 173.Pfeiffer E., Burkhardt B., Altemöller M., Podlech J., Metzler M. Activities of human recombinant cytochrome P450 isoforms and human hepatic microsomes for the hydroxylation ofAlternaria toxins. Mycotoxin Res. 2008;24:117–123. doi: 10.1007/BF03032337. [DOI] [PubMed] [Google Scholar]
- 174.Tiessen C., Ellmer D., Mikula H., Pahlke G., Warth B., Gehrke H., Zimmermann K., Heiss E., Fröhlich J., Marko D. Impact of phase I metabolism on uptake, oxidative stress and genotoxicity of the emerging mycotoxin alternariol and its monomethyl ether in esophageal cells. Arch. Toxicol. 2017;91:1213–1226. doi: 10.1007/s00204-016-1801-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Lemke A., Burkhardt B., Bunzel D., Pfeiffer E., Metzler M., Huch M., Kulling S.E., Franz C.M.A.P. Alternaria toxins of the alternariol type are not metabolised by human faecal microbiota. World Mycotoxin J. 2016;9:41–49. doi: 10.3920/WMJ2014.1875. [DOI] [Google Scholar]
- 176.Fleck S.C., Pfeiffer E., Podlech J., Metzler M. Epoxide Reduction to an Alcohol: A Novel Metabolic Pathway for Perylene Quinone-Type Alternaria Mycotoxins in Mammalian Cells. Chem. Res. Toxicol. 2014;27:247–253. doi: 10.1021/tx400366w. [DOI] [PubMed] [Google Scholar]
- 177.Burkhardt B., Wittenauer J., Pfeiffer E., Schauer U.M.D., Metzler M. Oxidative metabolism of the mycotoxins alternariol and alternariol-9-methyl ether in precision-cut rat liver slices in vitro. Mol. Nutr. Food Res. 2011;55:1079–1086. doi: 10.1002/mnfr.201000487. [DOI] [PubMed] [Google Scholar]
- 178.Pfeiffer E., Schmit C., Burkhardt B., Altemöller M., Podlech J., Metzler M. Glucuronidation of the mycotoxins alternariol and alternariol-9-methyl ether in vitro: Chemical structures of glucuronides and activities of human UDP-glucuronosyltransferase isoforms. Mycotoxin Res. 2009;25:3–10. doi: 10.1007/s12550-008-0001-z. [DOI] [PubMed] [Google Scholar]
- 179.Soukup S.T., Kohn B.N., Pfeiffer E., Geisen R., Metzler M., Bunzel M., Kulling S.E. Sulfoglucosides as Novel Modified Forms of the Mycotoxins Alternariol and Alternariol Monomethyl Ether. J. Agric. Food Chem. 2016;64:8892–8901. doi: 10.1021/acs.jafc.6b03120. [DOI] [PubMed] [Google Scholar]
- 180.Puntscher H., Marko D., Warth B. The fate of altertoxin ii during tomato processing steps at a laboratory scale. Front. Nutr. 2019;6:92. doi: 10.3389/fnut.2019.00092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Pfeiffer E., Herrmann C., Altemöller M., Podlech J., Metzler M. Oxidative in vitro metabolism of the Alternaria toxins altenuene and isoaltenuene. Mol. Nutr. Food Res. 2009;53:452–459. doi: 10.1002/mnfr.200700501. [DOI] [PubMed] [Google Scholar]
- 182.Rychlik M., Kircher F., Schusdziarra V., Lippl F. Absorption of the mycotoxin patulin from the rat stomach. Food Chem. Toxicol. 2004;42:729–735. doi: 10.1016/j.fct.2003.12.015. [DOI] [PubMed] [Google Scholar]
- 183.Rychlik M. Rapid degradation of the mycotoxin patulin in man quantified by stable isotope dilution assays. Food Addit. Contam. 2003;20:829–837. doi: 10.1080/0265203031000152424. [DOI] [PubMed] [Google Scholar]
- 184.Zheng X., Wei W., Rao S., Gao L., Li H., Yang Z. Degradation of patulin in fruit juice by a lactic acid bacteria strain Lactobacillus casei YZU01. Food Control. 2020;112:107147. doi: 10.1016/j.foodcont.2020.107147. [DOI] [Google Scholar]
- 185.Tannous J., Snini S.P., El Khoury R., Canlet C., Pinton P., Lippi Y., Alassane-Kpembi I., Gauthier T., El Khoury A., Atoui A., et al. Patulin transformation products and last intermediates in its biosynthetic pathway, E- and Z-ascladiol, are not toxic to human cells. Arch. Toxicol. 2017;91:2455–2467. doi: 10.1007/s00204-016-1900-y. [DOI] [PubMed] [Google Scholar]
- 186.Ianiri G., Idnurm A., Wright S.A.I., Durán-Patrón R., Mannina L., Ferracane R., Ritieni A., Castoria R. Searching for genes responsible for patulin degradation in a biocontrol yeast provides insight into the basis for resistance to this mycotoxin. Appl. Environ. Microbiol. 2013;79:3101–3115. doi: 10.1128/AEM.03851-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Xu R., Zhu H., Hu L., Yu B., Zhan X., Yuan Y., Zhou P. Characterization of the intestinal absorption of morroniside from Cornus officinalis Sieb. et Zucc via a Caco-2 cell monolayer model. PLoS ONE. 2020;15:e0227844. doi: 10.1371/journal.pone.0227844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Zhang J., Zheng N., Liu J., Li F.D., Li S.L., Wang J.Q. Aflatoxin B1 and aflatoxin M1 induced cytotoxicity and DNA damage in differentiated and undifferentiated Caco-2 cells. Food Chem. Toxicol. 2015;83:54–60. doi: 10.1016/j.fct.2015.05.020. [DOI] [PubMed] [Google Scholar]
- 189.Fogh J., Fogh J.M., Orfeo T. One Hundred and Twenty-Seven Cultured Human Tumor Cell Lines Producing Tumors in Nude Mice23. JNCI J. Natl. Cancer Inst. 1977;59:221–226. doi: 10.1093/jnci/59.1.221. [DOI] [PubMed] [Google Scholar]
- 190.Iftikhar M., Iftikhar A., Zhang H., Gong L., Wang J. Transport, metabolism and remedial potential of functional food extracts (FFEs) in Caco-2 cells monolayer: A Review. Food Res. Int. 2020;136:109240. doi: 10.1016/j.foodres.2020.109240. [DOI] [PubMed] [Google Scholar]
- 191.Kamiloglu S., Capanoglu E., Grootaert C., Van Camp J. Anthocyanin Absorption and Metabolism by Human Intestinal Caco-2 Cells—A Review. Int. J. Mol. Sci. 2015;16:21555–21574. doi: 10.3390/ijms160921555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Awortwe C., Fasinu P.S., Rosenkranz B. Application of Caco-2 cell line in herb-drug interaction studies: Current approaches and challenges. J. Pharm. Pharm. Sci. Publ. Can. Soc. Pharm. Sci. Société Can. Sci. Pharm. 2014;17:1–19. doi: 10.18433/J30K63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Wang X., Li L., Zhang G. Impact of deoxynivalenol and kaempferol on expression of tight junction proteins at different stages of Caco-2 cell proliferation and differentiation. RSC Adv. 2019;9:34607–34616. doi: 10.1039/C9RA06222J. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Wang X., Li L., Zhang G. A proteomic study on the protective effect of kaempferol pretreatment against deoxynivalenol-induced intestinal barrier dysfunction in a Caco-2 cell model. Food Funct. 2020;11:7266–7279. doi: 10.1039/D0FO01579B. [DOI] [PubMed] [Google Scholar]
- 195.Akbari P., Braber S., Varasteh S., Alizadeh A., Garssen J., Fink-Gremmels J. The intestinal barrier as an emerging target in the toxicological assessment of mycotoxins. Arch. Toxicol. 2017;91:1007–1029. doi: 10.1007/s00204-016-1794-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Alizadeh A., Akbari P., Varasteh S., Braber S., Malekinejad H., Fink-Gremmels J. Ochratoxin A challenges the intestinal epithelial cell integrity: Results obtained in model experiments with Caco-2 cells. World Mycotoxin J. 2019;12:399–407. doi: 10.3920/WMJ2019.2451. [DOI] [Google Scholar]
- 197.Pinton P., Nougayrède J.P., Del Rio J.C., Moreno C., Marin D.E., Ferrier L., Bracarense A.P., Kolf-Clauw M., Oswald I.P. The food contaminant deoxynivalenol, decreases intestinal barrier permeability and reduces claudin expression. Toxicol. Appl. Pharmacol. 2009;237:41–48. doi: 10.1016/j.taap.2009.03.003. [DOI] [PubMed] [Google Scholar]
- 198.Romero A., Ares I., Ramos E., Castellano V., Martínez M.M.A., Martínez-Larrañaga M.R., Anadón A., Martínez M.M.A. Mycotoxins modify the barrier function of Caco-2 cells through differential gene expression of specific claudin isoforms: Protective effect of illite mineral clay. Toxicology. 2016;353–354:21–33. doi: 10.1016/j.tox.2016.05.003. [DOI] [PubMed] [Google Scholar]
- 199.Sun H., Chow E.C.Y., Liu S., Du Y., Pang K.S. The Caco-2 cell monolayer: Usefulness and limitations. Expert Opin. Drug Metab. Toxicol. 2008;4:395–411. doi: 10.1517/17425255.4.4.395. [DOI] [PubMed] [Google Scholar]
- 200.Ferruzza S., Scarino M.L., Gambling L., Natella F., Sambuy Y. Biphasic effect of iron on human intestinal Caco-2 cells: Early effect on tight junction permeability with delayed onset of oxidative cytotoxic damage. Cell. Mol. Biol. 2003;49:89–99. [PubMed] [Google Scholar]
- 201.Videmann B., Tep J., Cavret S., Lecoeur S. Epithelial transport of deoxynivalenol: Involvement of human P-glycoprotein (ABCB1) and multidrug resistance-associated protein 2 (ABCC2) Food Chem. Toxicol. 2007;45:1938–1947. doi: 10.1016/j.fct.2007.04.011. [DOI] [PubMed] [Google Scholar]
- 202.Sergent T., Parys M., Garsou S., Pussemier L., Schneider Y.J., Larondelle Y. Deoxynivalenol transport across human intestinal Caco-2 cells and its effects on cellular metabolism at realistic intestinal concentrations. Toxicol. Lett. 2006;164:167–176. doi: 10.1016/j.toxlet.2005.12.006. [DOI] [PubMed] [Google Scholar]
- 203.Artursson P., Palm K., Luthman K. Caco-2 monolayers in experimental and theoretical predictions of drug transport. Adv. Drug Deliv. Rev. 2012;22:280–289. doi: 10.1016/j.addr.2012.09.005. [DOI] [PubMed] [Google Scholar]
- 204.Kadota T., Furusawa H., Hirano S., Tajima O., Kamata Y., Sugita-Konishi Y. Comparative study of deoxynivalenol, 3-acetyldeoxynivalenol, and 15-acetyldeoxynivalenol on intestinal transport and IL-8 secretion in the human cell line Caco-2. Toxicol. In Vitro. 2013;27:1888–1895. doi: 10.1016/j.tiv.2013.06.003. [DOI] [PubMed] [Google Scholar]
- 205.Sobral M.M.C., Faria M.A., Cunha S.C., Miladinovic B., Ferreira I.M. Transport of mycotoxins across human gastric NCI–N87 and intestinal Caco-2 cell models. Food Chem. Toxicol. 2019;131:110595. doi: 10.1016/j.fct.2019.110595. [DOI] [PubMed] [Google Scholar]
- 206.Tep J., Videmann B., Mazallon M., Balleydier S., Cavret S., Lecoeur S. Transepithelial transport of fusariotoxin nivalenol: Mediation of secretion by ABC transporters. Toxicol. Lett. 2007;170:248–258. doi: 10.1016/j.toxlet.2007.03.012. [DOI] [PubMed] [Google Scholar]
- 207.Pfeiffer E., Kommer A., Dempe J.S., Hildebrand A.A., Metzler M. Absorption and metabolism of the mycotoxin zearalenone and the growth promotor zeranol in Caco-2 cells in vitro. Mol. Nutr. Food Res. 2011;55:560–567. doi: 10.1002/mnfr.201000381. [DOI] [PubMed] [Google Scholar]
- 208.Cirlini M., Barilli A., Galaverna G., Michlmayr H., Adam G., Berthiller F., Dall’Asta C. Study on the uptake and deglycosylation of the masked forms of zearalenone in human intestinal Caco-2 cells. Food Chem. Toxicol. 2016;98:232–239. doi: 10.1016/j.fct.2016.11.003. [DOI] [PubMed] [Google Scholar]
- 209.Prosperini A., Meca G., Font G., Ruiz M.J. Study of the cytotoxic activity of beauvericin and fusaproliferin and bioavailability in vitro on Caco-2 cells. Food Chem. Toxicol. 2012;50:2356–2361. doi: 10.1016/j.fct.2012.04.030. [DOI] [PubMed] [Google Scholar]
- 210.De Angelis I., Friggè G., Raimondi F., Stammati A., Zucco F., Caloni F. Absorption of Fumonisin B1 and aminopentol on an in vitro model of intestinal epithelium; the role of P-glycoprotein. Toxicon. 2005;45:285–291. doi: 10.1016/j.toxicon.2004.10.015. [DOI] [PubMed] [Google Scholar]
- 211.Berger V., Gabriel A.F., Sergent T., Trouet A., Larondelle Y., Schneider Y.J. Interaction of ochratoxin A with human intestinal Caco-2 cells: Possible implication of a multidrug resistance-associated protein (MRP2) Toxicol. Lett. 2003;140–141:465–476. doi: 10.1016/S0378-4274(03)00043-2. [DOI] [PubMed] [Google Scholar]
- 212.De Walle J.V., Sergent T., Piront N., Toussaint O., Schneider Y.-J., Larondelle Y. Deoxynivalenol affects in vitro intestinal epithelial cell barrier integrity through inhibition of protein synthesis. Toxicol. Appl. Pharmacol. 2010;245:291–298. doi: 10.1016/j.taap.2010.03.012. [DOI] [PubMed] [Google Scholar]
- 213.De Nijs M., Van Den Top H.J., Portier L., Oegema G., Kramer E., Van Egmond H.P., Hoogenboom L.A.P. Digestibility and absorption of deoxynivalenol-3-ß-glucoside in in vitro models. World Mycotoxin J. 2012;5:319–324. doi: 10.3920/WMJ2012.1430. [DOI] [Google Scholar]
- 214.Schrickx J., Lektarau Y., Fink-Gremmels J. Ochratoxin A secretion by ATP-dependent membrane transporters in Caco-2 cells. Arch. Toxicol. 2006;80:243–249. doi: 10.1007/s00204-005-0041-5. [DOI] [PubMed] [Google Scholar]
- 215.Tuntiteerawit P., Jarukamjorn K., Porasuphatana S. The effect of green tea catechins on breast cancer resistance protein activity and intestinal efflux of aflatoxin B1 via breast cancer resistance protein in Caco-2 cells. Toxicol. Res. 2020;36:293–300. doi: 10.1007/s43188-019-00032-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Li X., Mu P., Wen J., Deng Y. Carrier-Mediated and Energy-Dependent Uptake and Efflux of Deoxynivalenol in Mammalian Cells. Sci. Rep. 2017;7:1–10. doi: 10.1038/s41598-017-06199-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Li X., Mu P., Qiao H., Wen J., Deng Y. JNK-AKT-NF-κB controls P-glycoprotein expression to attenuate the cytotoxicity of deoxynivalenol in mammalian cells. Biochem. Pharmacol. 2018;156:120–134. doi: 10.1016/j.bcp.2018.08.020. [DOI] [PubMed] [Google Scholar]
- 218.Ivanova L., Fæste C.K., Solhaug A. Role of P-glycoprotein in deoxynivalenol-mediated in vitro toxicity. Toxicol. Lett. 2018;284:21–28. doi: 10.1016/j.toxlet.2017.11.021. [DOI] [PubMed] [Google Scholar]
- 219.Anderle P., Niederer E., Rubas W., Hilgendorf C., Spahn-Langguth H., Wunderli-Allenspach H., Merkle H.P., Langguth P. P-glycoprotein (P-gp) mediated efflux in Caco-2 cell monolayers: The influence of culturing conditions and drug exposure on P-gp expression levels. J. Pharm. Sci. 1998;87:757–762. doi: 10.1021/js970372e. [DOI] [PubMed] [Google Scholar]
- 220.Videmann B., Mazallon M., Prouillac C., Delaforge M., Lecoeur S. ABCC1, ABCC2 and ABCC3 are implicated in the transepithelial transport of the myco-estrogen zearalenone and its major metabolites. Toxicol. Lett. 2009;190:215–223. doi: 10.1016/j.toxlet.2009.07.021. [DOI] [PubMed] [Google Scholar]
- 221.Xu R., Karrow N.A., Shandilya U.K., Sun L.H., Kitazawa H. In-vitro cell culture for efficient assessment of mycotoxin exposure, toxicity and risk mitigation. Toxins. 2020;12:146. doi: 10.3390/toxins12030146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Gao Y., Li S., Wang J., Luo C., Zhao S., Zheng N. Modulation of intestinal epithelial permeability in differentiated caco-2 cells exposed to aflatoxin M1 and ochratoxin a individually or collectively. Toxins. 2018;10:13. doi: 10.3390/toxins10010013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Marin D., Motiu M., Taranu I. Food Contaminant Zearalenone and Its Metabolites Affect Cytokine Synthesis and Intestinal Epithelial Integrity of Porcine Cells. Toxins. 2015;7:1979–1988. doi: 10.3390/toxins7061979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Bouhet S., Hourcade E., Loiseau N., Fikry A., Roselli M., Galtier P., Mengheri E., Oswald I.P., Martinez S., Roselli M., et al. The mycotoxin fumonisin B1 alters the proliferation and the barrier function of porcine intestinal epithelial cells. Toxicol. Sci. 2004;77:165–171. doi: 10.1093/toxsci/kfh006. [DOI] [PubMed] [Google Scholar]
- 225.Pinton P., Braicu C., Nougayrede J.-P., Laffitte J., Taranu I., Oswald I.P. Deoxynivalenol Impairs Porcine Intestinal Barrier Function and Decreases the Protein Expression of Claudin-4 through a Mitogen-Activated Protein Kinase-Dependent Mechanism. J. Nutr. 2010;140:1956–1962. doi: 10.3945/jn.110.123919. [DOI] [PubMed] [Google Scholar]
- 226.Madara J.L. Regulation of the Movement of Solutes Across Tight Junctions. Annu. Rev. Physiol. 1998;60:143–159. doi: 10.1146/annurev.physiol.60.1.143. [DOI] [PubMed] [Google Scholar]
- 227.Gao Y.N., Wang J.Q., Li S.L., Zhang Y.D., Zheng N. Aflatoxin M1 cytotoxicity against human intestinal Caco-2 cells is enhanced in the presence of other mycotoxins. Food Chem. Toxicol. 2016;96:79–89. doi: 10.1016/j.fct.2016.07.019. [DOI] [PubMed] [Google Scholar]
- 228.Fleck S.C., Pfeiffer E., Metzler M. Permeation and metabolism of Alternaria mycotoxins with perylene quinone structure in cultured Caco-2 cells. Mycotoxin Res. 2014;30:17–23. doi: 10.1007/s12550-013-0180-0. [DOI] [PubMed] [Google Scholar]
- 229.González-Arias C.A., Marín S., Rojas-García A.E., Sanchis V., Ramos A.J. UPLC-MS/MS analysis of ochratoxin A metabolites produced by Caco-2 and HepG2 cells in a co-culture system. Food Chem. Toxicol. 2017;109:333–340. doi: 10.1016/j.fct.2017.09.011. [DOI] [PubMed] [Google Scholar]
- 230.Matter K., Balda M.S. Signalling to and from tight junctions. Nat. Rev. Mol. Cell Biol. 2003;4:225–237. doi: 10.1038/nrm1055. [DOI] [PubMed] [Google Scholar]
- 231.Moon Y. Vomitoxin-Induced Cyclooxygenase-2 Gene Expression in Macrophages Mediated by Activation of ERK and p38 but Not JNK Mitogen-Activated Protein Kinases. Toxicol. Sci. 2002;69:373–382. doi: 10.1093/toxsci/69.2.373. [DOI] [PubMed] [Google Scholar]
- 232.Zhou H.-R., Jia Q., Pestka J.J. Ribotoxic Stress Response to the Trichothecene Deoxynivalenol in the Macrophage Involves the Src Family Kinase Hck. Toxicol. Sci. 2005;85:916–926. doi: 10.1093/toxsci/kfi146. [DOI] [PubMed] [Google Scholar]
- 233.Pinton P., Graziani F., Pujol A., Nicoletti C., Paris O., Ernouf P., Di Pasquale E., Perrier J., Oswald I.P., Maresca M. Deoxynivalenol inhibits the expression by goblet cells of intestinal mucins through a PKR and MAP kinase dependent repression of the resistin-like molecule β. Mol. Nutr. Food Res. 2015;59:1076–1087. doi: 10.1002/mnfr.201500005. [DOI] [PubMed] [Google Scholar]
- 234.Morrison D.K. MAP Kinase Pathways. Cold Spring Harb. Perspect. Biol. 2012;4:a011254. doi: 10.1101/cshperspect.a011254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Lee J.Y., Lim W., Park S., Kim J., You S., Song G. Deoxynivalenol induces apoptosis and disrupts cellular homeostasis through MAPK signaling pathways in bovine mammary epithelial cells. Environ. Pollut. 2019;252:879–887. doi: 10.1016/j.envpol.2019.06.001. [DOI] [PubMed] [Google Scholar]
- 236.Bouhet S., Le E., Peres S., Fairbrother J.M., Oswald I.P., Hyacinthe S. Mycotoxin fumonisin B 1 selectively down-regulates the basal IL-8 expression in pig intestine: In vivo and in vitro studies. Food Chem. Toxicol. 2006;44:1768–1773. doi: 10.1016/j.fct.2006.05.018. [DOI] [PubMed] [Google Scholar]
- 237.Beisl J., Pahlke G., Abeln H., Ehling-Schulz M., Del Favero G., Varga E., Warth B., Sulyok M., Abia W., Ezekiel C.N., et al. Combinatory effects of cereulide and deoxynivalenol on in vitro cell viability and inflammation of human Caco-2 cells. Arch. Toxicol. 2020;94:833–844. doi: 10.1007/s00204-020-02658-w. [DOI] [PubMed] [Google Scholar]
- 238.Pellegrina C.D., Perbellini O., Scupoli M.T., Tomelleri C., Zanetti C., Zoccatelli G., Fusi M., Peruffo A., Rizzi C., Chignola R. Effects of wheat germ agglutinin on human gastrointestinal epithelium: Insights from an experimental model of immune/epithelial cell interaction. Toxicol. Appl. Pharmacol. 2009;237:146–153. doi: 10.1016/j.taap.2009.03.012. [DOI] [PubMed] [Google Scholar]
- 239.Gao Y., Ye Q., Bao X., Huang X., Wang J., Zheng N. Transcriptomic and proteomic profiling reveals the intestinal immunotoxicity induced by aflatoxin M1 and ochratoxin A. Toxicon. 2020;180:49–61. doi: 10.1016/j.toxicon.2020.03.008. [DOI] [PubMed] [Google Scholar]
- 240.Trapecar M., Cencic A. Application of Gut Cell Models for Toxicological and Bioactivity Studies of Functional and Novel Foods. Foods. 2014;1:40–51. doi: 10.3390/foods1010040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Fu J., Cui Y. In vitro digestion/Caco-2 cell model to estimate cadmium and lead bioaccessibility/bioavailability in two vegetables: The influence of cooking and additives. Food Chem. Toxicol. 2013;59:215–221. doi: 10.1016/j.fct.2013.06.014. [DOI] [PubMed] [Google Scholar]
- 242.Ekmekcioglu C. A physiological approach for preparing and conducting intestinal bioavailability studies using experimental systems. Food Chem. 2002;76:225–230. doi: 10.1016/S0308-8146(01)00291-6. [DOI] [Google Scholar]
- 243.Seithel A., Karlsson J., Hilgendorf C., Bj A., Ungell A.-L., Björquist A., Ungell A.-L. Variability in mRNA expression of ABC- and SLC-transporters in human intestinal cells: Comparison between human segments and Caco-2 cells. Eur. J. Pharm. Sci. 2006;28:291–299. doi: 10.1016/j.ejps.2006.03.003. [DOI] [PubMed] [Google Scholar]
- 244.Taipalensuu J., Törnblom H., Lindberg G., Einarsson C., Sjöqvist F., Melhus H., Garberg P., Sjöström B., Lundgren B., Artursson P. Correlation of Gene Expression of Ten Drug Efflux Proteins of the ATP-Binding Cassette Transporter Family in Normal Human Jejunum and in Human Intestinal Epithelial Caco-2 Cell Monolayers. J. Pharmacol. Exp. Ther. 2001;299:164–170. [PubMed] [Google Scholar]
- 245.Lampen A., Bader A., Bestmann T., Winkler M., Witte L., Borlak J.T. Catalytic activities, protein- and mRNA-expression of cytochrome P450 isoenzymes in intestinal cell lines. Xenobiotica. 1998;28:429–441. doi: 10.1080/004982598239362. [DOI] [PubMed] [Google Scholar]
- 246.Caloni F., Cortinovis C., Pizzo F., De Angelis I. Transport of aflatoxin M1 in human intestinal Caco-2/TC7 cells. Front. Pharmacol. 2012;3:111. doi: 10.3389/fphar.2012.00111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Wu C., Gao Y., Li S., Huang X., Bao X., Wang J., Zheng N. Modulation of intestinal epithelial permeability and mucin mRNA (MUC2, MUC5AC, and MUC5B) expression and protein secretion in Caco-2/HT29-MTX co-cultures exposed to aflatoxin M1, ochratoxin A, and zearalenone individually or collectively. Toxicol. Lett. 2019;309:1–9. doi: 10.1016/j.toxlet.2019.03.010. [DOI] [PubMed] [Google Scholar]