

Abstract
Purpose
The aim of this study was to compare microRNA (miRNA) gene expression in saliva using miRNA polymerase chain reaction (PCR) arrays in healthy and aggressive periodontitis (AP) patients.
Methods
PCR arrays of 84 miRNAs related to the human inflammatory response and autoimmunity from the saliva samples of 4 patients with AP and 4 healthy controls were performed. The functions and diseases related to the miRNAs were obtained using TAM 2.0. Experimentally validated targets of differentially expressed miRNAs were obtained from mirTarBase. Gene ontology terms and pathways were analyzed using ConsensusPathDB.
Results
Four downregulated miRNAs (hsa-let-7a-5p, hsa-let-7f-5p, hsa-miR-181b-5p, and hsa-miR-23b-3p) were identified in patients with AP. These miRNAs are associated with cell death and innate immunity, and they target genes associated with osteoclast development and function.
Conclusions
This study is the first analysis of miRNAs in the saliva of patients with AP. Identifying discriminatory human salivary miRNA biomarkers reflective of periodontal disease in a non-invasive screening assay is crucial for the development of salivary diagnostics. These data provide a first step towards the discovery of key salivary miRNA biomarkers for AP.
Keywords: Aggressive periodontitis, Biomarkers, MicroRNAs, Saliva
Graphical Abstract
INTRODUCTION
MicroRNAs (miRNAs) are noncoding RNAs that are approximately 22 nucleotides long and can act as key regulators of diverse biological processes by mediating translational repression or messenger RNA (mRNA) degradation of their target genes [1,2,3,4]. The seed sequence of miRNAs specifically binds to the 3′ untranslated region of target mRNAs to exert its action [5,6]. Many studies have documented that miRNAs play a key role in certain infectious diseases [7], autoimmune diseases [8], chronic inflammatory diseases [9], cardiovascular diseases [10], cancers [11], and other diseases [12]. Furthermore, recent studies have reported that miRNAs have great potential as biomarkers for these diseases [13,14,15]. However, most of those studies analyzed miRNA levels in subjects' serum or plasma, which requires invasive collection methods. To overcome this drawback, some recent studies have attempted to analyze miRNAs collected noninvasively from saliva, reporting promise for detecting pancreatic cancer [16], oral cancer [17], Sjögren syndrome [18], and esophageal cancer [19].
Periodontal remodeling and inflammation are regulated by miRNAs. Several studies have demonstrated that miRNAs play a role in osteogenesis by regulating bone morphogenic protein 7 (BMP-7); runt-related transcription factor 2 (RUNX2); osterix; and osteocalcin; in osteoclastogenesis by regulating protein inhibitor of activated STAT3 (PIAS3); nuclear factor of activated T-cells, cytoplasmic, calcineurin-dependent 1 (NFATc1); nuclear factor kappa B (NF-κB); and receptor activator of NF-κB ligand (RANKL); and in the inflammatory process by regulating inflammatory mediators such as interleukin-1β (IL-1β); IL-6; IL-8; tumor necrosis factor-α (TNF-α); and prostaglandin E2 (PGE2) [5].
Other studies have found that miRNAs were differentially expressed in healthy and diseased human gingival tissues [20,21,22,23,24,25]. Xie et al. [20], Stoecklin-Wasmer et al. [21], and Ogata et al. [22] found differentially expressed miRNAs in chronic periodontitis (CP) tissue using microarrays. Lee et al. [23], Perri et al. [24], and Na et al. [25] identified differentially expressed miRNAs in CP tissue using quantitative polymerase chain reaction (PCR) arrays.
Microarray techniques have been used widely to profile miRNAs. The expression levels of thousands of miRNAs can be determined simultaneously in a single microarray experiment. However, the unique characteristics of miRNAs, such as their short length, differing melting temperatures, and the highly similar sequences between miRNA family members, make the design of microarray probes difficult [26]. Furthermore, a large amount of miRNAs are needed for microarray techniques [27], while PCR-based techniques can detect nanograms of miRNA with high sensitivity and specificity. Furthermore, more recently developed quantitative PCR-based arrays combine the high sensitivity of PCR-based techniques and the ability of microarray techniques to profile a large number of miRNA samples in a single experiment [27,28].
The studies discussed above used gingival biopsies, which are very invasive. To the best of our knowledge, only 1 study has evaluated the feasibility of using salivary miRNAs as a non-invasive diagnostic tool for CP [29].
Aggressive periodontitis (AP) is the most destructive type of periodontal disease [30]. While there are more genetic risk factors for AP than for CP, few studies have evaluated miRNA expression profiles in AP. Amaral et al. compared the expression patterns of miRNAs between CP and AP using gingival biopsies and PCR arrays [31], and Ghotloo et al. [32] compared expression levels of hsa-miR-146a between AP patients and healthy controls using gingival biopsies and real-time PCR. To our best knowledge, no study has utilized salivary miRNAs to identify AP. The aim of this study was to compare the miRNA gene expression profile in saliva using miRNA PCR arrays in healthy and AP patients.
MATERIALS AND METHODS
Study design
Eight subjects who visited the Department of Periodontics at Asan Medical Center between March and August 2012 were enrolled in this study. The test group for this study consisted of 4 patients who were diagnosed with AP, and the control group consisted of 4 patients with healthy periodontia. The inclusion criteria for the test group were as follows: 1) subjects who were younger than 40 years old and 2) subjects who were diagnosed with AP before 35 years of age in accordance with the classification system defined by the American Academy of Periodontology [33]. The inclusion criteria for the control group were as follows: 1) subjects who were younger than 40-years old and 2) subjects who had a healthy periodontium and who were not diagnosed with AP (had more than 20 permanent teeth, all probing pocket depths were 4 mm or less, and all clinical attachment levels were 2 mm or less). The exclusion criteria for both the control and test groups were as follows: 1) subjects who received periodontal treatment or antibiotic treatment within 3 months prior to participating in this study, 2) subjects who regularly received medication known to affect periodontal status, 3) subjects who required prophylactic antibiotics, 4) subjects who were pregnant or breastfeeding, 5) subjects who were alcoholics or who had a history of drug abuse, and 6) subjects with systemic diseases. This study was approved by the Institutional Review Board of Asan Medical Center (IRB No. 2012-0120). Informed consent was obtained from all subjects before they enrolled in the study.
Saliva collection
Unstimulated saliva samples were collected using a previously defined protocol [34]. Briefly, subjects were asked not to consume any food or water for at least 1 hour prior to saliva collection, and they were asked to rinse their mouth with sterile water for a minute. After 5 minutes, the subjects were asked to sit upright, and 2–3 mL of unstimulated whole saliva was collected in plastic tubes from each subject in the control and test groups. The whole saliva samples were stored at −70°C until use.
MicroRNA PCR array experiment
The miRNAs were isolated from 400-µL saliva samples using an Ambion mirVana miRNA isolation kit (Life Technologies, Carlsbad, CA, USA) according to the manufacturer's instruction. Complementary DNA (cDNA) was synthesized with a miScript II RT kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. Finally, PCR array experiments were performed using a miScript miRNA PCR Array for Human Inflammatory Response and Autoimmunity and miScript SYBR Green PCR kit (Qiagen). The miScript miRNA PCR Array for Human Inflammatory Response and Autoimmunity identifies 84 miRNAs related to inflammatory response and autoimmunity. A total volume of 1,100 µL of PCR reaction mix was prepared (550 µL of 2× QuantiTect SYBR Green PCR master mix, 110 µL of 10× miScript universal primer, 340 µL of RNase-free water and 100 µL of template cDNA) before addition of 10 µL of reaction mix to each well of the miScript miRNA PCR Array. Optical adhesive film was then sealed onto the array. The array was centrifuged for 1 minute at 1,000 ×g at room temperature to remove bubbles. The PCR real-time cycler was programmed as follows: the initial activation step was carried out for 15 minutes at 95°C, followed by 40 cycles of denaturation (15 seconds, 94°C), annealing (30 seconds, 55°C), and extension (30 seconds, 70°C).
Data analysis
Normalized gene expression values and fold changes between the test and control groups for each miRNA were calculated using the ΔΔCt method with the data from the PCR array. The Student t-test was used to evaluate the significance of differences in normalized gene expression levels between the test and control groups. Differences were considered significant when P<0.05. The enriched miRNA clusters, families, miRNA-associated diseases, and functions of the differentially expressed miRNAs were determined using TAM 2.0 [35]. Experimentally validated target mRNAs were obtained from miRTarBase [36]. Finally, gene ontology (GO) terms and pathways were analyzed with ConsensusPathDB [37] using the selected target genes obtained from miRTarBase.
RESULTS
The demographic data for the control and test groups are summarized in Table 1.
Table 1. Demographic data.
| Characteristics | Control group (n=4) | Test group (n=4) |
|---|---|---|
| Age (years) | 31.3±3.8 | 29.5±3.1 |
| Sex (M/F) | 2/2 | 2/2 |
| PPD (mm) | 2.47±0.15 | 4.55±1.01 |
| Diagnosis (GAP/LAP) | -/- | 2/2 |
| Smoking history | 0% | 25% |
PPD: probing pocket depth, GAP: generalized aggressive periodontitis, LAP: localized aggressive periodontitis.
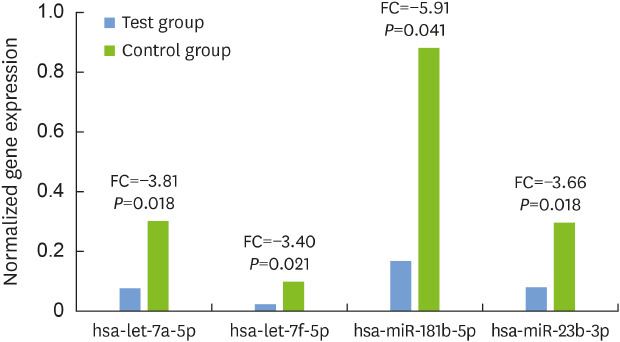
We identified 4 significantly downregulated miRNAs in subjects with AP compared to healthy subjects (Fig. 1). The 4 miRNAs were hsa-let-7a-5p, hsa-let-7f-5p, hsa-miR-181b-5p, and hsa-miR-23b-3p. The fold change of those miRNAs was −3.81, −3.40, −5.91, and −3.66, respectively.
Figure 1. Differentially expressed miRNAs. FC of statistically significantly downregulated miRNAs in subjects with aggressive periodontitis (P<0.05).

miRNA: microRNA, FC: fold changes.
The enriched miRNA clusters, families, miRNA-associated diseases, and functions for these 4 miRNAs were determined by TAM 2.0. Table 2 shows a total of 26 miRNA categories enriched by the 4 differentially expressed miRNAs (both adjusted P values less than 0.05 after adjustment for multiple testing by the Bonferroni and false discovery rate methods). Interestingly, 8 family members (hsa-let-7a-1, hsa-let-7a-2, hsa-let-7a-3, hsa-let-7f-1, hsa-let-7f-2, hsa-mir-181b-1, hsa-mir-181b-2, and hsa-mir-23b) of all differentially expressed miRNAs (hsa-let-7a-5p, hsa-let-7f-5p, hsa-miR181b-5p, and hsa-miR-23b-3p) were associated with the cell death function, and 6 family members (hsa-let-7a-1, hsa-let-7a-2, hsa-let-7f-1, hsa-let-7f-2, hsa-mir-181b-1, and hsa-mir-181b-2) of 3 miRNAs (hsa-let-7a-5p, hsa-let-7f-5p, and hsa-miR181b-5p) were associated with innate immunity. There were no enriched disease categories known to be either directly or indirectly related to AP except diabetes mellitus.
Table 2. Identified enriched miRNA clusters, families, miRNA-associated diseases, and functions (both adjusted P<0.05).
| Variables | Factor | # of miRNAs | % | P value | Adj. P (Bonferroni) | Adj. P (FDR) |
|---|---|---|---|---|---|---|
| Cluster | hsa-let-7a-1 cluster | 2 | 66.7 | <0.001 | 0.016 | 0.005 |
| Family | let-7 family | 5 | 41.7 | <0.001 | <0.001 | <0.001 |
| Function | Cell death | 8 | 10.3 | <0.001 | <0.001 | <0.001 |
| Innate immunity | 6 | 14.3 | <0.001 | <0.001 | <0.001 | |
| Adipocyte differentiation | 5 | 12.2 | <0.001 | <0.001 | <0.001 | |
| Neural stem cell differentiation | 3 | 50.0 | <0.001 | <0.001 | <0.001 | |
| Cell differentiation | 5 | 8.9 | 0.014 | 0.002 | 0.001 | |
| Aging | 5 | 7.9 | 0.003 | 0.003 | 0.002 | |
| Folliculogenesis | 3 | 27.3 | <0.001 | 0.005 | 0.002 | |
| Cell division | 3 | 17.6 | <0.001 | 0.020 | 0.006 | |
| Cardiac regeneration | 3 | 13.6 | <0.001 | 0.044 | 0.014 | |
| Disease | Vascular hypertrophy | 5 | 23.8 | <0.001 | <0.001 | <0.001 |
| Psoriasis | 5 | 15.2 | <0.001 | <0.001 | <0.001 | |
| Diabetes mellitus | 6 | 8.1 | <0.001 | <0.001 | <0.001 | |
| Asthma | 5 | 12.2 | <0.001 | <0.001 | <0.001 | |
| Hepatitis B virus infection | 5 | 7.5 | <0.001 | 0.005 | 0.002 | |
| ACTH-secreting pituitary adenoma | 3 | 27.3 | <0.001 | 0.005 | 0.002 | |
| Human papilloma virus infection | 4 | 11.8 | <0.001 | 0.006 | 0.003 | |
| Autoimmune diseases | 4 | 11.1 | <0.001 | 0.008 | 0.003 | |
| Spinal cord injuries | 3 | 23.1 | <0.001 | 0.008 | 0.003 | |
| Macular degeneration | 3 | 21.4 | <0.001 | 0.011 | 0.004 | |
| Dengue virus infection | 3 | 20.0 | <0.001 | 0.013 | 0.005 | |
| Diabetes mellitus, type 2 | 6 | 4.1 | <0.001 | 0.016 | 0.005 | |
| Alzheimer's disease | 5 | 5.0 | <0.001 | 0.032 | 0.010 | |
| Hirschsprung's disease | 3 | 13.6 | <0.001 | 0.044 | 0.014 | |
| Liver cirrhosis | 4 | 7.0 | <0.001 | 0.048 | 0.014 |
miRNA: microRNA, FDR: false discovery rate.
Experimentally validated target mRNAs were obtained from miRTarBase (Table 3). A total of 46 target genes that were targeted by more than 2 differentially expressed miRNAs and whose adjusted P values were lower than 0.05 were identified. Table 3 shows target genes that had adjusted P values lower than 0.02. CCNB2, SNRPC, TAB2, EIF3C, HNRNPUL1, and E2F1 were targeted by 3 differentially expressed miRNAs.
Table 3. Experimentally validated target mRNAs using miRTarBase (adjusted P<0.02).
| Gene symbol | Gene name | # of miRNAs | P value | Adj. P (FDR) |
|---|---|---|---|---|
| CCNB2 | Cyclin B2 | 3 | <0.001 | <0.001 |
| SNRPC | Small nuclear ribonucleoprotein polypeptide C | 3 | <0.001 | 0.002 |
| TAB2 | TGF-beta activated kinase 1/MAP3K7 binding protein 2 | 3 | <0.001 | 0.002 |
| NUDT4 | Nudix (nucleoside diphosphate linked moiety X)-type motif 4 | 2 | <0.001 | 0.005 |
| EIF3C | Eukaryotic translation initiation factor 3, subunit C | 3 | <0.001 | 0.013 |
| HNRNPUL1 | Heterogeneous nuclear ribonucleoprotein U-like 1 | 3 | <0.001 | 0.014 |
| E2F1 | E2F transcription factor 1 | 3 | <0.001 | 0.014 |
| ALDH7A1 | Aldehyde dehydrogenase 7 family, member A1 | 2 | <0.001 | 0.014 |
| WWP2 | WW domain containing E3 ubiquitin protein ligase 2 | 2 | <0.001 | 0.014 |
| GBF1 | Golgi brefeldin A resistant guanine nucleotide exchange factor 1 | 2 | <0.001 | 0.017 |
| UBAP2 | Ubiquitin associated protein 2 | 2 | <0.001 | 0.017 |
| CEP170B | Centrosomal protein 170B | 2 | <0.001 | 0.017 |
mRNA: messenger RNA, miRNA: microRNA, FDR: false discovery rate.
GO terms and pathways were identified with ConsensusPathDB using the 46 selected target genes from the above miRTarBase results (Tables 4 and 5).
Table 4. GO terms with ConsensusPathDB (category BP level 4) (adjusted P<0.02).
| GO term | Set size | Candidates contained | P value | Adj. P (FDR) | |
|---|---|---|---|---|---|
| GO:0090304 | Nucleic acid metabolic process | 5,174 | 24 (0.5%) | <0.001 | 0.005 |
| GO:0034645 | Cellular macromolecule biosynthetic process | 4,867 | 23 (0.5%) | <0.001 | 0.005 |
| GO:0009889 | Regulation of biosynthetic process | 4,246 | 21 (0.5%) | <0.001 | 0.005 |
| GO:0009059 | Macromolecule biosynthetic process | 5,010 | 23 (0.5%) | <0.001 | 0.005 |
| GO:0044271 | Cellular nitrogen compound biosynthetic process | 4,906 | 22 (0.5%) | <0.001 | 0.009 |
| GO:0072359 | Circulatory system development | 1,033 | 9 (0.9%) | <0.001 | 0.011 |
| GO:0010467 | Gene expression | 5,440 | 23 (0.4%) | <0.001 | 0.011 |
| GO:0043902 | Positive regulation of multi-organism process | 161 | 4 (2.5%) | <0.001 | 0.011 |
| GO:0006338 | Chromatin remodeling | 167 | 4 (2.4%) | <0.001 | 0.011 |
| GO:0051851 | Modification by host of symbiont morphology or physiology | 75 | 3 (4.0%) | <0.001 | 0.011 |
| GO:0016569 | Covalent chromatin modification | 480 | 6 (1.2%) | <0.001 | 0.011 |
| GO:0044772 | Mitotic cell cycle phase transition | 495 | 6 (1.2%) | <0.001 | 0.012 |
| GO:0034654 | Nucleobase-containing compound biosynthetic process | 4,222 | 19 (0.5%) | <0.001 | 0.013 |
| GO:0016032 | Viral process | 721 | 7 (1.0%) | <0.001 | 0.013 |
| GO:0018130 | Heterocycle biosynthetic process | 4,285 | 19 (0.4%) | 0.001 | 0.013 |
| GO:0019438 | Aromatic compound biosynthetic process | 4,296 | 19 (0.4%) | 0.001 | 0.013 |
| GO:0043903 | Regulation of symbiosis, encompassing mutualism through parasitism | 206 | 4 (1.9%) | 0.001 | 0.013 |
| GO:0051276 | Chromosome organization | 1,211 | 9 (0.7%) | 0.001 | 0.013 |
| GO:0051171 | Regulation of nitrogen compound metabolic process | 5,904 | 23 (0.4%) | 0.001 | 0.014 |
| GO:1901362 | Organic cyclic compound biosynthetic process | 4,434 | 19 (0.4%) | 0.002 | 0.016 |
| GO:0044774 | Mitotic DNA integrity checkpoint | 106 | 3 (2.8%) | 0.002 | 0.016 |
| GO:0051817 | Modification of morphology or physiology of other organism involved in symbiotic interaction | 111 | 3 (2.7%) | 0.002 | 0.017 |
| GO:0080090 | Regulation of primary metabolic process | 6,071 | 23 (0.4%) | 0.002 | 0.018 |
| GO:0044843 | Cell cycle G1/S phase transition | 252 | 4 (1.6%) | 0.002 | 0.018 |
| GO:0071826 | Ribonucleoprotein complex subunit organization | 253 | 4 (1.6%) | 0.002 | 0.018 |
| GO:0031323 | Regulation of cellular metabolic process | 6,130 | 23 (0.4%) | 0.002 | 0.018 |
| GO:0006974 | Cellular response to DNA damage stimulus | 850 | 7 (0.8%) | 0.003 | 0.018 |
| GO:0060255 | Regulation of macromolecule metabolic process | 6,156 | 23 (0.4%) | 0.003 | 0.018 |
| GO:0051101 | Regulation of DNA binding | 127 | 3 (2.4%) | 0.003 | 0.019 |
GO: gene ontology, BP: blood pressure, FDR: false discovery rate.
Table 5. Pathway analysis with ConsensusPathDB (adjusted P<0.02).
| Pathway | Source | Percentage | P value | Adj. P (FDR) |
|---|---|---|---|---|
| Direct p53 effectors | PID | 6 (4.2%) | <0.001 | <0.001 |
| E2F transcription factor network | PID | 4 (5.3%) | <0.001 | 0.002 |
| Regulation of TP53 expression and degradation | Wikipathways | 2 (50.0%) | <0.001 | 0.002 |
| FOXM1 transcription factor network | PID | 3 (7.1%) | <0.001 | 0.006 |
| HATs acetylate histones | Reactome | 4 (2.8%) | <0.001 | 0.014 |
| Integrated breast cancer pathway | Wikipathways | 3 (4.5%) | <0.001 | 0.016 |
| Polo-like kinase mediated events | Reactome | 2 (12.5%) | <0.001 | 0.016 |
| Herpes simplex infection - Homo sapiens (human) | KEGG | 4 (2.2%) | 0.001 | 0.018 |
| Cellular senescence | Reactome | 4 (2.1%) | 0.001 | 0.018 |
| C-MYB transcription factor network | PID | 3 (3.5%) | 0.001 | 0.018 |
| miRNA regulation of DNA damage response | Wikipathways | 3 (3.4%) | 0.001 | 0.018 |
| Cellular responses to stress | Reactome | 5 (1.5%) | 0.002 | 0.018 |
| B-WICH complex positively regulates rRNA expression | Reactome | 3 (3.2%) | 0.002 | 0.018 |
| Formation of the beta-catenin: TCF transactivating complex | Reactome | 3 (3.2%) | 0.002 | 0.018 |
FDR: false discovery rate, PID: Pathway Interaction Database, KEGG: Kyoto Encyclopedia of Genes and Genomes.
DISCUSSION
In this study, 4 differentially expressed miRNAs were identified in saliva samples of patients with AP using PCR arrays. Common characteristics of the differentially expressed miRNAs, such as related functions, diseases, and their potential target genes, were analyzed using TAM 2.0 and mirTarBase. Based on the target genes, GO terms and pathways were further analyzed using ConsensusPathDB. To the best of our knowledge, this is the first study to identify differentially expressed miRNAs in AP using saliva samples.
The 4 miRNAs (hsa-let-7a-5p, hsa-let-7f-5p, hsa-miR-181b-5p, and hsa-miR-23b-3p), which play a role in cell death and innate immunity by targeting CCNB2, SNRPC, TAB2, EIF3C, HNRNPUL1, and E2F1, were found to be significantly downregulated in AP in this study. Among these genes, the TAB2 gene encodes the activator of MAP3K7/TAK1, which is required for IL-1 induced activation of NF-κB and MAPK8/JNK. This protein participates in the signal transduction induced by RANKL through the activation of RANK, which regulates the development and function of osteoclasts [38]. Thus, the differentially expressed miRNAs in this study may play a role in the development of AP by regulating osteoclast development and function.
Some studies have identified differentially expressed miRNAs in healthy and diseased human gingival tissue. Xie et al. [20] found that hsa-miR-126, hsa-miR-20a, hsa-miR-142-3p, hsa-miR-19-a, hsa-let-7f, hsa-miR-203, hsa-miR-17, hsa-miR-223, hsa-miR-146b, and hsa-miR-146a were upregulated more than 2-fold, and hsa-miR-155 and hsa-miR-205 were downregulated more than 2-fold in CP tissue compared to healthy gingiva using a microarray. Stoecklin-Wasmer et al. [21] found that that hsa-miR-451, hsa-miR-223, hsa-miR-486-5p, and hsa-miR-3917 were upregulated, and hsa-miR-1246, hsa-miR-1260, hsa-miR-141, hsa-miR-1260b, hsa-miR-203, hsa-miR-210, and hsa-miR-205 were downregulated more than 2-fold in CP tissue using a microarray. Ogata et al. [22] reported that hsa-miR-150, hsa-miR-223, and hsa-miR-200b were upregulated, and that hsa-miR-379, hsa-miR-199a-5p, and hsa-miR-214 were downregulated in CP tissue using a microarray. Lee et al. [23] showed that hsa-let-7a, hsa-let-7c, hsa-miR-130a, hsa-miR-301a, hsa-miR-520d, and hsa-miR-548a were upregulated more than 8-fold in CP tissue compared to healthy gingiva using a PCR array. Perri et al. [24] showed hsa-miR-30e and hsa-miR-106b were upregulated in periodontitis tissue. Finally, Na et al. [25] showed that has-miR-128, has-miR-34a, and has-miR-381 were upregulated, and that has-miR-15b, has-miR-211, hsa-miR-372 and has-miR-656 were downregulated in periodontitis tissue.
Although genetics play a more important role in AP than CP [30], only a few studies have demonstrated associations between miRNA expression and AP, mostly using gingival biopsies. Amaral et al. [31] compared the expression patterns of 754 miRNAs between CP and AP, finding that hsa-miR-1274b, hsa-let-7b-5p, hsa-miR-24-3p, hsa-miR-19b-3p, hsa-miR-720, hsa-miR-126-3p, hsa-miR-17-3p, and hsa-miR-21-3p were upregulated in both groups, suggesting that there were no significant differences in miRNA expression patterns between CP and AP. Ghotloo et al. [32] found an association between hsa-miR-146a and generalized aggressive periodontitis (GAP) and suggested that hsa-miR-146a may serve as an indicator of disease activity.
Recently, Fujimori et al. [29] performed the only previous study that utilized saliva samples as a non-invasive diagnostic tool for CP. They found that salivary hsa-miR-381-3p was upregulated in patients with severe periodontitis when compared to those with no or mild periodontitis and that it was positively correlated with the mean probing pocket depth.
The studies mentioned above tried to distinguish miRNA expression profiles between patients with periodontitis and healthy patients. However, their results are not substantially consistent with each other or with our findings. Among these studies, hsa-miR-223 [20,21] and hsa-miR-381 [25,29] were found to be upregulated in periodontitis in only 2 studies, respectively, while hsa-miR-205 [20,21] was found to be downregulated in periodontitis in only 2 studies. These inconsistencies among studies may result from variation in the inclusion criteria and differences in experimental designs. It is strongly recommended that genome-wide investigations using miRNAs in the future should be performed using standardized experimental designs and protocols.
The current pilot study has some limitations. The sample size of each group was small, so that the reliability of our study may be limited. The results of this study should be confirmed with a larger number of subjects in the future. The test group consisted of patients with GAP and localized aggressive periodontitis (LAP). In fact, 2 of the 4 test subjects were diagnosed with GAP, while the others were diagnosed with LAP. In patients with LAP, the miRNAs differentially expressed in AP in the whole saliva sample may be diluted by the miRNAs from healthy regions of the periodontium. The contents of the saliva samples might be affected by food or water intake, even though we instructed subjects not to consume any food or water for at least 1 hour prior to saliva collection. The effect of fasting time duration on salivary contents should be further investigated. Smoking history was not controlled, which might affect the results of this study. The correlation of smoking history and miRNA expression levels should be confirmed in the near future. Furthermore, few miRNA studies have been conducted in the field of dental research, so the databases utilized in this study (TAM 2.0, mirTarBase, and ConsensusPathDB) were biased towards medical terms, especially for pathological terms. In the future, dental research focusing on miRNAs should be performed, so that the databases can be gradually supplemented with dental terms. Finally, the results from saliva samples should be compared with those from other methods, such as gingival biopsies or gingival crevicular fluid, to show whether salivary diagnostics of AP could be a reliable method.
This study is the first analysis of miRNAs in the saliva of patients with AP. Identifying discriminatory human salivary miRNA biomarkers reflective of periodontal disease in a non-invasive screening assay is crucial for the development of salivary diagnostics. These data provide a first step towards the discovery of key salivary miRNA biomarkers for AP.
Footnotes
Funding: This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT and Future Planning (NRF-2013R1A1A2011974).
- Conceptualization: Won-Kyung Kim, Young-Kyoo Lee, Su-Hwan Kim.
- Formal analysis: Young-Sung Kim, Nam-Hun Lee.
- Investigation: Nam-Hun Lee, Eunhye Lee, Su-Hwan Kim.
- Methodology: Nam-Hun Lee, Eunhye Lee.
- Project administration: Eunhye Lee, Su-Hwan Kim.
- Writing - original draft: Nam-Hun Lee, Su-Hwan Kim.
- Writing - review & editing: Young-Sung Kim, Su-Hwan Kim.
Conflict of Interest: No potential conflict of interest relevant to this article was reported.
References
- 1.Huntzinger E, Izaurralde E. Gene silencing by microRNAs: contributions of translational repression and mRNA decay. Nat Rev Genet. 2011;12:99–110. doi: 10.1038/nrg2936. [DOI] [PubMed] [Google Scholar]
- 2.Pasquinelli AE. MicroRNAs and their targets: recognition, regulation and an emerging reciprocal relationship. Nat Rev Genet. 2012;13:271–282. doi: 10.1038/nrg3162. [DOI] [PubMed] [Google Scholar]
- 3.Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. doi: 10.1016/s0092-8674(04)00045-5. [DOI] [PubMed] [Google Scholar]
- 4.Selbach M, Schwanhäusser B, Thierfelder N, Fang Z, Khanin R, Rajewsky N. Widespread changes in protein synthesis induced by microRNAs. Nature. 2008;455:58–63. doi: 10.1038/nature07228. [DOI] [PubMed] [Google Scholar]
- 5.Irwandi RA, Vacharaksa A. The role of microRNA in periodontal tissue: a review of the literature. Arch Oral Biol. 2016;72:66–74. doi: 10.1016/j.archoralbio.2016.08.014. [DOI] [PubMed] [Google Scholar]
- 6.Kim SH, Lee SY, Lee YM, Lee YK. MicroRNAs as biomarkers for dental diseases. Singapore Dent J. 2015;36:18–22. doi: 10.1016/j.sdj.2015.09.001. [DOI] [PubMed] [Google Scholar]
- 7.Eulalio A, Schulte L, Vogel J. The mammalian microRNA response to bacterial infections. RNA Biol. 2012;9:742–750. doi: 10.4161/rna.20018. [DOI] [PubMed] [Google Scholar]
- 8.Pauley KM, Cha S, Chan EK. MicroRNA in autoimmunity and autoimmune diseases. J Autoimmun. 2009;32:189–194. doi: 10.1016/j.jaut.2009.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Alexander M, O’Connell RM. Noncoding RNAs and chronic inflammation: micro-managing the fire within. BioEssays. 2015;37:1005–1015. doi: 10.1002/bies.201500054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Romaine SP, Tomaszewski M, Condorelli G, Samani NJ. MicroRNAs in cardiovascular disease: an introduction for clinicians. Heart. 2015;101:921–928. doi: 10.1136/heartjnl-2013-305402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Peng Y, Croce CM. The role of MicroRNAs in human cancer. Signal Transduct Target Ther. 2016;1:15004. doi: 10.1038/sigtrans.2015.4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Luan X, Zhou X, Naqvi A, Francis M, Foyle D, Nares S, et al. MicroRNAs and immunity in periodontal health and disease. Int J Oral Sci. 2018;10:24. doi: 10.1038/s41368-018-0025-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Wang J, Chen J, Sen S. MicroRNA as biomarkers and diagnostics. J Cell Physiol. 2016;231:25–30. doi: 10.1002/jcp.25056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ai J, Zhang R, Li Y, Pu J, Lu Y, Jiao J, et al. Circulating microRNA-1 as a potential novel biomarker for acute myocardial infarction. Biochem Biophys Res Commun. 2010;391:73–77. doi: 10.1016/j.bbrc.2009.11.005. [DOI] [PubMed] [Google Scholar]
- 15.Mitchell PS, Parkin RK, Kroh EM, Fritz BR, Wyman SK, Pogosova-Agadjanyan EL, et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc Natl Acad Sci U S A. 2008;105:10513–10518. doi: 10.1073/pnas.0804549105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Xie Z, Yin X, Gong B, Nie W, Wu B, Zhang X, et al. Salivary microRNAs show potential as a noninvasive biomarker for detecting resectable pancreatic cancer. Cancer Prev Res (Phila) 2015;8:165–173. doi: 10.1158/1940-6207.CAPR-14-0192. [DOI] [PubMed] [Google Scholar]
- 17.Park NJ, Zhou H, Elashoff D, Henson BS, Kastratovic DA, Abemayor E, et al. Salivary microRNA: discovery, characterization, and clinical utility for oral cancer detection. Clin Cancer Res. 2009;15:5473–5477. doi: 10.1158/1078-0432.CCR-09-0736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Alevizos I, Alexander S, Turner RJ, Illei GG. MicroRNA expression profiles as biomarkers of minor salivary gland inflammation and dysfunction in Sjögren's syndrome. Arthritis Rheum. 2011;63:535–544. doi: 10.1002/art.30131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Xie ZJ, Chen G, Zhang XC, Li DF, Huang J, Li ZJ. Saliva supernatant miR-21: a novel potential biomarker for esophageal cancer detection. Asian Pac J Cancer Prev. 2012;13:6145–6149. doi: 10.7314/apjcp.2012.13.12.6145. [DOI] [PubMed] [Google Scholar]
- 20.Xie YF, Shu R, Jiang SY, Liu DL, Zhang XL. Comparison of microRNA profiles of human periodontal diseased and healthy gingival tissues. Int J Oral Sci. 2011;3:125–134. doi: 10.4248/IJOS11046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Stoecklin-Wasmer C, Guarnieri P, Celenti R, Demmer RT, Kebschull M, Papapanou PN. MicroRNAs and their target genes in gingival tissues. J Dent Res. 2012;91:934–940. doi: 10.1177/0022034512456551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ogata Y, Matsui S, Kato A, Zhou L, Nakayama Y, Takai H. MicroRNA expression in inflamed and noninflamed gingival tissues from Japanese patients. J Oral Sci. 2014;56:253–260. doi: 10.2334/josnusd.56.253. [DOI] [PubMed] [Google Scholar]
- 23.Lee YH, Na HS, Jeong SY, Jeong SH, Park HR, Chung J. Comparison of inflammatory microRNA expression in healthy and periodontitis tissues. Biocell. 2011;35:43–49. [PubMed] [Google Scholar]
- 24.Perri R, Nares S, Zhang S, Barros SP, Offenbacher S. MicroRNA modulation in obesity and periodontitis. J Dent Res. 2012;91:33–38. doi: 10.1177/0022034511425045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Na HS, Park MH, Song YR, Kim S, Kim HJ, Lee JY, et al. Elevated microRNA-128 in periodontitis mitigates tumor necrosis factor-α response via p38 signaling pathway in macrophages. J Periodontol. 2016;87:e173–e182. doi: 10.1902/jop.2016.160033. [DOI] [PubMed] [Google Scholar]
- 26.Barad O, Meiri E, Avniel A, Aharonov R, Barzilai A, Bentwich I, et al. MicroRNA expression detected by oligonucleotide microarrays: system establishment and expression profiling in human tissues. Genome Res. 2004;14:2486–2494. doi: 10.1101/gr.2845604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Chen Y, Gelfond JA, McManus LM, Shireman PK. Reproducibility of quantitative RT-PCR array in miRNA expression profiling and comparison with microarray analysis. BMC Genomics. 2009;10:407. doi: 10.1186/1471-2164-10-407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Mestdagh P, Feys T, Bernard N, Guenther S, Chen C, Speleman F, et al. High-throughput stem-loop RT-qPCR miRNA expression profiling using minute amounts of input RNA. Nucleic Acids Res. 2008;36:e143. doi: 10.1093/nar/gkn725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Fujimori K, Yoneda T, Tomofuji T, Ekuni D, Azuma T, Maruyama T, et al. Detection of salivary miRNAs reflecting chronic periodontitis: a pilot study. Molecules. 2019;24:1034. doi: 10.3390/molecules24061034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Van der Velden U. What exactly distinguishes aggressive from chronic periodontitis: is it mainly a difference in the degree of bacterial invasiveness? Periodontol 2000. 2017;75:24–44. doi: 10.1111/prd.12202. [DOI] [PubMed] [Google Scholar]
- 31.Amaral SA, Pereira TS, Brito JA, Cortelli SC, Cortelli JR, Gomez RS, et al. Comparison of miRNA expression profiles in individuals with chronic or aggressive periodontitis. Oral Dis. 2019;25:561–568. doi: 10.1111/odi.12994. [DOI] [PubMed] [Google Scholar]
- 32.Ghotloo S, Motedayyen H, Amani D, Saffari M, Sattari M. Assessment of microRNA-146a in generalized aggressive periodontitis and its association with disease severity. J Periodontal Res. 2019;54:27–32. doi: 10.1111/jre.12538. [DOI] [PubMed] [Google Scholar]
- 33.Lang N, Bartold PM, Cullinan M, Jeffcoat M, Mombelli A, Murakami S, et al. Consensus report: aggressive periodontitis. Ann Periodontol. 1999;4:53. [Google Scholar]
- 34.Yoshizawa JM, Wong DT. Salivary microRNAs and oral cancer detection. Methods Mol Biol. 2013;936:313–324. doi: 10.1007/978-1-62703-083-0_24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Li J, Han X, Wan Y, Zhang S, Zhao Y, Fan R, et al. TAM 2.0: tool for MicroRNA set analysis. Nucleic Acids Res. 2018;46:W180–W185. doi: 10.1093/nar/gky509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Chou CH, Shrestha S, Yang CD, Chang NW, Lin YL, Liao KW, et al. miRTarBase update 2018: a resource for experimentally validated microRNA-target interactions. Nucleic Acids Res. 2018;46:D296–D302. doi: 10.1093/nar/gkx1067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kamburov A, Stelzl U, Lehrach H, Herwig R. The ConsensusPathDB interaction database: 2013 update. Nucleic Acids Res. 2013;41:D793–D800. doi: 10.1093/nar/gks1055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.TAB2: TGF-beta activated kinase 1 (MAP3K7) binding protein 2 [Homo sapiens (human)] [Internet] Bethesda (MD): National Center for Biotechnology Information; 2019. [cited 2019 Sep 1]. Available from: https://www.ncbi.nlm.nih.gov/gene/23118. [Google Scholar]