

Abstract
Strategies to personalize psychopharmacological treatment promise to improve efficacy and tolerability. We measured serotonin transporter occupancy immediately after infusion of the widely prescribed P-glycoprotein substrate citalopram and assessed to what extent variants of the ABCB1 gene affect drug target engagement in the brain in vivo. A total of 79 participants (39 female) including 31 patients with major depression and 48 healthy volunteers underwent two PET/MRI scans with the tracer [11C]DASB and placebo-controlled infusion of citalopram (8 mg) in a cross-over design. We tested the effect of six ABCB1 single nucleotide polymorphisms and found lower SERT occupancy in ABCB1 rs2235015 minor allele carriers (n = 26, MAF = 0.18) compared to major allele homozygotes (t73 = 2.73, pFWE < 0.05) as well as in men compared to women (t73 = 3.33, pFWE < 0.05). These effects were robust to correction for citalopram plasma concentration, age and diagnosis. From occupancy we derived the ratio of occupied to unoccupied SERT, because in theory this measure is equal to the product of drug affinity and concentration at target sites. A model combining genotype with basic clinical variables, predicted that, at the same dosage, occupied to unoccupied SERT ratio was −14.48 ± 5.38% lower in rs2235015 minor allele carriers, +19.10 ± 6.95% higher in women, −4.83 ± 2.70% lower per 10 kg bodyweight, and −2.68 ± 3.07% lower per 10 years of age. Our results support the exploration of clinical algorithms with adjustment of initial citalopram dosing and highlight the potential of imaging-genetics for precision pharmacotherapy in psychiatry.
Subject terms: Predictive markers, Neuroscience
Introduction
Non-response to pharmacotherapy is a major challenge in the treatment of major depressive disorder (MDD). While roughly 30% percent of patients achieve remission, about 60% do not respond sufficiently to first-line treatment with selective serotonin reuptake inhibitors (SSRIs) [1, 2]. Moreover, a considerable number of patients experience adverse effects. Thus, most patients require modifications of their initial treatment [3]. Large-scale collaborative research projects pursue the characterization of predictors for treatment resistance that may guide initial treatment strategies [2]. Therapeutic drug monitoring can inform dose adaptations to achieve drug concentrations in blood associated with the highest possible probability of response and the lowest possible risk of adverse events. This approach may be supplemented by pharmacogenetic testing, such as determination of cytochrome P450 (CYP) genotype and consideration of basic pharmacokinetic variables [4]. While the cost-effectiveness of this approach has been demonstrated previously [5], predictive markers of individual response are highly anticipated to accelerate remission.
Binding of SSRIs to their molecular target, the serotonin transporter (SERT), correlates with their concentration in the brain and antidepressant efficacy may be associated therewith [6–8]. Next to plasma concentration, active efflux transport at the blood-brain barrier (BBB) might influence availability of antidepressants at target sites [9, 10]. While drug metabolism and basic pharmacokinetic variables directly affect plasma concentration, BBB permeability has been demonstrated to modify the association between plasma levels and cerebral concentration [11]. P-glycoprotein (P-gp), encoded by the ATP-binding cassette transporter B1 (ABCB1) gene, represents one of the principal efflux mechanisms at the BBB protecting the brain against potentially toxic xenobiotics [12, 13]. Among other pharmaceuticals, several psychotropic substances including SSRIs were shown to be substrates of the P-gp [10, 14]. (Es)citalopram is the most widely used SSRI [15, 16] and one of the strongest P-gp substrates within this drug class. Preclinical studies demonstrated 1.9–3.7 times higher citalopram brain concentrations in P-gp knockout mice after acute and chronic administration [17–21]. Therefore, genotyping of ABCB1 holds promise as a predictive pharmacogenetic marker of antidepressant response based on the hypothesis that genetic variants might affect P-gp expression or function, and thus antidepressant concentration in the brain [9, 22]. Uhr et al. were the first to demonstrate an association between ABCB1 genotype and clinical efficacy of antidepressants in MDD [18]. However, results of consecutive studies are equivocal and interpretation is hampered by methodological heterogeneity [23–35]. Inconsistencies may be explained by substrate specific effects of variants, disorder heterogeneity and non-uniform outcome definition. Besides clinical efficacy, a large body of evidence suggests that ABCB1 genotype is associated with tolerability of antidepressants based on the assumption of genotype dependent alterations in cerebral drug accumulation (for review, see Brückl et al. [9]). Moreover, implications of ABCB1 genotype in the treatment of other neuropsychiatric disorders, such as schizophrenia [36–38] and epilepsy [39] highlight its role whenever P-gp substrates are administered.
Nevertheless, progress in the determination of the utility of ABCB1 genotyping by means of clinical trials is hampered by the need for large sample sizes and standardized treatment protocols. Adding to this, in light of the inherent heterogeneity of psychiatric disorders and the diversity of outcomes to define efficacy, the prospects of detecting and confirming relevant variants for each disorder and intervention appear distant. However, under the assumption that ABCB1 variants affect cerebral drug availability, measurement of target engagement in the brain could be exploited as an intermediate phenotype to identify pharmacogenetic variants with trans-diagnostic relevance. Positron emission tomography (PET) enables the in vivo quantification of SERT occupancy, i.e., the proportion of transporter sites blocked by medication and, thus, may provide an indirect measure of cerebral drug availability. In this imaging-pharmacogenetics study, we aimed to assess the impact of ABCB1 genotype on SERT occupancy by citalopram as a proxy of intracerebral drug availability in a large sample of healthy controls and patients diagnosed with MDD using gold standard SERT quantification procedures. Acute pharmacological challenge with intravenous citalopram was used to allow for the most direct assessment of the impact of efflux transport at the BBB on cerebral drug availability, because significant drug metabolism, first-pass effects, adaptations, and ceiling effects after prolonged therapy could be ruled out. This enabled the identification of variables with the potential to aid antidepressant dose adjustment and in this way contribute to precision pharmacotherapy in psychiatry.
Methods
Participants and study design
Thirty-one patients with MDD (age ± SD: 29.0 ± 9.0) and 48 healthy volunteers (age ± SD: 28.0 ± 8.7, see Table 1) were included in these analyses. Participants underwent two PET/MR scans with the radioligand [11C]-N,N-dimethyl-2-(2-amino-4-cyanophenylthio)-benzylamine ([11C]DASB) during which double-blind intravenous pharmacological challenge with the SSRI citalopram (8 mg) or placebo was performed in a randomized cross-over study design (see Figure S1). Scanning procedures were performed using established protocols as detailed in the supplement [40]. In short, [11C]DASB was applied as bolus plus constant infusion according to a protocol designed to rapidly attain equilibrium in high-binding regions [41]. Good agreement with conventional bolus application protocols was demonstrated using this approach [42]. Drug or placebo infusion was performed over 8 min starting 70 min after initiation of radioligand application. Occurrence and severity of frequently observed side-effects to SSRIs were assessed during scans. Arterial blood samples were drawn throughout the measurement and assessment of the concentration of [11C]DASB, radioactive metabolites and citalopram in plasma was performed [41, 43]. Time activity curves and metabolite corrected plasma activity are displayed in Fig. 1.
Table 1.
Demographics, clinical characteristics, pharmacokinetic and imaging parameters of study participants are shown.
| Diagnosis | ABCB1rs2235015 | |||||
|---|---|---|---|---|---|---|
| Controls | MDD | p | A + AC | C | p | |
| Group size | 48 | 31 | 26 | 53 | ||
| Age (y) | 28.0 ± 8.7 | 29.0 ± 9.0 | 0.61a | 29.0 ± 9.7 | 28.1 ± 8.3 | 0.69a |
| Sex (f/m) | 24/24 | 15/16 | 1b | 11/15 | 28/25 | 0.52b |
| Bodyweight (kg) | 70.3 ± 12.0 | 68.8 ± 13.2 | 0.63a | 70.1 ± 11.9 | 69.5 ± 12.8 | 0.83a |
| Citalopram AUC ((mg/ml) x sec) | 94.4 ± 19.6 | 85.2 ± 21.0 | 0.06a | 89.4 ± 20.1 | 91.5 ± 20.9 | 0.66a |
| HRSD | – | 22.7 ± 5.1 | – | – | ||
| BDI | – | 28.9 ± 8.2 | – | – | ||
| CGI | – | 4.7 ± 0.7 | – | – | ||
| Placebo SERT BPP (thalamus) | 23.3 ± 4.2 | 24.2 ± 4.1 | 0.39a | 23.5 ± 4.7 | 23.8 ± 3.9 | 0.78a |
| SERT occupancy (%) | 67.3 ± 7.1 | 66.8 ± 4.9 | 0.75a | 64.5 ± 6.4 | 68.4 ± 5.9 | 0.01a |
| O/U SERT | 2.1 ± 0.7 | 2.1 ± 0.5 | 0.41a | 1.9 ± 0.5 | 2.3 ± 0.6 | <0.01a |
Participants were grouped according to diagnosis (Controls/MDD) and ABCB1rs2235015 genotype (A + AC/C). Groups did not differ in terms of age, sex, bodyweight, AUC and imaging parameters. Minor allele carriers and major allele homozygotes did not differ in age, sex, bodyweight, AUC and baseline SERT binding potential. Significantly lower SERT occupancy was revealed in minor allele carriers when compared to major allele homozygotes.
MDD major depressive disorder, A + AC minor allele carriers, C major allele homozygotes, HRSD Hamilton Rating Scale for Depression, BDI Beck Depression Inventory, CGI Clinical Global Impression, SERT serotonin transporter, BPP binding potential.
aIndependent two-sample t-test.
bChi-square test.
Fig. 1. Quantification of serotonin transporter (SERT) binding potential (BPP).
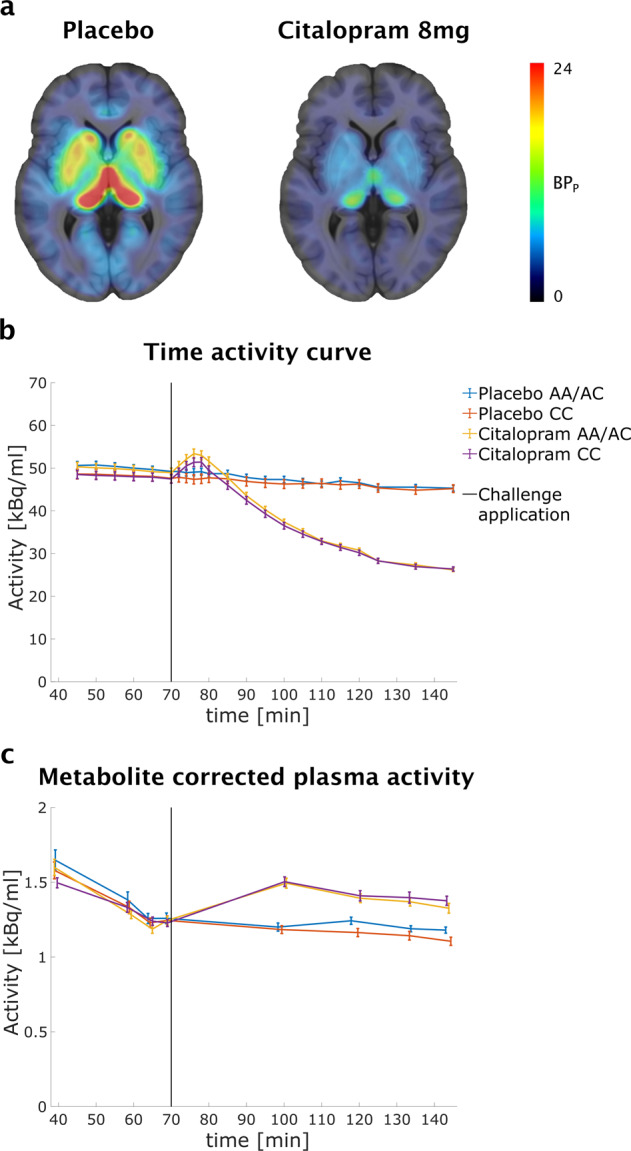
a Average SERT BPP from 79 participants included in this study are displayed in transversal planes overlaid on a MR template for placebo (left) and citalopram (right) scans. High baseline binding was observed in regions rich in SERT such as thalamus and striatum. Time activity curves (b) for the thalamus and metabolite corrected plasma activity (c) ±SE for PET/MR scans with citalopram and placebo challenge are plotted for groups defined by ABCB1rs2235015 genotype.
After completion of scanning procedures, antidepressant treatment with the SSRI escitalopram 10 mg (Cipralex, Lundbeck) was initiated in patients. Details on recruitment, inclusion criteria, clinical management and follow-up are outlined in the supplement.
Genotyping and selection of single nucleotide polymorphisms (SNPs)
Eight ABCB1 SNPs were considered for genotyping based on previously reported association with antidepressant treatment response to P-gp substrates or on association studies with ABCB1 expression [18, 24, 27, 28, 30, 44–50]. Genotyping procedures are described in detail in the supplement [51, 52]. Genotype frequencies were found to be distributed according to the Hardy–Weinberg equilibrium (see Table 2). The following six tag SNPs (mean r2 = 0.97) were selected and used for further statistical analyses: rs1128503, rs2235015, rs10245483, rs28373093, rs2032583, rs1045642 (see Fig. S2). Participants were grouped into minor allele carriers and major allele homozygotes for all statistical analyses.
Table 2.
Position of SNPs, minor allele frequency (MAF) and p value of Hardy–Weinberg statistics.
| Polymorphism | Position | Minor allele (Major Allele) | MAF | Hardy–Weinberg p value |
|---|---|---|---|---|
| rs1045642 | 87509329 | G(a) | 0.48 | 0.52 |
| rs2032583 | 87531245 | G(a) | 0.14 | 0.92 |
| rs2032582 | 87531302 | A(c) | 0.39 | 0.81 |
| rs2235033 | 87549827 | A(g) | 0.40 | 0.58 |
| rs1128503 | 87550285 | A(g) | 0.38 | 0.99 |
| rs2235015 | 87570248 | A(c) | 0.18 | 0.90 |
| rs28373093 | 87714707 | C(g) | 0.42 | 0.46 |
| rs10245483 | 90008294 | G(t) | 0.49 | 0.78 |
Effect of ABCB1 variants on SERT occupancy and clinical response
The thalamus was chosen for quantification of occupancy because our tracer application protocol is optimized for rapid equilibration in this region with high SERT expression [41]. Binding potentials (BPP) were obtained by subtracting the distribution volume of the reference region (cerebellar gray matter) from the thalamic distribution volume. SERT occupancy (ΔBPP) was calculated as the relative decrease in binding potentials between drug and placebo scans (see Fig. 1). R (v4.0.2, https://www.R-project.org/) was used for statistical analyses. To assess if ABCB1 genotype affects SERT occupancy, multiple linear regression models were built for each tag SNP with participant’s age, sex, citalopram under the curve (AUC) in plasma and diagnosis as covariates. Age, sex and AUC were included based on pharmacokinetic assumptions, prior evidence of higher plasma levels in females and elderly patients and the association between SERT occupancy and SSRI plasma levels [11, 53–55]. Correction for family wise error (FWE) was performed using the Bonferroni method for six SNPs at alpha = 0.05. Furthermore, we performed mediation analyses using the PROCESS procedure to assess if effects of variables on SERT occupancy were mediated by citalopram plasma AUC. In an exploratory analysis, the effect of significant ABCB1 variants and sex on treatment response (relative decrease in HDRS scores after six weeks of escitalopram treatment) was probed using independent t-tests. Pearson’s correlation coefficients were used to assess the association between SERT occupancy and treatment response. Moreover, we assessed the effect of genotype, sex and SERT occupancy on side-effect scores obtained during scans.
Modeling SERT binding based on clinical variables and ABCB1 genotype
While SERT occupancy (ΔBPP) facilitates comparison of our results with published findings, it asymptotically approaches 100% with increasing concentration, which hampers linear regression modeling and applicability of resulting models beyond the range covered by the data. Therefore, we calculated the ratio of occupied to unoccupied (O/U) SERT, which is directly derived from ΔBPP for further analyses (Fig. S3), hereafter referred to as occupied SERT ratio or O/U SERT:
| 1 |
This alternative measure of drug target engagement can be used to approximate the product of drug affinity (1/Ki) and free drug concentration at target sites ([I]) assuming ideal conditions as approached in vitro using equilibrium binding models and disregarding occupancy by endogenous ligands:
| 2 |
[SERT] and [SERT.I] signify the concentrations of free SERT and SERT-inhibitor complexes, respectively. The proportionality of O/U SERT to [I]/Ki enables modeling of linear effects of variables affecting SERT binding via changes in cerebral drug availability or affinity. A multiple linear regression model including weight, sex and age as predictors and occupied SERT ratio as the dependent variable was built. These basic parameters affecting pharmacokinetics are readily available and can be taken into account in clinical practice at the initiation of treatment. This model was compared to a model containing significant ABCB1 variants using analysis of variance and the Akaike information criterion (AIC). For comparison, models calculated using SERT occupancy are included in the supplement.
Results
Effect of ABCB1 variants on SERT occupancy and clinical response
Demographics, clinical characteristics, pharmacokinetic and imaging parameters are listed in Table 1. Average SERT BPP for placebo and citalopram condition are displayed in Fig. 1a. Regression analyses revealed a significant effect of rs2235015 on SERT occupancy (t73 = 2.73, pFWE < 0.05) with lower occupancy in minor allele carriers compared to major allele homozygotes (Fig. 2a). This effect was robust to correction for citalopram plasma AUC (Fig. 2b), sex, age and diagnosis. Moreover, we detected a significant effect of sex with higher SERT occupancy in female compared to male participants (t73 = 3.33, pFWE < 0.05). Notably, SERT BPP was higher in female compared to male participants in our sample (mean ± SD: 25.56 ± 4.19 vs 21.80 ± 3.29). While the effect of rs2235015 was largely unaffected (t72 = 2.84, pFWE < 0.05), the effect of sex on SERT occupancy was markedly decreased when additionally including placebo BPP as a covariate (t72 = 2.03, p < 0.05, uncorrected). Results for non-significant SNPs are provided in the supplement (Table S1). Mediation analysis indicated a direct effect (b = −0.16, SE = 0.07) but no indirect effect of age mediated by citalopram AUC in plasma on SERT occupancy (indirect: −0.05, SE = 0.04). However, we found direct and indirect effects mediated by citalopram AUC in plasma of sex on SERT occupancy (direct: b = 5.47, SE = 1.33; indirect: b = 1.45, SE = 0.70). No significant effect of weight on SERT occupancy was detected after sex-wise mean centering.
Fig. 2. The effect of genotype, sex and citalopram plasma concentrtion on serotonin transporter occupancy.

a Approximately one hour after infusion of 8 mg citalopram, lower serotonin transporter occupancy was observed in ABCB1rs2235015 minor allele carriers (A + AC) compared to major allele homozygotes (C) and female compared to male participants. b SERT occupancy is plotted against the area under the curve (AUC) of citalopram in plasma. Regression lines are plotted for participants grouped according to ABCB1rs2235015 genotype. Grey ribbons indicate 95% confidence intervals.
In 25 patients with follow-up data, we observed a response rate of 68% (≥50% reduction in HDRS score) after six weeks of treatment with escitalopram. Antidepressant treatment response to escitalopram did not differ significantly between ABCB1rs2235015 minor allele carriers and major allele homozygotes. No association between sex and antidepressant response was revealed. Furthermore, we did not detect a significant correlation between SERT occupancy and antidepressant treatment response. Lastly, there was no statistically significant effect of ABCB1rs2235015 genotype, sex or SERT occupancy on side-effects measured during scans, which were not different between placebo and citalopram 8 mg (see supplement for further details).
Modeling SERT binding based on clinical variables and ABCB1 genotype
In order to provide a model for estimating the effect of different clinical variables and ABCB1 genotype on SERT binding, we transformed SERT occupancy into the ratio of occupied to unoccupied SERT (O/U SERT) which is proportional to the product of drug affinity and concentration at target sites. Effects are reported as percentages of the sample’s mean O/U SERT to improve interpretability. Adding rs2235015 to a model for prediction of O/U SERT using a combination of the basic clinical variables sex, age and weight improved the fit to the data (adjusted R2 = 0.35 vs. 0.30) with a lower AIC (722.98 vs 728.01). This model predicted that, at the same dosage, occupied SERT ratio was −14.48 ± 5.38% (SE) lower in rs2235015 minor allele carriers, +19.10 ± 6.95% higher in women, −4.83 ± 2.70% lower per 10 kg bodyweight, and −2.68 ± 3.07% lower per 10 years of age. Based on this model, predictions of occupied SERT ratio and SERT occupancy across different combinations of rs2235015 genotype, sex, weight and age are illustrated in Fig. 3. As an example, when comparing a 20-year-old female patient genotyped as rs2235015 major allele homozygote with a bodyweight of 50 kg with a 40-year-old rs2235015 male minor allele carrier with a bodyweight of 100 kg, O/U SERT in the female patient can be expected to be twice as high as in the male patient.
Fig. 3. Modeling SERT binding based on clinical variables and ABCB1 genotype.

a The ratio of occupied to unoccupied (O/U) SERT after citalopram infusion was predicted for different combinations of sex, ABCB1rs2235015 genotype, age and weight based on a linear regression model fitted to the study data. O/U SERT is directly proportional to the product of drug affinity and concentration at target sites in equilibrium and is reported as a percentage of the sample’s average to aid interpretation of effects. O/U SERT was predicted to be −14.48 ± 5.38% lower in rs2235015 minor allele carriers, +19.10 ± 6.95% higher in women, −4.83 ± 2.70% lower per 10 kg bodyweight, and −2.68 ± 3.07% lower per 10 years of age. b Predictions of O/U SERT were transformed into SERT occupancy after citalopram 8 mg infusion based on equation 1 after multiplication with the sample’s mean O/U SERT.
Discussion
Research on the impact of ABCB1 gene variants on in vivo brain concentration of psychopharmaceuticals is methodologically challenging and evidence is currently limited. Molecular neuroimaging studies investigating target occupancy may be used to indirectly assess the impact of patient characteristics on drug availability in the brain if drug affinity is not correlated with the variables of interest. Recently, Simoons et al. demonstrated significantly higher paroxetine SERT occupancy at the same plasma drug concentration after six weeks of therapy in ABCB1 rs1128503 and rs2032582 minor allele carriers using [123I]β-CIT SPECT [11]. No association between SERT occupancy and rs1128503 genotype was found in our study. Increases in dopamine transporter binding following chronic paroxetine administration as shown in prior studies [56, 57] may hinder interpretation of results of the SPECT study considering non-selectivity of the applied radiotracer. Acute pharmacological challenge during PET measurements allows to neglect drug metabolism, first-pass and secondary effects, such as SERT up- and downregulation. Moreover, paroxetine applied in the SPECT study was demonstrated to be a relatively weak substrate of the P-gp while citalopram is one of the strongest P-gp substrates among SSRIs [58].
The intronic SNP rs2235015 is among the most frequently studied ABCB1 SNPs in the context of antidepressant treatment [9]. Two studies support an association of rs2235015 with antidepressant response [18, 59]. Although these findings are in contrast to our results, the power for the investigation of the genetic impact on antidepressant response was limited due to the relatively high response rate and small sample with clinical follow-up. In the study by Uhr et al. rs2235015 minor allele carriers were more likely to remit after a six-week treatment trial with a P-gp substrate when compared to non-carriers [18]. Under the hypothesis of a direct relationship between target tissue drug concentration and treatment response, this finding is at odds with the results of our study that demonstrates lower SERT occupancy in minor allele carriers. This discrepancy may be interpreted in light of heterogeneous antidepressant treatment in the study by Uhr et al. and previously reported substrate-specific differences in the direction of effects of SNPs [30]. Moreover, direct comparisons of our results with those obtained after chronic oral drug administration neglects the influence of ABCB1 variants on intestinal drug absorption [60]. Breitenstein et al. did not detect an association of rs2235015 genotype on treatment response but a significant genotype x plasma level interaction [59]. Interestingly, minor allele carriers had numerically higher antidepressant plasma concentrations when compared to non-carriers. An interpretation that might reconcile these findings with our results is that P-gp has a higher affinity for citalopram in rs2235015 minor allele carriers. This might result in higher efflux of citalopram at low concentrations, as observed after acute challenge of a low dose in our study, but lower efflux at high concentrations during chronic treatment due to a lower transport capacity of P-gp.
We detected a significant effect of sex on SERT occupancy. While previous studies did not assess sex specific differences in cerebral drug concentration, this finding is in line with known pharmacokinetic differences during antidepressant treatment between men and women [61–63]. Lower body weight and blood volume may account for higher plasma drug levels and, subsequently, higher cerebral concentration in women [63]. Mediation analysis indicated that a part of sex differences in SERT binding was due to differences in citalopram AUC in plasma. The effect of plasma concentration might be underestimated by using AUC, but equilibrium citalopram concentration in plasma was in many cases below the sensitivity of our quantification method. While gender-specific effects on CYP2C19 activity, the primarily responsible enzyme for degradation of citalopram, were previously reported, drug metabolism following intravenous administration can be neglected within our measurement timeframe. Sex differences in SERT occupancy were also associated with differences in placebo BP in our study. Although our results are in line with previously reported sex differences in SERT binding, more recent findings are equivocal [64, 65]. Higher BP in women in our study might have resulted in higher measures of occupancy due to regression to the mean [66]. The persistence of sex effects to correction for BP, indicates that there were sex differences in either cerebral citalopram concentration or SERT affinity. In line with our results, lower P-gp function was demonstrated in young women when compared to young men in a recent (R)-[11C]verapamil PET study [67].
Our findings may guide individual antidepressant dosing based on sex and ABCB1 genotype. While several studies suggest a dose–response relationship, higher doses come with the cost of reduced tolerability [68]. Our results may reassure patients and clinicians that the practice of lower starting doses of citalopram and escitalopram to avoid initial side effects need not to come at the cost of SERT occupancy when performed in a targeted manner taking into account basic clinical variables and genotype. While the effects of dose adjustments need to be investigated in further clinical trials, this approach holds promise to reduce the rate of initial side effects and, thus, increase adherence during SSRI treatment. Our model employing occupied SERT ratio can be directly applied as a guidance in this process if assuming a linear relationship between drug dosage and concentration at target sites [69, 70]. As described above, O/U SERT ratio is expected to be directly proportional to the product of drug affinity and concentration. Furthermore, considering that citalopram is a competitive inhibitor of SERT, its effect on serotonin (5-HT) uptake in vitro is described by the following Michaelis-Menten equation (Km, Michaelis constant):
| 3 |
This illustrates that the 5-HT concentration at which the half maximal velocity of SERT uptake (V) relative to the maximum rate (Vmax) is achieved is increased by a multiple of [I]/Ki in the presence of an inhibitor (I), which underlines the utility of O/U SERT as an outcome measure.
While our results may be applicable to drug dosing during the initiation of treatment, the effects of clinical variables and P-gp variants during prolonged treatment need to be established. For example, variants that lead to a higher affinity of P-gp and increased clearance of drug from the brain at low concentrations may lead to P-gp saturation at higher concentrations and thus inversely affect concentration in the brain during different phases of treatment [71].
Strengths of this investigation include the use of state-of-the-art PET imaging procedures that allow for highly specific and reliable SERT quantification in the context of a randomized, placebo-controlled trial. Participants were free from psychopharmacological medication, which allows for the investigation SERT occupancy in the absence of interactions. A relatively large sample of 79 participants was enrolled in this study which is seldom attained in resource intensive PET studies. Assessment of SERT occupancy following intravenous citalopram challenge allows for probing the genetic influence on active efflux transport at the BBB unaffected by drug metabolism and receptor up- and downregulation as observed during chronic treatment and, thus, is mainly dependent on cerebral drug availability and affinity [57].
Clinical implications of our findings remain to be determined in clinical trials with larger sample sizes. The assessed SNPs in our study were common (MAF ≥ 0.05), however, investigation of minor allele homozygotes could not be performed as group sizes did not provide sufficient statistical power. Generalizability of our results along a broader age spectrum need to be ascertained as mainly young participants were investigated in this trial. We observed differences in the arterial input function between genotypes. Based on the available literature we cannot provide a conclusive explanation for this phenomenon. However, preclinical data suggests that [11C]DASB does not interact with the P-gp at the BBB and, thus, no interferences with quantification of citalopram SERT occupancy are expected [72]. Nevertheless, possible interactions of [11C]DASB with the P-gp may be considered a limitation as in vivo data is currently lacking.
In conclusion, we demonstrate significant differences in SERT occupancy associated with ABCB1 genotype and sex. Our study supports the further exploration of pharmacogenetic testing for ABCB1 genotype during SSRI treatment. These results suggest that initial dosing of SSRI may be modified a priori based on patient characteristics and genotype, thus underscoring the potential of precision pharmacotherapy in psychiatry. While genetic testing of the ABCB1 gene may be restricted to specialized centers, dose-adjustments based on sex could be performed easily in clinical practice.
Supplementary information
Acknowledgements
We thank Markus Hartenbach, Tatjana Traub-Weidinger, Edda Winkler-Pjrek, Dietmar Winkler, Marius Hienert, Thomas Vanicek, Alexander Kautzky, Johannes Jungwirth, Paul Michenthaler, and Alim Emre Başaran for their medical support and Georg Kranz for his assistance in randomization of participants. We are grateful to Karoline Einenkel, Elisa Sittenberger and Vera Ritter for their administrative support and Gregory M. James, Murray B. Reed and Matej Murgas for technical support. We gratefully acknowledge the support of Verena Pichler, Neydher Berroterán-Infante, Theresa Balber, Markus Mitterhauser and Eva-Maria Klebermass in tracer synthesis and metabolite analysis.
Author contributions
SK and RL served as principal investigators, GG and RL designed the study, GG and LRS performed statistical analyses and drafted the first versions of the manuscript. GG, LRS, JU, GMG, DP carried out recruitment and clinical management of participants. MK and LR performed PET/MRI scans and data processing under supervision of AH. CV, CP, LN performed radiosynthesis and analysis of radioactive metabolites under supervision of WW. Medical supervision of the study was provided by SK and MH. TS performed analysis of plasma levels of citalopram. AMH and DR performed genetic analyses. All authors revised the manuscript and approved the contents.
Funding information
This research was funded in part by the Austrian Science Fund (FWF) [KLI 504] and by a grant from the Else Kröner-Fresenius-Stiftung (2014_A192) to Rupert Lanzenberger. For the purpose of open access, the author has applied a CC BY public copyright license to any Author Accepted Manuscript version arising from this submission. Icons in Fig. 3 were created with BioRender.com. Further, this study was supported partly through a research agreement between the Medical University of Vienna and the Siemens Healthcare GmbH. Leo R. Silberbauer, Gregor Gryglewski, Lucas Rischka, and Manfred Klöbl were recipients of DOC fellowships of the Austrian Academy of Sciences at the Department of Psychiatry and Psychotherapy, Medical University of Vienna. Daniel Pacher was supported by the MDPhD Excellence Program of the Medical University of Vienna. This scientific project was performed with the support of the Medical Imaging Cluster of the Medical University of Vienna.
Competing interests
RL received travel grants and/or conference speaker honoraria from Bruker BioSpin within the last three years and investigator-initiated research funding from Siemens Healthcare regarding clinical research using PET/MR. He is a shareholder of the start-up company BM Health GmbH since 2019. SK received grants/ research support, consulting fees and/or honoraria within the last three years. He received grant/research support from Lundbeck; he has served as a consultant or on advisory boards for Celegne, IQVIA, Janssen, Lundbeck, Mundipharma, Recordati, Takeda and Schwabe; and he has served on speaker bureaus for Angelini, Aspen Farmaceutica S.A., Janssen, Krka Pharma, Lundbeck, Medichem Pharmaceuticals Inc., Neuraxpharma, OM Pharma, Pierre Fabre, Sanofi, Servier, Schwabe, Sun Pharma. Without relevance to this work, WW is a part-time employee of CBmed GmbH, Graz, Austria and a co-founder/co-owner of MINUTE medical GmbH, Vienna, Austria. Within the last years, he received research grants from ITM, Garching, Germany, and Scintomics, Fürstenfeldbruck, Germany. The remaining authors declare no potential conflict of interest with respect to the research, authorship, and/ or publication of this article.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
The online version contains supplementary material available at 10.1038/s41380-022-01733-1.
References
- 1.Bauer M, Pfennig A, Severus E, Whybrow PC, Angst J, Möller H-J. World Federation of Societies of Biological Psychiatry (WFSBP) Guidelines for Biological Treatment of Unipolar Depressive Disorders, Part 1: Update 2013 on the acute and continuation treatment of unipolar depressive disorders. World J Biol Psychiatry. 2013;14:334–85. doi: 10.3109/15622975.2013.804195. [DOI] [PubMed] [Google Scholar]
- 2.Bartova L, Dold M, Kautzky A, Fabbri C, Spies M, Serretti A, et al. Results of the European Group for the Study of Resistant Depression (GSRD) - basis for further research and clinical practice. World J Biol Psychiatry. 2019;20:427–48. doi: 10.1080/15622975.2019.1635270. [DOI] [PubMed] [Google Scholar]
- 3.Fava M. Diagnosis and definition of treatment-resistant depression. Biol Psychiatry. 2003;53:649–59. doi: 10.1016/s0006-3223(03)00231-2. [DOI] [PubMed] [Google Scholar]
- 4.Hiemke C, Bergemann N, Clement HW, Conca A, Deckert J, Domschke K, et al. Consensus guidelines for therapeutic drug monitoring in neuropsychopharmacology: update 2017. Pharmacopsychiatry. 2018;51:9–62. doi: 10.1055/s-0043-116492. [DOI] [PubMed] [Google Scholar]
- 5.Touw DJ, Neef C, Thomson AH, Vinks AA. Cost-effectiveness of therapeutic drug monitoring: a systematic review. Ther Drug Monit. 2005;27:10–17. doi: 10.1097/00007691-200502000-00004. [DOI] [PubMed] [Google Scholar]
- 6.Liu X, Vilenski O, Kwan J, Apparsundaram S, Weikert R. Unbound brain concentration determines receptor occupancy: a correlation of drug concentration and brain serotonin and dopamine reuptake transporter occupancy for eighteen compounds in rats. Drug Metab Dispos. 2009;37:1548–56. doi: 10.1124/dmd.109.026674. [DOI] [PubMed] [Google Scholar]
- 7.Bundgaard C, Sveigaard C, Brennum LT, Stensbøl TB. Associating in vitro target binding and in vivo CNS occupancy of serotonin reuptake inhibitors in rats: the role of free drug concentrations. Xenobiotica. 2012;42:256–65. doi: 10.3109/00498254.2011.618953. [DOI] [PubMed] [Google Scholar]
- 8.Henry ME, Moore CM, Kaufman MJ, Michelson D, Schmidt ME, Stoddard E, et al. Brain kinetics of paroxetine and fluoxetine on the third day of placebo substitution: a fluorine MRS study. Am J Psychiatry. 2000;157:1506–8. doi: 10.1176/appi.ajp.157.9.1506. [DOI] [PubMed] [Google Scholar]
- 9.Brückl TM, Uhr M. ABCB1 genotyping in the treatment of depression. Pharmacogenomics. 2016;17:2039–69. doi: 10.2217/pgs.16.18. [DOI] [PubMed] [Google Scholar]
- 10.O’Brien FE, Dinan TG, Griffin BT, Cryan JF. Interactions between antidepressants and P-glycoprotein at the blood-brain barrier: clinical significance of in vitro and in vivo findings. Br J Pharmacol. 2012;165:289–312. doi: 10.1111/j.1476-5381.2011.01557.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Simoons M, Mulder H, Appeldoorn JTY, Risselada AJ, Schene AH, van Schaik RHN, et al. Modification of the association between paroxetine serum concentration and SERT-occupancy by ABCB1 (P-glycoprotein) polymorphisms in major depressive disorder. Psychiatr Genet. 2020;30:19–29. doi: 10.1097/YPG.0000000000000244. [DOI] [PubMed] [Google Scholar]
- 12.Schinkel AH. P-Glycoprotein, a gatekeeper in the blood-brain barrier. Adv drug Deliv Rev. 1999;36:179–94. doi: 10.1016/s0169-409x(98)00085-4. [DOI] [PubMed] [Google Scholar]
- 13.Cordon-Cardo C, O’Brien JP, Casals D, Rittman-Grauer L, Biedler JL, Melamed MR, et al. Multidrug-resistance gene (P-glycoprotein) is expressed by endothelial cells at blood-brain barrier sites. Proc Natl Acad Sci USA. 1989;86:695–8. doi: 10.1073/pnas.86.2.695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Linnet K, Ejsing TB. A review on the impact of P-glycoprotein on the penetration of drugs into the brain. Focus on psychotropic drugs. Eur Neuropsychopharmacol. 2008;18:157–69. doi: 10.1016/j.euroneuro.2007.06.003. [DOI] [PubMed] [Google Scholar]
- 15.Dold M, Kautzky A, Bartova L, Rabl U, Souery D, Mendlewicz J, et al. Pharmacological treatment strategies in unipolar depression in European tertiary psychiatric treatment centers – A pharmacoepidemiological cross-sectional multicenter study. Eur Neuropsychopharmacol. 2016;26:1960–71. doi: 10.1016/j.euroneuro.2016.10.005. [DOI] [PubMed] [Google Scholar]
- 16.Treviño LA, Ruble MW, Treviño K, Weinstein LM, Gresky DP. Antidepressant medication prescribing practices for treatment of major depressive disorder. Psychiatr Serv. 2017;68:199–202. doi: 10.1176/appi.ps.201600087. [DOI] [PubMed] [Google Scholar]
- 17.Uhr M, Grauer MT. abcb1ab P-glycoprotein is involved in the uptake of citalopram and trimipramine into the brain of mice. J Psychiatr Res. 2003;37:179–85. doi: 10.1016/s0022-3956(03)00022-0. [DOI] [PubMed] [Google Scholar]
- 18.Uhr M, Tontsch A, Namendorf C, Ripke S, Lucae S, Ising M, et al. Polymorphisms in the drug transporter gene ABCB1 predict antidepressant treatment response in depression. Neuron. 2008;57:203–9. doi: 10.1016/j.neuron.2007.11.017. [DOI] [PubMed] [Google Scholar]
- 19.Doran A, Obach RS, Smith BJ, Hosea NA, Becker S, Callegari E, et al. The impact of P-glycoprotein on the disposition of drugs targeted for indications of the central nervous system: evaluation using the MDR1A/1B knockout mouse model. Drug Metab Dispos. 2005;33:165–74. doi: 10.1124/dmd.104.001230. [DOI] [PubMed] [Google Scholar]
- 20.Bundgaard C, Jensen CJ, Garmer M. Species comparison of in vivo P-glycoprotein-mediated brain efflux using mdr1a-deficient rats and mice. Drug Metab Dispos. 2012;40:461–6. doi: 10.1124/dmd.111.043083. [DOI] [PubMed] [Google Scholar]
- 21.Karlsson L, Carlsson B, Hiemke C, Ahlner J, Bengtsson F, Schmitt U, et al. Altered brain concentrations of citalopram and escitalopram in P-glycoprotein deficient mice after acute and chronic treatment. Eur Neuropsychopharmacol. 2013;23:1636–44. doi: 10.1016/j.euroneuro.2013.01.003. [DOI] [PubMed] [Google Scholar]
- 22.Holsboer F. How can we realize the promise of personalized antidepressant medicines? Nat Rev Neurosci. 2008;9:638–46. doi: 10.1038/nrn2453. [DOI] [PubMed] [Google Scholar]
- 23.Gex-Fabry M, Eap CB, Oneda B, Gervasoni N, Aubry JM, Bondolfi G, et al. CYP2D6 and ABCB1 genetic variability: influence on paroxetine plasma level and therapeutic response. Ther Drug Monit. 2008;30:474–82. doi: 10.1097/FTD.0b013e31817d6f5d. [DOI] [PubMed] [Google Scholar]
- 24.Kato M, Fukuda T, Serretti A, Wakeno M, Okugawa G, Ikenaga Y, et al. ABCB1 (MDR1) gene polymorphisms are associated with the clinical response to paroxetine in patients with major depressive disorder. Prog neuro-Psychopharmacol Biol psychiatry. 2008;32:398–404. doi: 10.1016/j.pnpbp.2007.09.003. [DOI] [PubMed] [Google Scholar]
- 25.Nikisch G, Eap CB, Baumann P. Citalopram enantiomers in plasma and cerebrospinal fluid of ABCB1 genotyped depressive patients and clinical response: a pilot study. Pharm Res. 2008;58:344–7. doi: 10.1016/j.phrs.2008.09.010. [DOI] [PubMed] [Google Scholar]
- 26.Sarginson JE, Lazzeroni LC, Ryan HS, Ershoff BD, Schatzberg AF, Murphy GM., Jr ABCB1 (MDR1) polymorphisms and antidepressant response in geriatric depression. Pharmacogenet Genomics. 2010;20:467–75. doi: 10.1097/FPC.0b013e32833b593a. [DOI] [PubMed] [Google Scholar]
- 27.Lin KM, Chiu YF, Tsai IJ, Chen CH, Shen WW, Liu SC, et al. ABCB1 gene polymorphisms are associated with the severity of major depressive disorder and its response to escitalopram treatment. Pharmacogenet Genomics. 2011;21:163–70. doi: 10.1097/FPC.0b013e32833db216. [DOI] [PubMed] [Google Scholar]
- 28.Singh AB, Bousman CA, Ng CH, Byron K, Berk M. ABCB1 polymorphism predicts escitalopram dose needed for remission in major depression. Transl psychiatry. 2012;2:e198. doi: 10.1038/tp.2012.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Huang X, Yu T, Li X, Cao Y, Li X, Liu B, et al. ABCB6, ABCB1 and ABCG1 genetic polymorphisms and antidepressant response of SSRIs in Chinese depressive patients. Pharmacogenomics. 2013;14:1723–30. doi: 10.2217/pgs.13.151. [DOI] [PubMed] [Google Scholar]
- 30.Schatzberg AF, DeBattista C, Lazzeroni LC, Etkin A, Murphy GM, Jr., Williams LM. ABCB1 genetic effects on antidepressant outcomes: a report from the iSPOT-D trial. Am J Psychiatry. 2015;172:751–9. doi: 10.1176/appi.ajp.2015.14050680. [DOI] [PubMed] [Google Scholar]
- 31.Peters EJ, Slager SL, Kraft JB, Jenkins GD, Reinalda MS, McGrath PJ, et al. Pharmacokinetic genes do not influence response or tolerance to citalopram in the STAR*D sample. PloS one. 2008;3:e1872. doi: 10.1371/journal.pone.0001872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Mihaljevic Peles A, Bozina N, Sagud M, Rojnic Kuzman M, Lovric M. MDR1 gene polymorphism: therapeutic response to paroxetine among patients with major depression. Prog neuro-Psychopharmacol Biol psychiatry. 2008;32:1439–44. doi: 10.1016/j.pnpbp.2008.03.018. [DOI] [PubMed] [Google Scholar]
- 33.Menu P, Gressier F, Verstuyft C, Hardy P, Becquemont L, Corruble E. Antidepressants and ABCB1 gene C3435T functional polymorphism: a naturalistic study. Neuropsychobiology. 2010;62:193–7. doi: 10.1159/000319361. [DOI] [PubMed] [Google Scholar]
- 34.Perlis RH, Fijal B, Dharia S, Heinloth AN, Houston JP. Failure to replicate genetic associations with antidepressant treatment response in duloxetine-treated patients. Biol Psychiatry. 2010;67:1110–3. doi: 10.1016/j.biopsych.2009.12.010. [DOI] [PubMed] [Google Scholar]
- 35.Uher R, Perroud N, Ng MY, Hauser J, Henigsberg N, Maier W, et al. Genome-wide pharmacogenetics of antidepressant response in the GENDEP project. Am J Psychiatry. 2010;167:555–64. doi: 10.1176/appi.ajp.2009.09070932. [DOI] [PubMed] [Google Scholar]
- 36.Jovanović N, Božina N, Lovrić M, Medved V, Jakovljević M, Peleš AM. The role of CYP2D6 and ABCB1 pharmacogenetics in drug-naïve patients with first-episode schizophrenia treated with risperidone. Eur J Clin Pharmacol. 2010;66:1109–17. doi: 10.1007/s00228-010-0850-1. [DOI] [PubMed] [Google Scholar]
- 37.Xing Q, Gao R, Li H, Feng G, Xu M, Duan S, et al. Polymorphisms of the ABCB1 gene are associated with the therapeutic response to risperidone in Chinese schizophrenia patients. Pharmacogenomics. 2006;7:987–93. doi: 10.2217/14622416.7.7.987. [DOI] [PubMed] [Google Scholar]
- 38.Nikisch G, Baumann P, Oneda B, Kiessling B, Weisser H, Mathé AA, et al. Cytochrome P450 and ABCB1 genetics: association with quetiapine and norquetiapine plasma and cerebrospinal fluid concentrations and with clinical response in patients suffering from schizophrenia. A pilot study. J Psychopharmacol (Oxf, Engl) 2011;25:896–907. doi: 10.1177/0269881110389208. [DOI] [PubMed] [Google Scholar]
- 39.Zimprich F, Sunder-Plassmann R, Stogmann E, Gleiss A, Dal-Bianco A, Zimprich A, et al. Association of an ABCB1 gene haplotype with pharmacoresistance in temporal lobe epilepsy. Neurology. 2004;63:1087–9. doi: 10.1212/01.wnl.0000141021.42763.f6. [DOI] [PubMed] [Google Scholar]
- 40.Gryglewski G, Klobl M, Berroteran-Infante N, Rischka L, Balber T, Vanicek T et al. Modeling the acute pharmacological response to selective serotonin reuptake inhibitors in human brain using simultaneous PET/MR imaging. Eur Neuropsychopharmacol. 2019;29:711–9. [DOI] [PubMed]
- 41.Gryglewski G, Rischka L, Philippe C, Hahn A, James GM, Klebermass E, et al. Simple and rapid quantification of serotonin transporter binding using [(11)C]DASB bolus plus constant infusion. NeuroImage. 2017;149:23–32. doi: 10.1016/j.neuroimage.2017.01.050. [DOI] [PubMed] [Google Scholar]
- 42.Silberbauer LR, Gryglewski G, Berroterán-Infante N, Rischka L, Vanicek T, Pichler V et al. Serotonin transporter binding in the human brain after pharmacological challenge measured using PET and PET/MR. Front in Mol Neurosc. 2019;12:172. [DOI] [PMC free article] [PubMed]
- 43.Ginovart N, Wilson AA, Meyer JH, Hussey D, Houle S. Positron emission tomography quantification of [(11)C]-DASB binding to the human serotonin transporter: modeling strategies. J Cereb Blood Flow Metab: Off J Int Soc Cereb Blood Flow Metab. 2001;21:1342–53. doi: 10.1097/00004647-200111000-00010. [DOI] [PubMed] [Google Scholar]
- 44.Roberts RL, Joyce PR, Mulder RT, Begg EJ, Kennedy MA. A common P-glycoprotein polymorphism is associated with nortriptyline-induced postural hypotension in patients treated for major depression. Pharmacogenomics J. 2002;2:191–6. doi: 10.1038/sj.tpj.6500099. [DOI] [PubMed] [Google Scholar]
- 45.Noordam R, Aarts N, Hofman A, van Schaik RH, Stricker BH, Visser LE. Association between genetic variation in the ABCB1 gene and switching, discontinuation, and dosage of antidepressant therapy: results from the Rotterdam Study. J Clin Psychopharmacol. 2013;33:546–50. doi: 10.1097/JCP.0b013e318291c07b. [DOI] [PubMed] [Google Scholar]
- 46.de Klerk OL, Nolte IM, Bet PM, Bosker FJ, Snieder H, den Boer JA, et al. ABCB1 gene variants influence tolerance to selective serotonin reuptake inhibitors in a large sample of Dutch cases with major depressive disorder. Pharmacogenomics J. 2013;13:349–53. doi: 10.1038/tpj.2012.16. [DOI] [PubMed] [Google Scholar]
- 47.Perroud N, Bondolfi G, Uher R, Gex-Fabry M, Aubry JM, Bertschy G, et al. Clinical and genetic correlates of suicidal ideation during antidepressant treatment in a depressed outpatient sample. Pharmacogenomics. 2011;12:365–77. doi: 10.2217/pgs.10.189. [DOI] [PubMed] [Google Scholar]
- 48.Gassó P, Rodríguez N, Mas S, Pagerols M, Blázquez A, Plana MT, et al. Effect of CYP2D6, CYP2C9 and ABCB1 genotypes on fluoxetine plasma concentrations and clinical improvement in children and adolescent patients. Pharmacogenomics J. 2014;14:457–62. doi: 10.1038/tpj.2014.12. [DOI] [PubMed] [Google Scholar]
- 49.Chang HH, Chou CH, Yang YK, Lee IH, Chen PS. Association between ABCB1 polymorphisms and antidepressant treatment response in Taiwanese major depressive patients. Clin Psychopharmacol Neurosci. 2015;13:250–5. doi: 10.9758/cpn.2015.13.3.250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Bly MJ, Bishop JR, Thomas KL, Ellingrod VL. P-glycoprotein (PGP) polymorphisms and sexual dysfunction in female patients with depression and SSRI-associated sexual side effects. J Sex Marital Ther. 2013;39:280–8. doi: 10.1080/0092623X.2011.615896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Baldinger P, Hahn A, Mitterhauser M, Kranz GS, Friedl M, Wadsak W, et al. Impact of COMT genotype on serotonin-1A receptor binding investigated with PET. Brain Struct Funct. 2014;219:2017–28. doi: 10.1007/s00429-013-0621-8. [DOI] [PubMed] [Google Scholar]
- 52.Sigurdardottir HL, Kranz GS, Rami-Mark C, James GM, Vanicek T, Gryglewski G, et al. Effects of norepinephrine transporter gene variants on NET binding in ADHD and healthy controls investigated by PET. Hum brain Mapp. 2016;37:884–95. doi: 10.1002/hbm.23071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Unterecker S, Riederer P, Proft F, Maloney J, Deckert J, Pfuhlmann B. Effects of gender and age on serum concentrations of antidepressants under naturalistic conditions. J neural Transm (Vienna, Austria: 1996) 2013;120:1237–46. doi: 10.1007/s00702-012-0952-2. [DOI] [PubMed] [Google Scholar]
- 54.Reis M, Aamo T, Spigset O, Ahlner J. Serum concentrations of antidepressant drugs in a naturalistic setting: compilation based on a large therapeutic drug monitoring database. Ther Drug Monit. 2009;31:42–56. doi: 10.1097/FTD.0b013e31819114ea. [DOI] [PubMed] [Google Scholar]
- 55.Sogaard B, Mengel H, Rao N, Larsen F. The pharmacokinetics of escitalopram after oral and intravenous administration of single and multiple doses to healthy subjects. J Clin Pharmacol. 2005;45:1400–6. doi: 10.1177/0091270005280860. [DOI] [PubMed] [Google Scholar]
- 56.Booij J, de Jong J, de Bruin K, Knol R, de Win MM, van Eck-Smit BL. Quantification of striatal dopamine transporters with 123I-FP-CIT SPECT is influenced by the selective serotonin reuptake inhibitor paroxetine: a double-blind, placebo-controlled, crossover study in healthy control subjects. J Nucl Med. 2007;48:359–66. [PubMed] [Google Scholar]
- 57.Kugaya A, Seneca NM, Snyder PJ, Williams SA, Malison RT, Baldwin RM, et al. Changes in human in vivo serotonin and dopamine transporter availabilities during chronic antidepressant administration. Neuropsychopharmacology. 2003;28:413–20. doi: 10.1038/sj.npp.1300036. [DOI] [PubMed] [Google Scholar]
- 58.Uhr M, Grauer MT, Holsboer F. Differential enhancement of antidepressant penetration into the brain in mice with abcb1ab (mdr1ab) P-glycoprotein gene disruption. Biol Psychiatry. 2003;54:840–6. doi: 10.1016/s0006-3223(03)00074-x. [DOI] [PubMed] [Google Scholar]
- 59.Breitenstein B, Scheuer S, Brückl TM, Meyer J, Ising M, Uhr M, et al. Association of ABCB1 gene variants, plasma antidepressant concentration, and treatment response: Results from a randomized clinical study. J Psychiatr Res. 2016;73:86–95. doi: 10.1016/j.jpsychires.2015.11.010. [DOI] [PubMed] [Google Scholar]
- 60.Hoffmeyer S, Burk O, von Richter O, Arnold HP, Brockmöller J, Johne A, et al. Functional polymorphisms of the human multidrug-resistance gene: multiple sequence variations and correlation of one allele with P-glycoprotein expression and activity in vivo. Proc Natl Acad Sci USA. 2000;97:3473–8. doi: 10.1073/pnas.050585397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kokras N, Dalla C, Papadopoulou-Daifoti Z. Sex differences in pharmacokinetics of antidepressants. Expert Opin Drug Metab Toxicol. 2011;7:213–26. doi: 10.1517/17425255.2011.544250. [DOI] [PubMed] [Google Scholar]
- 62.Bigos KL, Pollock BG, Stankevich BA, Bies RR. Sex differences in the pharmacokinetics and pharmacodynamics of antidepressants: an updated review. Gend Med. 2009;6:522–43. doi: 10.1016/j.genm.2009.12.004. [DOI] [PubMed] [Google Scholar]
- 63.Marazziti D, Baroni S, Picchetti M, Piccinni A, Carlini M, Vatteroni E, et al. Pharmacokinetics and pharmacodinamics of psychotropic drugs: effect of sex. CNS Spectr. 2013;18:118–27. doi: 10.1017/S1092852912001010. [DOI] [PubMed] [Google Scholar]
- 64.Staley JK, Krishnan-Sarin S, Zoghbi S, Tamagnan G, Fujita M, Seibyl JP, et al. Sex differences in [123I]beta-CIT SPECT measures of dopamine and serotonin transporter availability in healthy smokers and nonsmokers. Synap (N. Y, NY) 2001;41:275–84. doi: 10.1002/syn.1084. [DOI] [PubMed] [Google Scholar]
- 65.Jovanovic H, Lundberg J, Karlsson P, Cerin Å, Saijo T, Varrone A, et al. Sex differences in the serotonin 1A receptor and serotonin transporter binding in the human brain measured by PET. NeuroImage. 2008;39:1408–19. doi: 10.1016/j.neuroimage.2007.10.016. [DOI] [PubMed] [Google Scholar]
- 66.Baldinger P, Kranz GS, Haeusler D, Savli M, Spies M, Philippe C, et al. Regional differences in SERT occupancy after acute and prolonged SSRI intake investigated by brain PET. NeuroImage. 2014;88:252–62. doi: 10.1016/j.neuroimage.2013.10.002. [DOI] [PubMed] [Google Scholar]
- 67.van Assema DM, Lubberink M, Boellaard R, Schuit RC, Windhorst AD, Scheltens P, et al. P-glycoprotein function at the blood-brain barrier: effects of age and gender. Mol Imaging Biol: MIB: Off Publ Acad Mol Imaging. 2012;14:771–6. doi: 10.1007/s11307-012-0556-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Jakubovski E, Varigonda AL, Freemantle N, Taylor MJ, Bloch MH. Systematic review and meta-analysis: dose-response relationship of selective serotonin reuptake inhibitors in major depressive disorder. Am J Psychiatry. 2016;173:174–83. doi: 10.1176/appi.ajp.2015.15030331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Lundberg J, Tiger M, Landen M, Halldin C, Farde L. Serotonin transporter occupancy with TCAs and SSRIs: a PET study in patients with major depressive disorder. Int J Neuropsychopharmacol. 2012;15:1167–72. doi: 10.1017/S1461145711001945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Sørensen A, Ruhé HG, Munkholm K. The relationship between dose and serotonin transporter occupancy of antidepressants-a systematic review. Mol psychiatry. 2022;27:192–201. doi: 10.1038/s41380-021-01285-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Weiss J, Dormann SM, Martin-Facklam M, Kerpen CJ, Ketabi-Kiyanvash N, Haefeli WE. Inhibition of P-glycoprotein by newer antidepressants. J Pharmacol Exp therapeutics. 2003;305:197–204. doi: 10.1124/jpet.102.046532. [DOI] [PubMed] [Google Scholar]
- 72.Vraka C, Dumanic M, Racz T, Pichler F, Philippe C, Balber T, et al. A new method measuring the interaction of radiotracers with the human P-glycoprotein (P-gp) transporter. Nucl Med Biol. 2018;60:29–36. doi: 10.1016/j.nucmedbio.2018.02.002. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.