


Abstract
Coordinating multiple activities of complex enzymes is critical for life, including transcribing, replicating and repairing DNA. Bacterial RecBCD helicase–nuclease must coordinate DNA unwinding and cutting to repair broken DNA. Starting at a DNA end, RecBCD unwinds DNA with its fast RecD helicase on the 5′-ended strand and its slower RecB helicase on the 3′-ended strand. At Chi hotspots (5′ GCTGGTGG 3′), RecB’s nuclease cuts the 3′-ended strand and loads RecA strand-exchange protein onto it. We report that a small molecule NSAC1003, a sulfanyltriazolobenzimidazole, mimics Chi sites by sensitizing RecBCD to cut DNA at a Chi-independent position a certain percent of the DNA substrate's length. This percent decreases with increasing NSAC1003 concentration. Our data indicate that NSAC1003 slows RecB relative to RecD and sensitizes it to cut DNA when the leading helicase RecD stops at the DNA end. Two previously described RecBCD mutants altered in the RecB ATP-binding site also have this property, but uninhibited wild-type RecBCD lacks it. ATP and NSAC1003 are competitive; computation docks NSAC1003 into RecB’s ATP-binding site, suggesting NSAC1003 acts directly on RecB. NSAC1003 will help elucidate molecular mechanisms of RecBCD-Chi regulation and DNA repair. Similar studies could help elucidate other DNA enzymes with activities coordinated at chromosomal sites.
INTRODUCTION
Complex, multi-subunit enzymes are required for most activities on nucleic acids, including replication, recombination, DNA repair, transcription, RNA splicing and protein synthesis. Particularly important is the repair of DNA double-strand breaks (DSBs) because when their DNA is broken, cells must repair it or they die. Faithful DSB repair requires DNA helicases and nucleases to prepare the broken DNA for interaction with an intact homologous DNA molecule. If the interacting DNA molecules differ genetically, DSB repair can also produce genetic recombinants and thereby propel evolution. Understanding the molecular basis of DSB repair and recombination requires understanding how complex, multi-functional enzymes act on DNA. Here, we describe a small organic molecule that alters the activity of a DNA helicase–nuclease complex in a novel way that lends new insights into the enzyme's mechanism and regulation.
In Escherichia coli and other enteric bacteria, DSB repair and recombination require RecBCD, a complex three-subunit 330 kDa enzyme with both DNA helicase and DNA nuclease activities (1,2,3). Its multiple activities are involved in the initial steps of DSB repair and recombination (Figure 1A). RecBCD binds tightly to a ds DNA end and unwinds the DNA rapidly (up to ∼1.5 kb/s) and with high processivity (up to 100 kb without dissociating) (4). RecB helicase binds to the 3′ end, and RecD to the 5′ end (5,6). Each helicase subunit hydrolyses ATP and moves along its bound strand (7). Because RecB is slower than RecD, a single-stranded (ss) DNA loop accumulates, presumably ahead of the RecB subunit (4). This loop grows with time of incubation, as do the short and long ss tails extending behind the enzyme. Annealing of the tails forms ds DNA and a second loop; the two loops continue to grow and move with time of incubation. Upon encountering a Chi site (5′ GCTGGTGG 3′), a hotspot of recombination, the RecB nuclease cuts the strand with this sequence (8). Unwinding continues, and RecA strand-exchange protein is loaded onto the newly generated 3′-ended strand likely by the RecB nuclease domain (9,10). The RecA–ssDNA filament invades intact homologous DNA to form a displacement (D)-loop (11). Subsequent reactions involve formation and resolution of a Holliday junction into reciprocal recombinants or priming of DNA synthesis to form a non-reciprocal recombinant (12,13).
Figure 1.

Models for RecBCD enzyme and its Chi-dependent promotion of DNA break repair and genetic recombination. (A) RecBCD pathway of genetic recombination. See Introduction for explanation. Note at step b, the long 5′ tail made by RecD and the short 3′ tail and loop made by RecB. At step c, these tails pair and form a second loop; the two loops continue to grow as RecBCD continues unwinding (4). (B) Signal-transduction model for Chi's control of RecBCD enzyme (14). When Chi is in the RecC tunnel, RecC signals RecD to stop unwinding DNA; RecD signals RecB to nick the DNA and to begin loading RecA. (C) Crystal structure of RecBCD bound to a ds DNA hairpin [PDB 1W36; (6)]. RecB (orange) contains helicase and nuclease domains connected by a tether. RecD (green) is held to RecB via RecC (blue). During unwinding, the 3′-ended strand passes through a tunnel (yellow dashed line) in RecC and into the nuclease active site when Chi is encountered. (D) Nuclease-swing model for Chi's control of RecBCD enzyme (15). Before DNA is bound (D1), RecBCD in solution assumes its conformation in the published structures (PDB 1W36 and PDB 5LD2). Upon binding DNA (D2), the nuclease domain swings away from the exit of the RecC tunnel (15). When Chi is encountered during unwinding (D3), the nuclease domain swings back, cuts the DNA at Chi, and begins loading RecA protein, perhaps after rotating to prevent further nuclease action. Modified from (16).
The molecular mechanism by which a Chi hotspot signals RecBCD to cut DNA at Chi and to begin loading RecA remains to be fully elucidated. Based on the behavior of two mutants altered in the RecB helicase domain, Amundsen et al. (14) proposed a ‘signal-transduction’ model (Figure 1B). In this model when Chi is in a tunnel in RecC (Figure 1C), RecC signals RecD to stop; RecD then signals RecB to cut the DNA at Chi and to load RecA. Subsequent analysis suggested that upon receiving the signal from RecD, RecB’s nuclease domain swings on its 19-amino-acid tether from an inactive position (on the ‘left’ side of RecC) to its nuclease-active position (at the exit of the tunnel in RecC) (Figure 1D) (15,16). Extensive mutational analyses of the RecC tunnel and of the RecB tether support this model (17,18,16). Mutants with altered contacts between RecC–RecD, RecD–RecB and RecB–RecC indicate that each of these contacts is important for Chi hotspot activity (19).
Here, we report the action of a small organic molecule on RecBCD, which mimics the behavior of the two RecB helicase mutants noted above (14). This small molecule, NSAC1003 (MW = 431; Figure 2A), was discovered by Achaogen in a screen for inhibitors of the helicase–nuclease activity of RecBCD. In the current studies, we noted that it induces RecBCD to cut DNA at a novel position that depends on the NSAC1003 concentration and on the length of the substrate. The two RecB helicase mutants mentioned above are altered in single amino acids (Y803H and V804E) near RecB’s ATP-binding site in a cryoEM structure (20) (see Discussion and Figure 3). These two helicase mutants also cut DNA at novel positions that depend on the substrate length (14). From these and other data, we infer that NSAC1003 has two effects: it slows the RecB helicase, relative to RecD, in a concentration-dependent manner, and it sensitizes the enzyme to cut DNA when RecD stops unwinding DNA at the end of the substrate. We discuss a molecular basis for these effects and how related compounds may also act on RecBCD and other complex enzymes.
Figure 2.
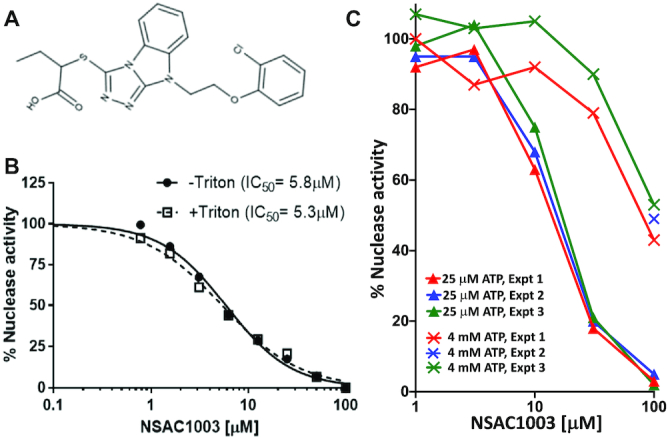
NSAC1003 inhibits RecBCD Chi-independent nuclease activity in an ATP-competitive manner. (A) Structure of NSAC1003, a sulfanyltriazolobenzimidazole. The IUPAC name is 2-({7-[2-(2-chlorophenoxy)ethyl]-2,4,5,7-tetraazatricyclo[6.4.0.02,6]dodeca-1(8),3,5,9,11-pentaen-3-yl}sulfanyl)butanoic acid. (B) RecBCD nuclease activity was assayed, using [3H] phage T7 DNA, in the presence of the indicated concentration of NSAC1003 with and without Triton X-100 (0.01%). See also Supplementary Figure S1 for duplicate assays without Triton X-100, in which the IC50 was 6.3 and 6.7 μM. (C) RecBCD nuclease activity was assayed in the presence of the standard 25 μM ATP (▴) or 4 mM ATP (X) similar to the 5 mM ATP used in RecBCD unwinding-cutting experiments in Figures 4 and 5. RecBCD was added before DNA in experiments 1 and 2, but DNA before RecBCD in experiment 3. See also Supplementary Figures S1 and S2.
Figure 3.

RecB helicase mutants and NSAC1003 docking site are at the RecB ATP-binding site. (A) RecBCD (RecB, orange; RecC, blue; RecD, green; surface view of PDB 5LD2) is shown with NSAC1003 (green sticks) in the RecB ATP-binding site (circled). NSAC1003 was docked with Docking Server (23) to a 20 Å cube of RecB centered on the ATP-binding site but without ADPNP or Mg2+ for the calculation. RecB’s Walker A box K29 (red), catalytic Mg2+ (cyan sphere), and the amino acids (F31 and W447; magenta) sandwiching the aromatic tricyclic core of NSAC1003 are also shown, as is the RecC tunnel (dashed yellow line) in which Chi is recognized. (B) Expanded view of part of panel A. (C) As in panel B with RecB as cartoon view except for K29 (red), F31 and W447 (magenta), and two helicase mutants Y803 and V804 (yellow) as sticks. Y803 and V804 are not visible in panels A and B. (D) As in panel C with ADPNP (blue sticks) superimposed. See also Supplementary Figure S3.
MATERIALS AND METHODS
Materials
RecBCD enzyme (200 000 units/mg) (15, 22), pBR322 (χo and χ+E224) (21), and [3H] phage T7 DNA (22) were prepared as described. Restriction enzymes and polynucleotide kinase were purchased from New England BioLabs (Ipswich, MA, USA), [γ-32P] ATP from Perkin-Elmer (Waltham, MA, USA), Triton X-100 from Sigma-Aldrich (St. Louis, MO, USA) and NSAC1003 and compound 1 from Nanosyn (Santa Clara, CA, USA). NSAC1003 was dissolved in DMSO at 10 mM; pure DMSO was added to experimental reactions to make a total concentration of 5% (nuclease activity in Figure 2 and Supplementary Figure S1) or 2% (unwinding and DNA cutting in Figure 4).
Figure 4.

NSAC1003 induces Chi-independent DNA cuts. (A) Diagram of 5′-labeled (*) ds DNA substrates of various lengths. (B) Diagram of RecBCD unwinding DNA with production of a ss DNA loop and two tails (4). x is the rate of the slower RecB helicase on the 3′-ended (top) strand, and y the rate of the faster RecD helicase on the 5′-ended (bottom) strand. When RecD reaches the left end of the DNA, in the presence of NSAC1003 RecB makes a novel cut where it is at that moment, or x/y of the substrate length, measured from RecBCD’s entry point. The length of the labeled product [S(1 – x/y)] is a nearly constant fraction independent of the initial substrate length S (see Figure 5A). NSAC1003 inhibits RecB helicase more than RecD helicase (see Results); thus, x/y decreases as [NSAC1003] increases and the cut product is longer. (C) pBR322 DNA (χo or χ+E224) was cut with HindIII and labeled at the 5′ end; portions were cut with ClaI, NdeI or StyI to produce singly labeled DNA ∼4350, 2270 or 1340 bp long (panel A). DNA was reacted with RecBCD enzyme with the indicated concentrations of NSAC1003 for 60 sec. Reaction products were examined by gel electrophoresis as described in Materials and methods. M, ss DNA markers; N, native (ds) substrate; B, boiled (ss) substrate; *, Chi-dependent product; -, χo substrate; +, χ+E224 substrate. Similar results were obtained in two additional experiments done on separate days. The upper left corner of the middle gel was marred, obscuring some marker bands.
Enzymatic methods
RecBCD ‘general’ (Chi-independent) nuclease activity (Figure 2 and Supplementary Figure S1) was assayed as described (22) as solubilization of uniformly labeled phage T7 DNA; a dye-displacement assay was used for the assays in Supplementary Figure S2. For Figure 2 and Supplementary Figure S1, reaction mixtures (50 μl) contained 50 mM Tris–HCl (pH 8.5), 10 mM MgCl2, polyvinylpyrrolidone (1 mg/ml), 1 mM dithiothreitol, 25 μM or 4 mM ATP, RecBCD enzyme (4 units/ml = 20 ng/ml = 0.061 nM in Figure 2B and Supplementary Figure S1; 3 units/ml = 15 ng/ml = 0.045 nM in Figure 2C), [3H] phage T7 DNA (2 μg/ml = 0.077 nM DNA molecules in Figure 2B and Supplementary Figure S1; 5.5 μg/ml = 0.21 nM in Figure 2C), and NSAC1003 at the indicated concentration; DMSO, the solvent for NSAC1003, was added to 5% final concentration in all reactions. Where indicated, Triton X-100 was added to 0.01%. Reactions in Figure 2B and Supplementary Figure S1 were assembled on ice and transferred to a 37°C bath to initiate the reaction; those in Figure 2C were assembled at 37°C and initiated by addition of ATP. After 20 min at 37°C, the reaction was stopped by addition of 25 μl of calf thymus DNA (0.2 mg/ml) and 250 μl of 5% trichloroacetic acid (TCA). After 10 min on ice, the tubes were centrifuged for 5 min at 13 400 RPM, and 300 μl of the supernate were assayed with a scintillation counter.
Chi-cutting activity (Figure 4C) was assayed as described (14). Reaction mixtures (15 μl) contained 25 mM Tris–HCl (pH 7.5), 2.5 mM MgCl2, 1 mM dithiothreitol, 0.2 nM [5′-32P] DNA (0.56, 0.30 and 0.18 μg/ml for the three substrates), 0.4 nM (32 units/ml = 130 ng/ml) RecBCD enzyme and NSAC1003 at the indicated concentration. DMSO was present at 2% in all reactions, which were incubated at 37°C for 10 min before initiating the reaction by addition of ATP to 5 mM. After 60 s at 37°C, the reaction was stopped by addition of 5 μl of stop buffer, and the products analyzed by electrophoresis on a 1.5% agarose gel (22 cm long) in TAE buffer (110 V for 3 h). The gel contents were analyzed by Typhoon Trio PhosphorImager (GE Lifesciences) and ImageQuant TL software (GE Lifesciences).
Docking methods
Using DockingServer (23), NSAC1003 was docked to a 20 Å cube containing the RecB or RecD ATP site in cryoEM structure PDB 5LD2 (24) with ADPNP and Mg2+ removed. Gasteiger partial charges were added to the ligand atoms. Non-polar hydrogen atoms were merged, and rotatable bonds were defined. Essential hydrogen atoms, Kollman united atom-type charges, and solvation parameters were added with the aid of AutoDock tools (25). Affinity (grid) maps of 20 Å grid points and 0.375 Å spacing were generated using the Autogrid program (25). During the search, a translational step of 0.2 Å and quaternion and torsion steps of 5 were applied. AutoDock parameter set- and distance-dependent dielectric functions were used in the calculation of van der Waals and electrostatic terms, respectively. Docking simulations were performed using the Lamarckian genetic algorithm (LGA) and a local search method (26). Initial position, orientation, and torsions of the ligand molecules were set randomly. All rotatable torsions were released during docking. Each docking experiment was derived from 100 different runs set to terminate after a maximum of 2 500 000 energy evaluations. The population size was set to 150. The dockings with the highest affinity scores are shown in Figure 3 and Supplementary Figure S3.
RESULTS
NSAC1003 inhibits RecBCD nuclease activity
RecBCD has potent Chi-independent (‘general’) nuclease activity under conditions with excess Mg2+ relative to ATP (22). In standard assays of RecBCD nuclease activity with 10 mM Mg2+ and 25 μM ATP, we found that NSAC1003 inhibited with an IC50 of 6.3 ± 0.3 μM (Figure 2B and Supplementary Figure S1). In other experiments, we compared NSAC1003 inhibition with ATP at 25 μM and 4 mM, side-by-side (Figure 2C); the IC50 was ∼10 and ∼100 μM, respectively, indicating that NSAC1003 and ATP compete with each other. A related compound, a hexanoic acid derivative otherwise identical to the butanoic acid derivative (NSAC1003), also inhibited RecBCD nuclease in an ATP-competitive manner (Supplementary Figure S2). These results suggest that these compounds inhibit ATP hydrolysis, which is required for helicase activity and thus nuclease activity (22,28,29,7).
Because some organic compounds found in screens for antibiotics form microcrystals or aggregates and may sequester an enzyme rather than simply inhibit it (27), we repeated the assays in the presence of 0.01% Triton X-100, which is thought to counter the effect of microcrystals. NSAC1003 inhibition was indistinguishable in the presence and absence of Triton (Figure 2B). Furthermore, addition of NSAC1003 (5–80 μM) to RecBCD, either without DNA or with DNA in an active reaction, followed by centrifugation (14 000 × g, 15 min) did not remove RecBCD from solution (assayed by western blots for RecB and RecC; unpublished data). Thus, we conclude that NSAC1003 inhibits RecBCD by direct binding, perhaps to the ATP-binding site(s) in accord with the competition results above.
NSAC1003 is predicted to bind tightly to the RecB ATP site
To test more directly the idea that NSAC1003 inhibits RecBCD by binding to an ATP-binding site, we computationally docked it to each of the two ATP sites, one in the RecB helicase domain and one in RecD. Docking server (23) indicated that NSAC1003 binds to the RecB ATP site with high affinity (KD ∼ 0.3 μM) and to the RecD ATP site with lower affinity (KD ∼ 2.2 μM) (Figure 3 and Supplementary Figure S3). The predicted binding affinities are lower than the measured IC50 of NSAC1003 for nuclease activity even at the lowest ATP concentration tested (25 μM) (Figure 2, Supplementary Figures S1 and S2), likely because ATP was not present in the computational docking. These results support the idea the NSAC1003 blocks the RecB ATPase and thus slows the RecB helicase relative to the RecD helicase (see Discussion). They are also consistent with the recB29 (K29Q) ATPase-negative mutant being ds nuclease-negative, but the recD2177 (K177Q) ATPase-negative mutant retaining significant nuclease activity (28,29,7).
NSAC1003 induces RecBCD to cut DNA at novel positions
Under conditions with excess ATP relative to Mg2+, RecBCD unwinds DNA and nicks one strand near Chi to generate a hotspot of recombinational exchange at Chi, as in cells (1). Using 5 mM ATP and 2.5 mM Mg2+, we tested the effect of NSAC1003 on RecBCD’s DNA unwinding and Chi-cutting activities. The linear ds DNA substrate was labeled at one 5′-end with 32P and contained, or not, an internal Chi site (χ+E224) (Figure 4A). After brief reaction, the products were analyzed by gel electrophoresis in comparison with ds and ss DNA length standards. In the absence of NSAC1003 RecBCD unwound some of the Chi-containing DNA (4.35 kb long; top panel) and produced a radioactive ss DNA fragment ∼970 nucleotides long, as expected from the Chi site being ∼970 bp from the 5′-32P label; also as expected, this fragment was not detected with DNA substrate lacking Chi (Figure 4B, C). [RecBCD entering the DNA from the right but not from the left, as drawn, cuts at Chi (30).] Similar results were found with 25 μM NSAC1003. With 50–400 μM NSAC1003, however, a Chi-dependent fragment was not detected but was replaced with a longer fragment indicative of cutting before Chi. This fragment's length increased with increasing NSAC1003 concentration and was observed with and without Chi. We interpret these results below.
With shorter DNA substrates, the results changed in an interesting way. With the 2.27 kb substrate (middle panel), the Chi-dependent fragment (∼970 nucleotides long) was detected with 0, 25 and 50 μM NSAC1003 but not with 100, 200 or 400 μM NSAC1003. At these higher NSAC1003 concentrations, radioactive fragments of increasing length were produced with DNA containing Chi or not. The length of these fragments increased with increasing NSAC1003 concentration, as noted above with the 4.35 kb substrate. With the 1.34 kb substrate (bottom panel), the Chi-dependent fragment was detected with 0–100 μM NSAC1003. An additional, Chi-independent fragment was produced with 50–400 μM NSAC1003, and as above its length increased with increasing NSAC1003 concentration.
The positions of NSAC1003-induced cuts depend on the NSAC1003 concentration and on the substrate length
The lengths of the Chi-independent fragments noted above depended on the NSAC1003 concentration, with an apparent half-maximal effect at ∼50–100 μM (Figure 5A). This value is comparable to the IC50 for NSAC1003 inhibition of the ‘general’ (Chi-independent) nuclease activity at the higher ATP concentration for these unwinding-cutting experiments (Figure 2, Supplementary Figures S1 and S2).
Figure 5.

The length of the NSAC1003-induced cut product is a linear function of substrate length, dependent on the NSAC1003 concentration. Three experiments of the type shown in Figure 4C were analyzed; data are the mean ± SEM (not visible in most cases). The length of the cut product (middle of the band) was estimated by interpolation against the indicated size markers (boiled substrates and products of digestion with SalI, BamHI or AlwNI). (A) Labeled product length increases with increasing NSAC1003 concentration. (B) Labeled product length is a linear function of substrate length. Note that the extrapolate to zero substrate length is slightly negative (–250 bases at 50 μM; –370 at 100 μM; –290 at 200 μM; and –200 at 400 μM) (see Discussion).
The radioactive product lengths were a linear function of the length of the DNA substrate (Figures 4B and 5B). These data show that NSAC1003 induces RecBCD to cut the DNA ∼30–70% of the distance from the entry point (the unlabeled end of the substrate) (Figure 4). In other words, at a low concentration (25 μM) of NSAC1003 the enzyme cuts at ∼70% of the substrate length from the entry point, and at higher concentration it cuts at ∼30% of the substrate length. This outcome is nearly the same for each substrate length tested. These results suggest that NSAC1003 slows RecB helicase, relative to RecD, and induces RecB nuclease to cut the DNA where it is when RecD reaches the end of the DNA (see Figure 4B and Discussion).
DISCUSSION
Our results presented here show a remarkable similarity between the effect of NSAC1003 inhibitor and the effect of two mutations altering amino acids in the RecB helicase very near its ATP-binding site (Figure 3). Molecular docking (23) indicates that NSAC1003 binds to the ATP-binding site in RecB (Figure 3) with high affinity (KD ∼ 0.3 μM) and with two consequences: the RecB helicase is slowed, relative to RecD, likely reflecting the competition with ATP binding and hydrolysis by RecB, and the nuclease is sensitized to cut the DNA when RecD helicase stops, perhaps because NSAC1003 alters the RecBCD conformation in the same way as the two mutations in the ATP-binding site of RecB (Y803H and V804E) (14). These effects of NSAC1003 indicate that this compound will be informative in further elucidating Chi's control of RecBCD enzyme and other complex, multi-activity enzymes, as discussed below.
Our interpretations of the data in Figures 4 and 5 reflect those of the two RecB helicase mutants that cut DNA at a certain fraction of the length of the DNA substrate (14). Initially, it was mysterious how these mutants could measure the length of the substrate, calculate a certain percent of that length (∼19% for Y803H and ∼6% V804E), and cut the DNA at that position. Analysis of the rates of the mutant RecB and RecD helicases by electron microscopy of partially unwound DNA molecules showed that the ratio of the RecB:RecD helicase rates was nearly the same as the fraction of the substrate length (from the RecBCD entry point) at which cuts occur. This led to the conclusion that, when RecD stops at the end of the DNA, it signals RecB to cut where it is at that moment. We propose the same interpretation for the effect of NSAC1003—it slows RecB, relative to RecD, in a concentration-dependent manner, and sensitizes RecB to cut the DNA when RecD stops at the end of the DNA. The high similarity in the effects of the mutations and of NSAC1003 is consistent with the altered amino acids being very near the ATP binding site in RecB and, we infer, NSAC1003 binding to the RecB ATP site in a competitive manner (Figure 3). Slowing of the RecB helicase by the mutations and by NSAC1003 is thereby readily accounted for.
The mechanism for sensitization of RecB to cut the DNA when RecD stops is less obvious. Recent analysis of mutations in each subunit that reduce or block Chi hotspot activity shows that amino acids widely scattered throughout the large RecBCD complex are important for Chi to signal DNA cutting (19). Three points of contact (RecC–RecD, RecD–RecB and RecB–RecC) appear to act sequentially to transmit the Chi signal from the RecC tunnel to the RecB nuclease domain (Figure 1B–D). NSAC1003 and the RecB helicase mutants could bypass the early steps of this cascade (Chi recognition in the RecC tunnel, and Chi-bound RecC signaling RecD to stop) and directly sensitize RecB to cut the DNA when RecD stops in the same way that Chi signals wild-type RecD, when stopped, to induce RecB to cut where it is at that moment [∼4–6 nucleotides to the 3′ side of the Chi sequence bound in the RecC tunnel (Figures 1 and 4)] (30).
Two features of the cut products lead to further insights into the control of RecBCD nuclease activity. NSAC1003-induced cutting, like Chi-independent cutting by the RecB helicase mutants (Y803H and V804E), produced a smear of DNA fragments differing in length by ∼200–300 nucleotides (Figure 4C) (14). Chi-dependent cutting, however, occurs over only a 2- to 3-nucleotide range, ∼4–6 nucleotides 3′ of the Chi sequence (30). We infer that the signal from Chi acts much more quickly than the signal from RecD stopping at the end of DNA (by NSAC1003 or by the RecB helicase mutants). Specifically, we propose that the time it takes for the RecB nuclease domain to swing from its position on the ‘left’ side of RecBCD to the DNA exit of the tunnel in RecC (Figure 1D) is ∼1 ms after Chi's encounter but ∼200 ms after RecD stops in the RecB helicase mutants or in the presence of NSAC1003. These estimates follow from RecBCD’s unwinding rate of ∼1 bp/ms (4). Upon encountering Chi, the nuclease would swing to the tunnel exit in ∼1 ms, with variability from molecule to molecule to account for the spread of Chi-dependent cuts over ∼3 nucleotides. With NSAC1003 or the RecB helicase mutants, the swing would take much longer (∼200–300 ms) and allow RecB to advance ∼200–300 nucleotides before cutting. This interpretation accounts for the Chi-dependent band being much sharper than the NSAC1003- or RecB helicase mutant-dependent bands (Figure 4) (14). An alternative interpretation is that Chi may form a kinked configuration in the RecC tunnel (20) and slow or stop DNA translocation until the DNA is cut near Chi; if so, a longer nuclease swing-time would still result in cuts only a few nucleotides from the Chi sequence. In experiments without Chi, the DNA in the RecC tunnel would not typically be kinked when RecD reaches the DNA end, and NSAC1003- or helicase mutant-induced cuts would be spread over a larger region from the same nuclease swing-time. Whether Chi is kinked in the RecC tunnel at the moment of DNA cutting in an active RecBCD reaction has not been reported.
The second feature of the Chi-independent cuts is the non-zero extrapolate of the position of the cuts. For both NSAC1003 and the RecB helicase mutants, the extrapolate to zero substrate length is ∼–200 to –300 nucleotides (Figure 5B) (14). Thus, the cuts occur ∼200–300 nucleotides farther along the DNA than expected from the cuts being exactly a constant fraction of the substrate length. This distance is that expected from the rather slow movement of the nuclease domain inferred from the smear distribution noted above. Both features concur in predicting that RecB advances, after RecD stops at the DNA end, ∼200–300 nucleotides before cutting in the presence of NSAC1003 or in the RecB helicase mutants.
These features of NSAC1003 action on RecBCD should enable further biophysical analysis of RecBCD and AddAB helicase–nucleases, closely related DNA repair enzymes ubiquitous among bacteria but not reported in eukaryotes (31). In particular, the predicted relatively slow speed of swinging of the RecB nuclease domain should be easier to detect with NSAC1003 or the RecB helicase mutants than with Chi and wild-type RecBCD. RecBCD is a member of the superfamily 1 helicases, which, like many ATPases, have well-conserved motifs at their ATP-binding sites (32,33). RecB and RecD have a so-called Walker A box, which binds to ATP (Figure 3 and Supplementary Figure S3); mutations in each Walker A box, such as RecB K29Q and RecD K177Q (Figure 3 and Supplementary Figure S3), show that both are required for ATPase and helicase activity (28,29,7). Our inference that NSAC1003 binds to the RecB and RecD ATP-binding sites (Figures 2 and 3, Supplementary Figures S2 and S3) therefore predicts that NSAC1003 would inhibit other helicases. If so, this compound may be broadly useful in studying ATP-hydrolyzing molecular motors and ATPases in general, one of the largest classes of enzymes, including DNA and RNA polymerases.
Supplementary Material
ACKNOWLEDGEMENTS
We are grateful to Sue Amundsen for valuable advice on RecBCD and preparation of Figure 2C, and to Sue Amundsen, Randy Hyppa, Brett Kaiser and Rasi Subramaniam for helpful comments on the manuscript.
Notes
Present address: Ahmet C. Karabulut, Stowers Institute for Medical Research, Kansas City, MO, USA.
Present address: Ryan T. Cirz, Revagenix, San Diego, CA, USA.
Contributor Information
Ahmet C Karabulut, Division of Basic Sciences, Fred Hutchinson Cancer Research Center, Seattle, WA, USA.
Ryan T Cirz, Achaogen, Inc., San Francisco, CA, USA.
Andrew F Taylor, Division of Basic Sciences, Fred Hutchinson Cancer Research Center, Seattle, WA, USA.
Gerald R Smith, Division of Basic Sciences, Fred Hutchinson Cancer Research Center, Seattle, WA, USA.
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online.
FUNDING
National Institutes of Health of the United States of America [R01 GM031693, R35 GM118120 to G.R.S., P30 CA015704 to the Fred Hutchinson Cancer Research Center]. Funding for open access charge: NIH [R35 GM118120].
Conflict of interest statement. None declared.
REFERENCES
- 1. Smith G.R. How RecBCD and Chi promote DNA break repair and recombination – a molecular biologist's view. Microbiol. Mol. Biol. Rev. 2012; 76:217–228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Wigley D.B. Bacterial DNA repair: recent insights into the mechanism of RecBCD, AddAB and AdnAB. Nat. Rev. Microbiol. 2013; 11:9–13. [DOI] [PubMed] [Google Scholar]
- 3. Lohman T.M., Fazio N.T.. How does a helicase unwind DNA? Insights from RecBCD helicase. Bioessays. 2018; 40:e1800009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Taylor A., Smith G.R.. Unwinding and rewinding of DNA by the RecBC enzyme. Cell. 1980; 22:447–457. [DOI] [PubMed] [Google Scholar]
- 5. Ganesan S., Smith G.R.. Strand-specific binding to duplex DNA ends by the subunits of Escherichia coli RecBCD enzyme. J. Mol. Biol. 1993; 229:67–78. [DOI] [PubMed] [Google Scholar]
- 6. Singleton M.R., Dillingham M.S., Gaudier M., Kowalczykowski S.C., Wigley D.B.. Crystal structure of RecBCD enzyme reveals a machine for processing DNA breaks. Nature. 2004; 432:187–193. [DOI] [PubMed] [Google Scholar]
- 7. Taylor A.F., Smith G.R.. RecBCD enzyme is a DNA helicase with fast and slow motors of opposite polarity. Nature. 2003; 423:889–893. [DOI] [PubMed] [Google Scholar]
- 8. Ponticelli A.S., Schultz D.W., Taylor A.F., Smith G.R.. Chi-dependent DNA strand cleavage by RecBC enzyme. Cell. 1985; 41:145–151. [DOI] [PubMed] [Google Scholar]
- 9. Anderson D.G., Kowalczykowski S.C.. The translocating RecBCD enzyme stimulates recombination by directing RecA protein onto ssDNA in a χ regulated manner. Cell. 1997; 90:77–86. [DOI] [PubMed] [Google Scholar]
- 10. Spies M., Kowalczykowski S.C.. The RecA binding locus of RecBCD is a general domain for recruitment of DNA strand exchange proteins. Mol. Cell. 2006; 21:573–580. [DOI] [PubMed] [Google Scholar]
- 11. Dixon D.A., Kowalczykowski S.C.. Homologous pairing in vitro stimulated by the recombination hotspot, Chi. Cell. 1991; 66:361–371. [DOI] [PubMed] [Google Scholar]
- 12. Smith G.R. Conjugational recombination in E. coli: myths and mechanisms. Cell. 1991; 64:19–27. [DOI] [PubMed] [Google Scholar]
- 13. Smith G.R. Homologous recombination near and far from DNA breaks: alternative roles and contrasting views. Annu. Rev. Genet. 2001; 35:243–274. [DOI] [PubMed] [Google Scholar]
- 14. Amundsen S.K., Taylor A.F., Reddy M., Smith G.R.. Intersubunit signaling in RecBCD enzyme, a complex protein machine regulated by Chi hot spots. Genes Dev. 2007; 21:3296–3307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Taylor A.F., Amundsen S.K., Guttman M., Lee K.K., Luo J., Ranish J., Smith G.R.. Control of RecBCD enzyme activity by DNA binding- and Chi hotspot-dependent conformational changes. J. Mol. Biol. 2014; 426:3479–3499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Amundsen S.K., Smith G.R.. The RecB helicase–nuclease tether mediates Chi hotspot control of RecBCD enzyme. Nucleic Acids Res. 2019; 47:197–209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Handa N., Yang L., Dillingham M.S., Kobayashi I., Wigley D.B., Kowalczykowski S.C.. Molecular determinants responsible for recognition of the single-stranded DNA regulatory sequence, chi, by RecBCD enzyme. Proc. Natl. Acad. Sci. U.S.A. 2012; 109:8901–8906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Amundsen S.K., Sharp J.W., Smith G.R.. RecBCD enzyme ‘Chi recognition’ mutants recognize Chi recombination hotspots in the right DNA context. Genetics. 2016; 204:139–152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Amundsen S.K., Taylor A.F., Smith G.R.. Chi hotspot control of RecBCD helicase-nuclease by long-range intramolecular signaling. 2020; bioRxiv doi:16 June 2020, preprint: not peer reviewed 10.1101/2020.05.04.077495. [DOI] [PMC free article] [PubMed]
- 20. Cheng K., Wilkinson M., Chaban Y., Wigley D.B.. A conformational switch in response to Chi converts RecBCD from phage destruction to DNA repair. Nat. Struct. Mol. Biol. 2020; 27:71–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Smith G.R., Kunes S.M., Schultz D.W., Taylor A., Triman K.L.. Structure of Chi hotspots of generalized recombination. Cell. 1981; 24:429–436. [DOI] [PubMed] [Google Scholar]
- 22. Eichler D.C., Lehman I.R.. On the role of ATP in phosphodiester bond hydrolysis catalyzed by the RecBC deoxyribonuclease of Escherichia coli. J. Biol. Chem. 1977; 252:499–503. [PubMed] [Google Scholar]
- 23. Bikadi Z., Hazai E.. Application of the PM6 semi-empirical method to modeling proteins enhances docking accuracy of AutoDock. J. Cheminform. 2009; 1:15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Wilkinson M., Chaban Y., Wigley D.B.. Mechanism for nuclease regulation in RecBCD. Elife. 2016; 5:e18227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Morris C.M., Goodsell D.S.. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J. Comp. Chem. 1998; 19:1639–1662. [Google Scholar]
- 26. Solis F.J., Wets R.J.B.. Minimization by random search techniques. Math. Oper. Res. 1981; 6:19–30. [Google Scholar]
- 27. Feng B.Y., Shoichet B.K.. A detergent-based assay for the detection of promiscuous inhibitors. Nat. Protoc. 2006; 1:550–553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Hsieh S., Julin D.A.. Alteration by site-directed mutagenesis of the conserved lysine residue in the consensus ATP-binding sequence of the RecB protein of Escherichia coli. Nucleic Acids Res. 1992; 20:5647–5653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Korangy F., Julin D.A.. Enzymatic effects of a lysine-to-glutamine mutation in the ATP-binding consensus sequence in the RecD subunit of the RecBCD enzyme from Escherichia coli. J. Biol. Chem. 1992; 267:1733–1740. [PubMed] [Google Scholar]
- 30. Taylor A.F., Schultz D.W., Ponticelli A.S., Smith G.R.. RecBC enzyme nicking at Chi sites during DNA unwinding: location and orientation dependence of the cutting. Cell. 1985; 41:153–163. [DOI] [PubMed] [Google Scholar]
- 31. Cromie G.A. Phylogenetic ubiquity and shuffling of the bacterial RecBCD and AddAB recombination complexes. J. Bacteriol. 2009; 191:5076–5084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Singleton M.R., Dillingham M.S., Wigley D.B.. Structure and mechanism of helicases and nucleic acid translocases. Annu. Rev. Biochem. 2007; 76:23–50. [DOI] [PubMed] [Google Scholar]
- 33. Fairman-Williams M.E., Guenther U.P., Jankowsky E.. SF1 and SF2 helicases: family matters. Curr. Opin. Struct. Biol. 2010; 20:313–324. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.