

Abstract
The spiral ganglion neurons (SGNs) are the primary afferent neurons in the spiral ganglion (SG), while their degeneration or loss would cause sensorineural hearing loss. As a cardiac-derived hormone, atrial natriuretic peptide (ANP) plays a critical role in cardiovascular homeostasis through binding to its functional receptors (NPR-A and NPR-C). ANP and its receptors are widely expressed in the mammalian nervous system where they could be implicated in the regulation of multiple neural functions. Although previous studies have provided direct evidence for the presence of ANP and its functional receptors in the inner ear, their presence within the cochlear SG and their regulatory roles during auditory neurotransmission and development remain largely unknown. Based on our previous findings, we investigated the expression patterns of ANP and its receptors in the cochlear SG and dissociated SGNs and determined the influence of ANP on neurite outgrowth in vitro by using organotypic SG explants and dissociated SGN cultures from postnatal rats. We have demonstrated that ANP and its receptors are expressed in neurons within the cochlear SG of postnatal rat, while ANP may promote neurite outgrowth of SGNs via the NPR-A/cGMP/PKG pathway in a dose-dependent manner. These results indicate that ANP would play a role in normal neuritogenesis of SGN during cochlear development and represents a potential therapeutic candidate to enhance regeneration and regrowth of SGN neurites.
1. Introduction
Sensorineural hearing loss (SNHL) is a major health problem which affects millions of individuals worldwide. SNHL is associated with irreversible degeneration of the cochlear sensory cells within the auditory portion of the inner ear, including hair cells (HCs) and spiral ganglion neurons (SGNs). In the mammalian inner ear, the HCs in the organ of Corti function in transducing the sound mechanical stimulation into the primary acoustic signals [1–3], while the SGNs are the primary afferent neurons in the spiral ganglion (SG) and play a critical role in hearing, transmitting primary acoustic information from HCs to the higher auditory centers of the central nervous system (CNS) [4–6]. Loss of HCs, primarily resulting from noise trauma, ototoxic drugs, infections, aging, and genetic mutations, with sequential degeneration of SGNs, ultimately leads to permanent SNHL [7–15]. The current preferred treatment of SNHL for patients with profound HC loss and mostly intact SGNs includes cochlear implants, which uses electrode arrays to substitute for mechanosensory HCs in generating electrical impulses to the auditory nerve [16]. In order to promote regeneration and guiding of neurites from residual auditory neurons, many potential guidance cues are under research, due to their influence on neurite outgrowth behavior and subsequent performance of cochlear implants [6, 17–23].
SG is a peripheral cluster of both neurons and glial cells located in Rosenthal's canal, which coils around the cochlear modiolus and forms the auditory nerve. SGNs are divided into two subpopulations, type I and type II, according to their different morphologies, synaptic connections, and functions. Approximately 95% of SGNs are larger, bipolar, and myelinated type I neurons, which can be further subdivided into three subtypes (Type IA, Type IB, and Type IC), innervating the inner HCs with their peripheral dendrites to principally encode the auditory signals [24–26]. The remaining 5% of SGNs are smaller, pseudomonopolar, and nonmyelinated type II neurons, which innervate the outer HCs and some of the supporting cells to provide sensory feedback, controlling the sensitivity of the auditory epithelium to specific sound stimuli. Additionally, the perikaryons of all types of SGNs are enveloped by satellite glial cells, forming loose myelin around the type I neuronal somata. Both peripheral dendrites and intracochlear axons of type I SGNs are myelinated by Schwann cells, whereas their central axons from the peripheral-central glial transition zone (glia limitans) to the terminal synapses in the cochlear nucleus are myelinated by oligodendrocytes and astrocytes [27, 28]. It is necessary to understand the expression, function, and signaling interactions of the regulatory substances which affect axonal development and neuronal plasticity of primary auditory neurons, to offer optimal strategies of manipulating connections between sensory epithelium or implanted electrodes and neurites of SGNs, and eventually provide promising pharmacological targets that facilitate new and effective therapies for hearing impairment.
Atrial natriuretic peptide (ANP) is a 28 amino acid peptide predominantly synthesized and secreted by the cardiac atria and is the first member of the natriuretic peptide family [29], which also includes brain natriuretic peptide (BNP) and C-type natriuretic peptide (CNP). ANP interacts with two specific, high affinity natriuretic peptide receptors, NPR-A and NPR-C, on the plasma membrane of target cells to mediate its physiological effects [30–32]. The natriuretic peptide receptor-A (NPR-A, also known as NPR1 or GC-A) is a transmembrane receptor coupled to the particulate guanylyl cyclase (GC) which catalyzes the synthesis of the second messenger cyclic guanosine-3′,5′-monophosphate (cGMP). cGMP modulates the activity of specific effector molecules including cGMP-regulated isoforms of phosphodiesterases, cyclic-nucleotide-gated ion channels, and cGMP-dependent protein kinases G (PKG), which in turn regulate diverse biological responses associated with blood vessel tone, transepithelial ion transportation, neuronal excitability, neuronal development, and neurite pathfinding, and the sensory transduction pathways underlying olfaction and vision [31, 32]. The natriuretic peptide receptor-C (NPR-C), which lacks the GC domain, contributes to the clearance of ANP and other natriuretic peptides from the circulation through receptor-mediated internalization and degradation. In addition, evidence has shown that NPR-C can also affect other second messenger signaling by activating phospholipase C and inhibiting adenylyl cyclase [33].
In addition to the cardiovascular system, the tissue-specific distribution and function of ANP, NPR-A, and NPR-C have been established in several tissues including the kidney, adrenal, lung, adipose tissue, and retina. Furthermore, ANP and its receptors have been found in the CNS, leading us to speculate that ANP may function as a neuromodulator or neuropeptide involved in neuronal and glial functions [34–36]. Importantly, their presence in the secretory and sensory compartments of the rodent inner ear is well documented, suggesting ANP may act as a local hormone regulating the fluid and electrolyte balance in the inner ear [37–51]. Previous reports revealed that ANP receptors have been localized to the cochlear modiolus of the guinea pig [42] and rat SG [51]. However, little is known regarding the localization and functional roles of ANP and its receptors in the inner ear, and here, we have focused our attention on them.
In our previous study, we have already investigated the expression patterns of ANP and its receptors, which provided direct evidence for the presence and synthesis of ANP as well as its receptors in the cochlear SG [52, 53]. In our current study, we reassessed the distribution of ANP and its receptors in the cochlear SG as well as in dissociated SGNs, and determined the influence of ANP on neurite outgrowth in vitro by using organotypic SG explants and dissociated SGN cultures from postnatal rats. We have demonstrated that ANP and its receptors are expressed in neurons within the cochlear SG of postnatal rat, while ANP may promote neurite outgrowth of SGNs via the NPR-A/cGMP/PKG pathway in a dose-dependent manner.
2. Materials and Methods
2.1. Animals and Tissue Preparation
All experiments were approved by the Animal Care Committee of Fourth Military Medical University, China, on the care and use of Laboratory Animal for Research Purposes. All cochleae used in this investigation were obtained from postnatal day 3 (P3) or day 14 (P14) Sprague-Dawley rats provided by the Laboratory Animal Center of the Fourth Military Medical University. All rat pups were sacrificed by decapitation, and the skulls were opened midsagitally. With the aid of a dissecting microscope (SZX16; Olympus, Japan), the rat cochleae were removed from the temporal bone, washed in ice-cold Hank's Balanced Salt Solution (HBSS; Thermo Fisher Scientific, USA), and collected for further use.
2.2. Preparation of Cochlear Sections and Spiral Ganglion Neurons Culture
For cochlear cryosections, the cochleae from P14 rats were fixed with 4% paraformaldehyde (PFA) in phosphate buffer (PB; 0.1 M, pH 7.2) by perfusion via the round and oval windows and then incubated with the same fixative overnight at 4°C. The cochleae were decalcified in a 5% EDTA solution for 2 days, followed by cryoprotection in 30% sucrose solution overnight at 4°C. The samples were then embedded in Tissue-Tek OCT compound (Sakura Finetek, USA) at -20°C, sectioned into 12 μm thick midmodiolar cross-sections using a cryostat microtome (CM1850; Leica, Germany) and mounted on poly-L-lysine-coated slides.
Dissociated cultures of SGNs were prepared from P3 rat pups and maintained as described previously [54–56]. Briefly, each SG was isolated from the cochlea in ice-cold HBSS by sequential removal of the bony cochlear capsule, the spiral ligament, and the organ of the Corti, leaving the SGNs within the modiolus. The modiolus tissues were transferred into Ca2+/Mg2+ free HBSS with 0.25% trypsin and 0.1% collagenase type IV (all Thermo Fisher Scientific) at 37°C for 20 min to enzymatically dissociate the cells. The enzymatic reaction was quenched by the addition of 10% fetal bovine serum (FBS; Thermo Fisher Scientific). After three washes with culture medium, the tissues were mechanically dissociated by trituration with a fame-polished Pasteur pipette. The dissociated cells were resuspended in a neural maintenance medium consisting of Dulbecco's modified Eagle medium/Ham's F12 medium (DMEM/F12) supplemented with 1x B27, 1x N2, and 1% penicillin-streptomycin (all Thermo Fisher Scientific) and plated at a density of 1.0 × 106 cells/glass bottom dish previously coated with poly-L-lysine (0.1 mg/mL in 10 mM borate buffer, pH 8.4; Thermo Fisher Scientific) to adhere for 4 h at 37°C, 5% CO2, and 95% humidity. After attachment was confirmed under an inverted microscope (Eclipse TE2000-U; Nikon, Japan), 1 mL of neural maintenance medium was added in each neuronal cell culture and incubated for 48 h prior to fixation with 4% PFA for 20 min at room temperature (RT).
2.3. Expression Pattern Analysis of ANP and Its Receptors by Immunofluorescence
For immunohistochemistry, the cochlear sections and fixed SGNs cultures were washed with phosphate-buffered saline (PBS; 0.01 M, pH 7.4), blocked with 5% bovine serum albumin (BSA; Sigma-Aldrich, USA) and 0.1% Triton X-100 in PBS for 40 min at 37°C, and incubated overnight at 4°C with the following primary antibodies diluted in antibody solution (1% BSA and 0.1% Triton X-100 in PBS): polyclonal rabbit anti-ANP antibody (1 : 500; Cat# PA5-29559, Thermo Fisher Scientific), polyclonal rabbit anti-NPR-A antibody (1 : 500; Cat# PA5-29049, Thermo Fisher Scientific), polyclonal rabbit anti-NPR-C antibody (1 : 500; Cat# PA5-96947, Thermo Fisher Scientific), and monoclonal mouse anti-Tubulin β-III (TUJ1) antibody (1 : 500; Cat# ab78078, Abcam, UK). After washing, samples were treated with the appropriate secondary antibodies diluted in antibody solution for 2 h at RT: Alexa Fluor 488-conjugated donkey anti-mouse IgG (1 : 500; Cat# A-21202, Thermo Fisher Scientific) and Alexa Fluor 594-conjugated donkey anti-rabbit IgG (1 : 500; Cat# A-21207, Thermo Fisher Scientific). Each experiment also included a negative control where the primary antibody was omitted. After rinsing, specimens were treated with the nuclear stain, 4′,6-diamidino-2-phenylindole (DAPI; diluted 1 : 1000; Thermo Fisher Scientific) for 15 min at RT, mounted with Prolong Gold anti-fading mounting medium (Thermo Fisher Scientific), and subsequently examined under a spectral scanning laser confocal microscope (FV1000; Olympus, Japan). All images were saved as TIFF files using Olympus confocal software (FV10-ASW 4.2; Olympus) and processed with Adobe Photoshop CS6 (Adobe Systems, USA) for adjustments of brightness and/or contrast.
2.4. Spiral Ganglion Explants and Spiral Ganglion Neuron Cultures
The cochlea dissection procedures were performed as described in previous studies with slight modifications [56–59]. The cochleae from P3 rats were immersed in ice-cold HBSS; then, the cochlear capsule was opened by fine forceps and the membranous labyrinth was removed from the modiolus under a dissecting microscope. The spiral lamina containing the SG was carefully separated from the modiolus and cut into equal portions of 300~500 μm before being transferred to the culture dish. The Cell-Tak Cell and Tissue Adhesive (Corning, USA) precoated 15 mm glass bottom culture dishes (Advance BioScience, USA) were loaded with 100 μL primary attachment medium consisting of DMEM, 10% FBS, 25 mM HEPES buffer, and 1% penicillin-streptomycin (all Thermo Fisher Scientific). Then, each dissected explant was plated onto single-glass bottom dish, and the culture medium was carefully aspirated from dishes containing SG explants, leaving only 10 μL to allow the tissue to settle for 3 ~ 5 min. For three-dimensional culture, 100 μL of a 20% Matrigel (Corning) mixture diluted in the primary attachment medium was dropped on to the tissue explants directly and left the tissues adhering overnight at 37°C, 5% CO2, and 95% humidity. After attachment was confirmed, SG explants incubated in neural maintenance medium with or without 20 ng/mL recombinant brain-derived neurotrophic factor (BDNF; PeproTech, USA) were served as control cultures. Experimental cultures were incubated in neural maintenance medium supplemented with 100 nM or 1 μM of ANP (Caymanchem, USA), 1 μM of the membrane-permeable cGMP analogue 8-(4-chlorophenylthio) guanosine-3′,5′-cyclic monophosphate (8-pCPT-cGMP; Sigma-Aldrich), or 1 μM ANP plus 1 μM of the PKG inhibitor KT5823 (Sigma-Aldrich), respectively. For each condition, three cochlear neural explants were cultured for 7 days in a 37°C humidified incubator containing 5% CO2 prior to fixation for neurite outgrowth study, and culture medium was changed every other day.
The dissociated SGNs prepared as described above, were resuspended in neural maintenance medium and plated at a density of 2.0 × 105 cells/coated glass bottom dish. After attachment, cells from different experimental cultures were incubated in neural maintenance medium in the absence or in the presence of pharmacological reagents identical to those in SG explant cultures: 20 ng/mL BDNF, 100 nM ANP, 1 μM ANP, 1 μM 8-pCPT-cGMP, or 1 μM ANP plus 1 μM KT5823. For each condition, three culture dishes seeded with dissociated SGNs were fed with fresh medium every other day and cultured for 5 days prior to fixation.
2.5. Immunofluorescent Analysis of Neurite Outgrowth Spiral Ganglion Explants and Neurons
After the culture period, SG explants or SGN cells were fixed in 4% PFA for 20 min at RT. Both explants and cells were blocked with 5% BSA and 0.1% Triton X-100 in PBS, followed by incubation with anti-Tubulin β-III primary antibody (diluted 1 : 500) and Alexa Fluor 488-conjugated donkey anti-mouse IgG (diluted 1 : 500) to selectively stain the neural components of the explants or cells, then counterstained with DAPI (diluted 1 : 1000) to visualize the nuclei. All specimens in the glass bottom dish were mounted with Prolong Gold medium and examined under the confocal microscope.
In vitro Images of the immunostained cultures were analyzed by using ImageJ software (version 1.46r; NIH, USA) according to a previous study [60]. Neurite tracing was performed by using the “Analyze-Set Scale” function, the pixel unit of neurite length measurement was set in micrometers. Images were rendered with segmentation function, and neurite tracer function was applied choosing the starting point at the SGN cell bodies, resulting in a compiled skeleton render of all measured neurites. Only those neurites contained entirely within the image were analyzed. Neurite outgrowth from the SG explants was evaluated by measuring the number and lengths of the processes. Total number and the neurite length of the dissociated SGNs were also analyzed. Statistical analysis was performed using a one-way analysis of variance (ANOVA) followed by Bonferroni's post hoc test. Data presented in the text and figures are means and standard error of the mean (means ± SEM). Analysis was performed using the Statistical Program for Social Science software (SPSS, version 22.0; IBM Inc., USA). P values less than 0.05 (P < 0.05) were considered statistically significant.
3. Results
3.1. Distribution of ANP and Its Receptors in Spiral Ganglion and Spiral Ganglion Neurons of Postnatal Rats
To investigate the localizations of ANP and its receptors within the SG, we coimmunostained cochlear sections from P14 rats with antibodies against a neuronal marker, class III β-tubulin. No noticeable apical-to-basal gradients of ANP and its receptors immunoreactivities were observed in the SG regions along the length of the cochlear tonotopic axis. Hence, the midcochlear turn was taken as a representative of the entire length of the cochlea for analysis of the cellular localizations of ANP and its receptors. As shown in Figure 1, the immunoreactivity of ANP, NPR-A, or NPR-C was colocalized with β-III tubulin-positive somata of SGNs, respectively. ANP was predominantly immunoreactive in the neuronal perikarya, including the plasma membrane and cytoplasm of SGNs (Figure 1(a)). The distribution of NPR-A (Figure 1(b)) and NPR-C (Figure 1(c)) in SGNs was rather similar, and they were predominantly immunoreactive in the plasma membrane and cytoplasm of SGNs and appeared more pronounced in the cellular membrane. Some heterogeneity in the levels of immunoreactivities of ANP, NPR-A, and NPR-C were also observed, with less colocalization in a subpopulation of SGNs. No immunoreactivity was observed in the negative controls where the primary antibody was omitted (data not shown).
Figure 1.
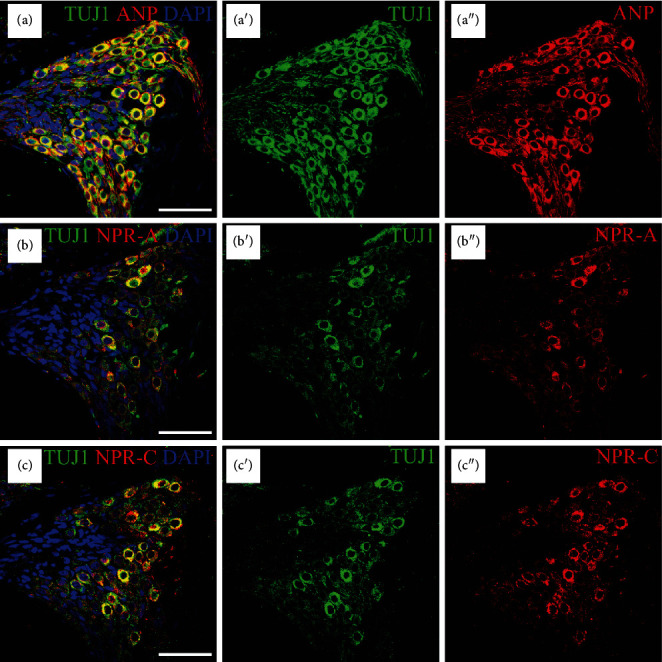
Immunolocalization of ANP, NPR-A, and NPR-C in SGNs within the SG of P14 rats. Merge and single-channel images of cochlear sections triple labeled with antibodies against neural marker TUJ1 (green), ANP/NPR-A/NPR-C (red), and DAPI (blue). (a) ANP was predominantly immunoreactive in the perikarya of SGNs. NPR-A (b) and NPR-C (c) were predominantly immunoreactive in the plasma membrane and cytoplasm of SGNs and appeared more pronounced in the cellular membrane. Scale bars = 50 μm.
To confirm the distribution pattern of ANP and its receptors in SGNs, we also performed immunohistochemistry on SG cell cultures using the same neuronal marker. As shown in Figure 2, the immunoreactivity of ANP, NPR-A, or NPR-C was colocalized with β-III tubulin-positive SGNs, respectively. In detail, the distribution of ANP and its receptors were immunoreactive all over each neuron including soma and neurites. No immunoreactivity was also observed in the negative controls where the primary antibody was omitted (data not shown).
Figure 2.
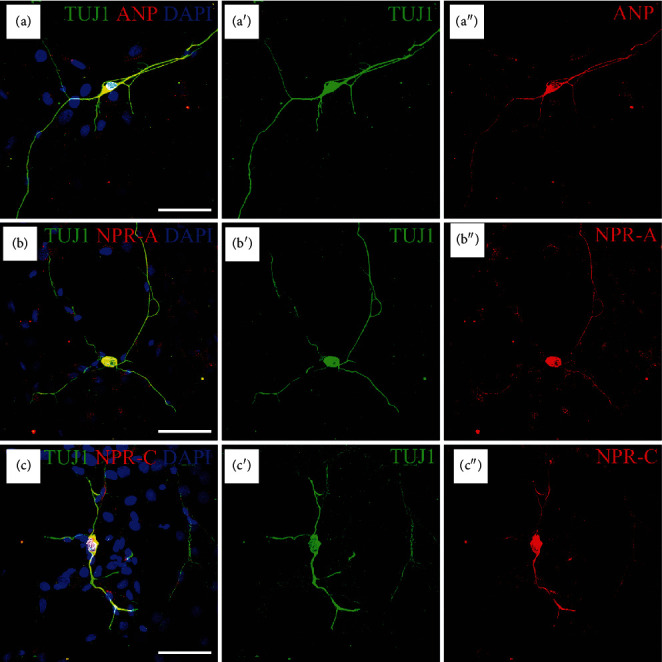
Immunolocalization of ANP, NPR-A, and NPR-C in dissociated SGNs from P3 rats. Merge and single-channel images of SG cells triple labeled with antibodies against TUJ1 (green), ANP/NPR-A/NPR-C (red), and DAPI (blue). The immunoreactivity of ANP (a), NPR-A (b), or NPR-C (c) was colocalized with β-III tubulin-positive SGNs, respectively, distributed in the neuronal soma and neurites. Scale bars = 50 μm.
3.2. Influence of ANP on Neurite Number and Neurite Length in Spiral Ganglion Explants
To determine the possible role of ANP in affecting neurite outgrowth of SGNs, we firstly quantified the number and length of neurites from SG explants from P3 rats maintained in culture medium supplemented with different reagents in vitro for 7 d. Representative images from different experimental cultures were shown in Figure 3. The SG explants incubated in culture medium without any supplement were used as a negative/baseline control, while explants incubated in culture medium supplemented with BDNF for trophic support of neurite outgrowth of SGNs were served as a positive control. Immunofluorescence and quantitative analysis of neuron-specific β-III tubulin-positive neurites revealed the number of neurites per explant was 46.0 ± 3.8, and the average neurite length was 1002.8 ± 155.5 μm for negative control samples (Figure 3(a)). As expected, abundant neurite sprouting and elongating were seen in explants treated with 20 ng/mL BDNF, and the average number and length of neurites from positive control samples were 191.7 ± 5.8 and 1645.8 ± 58.1 μm, respectively (Figure 3(b)). Strikingly, robust neurite extension from SG explants was seen after treatment with different dosage of ANP. The number and length of neurites were 92.0 ± 1.7 and 1438.3 ± 141.0 μm for 100 nM ANP-treated samples (Figure 3(c)) and were 151.0 ± 2.3 and 1551.2 ± 109.1 μm for 1 μM ANP-treated samples (Figure 3(d)), respectively. At any dosage of ANP (100 nM and 1 μM), a statistically significant increase in neurite outgrowth compared to the negative control was observed, while a significant difference was also found when compared to the positive control. To gain insight into the mechanism of ANP in promoting neurite outgrowth, we investigated whether this peptide acts through the GC-coupled receptors, NPR-A, which could elicit cGMP production. Results indicated that number (133.0 ± 3.79) and length (1374.8 ± 102.5 μm) of elongating neurites from SG explants treated with 1 μM 8-pCPT-cGMP, a cGMP analogue, were also significantly different from the negative and positive controls (Figure 3(e)). Treatment with 1 μM KT5823, a selective inhibitor of PKG, appeared to abrogate SGN neurite sprouting and outgrowth (47.3 ± 5.2 and 955.2 ± 109.2 μm) in the presence of 1 μM ANP (Figure 3(f)), and the resulting neurite outgrowth did not significantly differ from the negative control. Taken together, these results indicated that ANP could promote SG neurite outgrowth via the NPR-A/cGMP/PKG pathway in a dose-dependent manner.
Figure 3.
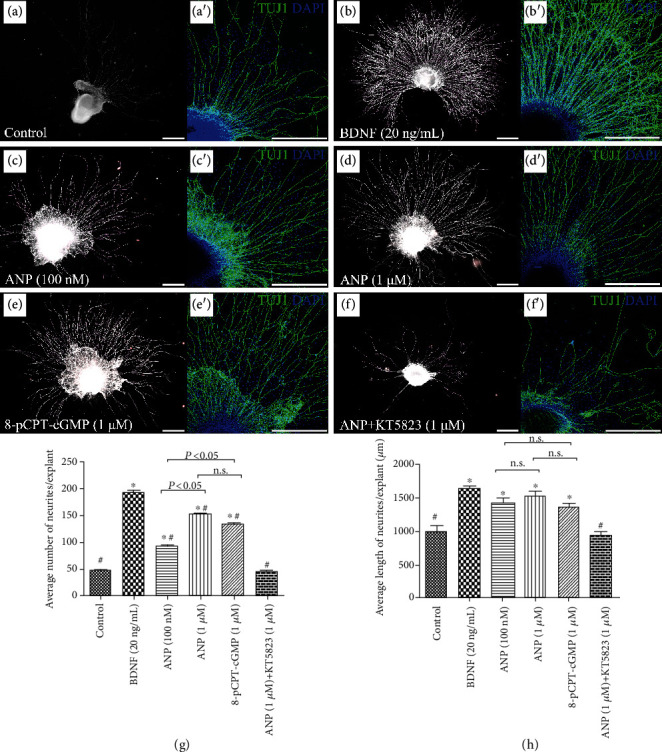
ANP promotes neurite outgrowth of SG explants in vitro. Immunohistochemical analysis of cochlear SG explants maintained in culture medium alone as a control (a), or treated with 20 ng/mL BDNF (b), 100 nM ANP (c), 1 μM ANP (d), 1 μM 8-pCPT-cGMP (e), or 1 μM ANP plus 1 μM KT5823 (f) for 7 d. Neurons were labeled with TuJ1 (green), while nuclei were stained with DAPI (blue). Scale bars = 500 μm. In each experimental culture, three cochlear neural explants were studied for neurite outgrowth, and the number of neurite outgrowth (g) and average neurite outgrowth length (h) of explants were calculated. Results are expressed as mean ± SEM (∗P < 0.05, versus negative control samples; #P < 0.05, versus positive control samples/BDNF; n.s., P > 0.05).
3.3. Influence of ANP on Cell Number and Neurite Length of Dissociated Spiral Ganglion Neurons
To validate the influence of ANP on the survival and neurite outgrowth of SGNs, we subsequently quantified the cell number and neurite length of dissociated SGNs from P3 rats maintained in culture medium with additives identical to those in explant cultures in vitro for 5 d. The dissociated neurons incubated in culture medium supplemented with or without 20 ng/mL BDNF were used as a positive or negative control, respectively. Representative images from different experimental cultures were shown in Figure 4. The average number of neurons per culture dish was 54.0 ± 1.5, and the average neurite length per neuron was 209.5 ± 19.5 μm in negative control samples (Figure 4(a)). Significantly increased number of neurons (78.3 ± 2.0) and elongating neurite outgrowth (763.2 ± 84.4 μm) were seen in cell cultures treated with 20 ng/mL BDNF (Figure 4(b)). The number of neurons and length of neurite were 69.0 ± 1.2 and 410.4 ± 29.5 μm for 100 nM ANP-treated neurons (Figure 4(c)), 70.0 ± 1.3 and 543.5 ± 31.5 μm for 1 μM ANP-treated neurons (Figure 4(d)), and 65.7 ± 2.9 and 475.2 ± 33.4 μm for 1 μM 8-pCPT-cGMP-treated neurons (Figure 4(e)), respectively. These observations were all significantly different from either negative or positive control samples. As expected, 1 μM ANP failed to either increase neuronal number or induce neurite outgrowth of SGNs (53.6 ± 1.2 and 255.5 ± 18.7 μm) in the presence of 1 μM KT5823 (Figure 4(f)). Collectively, these results indicated that the ANP/NPR-A/cGMP/PKG pathway may promote neuronal survival to some extent and would enhance neurite outgrowth of SGNs in a dose-dependent manner.
Figure 4.
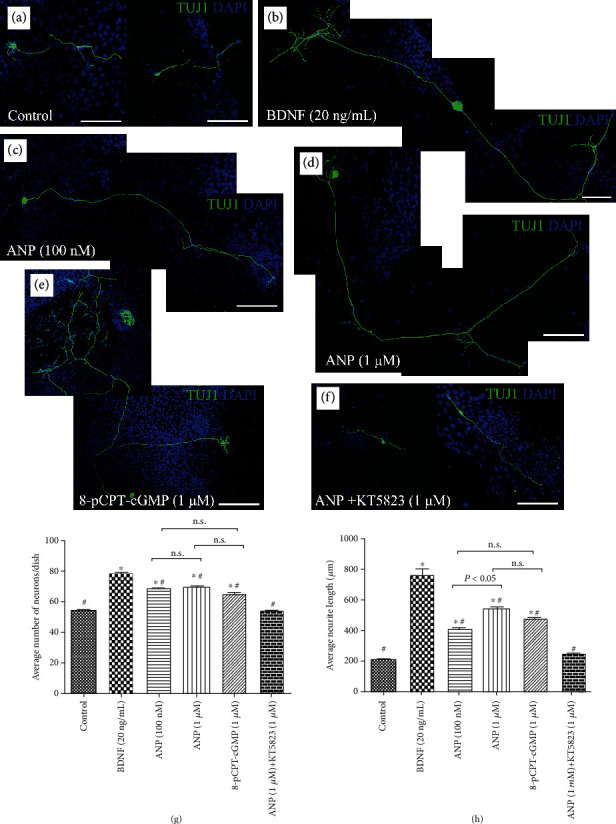
ANP promotes neurite outgrowth of dissociated SGNs in vitro. Immunohistochemical analysis of dissociated SGNs maintained in culture medium alone (a), or treated with 20 ng/mL BDNF (b), 100 nM ANP (c), 1 μM ANP (d), 1 μM 8-pCPT-cGMP (e), or 1 μM ANP plus 1 μM KT5823 (f) for 5 d. Neurons were labeled with TuJ1 (green), while nuclei were stained with DAPI (blue). Scale bars = 100 μm. In each experimental culture, three SGNs seeded-culture dishes were studied for neurite outgrowth, and the cell number (g) and average neurite outgrowth length (h) of dissociated SGNs were calculated. Results are expressed as mean ± SEM (∗P < 0.05, versus negative control samples; #P < 0.05, versus positive control samples/BDNF; n.s., P > 0.05).
Interestingly, when treated with ANP, a number of dissociated SGNs made more than one branches from a single neurite. This effect could be apparently seen in BDNF, ANP, and cGMP analogue 8-pCPT-cGMP-treated neurons, indicating that neurotrophins and ANP signaling could exert a trophic support for neurite outgrowth and axon branching (Figures 4(b)–4(e)). Thus, the effect of axonal branching or pathfinding mediated by the ANP/NPR-A/cGMP/PKG pathway still need more comprehensive evaluation during cochlear development for better understanding the exquisite mechanisms underlying the assembly of auditory circuits.
4. Discussion
To promote the regeneration of SGN and to guide the neurite outgrowth of SGN is a critical scientific question in the hearing research fields. In recent years, many previous reports used transcription regulation, biomaterials, electrical stimulation, and magnetic regulation to promote the regeneration and maturation of SGNs and other nervous tissues [23, 57, 60–67]. In our previous researches, we have already studied the expression patterns of ANP and its receptors by immunohistochemical and molecular biological analyses, with the aim of identifying their cellular localizations, expression levels, and the potential functions within the SG of postnatal rat. The immunoreactivity of ANP and its receptors distributed in SGNs and perineuronal glial cells, with differential expression levels of both mRNA and protein in the rat SG during postnatal development. All these data provided a general idea about the temporal and spatial expression profiles of ANP and its receptors within the cochlear SG, suggesting possible roles for ANP in modulating neuronal and glial functions within the SG during auditory neurotransmission and development [52, 53].
In our current study, we performed immunofluorescent analysis on cochlear cryosections as well as dissociated SGNs to validate the distribution of ANP and its receptors in SGNs. Additionally, the influence of ANP on neurite outgrowth was also determined by using organotypic SG explants and dissociated SGN cultures from postnatal rats in vitro. We have demonstrated that ANP and its receptors localize in SGNs within cochlear sections and dissociated cultures. ANP, NPR-A, and NPR-C were predominantly immunoreactive in the neuronal perikarya of SGNs, which was consistent with our previous results [52, 53].
ANP, as well as the other two major components of the natriuretic peptide family, BNP and CNP, together with their receptors (NPRs), are widely expressed in the neuronal and glial elements within rodent and mammalian CNS. Additionally, circumstantial evidences indicate that natriuretic peptides and receptors are also distributed in the neural region of peripheral sensory organs, particularly in the sensory ganglia including the dorsal root ganglion (DRG), as well as the ganglia in the retina and inner ear [42, 51, 68–73]. All they could potentially be involved in the regulation of several aspects of neuronal functions such as synaptic transmission and information processing, neural development, neuroprotection, neuroinflammation, neurovascular and blood-brain barrier integrity, and brain fluid homeostasis [34–36]. Especially, many recent researches have shown the significance of the cGMP signaling pathway for neuronal development and neurite pathfinding both in the central and peripheral nervous system, thus providing some hints on the possible functions of natriuretic peptides in axonal development. cGMP signaling elicited by activation of the transmembrane GC-coupled natriuretic peptide receptor NPR-B (also known as NPR2 or GC-B) by the ligand CNP control sensory axon bifurcation of DRG neurons entering the spinal cord [74–83]. Interestingly, the same phenomenon was also found in the cranial sensory ganglion neurons entering the hindbrain including cochlear SGNs [84–88]. Likewise, the mouse lacking cGMP-dependent protein kinase 1 (PKG1) or NPR-B has a defect in the central axonal projection of the DRG sensory neurons [74–78, 81–83] or SGNs [84–88], respectively. Consequently, all these results emphasize a strong significance of the CNP/NPR-B/cGMP/PKG1 pathway regulating neurite outgrowth or pathfinding during neuronal development.
Taking together the results of previous studies and our current work demonstrating colocalization of ANP and its receptors in cochlear SGNs, we hypothesize that ANP may be involved in the regulation of axonal development in the auditory circuits, since ANP also activates a cGMP-dependent signaling cascade upon binding to NPR-A. To test this hypothesis, the influence of ANP on neurite outgrowth was determined in organotypic SG explants and dissociated SGN cultures from postnatal rats in vitro. Since neurotrophins play a critical role in SGN development and maintenance and have been shown to promote SGN survival and enhance neurite elongation [89, 90], SG explants and dissociated SGN cultures incubated with BDNF additive were used as a positive control. Without any additional trophic support, neurite outgrowth from SGNs from explants or dissociated cultures in negative control samples is very limited. The observed neurite outgrowth induced by ANP is dose dependent and significantly different from that of negative control cultures. Furthermore, additional experiments using cell permeable cGMP analogue or a PKG inhibitor further confirmed the role of the ANP/NPRA/cGMP/PKG pathway in neurite elongation, since this activity can be replicated by using 8-pCPT-cGMP and abolished by KT-5823. Taken together, all our data indicated that ANP could enhance neurite outgrowth of SGNs via the NPR-A/cGMP/PKG pathway in a dose-dependent manner and may promote neuronal survival to some extent.
In conclusion, we have demonstrated that ANP and its receptors are expressed in neurons within the cochlear SG of postnatal rat, while ANP may promote neurite outgrowth of SGNs via the NPR-A/cGMP/PKG pathway in a dose-dependent manner. These results indicate that ANP would play a role in normal neuritogenesis of SGN during cochlear development and represents a potential therapeutic candidate to enhance regeneration and regrowth of SGN neurites. Our data offer a good starting point to identify these potential elements and generate new hypotheses; functional experiments must be performed for any gene of interest in such signaling pathway, as we have for NPR1 (NPR-A), which inspire us to investigate the modulatory effects of ANP on several types of neuronal functions within the SG may be involved in through its cell surface receptors. The role of ANP in the regulation of neural development may support it as a key regulator for auditory development and regeneration within the cochlear SG. Manipulation of cGMP levels and activation of PKG by activating ANP and receptor signals represent a potential therapeutic approach to support SGN survival as well as enhance regeneration and regrowth of SGN neurites, which promise to be a fruitful area for developing new and effective therapies for hearing impairment.
Acknowledgments
This research was supported by grants from the National Basic Research Program of China (973 Project, 2014CB541700), National Natural Science Foundation of China (81870732 and 81803187), Key Research and Development Program of Shaanxi Province (2018PT-01), Innovation Capability Support Program of Shaanxi Province (2017ZDXM-SF-061), Natural Science Foundation of Shaanxi Province (2019JQ-434), and Disciplinary Research Startup Program of Xijing Hospital in Fourth Military Medical University (XJZT19ML23 and XJZT18ML54).
Contributor Information
Jian-hua Qiu, Email: qiujh@fmmu.edu.cn.
Ding-jun Zha, Email: zhadjun@fmmu.edu.cn.
Data Availability
The data used to support the findings of this study are available from the corresponding author upon request.
Conflicts of Interest
The authors declare that there are no conflicts of interest regarding the publication of this article.
Authors' Contributions
Fei Sun, Ke Zhou, Ke-yong Tian, and Jie Wang contributed equally to this work.
References
- 1.Liu Y., Qi J., Chen X., et al. Critical role of spectrin in hearing development and deafness. Science Advances. 2019;5(4, article eaav7803) doi: 10.1126/sciadv.aav7803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Wang Y., Li J., Yao X., et al. Loss of CIB2 causes profound hearing loss and abolishes mechanoelectrical transduction in mice. Frontiers in Molecular Neuroscience. 2017;10:p. 401. doi: 10.3389/fnmol.2017.00401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zhu C., Cheng C., Wang Y., et al. Loss of ARHGEF6 causes hair cell stereocilia deficits and hearing loss in mice. Frontiers in Molecular Neuroscience. 2018;11:p. 362. doi: 10.3389/fnmol.2018.00362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Nayagam B. A., Muniak M. A., Ryugo D. K. The spiral ganglion: connecting the peripheral and central auditory systems. Hearing Research. 2011;278(1-2):2–20. doi: 10.1016/j.heares.2011.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Rusznák Z., Szucs G. Spiral ganglion neurones: an overview of morphology, firing behaviour, ionic channels and function. Pflügers Archiv. 2009;457(6):1303–1325. doi: 10.1007/s00424-008-0586-2. [DOI] [PubMed] [Google Scholar]
- 6.Zhang K. D., Coate T. M. Recent advances in the development and function of type II spiral ganglion neurons in the mammalian inner ear. Seminars in Cell & Developmental Biology. 2017;65:80–87. doi: 10.1016/j.semcdb.2016.09.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Shepherd R. K., Hardie N. A. Deafness-induced changes in the auditory pathway: implications for cochlear implants. Audiology & Neuro-Otology. 2001;6(6):305–318. doi: 10.1159/000046843. [DOI] [PubMed] [Google Scholar]
- 8.He Z. H., Zou S. Y., Li M., et al. The nuclear transcription factor FoxG1 affects the sensitivity of mimetic aging hair cells to inflammation by regulating autophagy pathways. Redox Biology. 2020;28, article 101364 doi: 10.1016/j.redox.2019.101364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Gao S., Cheng C., Wang M., et al. Blebbistatin inhibits neomycin-induced apoptosis in hair cell-like HEI-OC-1 cells and in cochlear hair cells. Frontiers in Cellular Neuroscience. 2020;13:p. 590. doi: 10.3389/fncel.2019.00590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zhang Y., Li W., He Z., et al. Pre-treatment with fasudil prevents neomycin-induced hair cell damage by reducing the accumulation of reactive oxygen species. Frontiers in Molecular Neuroscience. 2019;12:p. 264. doi: 10.3389/fnmol.2019.00264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Liu W., Xu X., Fan Z., et al. Wnt signaling activates TP53-induced glycolysis and apoptosis regulator and protects against cisplatin-induced spiral ganglion neuron damage in the mouse cochlea. Antioxidants & Redox Signaling. 2019;30(11):1389–1410. doi: 10.1089/ars.2017.7288. [DOI] [PubMed] [Google Scholar]
- 12.Liu L., Chen Y., Qi J., et al. Wnt activation protects against neomycin-induced hair cell damage in the mouse cochlea. Cell Death & Disease. 2016;7(3, article e2136) doi: 10.1038/cddis.2016.35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Li H., Song Y., He Z., et al. Meclofenamic acid reduces reactive oxygen species accumulation and apoptosis, inhibits excessive autophagy, and protects hair cell-like HEI-OC1 cells from cisplatin-induced damage. Frontiers in Cellular Neuroscience. 2018;12:p. 139. doi: 10.3389/fncel.2018.00139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zhang S., Zhang Y., Dong Y., et al. Knockdown of Foxg1 in supporting cells increases the trans-differentiation of supporting cells into hair cells in the neonatal mouse cochlea. Cellular and Molecular Life Sciences. 2020;77(7):1401–1419. doi: 10.1007/s00018-019-03291-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.He Z., Guo L., Shu Y., et al. Autophagy protects auditory hair cells against neomycin-induced damage. Autophagy. 2017;13(11):1884–1904. doi: 10.1080/15548627.2017.1359449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Guo R., Ma X., Liao M., et al. Development and application of cochlear implant-based electric-acoustic stimulation of spiral ganglion neurons. ACS Biomaterials Science & Engineering. 2019;5(12):6735–6741. doi: 10.1021/acsbiomaterials.9b01265. [DOI] [PubMed] [Google Scholar]
- 17.Li G., Chen K., You D., et al. Laminin-coated electrospun regenerated silk fibroin mats promote neural progenitor cell proliferation, differentiation, and survival in vitro. Frontiers in Bioengineering and Biotechnology. 2019;7:p. 190. doi: 10.3389/fbioe.2019.00190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Fan K. Q., Li Y. Y., Wang H. L., et al. Stress-induced metabolic disorder in peripheral CD4+ T cells leads to anxiety-like behavior. Cell. 2019;179(4):864–879.e19. doi: 10.1016/j.cell.2019.10.001. [DOI] [PubMed] [Google Scholar]
- 19.Fang Q., Zhang Y., Chen X., et al. Three-dimensional graphene enhances neural stem cell proliferation through metabolic regulation. Frontiers in Bioengineering and Biotechnology. 2020;7:p. 436. doi: 10.3389/fbioe.2019.00436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhao J., Tang M., Cao J., et al. Structurally tunable reduced graphene oxide substrate maintains mouse embryonic stem cell pluripotency. Advanced Science. 2019;6(12, article 1802136) doi: 10.1002/advs.201802136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Li D., Yan X., Hu Y., et al. Two-photon image tracking of neural stem cells via iridium complexes encapsulated in polymeric nanospheres. ACS Biomaterials Science & Engineering. 2019;5(3):1561–1568. doi: 10.1021/acsbiomaterials.8b01231. [DOI] [PubMed] [Google Scholar]
- 22.Han S., Xu Y., Sun J., et al. Isolation and analysis of extracellular vesicles in a Morpho butterfly wing- integrated microvortex biochip. Biosensors and Bioelectronics. 2020;154, article 112073 doi: 10.1016/j.bios.2020.112073. [DOI] [PubMed] [Google Scholar]
- 23.Tang M., Li J., He L., et al. Transcriptomic profiling of neural stem cell differentiation on graphene substrates. Colloids and Surfaces B: Biointerfaces. 2019;182:p. 110324. doi: 10.1016/j.colsurfb.2019.06.054. [DOI] [PubMed] [Google Scholar]
- 24.Sun S., Babola T., Pregernig G., et al. Hair cell mechanotransduction regulates spontaneous activity and spiral ganglion subtype specification in the auditory system. Cell. 2018;174(5):1247–1263.e15. doi: 10.1016/j.cell.2018.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Shrestha B. R., Chia C., Wu L., Kujawa S. G., Liberman M. C., Goodrich L. V. Sensory neuron diversity in the inner ear is shaped by activity. Cell. 2018;174(5):1229–1246.e17. doi: 10.1016/j.cell.2018.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Petitpré C., Wu H., Sharma A., et al. Neuronal heterogeneity and stereotyped connectivity in the auditory afferent system. Nature Communications. 2018;9(1, article 3691) doi: 10.1038/s41467-018-06033-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Toesca A. Central and peripheral myelin in the rat cochlear and vestibular nerves. Neuroscience Letters. 1996;221(1):21–24. doi: 10.1016/S0304-3940(96)13273-0. [DOI] [PubMed] [Google Scholar]
- 28.Jeon E. J., Xu N., Xu L., Hansen M. R. Influence of central glia on spiral ganglion neuron neurite growth. Neuroscience. 2011;177:321–334. doi: 10.1016/j.neuroscience.2011.01.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.de Bold A. J., Borenstein H. B., Veress A. T., Sonnenberg H. A rapid and potent natriuretic response to intravenous injection of atrial myocardial extract in rats. Life Sciences. 1981;28(1):89–94. doi: 10.1016/0024-3205(81)90370-2. [DOI] [PubMed] [Google Scholar]
- 30.Levin E. R., Gardner D. G., Samson W. K. Natriuretic peptides. The New England Journal of Medicine. 1998;339(5):321–328. doi: 10.1056/NEJM199807303390507. [DOI] [PubMed] [Google Scholar]
- 31.Potter L. R., Abbey-Hosch S., Dickey D. M. Natriuretic peptides, their receptors, and cyclic guanosine monophosphate-dependent signaling functions. Endocrine Reviews. 2006;27(1):47–72. doi: 10.1210/er.2005-0014. [DOI] [PubMed] [Google Scholar]
- 32.Potter L. R., Yoder A. R., Flora D. R., Antos L. K., Dickey D. M. Natriuretic peptides: their structures, receptors, physiologic functions and therapeutic applications. In: Schmidt H. H. H. W., Hofmann F., Stasch J. P., editors. cGMP: Generators, Effectors and Therapeutic Implications. Handbook of Experimental Pharmacology, vol 191. Berlin, Heidelberg: Springer; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Rose R. A., Giles W. R. Natriuretic peptide C receptor signalling in the heart and vasculature. The Journal of Physiology. 2008;586(2):353–366. doi: 10.1113/jphysiol.2007.144253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Cao L.-H., Yang X.-L. Natriuretic peptides and their receptors in the central nervous system. Progress in Neurobiology. 2008;84(3):234–248. doi: 10.1016/j.pneurobio.2007.12.003. [DOI] [PubMed] [Google Scholar]
- 35.Prado J., Baltrons M. A., Pifarré P., García A. Glial cells as sources and targets of natriuretic peptides. Neurochemistry International. 2010;57(4):367–374. doi: 10.1016/j.neuint.2010.03.004. [DOI] [PubMed] [Google Scholar]
- 36.Mahinrad S., de Craen A. J. M., Yasar S., van Heemst D., Sabayan B. Natriuretic peptides in the central nervous system: novel targets for cognitive impairment. Neuroscience & Biobehavioral Reviews. 2016;68:148–156. doi: 10.1016/j.neubiorev.2016.05.022. [DOI] [PubMed] [Google Scholar]
- 37.Yoon Y. J., Lee E. J., Kim S. H. Synthesis of atrial natriuretic peptide in the rabbit inner ear. The Laryngoscope. 2012;122(7):1605–1608. doi: 10.1002/lary.23235. [DOI] [PubMed] [Google Scholar]
- 38.Koch T., Gloddek B., Gutzke S. Binding sites of atrial natriuretic peptide (ANP) in the mammalian cochlea and stimulation of cyclic GMP synthesis. Hearing Research. 1992;63(1-2):197–202. doi: 10.1016/0378-5955(92)90085-2. [DOI] [PubMed] [Google Scholar]
- 39.Krause G., Meyer zum Gottesberge A. M., Wolfram G., Gerzer R. Transcripts encoding three types of guanylyl-cyclase-coupled trans-membrane receptors in inner ear tissues of guinea pigs. Hearing Research. 1997;110(1-2):95–106. doi: 10.1016/S0378-5955(97)00064-6. [DOI] [PubMed] [Google Scholar]
- 40.Meyer zum Gottesberge A., Lamprecht J. Localization of the atrial natriuretic peptide binding sites in the inner ear tissue—possibly an additional regulating system. Acta Oto-Laryngologica. Supplementum. 1989;468:53–57. doi: 10.3109/00016488909139021. [DOI] [PubMed] [Google Scholar]
- 41.Meyer zum Gottesberge A. M., Gagelmann M., Forssmann W. G. Atrial natriuretic peptide-like immunoreactive cells in the guinea pig inner ear. Hearing Research. 1991;56(1-2):86–92. doi: 10.1016/0378-5955(91)90157-5. [DOI] [PubMed] [Google Scholar]
- 42.Lamprecht J., Meyer zum Gottesberge A. M. The presence and localization of receptors for atrial natriuretic peptide in the inner ear of the guinea pig. Archives of Oto-Rhino-Laryngology. 1988;245(5):300–301. doi: 10.1007/BF00464636. [DOI] [PubMed] [Google Scholar]
- 43.Meyer zum Gottesberge A., Schleicher A., Drummer C., Gerzer R. The volume protective natriuretic peptide system in the inner ear comparison between vestibular and cochlear compartments. Acta Oto-Laryngologica. Supplementum. 1995;520, Part 1:170–173. doi: 10.3109/00016489509125219. [DOI] [PubMed] [Google Scholar]
- 44.Qiao L., Han Y., Zhang P., Cao Z., Qiu J. Detection of atrial natriuretic peptide and its receptor in marginal cells and cochlea tissues from the developing rats. Neuro Endocrinology Letters. 2011;32(2):187–192. [PubMed] [Google Scholar]
- 45.Seebacher T., Beitz E., Kumagami H., Wild K., Ruppersberg J. P., Schultz J. E. Expression of membrane-bound and cytosolic guanylyl cyclases in the rat inner ear. Hearing Research. 1999;127(1-2):95–102. doi: 10.1016/S0378-5955(98)00176-2. [DOI] [PubMed] [Google Scholar]
- 46.Suzuki M., Kitano H., Kitanishi T., et al. RT-PCR analysis of mRNA expression of natriuretic peptide family and their receptors in rat inner ear. Molecular Brain Research. 1998;55(1):165–168. doi: 10.1016/S0169-328X(98)00016-3. [DOI] [PubMed] [Google Scholar]
- 47.Yoon Y.-J., Hellström S. Immunohistochemical localization of α-atrial natriuretic polypeptide in the rat cochlea. Acta Oto-Laryngologica. 2009;112(4):604–610. doi: 10.3109/00016489209137448. [DOI] [PubMed] [Google Scholar]
- 48.Yoon Y. J., Anniko M. Distribution of alpha-ANP in the cochlea and the vestibular organs. ORL. 1994;56(2):73–77. doi: 10.1159/000276613. [DOI] [PubMed] [Google Scholar]
- 49.Yoon Y. J., Lee E. J., Hellstrom S., Kim J. S. Atrial natriuretic peptide modulates auditory brainstem response of rat. Acta Oto-Laryngologica. 2015;135(12):1293–1297. doi: 10.3109/00016489.2015.1073354. [DOI] [PubMed] [Google Scholar]
- 50.Chen H. X., Wang J. L., Liu Q. C., Qiu J. H. Distribution and location of immunoreactive atrial natriuretic peptides in cochlear stria vascularis of guinea pig. Chinese Medical Journal. 1994;107(1):53–56. [PubMed] [Google Scholar]
- 51.Furuta H., Mori N., Luo L., Ryan A. F. Detection of mRNA encoding guanylate cyclase A/atrial natriuretic peptide receptor in the rat cochlea by competitive polymerase chain reaction and in situ hybridization. Hearing Research. 1995;92(1-2):78–84. doi: 10.1016/0378-5955(95)00203-0. [DOI] [PubMed] [Google Scholar]
- 52.Sun F., Zhou K., Wang S., et al. Expression and localization of atrial natriuretic peptide and its receptors in rat spiral ganglion neurons. Brain Research Bulletin. 2013;95:28–32. doi: 10.1016/j.brainresbull.2013.04.001. [DOI] [PubMed] [Google Scholar]
- 53.Sun F., Zhou K., Wang S., Liang P., Zhu M., Qiu J. Expression patterns of atrial natriuretic peptide and its receptors within the cochlear spiral ganglion of the postnatal rat. Hearing Research. 2014;309:103–112. doi: 10.1016/j.heares.2013.11.010. [DOI] [PubMed] [Google Scholar]
- 54.Xu N., Engbers J., Khaja S., Xu L., Clark J. J., Hansen M. R. Influence of cAMP and protein kinase A on neurite length from spiral ganglion neurons. Hearing Research. 2012;283(1-2):33–44. doi: 10.1016/j.heares.2011.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Atkinson P. J., Cho C. H., Hansen M. R., Green S. H. Activity of all JNK isoforms contributes to neurite growth in spiral ganglion neurons. Hearing Research. 2011;278(1-2):77–85. doi: 10.1016/j.heares.2011.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Sun G., Liu W., Fan Z., et al. The three-dimensional culture system with Matrigel and neurotrophic factors preserves the structure and function of spiral ganglion neuron in vitro. Neural Plasticity. 2016;2016:15. doi: 10.1155/2016/4280407.4280407 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Waqas M., Sun S., Xuan C., et al. Bone morphogenetic protein 4 promotes the survival and preserves the structure of flow-sorted Bhlhb5+ cochlear spiral ganglion neurons in vitro. Scientific Reports. 2017;7(1, article 3506) doi: 10.1038/s41598-017-03810-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Meas S. J., Nishimura K., Scheibinger M., Dabdoub A. In vitro methods to cultivate spiral ganglion cells, and purification of cellular subtypes for induced neuronal reprogramming. Frontiers in Neuroscience. 2018;12:p. 822. doi: 10.3389/fnins.2018.00822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Leitmeyer K., Glutz A., Setz C., et al. Simvastatin results in a dose-dependent toxic effect on spiral ganglion neurons in an in vitro organotypic culture assay. BioMed Research International. 2016;2016:7. doi: 10.1155/2016/3580359.3580359 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Kempfle J. S., Nguyen K., Hamadani C., et al. Bisphosphonate-linked TrkB agonist: cochlea-targeted delivery of a neurotrophic agent as a strategy for the treatment of hearing loss. Bioconjugate Chemistry. 2018;29(4):1240–1250. doi: 10.1021/acs.bioconjchem.8b00022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Han S., Sun J., He S., Tang M., Chai R. The application of graphene-based biomaterials in biomedicine. American Journal of Translational Research. 2019;11(6):3246–3260. [PMC free article] [PubMed] [Google Scholar]
- 62.Yan W., Liu W., Qi J., et al. A three-dimensional culture system with Matrigel promotes purified spiral ganglion neuron survival and function in vitro. Molecular Neurobiology. 2018;55(3):2070–2084. doi: 10.1007/s12035-017-0471-0. [DOI] [PubMed] [Google Scholar]
- 63.Yang Y., Zhang Y., Chai R., Gu Z. Designs of biomaterials and microenvironments for neuroengineering. Neural Plasticity. 2018;2018:10. doi: 10.1155/2018/1021969.1021969 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Liu Z., Tang M., Zhao J., Chai R., Kang J. Looking into the future: toward advanced 3D biomaterials for stem-cell-based regenerative medicine. Advanced Materials. 2018;30(17, article e1705388) doi: 10.1002/adma.201705388. [DOI] [PubMed] [Google Scholar]
- 65.Xia L., Zhu W., Wang Y., He S., Chai R. Regulation of neural stem cell proliferation and differentiation by graphene- based biomaterials. Neural Plasticity. 2019;2019:11. doi: 10.1155/2019/3608386.3608386 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Guo R., Zhang S., Xiao M., et al. Accelerating bioelectric functional development of neural stem cells by graphene coupling: implications for neural interfacing with conductive materials. Biomaterials. 2016;106:193–204. doi: 10.1016/j.biomaterials.2016.08.019. [DOI] [PubMed] [Google Scholar]
- 67.Shang L., Yu Y., Gao W., et al. Bio-inspired anisotropic wettability surfaces from dynamic ferrofluid assembled templates. Advanced Functional Materials. 2018;28(7, article 1705802) doi: 10.1002/adfm.201705802. [DOI] [Google Scholar]
- 68.Abdelalim E. M., Masuda C., Tooyama I. Expression of natriuretic peptide-activated guanylate cyclases by cholinergic and dopaminergic amacrine cells of the rat retina. Peptides. 2008;29(4):622–628. doi: 10.1016/j.peptides.2007.11.021. [DOI] [PubMed] [Google Scholar]
- 69.Abdelalim E. M., Bellier J. P., Tooyama I. Expression of NPR-B in neurons of the dorsal root ganglia of the rat. Peptides. 2013;43:56–61. doi: 10.1016/j.peptides.2013.02.013. [DOI] [PubMed] [Google Scholar]
- 70.Li Z. W., Wu B., Ye P., Tan Z. Y., Ji Y. H. Brain natriuretic peptide suppresses pain induced by BmK I, a sodium channel-specific modulator, in rats. The Journal of Headache and Pain. 2016;17(1):p. 90. doi: 10.1186/s10194-016-0685-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Marchenkova A., Vilotti S., Fabbretti E., Nistri A. Brain natriuretic peptide constitutively downregulates P2X3 receptors by controlling their phosphorylation state and membrane localization. Molecular Pain. 2015;11:p. s12990-015-0074. doi: 10.1186/s12990-015-0074-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Xu G. Z., Tian J., Zhong Y. M., Yang X. L. Natriuretic peptide receptors are expressed in rat retinal ganglion cells. Brain Research Bulletin. 2010;82(3-4):188–192. doi: 10.1016/j.brainresbull.2010.03.004. [DOI] [PubMed] [Google Scholar]
- 73.Abdelalim E. M., Tooyama I. NPR-C is expressed in the cholinergic and dopaminergic amacrine cells in the rat retina. Peptides. 2010;31(1):180–183. doi: 10.1016/j.peptides.2009.10.014. [DOI] [PubMed] [Google Scholar]
- 74.Dumoulin A., Ter-Avetisyan G., Schmidt H., Rathjen F. G. Molecular analysis of sensory axon branching unraveled a cGMP-dependent signaling cascade. International Journal of Molecular Sciences. 2018;19(5, article 1266) doi: 10.3390/ijms19051266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Schmidt H., Werner M., Heppenstall P. A., et al. cGMP-mediated signaling via cGKIα is required for the guidance and connectivity of sensory axons. The Journal of Cell Biology. 2002;159(3):489–498. doi: 10.1083/jcb.200207058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Schmidt H., Dickey D. M., Dumoulin A., et al. Regulation of the natriuretic peptide receptor 2 (Npr 2) by phosphorylation of juxtamembrane serine and threonine residues is essential for bifurcation of sensory axons. The Journal of Neuroscience. 2018;38(45):9768–9780. doi: 10.1523/JNEUROSCI.0495-18.2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Schmidt H., Stonkute A., Jüttner R., Koesling D., Friebe A., Rathjen F. G. C-type natriuretic peptide (CNP) is a bifurcation factor for sensory neurons. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(39):16847–16852. doi: 10.1073/pnas.0906571106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Schmidt H., Stonkute A., Jüttner R.´., et al. The receptor guanylyl cyclase Npr 2 is essential for sensory axon bifurcation within the spinal cord. The Journal of Cell Biology. 2007;179(2):331–340. doi: 10.1083/jcb.200707176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Kishimoto I., Tokudome T., Horio T., et al. C-type natriuretic peptide is a Schwann cell-derived factor for development and function of sensory neurones. Journal of Neuroendocrinology. 2008;20(11):1213–1223. doi: 10.1111/j.1365-2826.2008.01778.x. [DOI] [PubMed] [Google Scholar]
- 80.Xia C., Nguyen M., Garrison A. K., et al. CNP/cGMP signaling regulates axon branching and growth by modulating microtubule polymerization. Developmental Neurobiology. 2013;73(9):673–687. doi: 10.1002/dneu.22078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Zhao Z., Ma L. Regulation of axonal development by natriuretic peptide hormones. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(42):18016–18021. doi: 10.1073/pnas.0906880106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Zhao Z., Wang Z., Gu Y., Feil R., Hofmann F., Ma L. Regulate axon branching by the cyclic GMP pathway via inhibition of glycogen synthase kinase 3 in dorsal root ganglion sensory neurons. The Journal of Neuroscience. 2009;29(5):1350–1360. doi: 10.1523/JNEUROSCI.3770-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Schmidt H., Peters S., Frank K., Wen L., Feil R., Rathjen F. G. Dorsal root ganglion axon bifurcation tolerates increased cyclic GMP levels: the role of phosphodiesterase 2A and scavenger receptor Npr 3. The European Journal of Neuroscience. 2016;44(12):2991–3000. doi: 10.1111/ejn.13434. [DOI] [PubMed] [Google Scholar]
- 84.Lu C. C., Appler J. M., Houseman E. A., Goodrich L. V. Developmental profiling of spiral ganglion neurons reveals insights into auditory circuit assembly. The Journal of Neuroscience. 2011;31(30):10903–10918. doi: 10.1523/JNEUROSCI.2358-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Ter-Avetisyan G., Rathjen F. G., Schmidt H. Bifurcation of axons from cranial sensory neurons is disabled in the absence of Npr2-induced cGMP signaling. The Journal of Neuroscience. 2014;34(3):737–747. doi: 10.1523/JNEUROSCI.4183-13.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Lu C. C., Cao X. J., Wright S., Ma L., Oertel D., Goodrich L. V. Mutation of Npr2 leads to blurred tonotopic organization of central auditory circuits in mice. PLoS Genetics. 2014;10(12, article e1004823) doi: 10.1371/journal.pgen.1004823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Wolter S., Möhrle D., Schmidt H., et al. GC-B deficient mice with axon bifurcation loss exhibit compromised auditory processing. Frontiers in Neural Circuits. 2018;12:p. 65. doi: 10.3389/fncir.2018.00065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Schmidt H., Fritzsch B. Npr2 null mutants show initial overshooting followed by reduction of spiral ganglion axon projections combined with near-normal cochleotopic projection. Cell and Tissue Research. 2019;378(1):15–32. doi: 10.1007/s00441-019-03050-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Evans A. J., Thompson B. C., Wallace G. G., et al. Promoting neurite outgrowth from spiral ganglion neuron explants using polypyrrole/BDNF-coated electrodes. Journal of Biomedical Materials Research. Part A. 2009;91(1):241–250. doi: 10.1002/jbm.a.32228. [DOI] [PubMed] [Google Scholar]
- 90.Stankovic K., Rio C., Xia A., et al. Survival of adult spiral ganglion neurons requires erbB receptor signaling in the inner ear. The Journal of Neuroscience. 2004;24(40):8651–8661. doi: 10.1523/JNEUROSCI.0733-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data used to support the findings of this study are available from the corresponding author upon request.