

Abstract
Background
Poultry feed consists mainly of conventional grains and protein supplements, however, using treated unconventional agro-industrial by-products as replacements of corn soybean-based diet can minimize production costs and improve productivity. Therefore, in this study, the effects of fermented or enzymatically treated dried brewer grains (DBG) on growth, expression of digestive enzymes and nutrient transporters genes and the profitability of the rations were evaluated. A total of 1600 one-day-old Ross 308 broiler chicks were randomly distributed in 2 × 4 factorial arrangement (eight treatments with ten replicates, 20 birds/replicate). Experimental diets included two controls; negative control (basal corn-soybean diet; NC) and positive control (basal corn-soybean diet with exogenous enzymes; PC), and six diets in which basal diet was replaced by three levels of fermented DBG (FDBG; 5, 10 or 15%), or enzyme-treated DBG (DBG 5, 10 or 15%+Enz), for 38 days.
Results
The results described that feeding FDBG (three levels) or DBG5%+Enz improved (P < 0.05) BW gain and feed efficiency of broilers. Also, feeding FDBG10% yielded the best improvement in weight gain (10%), compared to NC group. Increasing the inclusion levels of DBG either fermented or enzymatically treated up-regulated (p < 0.01) expression of digestive-genes in proventriculus (PGC and PGA5, range 1.4–1.8 fold), pancreas (AMY2A, PNLIP, CELA1, and CCK; range 1.2–2.3 fold) and duodenum (CAT1, CAT2, GLUT1, GLUT2, LAT1, Pep1; range 1.3-3 fold) when compared to NC group. Feeding treated DBG significantly increased (p < 0.05, range 4.5–13.6%) gizzard relative weight compared to NC and PC groups. An additional benefit was lower (p < 0.01) cholesterol content from 66.9 mg/100 mg (NC) to 62.8 mg/100 mg (FDBG5 or 10%) in thigh meat. Furthermore, the least cost feed/kg body gain was achieved in FDBG10% and DBG5%+Enz groups, with approx. 16% reduction compared to NC cost, leading to increasing the income gross margin by 47% and 40% in FDBG10% and DBG5%+Enz groups, respectively.
Conclusions
Substitution of corn-soybean based diet with 10% FDBG or 5% DBG+Enz resulted in better growth and higher economic efficiency of broilers chickens.
Keywords: Fermented dried brewer’s grain, Gene expression, Digestion gene, Nutrient transporter, Profitability
Background
In commercial poultry, the feeding costs of broilers contribute up to 70% of the total production costs. And, since, global feed prices are increasing, it is important to explore alternative or unconventional feed ingredients to achieve cost-effective poultry production [1]. However, the high fiber and low protein contents and presence of antinutritional factors (ANF) in unconventional feed represent a critical challenge for their use in poultry feed. One of the most prominent plant by-products arise mainly from oilseed mills and brewing industries [2]; called dried brewer’s grains (DBG). The latter could be an appropriate low-cost replacer for traditional feedstuffs (e.g., corn and soybean meal), as it contains a fair amount of feed residues following the brewing process [3–6]. It consists of around 20% crude protein, 6% ether extract, 15% crude fiber and 4% ash, in addition it has an adequate amount of essential AA (0.4% methionine, 0.9% lysine, 1.2% phenylalanine, 0.4% tryptophan, 1.1% threonine and 1.6% valine) [7].
However, the use of DBG in poultry feed has certain limitations such as elevated moisture content, non-starch polysaccharides (NSP) and tannins hindering feed utilization and inhibiting the digestive enzymes and nutrients absorption [8]. Adding different exogenous enzymes is one option to mitigate ANF and improve DBG utilization, in addition to supporting bird’s endogenous enzymes and nutrient absorption thus improving digestibility and availability of nutrients [9, 10]. However, supplementation of exogenous enzymes in poultry feed still have limitations which reduce their beneficial effects, such as the wide range of pH along bird’s GIT (while their optimum pH is between 4 and 6), the shorter time for digesta retention in proximal GIT (their main site of action) and the possibility of hydrolysis by endogenous proteolytic enzymes [11].
The other possible strategy for improving DBG quality in broiler diet is through the fermentation approach. Fermentation is a dynamic process containing microorganisms, substrates and environmental conditions to transform complex substrates into simpler ones [12]. As the fermentation intensity depends on the number of microorganisms, thus adding fungi or bacterial strains can be a favorable aid during fermentation. It is known that fermentation improves microbial and nutritional quality of feedstuffs by increasing protein and lipids and lowering fiber content [13]. As well as, improving the availability of vitamins and AAs [14, 15] and reducing several ANF and toxic compounds [16]. Fermentation has also been proven to enhance nutrient digestibility of organic matter, fiber and calcium [17], and the palatability of feedstuff [18]. In addition, Tan et al. [19] confirmed an enhanced nutritional content of fermented DBG through biological fermentation by adding certain fungi and bacteria.
The nature of broiler’s diet can modulate the expression of important genes associated with nutrient transport and digestive enzymes [20, 21]. In fact, improving nutrient transport, through the up-regulation of the transporter encoding genes, lead to accelerated nutrient entry into the intestinal epithelium and then to the circulation [22]. It is identified that transporters of glucose (GLUT1 and GLUT2), cationic amino acid (CAT1 and CAT2) and peptide (PepT1 and PepT2) in the intestinal epithelium are associated with the capacity of nutrient absorption [21, 23–25].
The effects of different DBG treatments (fermentation or added enzymes) on expression of genes controlling digestive enzymes and nutrients transporters in broilers remains not fully understood. In addition, the economic efficiency of replacing costly conventional feed ingredients should promote the use of low-priced unconventional fermented feedstuffs as DBG in broiler diets. Therefore, the present study investigated the impact of feeding different levels of DBG either fermented or enzyme-treated on the gene expression of selected digestive enzymes and nutrient transporters, growth performance and economic efficiency of broiler chickens.
Results
Growth indices
The highest body weight and weight gain (P < 0.05) were observed in FDBG10% group followed by FDBG5% and DBG5%+Enz groups, Table 1. The lowest total feed intake (P < 0.05) was observed in group fed FDBG15% followed by groups fed on DBG10%+Enz and negative control. The feed conversion ratio (FCR) was improved (P < 0.05) in groups fed FDBG5%, FDBG10% and DBG5%+Enz when compared with NC and PC groups.
Table 1.
Effects of different levels of fermented or enzymatically treated dried brewers’ grains on growth performance and carcass traits of broilers1
| Treatmnets2 | NC | PC | FDBG15% | FDBG10% | FDBG15% | DBG5%+Enz | DBG10%+Enz | DBG15%+Enz | SEM | P-value |
|---|---|---|---|---|---|---|---|---|---|---|
| Allover growth performance parameters3 | ||||||||||
| BW, g/bird | 2414e | 2559c | 2587b | 2690a | 2432d | 2593b | 2332f | 2188g | 9.26 | < 0.001 |
| BWG, g/bird | 2369e | 2514c | 2542b | 2645a | 2387d | 2549b | 2287f | 2143g | 9.20 | < 0.001 |
| FI, g/bird | 3939d | 4109a | 4052b | 4001c | 3883e | 3993c | 3915de | 4067b | 59.82 | < 0.001 |
| FCR | 1.66c | 1.63cd | 1.57e | 1.51f | 1.63d | 1.57e | 1.71b | 1.90a | < 0.006 | < 0.001 |
| Carcass traits4 | ||||||||||
| Dressed weight | 1458c | 1576b | 1593b | 1641a | 1547b | 1603a | 1422c | 1321c | 0.75 | < 0.001 |
| Liver, % | 2.52 | 2.36 | 2.5 | 2.32 | 2.48 | 2.63 | 2.26 | 2.61 | 0.02 | 0.11 |
| Gizzard, % | 2.20c | 2.26bc | 2.31b | 2.49a | 2.50a | 2.36b | 2.40ab | 2.43a | 0.10 | 0.03 |
| Spleen, % | 0.17 | 0.14 | 0.15 | 0.14 | 0.15 | 0.14 | 0.13 | 0.14 | 0.01 | 0.27 |
| Thymus, % | 0.34 | 0.31 | 0.38 | 0.34 | 0.34 | 0.37 | 0.37 | 0.33 | 0.05 | 0.08 |
| Bursa, % | 0.06 | 0.67 | 0.07 | 0.08 | 0.07 | 0.07 | 0.08 | 0.09 | 0.01 | 0.5 |
| Abdominal fat, % | 1.69 | 1.72 | 1.70 | 1.74 | 1.74 | 1.72 | 1.78 | 1.76 | 0.30 | <0.32 |
a-gMeans within the same row carrying different superscripts are significantly different at P < 0.05. 1Data represent the mean value of ten replicate pens of 20 birds. 2Treatments include: NC (negative control), PC (positive control), FDBG5% (5% fermented dried brewers grains), FDBG10% (10% fermented dried brewer grain), FDBG15% (15% fermented dried brewers grains), DBG5%+Enz (5%dried brewers grains mixed with enzymes), DBG10%+Enz (10% dried brewers grains mixed with enzymes), DBG15%+Enz (15% dried brewers grains mixed with enzymes).3BW Body weight, BWG Body weight gain, FI Feed intake, FCR Feed conversion ratio.4Carcass traits (n=5/treatment)
Carcass traits
The highest dressing percentage (P < 0.05) was observed in groups fed FDBG10% and DBG5%+Enz, Table 1. Feeding on DBG either fermented or enzymatically treated significantly increased (P < 0.05) gizzard relative weight. Feeding of broiler chicks on treated DBG had no effect on the relative weights of liver, spleen, thymus, bursa and abdominal fat.
Nutrient transporter and digestive enzyme genes expression
Proventriculus pepsinogen (PGC and PGA5) genes expression (Fig. 1)
Fig. 1.
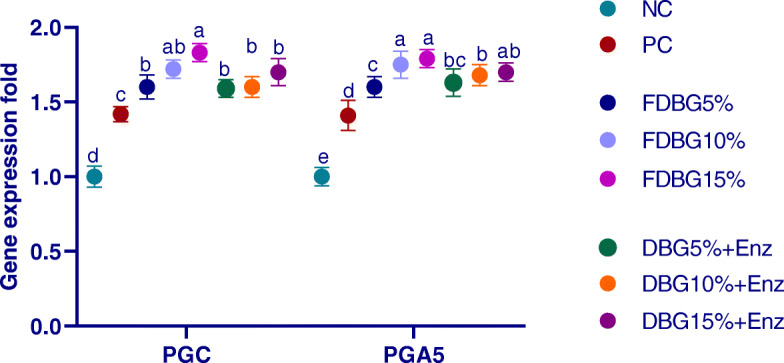
Effects of different level of fermented or enzymatically treated dried brewers’ grains on expression of proventriculus gene (PGC and PGA5). a-c Means within the same column carrying different superscripts are significantly different at P < 0.05
The upregulation of proventriculus pepsinogen (PGC and PGA5) genes was more prominent (P < 0.01) in groups with higher levels of DBG either fermented or enzymatically treated when compared with the NC and PC groups.
Pancreatic genes expression, AMY2A and PNLIP (Fig. 2a), CELA1 and CCK (Fig. 2b)
Fig. 2.

Effects of different level of fermented or enzymatically treated dried brewers’ grains on expression of pancreatic genes (AMY2A, PNLIP; (a), CCK and CELA1; (b). a-cMeans within the same column carrying different superscripts are significantly different at P < 0.05
The mRNA expression of pancreatic AMY2A, PNLIP and CELA1 was upregulated (p < 0.01) in response to all inclusion levels of treated DBG. The higher expression of AMY2A gene was more prominent in groups fed higher levels of FDBG. Regarding CCK expression level, groups fed FDBG10% and 15% had the highest expression level while FDBG5% and enzymatically treated DBG groups had the intermediate expression levels and control groups had the lowest expression level.
Duodenal genes expression, GLUT1 and GLUT2, (Fig. 3a); PepT1 and PepT2 (Fig. 3b) and CAT1, CAT2 (Fig. 4a) and LAT1 (Fig. 4b)
Fig. 3.

Effects of different level of fermented or enzymatically treated dried brewers’ grains on expression of duodenal genes, GLUT1, GLUT2; (a) and PEPT1 and PEPT2; (b). a-bMeans within the same column carrying different superscripts are significantly different at P < 0.05
Fig. 4.

Effects of different level of fermented or enzymatically treated dried brewers’ grains on expression of duodenal genes (CAT1, CAT2; (a) and LAT1; (b)). a-cMeans within the same column carrying different superscripts are significantly different at P < 0.05
The mRNA expression of GLUT1, GLUT2, were up regulated in response to the dietary inclusion of treated DBG. The higher levels of Broilers fed on Fermented or enzymatically treated DBG or PC significantly up-regulated (P < 0.01) the CAT1, CAT2, while PepT2 expression not affected by feeding on treated DBG (P > 0.05). Increasing inclusion of fermented or enzymatically treated DBG significantly up-regulated (P < 0.01) LAT1 gene expression. Moreover, the effect of fermentation on upregulation of previous genes was more prominent than enzymes treatment. Increasing the inclusion levels from DBG either fermented or enzymatically treated up-regulated (P < 0.05) the expression of GLUT1, GLUT2 genes than in control positive group.
Chemical composition of meat
The moisture, crude protein and cholesterol contents of breast meat were not different among all treatments, Table 2. However, the groups fed on FDBG5% and FDBG10% had the lowest cholesterol content (P < 0.05) in thigh meat .
Table 2.
Effects of different levels of fermented or enzymatically treated dried brewers’ grains on chemical composition of meat
| Treatments1 | Breast moisture % | Breast protein % | Breast cholesterol (mg/100mg) | Thigh cholesterol (mg/100mg) |
|---|---|---|---|---|
| NC | 70.60 | 21.97 | 62.36 | 66.91ab |
| PC | 70.81 | 22.23 | 62.25 | 66.85ab |
| FDBG5% | 70.93 | 22.80 | 61.46 | 62.84d |
| FDBG10% | 70.64 | 22.12 | 61.54 | 62.82d |
| FDBG15% | 70.56 | 23.00 | 61.34 | 64.33c |
| DBG5%+Enz | 71.00 | 22.94 | 61.83 | 65.67bc |
| DBG10%+Enz | 70.63 | 22.49 | 61.97 | 68.00a |
| DBG15%+Enz | 70.27 | 22.10 | 61.73 | 68.70a |
| SEM | 0.06 | 0.07 | 0.30 | 0.04 |
| P-value | 0.09 | <0.08 | <0.07 | <0.001 |
a-cMeans within the same column carrying different superscripts are significantly different at P < 0.05, n=5/treatment). 1Treatments include: NC (negative control), PC (positive control), FDBG5% (5% fermented dried brewers grains), FDBG10% (10% fermented dried brewer grain), FDBG15% (15% fermented dried brewers grains), DBG5%+Enz (5%dried brewers grains mixed with enzymes), DBG10%+ Enz (10% dried brewers grains mixed with enzymes), DBG15%+Enz (15% dried brewers grains mixed with enzymes)
Serum biochemical parameters
The concentrations of total protein, albumin, globulin, triglyceride, ALT and AST were not affected (P > 0 0.05) by any of the experimental diets, Table 3. However, the serum concentrations of total cholesterol and LDL were significantly lowered (P < 0.05) in broilers fed FDBG when compared with other groups.
Table 3.
Effects of different levels of fermented or enzymatically treated dried brewers’ grains on some serum biochemical parameters
| Treatments1 | Total protein (g/dl) | Albumin (g/dl) | Globulin (g/dl) | TC (mg/dl) | TAG (mg/dl) | HDL (mg/dl) | LDL (mg/dl) | AST (U/L) | ALT (U/L) |
|---|---|---|---|---|---|---|---|---|---|
| NC | 4.55 | 3.14 | 1.41 | 174.50a | 72.13 | 77.41 | 82.75a | 1.63 | 51.38 |
| PC | 4.87 | 3.28 | 1.59 | 165.16a | 77.22 | 76.03 | 73.68a | 1.51 | 51.30 |
| FDBG5% | 5.19 | 3.75 | 1.440 | 154.52b | 75.67 | 76.72 | 62.66b | 1.51 | 51.83 |
| FDBG10% | 5.13 | 3.07 | 2.06 | 156.94b | 72.27 | 78.36 | 64.13b | 1.64 | 51.67 |
| FDBG15% | 6.02 | 3.03 | 2.99 | 157.33b | 76.38 | 78.30 | 63.75b | 1.60 | 53.18 |
| DBG5%+Enz | 5.09 | 3.32 | 1.77 | 162.80ab | 79.14 | 73.21 | 73.75ab | 1.61 | 52.61 |
| DBG10%+Enz | 4.92 | 3.583 | 1.54 | 172.95a | 75.81 | 69.97 | 87.83a | 1.63 | 52.86 |
| DBG15%+Enz | 4.88 | 3.61 | 1.31 | 171.10a | 74.84 | 71.07 | 85.06a | 1.66 | 50.76 |
| SEM | 0.03 | 0.17 | 0.07 | 4.542 | 15.01 | 3.13 | 11.43 | 0.60 | 0.001 |
| P-value | 0.06 | 0.91 | 0.13 | <0.001 | 0.972 | 0.119 | <0.003 | 0.71 | 0.06 |
a-bMeans within the same column carrying different superscripts are significantly different at P < 0.05.1Treatments include: NC (negative control), PC (positive control), FDBG5% (5% fermented dried brewers grains), FDBG10% (10% fermented dried brewer grain), FDBG15% (15% fermented dried brewers grains), DBG5%+Enz (5%dried brewers grains mixed with enzymes), DBG10%+ Enz (10% dried brewers grains mixed with enzymes), DBG15%+Enz (15% dried brewers grains mixed with enzymes). TC (Total Cholesterol), TAG (Triglyceride), HDL (high density conjugated protein), LDL (low density protein), AST (Aspartate aminotransferase), ALT (alanine aminotransferase)
Economic indices
Total feed cost and total cost decreased significantly (P < 0.05) with increasing DBG levels, either fermented or mixed with enzymes, Table 4. The highest values of total expenses were recorded in NC (2.71$) and PC (2.80$) groups. While the highest gross margin (P < 0.05) was evident in broilers fed FDBG10% (1.58$) followed by DBG5%+Enz (1.50$). The lowest cost/kg BW gain of broilers and highest cost benefit ratio were achieved in groups fed FDBG10% and DBG5%+Enz.
Table 4.
Effects of different level of fermented or enzymatically treated dried brewers’ grains on the economic indices
| Treatments1 | Feed cost2 | Total expenses | Total revenue | Gross margin | benefit cost ratio | Cost/ kg BW gain |
|---|---|---|---|---|---|---|
| NC | 1.61b | 2.71b | 3.79c | 1.07g | 0.66e | 0.69a |
| PC | 1.70a | 2.80a | 4.01b | 1.21e | 0.70d | 0.68a |
| FDBG5% | 1.57c | 2.67c | 4.05b | 1.38c | 0.87b | 0.61bc |
| FDBG10% | 1.52d | 2.62d | 4.21a | 1.58a | 1.04a | 0.58d |
| FDBG15% | 1.44f | 2.53f | 3.80c | 1.28d | 0.89b | 0.60c |
| DBG5%+Enz | 1.46e | 2.56e | 4.06b | 1.50b | 1.02a | 0.56d |
| DBG10%+Enz | 1.42g | 2.56e | 3.66d | 1.14f | 0.80c | 0.62b |
| DBG15%+Enz | 1.46e | 2.52g | 3.44e | 0.88h | 0.60f | 0.68a |
| SEM | 2.59 | 2.59 | 0.003 | 0.006 | 0.006 | 1.15 |
| P-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
a-hMeans within the same column carrying different superscripts are significantly different at P < 0.05. 1Treatments include: NC (negative control), PC (positive control), FDBG5% (5% fermented dried brewers grains), FDBG10% (10% fermented dried brewer grain), FDBG15% (15% fermented dried brewers grains), DBG5%+Enz (5%dried brewers grains mixed with enzymes), DBG10%+ Enz (10% dried brewers grains mixed with enzymes), DBG15%+Enz (15% dried brewers grains mixed with enzymes).2Cost/kg diet$ = NC (0.411) , PC (0.415), FDBG5% (0.388), FDBG10% (0.381), FDBG15% (0.370), DBG5%+Enz (0.367), DBG10%+Enz (0.362), DBG15% +Enz (0.359). price/kg meat = $ 1.52. fixed costs = $ 1.1
Discussion
Microbial fermentation is a recent low-cost method to enhance the nutritional value of unconventional feed ingredients for broiler chickens. Also, there is a growing interest in the introduction of fermented feed to broiler diets due to its positive impacts on the gut health and growth performance [26, 27]. Thus, subjecting unconventional feed as DBG to microbial fermentation may be a sound alternate to improve DBG nutritive value. In the current study, microbial fermentation improved nutritive value of DBG which allowed its inclusion up to 10% in the formulated diet and led to better broiler’s performance. Similar studies have reported that microbial fermentation is an effective method to degrade ANFs and provide probiotics in feed, thus enhancing the nutritional quality and improving animal growth [28–30]. Improved nutritional quality due to fermentation process is through increasing crude protein and reducing crude fiber contents [29].
Therefore, increased protein content in fermented DBG may arise from (1) synthesis of microbial proteins and enzymes during fermentation [31], (2) increasing microbial population which is composed mainly from protein [32], and (3) enhancing the activities of proteolytic microorganisms which causes an increase in free amino acids and peptides in fermented products [33]. Also, fiber reduction after fermentation is due to increased microbial population which produce fiber degrading enzymes [34]. Also, addition of B. subtilis during fermentation enhanced growth and proliferation of lactic acid bacteria (LAB) and in turn reduced pH thus inhibited the growth of pathogenic bacteria [30]. Moreover, during anerobic fermentation process, the addition of S. cerevisiae was preferred due to its greater pH lowering capacity [35] and enzyme producing ability as β -glucanase, phytase and invertase [36–38].
The improved BW and FCR of broiler chickens during the whole grow-out period (d 1–38) in groups fed FDBG (up to 10%), is a reflection to the improved nutritional properties of FDBG compared to enzymatically treated DBG. Similarly, recent reports showed that the fermentation help in the production of functional feeds that can improve the microecology and health of broiler’s gut and their productive performance [39]. In addition, fermentation was associated with a high concentration of organic acids which increased the number of LAB leading to lowered gastric pH and pathogenic microbial activity [17, 40, 41]. All these previous characteristics of fermented feed should prevent pathogenic feed contamination prior to feeding [42], protect chicken gastrointestinal health [43, 44] and improve their growth [45].
Adding selected microbial strains as B.subtilis, L. rhamnosus and S. cerevisiae during fermentation of DBG not only improved DBG nutritional value but also enhanced birds growth rate. In accordance with our results, microbial fermentation with B. Subtilis can increase feed palatability [46], secrete digestive enzymes (proteases, lipases and amylases), stimulate good digestion and absorption of nutrients, and, produce active substances (bacitracin, polymyxin, nystatin, gramicidin) which inhibit endogenous pathogens [47]. The last authors also suggested that microbial phytase and protease enzymes may be accountable for decreasing the contents of phytic acid and allergic proteins, respectively. In addition, S. cerevisiae effects were attributed to maintaining beneficial microbial population and modifying metabolism by enhancing the activity of digestive enzymes [48].
Treating DBG with enzymes (especially the DBG5%+ENZ group) also had positive effects on improving broiler’s FCR agreeing with previous reports describing that inclusion of exogenous enzymes in conventional or unconventional broiler’s diet led to lower digesta viscosity, countered anti-nutritional factors and assisted in the development of important microbiota [49–51]. Also, the incorporation of multienzyme (xylanase, amylase, and protease) in broiler’s diet improved fibers utilization [52]. While, supplementing β-glucanase to barley-based diet reduced ileal viscosity and altered the concentration of short chain fatty acids in the crop and ceca thus increased broilers body weight gain [53, 54].
Increased gizzard weight observed in all DBG groups may be due to the presence of NSP in DBG, which was shown to beneficially affect nutrient utilization by (1) increased proventriculus and gizzard activity, (2) triggered hydrochloric acid and digestive enzyme secretion and (3) augmented hind gut bacterial fermentation [55]. Moreover, a well-developed gizzard musculature can produce strong reverse peristalsis contractions and increase the digesta refluxes in digestive tract thus re‐exposing the digesta to hydrochloric acid and pepsin and help their mixing [56–58].
The current study demonstrated that genes essential for digestion and nutrient transport were upregulated in the proventriculus, duodenum and pancreas of broilers in line with increased level of structural fiber components (by increasing DBG level; either fermented or enzymatically treated) in broiler’s diet. This effect was more prominent with FDBG than in enzymatically treated DBG. In accordance with the upregulated pepsinogen expression after feeding treated DBG in our study, it was shown that feeding on high fiber diet upregulated the expression of pepsinogen A and C in birds [55]. In addition, the high fiber content in DBG should stimulate frequent and powerful contractions of the gizzard and relocating the digesta back into the proventriculus leading to more stimulation of proventricular function, as previously reported in high fiber diets [55]. Additionally, the upregulation of pepsinogen A and C expression with higher inclusion levels of FDBG may be related to increased LAB, more lactic acid and lower pH, which accelerated the proliferation of gut epithelial cells [59, 60] and stimulated gastric chief cells to release pepsinogen [61]. While, feeding enzymatically treated DBG could enhance pepsinogen via the supplied exogenous enzymes which upregulated the expression of digestive enzymes [62]. So, in our study increasing gizzard contraction and pepsin production in proventriculus can result in a more active foregut and better nutrient digestion.
Also, in this study, the nature of diet affected the magnitude of pancreatic secretion, as high dietary carbohydrates or fat stimulated pancreatic secretions and increased amylase and lipase concentration in serum [63–65]. In the current study, the increased expression of pancreatic enzymes-related genes by using DBG was in accordance with previous reports which stated that insoluble fiber can increase pancreatic enzymes such as chymotrypsin [66, 67]. Similarly, oat hulls with high fiber content stimulated the secretion of pancreatic amylase and thus increased amylase activity in the jejunum [68]. Moreover, the activities of pancreatic α-amylase and lipase in broiler chickens were significantly increased by feeding wheat and barley-based diet compared to corn-based diet with enzymes supplementation [69]. Also, elevated CCK expression with increased levels of treated DBG was related to improved gut motility, gastro-duodenal reflux and secretion of pancreatic enzymes which in turn increased CCK release and improved the digestibility [70, 71]. Likewise, fermented feeds elevated pancreatic AMY2A and CCK expression which in turn increased the secretion of pancreatic amylase and cholecystokinin in broilers [44, 72]. Additionally, the upregulated expression of pancreatic lipase in all treated DBG groups led to higher lipase secretion and resulted in better fat absorption [73]. Furthermore, the probiotic properties of FDBG led to improving intestinal structure, absorptive surface area and expression of brush border enzymes, thus achieving boosted digestion and absorption [74, 75]. Equally, B. subtilis based-diet up-regulated digestive genes such as pancreatic lipase, carboxypeptidase and chymotrypsin-like elastase in the gut [76].
Nutrient absorption, in the small intestine, is mediated by transporter proteins expressed in enterocytes. Up-regulation of these transporters improved nutrient transport capability and accelerated nutrients influx into the intestinal epithelial cells and then to all body (Ruhnke et al., 2015). In our study, GLUT1, GLUT2, CAT1, CAT2 and PEPT1 were all upregulated after feeding on DBG (either fermented or enzymatically treated). In agreement with our results, the addition of exogenous dietary enzyme for broilers significantly upregulated the expression of intestinal PEPT1 and GLUT2 thus facilitated micronutrients absorption [62]. Similarly, inclusion of fiber-rich sugarcane bagasse upregulated CAT1, LAT2 and PepT2, which facilitated the bidirectional transfer of cationic amino acids, Na+−dependent neutral/cationic amino acid exchange, and di- and tri-peptides transport in intestine, respectively [55]. The increase in carbohydrates consumption led to higher expression levels of glucose transmitters which increased glucose absorption [77]. Also, the expression of GLUT2 and PEPT1 genes was upregulated after supplementation of xylanase which may indicate an improved absorption in birds [78, 79]. All this confirms that increased entry of nutrients from the intestinal lumen into the enterocyte resulted in improved tissue protein synthesis and enhanced feed utilization as previously highlighted [80].
No considerable changes were noticed in the activities of liver metabolic enzymes (ALT and AST) among different experimental groups indicating absence of any metabolic side effects. Additionally, there was no substantial impact of treated DBG on any of the serum constituents (Albumin, globulin, cholesterol, triglyceride and high-density lipoprotein), except for total cholesterol and low-density lipoprotein LDL which were decreased in FDBG groups compared to control or enzymatically treated groups. This FDBG-caused reduction in blood cholesterol levels could be through, (1) inhibiting enzymes involved in biosynthesis of cholesterol (as 3-hydroxy-3-methylglutaryl CoA reductase) (2) increasing bile acid production, or (3) decreasing cholesterol synthesis and absorption in the gastrointestinal tract [81]. In addition, fermentation-related LAB decreased Niemann-Pick C1-like 1 expression levels in enterocytes leading to less cholesterol absorption [82].
In the present study, reduced cholesterol levels in blood was also reflected in lower cholesterol contents in thigh meat, following FDBG, which agree with previous findings [83]. In addition, it may be attributed to increased total fecal excretion of cholesterol via the bile [84]. Also, similar results were found when cholesterol concentration in breast and thigh meat decreased in turkey fed on fermented feed by-products [85]. Likewise, using of 10% fermented feed in broiler diet significantly decreased fat content in breast muscle [86].
It is accepted that using of non-conventional dietary sources can reduce the feeding cost of broilers which is a main constraint for profit in broiler industry. As described in our study the feed cost and total costs were higher in control groups, which is clearly due to the costs of conventional ingredients. However, feed cost and cost/kg diet were decreased with increasing the level of DBG in the rations, this is mainly due to its lower purchase price compared to conventional ingredients in the control diet. This was reflected in better net profit and total return values in 10% FDBG and 5% DBG + enzymes groups, which showed a decrease in feed cost/kg together with better feed utilization and body weight gain compared to the remaining groups. Similarly, appreciable financial gain was reported after increasing the inclusion level of exogenous enzymes fortified dried brewer grain in the broilers diet [87]. Also, combining exogenous enzymes with non-conventional ingredients is a valid practice to reduce the cost of feeding and to allow better utilization of the non-conventional feed ingredients that are typically rich in fibers and are difficult to be utilized by poultry endogenous enzymes. This was achieved in this study in the group fed on enzymatically treated DBG at 5% inclusion level. Additionally, previous findings indicated that the total cost of production and feed cost/kg decreased with increasing BDGs level [88]. Moreover, the feed cost effectiveness in FDBG 10%, agreed with previous reports [89, 90] which reported that probiotic produced by solid substrate fermentation (SSF) is economically and environment-friendly. Furthermore, we confirm previous hypothesis that using fermented feed containing probiotics would be of economic value via improving the broiler feed conversion ratio [91]. Therefore, from an economic point of view, it is clear in our results that fermentation of DBG achieved better economic production than enzymatically treated DBG.
Conclusions
Introduction of new nutritional strategies (such as microbial fermentation) for broilers, lead to better utilization of unconventional feed as DBG and improving the revenue. Fermentation of DBG positively affected the expression of digestive and nutrient transporters genes which regulate nutrients and energy availability necessary for optimum bird-growth, more than enzymatic treated DBG. These outcomes should encourage the poultry feed industry to apply cost effective protocols by incorporating fermented unconventional products in broiler diets.
Methods
Microbial fermentation of Dried Brewer Grains
The dried brewer’s grains (DBG) were obtained from beer factory and dried for 12 h at 40 °C then finely ground, then were fermented by different microbes. Bacillus subtilis (NCIMB 15,204) was activated with nutrient broth (5 ml) at 37 °C for 24 h. Lactobacillus rhamnosus (ATCC 7469) cultures were anaerobically incubated in De Man, Rogosa and Sharpe (MRS) (Oxoid Ltd., Basingstoke, Hampshire, UK) at 37 °C for 24 h for activation. Saccharomyces cerevisae (CGMCC No. 2.1793) was activated by incubation at 37 °C for 48 h in glycerol and MRS broth. A mixed liquid culture of approximately 1 × 106 CFU/ml of each S. cerevisiae, L. rhamnosus and B. subtilis, were utilized by the same ratio and mixed with DBG. The mixture was packaged and sealed in polyethylene bags with a one-way valve to permit carbon dioxide release during fermentation. Through the initial phase of fermentation, S. cerevisae consumed the oxygen in the bag and generated significant amount of carbon dioxide. This process produced an anaerobic and acidic conditions which enabled both L. rhamnosus and B. subtilis to grow and germinate. The fermentation was held for 25 days. Samples of dried brewers’ grains (DBG) were collected for chemical analysis according to AOAC [92], and shown in Table 5.
Table 5.
Chemical analysis (% on DM basis) of unfermented (UFDBG) and fermented (FDBG) dried brewers’ grains
| Constituent (%) | 1UFDBG | 2FDBG |
|---|---|---|
| Crude protein | 28.20±0.15b | 29.60±0.10a |
| Ether extract | 6.20±0.20b | 6.90±0.16a |
| Crude fiber | 12.60±0.19a | 10.80±0.17b |
| 3NDF | 54.66±0.08a | 51.36±0.07b |
| 4ADF | 20.36±0.09a | 18.46±0.20b |
| Lignin | 5.26±0.10a | 4.00±0.20b |
a-bMeans within the same row carrying different superscripts are significantly different at P < 0.05. 1UDBG (un-fermented dried brewer grains), 2FDBG (fermented dried brewer grains).3NDF (Neutral detergent fiber), 4ADF Acid detergent fiber. Values are expressed as means± standard error
Birds, experimental design, and management
One-day-old male Ross 308 broiler chicks (n = 1600) were obtained from a commercial chick producer (Dakahlia Poultry, Mansora, Egypt) and weighted and randomly assigned to one of the eight experimental treatments (ten replicates/treatment, and 20 birds/replicate) in 2 × 4 factorial arrangement. Treatments consist of negative control (NC) birds received corn-soybean based diet; positive control (PC) birds received corn-soybean based diet with exogenous enzymes and in the remaining six diets, the basal diet was replaced by three levels of fermented DBG (FDBG; 5, 10 or 15%), or enzyme-treated DBG (DBG 5, 10 or 15%+Enz). A commercial multi-exogenous enzyme was added was added to PC and enzyme treated DBG diets at a concentration of 1 gm/kg diet.
The birds were raised in a naturally ventilated open house with sawdust as litter. The feed and water were available ad-libitum and the broilers were raised in floor pens over a period of 38 days. The temperature, relative humidity and lighting were adjusted following Ross 308 management guidelines [93]. All diets were offered in mash form and the basal diets were formulated (starter, grower-finisher) according to nutrition specification of Ross broiler handbook [93], Tables 6 and 7. The chemical analyses (moisture, crude protein, ether extract and crude fiber) of all feed ingredients were conducted according to AOAC [92]. A commercial multi-exogenous enzymes (KEMZYME® Plus Europa NV, Belgium), containing Xylanase, B-glucanase, cellulase, alpha-amylase and protease. After the study, all remaining birds were released.
Table 6.
The ingredients and nutrient level of diets during starter stage
| Diet composition (%) | Experimental dietsa | |||||||
|---|---|---|---|---|---|---|---|---|
| NC | PCb | FDBG 5% |
FDBG 10% |
FDBG 15% |
DBG5% +Enzb |
DBG10% +Enzb |
DBG15% +Enzb |
|
| Yellow corn | 57.50 | 57.50 | 55.25 | 53.00 | 50.50 | 54.55 | 51.30 | 48.45 |
| Soybean meal | 31.80 | 31.80 | 29.00 | 26.20 | 23.70 | 29.40 | 27.20 | 24.70 |
| Corn gluten | 3.30 | 3.30 | 3.30 | 3.30 | 3.30 | 3.30 | 3.30 | 3.30 |
| FDBGc | 0 | 0 | 5.00 | 10.00 | 15.00 | 0 | 0 | 0 |
| DBGd | 0 | 0 | 0 | 0 | 0 | 5.00 | 10.00 | 15.00 |
| Soybean oil | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 | 3.30 | 3.70 | 4.00 |
| Calcium carbonate | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Calcium diphasic phosphate | 1.50 | 1.50 | 1.50 | 1.50 | 1.50 | 1.50 | 1.50 | 1.50 |
| Common salt | 0.30 | 0.30 | 0.30 | 0.30 | 0.30 | 0.30 | 0.30 | 0.30 |
| Premixe | 0.90 | 0.90 | 0.90 | 0.90 | 0.90 | 0.90 | 0.90 | 0.90 |
| L-Lysine | 0.20 | 0.40 | 0.25 | 0.30 | 0.30 | 0.25 | 0.30 | 0.35 |
| DL-Methionine | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 |
| Choline chloride | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 |
| Anti-mycotoxin | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 |
| Calculated composition | ||||||||
| ME (Kcal/Kg) | 3105 | 3105 | 3105 | 3105 | 3101 | 3102 | 3103 | 3101 |
| CP (%) | 22.50 | 22.50 | 22.50 | 22.50 | 22.50 | 22.50 | 22.50 | 22.50 |
| EE % | 5.44 | 5.44 | 5.67 | 5.90 | 6.10 | 5.89 | 6.44 | 6.90 |
| CF (%) | 2.56 | 2.56 | 2.94 | 3.30 | 3.32 | 3.12 | 3.68 | 4.23 |
| Calcium (%) | 1.11 | 1.11 | 1.10 | 1.10 | 1.11 | 1.11 | 1.10 | 1.10 |
| Available phosphorous (%) | 0.51 | 0.51 | 0.49 | 0.47 | 0.46 | 0.49 | 0.48 | 0.46 |
| Lysine (%) | 1.27 | 1.27 | 1.27 | 1.28 | 1.26 | 1.27 | 1.28 | 1.28 |
| Methionine (%) | 0.57 | 0.57 | 0.58 | 0.58 | 0.59 | 0.58 | 0.59 | 0.60 |
aNC (negative control), PC (positive control), FDBG5% (5% fermented dried brewers grains), FDBG10% (10% fermented dried brewer grain), FDBG15% (15% fermented dried brewers grains), DBG5%+Enz (5%dried brewers grains mixed with enzymes), DBG10%+Enz (10% dried brewers grains mixed with enzymes), DBG15%+Enz (15% dried brewers grains mixed with enzymes). bCommercial multi-exogenous enzyme was added was added to PC and enzyme treated DBG diets at a concentration of 1 gm/kg diet. cFDBG (fermented dried brewer grains). dDBG (dried brewer grains). eVitamin premix supplied per kilogram of diet: vitamin A, 10,000 IU; vitamin D3, 2000 IU; vitamin E, 6500 IU; vitamin K3, 1 mg; vitamin B1, 2560 mg; vitamin B2, 5000 mg; vitamin B6, 1500 mg; B5, 8 mg; niacin, 20000 mg; biotin, 0.25 mg; folic acid, 1000 mg; vitamin B12, 60 mg; Cu, 8 mg; Fe, 80 mg; Mn, 60 mg; Zn, 40 mg; Se, 0.15 mg
Table 7.
The ingredients and nutrient level of diets during grower-finisher stage
| Diet composition (%) | Experimental dietsa | |||||||
|---|---|---|---|---|---|---|---|---|
| NC | PCb | FDBG 5% |
FDBG 10% |
FDBG 15% |
DBG5% +Enzb |
DBG10% +Enzb |
DBG15% +Enzb |
|
| Yellow corn | 62.25 | 62.25 | 60.20 | 57.90 | 55.60 | 59.30 | 56.45 | 53.40 |
| Soybean meal | 26.00 | 26.00 | 23.00 | 20.25 | 17.50 | 23.50 | 21.00 | 18.70 |
| Corn gluten | 3.90 | 3.90 | 3.90 | 3.90 | 3.90 | 3.90 | 3.90 | 3.90 |
| FDBG c | 0 | 0 | 5.00 | 10.00 | 15.00 | 0 | 0 | 0 |
| DBG d | 0 | 0 | 0 | 0 | 0 | 5.00 | 10.00 | 15.00 |
| Soybean oil | 3.60 | 3.60 | 3.60 | 3.60 | 3.60 | 4.00 | 4.30 | 4.60 |
| Calcium carbonate | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Calcium diphasic phosphate | 1.30 | 1.30 | 1.30 | 1.30 | 1.30 | 1.30 | 1.30 | 1.30 |
| Common salt | 0.30 | 0.30 | 0.30 | 0.30 | 0.30 | 0.30 | 0.30 | 0.30 |
| Premix e | 0.90 | 0.90 | 0.9 | 0.90 | 0.90 | 0.90 | 0.90 | 0.90 |
| L-Lysine | 0.25 | 0.25 | 0.3 | 0.35 | 0.4 | 0.3 | 0.35 | 0.40 |
| DL-Methionine | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 |
| Choline chloride | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 |
| Anti-mycotoxin | 0.10 | 0.10 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| Calculated composition | ||||||||
| ME (Kcal/Kg) | 3200 | 3200 | 3201 | 3201 | 3200 | 3203 | 3202 | 3200 |
| CP (%) | 20.5 | 20.5 | 20.5 | 20.50 | 20.50 | 20.50 | 20.50 | 20.50 |
| EE % | 6.15 | 6.15 | 6.38 | 6.62 | 6.85 | 6.76 | 7.16 | 7.61 |
| CF (%) | 2.46 | 2.46 | 2.84 | 3.23 | 3.61 | 3.02 | 3.58 | 4.14 |
| Calcium (%) | 1.05 | 1.05 | 1.04 | 1.03 | 1.03 | 1.04 | 1.04 | 1.03 |
| Available phosphorous (%) | 0.45 | 0.45 | 0.43 | 0.41 | 0.39 | 0.43 | 0.41 | 0.39 |
| Lysine (%) | 1.16 | 1.16 | 1.16 | 1.16 | 1.17 | 1.16 | 1.16 | 1.17 |
| Methionine (%) | 0.56 | 0.56 | 0.56 | 0.57 | 0.57 | 0.56 | 0.57 | 0.57 |
aNC (negative control), PC (positive control), FDBG5% (5% fermented dried brewers grains), FDBG10% (10% fermented dried brewer grain), FDBG15% (15% fermented dried brewers grains), DBG5%+Enz (5%dried brewers grains mixed with enzymes), DBG10%+Enz (10% dried brewers grains mixed with enzymes), DBG15%+Enz (15% dried brewers grains mixed with enzymes). bCommercial multi-exogenous enzyme was added was added to PC and enzyme treated DBG diets at a concentration of 1 gm/kg diet. cFDBG (fermented dried brewer grains). dDBG (dried brewer grains). eVitamin premix supplied per kilogram of diet: vitamin A, 10,000 IU; vitamin D3, 2000 IU; vitamin E, 6500 IU; vitamin K3, 1 mg; vitamin B1, 2560 mg; vitamin B2, 5000 mg; vitamin B6, 1500 mg; B5, 8 mg; niacin, 20000 mg; biotin, 0.25 mg; folic acid, 1000 mg; vitamin B12, 60 mg; Cu, 8 mg; Fe, 80 mg; Mn, 60 mg; Zn, 40 mg; Se, 0.15 mg
Samples collection
At the end of experimental period. The birds euthanized by cervical dislocation and the blood samples (n = 5 per replicate) were collected in sterilized tubes from the brachial vein and the serum was separated for biochemical analyses. Following blood collection, these birds were manually defeathered and eviscerated to calculate the carcass weight together with the weights of gizzard, liver, spleen, thymus, bursa and abdominal fat. Samples (n = 5 per replicate) from breast and thigh meat were quickly obtained and stored at -20ºC until analysis of moisture, protein, and cholesterol in breast and cholesterol content in thigh. Small pieces (n = 5 per replicate), approximately 2 cm in the middle of proventriculus and pancreas and 4 cm of duodenum (distal loop) were excised, flushed with phosphate buffer saline, collected into Eppendorf cap lock tube and stored in -80ºC for subsequent RNA extraction.
Measurement of growth performance and carcass traits:
Birds and feed intake in each pen were weighed weekly and body weight gain (BWG), feed intake (FI), and feed conversion ratio (FCR) were calculated. At the end of experimental period BWG, FI and FCR were calculated for total growing period (days 1–38). The carcass weight (dressing yield) was calculated as the percentage of live weight and weight of gizzard, liver, spleen, thymus, bursa and abdominal fat was calculated as a percentage of carcass weight.
RNA Extraction, Reverse Transcription, and Quantitative Real-Time PCR
For each sample, total RNA was extracted from the proventriculus, pancreas and duodenum by RNeasy Mini Kit (Qiagen, Cat. No. 74,104) according to the manufacturer’s directions. The quantity and purity of total RNA was determined using a NanoDrop (ND-8000 spectrophotometer, Thermo Fisher Scientific, Waltham, USA). The isolated RNA of each sample was reverse transcribed using RevertAid™ H Minus kits (Fermentas Life Science, Pittsburgh, PA, USA). The cDNA (One µL) was mixed with 2x SYBR® Green PCR mix (12.5 µL), and RNase free water (5.5 µL), then 0.5 µL of each forward and reverse primer for the selected genes were added. The primers’ sequences of selected digestive and nutrient transporters genes were designed as previously described in Kheravii et al. [55], Table 8. The GAPDH was used as an internal control to normalize target gene expression levels [94]. The real-time PCR amplification was done with Rotor-Gene Q2 plex (Qiagen Inc., Valencia, CA, USA).
Table 8.
Primers Sequences and target genes used for quantitative real-time PCR
| Genesa | Gene full name | Primer sequence (5′-3′) | Accession No. |
|---|---|---|---|
| PGA5 | Pepsinogen A |
F-TCCGTCTACCTGAGCAAGGAT R- AAGCAGGCGACGTACTTGTT |
NM_204878.1 |
| PGC | Pepsinogen C |
F-ATCGGGATTGAGGA↓CTTCGC R- TGAAGACCTGGTTGGGAACG |
NM_204877.2 |
| AMY2A | Pancreatic alpha 2A amylase |
F-CGGAGTG↓GATGTTAACGACTGG R-ATGTTCGCAGACCCAGTCATTG |
NM_001001473.2 |
| PNLIP | Pancreatic lipase |
F-GCATCTGGGAAG↓GAACTAGGG R- TGAACCACAAGCATAGCCCA |
NM_001277382.1 |
| CCK | Cholecystokinin |
F-AGGTTCCACTGGGAGGTTCT R-CGCCTGCTGTTCTTTAGGAG |
XM_015281332.1 |
| CELA1 | Chymotrypsin-like elastase family, member 1 |
F-AGCGTAAGGAAATGGGGTGG R-GTGGAGACCCCATGCAAGTC |
XM_015300368.1 |
| GLUT1 | Glucose transporter-1 (SLC2A1) |
F-TCCTCCTGATCAACCGCAAT R-TGTGCCCCGGAGCTTCT |
NM_205209.1 |
| GLUT2 | Glucose transporter-2 (SLC2A2) |
F-TGATCGTGGCACTGATGGTT R-CCACCAGGAAGAC↓GGAGATA |
NM_207178.1 |
| CAT1 | Cationic amino acid transporter-1 (SLC7A1) |
F-CAAGAGGAAAACTCCAGTAATTGCA R- AAGTCGAAGAGGAAGGCCATAA |
XM_015277945.1 |
| CAT2 | Cationic amino acid transporter-2 (SLC7A2) |
F-TGCTCGCGTTCCCAAGA R- GGCCCACAGTTCACCAACAG |
XM_015285435.1 |
| LAT1 | L-type amino acid transporter-1 (SLC7A7) |
F-GATTGCAACGGGTGATGTGA R- CCCCACACCCACTTTTGTTT |
KT876067.1 |
| PepT1 | Peptide transporter-1 (SLC15A1) |
F-TACGCATACTGTCACCATCA R-TCCTGAGAACGGACTGTAAT |
AY029615.1 |
| PepT2 | Peptide transporter-2 (SLC15A2) |
F-TGACTGGGCATCGGAACAA R-ACCCGTGTCACCATTTTAACCT |
NM_001319028.1 |
| GAPDH | Glyceraldahyde -3-phosphate dehydrogenase |
F-GGTGGTGCTAAGCGTGTTA R-CCCTCCACAATGCCAA |
NM205518 |
aThe genes analyzed in the tissues are listed as follow: PGA5 and, PGC in proventriculus; AMY2A, CCK1R, CCK, CELA1, PNLIP, in pancreas; and, GLUT1, GLUT2, CAT1, CAT2, LAT1, PepT1, PepT2 in duodenum
Biochemical analyses
The serum biochemical indices: total plasma proteins, albumin, globulin, total cholesterol, triglycerides, high density conjugated protein (HDL), low density protein (LDL), alanine aminotransferase (ALT) and aspartate amino transferase (AST) were measured using diagnostic kits (Spinreact, Santa Coloma, Spain).
Analysis of cholesterol content in breast and thigh meat
The moisture and protein content of breast were calculated according to AOAC [92]. The total cholesterol in breast and thigh meat was determined by gas chromatography, as previously reported by Allain et al. [95].
Economic analysis
The economic efficiency of substituting corn-soybean by fermented or enzyme-treated DBG was calculated from the input-output analysis (as per the prevailing value of the experimental diets and also the broiler body weight during the experimental period) as follows:
Total feed cost = total feed intake per bird × cost of one kg diet [96].
Feed cost/kg BW gain = feed conversion × cost of one kg diet [97].
Total expenses = total feed cost + average fixed costs [2].
Total revenue = live body weight × price/kg [98].
Gross margin = total revenue – total expenses [98].
Benefit-cost ratio = gross margin/total feed cost [99].
Statistical analyses
The data were analyzed using GLM procedure of SPSS, after confirming the homogeneity among experimental groups using Levene’s test and normality using Shapiro-Wilk’s test. Tukey’s post hoc was used to test for significant differences between the mean values. Variation in the data was expressed as standard error of the mean (SEM) and the significance was set at 0.05. Relative fold changes in the expression of target genes calculated by the 2−ΔΔCt method as described by Livak and Schmittgen [100].
Acknowledgements
Not applicable.
Abbreviations
- DBG
Dried brewer grains
- FDBG
Fermented dried brewer grains
- NC
Negative control
- PC
Positive control
- Enz
Enzyme
- PGC
Pepsinogen C
- PGA5
Pepsinogen A
- AMY2A
Pancreatic alpha 2A amylase
- PNLIP
Pancreatic lipase
- CELA1
Chymotrypsin-like elastase family, member 1
- CCK
Cholecystokinin
- CAT1
Cationic amino acid transporter-1 (SLC7A1)
- CAT2
Cationic amino acid transporter-1 (SLC7A2)
- GLUT1
Glucose transporter-1 (SLC2A1)
- GLUT2
Glucose transporter-2 (SLC2A2)
- LAT1
L-type amino acid transporter-1
- Pep1
Peptide transporter-1 (SLC15A1)
- Pep2
Peptide transporter-2 (SLC15A2)
- GAPDH
Glyceraldahyde − 3-phosphate dehydrogenase
- ANF
Antinutritional factors
- NSP
Non-starch polysaccharides
- LAB
Lactic acid bacteria
- BWG
Body weight gain
- FI
Feed intake
- FCR
Feed conversion ratio
- HDL
High density conjugated protein
- LDL
Low density protein
- ALT
Alanine aminotransferase and
- AST
Aspartate amino transferase
- GLM
General linear method
- SEM
Standard error of the mean
- AA
Amino acids
Authors' contributions
All authors have read and approved the manuscript. Conceptualization, D.I, S.E.S, A.E.O, H.S.A, H.A.I and H.A.M.; methodology, D.I, S.E.S, A.E.O, H.S.A, H.A.I and H.A.M., software, D.I, S.E.S, H.A.M.; validation, D.I, S.E.S and A.E.O.; formal analysis, D.I, H.A.I and H.A.M.; investigation, .I, S.E.S, A.E.O, H.S.A, H.A.I and H.A.M.; resources, .I, S.E.S, A.E.O, H.S.A, H.A.I and H.A.M.; data curation, X.X.; writing—original draft preparation, D.I, S.E.S, A.E.O, H.S.A, H.A.I and H.A.M., writing—review and editing, D.I, S.E.S, A.E.O, H.S.A, and H.A.M visualization, D.I, S.E.S, A.E.O, H.S.A, H.A.I and H.A.M.; supervision, D.I, A.E.O, H.S.A, and H.A.M; funding acquisition, D.I, S.E.S, A.E.O, H.S.A, H.A.I and H.A.M. The authors have read and approved the manuscript.
Funding
This research received no external funding.
Ethics approval and consent to participate
All animal protocols and arrival guidelines were approved by the Institutional Animal Care and Use Committee (ZU-IACUC/3/M/2019), Poultry Research Unit, Faculty of Veterinary Medicine, Zagazig University.
Consent for publication
Not applicable.
Availability of data and materials
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Hanan S. Al-Khalaifah, Email: hkhalifa@safat.kisr.edu.kw
Doaa Ibrahim, Email: doibrahim@vet.zu.edu.eg.
References
- 1.Al-Sagheer AA, Abd El-Hack ME, Alagawany M, Naiel MA, Mahgoub SA, Badr MM, Hussein EO, Alowaimer AN, Swelum AA. Paulownia leaves as a new feed resource: Chemical composition and effects on growth, carcasses, digestibility, blood biochemistry, and intestinal bacterial populations of growing rabbits. Animals. 2019;9(3):95. doi: 10.3390/ani9030095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Fasuyi A, Aturamu O, Lawal A. Part II: Economic analyses and the growth performance of broiler finisher (29–58 day) birds on high fibre-low protein industrial plant by-products. Asian J Adv Agric Res. 2018;6(3):1–10. [Google Scholar]
- 3.Aregheore E, Abdulrazak S. Estimation of organic matter digestibility and metabolizable energy content of agro-industrial wastes using in vitro gas production. Nig J Anim Prod. 2005;32(1):79–87. [Google Scholar]
- 4.Khalili A, Tabeidian SA, Toghyani M, Ghalamkari G, Bahrami Y. Effect of different levels of brewer’s dried grains and enzyme on performance, protein digestibility, immune response and performance of broilers. Int J Acad Res. 2011;3(2):1153–7. [Google Scholar]
- 5.McCarthy AL. Novel ingredients from brewers’ spent grain-bioactivity in cell culture model systems and bioactivity retention in fortified food products. 2013. [Google Scholar]
- 6.Radzik-Rant A, Rant W, Niżnikowski R, Świątek M, Szymańska Ż, Ślęzak M, Niemiec T. The effect of the addition of wet brewers grain to the diet of lambs on body weight gain, slaughter valueand meat quality. Archiv fuer Tierzucht. 2018;61(2):245. [Google Scholar]
- 7.ME AE-H, Alagawany M, Patra A, Abdel-Latef M, Ashour E, Arif M, Farag M, Dhama K. Use of brewers dried grains as an un-conventional feed ingredient in the diets of broiler chickens: a review. Adv Anim Vet Sci. 2019;7(3):218–24. [Google Scholar]
- 8.Ashour E, El-Hack MA, El-Hindawy M, Attia A, Osman A, Swelum A, Alowaimer A, Saadeldin I, Laudadio V. Impacts of dietary inclusion of dried brewers’ grains on growth, carcass traits, meat quality, nutrient digestibility and blood biochemical indices of broilers. S Afr J Anim Sci. 2019;49(3):573–84. doi: 10.4314/sajas.v49i3.18. [DOI] [Google Scholar]
- 9.Isikwenu J. Performance and economic analysis of cockerel chicks fed enzyme supplemented brewer’s dried grains groundnut cake-based diets. Agric Biol J North Am. 2011;2:47–51. doi: 10.5251/abjna.2011.2.1.47.51. [DOI] [Google Scholar]
- 10.Alabi O, Atteh J, Adejumo I, Ogundele O (2014) Effects of dietary levels of brewers’ dried grain supplemented with commercial enzymes on performance, nutrient retention and gastro-intestinal tract characteristics of Arbor acres broilers. Int J Agric Innov Res 2(5):818–23.
- 11.Ravindran V. Feed enzymes: The science, practice, and metabolic realities. J Appl Poultry Res. 2013;22(3):628–36. doi: 10.3382/japr.2013-00739. [DOI] [Google Scholar]
- 12.Niba A, Beal J, Kudi A, Brooks P. Potential of bacterial fermentation as a biosafe method of improving feeds for pigs and poultry. Afr J Biotech . 2009;8(9):1758–1767. [Google Scholar]
- 13.Sugiharto S, Yudiarti T, Isroli I. Functional properties of filamentous fungi isolated from the Indonesian fermented dried cassava, with particular application on poultry. Mycobiology. 2015;43(4):415–22. doi: 10.5941/MYCO.2015.43.4.415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Borresen C, Erica J, Henderson A, Kumar A, L Weir T, Ryan P. Fermented foods: patented approaches and formulations for nutritional supplementation and health promotion. Recent Pat Food Nutr Agric. 2012;4(2):134–40. doi: 10.2174/2212798411204020134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sugiharto S, Yudiarti T, Isroli I. Performances and haematological profile of broilers fed fermented dried cassava (Manihot esculenta Crantz) Trop Anim Health Prod. 2016;48(7):1337–41. doi: 10.1007/s11250-016-1098-2. [DOI] [PubMed] [Google Scholar]
- 16.Xu F, Zeng X, Ding X. Effects of replacing soybean meal with fermented rapeseed meal on performance, serum biochemical variables and intestinal morphology of broilers. Asian-Australas J Anim Sci. 2012;25(12):1734. doi: 10.5713/ajas.2012.12249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Canibe N, Jensen BB. Fermented liquid feed—Microbial and nutritional aspects and impact on enteric diseases in pigs. Anim Feed Sci Technol. 2012;173(1–2):17–40. doi: 10.1016/j.anifeedsci.2011.12.021. [DOI] [Google Scholar]
- 18.Shahowna E, Mahala A, Mokhtar A, Amasaib E, Attaelmnan B. Evaluation of nutritive value of sugar cane bagasse fermented with poultry litter as animal feed. Afr J Food Sci Technol. 2013;4:106e109. [Google Scholar]
- 19.Tan YX, Mok WK, Lee J, Kim J, Chen WN. Solid state fermentation of brewers’ spent grains for improved nutritional profile using bacillus subtilis wx-17. Fermentation. 2019;5(3):52. doi: 10.3390/fermentation5030052. [DOI] [Google Scholar]
- 20.Poncet N, Taylor PM. The role of amino acid transporters in nutrition. Curr Opin Clin Nutr Metab Care. 2013;16(1):57–65. doi: 10.1097/MCO.0b013e32835a885c. [DOI] [PubMed] [Google Scholar]
- 21.Kheravii S, Morgan N, Swick RA, Choct M, Wu S-B. Roles of dietary fibre and ingredient particle size in broiler nutrition. Worlds Poult Sci J. 2018;74(2):301–16. doi: 10.1017/S0043933918000259. [DOI] [Google Scholar]
- 22.Ruhnke I, Röhe I, Goodarzi Boroojeni F, Knorr F, Mader A, Hafeez A, Zentek J. Feed supplemented with organic acids does not affect starch digestibility, nor intestinal absorptive or secretory function in broiler chickens. J Anim Physiol Anim Nutr (Berl) 2015;99:29–35. doi: 10.1111/jpn.12313. [DOI] [PubMed] [Google Scholar]
- 23.Fotiadis D, Kanai Y, Palacín M. The SLC3 and SLC7 families of amino acid transporters. Mol Aspects Med. 2013;34(2–3):139–58. doi: 10.1016/j.mam.2012.10.007. [DOI] [PubMed] [Google Scholar]
- 24.Chen H, Mao X, Yin J, Yu B, He J, Che L, Yu J, Huang Z, Zheng P, Michiels J (2015) Comparison of jejunal digestive enzyme activities, expression of nutrient transporter genes, and apparent fecal digestibility in weaned piglets fed diets with varied sources of fiber. J Anim Feed Sci 24(1):41–7.
- 25.Röder PV, Geillinger KE, Zietek TS, Thorens B, Koepsell H, Daniel H. The role of SGLT1 and GLUT2 in intestinal glucose transport and sensing. PLoS One. 2014;9(2):e89977. doi: 10.1371/journal.pone.0089977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Alshelmani MI, Loh TC, Foo HL, Sazili AQ, Lau WH. Effect of solid state fermentation on nutrient content and ileal amino acids digestibility of palm kernel cake in broiler chickens. Indian J Anim Sci. 2017;87:1135–40. [Google Scholar]
- 27.Zhang J, Zhu J, Sun J, Li Y, Wang P, Jiang R, Rahman MRT. Effect of fermented feed on intestinal morphology, immune status, carcass and growth performance of Emei Black chickens. FASEB J. 2016;30(1_supplement):lb240–0. [Google Scholar]
- 28.Shi C, Zhang Y, Lu Z, Wang Y. Solid-state fermentation of corn-soybean meal mixed feed with Bacillus subtilis and Enterococcus faecium for degrading antinutritional factors and enhancing nutritional value. J Anim Sci Biotechnol. 2017;8(1):50. doi: 10.1186/s40104-017-0184-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Khempaka S, Thongkratok R, Okrathok S, Molee W. An evaluation of cassava pulp feedstuff fermented with A. oryzae, on growth performance, nutrient digestibility and carcass quality of broilers. Poult Sci. 2013;51(1):71–79. doi: 10.2141/jpsa.0130022. [DOI] [Google Scholar]
- 30.Jazi V, Boldaji F, Dastar B, Hashemi S, Ashayerizadeh A. Effects of fermented cottonseed meal on the growth performance, gastrointestinal microflora population and small intestinal morphology in broiler chickens. Br Poult Sci. 2017;58(4):402–8. doi: 10.1080/00071668.2017.1315051. [DOI] [PubMed] [Google Scholar]
- 31.Zhang W-J, Xu Z-R, Zhao S-H, Sun J-Y, Yang X. Development of a microbial fermentation process for detoxification of gossypol in cottonseed meal. Anim Feed Sci Technol. 2007;135(1–2):176–86. doi: 10.1016/j.anifeedsci.2006.06.003. [DOI] [Google Scholar]
- 32.Sun H, Jiang-Wu T, Xiao-Hong Y, Yi-Fei W, Wang X, Feng J. Improvement of the nutritional quality of cottonseed meal by Bacillus subtilis and the addition of papain. Int J Agric Biol. 2012;14(4):563–568. [Google Scholar]
- 33.Sun H, Yao X, Wang X, Wu Y, Liu Y, Tang J, Feng J (2015) Chemical composition and in vitro antioxidant property of peptides produced from cottonseed meal by solid-state fermentation. CyTA-J Food 13(2):264–72.
- 34.Sugiharto S, Ranjitkar S. Recent advances in fermented feeds towards improved broiler chicken performance, gastrointestinal tract microecology and immune responses: A review. Animal nutrition. 2018;5(1):1. doi: 10.1016/j.aninu.2018.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Yeh RH, Hsieh CW, Chen KL. Screening lactic acid bacteria to manufacture two-stage fermented feed and pelleting to investigate the feeding effect on broilers. Poult Sci. 2018;97(1):236–46. doi: 10.3382/ps/pex300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ries EF, Alves Macedo G (2011) Improvement of phytase activity by a new Saccharomyces cerevisiae strain using statistical optimization. Enzyme Res 2011:796394. [DOI] [PMC free article] [PubMed]
- 37.Cenamor R, Molina M, Galdona J, Sánchez M (1987) Production and secretion of Saccharomyces cerevisiae β-glucanases: differences between protoplast and periplasmic enzymes. Microbiology 133(3):619–28.
- 38.Arumugam G, Sadiq AM, Nagalingam M, Panneerselvam A. Production of invertase enzymes from Saccharomyces cerevisiae strain isolated from sugarcane and grape juices. Eur J Exp Biol. 2014;4(5):29–32. [Google Scholar]
- 39.Sugiharto S, Ranjitkar S (2019) Recent advances in fermented feeds towards improved broiler chicken performance, gastrointestinal tract microecology and immune responses: A review. Anim Nutr 5(1):1–10. [DOI] [PMC free article] [PubMed]
- 40.Engberg RM, Hammersh⊘ j M, Johansen N, Abousekken MS, Steenfeldt S, Jensen BB. Fermented feed for laying hens: effects on egg production, egg quality, plumage condition and composition and activity of the intestinal microflora. Br Poult Sci. 2009;50(2):228–39. doi: 10.1080/00071660902736722. [DOI] [PubMed] [Google Scholar]
- 41.Cutler S, Rasmussen M, Hensley M, Wilhelms K, Griffith R, Scanes C. Effects of Lactobacilli and lactose on Salmonella typhimurium colonisation and microbial fermentation in the crop of the young turkey. Br Poult Sci. 2005;46(6):708–16. doi: 10.1080/00071660500393694. [DOI] [PubMed] [Google Scholar]
- 42.Niba A, Beal J, Kudi A, Brooks P. Bacterial fermentation in the gastrointestinal tract of non-ruminants: influence of fermented feeds and fermentable carbohydrates. Trop Anim Health Prod. 2009;41(7):1393. doi: 10.1007/s11250-009-9327-6. [DOI] [PubMed] [Google Scholar]
- 43.Missotten J, Michiels J, Dierick N, Ovyn A, Akbarian A, De Smet S. Effect of fermented moist feed on performance, gut bacteria and gut histo-morphology in broilers. Br Poult Sci. 2013;54(5):627–34. doi: 10.1080/00071668.2013.811718. [DOI] [PubMed] [Google Scholar]
- 44.Sun H, Tang J, Fang C, Yao X, Wu Y, Wang X, Feng J. Molecular analysis of intestinal bacterial microbiota of broiler chickens fed diets containing fermented cottonseed meal. Poult Sci. 2013;92(2):392–401. doi: 10.3382/ps.2012-02533. [DOI] [PubMed] [Google Scholar]
- 45.Xie Pj H, Lx, Zhang Ch. Nutrient assessment of olive leaf residues processed by solid-state fermentation as an innovative feedstuff additive. J Appl Microbiol. 2016;121(1):28–40. doi: 10.1111/jam.13131. [DOI] [PubMed] [Google Scholar]
- 46.Gao Z, Wu H, Shi L, Zhang X, Sheng R, Yin F, Gooneratne R (2017) Study of Bacillus subtilis on growth performance, nutrition metabolism and intestinal microflora of 1 to 42 d broiler chickens. Anim Nutr 3(2):109–13. [DOI] [PMC free article] [PubMed]
- 47.Li W, Bai J, Li Y, Qin Y, Yu D. Effects of Bacillus subtilis on meat quality, nutrient digestibility and serum biochemical parameters of broilers. Chin J Vet Sci. 2014;34(10):1682–5. [Google Scholar]
- 48.Shankar PA, Premavalli K, Omprakash A, Kirubakaran JJ, Hudson G, Vairamuthu S. Effect of Dietary Yeast Supplementation on Serum Biochemical Profile of Broiler Chicken. 2017;95(6):13–15. [Google Scholar]
- 49.Horvatovic M, Glamocic D, Zikic D, Hadnadjev T (2015) Performance and some intestinal functions of broilers fed diets with different inclusion levels of sunflower meal and supplemented or not with enzymes. Braz J Poultry Sci 17(1):25–30.
- 50.Almirall M, Francesch M, Perez-Vendrell AM, Brufau J, Esteve-Garcia E. The differences in intestinal viscosity produced by barley and β-glucanase alter digesta enzyme activities and ileal nutrient digestibilities more in broiler chicks than in cocks. J Nutr. 1995;125(4):947–55. doi: 10.1093/jn/125.4.947. [DOI] [PubMed] [Google Scholar]
- 51.Bedford M, Cowieson A. Exogenous enzymes and their effects on intestinal microbiology. Anim Feed Sci Technol. 2012;173(1–2):76–85. doi: 10.1016/j.anifeedsci.2011.12.018. [DOI] [Google Scholar]
- 52.Singh A, Tiwari U, Berrocoso J, Dersjant-Li Y, Awati A, Jha R. Effects of a combination of xylanase, amylase and protease, and probiotics on major nutrients including amino acids and non-starch polysaccharides utilization in broilers fed different level of fibers. Poult Sci. 2019;98(11):5571–81. doi: 10.3382/ps/pez310. [DOI] [PubMed] [Google Scholar]
- 53.Kalantar M, Khajali F, Yaghobfar A. Different dietary source of non-starch polysaccharides supplemented with enzymes affected growth and carcass traits, blood parameters and gut physicochemical properties of broilers. Global J Anim Sci Res. 2015;3:412–8. [Google Scholar]
- 54.Zeng Q, Huang X, Luo Y, Ding X, Bai S, Wang J, Xuan Y, Su Z, Liu Y, Zhang K. Effects of a multi-enzyme complex on growth performance, nutrient utilization and bone mineralization of meat duck. J Anim Sci Biotechnol. 2015;6(1):12. doi: 10.1186/s40104-015-0013-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kheravii S, Swick RA, Choct M, Wu S-B. Upregulation of genes encoding digestive enzymes and nutrient transporters in the digestive system of broiler chickens by dietary supplementation of fiber and inclusion of coarse particle size corn. BMC Genom. 2018;19(1):208. doi: 10.1186/s12864-018-4592-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ferket P. Feeding whole grains to poultry improves gut health. Feedstuffs. 2000;72(37):12–3. [Google Scholar]
- 57.González-Alvarado J, Jiménez-Moreno E, Valencia D, Lázaro R, Mateos G. Effects of fiber source and heat processing of the cereal on the development and pH of the gastrointestinal tract of broilers fed diets based on corn or rice. Poult Sci. 2008;87(9):1779–95. doi: 10.3382/ps.2008-00070. [DOI] [PubMed] [Google Scholar]
- 58.Svihus B. The gizzard: function, influence of diet structure and effects on nutrient availability. Worlds Poult Sci J. 2011;67(2):207–24. doi: 10.1017/S0043933911000249. [DOI] [Google Scholar]
- 59.Pacheco W, Stark C, Ferket P, Brake J. Effects of trypsin inhibitor and particle size of expeller-extracted soybean meal on broiler live performance and weight of gizzard and pancreas. Poult Sci. 2014;93(9):2245–52. doi: 10.3382/ps.2014-03986. [DOI] [PubMed] [Google Scholar]
- 60.Gabriel I, Mallet S, Leconte M, Travel A, Lalles J. Effects of whole wheat feeding on the development of the digestive tract of broiler chickens. Anim Feed Sci Technol. 2008;142(1–2):144–62. doi: 10.1016/j.anifeedsci.2007.06.036. [DOI] [Google Scholar]
- 61.Lan Y, Williams BA, Tamminga S, Boer H, Akkermans A, Erdi G, Verstegen MW. In vitro fermentation kinetics of some non-digestible carbohydrates by the caecal microbial community of broilers. Anim Feed Sci Technol. 2005;123:687–702. doi: 10.1016/j.anifeedsci.2005.04.027. [DOI] [Google Scholar]
- 62.Saleh AA, Ali H, Abdel-Latif MA, Emam MA, Ghanem R, El-Hamid HSA. Exogenous dietary enzyme formulations improve growth performance of broiler chickens fed a low-energy diet targeting the intestinal nutrient transporter genes. PLoS One. 2018;13(5):e0198085. doi: 10.1371/journal.pone.0198085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Brenes A, Smith M, Guenter W, Marquardt R. Effect of enzyme supplementation on the performance and digestive tract size of broiler chickens fed wheat-and barley-based diets. Poult Sci. 1993;72(9):1731–9. doi: 10.3382/ps.0721731. [DOI] [PubMed] [Google Scholar]
- 64.Zhao F, Hou S, Zhang H, Zhang Z. Effects of dietary metabolizable energy and crude protein content on the activities of digestive enzymes in jejunal fluid of Peking ducks. Poult Sci. 2007;86(8):1690–5. doi: 10.1093/ps/86.8.1690. [DOI] [PubMed] [Google Scholar]
- 65.Lin P, Shih B, Hsu J. Effects of different sources of dietary non-starch polysaccharides on the growth performance, development of digestive tract and activities of pancreatic enzymes in goslings. Br Poult Sci. 2010;51(2):270–7. doi: 10.1080/00071661003779124. [DOI] [PubMed] [Google Scholar]
- 66.Yokhana J, Parkinson G, Frankel T. Effect of insoluble fiber supplementation applied at different ages on digestive organ weight and digestive enzymes of layer-strain poultry. Poult Sci. 2016;95(3):550–9. doi: 10.3382/ps/pev336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Bogułsawska-Tryk M. Effect of different levels of cellulose in the diet on the proteolytic activity of the pancreas in broiler chickens. Folia Biol. 2005;53(4):19–23. doi: 10.3409/173491605775789353. [DOI] [Google Scholar]
- 68.Hetland H, Svihus B, Krogdahl Å. Effects of oat hulls and wood shavings on digestion in broilers and layers fed diets based on whole or ground wheat. Br Poult Sci. 2003;44(2):275–82. doi: 10.1080/0007166031000124595. [DOI] [PubMed] [Google Scholar]
- 69.Li W-F, Feng J, Xu Z-R, Yang C-M. Effects of non-starch polysaccharides enzymes on pancreatic and small intestinal digestive enzyme activities in piglet fed diets containing high amounts of barley. World J Gastroenterol. 2004;10(6):856. doi: 10.3748/wjg.v10.i6.856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Kheravii SK, Choct M, Swick R, Wu R. Improved performance of broilers by structural manipulation of feed: Evidence from gizzard development, nutrient digestibility, gut microflora and gene expression. 2017. [Google Scholar]
- 71.Kheravii S, Swick RA, Choct M, Wu S-B. Dietary sugarcane bagasse and coarse particle size of corn are beneficial to performance and gizzard development in broilers fed normal and high sodium diets. Poult Sci. 2017;96(11):4006–16. doi: 10.3382/ps/pex225. [DOI] [PubMed] [Google Scholar]
- 72.Feng J, Liu X, Xu Z, Wang Y, Liu J. Effects of fermented soybean meal on digestive enzyme activities and intestinal morphology in broilers. Poult Sci. 2007;86(6):1149–54. doi: 10.1093/ps/86.6.1149. [DOI] [PubMed] [Google Scholar]
- 73.Karásková K, Suchý P, Straková E. Current use of phytogenic feed additives in animal nutrition: a review. Czech J Anim Sci. 2015;60(12):521–30. doi: 10.17221/8594-CJAS. [DOI] [Google Scholar]
- 74.Pluske JR, Williams IH, Aherne FX. Nutrition of the neonatal pig. In: Varley M.A., editor. The neonatal pig - development and survival. Wallingford: CAB International; 1995. p. 187–235.
- 75.Chiang G, Lu W, Piao X, Hu J, Gong L, Thacker P. Effects of feeding solid-state fermented rapeseed meal on performance, nutrient digestibility, intestinal ecology and intestinal morphology of broiler chickens. Asian-Australas J Anim Sci. 2009;23(2):263–71. doi: 10.5713/ajas.2010.90145. [DOI] [Google Scholar]
- 76.Lee K-W, Kim DK, Lillehoj HS, Jang SI, Lee S-H. Immune modulation by Bacillus subtilis-based direct-fed microbials in commercial broiler chickens. Anim Feed Sci Technol. 2015;200:76–85. doi: 10.1016/j.anifeedsci.2014.12.006. [DOI] [Google Scholar]
- 77.Ferraris RP. Dietary and developmental regulation of intestinal sugar transport. Biochem J. 2001;360(2):265–76. doi: 10.1042/bj3600265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Lee SA, Wiseman J, Masey O’Neill HV, Scholey DV, Burton EJ, Hill SE (2017) Understanding the direct and indirect mechanisms of xylanase action on starch digestion in broilers. J World’s Poultry Res 7(2):35–47.
- 79.Hosseini SM, Manafi M, Nazarizadeh H. Effects of xylanase supplementation and citric acid on performance, ileal nutrients digestibility, and gene expression of intestinal nutrient transporters in broilers challenged with Clostridium perfringens. The Journal of Poultry Science. 2016;54(2):149–156. doi: 10.2141/jpsa.0160099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Wang Y, Gu Q. Effect of probiotic on growth performance and digestive enzyme activity of Arbor Acres broilers. Res Vet Sci. 2010;89(2):163–7. doi: 10.1016/j.rvsc.2010.03.009. [DOI] [PubMed] [Google Scholar]
- 81.Shang HM, Zhao JC, Guo Y, Zhang HX, Song H. Effects of supplementing feed with fermentation concentrate of Hericium caput-medusae (Bull.: Fr.) Pers. on cholesterol deposition in broiler chickens. Livestock Science. 2020. p. 104009.
- 82.Huang Y, Zheng Y. The probiotic Lactobacillus acidophilus reduces cholesterol absorption through the down-regulation of Niemann-Pick C1-like 1 in Caco-2 cells. Br J Nutr. 2010;103(4):473–8. doi: 10.1017/S0007114509991991. [DOI] [PubMed] [Google Scholar]
- 83.Ashayerizadeh A, Dastar B, Shargh MS, Mahoonak AS, Zerehdaran S (2018) Effects of feeding fermented rapeseed meal on growth performance, gastrointestinal microflora population, blood metabolites, meat quality, and lipid metabolism in broiler chickens. Livestock Sci 216:183–90.
- 84.Ramasamy K, Abdullah N, Wong MC, Karuthan C, Ho YW. Bile salt deconjugation and cholesterol removal from media by Lactobacillus strains used as probiotics in chickens. J Sci Food Agric. 2010;90(1):65–9. doi: 10.1002/jsfa.3780. [DOI] [PubMed] [Google Scholar]
- 85.Drażbo A, Kozłowski K, Ognik K, Zaworska A, Jankowski J. The effect of raw and fermented rapeseed cake on growth performance, carcass traits, and breast meat quality in turkey. Poult Sci. 2019;98(11):6161–9. doi: 10.3382/ps/pez322. [DOI] [PubMed] [Google Scholar]
- 86.Nie C, Zhang W, Ge W, Liu Y, Wang Y, Liu J (2015) Effect of cottonseed meal fermented with yeast on the lipid-related gene expression in broiler chickens. Braz J Poultry Sci 17(SPE):57–64.
- 87.Okorie K, Ehirim F, Gabriel O, Ikpe J, Okorie R, Okoro-Ugo C (2011) Effect of exogenous enzyme fortified palm kernel meal on the performance, carcass quality and biochemical profile of finisher broiler. Glob Res J Sci 1:86–92.
- 88.Adama T, Ogunbajo S, Mambo M (2007) Feed intake, growth performance and nutrient digestibility of broiler chicks fed diets containing varying levels of sorghum dried brewers’ grains. Int J Poultry Sci 6(8):592–598.
- 89.El-Bendary MA. Bacillus thuringiensis and Bacillus sphaericus biopesticides production. J Basic Microbiol. 2006;46(2):158–70. doi: 10.1002/jobm.200510585. [DOI] [PubMed] [Google Scholar]
- 90.Shim Y, Shinde P, Choi J, Kim J, Seo D, Pak J, Chae B, Kwon I. Evaluation of multi-microbial probiotics produced by submerged liquid and solid substrate fermentation methods in broilers. Asian-Australas J Anim Sci. 2010;23(4):521–9. doi: 10.5713/ajas.2010.90446. [DOI] [Google Scholar]
- 91.Naji SA, Al-Zamili I, Hasan S, Al-Gharawi J. The Effects of Fermented Feed on Broiler Production and Intestinal Morphology. Pertanika J Trop Agric Sci. 2016;39(4):597–607. [Google Scholar]
- 92.AOAC . Official methods of analysis of AOAC International, Association of Official Analytical Chemists. 2012. [Google Scholar]
- 93.Aviagen W. Ross 308: broiler’s management and nutrition specification. 2018. [Google Scholar]
- 94.Li YP, Bang DD, Handberg KJ, Jorgensen PH, Zhang MF. Evaluation of the suitability of six host genes as internal control in real-time RT-PCR assays in chicken embryo cell cultures infected with infectious bursal disease virus. Vet Microbiol. 2005;110(3–4):155–165. doi: 10.1016/j.vetmic.2005.06.014. [DOI] [PubMed] [Google Scholar]
- 95.Allain CC, Poon LS, Chan CS, Richmond W, Fu PC. Enzymatic determination of total serum cholesterol. Clin Chem. 1974;20(4):470–475. doi: 10.1093/clinchem/20.4.470. [DOI] [PubMed] [Google Scholar]
- 96.Surai P, Fisinin V. Selenium in poultry breeder nutrition: An update. Anim Feed Sci Technol. 2014;191:1–15. doi: 10.1016/j.anifeedsci.2014.02.005. [DOI] [Google Scholar]
- 97.Tufarelli V, Casalino E, D’Alessandro AG, Laudadio V. Dietary phenolic compounds: biochemistry, metabolism and significance in animal and human health. Curr Drug Metab. 2017;18(10):905–913. doi: 10.2174/1389200218666170925124004. [DOI] [PubMed] [Google Scholar]
- 98.Tag-el-Din SEI. Fixed vs. Variable Return Models: An Economic Analysis. Leicester: 8th Intensive Orientation Seminar on ÔIslamic Economics, Banking and Finance, Islamic; 1999. pp. 23–27. [Google Scholar]
- 99.Tabib MA, Karim M, Haque M, Khaliq Q, Solaiman A. Effect of planting arrangements on productivity of cotton + mungbean intercropping systems. Bangladesh Agronomy Journal. 2014;17(1):11–22. doi: 10.3329/baj.v17i1.23671. [DOI] [Google Scholar]
- 100.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2 – ∆∆CT method. Methods. 2001;25(4):402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.