

Publisher's Note: There is a Blood Commentary on this article in this issue.
Key Points
Inhibition of LSD1 promotes the propagation of human HSCs in vitro.
The LSD1-containing CoREST complex is a primary target of the HSC expansion molecule UM171.
Abstract
Culture conditions in which hematopoietic stem cells (HSCs) can be expanded for clinical benefit are highly sought after. Here, we report that inhibition of the epigenetic regulator lysine-specific histone demethylase 1A (LSD1) induces a rapid expansion of human cord blood–derived CD34+ cells and promotes in vitro propagation of long-term repopulating HSCs by preventing differentiation. The phenotype and molecular characteristics of cells treated with LSD1 inhibitors were highly similar to cells treated with UM171, an agent promoting expansion of HSCs through undefined mechanisms and currently being tested in clinical trials. Strikingly, we found that LSD1, as well as other members of the LSD1-containing chromatin remodeling complex CoREST, is rapidly polyubiquitinated and degraded upon UM171 treatment. CRISPR (clustered regularly interspaced short palindromic repeats)/Cas9 depletion of the CoREST core member, RCOR1, resulted in expansion of CD34+ cells similar to LSD1 inhibition and UM171. Taken together, LSD1 and CoREST restrict HSC expansion and are principal targets of UM171, forming a mechanistic basis for the HSC-promoting activity of UM171.
Visual Abstract

Introduction
Allogeneic hematopoietic stem cell (HSC) transplantation is a life-saving treatment for hematological disorders, but donor availability remains a major restriction. Umbilical cord blood (UCB) transplants offer several advantages, such as broader HLA compatibility and lower incidence of graft-versus-host disease1; however, they are primarily restricted to pediatric cases because of the limited number of hematopoietic stem and progenitor cells (HSPCs) present in a single cord blood unit.2 Therefore, successful development of strategies for ex vivo expansion of HSCs could enable curable HSC transplantation for large numbers of patients who are currently not eligible for transplantation therapy. This would further address current challenges in the generation of transplantable HSCs from embryonic stem cells and induced pluripotent stem cells, as well as the development of HSC gene editing and gene therapy applications for inherited hematological disorders.
Various in vitro expansion protocols have been proposed to facilitate the expansion of UCB HSPCs using combinations of growth factors and small molecule compounds. Small molecules targeting the aryl hydrocarbon receptor,3 p38 signaling,4 and histone deacetylase (HDAC) activity,5 as well as developmental factors, such as Notch ligand6 and the inflammation mediator prostaglandin E2,7 have all shown promising results in achieving HSPC expansion. Several of these factors are being exploited in clinical trials using transplantation of ex vivo–expanded UCB cells for the treatment of leukemia. In addition, UM171, a pyrimidoindole derivative, was shown to significantly expand the number of UCB HSCs; currently, UM171-expanded UCB HSPCs are being tested in a clinical trial for which preliminary results show improved early neutrophil recovery and reduced graft-versus-host disease following transplantation.8,9 However, the precise target of UM171, as well as the molecular mechanisms of its function, has not been defined.
Lysine-specific histone demethylase 1A (LSD1), also known as lysine (K)-specific demethylase 1A (KDM1A), regulates gene expression by specifically eliminating mono- and di-methyl groups on H3 lysine K4 and K9 residues.10,11 Studies in mice have shown that complete knockout of Lsd1 results in pancytopenia and functional impairment of HSCs,12 whereas conditional knockdown restricts hematopoietic differentiation and triggers an expansion of HSPCs in the bone marrow (BM).13 Lsd1 was shown to repress HSPC-specific gene expression, and loss of Lsd1 was associated with increased H3K4me1 and H3K4me2 marks on HSPC genes.12 Given that knockdown of Lsd1 expanded murine HSPCs in vivo, we hypothesized that targeting LSD1 in cultured human HSPCs might facilitate their expansion ex vivo.
In this study, we show that pharmacological inhibition of LSD1 promotes expansion of cultured human HSPCs and enhances the numbers of transplantable HSCs. We found that LSD1 inhibition triggered phenotypic and molecular responses that were highly similar to UM171 treatment and, remarkably, that UM171 treatment directly abrogates LSD1 by targeting the LSD1-containing CoREST complex. We conclude that LSD1 and CoREST are principal targets of UM171-mediated HSC expansion.
Methods
Cells
UCB samples were collected from maternity wards at Skåne University Hospital in Lund and Malmö, Sweden, as well as from Helsingborg General Hospital in Sweden. Normal BM samples were collected from healthy volunteers at Skåne University Hospital in Lund. All samples were collected after informed consent and approval by the regional ethical committee at Skåne University Hospital and Lund University. Mononuclear cell fractions from UBC and BM were isolated by density gradient centrifugation using Lymphoprep tubes (Alere Technologies; #1019818). CD34+ HSPCs were enriched by magnetic bead–based purification (Miltenyi Biotec; #130-046-703).
Flow cytometry
A BD LSRFortessa and a BD FACSCanto II were used to analyze samples. Cell sorting was performed using a BD FACSAria III. The following antibodies were used for fluorescence-activated cell sorting (FACS): CD34-FITC (#343604), EPCR-APC (#351906), CD11b-FITC (#367116), CD38-PE-Cy7 (#303516), CD45-APC (#304012), and CD33-PE (#303404; all from BioLegend; CD90-BV605 (#562685), CD38-PE (#345806), CD45RA-V450 (#560362), and CD19-BV605 (#562653; all from BD); and CD34-eFluor 450 (#48-0349-42) and CD3-PE-Cy7 (#25-0038-42; both from eBioscience). To measure cell proliferation, cells were labeled using carboxyfluorescein diacetate succinimidyl ester (CFSE) (eBioscience; #65-0850-84), and division history was measured with FACS.
In vitro culture and compounds
Cells were expanded in serum-free expansion medium (SFEM) (STEMCELL Technologies; #09650) supplemented with stem cell factor (SCF), thrombopoietin, and FMS-like tyrosine kinase 3 ligand at a final concentration of 100 ng/mL each (PeproTech; #300-07, #300-18, and #300-19). The LSD1 inhibitors 2-PCPA (#10010494), GSK-LSD1 (#16439), and RN-1 (#18124) were purchased from Cayman Chemical. UM171 (#72914) was obtained from STEMCELL Technologies. Compounds were dissolved in dimethyl sulfoxide (DMSO), as per the manufacturers’ recommendations.
LSD1 inhibitor screening assay
An LSD1 in vitro inhibition assay was performed using an LSD1 Inhibitor Screening Assay Kit (Cayman Chemical; #700120). The assay was performed as per the manufacturer’s instructions, and fluorescence was measured using a GloMAX Discover microplate reader (Promega; #GM3000).
Colony-forming unit assay
UCB CD34+ cells were expanded in vitro with DMSO, 2-PCPA, or UM171 for 6 days. From the expanded cells, 300 CD34+ cells were sorted per well and plated in MethoCult H4434 Classic (STEMCELL Technologies). Plates were incubated at 37°C for 14 days, and colonies were counted under a light microscope.
Transplantation
FACS-sorted CD34+ cells were cultured in SFEM supplemented with stem cell factor, thrombopoietin, and FMS-like tyrosine kinase 3 ligand, as well as with LSD1 inhibitors, UM171, or DMSO control, for 6 days and transplanted into sublethally irradiated (3 Gy) NSG mice. Peripheral blood samples from the tail vein were analyzed for human engraftment 5 weeks posttransplantation. BM samples were analyzed for different lineages and HSC markers at week 16. BM samples for limiting dilution analysis (LDA) assay were analyzed 18 weeks posttransplantation. Mice with human chimerism > 0.1% or human myeloid chimerism > 0.05% were regarded as positive, as used by other investigators.8 The frequency of long-term HSCs in each condition was calculated with an ELDA online tool using Poisson statistics.14 All animal studies were approved by the ethical committee at Lund University.
Microarray analysis
The UCB-derived HSC-enriched (CD34+CD38−CD90+CD45RA−) fraction was treated with DMSO or 2-PCPA for 24 hours. In the second experiment, the HSC-enriched fraction was treated with UM171 for 6 days, and CD34+CD90+ cells were FACS sorted and collected into RLT lysis buffer. RNA was isolated using a QIAGEN RNeasy Micro Kit (#74126), following the manufacturer’s instructions. RNA samples were amplified, labeled, and hybridized to Human Gene 2.0 ST array chips from Affymetrix (#902113). All of these experiments were performed at the Kompetenzzentrum Fluoreszents Bioanalytik (Regensburg, Germany). Raw data were processed and normalized using the Robust Multi-array Average algorithm.15 Gene set enrichment analysis (GSEA) was performed using the GSEA software tool from Broad Institute.16 Microarray data have been submitted to the Gene Expression Omnibus database (accession number GSE132747).
Western blot
CD34+ cord blood cells were treated with 2-PCPA, UM171, or DMSO for 24 hours. Cells were washed 3 times with phosphate-buffered saline and collected in IP Lysis Buffer (#87787) supplemented with protease inhibitor cocktail (both from Thermo Fisher Scientific; #87786). Samples were mixed with 2× Laemmli buffer and denatured for 5 minutes at 95°C. Western blotting was performed using a NuPAGE electrophoresis system, as per the manufacturer’s instructions (Thermo Fisher Scientific; Manual part no. IM-1001). The following primary antibodies were used: RCOR1 (#14567), LSD1 (#2184), HDAC1 (#34589T), HDAC2 (#5113T), H3K4me2 (#9725), and H3K9me2 (#4658; all from Cell Signaling Technology), Actin (#612656; from BD), and ANTI-FLAG M2 (#F3165; Sigma-Aldrich).
For polyubiquitination analysis, 10 million Flag-tagged RCOR1-expressing HL-60 cells were treated with UM171 and epoxomicin for 1.5 hours, and samples were processed as described.17 Flag-RCOR1 and endogenous LSD1 were enriched by immunoprecipitation (IP), and western blotting was performed as described above. Ubiquitin conjugates were detected with FK2 antibody (BML-PW0150-0025; Enzo Life Sciences). For co-IP experiments, 10 million HL-60 cells were lysed in IP Lysis Buffer (Pierce; #87788) supplemented with protease and phosphatase inhibitors (Thermo Fisher Scientific; #78441) and incubated overnight with shaking at 4°C with immunoglobulin G or LSD1. Antibody-protein complexes were captured with Protein A/G Magnetic Beads (Pierce; #26162).
Thermal proteome profiling analysis
Thermal proteome profiling (TPP) analysis was performed by the Chemical Proteomics core facility at Biomedicum (Karolinska Institute), part of the Chemical proteomics and Proteogenomics facility at SciLifeLab and of the Swedish National Infrastructure for Biological Mass Spectrometry. Two complete TPP experiments, each using duplicates of UM171-treated plus untreated controls, were performed, according to previously described protocols,18 in 2 acute myeloid leukemia (AML) cell lines (HL-60 and Kasumi-1). Cell cultures were treated with UM171 (1 µM) in complete culture medium for 2 hours. Proteomics sample processing was achieved by reduction with dithiothreitol, alkylation with iodoacetic acid, cold acetone precipitation, LysC and trypsin digestion, tandem mass tag 10 labeling, C18 desalting, and high pH reverse-phase fractionation. Mass spectrometry analysis was performed using a Q Exactive HF mass spectrometer equipped with an EASY-Spray Source and connected to an UltiMate 3000 RSLC Nano UPLC system (Thermo Fisher Scientific). Protein identification and quantification were performed using MaxQuant software19 and a UniProt complete human proteome database (UP000005640). The MaxQuant output was processed for TPP data analysis by an in-house–developed R library (in accordance with the original software package18) to fit sigmoid protein melting curves and derive the melting temperature difference between UM171-treated and untreated samples.
CRISPR (clustered regularly interspaced short palindromic repeats)/Cas9 gene editing
Three single guide RNAs (sgRNAs) targeting RCOR1 (sg1: GCCAGAATAAAAGTTTGGCGG, sg2: GACTTCGACCCCGGTGAGTAG, and sg3: GCGGAGAGGGAGGAACAACG) were designed and cloned into a modified lentiCRISPR v2 vector (Addgene plasmid #52961) in which Cas9 was replaced with GFP to monitor transduction. Lentivirus particles were produced as described before.20 CD34+ cells were transduced 24 hours after thawing and electroporated 72 hours after transduction with Cas9 messenger RNA (mRNA) using an ECM 830 Electroporation System (Harvard Apparatus). For LSD1 targeting, Cas9/sgRNA ribonucleoproteins, Cas9 protein (PNA Bio) in storage buffer, and synthetic sgRNA (Synthego #GKO_HS2_00000, KDM1A) in storage buffer (molar ratio 1:2.5) were incubated for 10 minutes at room temperature before electroporation.
Statistics
A 2-tailed unpaired Student t test was used to analyze groups of 2. Multiple groups were analyzed with 1-way analysis of variance. Results having a P value ≤ .05 were considered significant.
Results
LSD1 inhibition promotes expansion of human HSPCs in vitro
Studies in mice have shown that knockout of Lsd1 impairs HSCs in vivo,12 whereas Lsd1 knockdown triggers an expansion of HSPCs in the BM.13 To assess the consequences of loss of LSD1 in human HSPCs, we first targeted LSD1 in CD34+ cells by CRISPR and found that this led to severe impairment of proliferation, consistent with the mouse-knockout studies (supplemental Figure 1A; available on the Blood Web site). Next, to assess whether partial abrogation of LSD1 affects ex vivo expansion of human HSPCs, we treated UCB CD34+ cells with 3 LSD1 inhibitors (2-PCPA, GSK-LSD1, and RN-1) in a dose-titration experiment, and assessed the numbers of CD34+ cells following 6 days of culture (Figure 1A). Remarkably, all 3 compounds showed significantly increased numbers of CD34+ cells (Figure 1B), as well as of more immature CD34+CD90+ cells (supplemental Figure 1B), compared with DMSO-treated controls. Because recent studies by us and other investigators21,22 have shown that endothelial protein C receptor (EPCR) is a reliable cell surface marker to track functional HSCs during in vitro culture, we further evaluated the expansion of CD34+EPCR+ cells and observed a profound increase in this population, in particular by 2-PCPA and GSK-LSD1 (Figure 1C-D). Global transcriptome profiling of the CD34+EPCR+ and CD34+EPCR− cell fractions from 2-PCPA–treated cultures, and GSEA, confirmed a strong enrichment of HSC-associated gene sets23 in the CD34+EPCR+ cells (supplemental Figure 1C). Thus, LSD1 inhibition promotes the in vitro propagation of phenotypically defined HSCs. Further, using CFSE labeling, we found that 2-PCPA treatment did not significantly alter the cell division rate of the CD34+ cells, suggesting that their enhanced propagation is due to reduced differentiation and a preserved immature state, rather than increased proliferation (supplemental Figure 1D).
Figure 1.
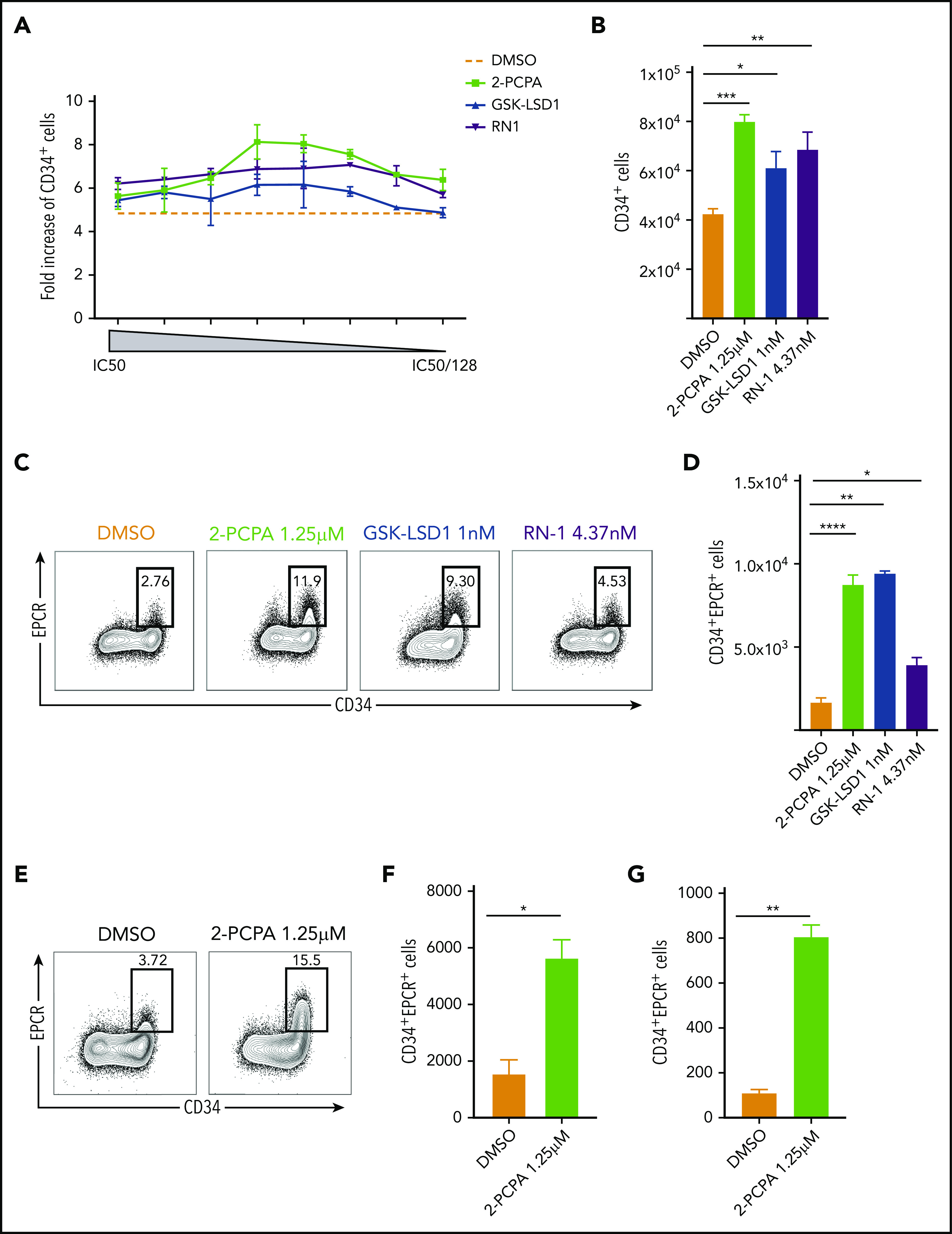
LSD1 inhibition promotes expansion of human HSPCs in vitro. (A) Dose-titration experiment showing the effect of 3 LSD1 inhibitors (2-PCPA, GSK-LSD1, or RN-1) on UCB-derived CD34+ HSPC expansion rate compared with the starting cell number (data from 2 repeats). (B) Total numbers of CD34+ cells obtained after treating 10 000 fresh CD34+ cells with 2-PCPA, GSK-LSD1, or RN-1 at their respective half-maximal inhibitory concentration (IC50)/16 concentration (2-PCPA-1.25 μM; GSK_LSD1-1 nM, RN1-4.37 nM) for 6 days. (C) FACS plots showing the frequency (C) and graph showing the total numbers (D) of CD34+EPCR+ cells obtained after treating 10 000 fresh CD34+ cells with 2-PCPA, GSK-LSD1 or RN-1, at their respective IC50/16 concentration, for 6 days. Data from 3 replicates from 1 of 5 independent experiments with similar results are shown. Frequency (E) and numbers (F) of CD34+EPCR+ cells obtained after treating UCB CD34+CD38−CD45RA−CD90+ cells with 2-PCPA for 6 days. (G) Total numbers of CD34+EPCR+ cells obtained after treating BM-derived CD34+CD38−CD45RA−CD90+ cells with 2-PCPA for 6 days. Data from 3 replicates from 1 of 2 independent experiments with similar results. *P ≤ .05, **P ≤ .01, ***P ≤ .001, ****P ≤ .0001. ns, not significant.
The UCB-derived CD34+ cell population is heterogeneous, and the enhanced expansion of CD34+EPCR+ cells through LSD1 inhibition could be due to direct or indirect mechanisms, such as altering paracrine signaling from more mature cells. To assess direct effects of LSD1 inhibition on UCB HSCs, we treated highly HSC-enriched CD34+CD38−CD45RA−CD90+ cells with 2-PCPA for 6 days. 2-PCPA treatment significantly increased the frequency and numbers of CD34+EPCR+ cells, suggesting that CD34+EPCR+ expansion mediated by LSD1 inhibition is a direct effect on the most immature HSPCs (Figure 1E-F). Moreover, we observed a similar effect on HSCs purified from adult human BM, indicating that LSD1 inhibition promotes ex vivo expansion of human HSPCs from different sources and stages of development (Figure 1G).
LSD1 inhibition enhances the ex vivo propagation of long-term repopulating HSCs
The pyrimidoindole derivative UM171 has been reported to efficiently expand transplantable human HSCs and is currently being tested in clinical trials.9 Specifically, it was shown to increase the CD34+EPCR+ population21; therefore, we decided to use UM171 as a reference and positive control in subsequent experiments. Indeed, we also found a strong expansion of CD34+EPCR+ cells by UM171 that was even more pronounced than that caused by 2-PCPA (supplemental Figure 2A-B).
To evaluate the functionality of the expanded HSPCs, we cultured UCB CD34+ cells for 6 days in the presence of 2-PCPA, UM171, or DMSO and measured the colony-forming capacity of the expanded cells. Although the total colony numbers were not significantly different, we observed multilineage colony forming unit–granulocyte erythrocyte monocyte megakaryocyte exclusively in 2-PCPA– and UM171-treated cultures (Figure 2A), indicating that the more immature progenitor populations are selectively supported under these conditions.
Figure 2.
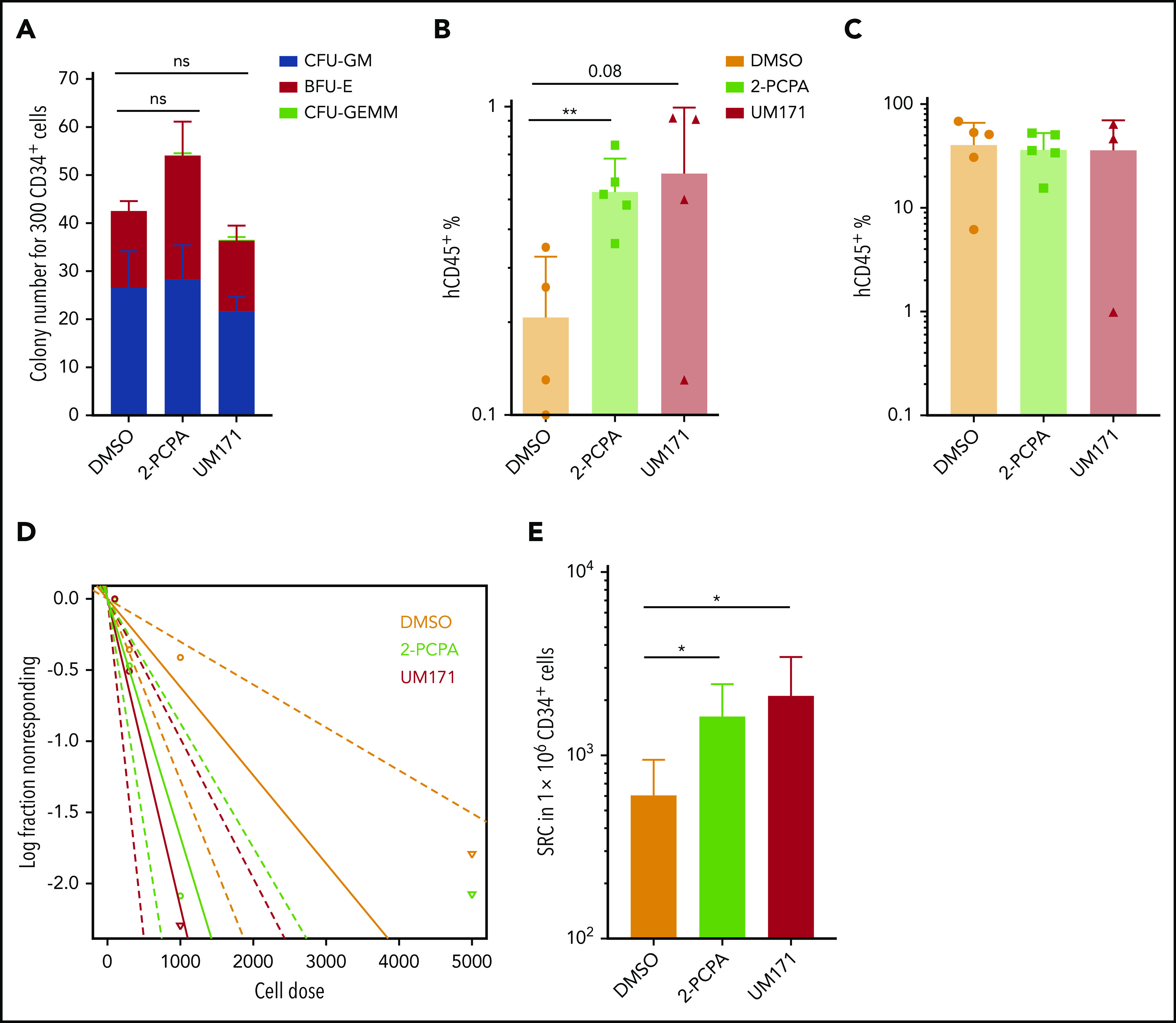
LSD1 inhibition enhances the ex vivo propagation of long-term repopulating HSCs. (A) Colony-forming capacity of CD34+ cells expanded with 2-PCPA or UM171 for 6 days. Accumulated data from 2 independent experiments with 3 replicates each. hCD45 engraftment in peripheral blood at week 5 (B) and hCD45 engraftment in BM at week 16 (C) in NSG mice transplanted with the progeny of 20 000 plated UCB CD34+ cells expanded with DMSO, 2-PCPA, or UM171 for 6 days. Five mice per each group was used as starting number. (D) Plot summarizing data obtained from LDA analysis. (E) Total number of SRCs obtained from LDA of cultures treated with DMSO, 2-PCPA, or UM171. Numbers are normalized to 1 million CD34+ cells before culture. *P ≤ .05, **P ≤ .01, generalized linear models.
Next, to evaluate the in vivo repopulating activity of 2-PCPA–expanded cultures, we performed xenotransplantation experiments using NOD.Cg-PrkdcscidIl2rgtm1Wjl/SzJ (NSG) mice. A total of 20 000 UCB CD34+ cells was plated and cultured for 6 days in the presence of 2-PCPA, UM171, or DMSO before being transplanted in bulk into sublethally irradiated (3 Gy) NSG mice. Five weeks after transplantation, 2-PCPA– and UM171-treated cells showed a nearly threefold higher chimerism in peripheral blood compared with the DMSO control, as well as a higher myeloid human (h)CD33+ cell frequency, indicating a faster myeloid recovery (Figure 2B; supplemental Figure 2C). However, the long-term engraftment levels measured 16 weeks after transplantation were very high for all conditions and were not significantly different among UM171, 2-PCPA, and DMSO (Figure 2C). Because human engraftment levels may have been saturated using the bulk transplantation model, we performed LDA to more accurately evaluate the changes in the number of SCID-repopulating cells (SRCs) following 2-PCPA and UM171 treatment. For LDA, CD34+ cells were cultured with 2-PCPA, UM171, or DMSO for 6 days, and the equivalent progeny of 100, 300, 1000, or 5000 plated cells were transplanted to NSG mice. Based on the analysis of human engraftment at 18 weeks posttransplantation, cells treated with 2-PCPA and UM171 showed a significant increase (2.7- and 3.5-fold, respectively) in SRCs compared with the DMSO control (Figure 2D-E; supplemental Table 1). We confirmed the supportive effects of LSD1 inhibition on long-term engrafting HSCs using a longer culture period (2 weeks) and an independent LSD1 inhibitor (GSK-LSD1) (supplemental Figure 2D-E).
Taken together, our data suggest that LSD1 inhibition, similar to UM171, supports the propagation of phenotypic and functional HSCs in culture by preserving their undifferentiated state.
LSD1 inhibition and UM171 trigger similar molecular responses
To explore the molecular consequences of LSD1 inhibition in ex vivo–cultured HSCs, we assayed the transcriptome of CD34+CD38−CD45RA−CD90+ cells following 24 hours of exposure to 2-PCPA. Using GSEA analysis, we found that HSC-specific gene sets24 were significantly enriched upon 2-PCPA treatment compared with DMSO control (Figure 3A). Additionally, genes specific for lineage-committed progenitors25 (erythroid, megakaryocytic, myeloid, and lymphoid) were all downregulated upon LSD1 inhibition (supplemental Figure 3A). This suggests that 2-PCPA induces immediate transcriptional changes that restrict differentiation and preserves the HSC state.
Figure 3.
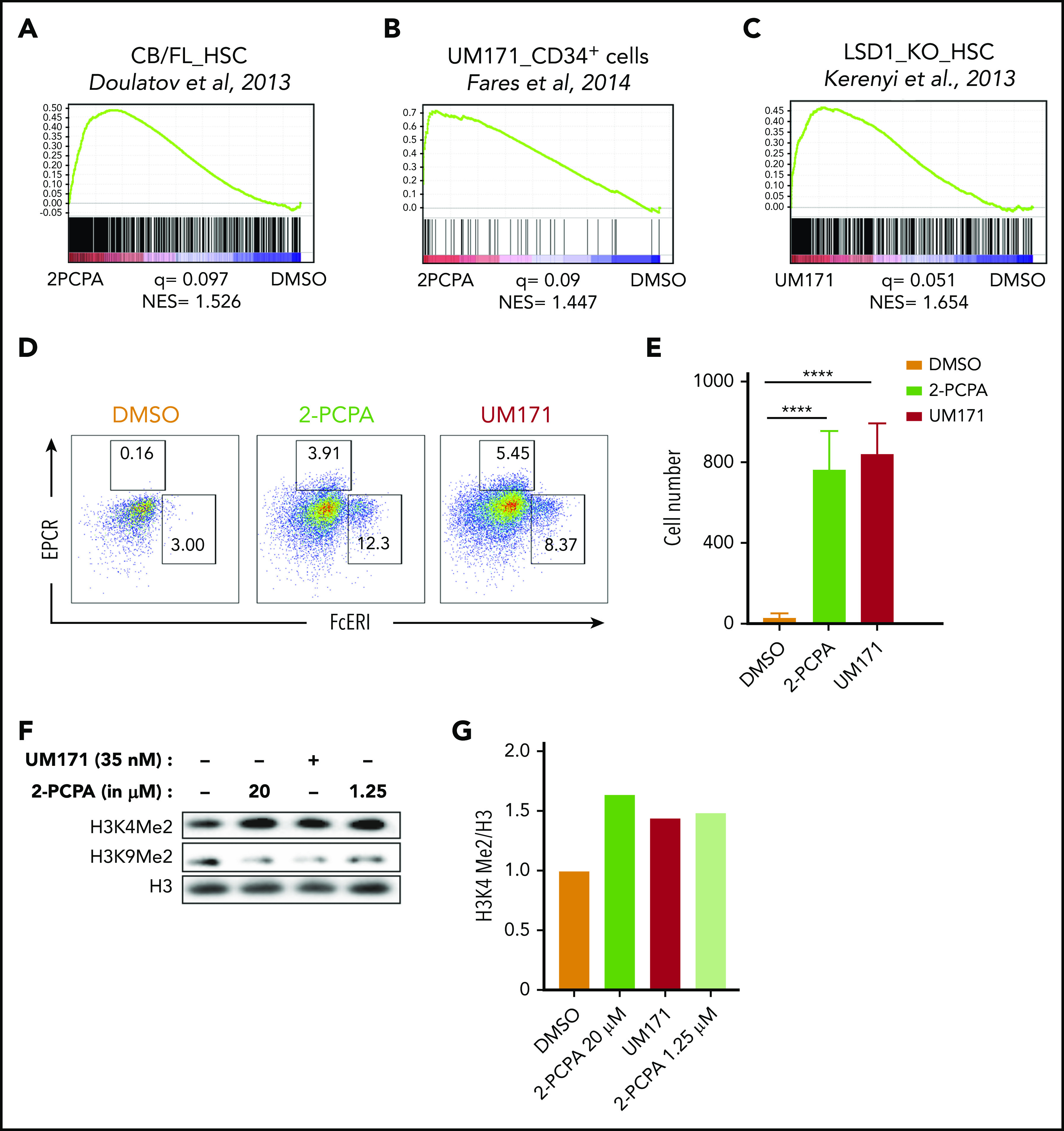
LSD1 inhibition and UM171 trigger similar molecular responses. GSEA plots showing HSC gene signature (A) and UM171-induced gene signature (B) in UCB HSC fraction treated with 2-PCPA for 24 hours. (C) LSD1-knockout (KO) gene signature in UCB HSC fraction treated with UM171 for 6 days. Microarray was performed in triplicates, and normalized enrichment score (NES) and q value were calculated with GSEA using reference gene sets from the indicated publications.8,12,24 FACS plots showing the frequency (D) and graphs showing the total numbers (E) of mast cell progenitors in cultures treated with 2-PCPA or UM171. Data from 4 replicates from 1 of 2 independent experiments with similar results. (F) Western blot showing H3K4me2 and H3K9me2 marks after 2-PCPA or UM171 treatment. (G) Relative increase in H3K4me2 compared with H3. ****P ≤ .0001.
Next, we compared the gene expression data for 2-PCPA treatment with gene signatures induced by early (12-72 hours) exposure to UM1718 and found a significant correlation (Figure 3B). Strikingly, there was also a strong correlation between gene expression signatures from UM171-cultured cells and those induced by LSD1 loss in mouse HSCs12 (Figure 3C). A distinct signature of UM171 treatment is the induction of mast cell–specific genes,26 and we found that both UM171 and 2-PCPA treatment induced a pronounced population of CD34+FcERI+ mast cell progenitors over 14 days of culture, suggesting overlapping functions of the 2 compounds (Figure 3D-E). In addition, given the known histone demethylase activity of LSD1, we assessed histone methyl marks to further compare the molecular features of 2-PCPA and UM171. As expected,27 we saw increased H3K4me2 marks upon LSD1 inhibition in CD34+ cells, but remarkably, also from UM171 treatment, further indicating overlapping molecular functions (Figure 3F-G; supplemental Figure 3B). Preserved methylation marks of H3K4 have been associated with genes expressed in more undifferentiated HSPCs.12 We did not observe any increase in H3K9me2 marks for 2-PCPA or UM171 (Figure 3F).
Taken together, LSD1 inhibition triggers distinct phenotypic and molecular features in HSPCs that are highly similar to UM171 treatment. This indicates a common mechanism and raises the question of whether UM171 may act on HSPCs through modulation of LSD1 activity.
LSD1 and the CoREST complex are targeted by UM171
Having seen similar and overlapping functions of 2-PCPA and UM171, we next asked whether LSD1 may be targeted by UM171. First, to assess whether UM171 is a direct inhibitor of LSD1 enzymatic activity, we performed an in vitro LSD1 inhibitor assay in which LSD1 activity is quantitatively coupled to a fluorescence readout. However, in contrast to 2-PCPA, UM171 did not inhibit LSD1 activity, even at increasing concentrations, arguing against a role in modifying the enzymatic function of LSD1 (Figure 4A). Recent studies have shown that LSD1 coordinates and binds to multiple epigenetic chromatin-remodeling complexes,28 so we next asked whether UM171 may selectively perturb any of these LSD1-containing protein complexes. To identify direct or immediate indirect targets of UM171, we performed TPP, which measures, at a proteome-wide level by quantitative proteomics and curve fitting, melting temperature differences of target proteins caused by stabilization or destabilization upon candidate drug binding.29 Because the assay requires a large number of cells, we used AML cell lines (HL-60 and Kasumi-1) instead of UCB CD34+ cells. By setting a melting temperature difference cutoff of ±1°C and a P value threshold of .05, TPP identified 130 and 44 candidates in HL-60 cells and Kasumi-1 cells, respectively (supplemental Table 2). Subsequent analysis identified 5 candidates who had significant alterations in both cell lines (Figure 4B), 1 of which was RCOR1, a core member of the LSD1-containing CoREST complex. To assess potential effects of UM171 treatment on this complex, we treated UCB CD34+ cells with UM171 for 24 hours and performed western blot by probing with anti-RCOR1 antibody. Remarkably, UM171 treatment almost completely abolished RCOR1 protein levels compared with DMSO and 2-PCPA treatment (Figure 4C). Further analysis of several other protein components of the CoREST complex (LSD1, HDAC1, HDAC2) revealed markedly reduced levels of LSD1 and HDAC2 upon UM171 treatment, whereas HDAC1 levels were unaltered (Figure 4D). Importantly, none of the proteins were perturbed at the mRNA level, suggesting a posttranslational mechanism for targeting of the protein complex by UM171 (supplemental Figure 4A). RCOR1 is exclusively associated with CoREST, whereas the other components, including LSD1, are part of other complexes as well, which may explain why RCOR1 was almost completely lost, and the levels of the other proteins were more variable. To further elucidate whether CoREST may be directly or indirectly targeted by UM171, we performed a time-course analysis in 3 AML cell lines and found clearly reduced levels of RCOR1 and LSD1 after only 3 hours of exposure to UM171 (Figure 4E). This time frame is consistent with the direct targeting of proteins for degradation, as shown previously.17 Next, we enriched the residual LSD1 present in UM171-treated HL-60 cells by co-IP and found that UM171 preferentially degrades CoREST-bound LSD1 compared with the fraction of LSD1 not bound to CoREST (supplemental Figure 4B-C). This suggests that UM171 perturbs LSD1 by selectively degrading the CoREST complex.
Figure 4.
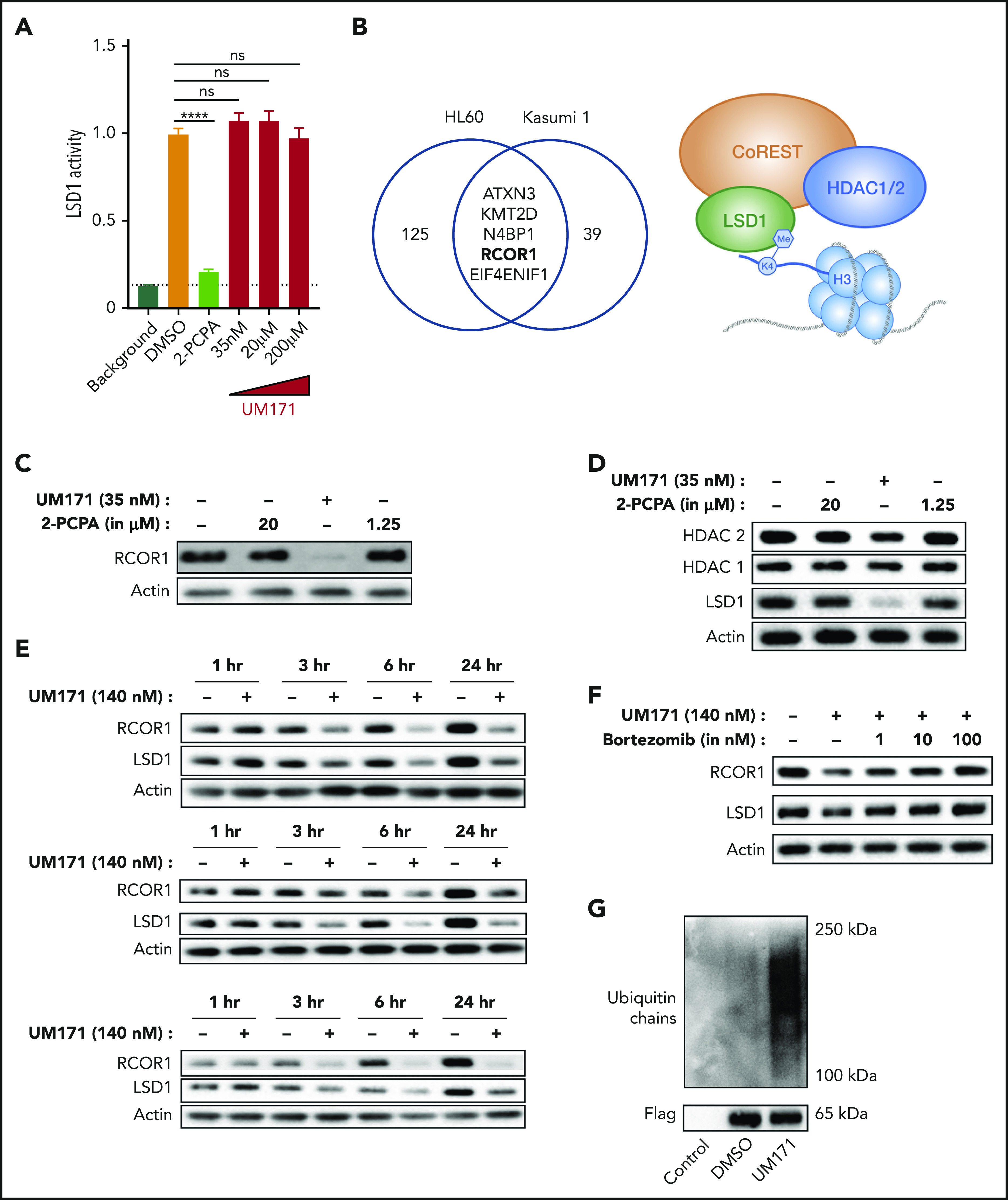
LSD1 and the CoREST complex are targeted by UM171. (A) In vitro assay measuring LSD1 enzymatic activity in the presence of 2-PCPA or UM171. Data from 3 to 6 repeats of the assay are represented. (B) Venn diagram showing common target proteins of UM171 between HL-60 and Kasumi-1 cells, identified by TPP. Western blots showing RCOR1 (C) and LSD1, HDAC1, and HDAC2 (D) in UCB CD34+ cells treated with 2-PCPA or UM171 for 24 hours. (E) Time kinetics experiment showing RCOR1 and LSD1 protein expression in OCI-AML3 (top panel), HL-60 (middle panel), and U937 (bottom panel) cell lines treated with DMSO or UM171. (F) RCOR1 and LSD1 protein expression in HL-60 cell line treated with UM171 and proteasomal inhibitor bortezomib for 3 hours. (G) Flag-tagged RCOR1-expressing HL-60 cells were treated with DMSO or UM171 for 1.5 hours, and RCOR1 was enriched with Flag IP and analyzed for polyubiquitination. ****P ≤ .0001.
Further, addition of the proteasome inhibitor bortezomib along with UM171 prevented reductions in RCOR1 and LSD1 proteins in a dose-dependent manner (Figure 4F), suggesting involvement of the ubiquitin proteasomal system. To establish involvement of the ubiquitin proteasomal system in the degradation of the CoREST complex, we performed polyubiquitination analysis of LSD1 and Flag-tagged RCOR1 post-UM171 treatment. With IP and western blotting, we found that RCOR1 and LSD1 were rapidly polyubiquitinated upon UM171 treatment (Figure 4G; supplemental Figure 4D-E). Taken together, these findings clearly suggest that the LSD1-containing CoREST protein complex is targeted and degraded by UM171, which provide a possible mechanistic basis for the HSC stimulatory effects mediated by UM171.
Finally, to directly assess whether abrogation of the CoREST complex can mediate HSPC expansion, we knocked out the core member, RCOR1, in CD34+ cells using CRISPR/Cas9. We used 3 separate sgRNAs targeting RCOR1 and delivered these by lentiviral transduction, whereas Cas9 mRNA was introduced through electroporation (Figure 5A). All 3 guides showed efficient knockout of RCOR1 (Figure 5B) and enhanced the numbers of CD34+ (threefold to fivefold) and CD34+EPCR+ (fivefold to 10-fold) cell populations over a course of 10 days compared with a nontargeting guide control (Figure 5C-E). Because of the more extensive manipulation of these cells by transduction and electroporation, the overall frequency and numbers of cells were lower compared with our other settings. We next ablated RCOR1 with CRISPR and assessed expansion in the presence or absence of UM171. Because GFP was coexpressed from the sgRNA vector, RCOR1-edited cells were labeled with GFP and could be compared with nonedited GFP− cells within the same cultures. We found that GFP+ RCOR1-deficient cells had a relative expansion advantage over GFP− cells that was completely cancelled out by UM171 (Figure 5F), suggesting a common mechanism for HSPC expansion for UM171 treatment and RCOR1 loss. Cells transduced with nontargeting guide did not show any expansion advantage and were not differentially affected by UM171 compared with their GFP− nontransduced counterparts (Figure 5F). Altogether, this shows that the functional loss of CoREST indeed induces HSPC expansion and constitutes a principal mechanistic component of UM171-mediated effects.
Figure 5.
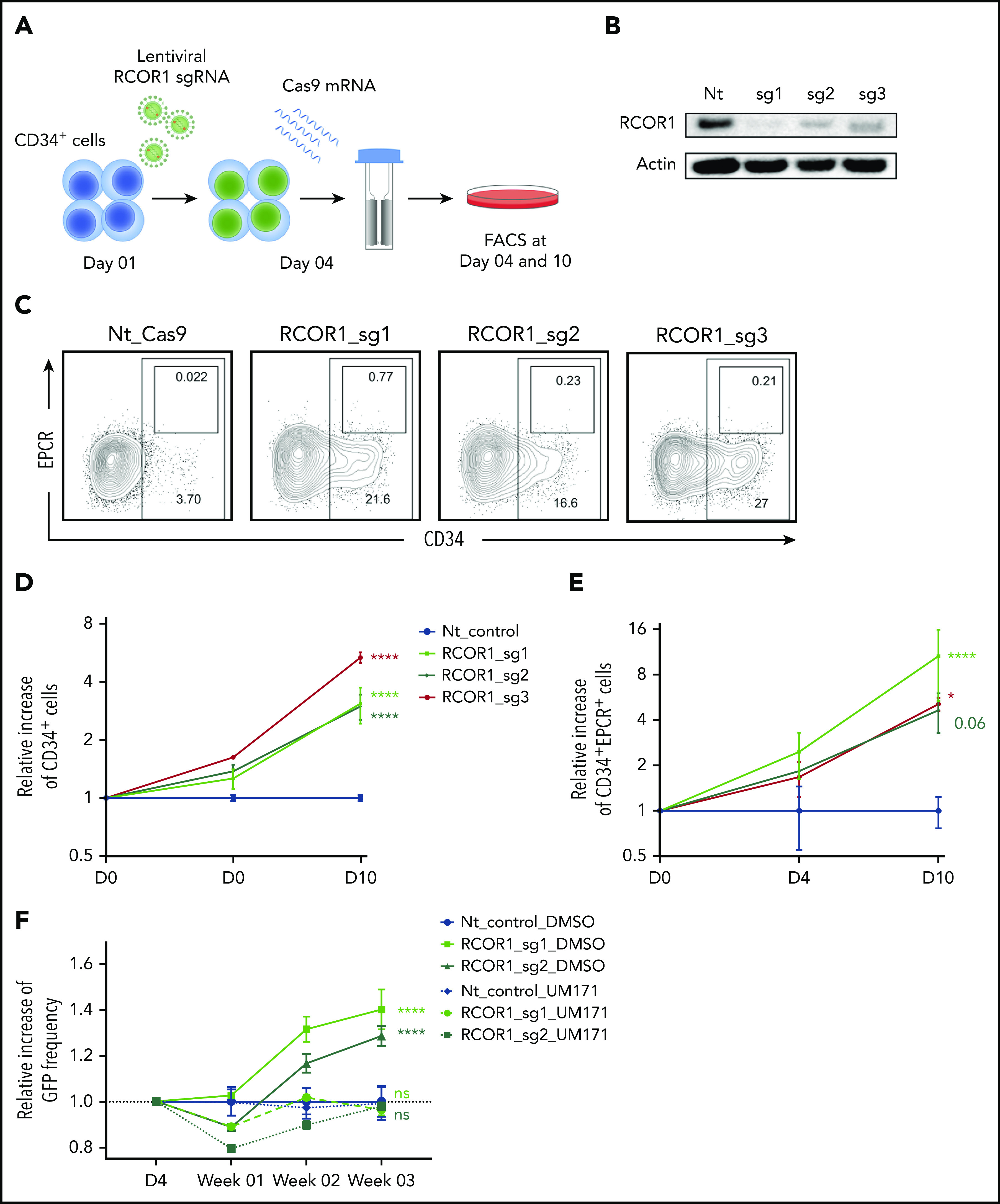
Loss of CoREST (RCOR1) induces HSPC expansion. (A) Schematic representation of the experimental strategy to knockout RCOR1. (B) RCOR1 protein levels in UCB CD34+ cells edited with nontargeting (Nt) control or RCOR1 guides (day 10). (C) FACS plots showing the frequency of CD34+ and EPCR+ populations in Nt control and RCOR1-knockout UCB HSPCs (day 10). The relative number of CD34+ cells (D) and CD34+EPCR+ cells (E) in RCOR1-knockout HSPCs compared with the Nt control–treated cells over time. Total cell numbers for each condition and time point were calculated and normalized to the mean of the control Nt sgRNA. Data from 3 replicates from 1 of 2 independent experiments with similar results. (F) Relative change in frequency of GFP+ cells over time in Nt control and RCOR1-edited samples compared with day 4 (D4) in DMSO- or UM171-treated cultures. Values are normalized to the NT DMSO control (n = 3). *P ≤ .05, ****P ≤ .0001.
Discussion
In this study, we report the identification of LSD1 as a modulator of human HSC propagation ex vivo and provide a mechanistic basis for the expansion phenotype of the pyrimidoindole compound UM171. Based on our findings, we propose that UM171-mediated HSC expansion involves targeting and depletion of the LSD1-containing CoREST complex.
In vitro culture conditions supplemented with cytokines tend to promote proliferation and differentiation of HSCs rather than expansion.30,31 Because the presence of cytokines is also essential for cell survival, it is important to identify cues that may counteract differentiation and push cell fate toward maintenance and renewal of the more immature state instead. External cues, along with transcription factors, influence chromatin states via epigenetic modifications, and the resulting transcriptional programs are major factors in regulating HSC maintenance and fate decisions.32 Epigenetic modifications are primarily introduced and altered through posttranslational modifications of DNA and histones. LSD1, when present in the CoREST complex, triggers histone modifications by executing the removal of histone mono- and dimethyl groups at H3K4 residues.10 Previous studies in murine models suggest that Lsd1 and Rcor1 are essential in silencing HSC genes during differentiation by demethylation of H3K4 residues.12,33 Therefore, loss of LSD1 and preservation of the methylation marks on H3K4, as we observed here, may maintain a global epigenetic landscape of more undifferentiated cells and keep HSC genes actively transcribed.12 In the context of human HSC expansion, it has also been shown that inhibition of pan-histone deacetylation using HDAC inhibitors, such as valproic acid, prevents differentiation and expands human HSCs in vitro; however, in addition, that results in aberrant activation of nonhematopoietic pluripotency genes like OCT4, SOX2, and NANOG.5 We did not observe such activation of pluripotency genes or other nonhematopoietic genes for LSD1 inhibition, indicating that HDAC inhibitors may induce more profound alterations of cell fate.
The discovery of UM171 as an HSC-promoting molecule was reported in 2014, and a >10-fold expansion of long-term repopulating HSCs was demonstrated.8 Despite an unresolved mechanism of action, the preclinical testing and subsequent establishment of clinical trials for UM171 have progressed rapidly since its initial discovery. The first reports from clinical trials using UCB cells expanded with UM171 for transplantation of leukemia patients show promising results.9 Although the expansion levels seen in this study for UM171 were more modest than originally reported,8 our culture time was significantly shorter (6 vs 12 days), and we did not use the HSC-promoting Fed-Batch culture system.34 We saw similar, yet slightly higher, SRC numbers for UM171 compared with 2-PCPA, which may indicate that targeting CoREST by UM171 is preferred over specifically abrogating LSD1 activity by 2-PCPA or other inhibitors. It is possible that loss of other functions of the CoREST complex, such as HDAC activity, could have HSC-promoting effects, in addition to the loss of LSD1-mediated demethylase activity. Alternatively, UM171 may have additional targets with HSC-promoting effects. For example, it was recently proposed that UM171 may stimulate HSC expansion by balancing pro- and anti-inflammatory signals.26
Perhaps the most striking finding from our study was the extremely rapid and efficient degradation of members of the CoREST complex by UM171. It is highly unusual to observe such selective degradation of specific proteins by small molecules. Our work clearly defines CoREST as a target of UM171, but it remains to be elucidated exactly how polyubiquitination and degradation are triggered, including the primary binding target of UM171. RCOR1 would be a strong candidate because it is a unique core member of the complex and was the most efficiently degraded component that we measured. LSD1, which unlike RCOR1, is also part of other complexes, was not completely degraded, and the more generic components HDAC1 and HDAC2 showed even less degradation. The precise mapping of a primary binding target and means of degradation of CoREST by UM171 may reveal principles that can be further generalized and exploited for purposes other than HSC expansion.
Supplementary Material
The online version of this article contains a data supplement.
Acknowledgments
The authors thank the staff at BMC FACS core and animal house facilities for excellent support; Karina Vidovic and Martin L. Olsson for help with western blotting, and Veronika Bendoriūtė for help with illustrations.
This work was supported by grants from the Swedish Research Council, the Swedish Cancer Foundation, the Swedish Pediatric Cancer Foundation, Knut och Alice Wallenbergs Stiftelse, and the European Research Council under the European Union’s Horizon 2020 research and innovation program (grant agreement no. 648894) (J.L.). It was also supported by the HematoLinné and StemTherapy programs at Lund University. A.S. was supported by the Swedish Cancer Foundation and the Lady Tata Memorial Trust.
Footnotes
Microarray data have been submitted to the Gene Expression Omnibus database (accession number GSE132747).
Data sharing requests should be sent to Jonas Larsson (jonas.larsson@med.lu.se).
The publication costs of this article were defrayed in part by page charge payment. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 USC section 1734.
Authorship
Contribution: A.S. and J.L. conceived the study, designed the experiments, analyzed the results, and wrote the manuscript; A.S., K.Ž., M.S.T., S.D., J.C., and M.V.J., performed transplantation experiments; D.Y. and A.B. performed CRISPR experiments; A.S. and K.Ž. performed western blot experiments; R.G. helped with GSEA analysis and editing of the manuscript; and M.G. and R.A.Z. performed TPP experiments and the corresponding data analysis.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Jonas Larsson, Molecular Medicine and Gene Therapy, Lund University, BMC A12, 221 84 Lund, Sweden; e-mail: jonas.larsson@med.lu.se.
REFERENCES
- 1.Laughlin MJ, Barker J, Bambach B, et al. Hematopoietic engraftment and survival in adult recipients of umbilical-cord blood from unrelated donors. N Engl J Med. 2001;344(24):1815-1822. [DOI] [PubMed] [Google Scholar]
- 2.Lund TC, Boitano AE, Delaney CS, Shpall EJ, Wagner JE. Advances in umbilical cord blood manipulation-from niche to bedside. Nat Rev Clin Oncol. 2015;12(3):163-174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Boitano AE, Wang J, Romeo R, et al. Aryl hydrocarbon receptor antagonists promote the expansion of human hematopoietic stem cells. Science. 2010;329(5997):1345-1348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Baudet A, Karlsson C, Safaee Talkhoncheh M, Galeev R, Magnusson M, Larsson J. RNAi screen identifies MAPK14 as a druggable suppressor of human hematopoietic stem cell expansion. Blood. 2012;119(26):6255-6258. [DOI] [PubMed] [Google Scholar]
- 5.Chaurasia P, Gajzer DC, Schaniel C, D’Souza S, Hoffman R. Epigenetic reprogramming induces the expansion of cord blood stem cells. J Clin Invest. 2014;124(6):2378-2395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Himburg HA, Muramoto GG, Daher P, et al. Pleiotrophin regulates the expansion and regeneration of hematopoietic stem cells. Nat Med. 2010;16(4):475-482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.North TE, Goessling W, Walkley CR, et al. Prostaglandin E2 regulates vertebrate haematopoietic stem cell homeostasis. Nature. 2007;447(7147):1007-1011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Fares I, Chagraoui J, Gareau Y, et al. Cord blood expansion. Pyrimidoindole derivatives are agonists of human hematopoietic stem cell self-renewal. Science. 2014;345(6203):1509-1512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cohen SRJ, Lachance S, Marinier A, et al. Single UM171 expanded cord blood transplant is feasible, safe, and permits transplantation of better HLA matched cords with very low transplant related mortality. Biol Blood Marrow Transplant. 2018;24(3):S190-S191. [Google Scholar]
- 10.Lee MG, Wynder C, Cooch N, Shiekhattar R. An essential role for CoREST in nucleosomal histone 3 lysine 4 demethylation. Nature. 2005;437(7057):432-435. [DOI] [PubMed] [Google Scholar]
- 11.Metzger E, Wissmann M, Yin N, et al. LSD1 demethylates repressive histone marks to promote androgen-receptor-dependent transcription. Nature. 2005;437(7057):436-439. [DOI] [PubMed] [Google Scholar]
- 12.Kerenyi MA, Shao Z, Hsu YJ, et al. Histone demethylase Lsd1 represses hematopoietic stem and progenitor cell signatures during blood cell maturation. eLife. 2013;2:e00633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Sprüssel A, Schulte JH, Weber S, et al. Lysine-specific demethylase 1 restricts hematopoietic progenitor proliferation and is essential for terminal differentiation. Leukemia. 2012;26(9):2039-2051. [DOI] [PubMed] [Google Scholar]
- 14.Hu Y, Smyth GK. ELDA: extreme limiting dilution analysis for comparing depleted and enriched populations in stem cell and other assays. J Immunol Methods. 2009;347(1-2):70-78. [DOI] [PubMed] [Google Scholar]
- 15.Irizarry RA, Hobbs B, Collin F, et al. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics. 2003;4(2):249-264. [DOI] [PubMed] [Google Scholar]
- 16.Subramanian A, Tamayo P, Mootha VK, et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci USA. 2005;102(43):15545-15550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Krönke J, Udeshi ND, Narla A, et al. Lenalidomide causes selective degradation of IKZF1 and IKZF3 in multiple myeloma cells. Science. 2014;343(6168):301-305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Franken H, Mathieson T, Childs D, et al. Thermal proteome profiling for unbiased identification of direct and indirect drug targets using multiplexed quantitative mass spectrometry. Nat Protoc. 2015;10(10):1567-1593. [DOI] [PubMed] [Google Scholar]
- 19.Cox J, Mann M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat Biotechnol. 2008;26(12):1367-1372. [DOI] [PubMed] [Google Scholar]
- 20.Galeev R, Karlsson C, Baudet A, Larsson J. Forward RNAi screens in human hematopoietic stem cells. Methods Mol Biol. 2017;1622:29-50. [DOI] [PubMed] [Google Scholar]
- 21.Fares I, Chagraoui J, Lehnertz B, et al. EPCR expression marks UM171-expanded CD34+ cord blood stem cells. Blood. 2017;129(25):3344-3351. [DOI] [PubMed] [Google Scholar]
- 22.Subramaniam A, Talkhoncheh MS, Magnusson M, Larsson J. Endothelial protein C receptor (EPCR) expression marks human fetal liver hematopoietic stem cells. Haematologica. 2019;104(2):e47-e50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Prashad SL, Calvanese V, Yao CY, et al. GPI-80 defines self-renewal ability in hematopoietic stem cells during human development. Cell Stem Cell. 2015;16(1):80-87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Doulatov S, Vo LT, Chou SS, et al. Induction of multipotential hematopoietic progenitors from human pluripotent stem cells via respecification of lineage-restricted precursors. Cell Stem Cell. 2013;13(4):459-470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Laurenti E, Doulatov S, Zandi S, et al. The transcriptional architecture of early human hematopoiesis identifies multilevel control of lymphoid commitment. Nat Immunol. 2013;14(7):756-763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Chagraoui J, Lehnertz B, Girard S, et al. UM171 induces a homeostatic inflammatory-detoxification response supporting human HSC self-renewal. PLoS One. 2019;14(11):e0224900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lee MG, Wynder C, Schmidt DM, McCafferty DG, Shiekhattar R. Histone H3 lysine 4 demethylation is a target of nonselective antidepressive medications. Chem Biol. 2006;13(6):563-567. [DOI] [PubMed] [Google Scholar]
- 28.Rudolph T, Beuch S, Reuter G. Lysine-specific histone demethylase LSD1 and the dynamic control of chromatin. Biol Chem. 2013;394(8):1019-1028. [DOI] [PubMed] [Google Scholar]
- 29.Savitski MM, Reinhard FB, Franken H, et al. Tracking cancer drugs in living cells by thermal profiling of the proteome. Science. 2014;346(6205):1255784. [DOI] [PubMed] [Google Scholar]
- 30.Srour EF, Tong X, Sung KW, et al. Modulation of in vitro proliferation kinetics and primitive hematopoietic potential of individual human CD34+CD38-/lo cells in G0. Blood. 2005;105(8):3109-3116. [DOI] [PubMed] [Google Scholar]
- 31.Norkin M, Lazarus HM, Wingard JR. Umbilical cord blood graft enhancement strategies: has the time come to move these into the clinic? Bone Marrow Transplant. 2013;48(7):884-889. [DOI] [PubMed] [Google Scholar]
- 32.Cui K, Zang C, Roh TY, et al. Chromatin signatures in multipotent human hematopoietic stem cells indicate the fate of bivalent genes during differentiation. Cell Stem Cell. 2009;4(1):80-93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Yao H, Goldman DC, Nechiporuk T, et al. Corepressor Rcor1 is essential for murine erythropoiesis. Blood. 2014;123(20):3175-3184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Csaszar E, Kirouac DC, Yu M, et al. Rapid expansion of human hematopoietic stem cells by automated control of inhibitory feedback signaling. Cell Stem Cell. 2012;10(2):218-229. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.