

Keywords: C-fibers, esophagus, nerve terminal, vagus nerve, voltage-gated sodium channels
Abstract
We investigated voltage-gated sodium channel (NaV1) subunits that regulate action potential initiation in the nerve terminals of vagal nodose C-fibers innervating the esophagus. Extracellular single fiber recordings were made from the nodose C-fibers, with mechanically sensitive nerve terminals in the isolated innervated guinea pig esophagus. NaV1 inhibitors were selectively delivered to the tissue-containing nerve terminals. Graded esophageal distention was used for mechanical stimulation. The NaV1.7 inhibitor PF-05089771 nearly abolished action potential initiation in response to low levels of esophageal distention but only partially inhibited the response to higher levels of esophageal distention. The PF-05089771-insensitive component of the response progressively increased (up to ≈50%) with increasing esophageal distention and was abolished by tetrodotoxin (TTX). In addition to NaV1.7, nodose C-fiber [transient receptor potential channel-vanilloid subfamily member 1 (TRPV1)-positive] neurons retrogradely labeled from the esophagus expressed mRNA for multiple TTX-sensitive NaV1s. The group NaV1.1, NaV1.2, and NaV1.3 inhibitor ICA-121431 inhibited but did not abolish the PF-05089771-insensitive component of the response to high level of esophageal distention. However, combination of ICA-121431 with compound 801, which also inhibits NaV1.7 and NaV1.6, nearly abolished the response to the high level of esophageal distention. Our data indicate that the action potential initiation in esophageal nodose C-fibers evoked by low (innocuous) levels of esophageal distention is mediated by NaV1.7. However, the response evoked by higher (noxious) levels of esophageal distention has a progressively increasing NaV1.7-independent component that involves multiple TTX-sensitive NaV1s. The stimulus intensity-dependent recruitment of NaV1s may offer novel opportunities for strategic targeting of NaV1 subunits for inhibition of nociceptive signaling in visceral C-fibers.
NEW & NOTEWORTHY We report that pharmacologically distinguishable voltage-gated sodium channels (NaV1) mediate action potential initiation at low (innocuous) versus high (noxious) intensity of esophageal distention in nerve terminals of vagal nodose C-fibers. Action potential initiation at low intensity is entirely dependent on NaV1.7; however, additional tetrodotoxin (TTX)-sensitive NaV1s are recruited at higher intensity of distention. This is the first demonstration that NaV1s underlying action potential initiation in visceral C-fibers depend on the intensity of the stimulus.
INTRODUCTION
Primary afferent nerve terminals in peripheral tissues are activated by physical and/or chemical stimuli. The activating stimulus directly or indirectly gates various ion channels that lead to a depolarization of the nerve terminal membrane, referred to as the generator potential. The generator potential leads to gating of voltage-gated sodium channels (NaV1s) and initiation of action potentials, which are then conducted to the central terminals, where they evoke the release of neurotransmitters at the central synapse. Therefore, NaV1 channels are the sine qua non of sensory neurotransmission.
The primary afferent nerves innervating internal organs (visceral afferent nerves) mediate reflex regulation and sensations, including pain from the viscera (8). In many diseases, inhibition of these nerves would help to alleviate symptoms resulting from altered motility and chronic pain. Indeed, voltage-gated sodium channels as potential drug targets have long been investigated in visceral afferent nerves (e.g., see Refs. 2, 12, and 22; reviewed in Ref. 6). Nonetheless, the information on NaV1s regulating action potential initiation in visceral afferent nerves is limited (7, 18).
In contrast to somatic skin mechanonociceptors, which respond to noxious high intensity but not to innocuous low-intensity mechanical stimuli, many visceral nerves implicated in nociception respond to and encode a wide range of mechanical stimuli from innocuous to noxious (reviewed in Ref. 11). For example, colonic afferent nerves are typically activated by low levels of colonic distention, and their activity continues to linearly increase as the distention continues to increase to noxious levels (e.g., see Refs. 4 and 13). Similar wide range is observed in other organs (3, 28, 33). Therefore, it is often thought that at lower levels of stimulation visceral afferent nerves initiate physiological reflex regulation, whereas at higher levels of stimulation these nerves initiate noxious reflexes and sensations such as discomfort and pain. Therefore, strategies that would preferentially inhibit the high levels of activation by noxious stimuli would be useful for treatment of pain. On the other hand, strategies that would preferentially inhibit low levels of activation would be useful for management of reflex dysregulation (altered motility).
We investigated action potential initiation in the nerve terminals of vagal nodose C-fibers innervating the esophagus. We used an ex vivo innervated esophagus preparation in which NaV1 inhibitors were delivered selectively to esophageal tissue containing the nerve terminals of vagal nodose C-fibers. Our data support the hypothesis that NaV1.7 is responsible for the action potential initiation at low physiological levels of esophageal distention while additional NaV1 subtypes are recruited at higher noxious levels of distention.
METHODS
Ethical approval.
The experiments were approved by the Johns Hopkins School of Medicine Animal Care and Use Committee. Male Hartley guinea pigs (Hilltop Laboratory Animals Inc., Scottsdale, PA) weighing 100–200 g were housed in an approved animal facility with controlled temperature and humidity, 24-h/day-night cycle, in groups in cages, with unrestricted access to food and water provided and species-appropriate environmental enrichment. The animals were euthanized by exposure to CO2 in rising concentration (CO2 was sourced from a compressed gas cylinder and administered at a flow rate measured by flowmeter, displacing ∼20–30% of euthanasia chamber volume/min conforming to experiments in Ref. 10) and exsanguination.
Extracellular recordings of single-fiber activity.
The ex vivo guinea pig esophagus preparation developed previously (5, 33) was modified to allow for selective delivery of drugs to esophageal tissue without exposing the vagus and recurrent laryngeal nerves to the drugs or for selective delivery of drugs to desheathed vagus nerve without exposing the esophageal tissue to the drugs (Fig. 1A). The esophagus with preserved bilateral extrinsic vagal innervation (including the nodose and jugular ganglia) was dissected. Esophageal mucosa was removed by partially circularly dissecting mucosa from the muscle at the oral end and then inserting a thread into the esophageal lumen, securing partially dissected mucosa to the thread, and then pulling the mucosa out through the inside of the esophagus. We and others have noted previously that in the guinea pig the mucosa can be easily separated from the muscle (33, 34). We have also reported that the drugs (e.g., capsaicin) delivered into the esophageal lumen are ineffective to activate esophageal C-fibers with the esophageal mucosa intact but are effective when esophageal mucosa is removed or disrupted (33). The preparation was placed in a two-compartment Sylgard-lined Perspex recording dish. The esophagus was mounted in the tissue compartment, and the rostral aspect of the vagus nerves, including the nodose ganglion, was pinned in the recording compartment. For experiments in which drugs were selectively delivered to the vagus nerve, a section of the vagus nerve was carefully desheathed, and the preparation was placed in the three-compartment recording dish (Fig. 1A). The esophagus was pinned in the tissue compartment, and the vagus nerve was pulled through the vagus compartment with the rostral end and nodose ganglion in the recording compartment (Fig. 1A). The openings between the compartments containing the vagus nerve were sealed with Vaseline. Each compartment was separately superfused (4–6 mL/min) with heated (35°C) Krebs bicarbonate solution (in mM: 118 NaCl, 5.4 KCl, 1 NaH2PO4, 1.2 MgSO4, 1.9 CaCl2, 25 NaHCO3, and 11 dextrose, gassed with 95% O2-5% CO2) containing indomethacin (3 μM). The esophagus was perfused through the lumen. Polyethylene tubing was inserted 3–5 mm into the cranial and caudal ends of the esophagus and secured for perfusion. Mechanical stimulation with isobaric esophageal distension was performed by the intraluminal pressure increase from resting condition (no intraluminal pressure applied, ≈0 mmHg) to desired pressure for 20 s, followed by the return to resting condition (≈0 mmHg). When multiple stimulations were applied successively (e.g., 10, 30, and 60 mmHg), there was a 2-min interval between the stimulations. The increase in isobaric pressure was generated by a gravity-driven fluid system. In separate experiments, the distensibility of the esophagus with and without mucosa was evaluated in a paired manner using the same approach (Fig. 1B, inset). The outer diameter was measured at the center of the lower, middle, and upper thirds of the esophagus by a water-resistant IP54 digital caliper (EAGems; http://edwardallengems.com/) and averaged in the esophagus with an intact mucosa; then, the mucosa was removed as described above, and the measurement was repeated.
Fig. 1.

Experimental setup for evaluation of the effects of voltage-gated sodium channel (NaV1) inhibitors on action potential initiation and conduction in esophageal nodose C-fibers. A: isolated esophagus with attached vagus nerve was placed in the custom-made dish with 3 separately superfused (37°C) compartments: the recording compartment containing vagal nodose and jugular ganglia, the vagal compartment containing the desheathed segment of the vagus nerve, and the tissue compartment containing the esophagus. To selectively deliver NaV1 inhibitors to esophageal tissue containing nodose C-fiber nerve terminals, esophageal mucosa was removed and NaV1 inhibitors were perfused through the esophageal lumen. To selectively deliver NaV1 inhibitors to the vagus nerve containing conducting axons of esophageal nodose C-fibers, the vagus nerve compartment was superfused with NaV1 inhibitor. B: comparison of the magnitude of mechanical response evoked by esophageal distention in the esophagus preparations with intact and removed esophageal mucosa. As expected, the distention with the same intraesophageal pressure evoked more activation in the more distensible esophageal preparations with removed mucosa as the removal of mucosa reduced tissue components opposing esophageal distention. Inset: comparison of distensibility of the esophagus with and without esophageal mucosa (esophageal outside diameter). C: solvent for NaV1 inhibitors DMSO in the highest concentration used in this study (0.2%, 30-min incubation) did not appreciably affect the response of nodose C-fibers to distention.
Extracellular recordings were made from vagal nodose neurons projecting distention-sensitive C-fibers into the esophagus in an isolated, perfused, vagally innervated guinea pig esophagus. An aluminosilicate glass microelectrode was filled with a 3 M sodium chloride solution (electrode resistance ≈2 MΩ). The electrode was placed in an electrode holder connected directly to the head stage (A-M Systems, Sequim, WA). A return silver-silver chloride wire electrode and earthed silver-silver chloride pellet were placed in the perfusion fluid of the recording compartment. The recorded signal was amplified (M1800; A-M Systems) and filtered (low cutoff: 0.3 kHz; high cutoff: 1 kHz), and the signal was displayed on an oscilloscope (Tektronix, Beaverton, OR) and fed through an A/D converter to an Apple computer. The software TheNerveOfIt (sampling frequency: 33 kHz; PHOCIS, Baltimore, MD) was used for data analysis. The recording electrode was randomly inserted into the nodose ganglion using a micromanipulator, and a distension-sensitive nerve fiber was identified if esophageal distension (to 60 mmHg for 5 s) evoked action potential discharge. One fiber per animal was recorded.
Experimental protocol.
First, the control response to esophageal distention was evaluated for reproducibility. Then NaV1 blocker was perfused through the esophageal limen for 30 min. The distention was then applied again in the presence of NaV1 blocker. Up to three different concentrations or combinations of NaV1 blockers were tested per experiment. The distention to 10–60 mmHg was used as indicated. Tetrodotoxin (0.3 µM or 1µM) and/or lidocaine (1 mM) was perfused through the esophageal lumen at the end of the experiment to confirm that the drugs applied into the lumen were able to diffuse to the investigated nerve fiber terminal in the esophageal wall. In experiments investigating the conduction block in vagal axons, the NaV1 inhibitor was superfused in the vagal compartment of recording chamber (Fig. 1A).
Single-cell RT-PCR.
We described this method previously (5, 18). The neurons innervating the esophagus were retrogradely labeled using retrograde tracer DiI (Invitrogen, Carlsbad, CA) in a solution (1% in DMSO diluted 1:10 in PBS). Guinea pigs were anesthetized by intraperitoneal ketamine (50 mg/mL) and xylazine (2.5 mg/mL). Using a sterile surgical approach, cervical esophagus was exposed through the midline, and the DiI was injected by using Hamilton syringe into two to three sites (2–3 µL/site) into the esophageal wall. Subcutaneous layers and the skin were sutured. After the surgery, animals recovered on a heated pad under close observation for 2–3 h. The animals were monitored for wound healing and the presence of distress and pain daily. No animal in this study met conditions under which early euthanasia would be considered as defined in our animal protocol; 14–21 days after the injection, the animals were euthanized by CO2 asphyxiation and exsanguination. Vagal nodose ganglia were harvested and enzymatically dissociated (2 mg/mL collagenase and 2 mg/mL dispase II dissolved in Ca- and Mg-free Hanks’ balanced salt solution) while repeatedly triturated with glass fire-polished Pasteur pipettes, washed by centrifugation, and suspended in L-15 medium containing 10% fetal bovine serum (FBS). The cell suspension was transferred onto poly-d-lysine/laminin-coated coverslips. After the suspended neurons had adhered to the coverslips for 2 h (37°C), the neuron-attached coverslips were flooded with the L-15 medium (10% FBS), stored at 37°C, and used within 6 h. For cell picking, the coverslips with dissociated neurons were mounted on the microscope stage and constantly perfused by PBS. The DiI-labeled cells were identified using epifluorescence, and labeled-clean single neurons were collected into a glass pipette (tip diameter 50–150 µm, pulled with a micropipette puller P-87; Sutter Instruments, Novato, CA) by applying negative pressure. The pipette tip was then broken in a PCR tube containing 1 µL of RNAse inhibitor (RNaseOUT; 2 U/µL). The tubes were immediately snap-frozen and stored on dry ice. Only one to four cells were collected from each coverslip. The major concern in this type of experiment is whether the sample that also contains some superfusing fluid is contaminated by RNA/DNA from adjacent and/or damaged/lysed cells. To control for this problem, we collected only the neurons that were clean and not attached to other neurons, other cell types, or debris. In addition, our approach using constant flow perfusion of the coverslips during the cell collection repeatedly proved effective to essentially eliminate any contamination. In each experiment, the samples of superfusing fluid from the vicinity of the collected neurons was obtained. These negative controls were found nearly uniformly negative (in a few instances when these controls were positive, all cells from the coverslip were discarded).
First-strand cDNA was synthesized from single neurons by using the Super-Script(tm) III Kit (Thermofisher) according to the manufacturer’s recommendations. Samples were defrosted and lysed (10 min, 75°C). Then, poly(dT) and random hexamer primers (Roche Applied Bioscience) were added. The samples were reverse transcribed by adding Superscript III RT for cDNA synthesis. Two microliters of each sample (cDNA, water control or bath control) was used for PCR amplification by the HotStar Taq Polymerase Kit (Qiagen) according to the manufacturer’s recommendations in a final volume of 20 µL. After an initial activation step 95°C for 15 min, cDNAs were amplified with custom-synthesized primers (Thermofisher) by 50 cycles of denaturation 94°C for 30 s, annealing at 60°C for 30 s, and extension 72°C for 1 min, followed by a final extension 72°C for 10 min. Products were visualized in ethidium bromide-stained 1.5% agarose gels with 50-bp or 100-bp DNA ladder. The figures were prepared by using Microsoft PowerPoint and Apple Preview.
Intron-spanning primers specific for each guinea pig NaV1 α-subunit were designed based on indicated PubMed sequences with the aid of the University of California Santa Cruz (UCSC) Genome Browser, using the Primer3 (version 0.4.0) program (25). These primers were reported previously (Table 1; also see Ref. 16). For most primers, no genomic product can be amplified because its predicted size of >1,000 bp is not achievable with the extension time of 30 s used for PCR. The selectivity of each primer was evaluated by aligning all NaV1 subunits with each primer using ClustalW. RNA for positive control was selected based on established NaV1 subunit expression pattern: whole brain RNA for NaV1.1, 1.2, 1.3, and 1.6; skeletal muscle RNA for NaV1.4; heart muscle RNA for NaV1.5; and dorsal root ganglia (DRG) or vagal sensory ganglia RNA for NaV1.7, 1.8, and 1.9. The guinea pig RNA was isolated by using RNAeasy mini kit (Qiagen).
Table 1.
Guinea pig primer sequences for single cell RT-PCR
| Target (Primer) | Sequence (5′ to 3′) | Product size, bp | Genomic size, bp | NCBI Reference Sequence |
|---|---|---|---|---|
| β-Actin | ||||
| Forward | TCTCTTCCAGCCCTCCTTC | 373 | NA | NT_176389.1 |
| Reverse | GTCCTCAAAGGTGCTGTGCT | |||
| TRPV1 | ||||
| Forward | CCAACAAGAAGGGGTTCACA | 168 | >1,000 | NM_001172652.1 |
| Reverse | ACAGGTCATAGAGCGAGGAG | |||
| NaV1.1 | ||||
| Forward | AAGGGTTTCGCTTCTCCATT | 206 | >1,000 | XM_023561064.1 |
| Reverse | GTCCTCAAAGGTGCTGTGCT | |||
| NaV1.2 | ||||
| Forward | GAGACTTCAGTGGTGCTGGTG | 265 | NA | XM_023561036.1 |
| Reverse | AGCAGAGACTGGTGTGGAGAA | |||
| NaV1.3 | ||||
| Forward | ACACAACAGAGGAGAGGCAGA | 171 | >1,000 | XM_023561157.1 |
| Reverse | CGTCTTGGGGAGAATAGGG | |||
| NaV1.4 | ||||
| Forward | GGCTCTCCCTCCACCATC | 159 | NA | XM_003465929.2 |
| Reverse | CAGCGGTTTCTTGCCATC | |||
| NaV1.5 | ||||
| Forward | GTCATTTTCCTGGGCTCCTT | 245 | >1,000 | XM_005001107.2 |
| Reverse | TTGCTCCTTCTCTCGTGGTT | |||
| NaV1.6 | ||||
| Forward | CCTCCTATGGACGAAAAGACA | 222 | >1,000 | XM_023567676.1 |
| Reverse | TGCAGATGGTGATAGCCAAG | |||
| NaV1.7 | ||||
| Forward | GAGGAAAAGGGAGATGATGAGA | 158 | >1,000 | XM_013142433.2 |
| Reverse | AACAGGGAGCCACGAATG | |||
| NaV1.8 | ||||
| Forward | CGGAAAGGTGACAATGGAG | 187 | >1,000 | XM_013146277.2 |
| Reverse | AGCAGGGACAGTAGCGAAGA | |||
| NaV1.9 | ||||
| Forward | GTTCCAGGTTCCCAAATCAA | 162 | >1,000 | XM_023562606.1 |
| Reverse | GAGGCAGAAGAGGGTGAGG |
NaV1, voltage-gated sodium channels; NA, no match with genomic in UCSC In-Silico PCR; TRPV1, transient receptor potential channel-vanilloid subfamily member 1.
Data analysis.
Nerve activity (action potential discharge) in response to esophageal distention was recorded. The response was quantified as the total number of action potentials (means ± SE) during the 20s of distention (baseline activity if present was subtracted). For comparison of the distention-response curves, the area under the curve (AUC) was used as described previously (5). AUC was calculated using standard geometrical formula (units omitted): AUC = 20 × (T10 + T30)/2 + 30 × (T30 + T60)/2, where T10, T30, and T60 are the total number of action potentials at the distention pressures of 10, 30, and 60 mmHg, respectively, and the coefficients 20 and 30 refer to difference between the tested pressures (i.e., 20 mmHg = 30–10 mmHg. Paired t test, unpaired t test, and ANOVA were used for statistical comparison as indicated in the text and figure legends. The significance was defined as P < 0.05.
Drugs and chemicals.
The compounds used were prepared as stock solutions stored at −20°C and were diluted in Krebs solution to their final concentrations on the day of use. Tetrodotoxin (Alomone Laboratories Jerusalem, Israel), bradykinin (Sigma-Aldrich, St. Louis, MO), and α,β-methylene-ATP (Sigma-Aldrich) were dissolved in distilled water to 1mM, 10 mM and 10 mM stock solutions, respectively. Lidocaine (Sigma-Aldrich) was dissolved in ethanol to 1 mM. ICA121431 (Tocris, Minneapolis, MN), PF05089771 (Tocris), A-803467 (Tocris) and Compound 801 (Lilly, Indianapolis, IN) (16) were dissolved in DMSO to 10 mM each.
RESULTS
The experiments were designed to deliver NaV1 inhibitors to mechanosensitive nerve terminals of nodose C-fibers in the esophagus but to prevent the exposure of their conducting axons in the recurrent laryngeal and vagus nerves to NaV1 inhibitors. The esophagus was preserved as a tube, the esophageal mucosa was gently removed (pulled out from inside), and the NaV1 inhibitors were perfused through the esophageal lumen (Fig. 1A). Mechanical stimulation was performed by graded esophageal distention (20 s) with increasing hydrostatic pressure. The removal of mucosa led to increased esophageal distensibility (Fig. 1B, inset). Thus, a given distention pressure in the esophagus preparations without mucosa evoked larger activation than the same distention pressure in the esophagus preparations with intact mucosa (Fig. 1B). Specifically, in the esophageal preparations without the mucosa, the distention to 10, 30, and 60 mmHg evoked activation comparable with that evoked by distention to ∼35, 60, and 90 mmHg in the preparations with the mucosa (Fig. 1B). This roughly correlated with the changes in esophageal distensibility (Fig. 1B, inset). Based on our previous studies, NaV1 inhibitors were perfused for 30 min to achieve equilibrium (16, 18).
We previously reported that NaV1.7 is required for action potential initiation in nodose C-fibers innervating the lungs (18). Therefore, we first evaluated the effect of the NaV1.7 inhibitor PF-05089771 (1, 30) on the mechanical response of esophageal nodose C-fibers to esophageal distention. By using a guinea pig NaV1.7 nerve tissue bioassay for NaV1.7 (16), we reported that IC50 for the PF-05089771 is ≈0.5µM and that a complete block of the guinea pig NaV1.7 is achieved at the concentration of 3µM (18). Therefore, we initially used 3 µM PF-05089771. The response to esophageal distention was highly reproducible in the time control experiments (n = 11 guinea pigs; Fig. 2B). PF-05089771 (3 µM, 30 min) nearly abolished the response to low levels of esophageal distention (10 mmHg) but only partially inhibited the response to higher levels of esophageal distention (30–60 mmHg) (Fig. 2C). Increasing the concentration of PF-05089771 to 10 µM that is supramaximal for the guinea pig NaV1.7 confirmed the NaV1.7-independent mechanical response in nodose C-fibers at higher levels of esophageal distention (Fig. 2D).
Fig. 2.

The voltage-gated sodium channel (NaV1.7) inhibitor PF-05089771 is more effective at inhibiting the action potential initiated by low levels than higher levels of esophageal distention in nodose C-fibers. A: representative traces of the response to increasing esophageal distention pressure in control conditions and following the incubation with PF-05089771 perfused through the esophageal lumen. #Esophageal mucosa removed. B: reproducibility of the response to esophageal distention (n = 11 guinea pigs). C and D: inhibition of the mechanical response by PF-05089771 (3 µM, n = 6 guinea pigs; *P < 0.05, paired t test, area under the curve) and by PF-05089771 (10 µM, n = 8 guinea pigs; **P < 0.01, paired t test, area under the curve). Note that subsequent application of tetrodotoxin (TTX; 0.3 µM) abolished mechanical response that persisted in the presence of PF-05089771. E: the degree of inhibition of mechanical response by PF-05089771 decreases with increasing distention pressure. ANOVA (P < 0.01) followed by post hoc Tukey test. **P < 0.01 as indicated.
These data show that a substantial proportion (>50%) of the mechanical response of esophageal nodose C-fibers is still present at higher levels of esophageal distention despite a complete block of the guinea pig NaV1.7 by PF-05089771. Importantly, the inhibition of distention-inducer response by PF-05089771(10 µM) diminished with the increasing distention pressure. The response was inhibited by 97%, 64%, and 45% at 10, 30, and 60 mmHg, respectively (Fig. 2E). The inhibitory effect of PF-05089771 on distention with 60 mmHg was concentration dependent. PF-05089771 caused the inhibition by 18 ± 6% (n = 6 guinea pigs), 39 ± 7% (n = 12 guinea pigs), and 48% ± 5 (n = 8 guinea pigs) at concentrations of 0.3 µM, 3 µM, and 10 µM, respectively (P < 0.05, ANOVA followed by post hoc Tukey test). Thus, PF-05089771-sensitive NaV1s are required for mechanical response at low levels of mechanical stimulation, but at higher levels of mechanical stimulation, nodose C-fibers have a PF-05089771-insensitive component that increases with increasing level of stimulation.
The PF-05089771-insensitive (NaV1.7-independent) proportion of mechanical response in nodose C-fibers was abolished by 0.3 µM tetrodotoxin (TTX; evaluated at the high level of esophageal distention 60 mmHg; Fig. 2, B and C). This indicated that the NaV1.7-independent response is mediated by TTX-sensitive NaV1s. Therefore, we evaluated the expression of mRNA for TTX-sensitive NaV1 subunits in the esophageal nodose C-fiber neurons by single-cell RT-PCR. In the guinea pig, the nodose ganglion neurons project two types of afferent fibers into the esophagus, the capsaicin-sensitive [transient receptor potential channel-vanilloid subfamily member 1 (TRPV1)-positive] C-fibers and the capsaicin-insensitive (TRPV1-negative) Aδ-fibers, which are non-nociceptive tension mechanoreceptors (17, 32, 33). Therefore, the neuronal cell bodies of the esophageal nodose C-fibers can be identified by retrograde labeling from the esophagus and the expression of the mRNA for the capsaicin receptor TRPV1. We found that the esophageal nodose C-fiber neurons expressed mRNA for multiple TTX-sensitive NaV1 subunits (Fig. 3). NaV1.7 was expressed in nearly all (95%, 14 of 15) esophageal TRPV1-positive neurons. However, a substantial proportion of these neurons also expressed mRNA for other neuronal TTX-sensitive NaV1s, including NaV1.2 (70%, 25/35), NaV1.6 (60%, 9/15), NaV1.3 (55%, 20/35), and NaV1.1 (45%, 16/35). As expected, the TTX-sensitive NaV1.4 subunit, which is selectively expressed in skeletal muscle, was absent from the esophageal nodose C-fibers neurons (detected only in 1 of 27 tested neurons). In agreement with previous observations in the guinea pig nodose neurons, we also noted that the majority of nodose C-fiber neurons expressed TTX-resistant NaV1 subunits NaV1.8 (80%, 12 of 15) and NaV1.9 (85%, 13 of 15) (19, 21).
Fig. 3.

The esophageal nodose transient receptor potential channel-vanilloid subfamily member 1 (TRPV1)-positive (C-fiber) neurons express mRNA for multiple tetrodotoxin (TTX)-sensitive voltage-gated sodium channel (NaV1) subunits. A: representative gels of NaV1 subunit expression in individual single nodose TRPV1-positive (C-fibers) neurons (numbered) retrogradely labeled from the esophagus detected by single cell RT-PCR. +, Positive control; −, negative bath control. B: proportions of the labeled TRPV1-positive neurons that expressed indicated NaV1s. Numbers in brackets indicate the ratio of positive neurons to tested neurons for each subunit. Neurons were obtained from 6 guinea pigs.
Because nearly 80% of analyzed nodose C-fiber neurons expressed NaV1.1, NaV1.2, and/or NaV1.3, we next evaluated the effect of the group NaV1.1, 1.2, and 1.3 inhibitor ICA-121431 (20) on high levels of esophageal distention (Fig. 4, A and B). ICA-121431 perfused through the esophageal lumen concentration-dependently inhibited the response to 60 mmHg, indicating that ICA-121431-sensitive NaV1s subunits contributed to action potential initiation in nodose C-fiber nerve terminals. In contrast to the inhibition by PF-05089771, which was more effective at low levels and less effective at higher levels of esophageal distention (Fig. 2, C and D), the inhibition by ICA-121431 was less effective at low levels and more effective at the higher levels of esophageal distention (Fig. 4C). Because ICA-121431 inhibited the response to esophageal distention and the pattern of this inhibition (Fig. 4C) was complementary to that of PF-05089771 (Fig. 2, C and D), we hypothesized that the combination of PF-05089771 and ICA-121431 will abolish the response to high level of esophageal distention. However, although the combination of PF-05089771 and ICA-121431 inhibited the response more than PF-05089771, ∼35% of the response was still preserved (Fig. 4D).
Fig. 4.

The effect of the voltage-gated sodium channel (NaV1.1), NaV1.2, and NaV1.3 group inhibitor ICA-121341 on the action potential initiation in the esophageal nodose C-fibers. A: representative traces. The action potential discharge was evoked by esophageal distention, and ICA-121341 was perfused through the esophageal lumen. #Esophageal mucosa removed. B: mean data (n = 6–11 guinea pigs). ANOVA (P < 0.01) followed by post hoc Tukey test. **P < 0.01 as indicated. C: ICA-121431 had only a limited effect at lower levels of esophageal distention but inhibited the response at higher levels of distention (n = 6). D: combination of PF-05089771 and ICA-1214311 inhibited the action potential initiation evoked by high levels of esophageal distention (60 mmHg#) more than PF-05089771 alone (unpaired t test; *P < 0.05). PF-05089771 (3 µM, n = 6 guinea pigs; or 10 µM, n = 8 guinea pigs) and PF-05089771 + ICA-121431 (3 µM and 10 µM, respectively, n = 4 guinea pigs, or 10 µM and 10 µM, respectively, n = 3 guinea pigs). TTX, tetrodotoxin.
These data indicate that a TTX-sensitive NaV1 subunit that is expressed by esophageal C-fiber neurons but not inhibited by ICA-121431 or PF-05089771 contributes to the action potential response to high levels of distention. A likely candidate for this is NaV1.6. To address this possibility, we took advantage of Compound 801, which we recently characterized (16). We reported that the Compound 801 inhibits both NaV1.6 and NaV1.7 (IC50 of for NaV1.6 is <10-fold higher than that of NaV1.7) (Table 1; also see Ref. 16). Compound 801 (10 µM) alone inhibited the response to 60 mmHg by ∼60% but failed to abolish it (Fig. 5). However, the combination of Compound 801 (10 µM) and ICA-121431 (10 µM) almost abolished (inhibited by 91%) the response (Fig. 5).
Fig. 5.

Combination of Compound 801 that inhibits both voltage-gated sodium channel (NaV1.6) and NaV1.7 and the group NaV1.1/1.2/1.3 inhibitor ICA-121431 nearly abolished the response evoked by high level of esophageal distention. A: an example of the effect of incubation with Compound 801 alone followed by combination of Compound 801 and ICA-121431. #Esophageal mucosa removed. B: mean data. Control (n = 20 guinea pigs), Compound 801 (10 µM; n = 8 guinea pigs), ICA-121431 (10 µM; n = 11 guinea pigs), and Compound 801 + ICA-121431 (10 µM each; n = 7 guinea pigs). ANOVA (P < 0.01) followed by post hoc Tukey test. **P < 0.01. Control 2 vs. Compound 801 and control 2 vs. ICA121431 were also significant (P < 0.01 and P < 0.05, respectively) but are not indicated in the figure for clarity.
As described above, the majority (≈80%) of the nodose C-fiber neurons expressed mRNA for TTX-resistant NaV1.8 subunit. Therefore, we also evaluated the effect of NaV1.8 inhibitor A-803467 on mechanical response to distention in nodose C-fibers (14). In the presence of A-803467 (30 µM, 30 min), the response to 60-mmHg distention was 102 ± 13% of the control response (n = 5 guinea pigs, P = 0.9; paired t test).
We have recently reported that the inhibition of NaV1.7 in the nerve terminals of nodose C-fibers in the lung virtually abolished the response to chemical activators (18). Therefore, we evaluated the response to bradykinin (1 µM) and the P2X receptors agonist α,β-methylene-ATP (30 µM) that were shown to activate esophageal nodose C-fibers (31, 33). Unpaired design was selected to avoid desensitization often observed with repeated application of certain chemical activators (31). In control conditions, three of four C-fibers responded to bradykinin (peak frequency: 3, 4, and 8 Hz). In the presence of PF-05089771 (3 µM), two of three C-fibers responded to bradykinin (peak frequency: 4 and 12 Hz). In control conditions, two of three C-fibers responded to α,β-methylene-ATP (peak frequency: 3 and 11 Hz). In the presence of PF-05089771 (3 µM), two of three C-fibers responded (peak frequency: 8 and 11 Hz). Thus, combined five of seven and four of six esophageal nodose C-fibers responded to chemical activators in control conditions and in the presence of PF-05089771, respectively, with similar magnitude of activation. These data indicate that NaV1.7 is not essential for the response to chemical activators in esophageal nodose C-fibers.
The experiments described thus far were designed to investigate action potential initiation in the nerve terminals of esophageal nerve fibers. Therefore, the esophagus was preserved as a tube, the esophageal mucosa was removed, and the NaV1 inhibitors were perfused through the esophageal lumen. In this design, the NaV1 inhibitors delivered into the lumen are predicted to act on the C-fiber nerve terminals in the esophageal wall. However, it cannot be ruled out that a portion of the nodose C-fiber conducting axon runs in the esophageal wall in such a way that it is accessible to NaV1 inhibitors applied into the lumen. Therefore, it is possible that the inhibition of the response to esophageal distention was in fact due to NaV1 inhibitors acting not on the nerve terminals but on the conducting axons of nodose C-fibers in the esophageal wall.
To address this issue, we designed experiments to evaluate the effect of PF-05089771 on action potential conduction in conducting axons of esophageal nodose C-fibers. In these experiments, PF-05089771 was applied selectively to a desheathed segment of the vagus nerve in a separate chamber, which did not contain the esophagus (Fig. 1A). We found that PF-05089771 (10 µM) applied to desheathed vagus nerve caused either no inhibition (in 4 of 9 nodose C-fibers) or a complete block of action potential conduction (in the remaining 5 of 9 nodose C-fibers, in these fibers the electrical stimulation of their receptive fields also failed to evoke action potentials) (Fig. 6, B and C). This type of effect contrasted with the effect of PF-05089771 (10 µM) applied into the esophageal lumen, which in all instances caused a partial inhibition of action potential initiation by 30% to 70% (n = 8 guinea pigs; Fig. 6, A and C). These results strongly argue against the possibility that the NaV1 inhibitors delivered into the esophageal lumen act on the conducting axon of nodose C-fibers in the esophageal wall. If this was the case, PF-05089771 applied into the esophageal lumen would either cause a complete block or have no effect. However, when PF-05089771 was applied into the lumen, it caused a partial inhibition that was never observed when PF-05089771 was applied to desheathed vagus nerve.
Fig. 6.

Distinct effects of PF-05089771 on action potential initiation in nerve terminals versus action potential conduction in the axons of nodose C-fibers. A: the voltage-gated sodium channel (NaV1.7) inhibitor PF-05089771 applied into the esophagus containing nerve terminals of nodose C-fibers caused a consistent partial inhibition of action potential initiation evoked by high levels of distention (60 mmHg) in all C-fibers tested (data from Fig. 2D). B: in contrast, PF-05089771 applied to the desheathed vagus nerve containing the axons of nodose C-fibers either completely blocked or had no effect on conduction of action potentials evoked by the same distention (60 mmHg) in nodose C-fibers. Horizontal bars in A and B indicate esophageal distention to 60 mmHg for 20 s. C: individual data points showing partial inhibition of action potential initiation in nerve terminals versus complete block or no effect of action potential conduction in axons by PF-05089771.
DISCUSSION
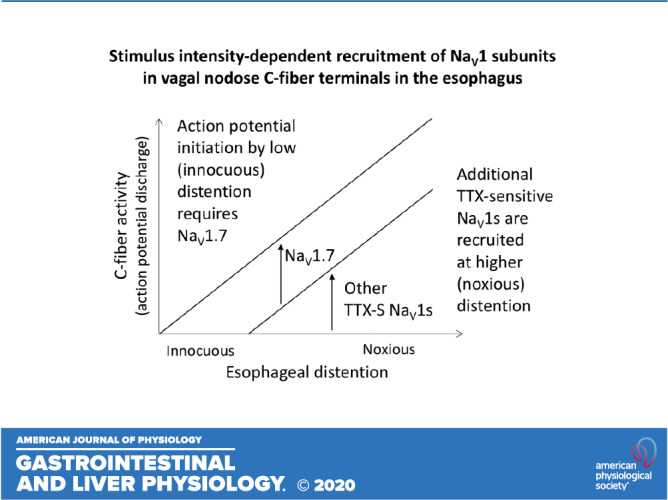
Our major finding is that the nature of the NaV1 subunits involved in action potential initiation evoked by mechanical stimulation (esophageal distention) in esophageal C-fiber terminals is dependent on the intensity of the stimulation. We found that action potential initiation at low levels of mechanical stimulation was entirely blocked by PF-05089771. However, action potential initiation at higher levels of mechanical stimulation had a substantial PF-05089771-insensitive component, which increased with increasing intensity of stimulation. We hypothesize that NaV1.7 is essential for action potential initiation by low levels of mechanical stimulation, but additional TTX-sensitive NaV1s are recruited for action potential initiation at higher levels of stimulation. Our main conclusion that at least one additional type of NaV1 subunit is recruited at higher level of mechanical stimulation is sufficiently supported by current data. PF-05089771 inhibited the action potential initiation at both low and high levels of stimulation (Fig. 2, B and C). In contrast, ICA-121431 did not inhibit the action potential initiation at low levels but inhibited it at higher levels of stimulation (Fig. 4C). If the effects of both PF-05089771 and ICA-121431 were due action on the same NaV1 subunits, then their effect would be similar at both low and high levels of mechanical stimulation.
The information on pharmacology of guinea pig NaV1s is limited (16, 18), and therefore, we base our discussion on the data from human NaV1s with the caveat that there are known species differences in NaV1 pharmacology (e.g., see Refs. 1 and 20). However, in some instances, the key structural determinants responsible for effects of NaV1 inhibitors are known and are similar between humans and guinea pigs, allowing for informed extrapolations. PF-05089771 in the concentration of 3 µM completely blocked the response mediated by NaV1.7 in the guinea pig nerve tissue bioassay (18). Thus, the proportion (≈50%) of the response to high level of esophageal distention that was preserved in the presence of PF-05089771 (3 µM) is independent of NaV1.7 (Fig. 2). We reported that Compound 801 in the concentration of 1µM also completely blocked the guinea pig NaV1.7 (16). Nonetheless, ∼40% of the response to high levels of esophageal distention was preserved in the presence of Compound 801 (10 µM; Fig. 4B). These data strongly support the conclusion that the response of esophageal nodose C-fibers to high levels of mechanical stimulation has a substantial component independent of NaV1.7.
The NaV1.7-independent response to high levels of esophageal distention was abolished by TTX (0.3 µM), indicating that TTX-sensitive NaV1.1, NaV1.2, NaV1.3, NaV1.4, and/or NaV1.6 are involved. We found that in addition to NaV1.7, the esophageal C-fiber neurons express various combinations of mRNAs for NaV1.1, NaV1.2, NaV1.3, and/or NaV1.6 (Fig. 3). This NaV1 expression profile was similar to that previously reported in visceral DRG neurons innervating the colon (13). Because of frequent expression of NaV1.1, NaV1.2, and NaV1.3 mRNA, we evaluated the effect of the group NaV1.1, NaV1.2, and NaV1.3 inhibitor ICA-121431. ICA-121431 inhibits human NaV1.1, NaV1.2, and NaV1.3, and this inhibition depends on three specific amino acid residues within the S2–S4 region of domain 4 (20). The comparison of the predicted guinea pig NaV1.1, NaV1.2, and NaV1.3 protein sequences with their human orthologs revealed that the guinea pig and human NaV1.1, NaV1.2, and NaV1.3 have identical amino acid residues in all of these three critical positions. Thus, it is probable that ICA-121431 inhibits the guinea pig NaV1.1, NaV1.2, and NaV1. In contrast, ICA-121431 does not inhibit human NaV1.6 (IC50 >10µM) and NaV1.7 (IC50 >10 µM), which have different amino acid residues in the three critical positions. Again, the human and the predicted guinea pig NaV1.6 and NaV1.7 have identical amino acid residues at those three positions, and thus ICA-121431 is not expected to inhibit the guinea pig NaV1.6 and NaV1.7. Indeed, we previously noted that ICA-121431 in the concentrations up to 10 µM had no effect on the guinea pig NaV1.7 (16).
Because ∼30% of the response to high levels of esophageal distention was still present in the presence of a combination of PF-05089771 and ICA-121431 (together expected to block NaV1.1, NaV1.2, NaV1.3, and NaV1.7), and this response was abolished by TTX, the only remaining TTX-sensitive channel expressed in C-fiber neurons, NaV1.6, was implicated. To address the involvement of NaV1.6, we took advantage of Compound 801, which is only approximately eightfold less potent to block NaV1.6 than to block NaV1.7 (16). Therefore, if Compound 801 in a concentration of 10 µM, which is 10 times higher than that required to completely block NaV1.7, would not block the remaining response, it would strongly argue against the role of NaV1.6. However, we found that Compound 801 (10 µM) inhibited the response to high level of esophageal distention more than PF-05089771 alone and that Compound 801 (10 µM) in combination with ICA-121431 caused >90% inhibition of the response to high level of esophageal distention (Fig. 5).
Excessive esophageal distention is most often caused by esophageal obstruction, triggers intense pain and reflex increases in motility and salivation, and, if unresolved, leads to esophageal rupture. Because it threatens integrity of the tissue and evokes pain, it is a bona fide noxious stimulus. In humans, physiological (innocuous) distention of <30 mmHg, such as during swallowing, rarely evokes perceptions; however, a high level of distention >50 mmHg invariably evokes pain that is linearly proportional to distention pressure (23, 24). In rats, esophageal distention at similarly high levels evokes pseudoaffecitve visceromotor and cardiovascular responses consistent with pain (9). Indeed, esophageal distention activates spinal DRG nociceptors (28, 29), and this activation is linearly dependent on distention pressure. The relevant distention pressure for activation of esophageal nerves is relatively similar between the species (27, 33). Therefore, irrespective of the nerve type studied, the relatively high level of esophageal distention evaluated in our studies can be safely considered noxious. Recognizing low levels of distention as innocuous and high levels as noxious puts our findings in the context of esophageal physiology.
Similar to many visceral afferent fibers implicated in nociception, including esophageal spinal DRG nociceptors (28, 29), esophageal nodose C-fibers encode both physiologically innocuous and noxious levels of esophageal distention (intact mucosa; Fig. 1B) (17, 32, 33). Note that because the esophageal mucosa was removed in this study, the distention to 10 mmHg evoked activation equivalent to distention with innocuous 35 mmHg in the esophagus with the mucosa, whereas distention to 60 mmHg without mucosa evoked activation equivalent to bona fide noxious distention to 90 mmHg in the esophagus with the intact mucosa (Fig. 1B). The ability to encode both innocuous and noxious distention suggests that nodose C-fibers contribute to both physiological regulation of esophageal motility as well as to nociceptive reflexes (e.g., nausea and vomiting) and possibly sensations evoked by noxious stimuli. Our data support the conclusion that the action potential initiation in response to innocuous mechanical stimuli requires PF-05089771-sensitive NaV1 (presumably NaV1.7), whereas both PF-05089771-sensitive and PF-05089771-insensitive NaV1s contribute to noxious mechanical stimulation. This may offer clinically relevant opportunities for refining inhibition strategies. Targeting NaV1.7, which mediates the response to low levels of mechanical stimulation (Fig. 2), should be useful in conditions when the goal is to improve reflex dysregulation (such as certain spastic motility disorders) while limiting the effects on nociception. Targeting the response to low levels of stimulation may also be useful in conditions with pronounced central sensitization when innocuous visceral stimuli are interpreted as noxious by CNS and lead to discomfort or pain (analogous to allodynia in somatosensory system). Indeed, central hypersensitivity of esophageal sensory pathways has been demonstrated in esophageal disorders such as noncardiac chest pain and gastresophageal reflux disease (15, 26). The inhibition of NaV1.7 may be accomplished by systemic administration of NaV1.7 inhibitor and possibly by emerging genetic silencing approaches of NaVs selectively in targeted nerve types in the future.
In contrast, inhibiting NaV1s that are recruited at higher levels of stimulation would largely preserve the response to low levels of mechanical stimulation but would inhibit, although not abolish, the response to higher levels of stimulation. In the case of esophageal nodose C-fibers, such inhibition is exemplified by ICA-121431 (Fig. 4C). This strategy may be useful in conditions when C-fibers are exposed to noxious stimuli causing excessive stimulation. By inhibiting NaV1s that are recruited at higher levels of stimulation, the response to innocuous stimuli and, therefore, physiological regulation would be largely preserved, but the response to noxious stimuli would be inhibited. Again, this may be accomplished by systemic administration of appropriate NaV1 inhibitors (selective or group inhibitors similar to ICA-121431) once NaV1s involved are identified or by emerging targeted genetic silencing approaches.
The mechanism by which NaV1s other than NaV1.7 are recruited at higher levels of stimulation cannot be deduced from our studies. We speculate that the distribution of NaV1.7 and other NaV1s in the nerve terminals together with their biophysical properties underlie this phenomenon. One possibility is that the terminal membrane close to mechanically gated channels contains mostly NaV1.7, whereas a more distant membrane also contains other NaV1s implicated in action potential initiation at higher level of stimulation. Therefore, a generator potential evoked by a low level of mechanical stimulation requires NaV1.7, whereas larger generator potential results also in gating other more distant NaV1s even when NaV1.7 inhibited. Regrettably, this must remain a mere speculation, as this hypothesis cannot be readily addressed at the level of afferent nerve terminals with the technologies presently available.
Isolated ex vivo preparations are particularly suited for the study of action potential initiation in nerve terminals because they can be designed to allow for pharmacological inhibition of NaV1s in the nerve terminals without inhibiting conducting axons and vice versa (Fig. 1A and Ref. 18). It remains to be investigated whether the recruitment of additional different NaV1 subunits for action potential initiation at higher levels of stimulation occurs in other types of visceral and somatic C-fibers. In the meantime, it would be prudent not to assume that the same NaV1 subunit(s) are responsible for action potential initiation at all levels of stimulation. Moreover, perhaps the recruitment of additional different NaV1 subunits may help to explain the puzzling lack of an apparent role of NaV1.7 in visceral pain (6, 13). Because genetic and neuromodulation strategies targeting specific neuronal population are expected to overcome the most limiting aspect of systemic NaV1 inhibition (side effects from unwanted inhibition of other nerve types), the need for knowledge of NaV1s regulating visceral nociception is becoming more urgent.
GRANTS
This study was supported by Eli Lilly and Company and National Institutes of Health Stimulating Peripheral Activity to Relieve Conditions Grant U01-DK-116311 (to M. Kollarik).
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
J.L.K. and M.K. conceived and designed research; F.R. and N.P. performed experiments; F.R., N.P., and B.J.U. analyzed data; F.R., J.L.K., J.S.M., B.J.U., and M.K. interpreted results of experiments; N.P. prepared figures; N.P., J.L.K., J.S.M., and M.K. drafted manuscript; N.P., B.J.U., and M.K. edited and revised manuscript; F.R., N.P., J.L.K., J.S.M., B.J.U., and M.K. approved final version of manuscript.
ACKNOWLEDGMENTS
Present address of J. L. Krajewski: Merck Research Laboratories, Kenilworth, NJ.
REFERENCES
- 1.Alexandrou AJ, Brown AR, Chapman ML, Estacion M, Turner J, Mis MA, Wilbrey A, Payne EC, Gutteridge A, Cox PJ, Doyle R, Printzenhoff D, Lin Z, Marron BE, West C, Swain NA, Storer RI, Stupple PA, Castle NA, Hounshell JA, Rivara M, Randall A, Dib-Hajj SD, Krafte D, Waxman SG, Patel MK, Butt RP, Stevens EB. Subtype-selective small molecule inhibitors reveal a fundamental role for Nav1.7 in nociceptor electrogenesis, axonal conduction and presynaptic release. PLoS One 11: e0152405, 2016. doi: 10.1371/journal.pone.0152405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bielefeldt K, Ozaki N, Gebhart GF. Experimental ulcers alter voltage-sensitive sodium currents in rat gastric sensory neurons. Gastroenterology 122: 394–405, 2002. doi: 10.1053/gast.2002.31026. [DOI] [PubMed] [Google Scholar]
- 3.Bielefeldt K, Zhong F, Koerber HR, Davis BM. Phenotypic characterization of gastric sensory neurons in mice. Am J Physiol Gastrointest Liver Physiol 291: G987–G997, 2006. doi: 10.1152/ajpgi.00080.2006. [DOI] [PubMed] [Google Scholar]
- 4.Brierley SM, Page AJ, Hughes PA, Adam B, Liebregts T, Cooper NJ, Holtmann G, Liedtke W, Blackshaw LA. Selective role for TRPV4 ion channels in visceral sensory pathways. Gastroenterology 134: 2059–2069, 2008. doi: 10.1053/j.gastro.2008.01.074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Brozmanova M, Mazurova L, Ru F, Tatar M, Hu Y, Yu S, Kollarik M. Mechanisms of the adenosine A2A receptor-induced sensitization of esophageal C fibers. Am J Physiol Gastrointest Liver Physiol 310: G215–G223, 2016. doi: 10.1152/ajpgi.00350.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Erickson A, Deiteren A, Harrington AM, Garcia-Caraballo S, Castro J, Caldwell A, Grundy L, Brierley SM. Voltage-gated sodium channels: (NaV )igating the field to determine their contribution to visceral nociception. J Physiol 596: 785–807, 2018. doi: 10.1113/JP273461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Feng B, Zhu Y, La JH, Wills ZP, Gebhart GF. Experimental and computational evidence for an essential role of NaV1.6 in spike initiation at stretch-sensitive colorectal afferent endings. J Neurophysiol 113: 2618–2634, 2015. doi: 10.1152/jn.00717.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gebhart GF, Bielefeldt K. Physiology of visceral pain. Compr Physiol 6: 1609–1633, 2016. doi: 10.1002/cphy.c150049. [DOI] [PubMed] [Google Scholar]
- 9.Gebhart GF, Sengupta JN. Evaluation of visceral pain. In: Handbook of Methods in Gastrointestinal Pharmacology, edited by Gaginella TS. Boca Raton, FL: CRC Press, 1996, p. 359–373. [Google Scholar]
- 10.Grundy D. Principles and standards for reporting animal experiments in The Journal of Physiology and Experimental Physiology. J Physiol 593: 2547–2549, 2015. doi: 10.1113/JP270818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Grundy L, Erickson A, Brierley SM. Visceral pain. Annu Rev Physiol 81: 261–284, 2019. doi: 10.1146/annurev-physiol-020518-114525. [DOI] [PubMed] [Google Scholar]
- 12.Hockley JR, Boundouki G, Cibert-Goton V, McGuire C, Yip PK, Chan C, Tranter M, Wood JN, Nassar MA, Blackshaw LA, Aziz Q, Michael GJ, Baker MD, Winchester WJ, Knowles CH, Bulmer DC. Multiple roles for NaV1.9 in the activation of visceral afferents by noxious inflammatory, mechanical, and human disease-derived stimuli. Pain 155: 1962–1975, 2014. doi: 10.1016/j.pain.2014.06.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hockley JR, González-Cano R, McMurray S, Tejada-Giraldez MA, McGuire C, Torres A, Wilbrey AL, Cibert-Goton V, Nieto FR, Pitcher T, Knowles CH, Baeyens JM, Wood JN, Winchester WJ, Bulmer DC, Cendán CM, McMurray G. Visceral and somatic pain modalities reveal NaV 1.7-independent visceral nociceptive pathways. J Physiol 595: 2661–2679, 2017. doi: 10.1113/JP272837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Jarvis MF, Honore P, Shieh CC, Chapman M, Joshi S, Zhang XF, Kort M, Carroll W, Marron B, Atkinson R, Thomas J, Liu D, Krambis M, Liu Y, McGaraughty S, Chu K, Roeloffs R, Zhong C, Mikusa JP, Hernandez G, Gauvin D, Wade C, Zhu C, Pai M, Scanio M, Shi L, Drizin I, Gregg R, Matulenko M, Hakeem A, Gross M, Johnson M, Marsh K, Wagoner PK, Sullivan JP, Faltynek CR, Krafte DS. A-803467, a potent and selective Nav1.8 sodium channel blocker, attenuates neuropathic and inflammatory pain in the rat. Proc Natl Acad Sci USA 104: 8520–8525, 2007. doi: 10.1073/pnas.0611364104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Knowles CH, Aziz Q. Visceral hypersensitivity in non-erosive reflux disease. Gut 57: 674–683, 2008. doi: 10.1136/gut.2007.127886. [DOI] [PubMed] [Google Scholar]
- 16.Kocmalova M, Kollarik M, Canning BJ, Ru F, Adam Herbstsomer R, Meeker S, Fonquerna S, Aparici M, Miralpeix M, Chi XX, Li B, Wilenkin B, McDermott J, Nisenbaum E, Krajewski JL, Undem BJ. Control of neurotransmission by NaV1.7 in human, guinea pig, and mouse airway parasympathetic nerves. J Pharmacol Exp Ther 361: 172–180, 2017. doi: 10.1124/jpet.116.238469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kollarik M, Ru F, Brozmanova M. Vagal afferent nerves with the properties of nociceptors. Auton Neurosci 153: 12–20, 2010. doi: 10.1016/j.autneu.2009.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kollarik M, Sun H, Herbstsomer RA, Ru F, Kocmalova M, Meeker SN, Undem BJ. Different role of TTX-sensitive voltage-gated sodium channel (NaV 1) subtypes in action potential initiation and conduction in vagal airway nociceptors. J Physiol 596: 1419–1432, 2018. doi: 10.1113/JP275698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kwong K, Carr MJ, Gibbard A, Savage TJ, Singh K, Jing J, Meeker S, Undem BJ. Voltage-gated sodium channels in nociceptive versus non-nociceptive nodose vagal sensory neurons innervating guinea pig lungs. J Physiol 586: 1321–1336, 2008. doi: 10.1113/jphysiol.2007.146365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.McCormack K, Santos S, Chapman ML, Krafte DS, Marron BE, West CW, Krambis MJ, Antonio BM, Zellmer SG, Printzenhoff D, Padilla KM, Lin Z, Wagoner PK, Swain NA, Stupple PA, de Groot M, Butt RP, Castle NA. Voltage sensor interaction site for selective small molecule inhibitors of voltage-gated sodium channels. Proc Natl Acad Sci USA 110: E2724–E2732, 2013. doi: 10.1073/pnas.1220844110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Muroi Y, Ru F, Kollarik M, Canning BJ, Hughes SA, Walsh S, Sigg M, Carr MJ, Undem BJ. Selective silencing of Na(V)1.7 decreases excitability and conduction in vagal sensory neurons. J Physiol 589: 5663–5676, 2011. doi: 10.1113/jphysiol.2011.215384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Osteen JD, Herzig V, Gilchrist J, Emrick JJ, Zhang C, Wang X, Castro J, Garcia-Caraballo S, Grundy L, Rychkov GY, Weyer AD, Dekan Z, Undheim EA, Alewood P, Stucky CL, Brierley SM, Basbaum AI, Bosmans F, King GF, Julius D. Selective spider toxins reveal a role for the Nav1.1 channel in mechanical pain. Nature 534: 494–499, 2016. doi: 10.1038/nature17976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Pedersen J, Reddy H, Funch-Jensen P, Arendt-Nielsen L, Gregersen H, Drewes AM. Differences between male and female responses to painful thermal and mechanical stimulation of the human esophagus. Dig Dis Sci 49: 1065–1074, 2004. doi: 10.1023/B:DDAS.0000037789.25734.06. [DOI] [PubMed] [Google Scholar]
- 24.Remes-Troche JM, Attaluri A, Chahal P, Rao SS. Barostat or dynamic balloon distention test: which technique is best suited for esophageal sensory testing? Dis Esophagus 25: 584–589, 2012. doi: 10.1111/j.1442-2050.2011.01294.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Rozen S, Skaletsky J. Primer3 on the WWW for general users and for biologist programmers. In: Bioinformatics Methods and Protocols: Methods in Molecular Biology, edited by Krawetz S and Misener S. Totowa, NJ: Humana Press, 2000, p. 365–386. [DOI] [PubMed] [Google Scholar]
- 26.Sarkar S, Aziz Q, Woolf CJ, Hobson AR, Thompson DG. Contribution of central sensitisation to the development of non-cardiac chest pain. Lancet 356: 1154–1159, 2000. doi: 10.1016/S0140-6736(00)02758-6. [DOI] [PubMed] [Google Scholar]
- 27.Sengupta JN, Kauvar D, Goyal RK. Characteristics of vagal esophageal tension-sensitive afferent fibers in the opossum. J Neurophysiol 61: 1001–1010, 1989. doi: 10.1152/jn.1989.61.5.1001. [DOI] [PubMed] [Google Scholar]
- 28.Sengupta JN, Saha JK, Goyal RK. Differential sensitivity to bradykinin of esophageal distension-sensitive mechanoreceptors in vagal and sympathetic afferents of the opossum. J Neurophysiol 68: 1053–1067, 1992. doi: 10.1152/jn.1992.68.4.1053. [DOI] [PubMed] [Google Scholar]
- 29.Sengupta JN, Saha JK, Goyal RK. Stimulus-response function studies of esophageal mechanosensitive nociceptors in sympathetic afferents of opossum. J Neurophysiol 64: 796–812, 1990. doi: 10.1152/jn.1990.64.3.796. [DOI] [PubMed] [Google Scholar]
- 30.Swain NA, Batchelor D, Beaudoin S, Bechle BM, Bradley PA, Brown AD, Brown B, Butcher KJ, Butt RP, Chapman ML, Denton S, Ellis D, Galan SRG, Gaulier SM, Greener BS, de Groot MJ, Glossop MS, Gurrell IK, Hannam J, Johnson MS, Lin Z, Markworth CJ, Marron BE, Millan DS, Nakagawa S, Pike A, Printzenhoff D, Rawson DJ, Ransley SJ, Reister SM, Sasaki K, Storer RI, Stupple PA, West CW. Discovery of clinical candidate 4-[2-(5-amino-1H-pyrazol-4-yl)-4-chlorophenoxy]-5-chloro-2-fluoro-N-1,3-thiazol-4-ylbenzenesulfonamide (PF-05089771): design and optimization of diaryl ether aryl sulfonamides as selective inhibitors of NaV1.7. J Med Chem 60: 7029–7042, 2017. doi: 10.1021/acs.jmedchem.7b00598. [DOI] [PubMed] [Google Scholar]
- 31.Yu S, Ouyang A. TRPA1 in bradykinin-induced mechanical hypersensitivity of vagal C fibers in guinea pig esophagus. Am J Physiol Gastrointest Liver Physiol 296: G255–G265, 2009. doi: 10.1152/ajpgi.90530.2008. [DOI] [PubMed] [Google Scholar]
- 32.Yu S, Ru F, Ouyang A, Kollarik M. 5-Hydroxytryptamine selectively activates the vagal nodose C-fibre subtype in the guinea-pig oesophagus. Neurogastroenterol Motil 20: 1042–1050, 2008. doi: 10.1111/j.1365-2982.2008.01136.x. [DOI] [PubMed] [Google Scholar]
- 33.Yu S, Undem BJ, Kollarik M. Vagal afferent nerves with nociceptive properties in guinea-pig oesophagus. J Physiol 563: 831–842, 2005. doi: 10.1113/jphysiol.2004.079574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Zagorodnyuk VP, Brookes SJ. Transduction sites of vagal mechanoreceptors in the guinea pig esophagus. J Neurosci 20: 6249–6255, 2000. doi: 10.1523/JNEUROSCI.20-16-06249.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]