

Abstract
The slack length of a relaxed skeletal muscle can be reduced by isometric contraction at short lengths (“contract-short conditioning”). This study explored how the effect of contract-short conditioning on muscle slack length is modified by 1) the intensity of the contraction, 2) the delay between the contraction and measurement of slack length, and 3) the amplitude of a stretch delivered to the relaxed muscle after the contraction. Muscle fascicles in the human vastus lateralis muscle were observed with ultrasound imaging while the relaxed muscle was lengthened by flexing the knee. The knee angle at which muscle fascicle slack was taken up was used as a proxy for muscle slack length. Conditioning the muscle with voluntary isometric (fixed-end) contractions at short muscle lengths reduced vastus lateralis muscle slack length, measured 60 s later, by a mean of 10°. This effect was independent of contraction intensity from 5% to 100% maximal voluntary contraction. The effect was largest when first observed 5 s after the contraction, decayed about one-third by 60 s, and then remained nearly constant until the last observation 5 min after the contraction. A slow stretch given to the relaxed muscle after contract-short conditioning increased slack length (i.e., reduced the effect of contract-short conditioning). Slack length increased nonlinearly with stretch amplitude. Very large stretches (>30°, possibly as large as 90°) were required to abolish the effect of contract-short conditioning. The phenomena described here share some characteristics with, and may involve similar mechanisms to, passive force enhancement and muscle thixotropy.
NEW & NOTEWORTHY The slack length of a relaxed human skeletal muscle is not fixed; it can be modified by contraction and stretch. Contraction of the human vastus lateralis muscle at short lengths reduces the muscle’s slack length. Even very weak contractions are sufficient to induce this effect. The effect persists for at least 5 min but can be reduced or abolished with a large-amplitude passive stretch.
Keywords: muscle, passive force enhancement, passive muscle properties, slack length, thixotropy
INTRODUCTION
The amount of tension generated by a skeletal muscle at any moment in time depends not just on the muscle’s current length and activation, but also on the lengths and activation levels that the muscle was exposed to in its recent past. Thus the mechanical properties of muscle are said to be “history dependent.”
Relaxed muscles demonstrate a range of history-dependent behaviors. These include viscous behaviors such as stress relaxation, creep, and hysteresis (40), as well as muscle thixotropy and passive force enhancement. Muscle thixotropy manifests in several ways, one of which is that a relaxed muscle subjected to a large-amplitude quick stretch two times in quick succession is less stiff on the second stretch (5). Another manifestation of muscle thixotropy is that passive tension exerted by a relaxed muscle is lower after the muscle has been conditioned with an isometric contraction at stretched lengths than after the muscle has been conditioned with an isometric contraction at a shorter length (43). Passive force enhancement refers to the observation that passive tension at a particular muscle length is greater after active stretch than after a passive stretch or an isometric (fixed-end) contraction at that length (12).
The molecular mechanisms that mediate these and other history-dependent phenomena in relaxed skeletal muscle are poorly understood. Viscous behaviors in muscle could be caused by the unfolding and refolding of titin molecules (28, 31). Campbell and Lakie hypothesize that muscle thixotropy is partly mediated by actin-myosin cross-bridges which form spontaneously in resting muscle but which are broken by muscle contraction or large-amplitude stretches (5), and partly by non-cross-bridge mechanisms, possibly involving titin or myosin-binding protein C (24). Herzog has hypothesized that passive force enhancement occurs because muscle contraction exposes attachment sites on actin filaments that allow titin to bind to actin, reducing titin’s free-spring length and increasing passive tension in myofibrils (13). There is some debate about whether titin contributes significantly to the stiffness of relaxed mammalian skeletal muscles (3, 30) but, because titin is a “tunable spring” (i.e., its length and stiffness can be rapidly modified; see Ref. 34), and because cleavage of titin greatly reduces the stiffness of mammalian muscle fiber bundles at physiological sarcomere lengths (38), titin appears well suited to explain the history dependence of relaxed muscle.
We recently developed a simple procedure to measure the slack length of single human skeletal muscles in vivo (39). To measure the slack length of the vastus lateralis muscle we first passively extend the participant’s knee. With the knee nearly fully extended, the relaxed vastus lateralis muscle is short and at this length it falls slack (i.e., it exerts no passive tension). The length of muscle fascicles in the vastus lateralis muscle are observed in real time using ultrasound imaging. When, subsequently, the relaxed muscle is slowly lengthened by flexing the knee, muscle fascicle length initially does not change. Only when the muscle has been lengthened enough to take up the slack do the muscle fascicles begin to lengthen (9, 10). The objective of this procedure is to measure the muscle length at which slack is taken up. However, it is easier to measure joint angle than muscle length, and joint angle is a good proxy for the length of a relaxed muscle-tendon unit. So, for convenience, we refer to the knee joint angle at which the slack is taken up as the muscle slack length.
Using this method, we demonstrated history dependence of the slack length of the human vastus lateralis muscle in vivo (39). After a standard conditioning protocol, mean muscle slack length occurred at a knee angle of 24° flexion. When, however, participants performed a strong voluntary isometric contraction of the vastus lateralis muscle with the muscle at a short length, mean muscle slack length was reduced to 10°. (Here and elsewhere in this paper the term “isometric” is used to mean “fixed-end.” Muscle fibers and sarcomeres may shorten during fixed-end contractions and then lengthen as the muscle relaxes.) Thus muscle slack length is reduced by “contract-short conditioning.” Stretching the relaxed muscle by passively flexing the knee through a 90° range after the conditioning contraction completely abolished the effect of contract-short conditioning.
In this study we sought to explore how three factors—the intensity of the conditioning contraction, the period between the conditioning contraction and measurement of muscle slack length, and the amplitude of a stretch delivered after the conditioning contraction—influence the effect of contract-short conditioning on the slack length of the human vastus lateralis muscle in vivo.
METHODS
Ethical Approval
The study was approved by the University of New South Wales Human Research Ethics Committee (approval no. 15/006) and conducted in accordance with the Declaration of Helsinki (2013) except for registration in a database. Participants provided informed consent in writing.
Participants
Fourteen healthy adults with no known lower limb musculoskeletal pathology each participated in up to three experiments. Ten people [3 women and 7 men, mean age 26.4 ± 5.9 (SD) yr] participated in experiment 1 and experiment 3, and ten people (6 women and 4 men, mean age 26.4 ± 6.1 yr) participated in experiment 2. Six people participated in all three experiments. Experiments 1 and 3 were completed on the same day in random order.
Experimental Set-Up
Participants were seated with the left shank firmly strapped to a dynamometer (Cybex Norm with Humac; CSMi, Stoughton, MA) that flexed and extended the knee at 5°/s from maximum comfortable extension (experiments 1 and 3: 4.1 ± 3.7°; experiment 2: 2.5 ± 2.3°) to ~90° of flexion. The knee angle was measured as the angle between the anterior aspect of the tibia and a line drawn from the lateral condyle to the greater tubercle of the femur. For each participant in each experiment, knee range of motion was kept constant across conditions. Pilot studies showed that even slight misalignment of the axes of rotation of the dynamometer and knee joint caused substantial discrepancies between the amount of rotation of the dynamometer and the knee. For this reason, we repositioned the axis of the dynamometer until changes in the inclination of the shank and the dynamometer input arm, measured with a digital inclinometer, were the same (±2°). Pilot studies also showed that the knee extended by as much as 10° during maximal knee extension contractions due to compression of the seat cushion. Consequently, the cushion was replaced with a steel plate. With the steel plate in place, the maximal change in knee angle during maximal knee extension contractions was ~1°. The dynamometer generated a knee angle signal that was sampled at 50 Hz (16 bit DAQ, 1401; CED, Cambridge, UK).
Electromyography (EMG) signals were recorded from the proximal and distal ends of the left vastus lateralis muscle with pairs of Ag-AgCl electrodes (diameter 10 mm, spacing 30 mm; Cleartrace; ConMed Corporation, Utica, NY). The ground electrode was placed over the patella. EMG signals were amplified (×3,000), band-pass filtered (20–500 Hz; isolated preamplifier, 1902; CED, Cambridge, UK), and sampled at 2 kHz.
Ultrasound images of vastus lateralis muscle fascicles were obtained using two 46-mm linear array ultrasound transducers (MyLab25 with LA522E transducers, operating at 12 MHz; Esaote, Firenze, Italy) held together in a custom-built mold (9). The use of two ultrasound transducers increased the field of view. The transducers were placed over the midbelly of the vastus lateralis muscle, and the orientation of the transducers was adjusted to generate the clearest possible image of the muscle fascicles (23). Tape was placed on the skin to facilitate consistent placement of the transducer throughout the experiment. Images from the two transducers were captured synchronously with a dual-channel video capture card at 15 Hz (i.e., 15 complete images/s) with Spike2 software and the S2video plug-in (CED, Cambridge, UK).
Except when performing the voluntary knee extension contractions that were part of the experimental protocols (see below), participants were instructed to stay completely relaxed. Vastus lateralis EMG signals and ultrasound images were monitored. Trials with overt muscle activity (e.g., trials in which there was a clear burst of activity on EMG recordings or twitch-like fascicle shortening visible on ultrasound) were stopped and repeated.
Experimental Protocol
There were three experiments (Fig. 1). In each experiment, isometric contraction of the knee extensor muscles at short muscle lengths (the “contract-short” condition) was used to reduce the slack length of the vastus lateralis muscle compared with when the muscle did not contract at short lengths (the “reference” condition).
Fig. 1.

Experimental protocols. A: experiment 1; B: experiment 2; C: experiment 3. All three experiments included one or more reference conditions (left) and several contract-short conditions (right). For all protocols, the horizontal axis represents time, and the vertical axis represents knee angle (a surrogate for muscle length). All trials started with participants performing a 2- to 3-s maximum voluntary isometric knee extension contraction (MVC) at a long muscle length (knee flexed ~90°). The knee was then passively extended to the maximum comfortable knee extension and stayed in this position until muscle slack length was measured with ultrasound (the start of which is indicated with triangles marked “US”). For contract-short conditions, an additional isometric knee extension contraction was performed when the knee first reached the extended knee position. For the contract-short and reference conditions of each experiment, the time between first reaching the extended knee position and the measurement of muscle slack length was the same. A: experiment 1 included one reference condition and four contract-short conditions each with a different intensity of conditioning contraction (5, 25, 50, or 100% maximum electromyography amplitude). B: experiment 2 included four reference conditions and four contract-short conditions each with a different delay (5, 60, 180, or 300 s) between the conditioning contraction and measurement of muscle slack length. C: experiment 3 included one reference condition and four contract-short conditions each with four different amplitude passive knee flexion-extension cycles (0°, 5°, 30°, or 90°) after the conditioning contraction.
For the reference condition, the knee was passively positioned in 90° of flexion (i.e., with the muscle moderately stretched). The participant performed a 2–3 s maximal voluntary isometric knee extension contraction in this position. Next, the knee was passively extended as far as the participant comfortably tolerated, taking the muscle to a short length. The knee remained relaxed in this position until muscle slack length was tested. The contract-short condition was the same as the reference condition except that when the knee reached the extended position participants performed an additional 2–3 s voluntary isometric knee extension contraction.
After each trial of each condition in all experiments, measurements of the slack length of the vastus lateralis muscle were obtained. To measure muscle slack length, the knee was passively flexed (i.e., the vastus lateralis muscle was passively lengthened) at 5°/s from the maximally extended position (i.e., from a short muscle length). Ultrasound images were obtained as the knee was flexed. The ultrasound images were used to determine muscle slack length (see Data Analysis). The effect of contract-short conditioning was calculated by subtracting the muscle slack length obtained in the contract-short condition from the muscle slack length obtained in the reference condition.
Experiment 1 investigated the effect of the intensity of the conditioning contraction. Experiment 2 investigated how long the effect of contract-short conditioning lasted. Experiment 3 investigated the effect of different amplitude stretches applied after the contract-short conditioning. In each of the three experiments, three trials of each condition were recorded. There was a 2-min rest between trials. The order of conditions was randomized across experimental conditions and participants.
Experiment 1: Effect of contraction intensity.
In this experiment, participants performed conditioning contractions of different contraction intensities (Fig. 1A). Contraction intensity was measured with EMG. To determine maximum vastus lateralis muscle EMG amplitude, participants performed three maximum voluntary knee extension contractions with the knee in an extended position. A 60-s rest period was provided between contractions. The EMG signal from the pair of electrodes over the distal part of the vastus lateralis muscle was digitally rectified and smoothed (0.05-s time constant), and the maximum EMG amplitude across all three contractions was determined.
For the reference condition, participants remained relaxed with the knee in an extended position for the 30 s before measurement of muscle slack length. The contract-short conditions were identical except that the participant performed a 2- to 3-s voluntary isometric knee extension contraction to target intensities of either 5, 25, 50, or 100% of the maximum EMG amplitude [i.e., 5, 25, 50, or 100% of a maximal voluntary contraction (MVC)] shortly after being put in the extended knee position. Participants were provided with visual feedback of the smoothed and rectified EMG and target EMG level during the contractions.
Experiment 2: Effect of time.
In this experiment, the effect of contract-short conditioning was assessed at different times after the conditioning contraction (Fig. 1B). For the reference conditions, the knee remained in the extended position for either 5, 60, 180, or 300 s before measurement of muscle slack length. The contract-short conditions were identical except that the participant performed a 2- to 3-s maximal voluntary isometric knee extension contraction shortly after being put in the extended knee position.
Experiment 3: Effect of passive stretch amplitude.
In this experiment, slow stretches (lengthening-shortening cycles) of different amplitudes were applied to the relaxed muscle after the contract-short conditioning (Fig. 1C). For the reference condition, the knee was extended and remained in this position for 60 s before measurement of muscle slack length. For each of the four contract-short conditions, the knee was extended, and then participants performed a 2- to 3-s maximal voluntary isometric knee extension. The knee then stayed in this position (i.e., it was passively flexed and extended by 0°) or it was passively flexed 5°, 30°, or 90° before being returned to the extended knee position. The timing of these movements was adjusted so that the peak of the stretch occurred at the same time regardless of the amplitude of the stretch. Slack length was always measured 60 s after reaching the extended position.
Data Analysis
The ultrasound image sequences obtained from the two transducers were stitched together to provide a composite image sequence with a 110-mm field of view. The composite images had an 18-mm gap between the two transducers. Muscle fascicles were tracked on the composite image sequence using a semiautomated procedure (9). There is little between-fascicle variability in slack lengths (10), so just one clearly visible muscle fascicle from each image sequence was tracked. Tracking involved identifying the location of the proximal and distal ends of the muscle fascicle in each frame of the image sequence. Muscle fascicle length was defined as the distance between the proximal and distal ends of the muscle fascicle. Fascicle length was plotted as a function of knee angle.
A semiautomated algorithm determined the muscle slack length (39). The algorithm was supervised by two investigators who were blind to experimental condition. If either assessor thought the slack length selected by the algorithm differed by >5° from the slack length selected by visual inspection, the trial was flagged for manual slack length identification. This occurred in 9% of trials from experiment 1, 7% of trials from experiment 2, and 6% of trials from experiment 3.
Preliminary analysis of data from experiment 1 indicated that even the weakest conditioning contraction (5% MVC) had a large effect on muscle slack length. This was surprising and raised concerns that any level of muscle activity, regardless of size, would influence muscle slack length. Because EMG signals were monitored during the experiment, no trials displayed muscle activity >1% of maximum EMG amplitude. Nevertheless, given the preliminary results from experiment 1, we reinspected raw vastus lateralis EMG signals from all trials from all three experiments, blinded to experimental conditions, and excluded trials with any evidence of contraction (which was as little as 4 motor unit action potentials from a single motor unit). Although this process excluded dozens of trials, especially trials in experiment 2 where participants had to remain relaxed for as long as 300 s, it ensured that participants were, as far as possible, completely relaxed before and during the measurement of muscle slack length.
Statistical Analysis
An estimation approach was used to quantify the effects of passive stretch amplitude, contraction intensity, and time on the history dependence of muscle slack length (4). Each participant performed multiple trials of each condition in each experiment. To ensure that the assumption of independence of observations was met, the unit of observation for all statistical analyses was the mean slack length for each participant for each condition. Ninety-five percent confidence intervals were computed for the mean (across participants) of each condition using the relevant t distribution. The Python programming language (Python Software Foundation, version 3.7) was used for statistical analyses except where otherwise indicated.
RESULTS
The number of trials and excluded trials, as well as reasons for exclusion, are presented in Table 1.
Table 1.
Number of total, excluded, and analyzed trials
| Excluded Trials |
|||||
|---|---|---|---|---|---|
| Experiment No. | Total Trials | Contraction on ultrasound | Ultrasound not trackable | EMG activity | Trials Analyzed |
| 1 | 150 | 2 | 5 | 14 | 129 |
| 2 | 240 | 3 | 6 | 57 | 176 |
| 3 | 150 | 1 | 5 | 11 | 133 |
EMG, electromyography.
Experiment 1: Effect of contraction intensity
As expected, contract-short conditioning reduced muscle slack length. The effect of contraction intensity on muscle slack length is shown in Fig. 2. Over the range of contraction intensities from 5 to 100% MVC, the effect of contract-short conditioning was, as far as could be discerned, independent of contraction intensity [5% MVC: 9.0° (95% CI 7.1–10.9]; 25% MVC: 8.7° (7.3–10.2); 50% MVC: 10.2° (8.5–11.6); 100% MVC: 10.2° (8.5–11.9)].
Fig. 2.
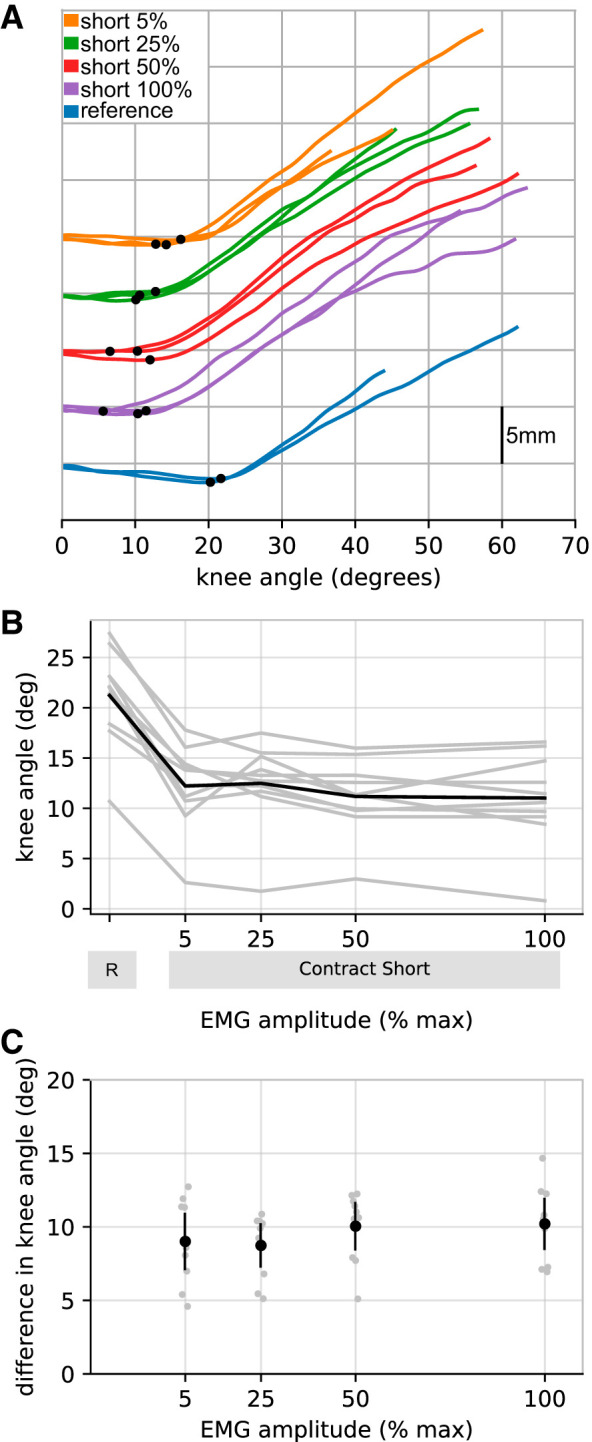
Experiment 1. Effect of contraction intensity. A: raw data (all trials from a single participant) showing how muscle slack length was determined. A different color has been used for each experimental condition. The vertical axis is muscle fascicle length, measured from composite ultrasound image sequences. For clarity, initial fascicle lengths have been offset vertically for each experimental condition. Muscle slack length is represented by the knee angle at which fascicle length first starts to increase, indicated by a black dot on each trace. B: mean muscle slack lengths for each participant (n = 10, gray lines) for the reference (R) condition and the four contract-short conditions [5, 25, 50, and 100% maximum electromyography (EMG) amplitude]. The black line joins the means of the participants’ means. C: difference in muscle slack lengths between the reference condition and each of the contract-short conditions. Individual participant means (gray circles) are shown, as are the means of participants’ means (black circles with 95% confidence intervals).
Experiment 2: Effect of time
Figure 3 shows how the effect of a conditioning contraction diminishes over time. The largest reduction in muscle slack length [16.1° (14.0–18.1)] was observed with the shortest measured delay between the conditioning contraction and the measurement of slack length (5 s). Smaller effects were observed after a delay of 60 s (10.8° [8.61–13.1]). This effect was maintained at 180 [10.6° (8.3–12.9)] and 300 [9.8° (8.0–11.6)] s.
Fig. 3.
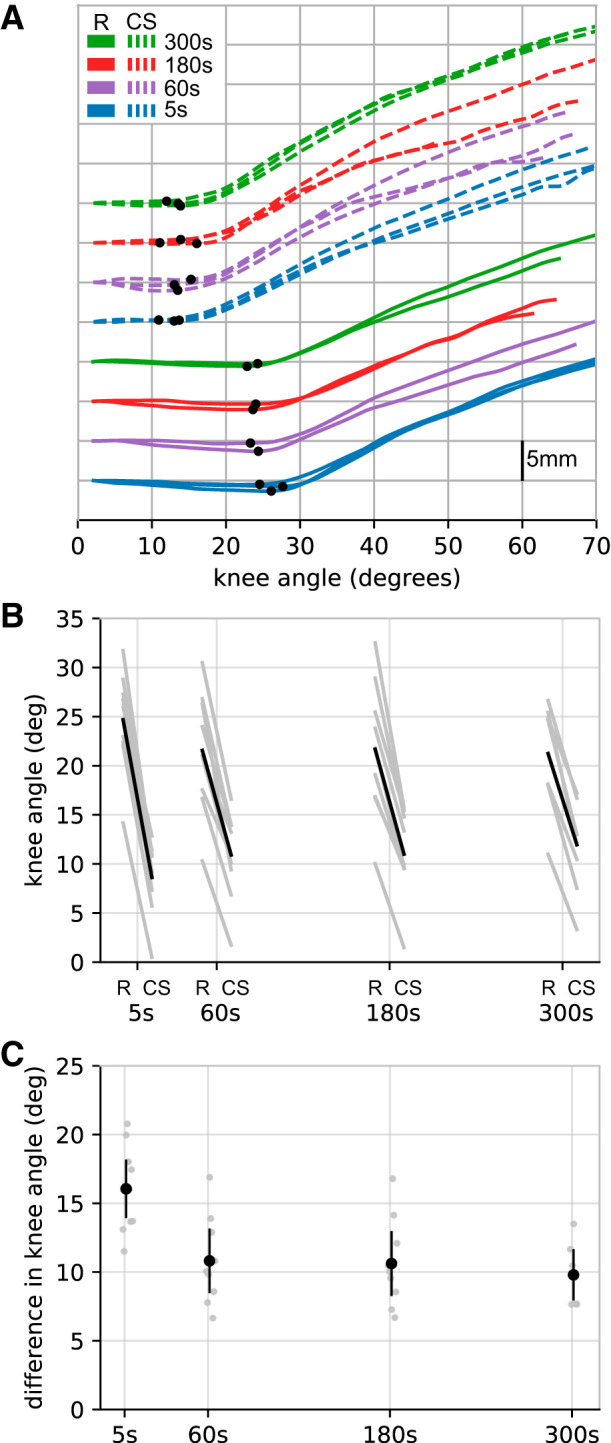
Experiment 2. Effect of time. A: raw data (all trials from a single participant). For clarity, initial fascicle lengths have been offset vertically for each experimental condition. A different color is used for each experimental condition. Solid lines refer to the reference condition (R), and dashed lines refer to the contract-short (CS) condition. B: mean muscle slack lengths for each participant (n = 10, gray lines) for the reference and contract-short condition after the four delays (5, 60, 180, and 300 s). The black line is the mean of the participants’ means. C: difference in muscle slack lengths between the reference condition and the corresponding contract-short condition for each delay. Individual participant means (gray circles) are shown, as are the means of participants’ means (black circles with 95% confidence intervals).
Because the effect of contract-short conditioning was measured as the difference in muscle slack lengths observed in the reference and contract-short conditions, the decay in effect of contract-short conditioning between 5 and 60 s after the conditioning contraction could be caused by either an increase in muscle slack length in the contract-short condition or a decrease in muscle slack length in the reference condition. In fact there was both an increase in muscle slack length in the contract-short condition [mean 2.2° (1.2–3.2)] and a decrease in muscle slack length in the reference condition [2.9° (1.4–4.4)] between 5 and 60 s.
Experiment 3: Effect of Stretch Amplitude
Figure 4 shows that the effect of contract-short conditioning on muscle slack length is reduced by a slow stretch and release delivered after the conditioning contraction. If no stretch was applied after the conditioning contraction, contract-short conditioning reduced muscle slack length by 9.9° (8.0–11.8) compared with the reference condition. Application of a small stretch by flexing and extending the knee 5° after the conditioning contraction appeared to reduce the mean effect of contract-short conditioning by 1.5° (−0.6−3.6) to 8.2° (7.3–9.6) although the confidence intervals do not completely preclude the possibility that the 5° stretch had no effect. A larger stretch, flexing and extending the knee 30°, greatly reduced the effect of contract-short conditioning [by 6.1° (4.6–7.5) to 3.9° (2.7–5.0)]. Flexing and extending the knee 90° reduced the effect of contract-short conditioning even more [by 9.6° (8.4–10.8)], enough to completely abolish the effect of contract-short conditioning (to 0.4° (−1.3 to 2.0)].
Fig. 4.
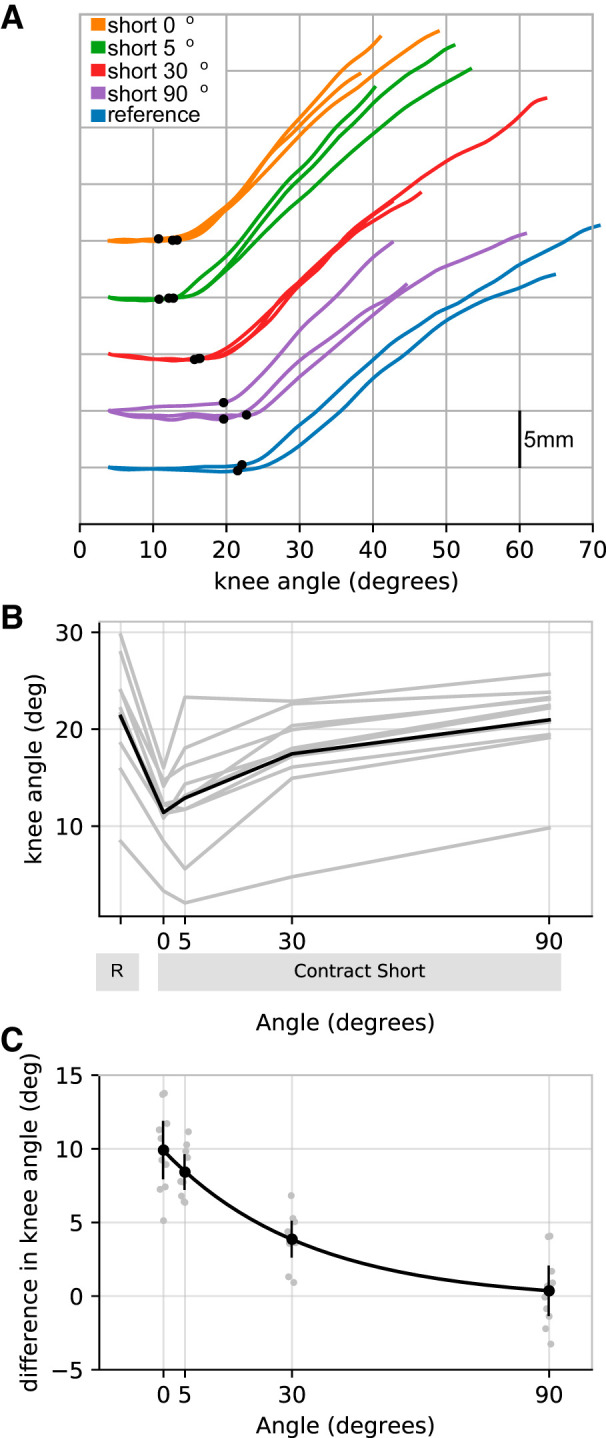
Experiment 3. Effect of stretch amplitude. A: raw data (all trials from a single participant). For clarity, initial fascicle lengths have been offset vertically for each experimental condition. B: mean muscle slack lengths for each participant (n = 10, gray lines) for the reference (R) and contract-short condition after stretches of 0, 5, 30, or 90°. The black line is the mean of the participants’ means. C: difference in muscle slack lengths between the reference condition and the corresponding contract-short condition for each stretch amplitude. Individual participant means (gray circles) are shown, as are the means of participants’ means (black circles with 95% confidence intervals). The black line is an exponential fit.
We evaluated whether the effect of stretch amplitude on the effect of contract-short conditioning was better fit with a linear or exponential model. This analysis was conducted by comparing the Bayesian information criterion of linear and exponential models using Stata 16 (StatCorp, College Station, TX). The exponential model (effect of contract-short conditioning = 9.91 × e−stretch amplitude in degrees/31) provided a better fit than the linear model. This exponential model implies that (every) 21.5° (14.3–28.7) of stretch reduces the effect of contract-short conditioning by a (further) one-half.
DISCUSSION
These experiments confirm the findings from our previous study which showed that, when compared with the same reference condition as used here, contract-short conditioning reduced the slack length of the human vastus lateralis muscle 60 s after the conditioning contraction by a mean of 9° (39). In experiments 2 and 3, which also measured muscle slack length 60 s after the conditioning contraction, contract-short conditioning reduced muscle slack length by means of 11° and 10°, respectively.
The three experiments reported here further characterize the effects of contract-short conditioning on muscle slack length. The first experiment showed that the effect of contract-short conditioning on muscle slack length is independent of the intensity of the conditioning contraction over the range of contraction intensities from 5 to 100% MVC. The second experiment showed that the effect of contract-short conditioning on muscle slack length is long lasting; the effect decays by about one-third in the first minute and then remains nearly constant until at least 5 min after the conditioning contraction. The third experiment showed that the effect of contract-short conditioning on muscle slack length can be partly or completely reversed by stretching the relaxed muscle after the conditioning contraction. The extent of the reversal depends nonlinearly on the amplitude of the stretch.
We previously showed that the contract-short conditioning protocol does not reduce muscle slack length if, instead of contracting the muscle at short lengths, the muscle remains relaxed (39). That finding is corroborated by experiment 2, which shows contract short-conditioning reduces muscle slack length compared with a reference condition in which the muscle is subjected to the same changes in length but does not contract. Thus the reduction in slack length is mediated by contraction-related events. This suggests, but does not definitively prove, that the mechanism that reduces muscle slack length occurs inside muscle fibers. If confirmed, that would imply muscle slack length is determined by intracellular structures, at least after contract-short conditioning.
What is the mechanism by which contract-short conditioning reduces muscle slack length? Two possible intracellular mechanisms involve titin filaments. Herzog and colleagues have hypothesized that (eccentric) muscle contraction at short lengths causes titin to bind with actin (15). Binding of titin to actin would reduce the length of the free (untethered) part of the titin molecule, which could reduce muscle slack length. Another possibility is that contraction at short muscle lengths could cause sustained folding of titin (37). Titin undergoes history-dependent folding and unfolding at physiological forces (28). The present experiments do not allow us to test these or other mechanisms.
Effect of Contraction Intensity
Experiment 1 showed that contract-short conditioning with weak contractions (5% MVC) produces similar reductions in muscle slack length to contract-short conditioning with maximal voluntary contractions.
In weak conditioning contractions, some motor units are recruited but others are not. Only muscle fibers from recruited motor units actively generate tension. If, as suggested above, the mechanism that reduces muscle slack length is mediated by contraction-related events inside muscle fibers, the reduction in slack observed after weak contractions would have to be mediated by events that occur only in those fibers activated during weak contractions.
Stronger conditioning contractions recruit more motor units but do not produce greater reductions in muscle slack length. There are two possible explanations. One explanation is that the mechanism by which the first-recruited muscle fibers reduce muscle slack length (binding of titin to actin, folding of titin, or some other mechanism) does not occur in later-recruited muscle fibers. A more plausible explanation is that the mechanism does occur in later-recruited muscle fibers but does not reduce muscle slack length any more than has already been achieved by fibers recruited at lower-contraction intensities.
Vastus lateralis muscle fibers, like the muscle fibers in most skeletal muscles, are arranged in series with elastic aponeuroses and tendons. As a consequence, muscle fibers shorten during isometric (fixed-end) contractions, and they shorten more during high-intensity isometric contractions than during low-intensity isometric contractions. Thus higher-intensity conditioning contractions cause the muscle to contract at shorter muscle fiber lengths. Our previous study had shown that the effect of conditioning contractions depends on the length at which the conditioning contractions are performed: contract-short conditioning reduced muscle slack length but contract-long conditioning does not (39). For that reason, we were surprised that contract-short conditioning with maximal contractions did not reduce muscle slack length more than contract-short conditioning with weak contractions. One explanation could be that most fiber shortening occurs at low contraction intensities, as it does in other muscles tested at longer lengths (11, 19), and there is not enough further shortening at higher-contraction intensities to create detectably greater effects of high-intensity contract-short conditioning. Another explanation could be that, while muscle fibers shorten during conditioning contractions, they subsequently lengthen when the contraction ceases (7). Thus it could be that high-intensity conditioning contractions produce greater reductions in muscle slack length during the contraction, but those greater reductions are subsequently abolished by the greater lengthening of the muscle fibers that occurs when the contraction ceases. This explanation is broadly consistent with the findings of the third experiment, which suggested that even quite a small (passive) stretch applied to muscle fibers after the conditioning contraction partly reverses the effect of the conditioning contraction.
The active lengthening of muscle fibers during the relaxation phase of an isometric contraction might be expected to cause passive force enhancement and reduce (rather than increase) slack length. This possibility was examined by Raiteri and Hahn (35). They found no evidence that fascicle lengthening in the relaxation phase of a fixed-end contraction caused passive force enhancement but did not rule out that possibility because of concerns they may not have had the resolution to detect such effects.
The standard protocol used to demonstrate passive force enhancement involves conditioning the muscle with a lengthening contraction. Passive tension at the final stretched length is higher after conditioning with a lengthening contraction than the passive tension observed at the same length after passive lengthening or after an isometric contraction at that length (12). In the absence of experimental evidence to the contrary, we hypothesize that muscle contraction of any type (concentric, isometric, or eccentric) can increase passive tension and reduce slack length, provided the contraction is at short muscle lengths. Furthermore, we hypothesize that large-amplitude stretches, whether passive or active, reduce passive force enhancement and increase slack length. If this hypothesis were true, large-amplitude lengthening contractions would be associated with greater passive force enhancement because the contraction was initiated at shorter lengths, despite, not because of, the large-amplitude stretch. This interpretation is consistent with the findings of Hagbarth and colleagues (8) who showed a “stiffening effect of a resisted shortening voluntary contraction” (see their Fig. 4). Moreover, this interpretation can be explained by invoking relatively simple mechanisms (e.g., contraction at short lengths causes formation of bonds between titin and actin at short lengths; passive or active stretch breaks those bonds); it is not necessary to identify a mechanism that is initiated by lengthening contractions but not isometric contractions, nor is it necessary to identify a mechanism that is initiated by active stretch but reversed by passive stretch. It should be possible to test this hypothesis with experiments on isolated muscle or muscle fibers, and perhaps even with experiments on human muscles in vivo.
Effect of Time
Experiment 2 showed that the effect of contract-short conditioning on muscle slack length declined by about one-third in the period between 5 and 60 s after the conditioning contraction. The effect then remained nearly constant until at least 5 min after the conditioning contraction.
We defined the effect of contract-short conditioning on muscle slack length as the difference between the muscle slack lengths observed after the contract-short and reference conditions. The reduction in the effect of contract-short conditioning between 5 and 60 s after the conditioning contraction was caused by both a 2° increase in muscle slack length in the contract-short condition and a 3° reduction in slack length in the reference condition. The small increase in muscle slack length observed in the period between 5 and 60 s after the conditioning contraction in the contract-short condition could be caused by the breaking of bonds between titin and actin, or by unfolding of titin filaments, or some other mechanism. Whatever the mechanism, it does not completely reverse the effect of contract-short conditioning: some of the effect of contract-short condition persists for at least 5 min. The small reduction in muscle slack length observed in the period between 5 and 60 s after the conditioning contraction in the reference condition could be due the progressive formation of bonds between actin and titin, progressive folding of titin, or another mechanism.
Effect of Passive Stretch
Experiment 3 shows that the effect of contract-short conditioning on slack length is reduced if, after the conditioning contraction, the muscle is slowly stretched and released. Larger-amplitude stretch releases produce greater reductions in the effect of contract-short conditioning (i.e., they produce greater increases in slack length).
When the muscle was subjected to contract-short conditioning but then not stretched, the mean muscle slack length was 11.4°. In this condition, no individual participant had a slack length >16° (see Fig. 2B). Therefore, when the muscle was subjected to contract-short conditioning and then stretched 30°, the stretch must have taken up the slack in the vastus lateralis muscle and substantially lengthened the muscle in all participants. Assuming a moment arm at the knee of ~48 mm (2, 41), 14° of knee rotation beyond the muscle’s slack length would lengthen the muscle by ~12 mm beyond its slack length. Despite this large stretch, over one-third of the effect of the preceding conditioning contraction persisted 60 s after the conditioning contraction.
This implies one of two things: either the 30° stretch was insufficient to completely reverse the mechanism by which contract-short conditioning reduced slack length, or the 30° stretch completely reversed the mechanism but it then recovered in the 23 s period after the stretch when the muscle was resting at its shortest in vivo length. If this second possibility were true, the effect of stretch amplitude would be confounded by the duration for which the muscle rested at short lengths. The time spent resting at short lengths after the 0°, 5°, 30°, and 90° stretch release cycles was 59, 28, 23, and 11 s, respectively. It could be that muscle slack length was shorter after the smaller-amplitude stretches because there was more time for recovery of the mechanism that reduced muscle slack length after the smaller stretches. Having considered this second possibility, we feel it is unlikely. Experiment 2 showed that, when in the reference condition the muscle rested at a short length for 60 s, muscle slack length was on average 2.9° shorter than when the muscle rested at a short length for 5 s. This effect of resting at short lengths is small compared with the 8.1° difference between the mean muscle slack lengths observed 28 s after the 5° stretch and 11 s after the 90° stretch. Hence, it seems unlikely that differences in time spent at short muscle lengths could account for the observed differences in muscle slack lengths following different-amplitude stretches.
Comparison with Other History-Dependent Phenomena
Physiologists and biomechanists have long understood that the slack lengths of many soft tissues are history dependent, at least under experimental conditions. When tendons or ligaments are subjected to repeated stretch-release cycles, slack length is frequently observed to increase, especially between the first and second stretch cycles. Slack length may slowly decrease when the tissue is returned to short lengths (20, 36). To our knowledge, these direct measurements of the history dependence of slack lengths have always been made on isolated tissue preparations in which small departures from truly physiological patterns of loading could cause low-level damage to the specimen. That cannot explain the history-dependent behaviors observed in the present experiment because the present experiment was conducted using noninvasive methods in vivo.
There are also reports of history dependence of slack length in relaxed skeletal muscle, including perfused mammalian skeletal muscles (16, 29). Muscles potentially exhibit more complex history-dependent behavior than other soft tissues because they have the ability to actively contract on short time scales, and contraction history can also influence muscle slack length. A number of references in the older muscle literature report that muscle contraction can “reset” muscle slack length in isolated muscle preparations (e.g., 1, 29 and references therein). Studies conducted on human muscles in vivo show similar effects of muscle contraction on resting joint angle and resting joint stiffness (8, 22). Using the same methods as used here, we previously demonstrated that isometric contractions at short lengths reduced the slack length of the human vastus lateralis muscle in vivo (39).
In some ways, the effect of contract-short conditioning on muscle slack length is similar to passive force enhancement. The effects of both contract-short conditioning and passive force enhancement are induced by muscle contraction, are more evident when the contraction is initiated at short muscle lengths (although passive force enhancement is only evident at final lengths greater than slack length), produce long-lasting effects, and can be abolished by a large-amplitude passive movement (13, 14, 18, 26). These similarities suggest the two phenomena could share the same molecular mechanisms.
The effect of contract-short conditioning on muscle slack length also shares similarities with muscle thixotropy. One way that muscle thixotropy manifests is that, when a relaxed muscle is held at a fixed length, its passive resistance to small-amplitude perturbations (i.e., its short-range stiffness; see Ref. 17) slowly increases over time. In human finger extensor muscles in vivo, stiffness increases at an exponentially decreasing rate, doubling in 1–3 min (25). In the reference condition of experiment 2, muscle slack length decreased by 2.9° between the 5th and 60th s.
Functional Consequences
The effects of contract-short conditioning on muscle slack length are large and were consistently observed in all participants. However it is not known if these effects confer any functional advantage or are simply epiphenomena. It is conceivable that the effects of contract-short conditioning could be exploited to improve sporting performance and that contract-short conditioning has a role in muscle injury or in the development of cramp or muscle contractures. To our knowledge, these possibilities have not been explored.
Study Strengths and Limitations
A strength of the approach used here is that the experiments were conducted in vivo, so the muscles were tested under physiological conditions without disrupting the tissues. Even though the experiments were conducted in vivo, the methods permitted sensitive measurement of the slack length of a single muscle. The procedures have proven to be highly reproducible, generating very similar results across experiments. Large effects of contract-short conditioning were demonstrated even with very weak conditioning contractions, suggesting the phenomenon is not an artifact induced by movement accompanying intense muscle contractions.
Participants were asked to keep their muscles relaxed while muscle slack length was measured. Contract-short conditioning could increase the response of muscle spindles to stretch, which might make it difficult for participants to keep their muscles relaxed. However several lines of evidence suggest that the measurement of muscle slack length was not distorted by increased resting discharge of muscle spindles. First, fast stretches are required to induce stretch reflexes in the relaxed muscles of neurologically unimpaired subjects (33), whereas slow stretches (5°/s) were applied here. Second, the shape of the relationship between fascicle length and muscle length was very consistent (see Figs. 2A, 3A, and 4A), which would seem unlikely if the stretch elicited stretch reflexes. Third, reflex contraction of the muscle at short lengths would cause shortening of muscle fascicles. This is unlikely to be confused with the abrupt increase in fascicle length that characterizes muscle slack length. Last, particular care was taken to exclude from the analysis any trials in which the participant was not completely relaxed; this was done blind to experimental condition.
A weakness of our approach is that we infer slack lengths from kinematic data, without measuring muscle tension. It is difficult to measure very low tensions in a single human muscle in vivo because measurements of joint torque must be used as a surrogate for muscle tension. Joint torques reflect tension in multiple muscles and other soft tissues, so are difficult to interpret. However, it may be possible to measure shear modulus with ultrasound elastography and use shear modulus as a proxy for muscle tension (32). Shear-wave elastography has been used to measure the slack lengths of human muscles in vivo (21). Another limitation of the in vivo model is that the muscle is potentially subjected to boundary forces, including friction, because it is sandwiched between adjacent anatomical structures and connected to them by epimuscular connective tissue and neurovascular tracts (27). However, there is evidence both that there is little or no transmission of force from the adjacent rectus femoris muscle (6) and that the length of the rectus femoris muscle does not affect vastus lateralis slack length (42). Further reassurance is provided by the observation that similar phenomena to those described here have been observed in isolated muscles (1, 29).
Future Studies
It might be informative to replicate the current experiments using very low intensity conditioning contractions (<5% MVC). It would also be interesting to determine if there is evidence of further decay of the effect of conditioning contractions beyond 5 min and if the effect of fast stretches differs from the effect of slow stretches.
Reduced muscle preparations such as single-fiber preparations and myofibril preparations potentially provide powerful ways to explore the molecular mechanisms of physiological phenomena like contract-short conditioning. We hope that physiologists who conduct such studies will further explore the mechanisms by which contract-short conditioning influences mechanical properties of relaxed skeletal muscle.
GRANTS
The study was supported by the Australian National Health and Medical Research Council (NHMRC; Program Grant APP1055084). RDH is supported by an NHMRC research fellowship.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
M.E.H., P.W.S., and R.D.H. conceived and designed research; M.E.H., I.A., S.N.V., and R.D.H. performed experiments; M.E.H., I.A., S.N.V., J.D., P.W.S., and R.D.H. analyzed data; M.E.H., I.A., S.N.V., P.W.S., and R.D.H. interpreted results of experiments; M.E.H. prepared figures; M.E.H., I.A., S.N.V., and R.D.H. drafted manuscript; M.E.H., I.A., S.N.V., J.D., P.W.S., and R.D.H. edited and revised manuscript; M.E.H., I.A., S.N.V., J.D., P.W.S., and R.D.H. approved final version of manuscript.
ACKNOWLEDGMENTS
We thank Prof. Janet Taylor, Dr. Brent Raiteri and two anonymous reviewers for comments that improved the manuscript.
REFERENCES
- 1.Alexander RS, Johnson PD Jr. Muscle stretch and theories of contraction. Am J Physiol 208: 412–416, 1965. doi: 10.1152/ajplegacy.1965.208.3.412. [DOI] [PubMed] [Google Scholar]
- 2.Bakenecker P, Raiteri B, Hahn D. Patella tendon moment arm function considerations for human vastus lateralis force estimates. J Biomech 86: 225–231, 2019. doi: 10.1016/j.jbiomech.2019.01.042. [DOI] [PubMed] [Google Scholar]
- 3.Brynnel A, Hernandez Y, Kiss B, Lindqvist J, Adler M, Kolb J, van der Pijl R, Gohlke J, Strom J, Smith J, Ottenheijm C, Granzier HL. Downsizing the molecular spring of the giant protein titin reveals that skeletal muscle titin determines passive stiffness and drives longitudinal hypertrophy. eLife 7: e40532, 2018. doi: 10.7554/eLife.40532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Calin-Jageman RJ, Cumming G. Estimation for better inference in neuroscience. eNeuro 6: 6, 2019. doi: 10.1523/ENEURO.0205-19.2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Campbell KS, Lakie M. A cross-bridge mechanism can explain the thixotropic short-range elastic component of relaxed frog skeletal muscle. J Physiol 510: 941–962, 1998. doi: 10.1111/j.1469-7793.1998.941bj.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Freitas SR, Antunes A, Salmon P, Mendes B, Firmino T, Cruz-Montecinos C, Cerda M, Vaz JR. Does epimuscular myofascial force transmission occur between the human quadriceps muscles in vivo during passive stretching? J Biomech 83: 91–96, 2019. doi: 10.1016/j.jbiomech.2018.11.026. [DOI] [PubMed] [Google Scholar]
- 7.Griffiths RI. Shortening of muscle fibres during stretch of the active cat medial gastrocnemius muscle: the role of tendon compliance. J Physiol 436: 219–236, 1991. doi: 10.1113/jphysiol.1991.sp018547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hagbarth K-E, Hägglund JV, Nordin M, Wallin EU. Thixotropic behaviour of human finger flexor muscles with accompanying changes in spindle and reflex responses to stretch. J Physiol 368: 323–342, 1985. doi: 10.1113/jphysiol.1985.sp015860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Herbert RD, Clarke J, Kwah LK, Diong J, Martin J, Clarke EC, Bilston LE, Gandevia SC. In vivo passive mechanical behaviour of muscle fascicles and tendons in human gastrocnemius muscle-tendon units. J Physiol 589: 5257–5267, 2011. doi: 10.1113/jphysiol.2011.212175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Herbert RD, Héroux ME, Diong J, Bilston LE, Gandevia SC, Lichtwark GA. Changes in the length and three-dimensional orientation of muscle fascicles and aponeuroses with passive length changes in human gastrocnemius muscles. J Physiol 593: 441–455, 2015. doi: 10.1113/jphysiol.2014.279166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Héroux ME, Stubbs PW, Herbert RD. Behavior of human gastrocnemius muscle fascicles during ramped submaximal isometric contractions. Physiol Rep 4: e12947, 2016. doi: 10.14814/phy2.12947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Herzog W, Schachar R, Leonard TR. Characterization of the passive component of force enhancement following active stretching of skeletal muscle. J Exp Biol 206: 3635–3643, 2003. doi: 10.1242/jeb.00645. [DOI] [PubMed] [Google Scholar]
- 13.Herzog W. Passive force enhancement in striated muscle. J Appl Physiol (1985) 126: 1782–1789, 2019. doi: 10.1152/japplphysiol.00676.2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Herzog W, Leonard TR. Force enhancement following stretching of skeletal muscle: a new mechanism. J Exp Biol 205: 1275–1283, 2002. [DOI] [PubMed] [Google Scholar]
- 15.Herzog W, Powers K, Johnston K, Duvall M. A new paradigm for muscle contraction. Front Physiol 6: 174, 2015. doi: 10.3389/fphys.2015.00174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hill AV. Is relaxation an active process? Proc R Soc Lond B Biol Sci 136: 420–435, 1949. doi: 10.1098/rspb.1949.0034. [DOI] [PubMed] [Google Scholar]
- 17.Hill DK. Tension due to interaction between the sliding filaments in resting striated muscle. The effect of stimulation. J Physiol 199: 637–684, 1968. doi: 10.1113/jphysiol.1968.sp008672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hisey B, Leonard TR, Herzog W. Does residual force enhancement increase with increasing stretch magnitudes? J Biomech 42: 1488–1492, 2009. doi: 10.1016/j.jbiomech.2009.03.046. [DOI] [PubMed] [Google Scholar]
- 19.Hodges PW, Pengel LH, Herbert RD, Gandevia SC. Measurement of muscle contraction with ultrasound imaging. Muscle Nerve 27: 682–692, 2003. doi: 10.1002/mus.10375. [DOI] [PubMed] [Google Scholar]
- 20.Hubbard RP, Soutas-Little RW. Mechanical properties of human tendon and their age dependence. J Biomech Eng 106: 144–150, 1984. doi: 10.1115/1.3138471. [DOI] [PubMed] [Google Scholar]
- 21.Hug F, Lacourpaille L, Maïsetti O, Nordez A. Slack length of gastrocnemius medialis and Achilles tendon occurs at different ankle angles. J Biomech 46: 2534–2538, 2013. doi: 10.1016/j.jbiomech.2013.07.015. [DOI] [PubMed] [Google Scholar]
- 22.Jahnke MT, Proske U, Struppler A. Measurements of muscle stiffness, the electromyogram and activity in single muscle spindles of human flexor muscles following conditioning by passive stretch or contraction. Brain Res 493: 103–112, 1989. doi: 10.1016/0006-8993(89)91004-4. [DOI] [PubMed] [Google Scholar]
- 23.Kwah LK, Pinto RZ, Diong J, Herbert RD. Reliability and validity of ultrasound measurements of muscle fascicle length and pennation in humans: a systematic review. J Appl Physiol (1985) 114: 761–769, 2013. doi: 10.1152/japplphysiol.01430.2011. [DOI] [PubMed] [Google Scholar]
- 24.Lakie M, Campbell KS. Muscle thixotropy-where are we now? J Appl Physiol (1985) 126: 1790–1799, 2019. doi: 10.1152/japplphysiol.00788.2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lakie M, Robson LG. Thixotropic changes in human muscle stiffness and the effects of fatigue. Q J Exp Physiol 73: 487–500, 1988. doi: 10.1113/expphysiol.1988.sp003169. [DOI] [PubMed] [Google Scholar]
- 26.Lee H-D, Herzog W. Force enhancement following muscle stretch of electrically stimulated and voluntarily activated human adductor pollicis. J Physiol 545: 321–330, 2002. doi: 10.1113/jphysiol.2002.018010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Maas H. Significance of epimuscular myofascial force transmission under passive muscle conditions. J Appl Physiol (1985) 126: 1465–1473, 2019. doi: 10.1152/japplphysiol.00631.2018. [DOI] [PubMed] [Google Scholar]
- 28.Mártonfalvi Z, Bianco P, Linari M, Caremani M, Nagy A, Lombardi V, Kellermayer M. Low-force transitions in single titin molecules reflect a memory of contractile history. J Cell Sci 127: 858–870, 2014. doi: 10.1242/jcs.138461. [DOI] [PubMed] [Google Scholar]
- 29.McCarter RJM, Nabarro FR, Wyndham CH. Reversibility of the passive length-tension relation in mammalian skeletal muscle. Arch Int Physiol Biochim 79: 469–479, 1971. doi: 10.3109/13813457109085331. [DOI] [PubMed] [Google Scholar]
- 30.Meyer G, Lieber RL. Frog muscle fibers bear a larger fraction of passive muscle tension than mouse fibers. J Exp Biol 221: jeb.182089, 2018. doi: 10.1242/jeb.182089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Minajeva A, Kulke M, Fernandez JM, Linke WA. Unfolding of titin domains explains the viscoelastic behavior of skeletal myofibrils. Biophys J 80: 1442–1451, 2001. doi: 10.1016/S0006-3495(01)76116-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Nordez A, Hug F. Muscle shear elastic modulus measured using supersonic shear imaging is highly related to muscle activity level. J Appl Physiol (1985) 108: 1389–1394, 2010. doi: 10.1152/japplphysiol.01323.2009. [DOI] [PubMed] [Google Scholar]
- 33.Pisano F, Miscio G, Colombo R, Pinelli P. Quantitative evaluation of normal muscle tone. J Neurol Sci 135: 168–172, 1996. doi: 10.1016/0022-510X(95)00291-9. [DOI] [PubMed] [Google Scholar]
- 34.Powers JD, Bianco P, Pertici I, Reconditi M, Lombardi V, Piazzesi G. Contracting striated muscle has a dynamic I-band spring with an undamped stiffness 100 times larger than the passive stiffness. J Physiol 598: 331–345, 2020. doi: 10.1113/JP278713. [DOI] [PubMed] [Google Scholar]
- 35.Raiteri BJ, Hahn D. A reduction in compliance or activation level reduces residual force depression in human tibialis anterior. Acta Physiol (Oxf) 225: e13198, 2019. doi: 10.1111/apha.13198. [DOI] [PubMed] [Google Scholar]
- 36.Rigby BJ, Hirai N, Spikes JD, Eyring H. The mechanical properties of rat tail tendon. J Gen Physiol 43: 265–283, 1959. doi: 10.1085/jgp.43.2.265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Rivas-Pardo JA, Eckels EC, Popa I, Kosuri P, Linke WA, Fernández JM. Work done by titin protein folding assists muscle contraction. Cell Rep 14: 1339–1347, 2016. doi: 10.1016/j.celrep.2016.01.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Rivas-Pardo JA, Li Y, Mártonfalvi Z, Tapia-Rojo R, Unger A, Fernández-Trasancos Á, Herrero-Galán E, Velázquez-Carreras D, Fernández JM, Linke WA, Alegre-Cebollada J. A HaloTag-TEV genetic cassette for mechanical phenotyping of proteins from tissues. Nat Commun 11: 2060, 2020. doi: 10.1038/s41467-020-15465-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Stubbs PW, Walsh LD, D’Souza A, Héroux ME, Bolsterlee B, Gandevia SC, Herbert RD. History-dependence of muscle slack length following contraction and stretch in the human vastus lateralis. J Physiol 596: 2121–2129, 2018. doi: 10.1113/JP275527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Taylor DC, Dalton JD Jr, Seaber AV, Garrett WE Jr. Viscoelastic properties of muscle-tendon units. The biomechanical effects of stretching. Am J Sports Med 18: 300–309, 1990. doi: 10.1177/036354659001800314. [DOI] [PubMed] [Google Scholar]
- 41.Visser JJ, Hoogkamer JE, Bobbert MF, Huijing PA. Length and moment arm of human leg muscles as a function of knee and hip-joint angles. Eur J Appl Physiol Occup Physiol 61: 453–460, 1990. doi: 10.1007/BF00236067. [DOI] [PubMed] [Google Scholar]
- 42.Whitaker RM. No Evidence of Epimuscular Myofascial Force Transmission between the Human Rectus Femoris and Vastus Lateralis Muscles in Passive Conditions (MSc thesis). Amsterdam, The Netherlands: Vrije Universiteit, 2019. [DOI] [PubMed] [Google Scholar]
- 43.Whitehead NP, Gregory JE, Morgan DL, Proske U. Passive mechanical properties of the medial gastrocnemius muscle of the cat. J Physiol 536: 893–903, 2001. doi: 10.1111/j.1469-7793.2001.00893.x. [DOI] [PMC free article] [PubMed] [Google Scholar]