

Abstract
BRCA1-Associated Protein 1 (BAP1) is a ubiquitin carboxy-terminal hydrolase that has been established as a tumor suppressor, utilizing its deubiquitinating activity to regulate a number of processes including DNA damage repair, cell cycle control, chromatin modification, programmed cell death, and the immune response. Mutations in the BAP1 gene commonly result in a number of aggressive cancers; predominantly uveal melanoma, malignant mesothelioma, renal cell carcinoma, and cutaneous melanoma. Importantly, germline mutations in the BAP1 gene have been established as a novel tumor predisposition syndrome, conferring an increased risk of hereditary, early-onset cancers. Current treatment options for cancers with BAP1 alterations are limited to standard therapies. However, several therapeutic avenues have been proposed to specifically target BAP1 alterations in cancer. Molecularly targeted approaches include histone deacetylase inhibitors and EZH2 inhibitors to target the role of BAP1 in chromatin modification and transcriptional regulation, respectively. PARP inhibitors and platinum chemotherapy agents have the potential to target BAP1-altered tumors, due to the role of BAP1 in DNA damage repair. Lastly, emerging reports suggest that BAP1-alterations in cancer confer distinct immunogenic phenotypes that may be particularly susceptible to novel cancer immunotherapies. This review aims to present a concise and up to date report on the BAP1 gene in cancer, surveying its functional roles, characteristics and clinical manifestations. Furthermore, we highlight the established and emerging therapeutic options for BAP1-mutated cancers.
Keywords: BAP1, Cancer, Targeted Therapy, Immunotherapy
INTRODUCTION
BRCA1-Associated Protein 1 (BAP1), encoded by the BAP1 gene, was originally discovered in 1998 as a novel ubiquitin carboxy-terminal hydrolase, an enzyme responsible for removing ubiquitin from protein substrates.[1] BAP1 was initially shown to localize to the nucleus where its primary interaction was binding to BRCA1 and enhancing its tumor suppressive activity.[1] In subsequent years, numerous research groups have revealed that BAP1 acts independently as a tumor suppressor, utilizing its deubiquitinating activity to regulate proteins involved in DNA damage repair, cellular differentiation, chromatin modulation, cell cycle control, and cell proliferation.[2] Clinical reports have demonstrated that BAP1 is commonly lost or inactivated in a variety of cancers.[3] Most notably, recent studies have indicated that germline mutations of BAP1 confer a novel tumor predisposition syndrome, characterized by a high incidence of early-onset malignancies consisting of uveal melanoma, malignant mesothelioma, cutaneous melanoma, and several other cancers.[4]
To date, the treatment options available for tumors with BAP1 mutations are limited to standard therapies. Some targeted approaches such as HDAC inhibitors, EZH2 inhibitors, and PARP inhibitors have been suggested.[5] Additionally, with the rapid development of cancer immunotherapy in recent years, several studies have investigated the association between BAP1 alterations and immunogenic phenotypes, with some proposing that BAP1 abnormalities may confer increased sensitivity to immune-targeted therapies.[6] These focused approaches represent a growing body of research surrounding BAP1 and may have future implications for therapy in BAP1-mutated cancers.
In this report, we aim to present a concise review of the BAP1 gene in cancer, surveying its functional roles, characteristics, and the clinical manifestations that have been elucidated over the last two decades since its discovery. Furthermore, we will highlight a growing body of research on novel therapeutic approaches for BAP1-mutated cancers including molecularly targeted therapies and most recently, cancer immunotherapy.
STRUCTURE AND FUNCTION OF BAP1
BAP1 is a multifunctional tumor suppressor involved in chromatin remodeling, DNA damage response through its relationship with BRCA1, cell cycle control, regulated cell death, and the immune response.[7–12]
Molecular Characteristics
The BAP1 gene is located on the short (p) arm of chromosome 3 at position 21.1 (3p21.1) and spans approximately 9 kilobases. Within the gene, there are 17 exons encoding 729 amino acids that comprise the 90 kDA BAP1 protein (Figure 1).[2] The primary catalytic domain of the BAP1 protein is a ubiquitin carboxy-terminal hydrolase domain located at its N-terminal region.[1] This ubiquitin carboxy-terminal hydrolase activity allows BAP1 to enact its primary function of removing ubiquitin signals from protein substrates. While protein ubiquitination was originally known to be associated with targeting proteins for proteasome degradation, it has since been established as a complex multifunctional cell signaling system.[13] Therefore, BAP1 utilizes its deubiquitinating activity to alter ubiquitin signaling and affect a wide variety of cellular processes (Figure 2). Other domains on BAP1 include protein binding motifs for Host Cell Factor 1 (HCF1), BRCA1, ASXL1/2, FOXk1/2, and YY1, which indicate an array of processes that BAP1 is involved in regulating (Figures 1 and 2).[2] Finally, at its C-terminal region, BAP1 contains two nuclear localization signals that pinpoint where its primary functions occur.[14]
Figure 1. Functional domains of BRCA1-Associated Protein 1 (BAP1).
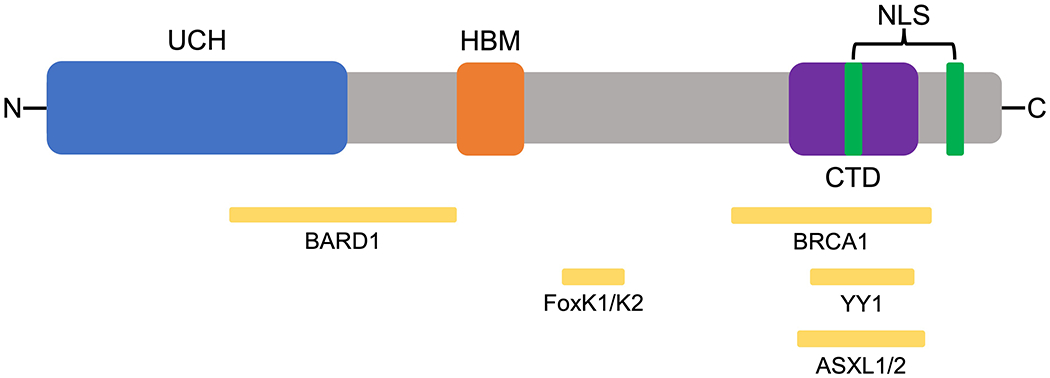
BAP1 is a 729 amino acid protein. Ubiquitin carboxy hydrolase (UCH) domain (1-250); BARD1-binding region (182-365); HCF1 binding (HBM) domain (365-385); FoxK1/K2 binding region (477-526); BRCA1 binding region (596-721); C-terminal binding domain (CTD) and ASKXL1/2 binding domain (635-693); Nuclear localization signals (NLS) (656-661 and 717-722); Ying Yang 1 (YY1) binding domain (642-686).[2]
Figure 2. Summary of the functional roles of BAP1.
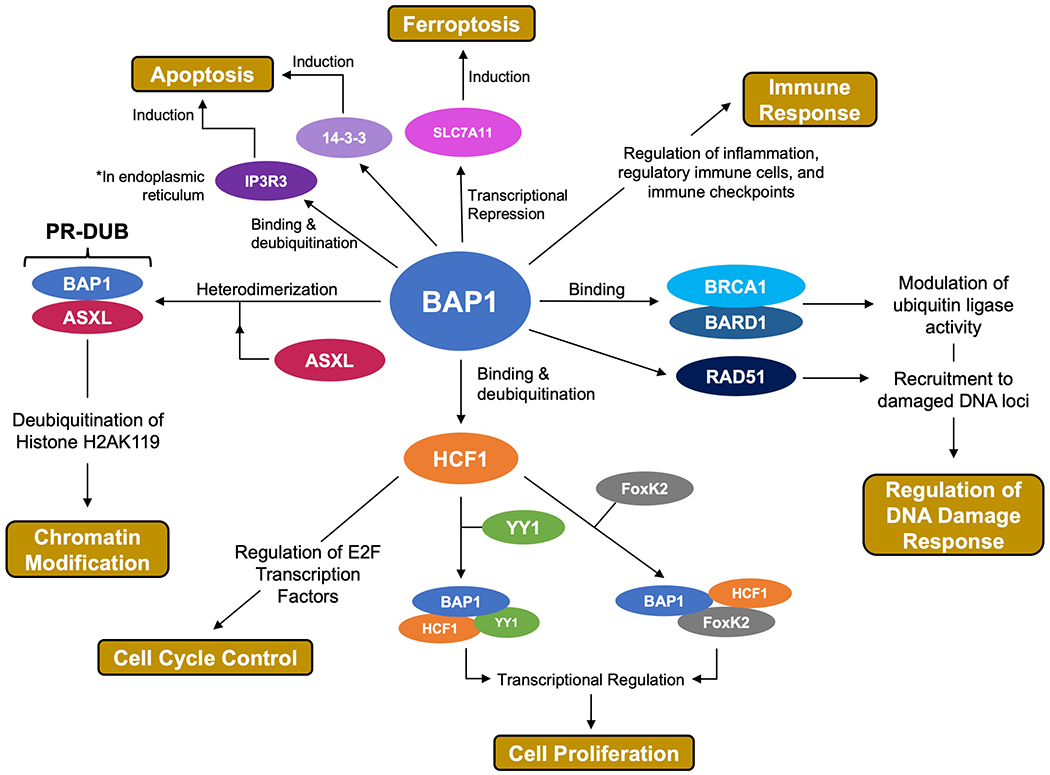
BAP1 regulates the DNA damage repair pathway through interactions with BRCA1, BARD1, and RAD51.[8,9,20] BAP1 interacts with host-cell control factor 1 (HCF1) in a number of processes involved in cell-cycle control and cell proliferation.[10,23,24] BAP1 binds to ASXL to form the PR-DUB complex, responsible for regulation of chromatin through Histone H2A deubiquitination.[7] BAP1 is associated in a number of regulated cell death pathways including apoptosis and ferroptosis.[11,25,26] BAP1 is implicated in immune regulation.[12,27,28]
Chromatin Modification
One of the fundamental biological roles of BAP1 is its involvement in epigenetic modification of chromatin. In 2010, Scheuermann et al. first discovered that in Drosophila, the Polycomb group (PcG) protein Calypso, a BAP1 homolog, binds to additional sex combs (ASX, homolog of ASXL1/2/3 in mammals) and forms the Polycomb repressive deubiquitinase (PR-DUB) complex.[7] The PR-DUB complex utilizes the catalytic activity of BAP1 to modulate chromatin by removing monoubiquitin from histone H2A at its lysine 119 residue (H2AK119ub1).[7] This function of PR-DUB, which is conserved in mammalian cells, was shown to directly oppose the function of another PcG protein, Polycomb repressive complex 1 (PRC1). PRC1 is an E3-ligase that facilitates the monoubiquitination of histone H2AK119, leading to chromatin modification and gene silencing.[15] Therefore, PR-DUB plays an antagonistic role to PRC1, by removing monoubiquitin from H2A, thus altering chromatin architecture and preventing PRC1-mediated gene silencing.[15] Polycomb group proteins are a well-known family of transcriptional regulators known to target genes involved in embryonic development, self-renewal, pluripotency, and differentiation.[16] Therefore, BAP1 and its role in PR-DUB deubiquitination of histone H2A directly regulates chromatin modification and subsequent transcription of these important PcG target genes. More broadly, histone H2AK119 ubiquitination is known to be a highly regulated modification that affects transcription initiation, elongation, and silencing of genes involved in many processes.[17] BAP1-mediated deubiquitination likely plays a widespread role in maintaining a delicate balance of H2A ubiquitination, regulating the chromatin architecture, and affecting expression of genes, many of which may be implicated in cancer pathways.
DNA Damage Response and Relationship to BRCA1
BAP1 stands for BRCA1-associated protein 1. The first study of BAP1 focused on its interaction with BRCA1, a well-known tumor suppressor responsible for coordinating the DNA damage response via homologous recombination (HR).[18] Further investigation has revealed that BAP1 in fact regulates the DNA damage response in a number of ways. Histone ubiquitination is known to play a significant role in the DNA damage response.[17] BAP1 was shown to bind the BRCA1/BARD1 complex, which is an E3 ubiquitin ligase that regulates the HR pathway of DNA damage repair.[8] BAP1 modulates the function of the BRCA1/BARD1 complex by binding to the RING finger domain on BARD1 and inhibiting its E3 ligase function. It also plays an antagonistic role by deubiquitinating sites of BRCA1/BARD1 ubiquitination.[8] Importantly, BAP1 inhibition by shRNA results in hypersensitivity of cells to ionizing radiation, suggesting that the interaction between BAP1 and BRCA1/BARD1 complex may in fact be necessary to mediate repair of DNA damage.[8] Other studies have shown that BAP1 is recruited to chromatin near DNA double stranded breaks and is necessary to recruit HR factors BRCA1 and RAD51, to these damaged loci in order to facilitate homologous recombination DNA repair.[19,20] Moreover, the BRCA1/BARD1 complex was recently shown to be directly involved in promoting RAD51-mediated homologous pairing.[9] Taken together, this evidence suggests that BAP1 plays an extensive role in coordinating BRCA1, BARD1 and RAD51 in the process of RAD51-mediated homologous recombination DNA repair, a process that maintains genome stability and is susceptible to dysregulation in cancer.
Cell-Cycle Control and Cell Proliferation
BAP1 is involved in binding and deubiquitinating Host Cell Factor 1 (HCF1), a transcriptional regulator involved in a number of processes that control the cell cycle and proliferation.[10] HCF1 is known to be involved in regulating transcription and promoting cell-cycle progression through the G1/S phase. It does so by recruiting histone modifying enzymes such as methyltransferases, acetyl transferases and deacetylases to modify the chromatin structure at promoters of key cell-cycle control transcription factors of the E2F family.[21] Studies have demonstrated that BAP1 function is necessary for the transition of cells from the G1 to S phase of the cell cycle, with knockdown of BAP1 causing cells to be arrested in the G1 phase.[22] Additionally, BAP1 was shown to bind to gene promoters targeted by E2F1 transcription factors and this localization is dependent on HCF1.[22] Furthermore, BAP1 is suggested to deubiquitinate H2AK119 at these sites, thus promoting transcriptional activation and proper progression through the G1/S phase of the cell cycle.[22] This evidence indicates that BAP1 plays a major part in regulating HCF1-mediated control of the cell cycle.
BAP1 was also shown to form a ternary complex with HCF1 and YY1, which control the expression of genes involved in cell proliferation.[23] BAP1 binds to the zinc fingers of YY1 through its coiled-coil motif and to HCF1 through its HBM domain, and this complex is recruited to target promoters, thus altering expression of genes such as COX7C, which is a component of the mitochondrial respiratory chain. More recently, another study demonstrated a different ternary complex between BAP1, HCF1, and FoxK2, a transcription factor involved in proper control of cell proliferation and cell cycle control.[24] This study showed that FoxK2 recruits BAP1 to its target genes leading to deubiquitination and gene silencing. In summary, it appears that BAP1 is involved in a number of multiprotein complexes that involve HCF1 and several other factors that are important for regulating cell-cycle control and proliferation.
Regulated Cell Death
Recent studies have suggested an emerging role of BAP1 in coordinating processes of regulated cell death. In 2017, Bononi et. al. presented a novel function of BAP1 in modulating type-3 inositol-1,4,5-triphosphate-receptor (IP3R3) and promoting apoptosis.[11] They showed that BAP1 localizes to the endoplasmic reticulum (ER), contrary to previous beliefs that it was purely nuclear-localized. Then BAP1 binds and deubiquitinates IP3R3 in the ER, leading to Calcium release into the cytosol and mitochondria, and initiation of apoptotic pathways. Moreover, when exposed to ionizing or UV radiation to impose DNA damage, BAP1+/− cells with reduced BAP1 expression, were resistant to executing apoptosis and exhibited increased survival and cellular transformation under genotoxic stress. Another study in 2018, showed that BAP1 interacts with the protein 14-3-3 in neuroblastoma cells causing the release of Bax and the decrease of pro-survival factors such as Bcl-2, to promote apoptosis via the intrinsic pathway.[25]
BAP1 is also involved in other forms of regulated cell death. Recently, Zhang et. al. demonstrated that BAP1 is involved in regulating the process of ferroptosis, a recently identified form of regulated cell death that is induced by metabolic stress from cystine reduction and increased reactive oxygen species.[26] They showed that BAP1 deubiquitinates H2Aub at the promoter of the cystine transporter SLC7A11. This epigenetic modification leads to reduced expression of SLC7A11, leading to decreased cystine uptake, increased lipid peroxide accumulation, and induction of ferroptosis. Moreover, BAP1 cancer-associated mutants show reduced repression of SLC7A11 and inability to promote ferroptosis, suggesting that BAP1 tumor suppression is mediated partly by induction of ferroptosis. In total, these recent studies linking BAP1 to multiple forms of regulated cell death, present additional mechanistic evidence for the involvement of BAP1 in cancer-associated pathways.
BAP1 Alterations and the Immune Response
A developing area of research on BAP1 has shifted focus towards its association with a number of immunological phenotypes, suggesting that BAP1 may play a role in regulating the immune response. In 2019, Shrestha et al. analyzed genomic, transcriptomic, and proteomic data from 19 patients with malignant peritoneal mesothelioma and found that BAP1-deficient tumors formed a unique molecular subtype characterized by an inflammatory tumor microenvironment, immune checkpoint activation, and distinct patterns of chromatin remodeling and DNA repair.[12] Additionally, BAP1-inactivated pleural mesothelioma tumors show significant increases in activated dendritic cells as well as slightly increased PD-L1 (CD274) expression, suggesting that BAP1-alterations promote an immunogenic phenotype in mesothelioma.
In uveal melanoma, Gezgin et al. (2017) found that loss of expression of BAP1 was associated with significantly increased infiltration of CD3+ T-cells and CD8+ T-cells, as well as increased expression of immune cell-attracting chemokines.[27] However, Figueiredo et. al recently reported evidence that despite these increases in tumor infiltrating lymphocytes, the loss of BAP1 expression in uveal melanoma is associated with an immunosuppressive microenvironment.[28] They demonstrated that decreased BAP1 expression correlates with upregulation of a number of immune regulatory gene networks that may alter immune cells to decrease anti-tumor immunity. Therefore, despite infiltration of lymphocytes and other immune cells in uveal melanoma, BAP1 loss potentially decreases cytotoxic effects and increases tumor immune-evasion.
In summary, these studies present emerging evidence that BAP1 loss is associated with distinct immunological phenotypes in cancer. Although the particular role of BAP1 in immune regulation remains unspecified, these tumor phenotypes may be of significance in the context of emerging cancer immunotherapies, discussed in further detail below.
BAP1 ALTERATIONS AND ASSOCIATED TUMORS
Given the widespread functional role of BAP1 in many cellular pathways implicated in cancer, it is not surprising that the BAP1 gene is altered in a variety of tumors.[3] BAP1 alterations observed in cancer are primarily inactivating mutations such as chromosomal deletions of the BAP1 gene locus at 3p21.1 or variants in the BAP1 gene itself, leading to loss of function.[3] BAP1 alterations occur in both germline and somatic form and are most commonly associated with uveal melanoma, malignant mesothelioma, cutaneous melanoma, renal cell carcinoma and several other malignancies (Tables 1 and 2).
Table 1.
Tumors Commonly Associated with BAP1 Germline Alterations (BAP1-Tumor Predisposition Syndrome (BAP1-TPDS)).[32]
| Patients with Germline BAP1 Alterations (Total, N=215) | Median Age of Onset (years) | |||
|---|---|---|---|---|
| Tumor Type | No. of tumors of specific class, N | % of patients with tumor of specific class | Tumors with Germline BAP1 Mutations | Sporadic Tumors |
| Uveal melanoma | 60 | 28% | 59 | 62 |
| Malignant mesothelioma | 48 | 22% | 46 | 74 |
| Cutaneous melanoma | 38 | 18% | 43 | 61 |
| Renal cell carcinoma | 20 | 9% | 51 | 64 |
| Basal cell carcinoma | 14 | 7% | 41 | 75 |
| BAP1-inactivated melanocytic tumor (BIMT)* | 40 | 75% | 31 | 24 |
Of the total 215 patients with BAP1-TPDS, 53 underwent the necessary total body skin examination to diagnose BIMT; of these, 40 patients (75%) had a positive diagnosis for one or more BIMTs.
Table 2.
Tumors with a High Incidence of Somatic BAP1 Alterations
| Tumors with Somatic BAP1 Mutations | |||||
|---|---|---|---|---|---|
| Tumor Type | Total no. of tumors, N | N | % | Comments | Ref. |
| Uveal melanoma | 142 | 43 | 30% | [37] | |
| 31 | 26 | 84% | Metastatic uveal melanoma | [38] | |
| Malignant mesothelioma | 42 | 20 | 48% | Most common mutation in malignant mesothelioma; BAP1 alterations are more common in pleural than in peritoneal mesotheliomas | [40] |
| 22 | 14 | 64% | [39] | ||
| Renal cell carcinoma | 445 | 39 | 9% | [42] | |
| Cutaneous melanoma | 60 | 3 | 5% | [31] | |
Germline Alterations of BAP1 and Cancer (Table 1)
Germline mutations of the BAP1 gene are inherited in an autosomal dominant pattern and have recently been reported by three independent research groups to constitute a novel tumor predisposition syndrome (BAP1-TPDS), conferring a high risk of hereditary cancers.[29–31] The cancers that BAP1-TPDS is most commonly associated with are uveal melanoma, malignant mesothelioma, cutaneous melanoma, and renal cell carcinoma.[32] Overall, roughly 85% of patients with BAP1-TPDS will develop ≥1 malignancies by a median age of 50 years old.[33] In addition to these cancers, BAP1-TPDS is commonly associated with a benign skin lesion, most recently termed a BAP1-inactivated melanocytic tumor (BIMT), that is commonly seen in BAP1-TPDS patients at an early age. This benign tumor has unique histomorphology co-occurring with loss of nuclear BAP1 expression and is discussed in further detail, below.
The most recent comprehensive studies of BAP1-TPDS account for a total of 215 patients from 87 families with germline BAP1 mutations (Table 1).[32,34] Of the predominant cancer types, uveal melanoma is the most commonly reported, being observed in 28% of patients with BAP1-TPDS. The median age of onset is 59 years (vs. 61 years, sporadic uveal melanoma)[32] and the tumors are generally more aggressive, have a higher rate of metastasis and lower overall survival.[34] Malignant mesothelioma is the second most common cancer identified with BAP1-TPDS, accounting for 22% of tumors. The median age of onset is 46 years, significantly earlier than that of sporadic malignant mesothelioma (74 years).[32] Interestingly, the survival rate for malignant mesothelioma with BAP1-TPDS has been reported to be 7-fold longer compared to patients with sporadic malignant mesothelioma.[35] Cutaneous melanoma is reported in 18% of tumors in BAP1-TPDS. The median age of onset is 43 years (vs. 61 years, sporadic cutaneous melanoma).[32] Renal cell carcinoma accounts for 9% of tumors in BAP1-TPDS and confers an earlier age of onset than the sporadic tumors (51 vs. 64 years)[32], more aggressive cancer, and a significantly reduced overall survival (31.2 months vs. 78.2 months).[32,34] Lastly, benign BAP1-inactivated melanocytic tumors (BIMTs) were present in 75% of the 53 BAP1-TPDS patients that reported undergoing a total-body skin examination, which was deemed a necessary criteria for diagnosis.[32] These BIMTs are diagnosed in BAP1-TPDS patients at a significantly earlier age (31 years) than the associated malignances (50 years).[32] In addition to these most commonly reported tumors with BAP1 germline mutations, there are ongoing studies within families that have also identified other associated tumors such as basal cell carcinoma, cholangiocarcinoma, meningioma, and several other cancers.[33,36]
Somatic Alterations of BAP1 and Cancer (Table 2)
Somatic BAP1 mutations appear in very similar types of tumors as seen in patients with germline mutations (Table 2). Recent reports identify somatic BAP1 mutations in approximately 30-40% of primary uveal melanoma.[36,37] More importantly, somatic BAP1 loss is found in 84% of metastatic uveal melanomas, suggesting a significant role of BAP1 in disease progression.[38] Indeed, BAP1 somatic mutations in uveal melanomas are associated with a much shorter time to metastasis and higher overall metastatic risk compared to both germline BAP1 mutation and mutation-negative uveal melanoma.[37] In malignant mesothelioma, several reports have indicated that BAP1 alterations are the most common somatic mutation; they occur in ~50% of tumors.[39,40] Similar to the situation with germline BAP1 mutations, malignant mesothelioma patients with somatic BAP1 mutations interestingly show significantly longer survival than BAP1 wild-type patients.[41] This common prognostic finding may suggest a particular protective role of BAP1 alterations in malignant mesothelioma. Somatic BAP1 mutations were found in 39 of the total 445 renal cell carcinoma cases (9%) in The Cancer Genome Atlas genomic database.[42] These tumors were associated with worse overall survival and disease-free survival in comparison to BAP1-wild type renal cell carcinoma. Lastly, somatic BAP1 mutations are estimated to be present in 5% of cutaneous melanoma.[31] Other cancers that show varying incidence of somatic BAP1 mutations are tumors of the prostate (0-6%), ovary (5%), large intestine (2-3%), breast (0-1%), and lung (0.4%).[3] In summary, tumor types that are associated with somatic BAP1 mutations show very similar patterns to those with germline BAP1 mutations. This may suggest common mechanisms of tumorigenesis and the potential for treatment strategies to target BAP1 in these frequently associated tumors.
SELECTED NEOPLASMS WITH HIGH PREVALENCE OF BAP1 ALTERATIONS
Uveal Melanoma
Uveal melanoma is a rare cancer that arises from melanocytes in the iris, ciliary body, or choroid of the eye.[43] The median age of diagnosis is in the early sixties, though it may be younger if there is a BAP1 germline alteration; however, the peak age range for diagnosis is between 70 and 79 years. Men are affected more commonly than women.
Early diagnosis and local treatment are critical, as survival correlates with primary tumor size. However, ~50% of patients will develop metastatic uveal melanoma, with survival of ~6–12 months from metastatic presentation.[44]
Over 80% of patients with uveal melanoma will have a BAP1 alteration (germline or somatic).[44] Overall, 1–2% of patients with uveal melanomas have germline BAP1 alterations;[45] ~28% of patients with germline BAP1 alterations will develop a uveal melanoma.[32] Importantly, GNAQ/GNA11 somatic mutations are also found in the majority of uveal melanoma tumors.[46]
Prognosis for patients with advanced uveal melanoma is very poor. Treatments include liver-directed therapies (e.g., hepatic arterial chemoembolization using multiple chemotherapy agents (fotemustine, BCNU, cisplatin)) and chemotherapy regimens adopted from cutaneous melanoma (e.g. dacarbazine, temozolomide, cisplatin, treosulfan, fotemustine, and assorted combinations), with response rates between 0% and 15%. Immunotherapy such as anti-PD1 and anti-CTLA4 agents, which have shown high success rates in cutaneous melanoma, demonstrate very low response rates (~5%) in uveal melanoma, perhaps because of their low tumor mutational burden.[44] However, combined anti-PD1 and anti-CTLA4 checkpoint blockade showed a response rate of ~15% with a median duration of response of about 25.5 months.[47]
Mesothelioma
Mesothelioma is a rare and insidious cancer with a long latency period (up to 40 years) after exposure to asbestos. Roughly 50% of patients with malignant pleural mesothelioma have a history of prior asbestos exposure. This neoplasm primarily occurs in the mesothelial surfaces of tissues in the pleura (~83% of cases) but can also occur in the peritoneum (~11%) and the tunica vaginalis (~1-2%).[40,48] Mesotheliomas are associated with poor prognosis, with a median survival of 12-16 months for advanced stage malignant pleural mesothelioma, and 12.5-31 months for peritoneal mesothelioma.[48,49]
Approximately 21-63% of malignant mesothelioma tumors harbor BAP1 alterations (germline or somatic).[40] BAP1 alterations tend to be more commonly associated with pleural than peritoneal mesothelioma. Overall, ~6% of patients with a malignant mesothelioma will have a BAP1 alteration that is germline in origin;[50] ~22% of patients with germline BAP1 alterations will develop a malignant mesothelioma.[32]
Patients with advanced or recurrent malignant mesothelioma commonly receive chemotherapy with platinum-based doublets. Cisplatin in combination with pemetrexed has been reported to improve clinical outcomes in malignant pleural mesothelioma when compared to cisplatin alone, with a response rate of 41.3% vs. 16.7% (P < 0.0001).[51] Immune checkpoint blockade has also been used. Scherpereel's team found that tumors shrank in approximately 18% of patients receiving nivolumab (anti-PD1) and 26% of those receiving nivolumab plus ipilimumab (anti-CTLA4) amongst 108 participants with malignant pleural mesothelioma.[52] The partial remission rate to pembrolizumab (anti-PD1 antibody) was similarly 24%.[53] In Japan, nivolumab has been approved as a salvage therapy in patients who do not respond to chemotherapy. In the United States, NCCN guidelines state that pembrolizumab (in programmed death ligand 1 (PD-L1) immunohistochemistry (IHC)-positive patients) and nivolumab, with or without ipilimumab (regardless of PD-L1 status), can be used as salvage therapies.[54]
BAP1-Inactivated Melanocytic Tumors (BIMTs)
BAP1-inactivated melanocytic tumors (BIMTs) were first described by Wiesner et. al. in 2011, who observed benign melanocytic tumors with unique histopathological features that were strongly associated with a nuclear loss of BAP1 expression.[31] These cutaneous lesions present as skin-colored, dome-shaped papules with an average size of 5 mm, appearing on the head, neck, trunk, and/or limbs. Histologically, they have a predominant composition of melanocytes that have a uniquely broad range of morphology including spitzoid, epithelioid, and rhabdoid cytologic features.[55] By definition, BIMTs show nuclear loss of BAP1 expression by immunohistochemistry. This can be a result of germline or somatic BAP1 mutations.[56] BIMTs can also occur sporadically with wild-type BAP1 status, and therefore diagnosis must be confirmed with immunohistochemistry of the lesion showing loss of BAP1 expression. One defining genomic feature of BIMTs is that they also present with a high frequency of BRAFV600E mutations.[56] This unique subset of histological and molecular features are reliable markers for diagnosis of BIMTs.
In general, BIMT lesions are described as having a benign course, but may have the potential to progress to malignancy, especially in the case of germline BAP1 mutations (i.e. BAP1 tumor predisposition syndrome).[31] Overall, the majority of BIMTs occur sporadically, but roughly 12% of BIMTs will be a result of a germline BAP1 mutation.[55,57] The exact prevalence of BIMTs with somatic BAP1 mutations is currently unclear. Germline BAP1 mutations are associated with a higher number of BIMT lesions per patient than sporadic BIMTs.[31] Furthermore, BIMTs are reported in ~75% of patients with BAP1 tumor predisposition syndrome (BAP1-TPDS), and the median age of diagnosis (32 years) is significantly earlier than the age-of-onset of the malignancies associated with the syndrome (50 years).[32] Therefore, the diagnosis of BIMTs is particularly pertinent in patients with BAP1-TPDS. Patients with multiple BIMTs and a family history of BAP1-TPDS-associated cancers should be tested for germline BAP1 mutations to potentially aid early diagnosis of BAP1-TPDS and preventative measures.[57]
BAP1 AND CANCER THERAPY
Given that BAP1 mutations are commonly associated with tumors that respond poorly to standard treatments, there have been a number of therapeutic strategies that aim to directly target the role of BAP1 in cancer (Table 3).
Table 3:
BAP1 Alterations and Cancer Therapy
| Tumor Phenotypes Related to BAP1-Alterations | Class of drugs that might impact BAP1 biologic effects | Comments |
|---|---|---|
| H2A ubiquitination mediated by BAP1 loss in uveal melanoma cells[58] | Histone deacetylase (HDAC) inhibitors | HDAC inhibitor vorinostat in phase 3 trial (placebo control arm) did not improve survival in malignant pleural mesothelioma (no selection for BAP1 alterations).[60] (NCT00128102) |
| EZH2 (enhancer of zeste 2 polycomb repressive complex 2) is upregulated in BAP1 deficient tumors[61,62] | EZH2 inhibitors | Two of 61 patients with malignant mesothelioma and concurrent BAP1 inactivation had a confirmed partial response with EZH2 inhibitor tazemostat.[63] (NCT02860286) |
| BAP1 regulates homologous recombination DNA damage repair[20] | Platinum agents | Overall survival was significantly longer for patients with malignant mesothelioma and loss-of-function mutations in BAP1 and DNA repair genes compared with patients with no such mutations who were treated with platinum chemotherapy.[65] |
| PARP inhibitors | Niraparib (NCT03207347) and olaparib (NCT03531840)—ongoing Phase II studies selecting for patients with BAP1 alterations and other DNA repair gene defects.[67,68] | |
| BAP1 alterations in mesothelioma correlate with tumor inflammation, infiltrating lymphocytes and increased PDL1 expression[6,12] | Anti-PD1/PDL1 compounds | Checkpoint blockade results in responses in ~20-30% of mesothelioma patients[52,53] Other chromatin remodeling genes similar to BAP1 have been associated with therapeutic response to Anti-PD1/PDL1 therapy.[74] |
| Loss of BAP1 expression in uveal melanoma shows upregulation of immunosuppressive genes[28] | Immune checkpoint inhibitors + Anti-CD38/Anti-CD74 compounds | Immune checkpoint inhibitors alone, result in 5-15% response rates in uveal melanoma.[44,47] Targeting other immunosuppressive markers such as CD38 and CD74, that are upregulated with BAP1-loss, may be viable adjuvants for immune checkpoint inhibitors.[28] |
| BAP1 mutations in renal cell carcinoma correlate with increased CCR5 expression and immunosuppression[70] | CCR5 inhibitors | CCR5 inhibition decreases immunosuppression and upregulates anti-tumor immunity in murine models of BAP1-mutant renal cell carcinoma.[70] |
HDAC inhibitors:
Histone deacetylase (HDAC) inhibitors have been shown to oppose the excess H2A ubiquitination mediated by BAP1 loss in uveal melanoma cells and inhibit tumor viability and growth.[58] Similarly, mesothelioma cells with loss of BAP1 show increased sensitivity to HDAC inhibitors.[59] Interestingly, a Phase 3 clinical trial of vorinostat, an HDAC inhibitor, in patients with malignant peritoneal mesothelioma showed only minimal improvement in overall survival compared to placebo.[60] However, this study did not select for BAP1 alterations or any other biomarkers. These findings suggest that future clinical studies of HDAC inhibitors may benefit from targeted selection of patients with BAP1 genomic alterations.
Targeting EZH2:
Another potential therapeutic strategy is targeting EZH2 (enhancer of zeste 2 polycomb repressive complex 2), an enhancer that is upregulated in BAP1 deficient tumors and is associated with poor prognosis.[61,62] EZH2 inhibitors have been proposed as a targeted treatment strategy in a number of cancer clinical trials. A phase 2 clinical trial of EZH2 inhibitor tazemetostat was recently conducted in malignant mesothelioma patients specifically harboring inactivating BAP1 alterations. The study reports that 2 of 61 patients had a confirmed partial response and the majority exhibited some level of disease control.[63] These results show support for the potential targeted use of EZH2 inhibitors in BAP1-mutated cancers.
Targeting DNA Damage Repair:
The role of BAP1 as a BRCA1-associated protein with its attendant role in regulating homologous recombination DNA damage repair suggests that targeting this pathway may have therapeutic value.[20] Therapies such as platinums as well as PARP inhibitors can be more effective in patients with BRCA-related DNA damage repair aberrations.
Cisplatin, a platinum agent, targets genomic DNA and causes widespread DNA lesions that block transcription and replication.[64] These platinum-induced DNA lesions strongly promote cell death when the DNA is unable to be repaired, such as in the case of mutations in BRCA1 or related DNA damage repair genes. In addition, selective forms of DNA damage such as nucleotide or base excision repair rely on repair pathways, in which poly-adenosyldiphosphate-ribose polymerase (PARP) plays a major role. Hence, patients with DNA damage repair gene defects, such as mutations in BAP1, may be more sensitive to platinum agents or to PARP inhibitors.
An evaluation of malignant mesotheliomas treated with platinum chemotherapy, revealed that patients who harbored inactivating mutations in BAP1 and other DNA repair genes experienced significantly longer overall survival in comparison to patients with no such mutations. This effect was observed in patients with pleural mesothelioma, but not for those with peritoneal mesothelioma, and remained significant after adjusting for gender and age at diagnosis.[65,66]
Other clinical trials are currently ongoing to investigate the efficacy of PARP1 inhibitors such as olaparib and niraparib in provoking synthetic lethality in various tumors.[67,68] These studies have selected for patients specifically with BAP1 mutations as well as several other DNA damage repair genes. Although there are currently no approved BAP1-specific therapies, these molecularly targeted strategies present viable options to target BAP1-mutated cancers.
Implications of BAP1 Alterations for Cancer Immunotherapy
Immunotherapies have recently demonstrated high efficacy in treating some aggressive tumors.[69] However, despite impressive durable responses in some patients, there is a high variability of response to these therapies and often serious side effects.[69] Thus, a growing body of research aims to identify genomic biomarkers that may help identify tumors that best respond to immunotherapy.[69]
BAP1 may be a potential candidate for such a predictive genomic biomarker. Tumors that respond to immunotherapy are commonly associated with increased tumor-infiltrating immune cells, PDL-1 expression, and inflammatory response.[69] As described above, BAP1 alterations have recently been associated with a number of similar immunological phenotypes in cancer.
In the case of malignant mesothelioma, tumors with BAP1-alterations are distinctly associated with an inflammatory tumor microenvironment, increased immune cell infiltration, and increased immune checkpoint activation.[6,12] This particular phenotype has the potential to be susceptible to immune-checkpoint inhibitors such as anti-PDL1 or anti-CTLA4 therapies.[6] To date, immune checkpoint inhibitor trials for mesothelioma have shown response rates ranging between 20-30%;[52,53] however, these trials do not consider genomic biomarker status. Therefore, BAP1 alterations should be considered as potential biomarkers in mesothelioma to predict response to immune checkpoint blockade.
Alternatively, BAP1 alterations, found in a large percentage of uveal melanomas, have been recently associated with activation of regulatory immune cells, thereby suppressing the immune response and enhancing tumor immune-evasion.[28] Thus far, immune checkpoint inhibitors such as anti-PDL1 or anti-CTLA4 therapies have shown fairly low efficacy in uveal melanoma with response rates ranging from 5-15%.[44,47] Figueiredo et. al. proposes that BAP1 loss in these tumors leads to upregulation of regulatory T-cells in the tumor microenvironment that suppress the cytotoxicity of effector T-cells.[28] They further suggest that inhibiting regulatory immune markers CD38 and CD74, which are upregulated with BAP1 loss, may be potential targets to reduce the immunosuppression and increase anti-tumor effects. These anti-CD38/anti-CD74 compounds can potentially be used as adjuvant therapies with immune checkpoint inhibition.[28]
Finally, in the case of renal cell carcinoma, Zhou et. al recently proposed a new therapeutic approach to BAP1-altered tumors by targeting CCR5.[70] CCR5 is an immune regulatory marker that shows consistently elevated expression in BAP1-mutant renal cell carcinomas.[70] They postulated that CCR5+ regulatory T cells are recruited in BAP1-mutated tumors and cause immune suppression and tumor immune-evasion. Subsequently, they demonstrated in murine tumor models that CCR5 blockade could induce anti-tumor immunity by decreasing regulatory immune suppression, thereby allowing the host immune system to inhibit tumor progression.[70] Thus, CCR5 blockade should be investigated as an alternate or adjuvant immunotherapy in the treatment of BAP1-mutated renal cell carcinoma.
These recent studies highlight that BAP1-alterations in cancer are associated with distinct immunological phenotypes that may be susceptible to various modalities of immunotherapy. However, more investigation into the underlying connection between BAP1-alterations and immune responses in cancer is necessary.
Chromatin Remodelers and Immunotherapy:
One potential mechanistic link between BAP1 and the immune response is through its role as a chromatin remodeler. Several proteins that participate in chromatin remodeling have been recently associated with immunotherapy response in cancer. In 2018, Pan et al. found that tumor cells that were exposed to experimental inactivation of genes in the SWI/SNF chromatin remodeling complex were more susceptible to T-cell mediated killing.[71] These genes include ARID2, PBRM1, and BRD7, three of the 29 genes that are known to code for the 15 subunits of the SWI/SNF complex. Genes of the SWI/SNF chromatin remodeling complex are mutated in over 20% of cancers, and several studies further suggest that tumors with deficiencies in these genes may show better responses to immunotherapy.[72] Loss of PBRM1 expression was shown to correlate with better response to immunotherapy in patients with clear cell renal carcinoma.[73] Similarly, ARID1A alterations were shown to predict better responses to checkpoint blockade immunotherapy across many cancers.[74]
BAP1 has an important connection with the SWI/SNF complex that may highlight a potential shared mechanism related to immunotherapy response. Recent studies have revealed an antagonistic relationship between the SWI/SNF complex and Polycomb repressive complexes (PRC1 and PRC2), the latter of which are involved in histone modification and gene silencing.[75,76] Mechanistically, the SWI/SNF complex was shown to reversibly evict PRC1 from chromatin, thereby reducing PRC1-mediated transcriptional silencing activity. As described above, one of the main functions of BAP1 is to deubiquitinate histone H2A, a direct antagonistic mechanism to the silencing function of PRC1, which ubiquitinates histone H2A. While BAP1 is not known to be a subunit of the SWI/SNF complex, it does share this antagonistic relationship to PRC1 as well as its own involvement in chromatin remodeling. Given the immunological phenotypes of tumor cells deficient in BAP1 and recent studies that showcase inactivated chromatin remodelers as potential markers for immunotherapy response, further investigation into an association between BAP1 alterations and the immune response in cancer is warranted.
CONCLUSIONS
BAP1 is a multifaceted tumor suppressor gene that regulates a wide variety of processes that are implicated in cancer. While the exact mechanisms of tumorigenesis remain uncertain, it is evident that BAP1 alterations are found in a common array of tumors, many of which show aggressive tumor phenotypes that respond poorly to standard treatments. Recent studies suggest that genomic selection of patients for clinical trials can be a useful approach for treatment.[77–84] In the case of malignancies harboring aberrant BAP1, directly targeting BAP1 functions using HDAC inhibitors and EZH2 inhibitors, as well as targeting DNA damage repair mechanisms with the use of platinums or PARP inhibitors may be a viable approach. Additionally, augmenting the immune response via checkpoint blockade and/or other modalities of immunotherapy are strategies that merit further investigation.
Furthermore, combination therapy may be a viable option for BAP1-altered tumors. Recent studies suggest that personalized combination therapies may be a more optimal approach, particularly for molecularly complex tumors.[78,83] Given that BAP1 is involved in a number of biological pathways and interacts with an array of cancer-associated genes, this type of combinatorial approach may be necessary. One strategy may be to combine a molecularly targeted therapy such as a PARP inhibitor with an immunotherapy drug such an anti-PD1/PDL1 checkpoint inhibitor to target multiple phenotypes associated with BAP1-altered tumors. Additionally, consideration of BAP1 as a therapeutic target in combination with other potential driver mutations should be a point of investigation for future precision cancer medicine strategies.
Highlights.
BAP1 is a tumor suppressor involved in multiple cell processes related to cancer.
Germline and somatic mutations of BAP1 are commonly found in a variety of cancers.
Molecularly targeted therapies have the potential for use in BAP1-mutated cancers.
Cancer immunotherapies may increase anti-tumor immunity in BAP1-mutated cancers.
ACKNOWLEDGEMENTS
This work was supported by the National Cancer Institute [P30 CA023100] and the Joan and Irwin Jacobs Fund (philanthropic fund).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Declaration of Interests
The authors declare the following financial interests/personal relationships which may be considered as potential competing interests:
Razelle Kurzrock has the following disclosure information: Stock and Other Equity Interests (IDbyDNA, CureMatch, Inc., and Soluventis); Consulting or Advisory Role (Gaido, LOXO, X-Biotech, Actuate Therapeutics, Roche, NeoMed, Soluventis, Pfizer, and Merck); Speaker’s fee (Roche); Research Funding (Incyte, Genentech, Merck Serono, Pfizer, Sequenom, Foundation Medicine, Guardant Health, Grifols, Konica Minolta, DeBiopharm, Boerhringer Ingelheim, and OmniSeq [All institutional]); Board Member (CureMatch, Inc., and CureMetrix, Inc.).
REFERENCES
- [1].Jensen DE, Proctor M, Marquis ST, Gardner HP, Ha SI, Chodosh LA, et al. BAP1: a novel ubiquitin hydrolase which binds to the BRCA1 RING finger and enhances BRCA1-mediated cell growth suppression. Oncogene 1998;16:1097–112. 10.1038/sj.onc.1201861. [DOI] [PubMed] [Google Scholar]
- [2].Wang A, Papneja A, Hyrcza M, Al-Habeeb A, Ghazarian D. Gene of the month: BAP1. Journal of Clinical Pathology 2016;69:750–3. 10.1136/jclinpath-2016-203866. [DOI] [PubMed] [Google Scholar]
- [3].Murali R, Wiesner T, Scolyer RA. Tumours associated with BAP1 mutations. Pathology 2013;45:116–26. 10.1097/PAT.0b013e32835d0efb. [DOI] [PubMed] [Google Scholar]
- [4].Pilarski R, Rai K, Cebulla C, Abdel-Rahman M. BAP1 Tumor Predisposition Syndrome In: Adam MP, Ardinger HH, Pagon RA, Wallace SE, Bean LJ, Stephens K, et al. , editors. GeneReviews®, Seattle (WA): University of Washington, Seattle; 1993. [PubMed] [Google Scholar]
- [5].Baas P, Schunselaar L. MS31.02 Clinical Implementation of BAP1 Inhibitors. Journal of Thoracic Oncology 2018;13:S305–6. 10.1016/j.jtho.2018.08.204. [DOI] [Google Scholar]
- [6].Ladanyi M, Sanchez Vega F, Zauderer M. Loss of BAP1 as a candidate predictive biomarker for immunotherapy of mesothelioma. Genome Medicine 2019; 11:18 10.1186/s13073-019-0631-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Scheuermann JC, de Ayala Alonso AG, Oktaba K, Ly-Hartig N, McGinty RK, Fraterman S, et al. Histone H2A deubiquitinase activity of the Polycomb repressive complex PR-DUB. Nature 2010;465:243–7. 10.1038/nature08966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Nishikawa H, Wu W, Koike A, Kojima R, Gomi H, Fukuda M, et al. BRCA1-Associated Protein 1 Interferes with BRCA1/BARD1 RING Heterodimer Activity. Cancer Res 2009;69:111–9. 10.1158/0008-5472.CAN-08-3355. [DOI] [PubMed] [Google Scholar]
- [9].Zhao W, Steinfeld JB, Liang F, Chen X, Maranon DG, Jian Ma C, et al. BRCA1-BARD1 promotes RAD51-mediated homologous DNA pairing. Nature 2017;550:360–5. 10.1038/nature24060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Machida YJ, Machida Y, Vashisht AA, Wohlschlegel JA, Dutta A. The deubiquitinating enzyme BAP1 regulates cell growth via interaction with HCF-1. J Biol Chem 2009;284:34179–88. 10.1074/jbc.M109.046755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Bononi A, Giorgi C, Patergnani S, Larson D, Verbruggen K, Tanji M, et al. BAP1 regulates IP3R3-mediated Ca2+ flux to mitochondria suppressing cell transformation. Nature 2017;546:549–53. 10.1038/nature22798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Shrestha R, Nabavi N, Lin Y-Y, Mo F, Anderson S, Volik S, et al. BAP1 haploinsufficiency predicts a distinct immunogenic class of malignant peritoneal mesothelioma. Genome Medicine 2019;11:8 10.1186/s13073-019-0620-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Welchman RL, Gordon C, Mayer RJ. Ubiquitin and ubiquitin-like proteins as multifunctional signals. Nature Reviews Molecular Cell Biology 2005;6:599–609. 10.1038/nrm1700. [DOI] [PubMed] [Google Scholar]
- [14].Ventii KH, Devi NS, Friedrich KL, Chernova TA, Tighiouart M, Meir EGV, et al. BRCA1-Associated Protein-1 Is a Tumor Suppressor that Requires Deubiquitinating Activity and Nuclear Localization. Cancer Res 2008;68:6953–62. 10.1158/0008-5472.CAN-08-0365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Campagne A, Lee M- K, Zielinski D, Michaud A, Le Corre S, Dingli F, et al. BAP1 complex promotes transcription by opposing PRC1-mediated H2A ubiquitylation. Nature Communications 2019;10:1–15. 10.1038/s41467-018-08255-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Di Croce L, Helin K. Transcriptional regulation by Polycomb group proteins. Nature Structural & Molecular Biology 2013;20:1147–55. 10.1038/nsmb.2669. [DOI] [PubMed] [Google Scholar]
- [17].Meas R, Mao P. Histone ubiquitylation and its roles in transcription and DNA damage response. DNA Repair (Amst) 2015;36:36–42. 10.1016/j.dnarep.2015.09.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Wu J, Lu L- Y, Yu X. The role of BRCA1 in DNA damage response. Protein Cell 2010;1:117–23. 10.1007/s13238-010-0010-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Ismail IH, Davidson R, Gagne J- P, Xu ZZ, Poirier GG, Hendzel MJ. Germline mutations in BAP1 impair its function in DNA double-strand break repair. Cancer Res 2014;74:4282–94. 10.1158/0008-5472.CAN-13-3109. [DOI] [PubMed] [Google Scholar]
- [20].Yu H, Pak H, Hammond-Martel I, Ghram M, Rodrigue A, Daou S, et al. Tumor suppressor and deubiquitinase BAP1 promotes DNA double-strand break repair. PNAS 2014;111:285–90. 10.1073/pnas.1309085110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Tyagi S, Chabes AL, Wysocka J, Herr W. E2F activation of S phase promoters via association with HCF-1 and the MLL family of histone H3K4 methyltransferases. Mol Cell 2007;27:107–19. 10.1016/j.molcel.2007.05.030. [DOI] [PubMed] [Google Scholar]
- [22].Pan H, Jia R, Zhang L, Xu S, Wu Q, Song X, et al. BAP1 regulates cell cycle progression through E2F1 target genes and mediates transcriptional silencing via H2A monoubiquitination in uveal melanoma cells. The International Journal of Biochemistry & Cell Biology 2015;60:176–84. 10.1016/j.biocel.2015.01.001. [DOI] [PubMed] [Google Scholar]
- [23].Yu H, Mashtalir N, Daou S, Hammond-Martel I, Ross J, Sui G, et al. The Ubiquitin Carboxyl Hydrolase BAP1 Forms a Ternary Complex with YY1 and HCF-1 and Is a Critical Regulator of Gene Expression. Molecular and Cellular Biology 2010;30:5071–85. 10.1128/MCB.00396-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Okino Y, Machida Y, Frankland-Searby S, Machida YJ. BRCA1-associated Protein 1 (BAP1) Deubiquitinase Antagonizes the Ubiquitin-mediated Activation of FoxK2 Target Genes. J Biol Chem 2015;290:1580–91. 10.1074/jbc.M114.609834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Sime W, Niu Q, Abassi Y, Masoumi KC, Zarrizi R, Kohler JB, et al. BAP1 induces cell death via interaction with 14–3-3 in neuroblastoma. Cell Death & Disease 2018;9:1 –16. 10.1038/s41419-018-0500-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Zhang Y, Shi J, Liu X, Feng L, Gong Z, Koppula P, et al. BAP1 links metabolic regulation of ferroptosis to tumor suppression. Nat Cell Biol 2018;20:1181–92. 10.1038/s41556-018-0178-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Gezgin G, Dogrusoz M, van Essen TH, Kroes WGM, Luyten GPM, van der Velden PA, et al. Genetic evolution of uveal melanoma guides the development of an inflammatory microenvironment. Cancer Immunol Immunother 2017;66:903–12. 10.1007/s00262-017-1991-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Figueiredo CR, Kalirai H, Sacco JJ, Azevedo RA, Duckworth A, Slupsky JR, et al. Loss of BAP1 expression is associated with an immunosuppressive microenvironment in uveal melanoma, with implications for immunotherapy development. The Journal of Pathology 2020;250:420–39. 10.1002/path.5384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Abdel-Rahman MH, Pilarski R, Cebulla CM, Massengill JB, Christopher BN, Boru G, et al. Germline BAP1 mutation predisposes to uveal melanoma, lung adenocarcinoma, meningioma, and other cancers. Journal of Medical Genetics 2011. ;48:856–9. 10.1136/jmedgenet-2011-100156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Testa JR, Cheung M, Pei J, Below JE, Tan Y, Sementino E, et al. Germline BAP1 mutations predispose to malignant mesothelioma. Nat Genet 2011;43:1022–5. 10.1038/ng.912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].Wiesner T, Obenauf AC, Murali R, Fried I, Griewank KG, Ulz P, et al. Germline mutations in BAP1 predispose to melanocytic tumors. Nat Genet 2011;43:1018–21. 10.1038/ng.910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Haugh AM, Njauw C- N, Bubley JA, Verzi AE, Zhang B, Kudalkar E, et al. Genotypic and Phenotypic Features of BAP1 Cancer Syndrome. JAMA Dermatol 2017; 153:999–1006. 10.1001/jamadermatol.2017.2330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Walpole S, Pritchard AL, Cebulla CM, Pilarski R, Stautberg M, Davidorf FH, et al. Comprehensive Study of the Clinical Phenotype of Germline BAP1 Variant-Carrying Families Worldwide. J Natl Cancer Inst 2018;110:1328–41. 10.1093/jnci/djy171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [34].Rai K, Pilarski R, Cebulla C m., Abdel-Rahman M h. Comprehensive review of BAP1 tumor predisposition syndrome with report of two new cases. Clinical Genetics 2016;89:285–94. 10.1111/cge.12630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Baumann F, Flores E, Napolitano A, Kanodia S, Taioli E, Pass H, et al. Mesothelioma patients with germline BAP1 mutations have 7-fold improved long-term survival. Carcinogenesis 2015;36:76–81. 10.1093/carcin/bgu227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Masoomian B, Shields JA, Shields CL. Overview of BAP1 cancer predisposition syndrome and the relationship to uveal melanoma. J Curr Ophthalmol 2018;30:102–9. 10.1016/j.joco.2018.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [37].Ewens KG, Lalonde E, Richards-Yutz J, Shields CL, Ganguly A. Comparison of Germline versus Somatic BAP1 Mutations for Risk of Metastasis in Uveal Melanoma. BMC Cancer 2018;18:1172 10.1186/s12885-018-5079-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Harbour JW, Onken MD, Roberson EDO, Duan S, Cao L, Worley LA, et al. Frequent mutation of BAP1 in metastasizing uveal melanomas. Science 2010;330:1410–3. 10.1126/science.1194472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Nasu M, Emi M, Pastorino S, Tanji M, Powers A, Luk H, et al. High Incidence of Somatic BAP1 Alterations in Sporadic Malignant Mesothelioma. Journal of Thoracic Oncology 2015;10:565–76. 10.1097/JTO.0000000000000471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].Kato S, Tomson BN, Buys TPH, Elkin SK, Carter JL, Kurzrock R. Genomic Landscape of Malignant Mesotheliomas. Mol Cancer Ther 2016;15:2498–507. 10.1158/1535-7163.MCT-16-0229. [DOI] [PubMed] [Google Scholar]
- [41].Farzin M, Toon CW, Clarkson A, Sioson L, Watson N, Andrici J, et al. Loss of expression of BAP1 predicts longer survival in mesothelioma. Pathology 2015;47:302–7. 10.1097/PAT.0000000000000250. [DOI] [PubMed] [Google Scholar]
- [42].Jin S, Wu J, Zhu Y, Gu W, Wan F, Xiao W, et al. Comprehensive Analysis of BAP1 Somatic Mutation in Clear Cell Renal Cell Carcinoma to Explore Potential Mechanisms in Silico. J Cancer 2018;9:4108–16. 10.7150/jca.27281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Krantz BA, Dave N, Komatsubara KM, Marr BP, Carvajal RD. Uveal melanoma: epidemiology, etiology, and treatment of primary disease. Clin Ophthalmol 2017; 11:279–89. 10.2147/OPTH.S89591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Yang J, Manson DK, Marr BP, Carvajal RD. Treatment of uveal melanoma: where are we now? Ther Adv Med Oncol 2018;10 10.1177/1758834018757175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [45].Gupta MP, Lane AM, DeAngelis MM, Mayne K, Crabtree M, Gragoudas ES, et al. Clinical Characteristics of Uveal Melanoma in Patients With Germline BAP1 Mutations. JAMA Ophthalmol 2015;133:881–7. 10.1001/jamaophthalmol.2015.1119. [DOI] [PubMed] [Google Scholar]
- [46].AJ P, V N, A G, K M, G F, R K. GNAS, GNAQ and GNA11 Alterations in Patients with Diverse Cancers. Cancer 2018;124:4080–9. 10.1002/cncr.31724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Heppt MV, Amaral T, Kahler KC, Heinzerling L, Hassel JC, Meissner M, et al. Combined immune checkpoint blockade for metastatic uveal melanoma: a retrospective, multi-center study. Journal for ImmunoTherapy of Cancer 2019;7:299 10.1186/s40425-019-0800-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Rusch VW, Giroux D, Kennedy C, Ruffini E, Cangir AK, Rice D, et al. Initial analysis of the international association for the study of lung cancer mesothelioma database. J Thorac Oncol 2012;7:1631–9. 10.1097/JTO.0b013e31826915f1. [DOI] [PubMed] [Google Scholar]
- [49].G S, H Y, Bm S, D C, Ph S. Results of treatment of 33 patients with peritoneal mesothelioma. Br J Surg 2000;87:1587–93. https://doi.org/10.1046/j.1365-2168.2000.01571.x. [DOI] [PubMed] [Google Scholar]
- [50].Ohar JA, Cheung M, Talarchek J, Howard SE, Howard TD, Hesdorffer M, et al. Germline BAP1 mutational landscape of asbestos-exposed malignant mesothelioma patients with family history of cancer. Cancer Res 2016;76:206–15. 10.1158/0008-5472.CAN-15-0295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Vogelzang NJ, Rusthoven JJ, Symanowski J, Denham C, Kaukel E, Ruffie P, et al. Phase III study of pemetrexed in combination with cisplatin versus cisplatin alone in patients with malignant pleural mesothelioma. J Clin Oncol 2003;21:2636–44. 10.1200/JCO.2003.11.136. [DOI] [PubMed] [Google Scholar]
- [52].Research AA for C Dual Checkpoint Blockade Takes Aim at Relapsed Mesothelioma. Cancer Discov 2017;7:OF7–OF7. 10.1158/2159-8290.CD-NB2017-087. [DOI] [PubMed] [Google Scholar]
- [53].Alley EW, Lopez J, Santoro A, Morosky A, Saraf S, Piperdi B, et al. Clinical safety and activity of pembrolizumab in patients with malignant pleural mesothelioma (KEYNOTE-028): preliminary results from a non-randomised, open-label, phase 1b trial. Lancet Oncol 2017;18:623–30. 10.1016/S1470-2045(17)30169-9. [DOI] [PubMed] [Google Scholar]
- [54].Forde PM, Scherpereel A, Tsao AS. Use of Immune Checkpoint Inhibitors in Mesothelioma. Curr Treat Options Oncol 2019;20:18 10.1007/s11864-019-0613-x. [DOI] [PubMed] [Google Scholar]
- [55].Garfield EM, Walton KE, Quan VL, VandenBoom T, Zhang B, Kong BY, et al. Histomorphologic spectrum of germline-related and sporadic BAP1-inactivated melanocytic tumors. Journal of the American Academy of Dermatology 2018;79:525–34. 10.1016/jJaad.2018.05.005. [DOI] [PubMed] [Google Scholar]
- [56].Wiesner T, Murali R, Fried I, Cerroni L, Busam K, Kutzner H, et al. A distinct subset of atypical Spitz tumors is characterized by BRAF mutation and loss of BAP1 expression. Am J Surg Pathol 2012;36:818–30. 10.1097/PAS.0b013e3182498be5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [57].Zhang AJ, Rush PS, Tsao H, Duncan LM. BRCA1-associated protein (BAP1)-inactivated melanocytic tumors. Journal of Cutaneous Pathology 2019;46:965–72. 10.1111/cup.13530. [DOI] [PubMed] [Google Scholar]
- [58].Landreville S, Agapova OA, Matatall KA, Kneass ZT, Onken MD, Lee RS, et al. Histone deacetylase inhibitors induce growth arrest and differentiation in uveal melanoma. Clin Cancer Res 2012;18:408–16. 10.1158/1078-0432.CCR-11-0946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Sacco JJ, Kenyani J, Butt Z, Carter R, Chew HY, Cheeseman LP, et al. Loss of the deubiquitylase BAP1 alters class I histone deacetylase expression and sensitivity of mesothelioma cells to HDAC inhibitors. Oncotarget 2015;6:13757–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].Krug LM, Kindler HL, Calvert H, Manegold C, Tsao AS, Fennell D, et al. Vorinostat in patients with advanced malignant pleural mesothelioma who have progressed on previous chemotherapy (VANTAGE-014): a phase 3, double-blind, randomised, placebo-controlled trial. Lancet Oncol 2015;16:447–56. 10.1016/S1470-2045(15)70056-2. [DOI] [PubMed] [Google Scholar]
- [61].LaFave LM, Beguelin W, Koche R, Teater M, Spitzer B, Chramiec A, et al. Loss of BAP1 function leads to EZH2-dependent transformation. Nat Med 2015;21:1344–9. 10.1038/nm.3947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [62].Sun C, Zhao C, Li S, Wang J, Zhou Q, Sun J, et al. EZH2 Expression is increased in BAP1-mutant renal clear cell carcinoma and is related to poor prognosis. J Cancer 2018;9:3787–96. 10.7150/jca.26275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [63].Zauderer MG, Szlosarek P, Le Moulec S, Popat S, Taylor P, Planchard D, et al. Phase 2, multicenter study of the EZH2 inhibitor tazemetostat as monotherapy in adults with relapsed or refractory (R/R) malignant mesothelioma (MM) with BAP1 inactivation. JCO 2018;36:8515–8515. 10.1200/JCO.2018.36.15_suppl.8515. [DOI] [Google Scholar]
- [64].Muggia F, Safra T. ‘BRCAness and its implications for platinum action in gynecologic cancer. Anticancer Res 2014;34:551–6. [PMC free article] [PubMed] [Google Scholar]
- [65].Hassan R, Morrow B, Thomas A, Walsh T, Lee MK, Gulsuner S, et al. Inherited predisposition to malignant mesothelioma and overall survival following platinum chemotherapy. Proc Natl Acad Sci U S A 2019;116:9008–13. 10.1073/pnas.1821510116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [66].Kobrinski DA, Yang H, Kittaneh M. BAP1: role in carcinogenesis and clinical implications. Transl Lung Cancer Res 2020;9:S60–6. 10.21037/tlcr.2019.11.24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [67].George TJ, DeRemer DL, Parekh HD, Lee J- H, Markham MJ, Daily KC, et al. Phase II trial of the PARP inhibitor, niraparib, in BAP1 and other DNA damage response (DDR) pathway deficient neoplasms including cholangiocarcinoma. JCO 2020;38:TPS591–TPS591. 10.1200/JCO.2020.38.4_suppl.TPS591. [DOI] [Google Scholar]
- [68].Hassan R, Mian I, Wagner C, Mallory Y, Agra M, Padiernos E, et al. Phase II study of olaparib in malignant mesothelioma (MM) to correlate efficacy with germline and somatic mutations in DNA repair genes. JCO 2020;38:9054–9054. 10.1200/JC0.2020.38.15_suppl.9054. [DOI] [Google Scholar]
- [69].Spencer KR, Wang J, Silk AW, Ganesan S, Kaufman HL, Mehnert JM. Biomarkers for Immunotherapy: Current Developments and Challenges. American Society of Clinical Oncology Educational Book 2016:e493–503. 10.1200/EDBK_160766. [DOI] [PubMed] [Google Scholar]
- [70].Zhou Q, Qi Y, Wang Z, Zeng H, Zhang H, Liu Z, et al. CCR5 blockade inflames antitumor immunity in BAP1-mutant clear cell renal cell carcinoma. J Immunother Cancer 2020;8 10.1136/jitc-2019-000228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [71].Pan D, Kobayashi A, Jiang P, LF de Andrade, Tay RE, Luoma AM, et al. A major chromatin regulator determines resistance of tumor cells to T cell-mediated killing. Science 2018;359:770–5. 10.1126/science.aao1710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [72].Sasaki M, Ogiwara H. Synthetic lethal therapy based on targeting the vulnerability of SWI/SNF chromatin remodeling complex-deficient cancers. Cancer Sci 2020; 111:774–82. 10.1111/cas.14311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [73].Miao D, Margolis CA, Gao W, Voss MH, Li W, Martini DJ, et al. Genomic correlates of response to immune checkpoint therapies in clear cell renal cell carcinoma. Science 2018;359:801–6. 10.1126/science.aan5951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [74].Okamura R, Kato S, Lee S, Jimenez RE, Sicklick JK, Kurzrock R. ARID1A alterations function as a biomarker for longer progression-free survival after anti-PD-1/PD-L1 immunotherapy. J Immunother Cancer 2020;8:e000438 10.1136/jitc-2019-000438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [75].Kadoch C, Williams RT, Calarco JP, Miller EL, Weber CM, Braun SMG, et al. Dynamics of BAF-Polycomb complex opposition on heterochromatin in normal and oncogenic states. Nat Genet 2017;49:213–22. 10.1038/ng.3734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [76].Stanton BZ, Hodges C, Calarco JP, Braun SMG, Ku WL, Kadoch C, et al. Smarca4 ATPase mutations disrupt direct eviction of PRC1 from chromatin. Nat Genet 2017;49:282–8. 10.1038/ng.3735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [77].Rodon J, Soria J- C, Berger R, Miller WH, Rubin E, Kugel A, et al. Genomic and transcriptomic profiling expands precision cancer medicine: the WINTHER trial. Nature Medicine 2019;25:751–8. 10.1038/s41591-019-0424-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [78].Sicklick JK, Kato S, Okamura R, Schwaederle M, Hahn ME, Williams CB, et al. Molecular profiling of cancer patients enables personalized combination therapy: the I-PREDICT study. Nat Med 2019;25:744–50. 10.1038/s41591-019-0407-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [79].Tsimberidou A- M, Iskander NG, Hong DS, Wheler JJ, Falchook GS, Fu S, et al. Personalized Medicine in a Phase I Clinical Trials Program: The MD Anderson Cancer Center Initiative. Clin Cancer Res 2012;18:6373–83. 10.1158/1078-0432.CCR-12-1627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [80].Tsimberidou A- M, Hong DS, Ye Y, Cartwright C, Wheler JJ, Falchook GS, et al. Initiative for Molecular Profiling and Advanced Cancer Therapy (IMPACT): An MD Anderson Precision Medicine Study. JCO Precis Oncol 2017;2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [81].Subbiah V, Gainor JF, Rahal R, Brubaker JD, Kim JL, Maynard M, et al. Precision Targeted Therapy With BLU-667 for RET-Driven Cancers. Cancer Discov 2018. https://doi.Org/10.1158/2159-8290.CD-18-0338. [DOI] [PubMed] [Google Scholar]
- [82].Tsimberidou A- M, Hong DS, Wheler JJ, Falchook GS, Janku F, Naing A, et al. Long-term overall survival and prognostic score predicting survival: the IMPACT study in precision medicine. J Hematol Oncol 2019; 12:145 https://doi.Org/10.1186/s13045-019-0835-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [83].Tsimberidou AM, Fountzilas E, Nikanjam M, Kurzrock R. Review of precision cancer medicine: Evolution of the treatment paradigm. Cancer Treat Rev 2020;86:102019 https://doi.Org/10.1016/j.ctrv.2020.102019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [84].Massard C, Michiels S, Ferte C, Deley M-CL, Lacroix L, Hollebecque A, et al. High-Throughput Genomics and Clinical Outcome in Hard-to-Treat Advanced Cancers: Results of the MOSCATO 01 Trial. Cancer Discov 2017. https://doi.Org/10.1158/2159-8290.CD-16-1396. [DOI] [PubMed] [Google Scholar]