

Abstract
A warm anomaly in the upper ocean, colloquially named “the Blob,” appeared in the Gulf of Alaska during the calm winter of 2013–2014, spread across the northern North Pacific (NP) Ocean, and shifted eastward and onto the Oregon shelf. At least 14 species of copepods occurred which had never been observed in shelf/slope waters off Oregon, some of which are known to have NP Gyre affinities, indicating that the source waters of the coastal “Blob” were likely of both offshore (from the west) and subtropical/tropical origin. The anomalously warm conditions were reduced during strong upwelling in spring 2015 but returned when upwelling weakened in July 2015 and transitioned to downwelling in fall 2015. The extended period of warm conditions resulted in prolonged effects on the ecosystem off central Oregon, lasting at least through 2016. Impacts to the lower trophic levels were unprecedented and include a novel plankton community composition resulting from increased copepod, diatom, and dinoflagellate species richness and increased abundance of dinoflagellates. Additionally, the multiyear warm anomalies were associated with reduced biomass of copepods and euphausiids, high abundance of larvaceans and doliolids (indictors of oligotrophic ocean conditions), and a toxic diatom bloom (Pseudo-nitzschia) throughout the California Current in 2015, thereby changing the composition of the food web that is relied upon by many commercially and ecologically important species.
1. Introduction
1.1. Background
During the winter of 2013–2014, the Aleutian Low was weaker than usual (+10 mbar anomaly) and cyclonic storms in the Gulf of Alaska (GOA) were infrequent [Bond et al., 2015]. Low wind speeds resulted in reduced cooling of surface waters by latent and sensible heat fluxes, and reduced mixing of cold deep waters to the sea surface. Thus, the upper layers of the northern portion of the North Pacific Ocean remained relatively warm throughout the winter. By January 2014, sea surface temperature (SST) anomalies were +3°C above the long-term average [Whitney, 2014; Bond et al., 2015]. This warming event was unprecedented in recent history: SST anomaly values in summer 2014 were ~4 standard deviations above climatology and 1.5°C greater in magnitude than historical observations from the North Pacific [Bond et al., 2015].
This warm water mass (first referred to as “the Blob” by State of Washington climatologist Nick Bond) expanded westward and northward during the summer of 2014, and anomalously warm waters were seen across a large portion of the North Pacific from North America to Japan as well as into the Bering Sea and the Sea of Okhotsk (Figure 1). In the temperature anomaly fields, warming extended to a depth of 100 m in the southern GOA as shown in oceanographic surveys completed in February and June 2014 along Line P, a transect along 50°N from 125°W to 145°W (M. Robert, Fisheries and Oceans Canada, personal communication, and shown in Peterson et al. [2015a]); the GOA along Line P remained warm through 2014 and into winter, spring, and summer of 2015 [Bond et al., 2015; Peterson et al., 2015b, 2016a]. The eastern side of the Blob began to shift toward the Pacific Northwest in spring 2014 and moved into continental shelf waters by mid-September 2014, raising SST off central Oregon by 6°C in 7 h, to 19.4°C, a +4.5°C anomaly [Peterson et al., 2015b]. By November 2014, the Blob occupied the upper 80 m of the water column over the shelf off Oregon.
Figure 1.
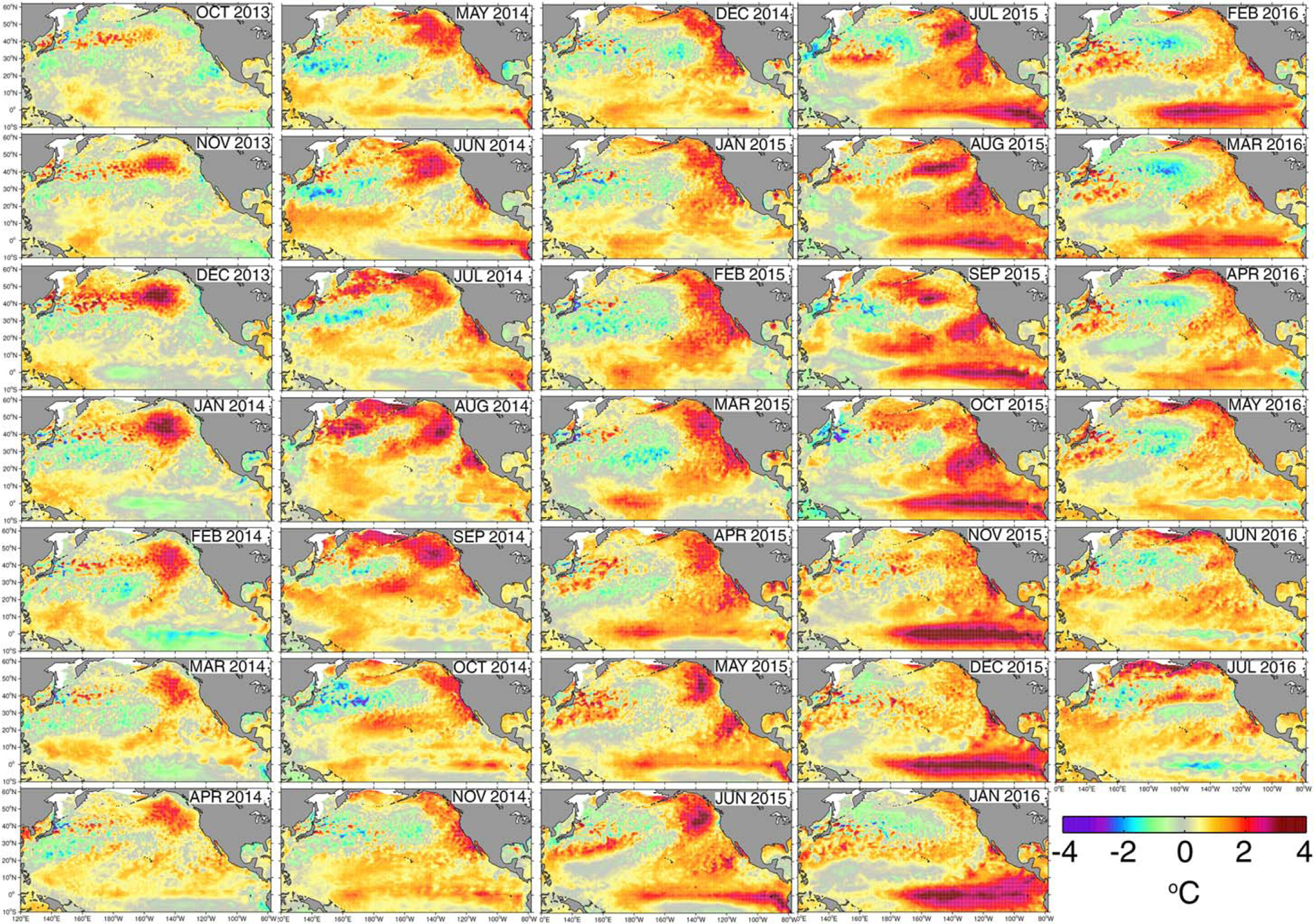
Monthly SST anomalies across the North Pacific from October 2013 through July 2016, showing conditions before the Blob began to form (October 2013), the interval when the Blob was present (2014 and early 2015), and the initiation of an El Niño at the equator (spring 2015).
While the warm anomaly was moving east in the North Pacific Current, an ENSO warm phase developed, creating the first pulse of warm water typical of an El Niño along the eastern equatorial Pacific in boreal spring of 2014. A resumption of westerly trade winds damped this signal [Levine and McPhaden, 2016] and resulted in only a weak El Niño in November 2014 (Figure 1) [McPhaden, 2015]. A stronger El Niño (as quantified by SST indices) developed in the eastern equatorial Pacific in 2015, as pulses of warm water moved eastward along the equator, resulting in extremely warm tropical sea surface temperatures. Equatorial waters were the warmest from November 2015 to January 2016 [Jacox et al., 2016; L’Heureux et al., 2107].
Several other ecologically significant warm-water events have occurred in the Northern California Current over the past 20 years but none as extensive as the warming in 2014–2016. These include the 1997–1998 and the 2009–2010 El Niño events, the extended warming of 2003–2006 associated with two moderate El Niño events [Fisher et al., 2015], and the warming during spring/summer 2005 due to a delay in the onset of the upwelling season. Each warming event was accompanied by SST anomalies of +1 to 2°C and impacts on the pelagic food chain. As an example, during each of these warm events the copepod community was dominated by warm-water copepod species [Peterson et al., 2002; Mackas et al., 2006; Hooff and Peterson, 2006; Peterson, 2009] and temporal changes in copepod community composition tracked linearly the changes in SST with lags of several months [Fisher et al., 2015].
To understand the significance of anomalously warm waters to the hydrography and biota of the Northern California Current (NCC) requires an understanding of the processes that create the range of “normal” conditions in this ecosystem. Key components of the biota are plankton. Many planktonic species are restricted to distinct water masses; thus, their presence/absence in the plankton indicates the presence/absence of specific water types. Such species are referred to as “indicator species” [Fager and McGowan, 1963; Fleminger, 1967; Brinton and Townsend, 1980]. Thus, one can use knowledge of the zoogeography of indicator species of copepods and euphausiids, for example, to describe the types, origin, and timing of arrival of different water types to shelf/slope waters of the NCC.
The water masses (and thus the zooplankton) that dominate shelf and slope waters vary seasonally. During summer, equatorward winds drive surface waters offshore (Ekman transport) to be replaced by the upwelling of cold nutrient-rich waters that move onto the continental shelf and fuel massive amounts of primary production. Equatorward winds also enhance the southward transport of water (and zooplankton) from the coastal GOA into the coastal NCC [Strub and James, 2002a]. These northern copepod species are referred to as “cold water” or “northern copepods” [Mackas et al., 2006; Hooff and Peterson, 2006]. During winter, winds reverse and the poleward Davidson current transports relatively warm coastal water from Southern California to the coastal NCC [Peterson and Miller, 1977] (Figure 2), bringing with it “warm water” or “southern copepods.”
Figure 2.
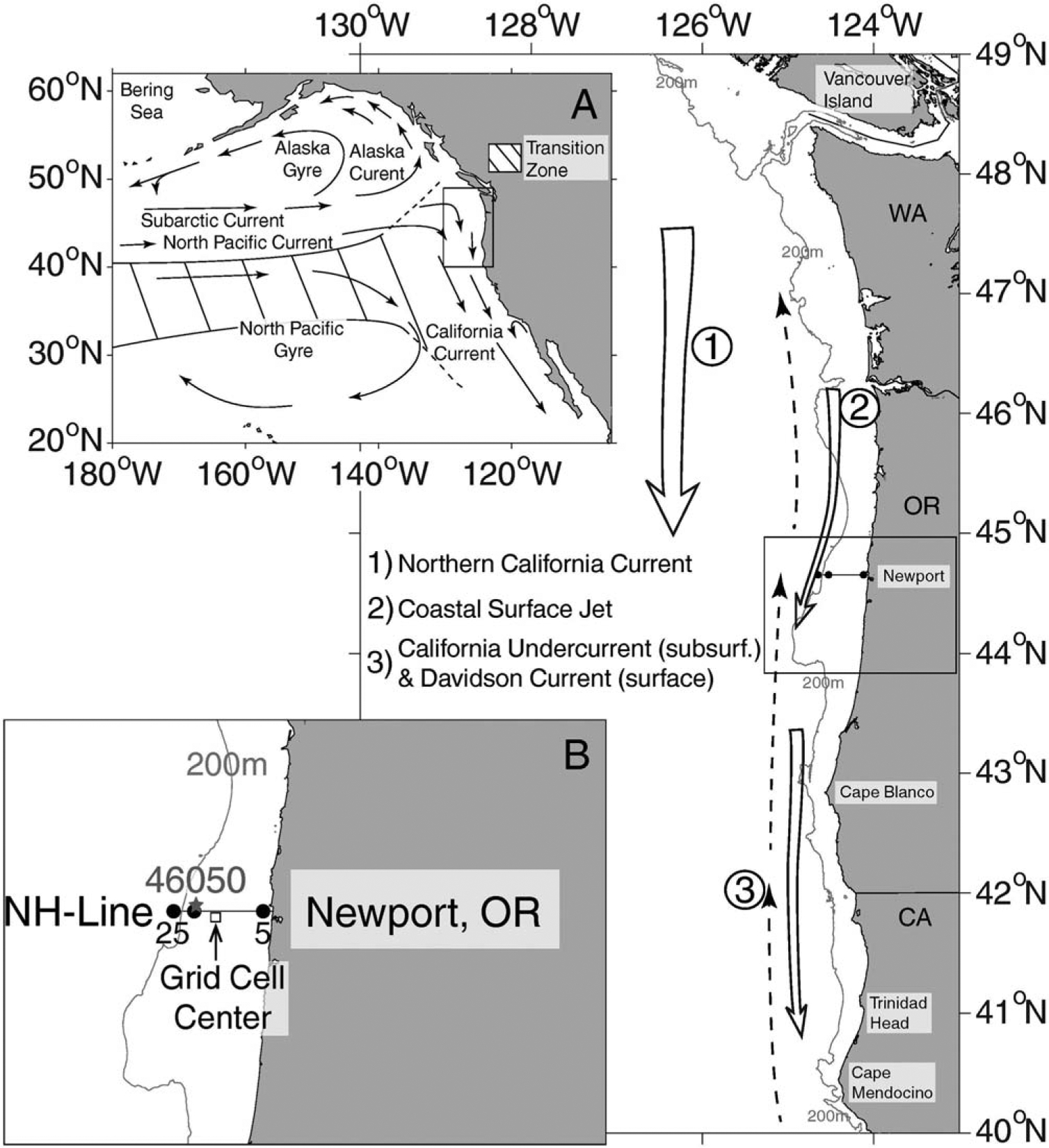
Chart of the NE Pacific showing the seasonal circulation in the California Current in relation to (a) the basin-scale circulation and (b) the study region (boxes).
There are differences in the lipid content of these groups of copepods—northern species are relatively lipid-rich whereas southern species are relatively lipid-poor. Thus, the bioenergetic content of the pelagic food web has a pronounced seasonal component, lipid rich in spring-summer and lipid poor in winter [Hooff and Peterson, 2006; Lee et al., 2006; Miller et al., 2017].
Cold-water lipid-rich northern copepods are also largely replaced by warm-water southern copepods during El Niño events [Miller et al., 1985; Peterson et al., 2002; Keister et al., 2005; Fisher et al., 2015] and during the positive phase of the Pacific Decadal Oscillation [Keister et al., 2011; Peterson et al., 2014]. Each of these events is associated with anomalously weak transport of cold subarctic waters from the north [Strub and James, 2002b; Keister et al., 2011; Bi et al., 2011] and/or increased transport of coastal subtropical waters from the south.
Disruptions of these seasonal and interannual patterns have implications for all trophic levels of the NCC ecosystem [Peterson and Schwing, 2003; Peterson et al., 2014] because a large biomass of predators undergo a feeding migration to the NCC in spring to take advantage of the typical lipid-rich food supply and fatten up for the coming winter. As examples, Pacific whiting and sardines migrate from Southern California to the NCC, humpback whales from the Eastern Tropical Pacific, gray whales from Baja California, sooty shearwaters from New Zealand and black-footed and Laysan albatross from Hawaii. At the same time, Pacific salmon migrate to the sea from coastal streams and large rivers of the Pacific Northwest and their survival is positively correlated with the availability of an abundant and lipid-rich food source when they first enter the ocean. Thus, atmospheric and oceanographic conditions that result in transport of specific water masses and either a “cold-water, lipid rich” or a “warm-water, lipid-poor” food chain can affect not only the biota of the California Current but long-distance migratory species as well.
1.2. Purpose
To determine the effects of anomalous atmospheric conditions and wide-spread warming associated with recent Blob and El Niño events on the pelagic ecosystem of the NCC, we examine the evolution of, and changes in, the physical and biogeochemical water properties of the coastal ocean, and the changes in the characteristics of the phytoplankton and mesozooplankton observed at 44.6°N (Newport, Oregon, United States). For this analysis, we compare ocean conditions during 2014–2016 to those during past warm events, observed during a 20 year biweekly-monthly time series of physical and biological observations off Newport, Oregon. We then examine the details of relationships between physical forcing and biological responses during the extratropical warming of 2014–2016. We hypothesize that the origins of the anomalously warm water that arrived on the Oregon shelf during the autumn 2014 Blob event was from a source farther offshore (west), and/or from farther south compared to past warm events which we have observed. What sets this contribution apart from others describing ocean conditions from 2014 to 2016 is our focus on species composition and abundance of phytoplankton, copepods, krill, and other zooplankton. We conclude with a discussion of how future warm events might affect/disrupt the transfer of energy from lower to upper trophic levels, to fisheries.
2. Methods
Meridional and zonal 10 m wind velocities presented here are derived from North American Regional Reanalysis (NARR; data available at http://www.esrl.noaa.gov/psd/data/gridded/data.narr.monolevel.html) model fields [Mesinger et al., 2006]. NARR, which is an extension of the NCEP (National Centers for Environmental Prediction) Global Reanalysis run over the North American region, has a 3 h temporal resolution and an approximately 32 km horizontal grid resolution. Each 3 h field was interpolated to a uniform 0.25° latitude × 0.25° longitude grid and vector averaged monthly for the period January 1982 to December 2016.
Daily, 0.25° latitude × 0.25° longitude, AVHRR-only, NOAA Optimum Interpolation Sea Surface Temperature fields (OISST) [Reynolds et al., 2007] (data available at https://www.ncdc.noaa.gov/oisst) were monthly averaged for the period January 1982 to December 2016. Daily SST data were also obtained from NOAA Buoy 46050, located approximately 36 km off the coast from Newport, Oregon.
Daily, blended sea surface height anomalies (SSHA) and geostrophic current velocity fields [Risien and Strub, 2016] (data available at https://doi.org/10.7267/N90Z7153) were averaged monthly for the period January 1993 to December 2016. The published SSHA data set [Risien and Strub, 2016], which blends gridded satellite altimeter sea level anomaly (SLA) fields with tide gauge observations, covers the period 1993–2014. The data set was extended through December 2016 using a combination of delayed-time (1 January 1993 to 11 September 2015) and near real-time (12 September 2015 to 31 December 2016) AVISO fields. As described in Risien and Strub [2016], the meridional component of the geostrophic currents at this location are reasonably well correlated (r = 0.73) with in situ ADCP observations collected at 16.5 m by the NH-10 mooring, located at approximately 44.64°N, 124.3°W [Kosro, 2002].
The seasonal cycles for each 0.25° × 0.25°grid cell were obtained using the same procedure applied to the OISST, NARR 10 m wind velocities, SSHA, and surface current velocity fields. Each multiyear time series of monthly means was fitted to a nine-parameter regression model consisting of a constant plus four harmonics [Risien and Chelton, 2008]. The seasonal cycles were then calculated from the regression coefficients for each 0.25° × 0.25° grid cell using the mean and first two harmonics (1 and 2 cycles per year). The OISST and NARR seasonal cycles are based on the 30 year period 1982–2011. Seasonal cycles for the SSHA and surface current velocity fields are based on the 20 year period 1993–2012.
In our analysis, we remove the seasonal cycles from gridded SSHA and the SST time series to produce non-seasonal anomalies, as is customary in discussions of El Niño along the U.S. West Coast (USWC). When we present the alongshore winds and altimeter-derived currents, however, we include the seasonal cycles and indicate those cycles as well as the actual individual monthly values. This allows us to see the actual direction of the winds and currents, as well as to assess the anomalies (difference from the seasonal cycles). We note that although the long-term mean is subtracted from the SLA fields (and thus from the alongshore currents), those mean currents over the Oregon continental shelf are poleward with magnitudes of only 0.02–0.04 m s−1 (due to the seasonal reversal of the currents [Strub et al., 1987]) and make no visible difference in plots of the alongshore currents.
The OISST, NARR, SSHA, and surface current velocity time series presented here are from the 0.25° latitude × 0.25° longitude grid cell centered at 44.625°N, 124.375°W. The Pacific Decadal Oscillation (PDO) and Oceanic Niño Index (ONI) are from http://research.jisao.washington.edu/pdo/PDO.latest and http://www.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ensoyears.shtml, respectively. The ONI is a 3 months running mean of SST anomalies in the Niño 3.4 region (5°N–5°S, 120°W–170°W), based on centered 30 year base periods updated every 5 years. Note that the eastern boundary of the ONI region is in the central Pacific, separated from the coast by 40° of longitude (4000 km). Thus, it is more useful as an indicator of the equatorial forcing of atmospheric teleconnections (hence its use for weather forecasting) than as an indicator of signals next to South America that move poleward into both hemispheres during El Niño events [Strub and James, 2002b].
Hydrographic and plankton data are from an ongoing 20+ year time series of hydrographic and plankton sampling in the Northern California Current, initiated in May 1996. Data are collected fortnightly to monthly along the Newport Hydrographic Line (NH, 44.6°N) at seven stations located from 1 to 46 km from shore (Figure 2), with water depths ranging from 30 to 300 m. Most of the data presented here are from three stations, NH-5 in mid-shelf waters, NH-20 in outer shelf waters, and NH-25, located just beyond the shelf break, 5, 20, and 25 nautical miles (9, 32, and 40 km) from shore in 60, 140, and 300 m water depth, respectively. Measurements include water column profiles of temperature and salinity from a Seabird CTD (SBE 19 or SBE 25), nitrogen (nitrate + nitrite), chlorophyll-a, and phytoplankton abundance and species composition from surface samples, and net tow samples to estimate the biomass and species composition of the zooplankton community, including euphausiids.
Not all time series of these data span the full time period of this study. The copepod time series is 20+ years in length (started May 1996); the CTD, nitrogen, and chlorophyll-a time series is 19+ years (started in February 1997); and the 15 year euphausiid biomass record started in 2001. Species of the diatom Pseudo-nitzschia (PN) were recorded starting in 2001, and phytoplankton community composition data collection began in 2009. Toxicity of PN blooms is assessed from data on domoic acid concentration in razor clams, collected, and published by the State of Oregon Department of Agriculture (http://www.oregon.gov/ODA/programs/FoodSafety/ShellfishClosures.aspx).
Details on methods of sampling and sample processing are found in the following references: nutrients, chlorophyll-a, and phytoplankton in Du et al., [2015], copepods and doliolids in Peterson and Keister [2003], and euphausiids in Shaw et al. [2010]. Copepod data are summarized in this paper as species richness (the number of copepod species in a sample) and biomass anomalies of the warm-water and cold-water copepod species from Fisher et al. [2015]. Additionally, to objectively determine whether changes in the copepod community occurred, we conducted hierarchical cluster analysis followed by a similarity profile test to determine which clusters were significantly different (α 0.01 [Clark and Warwick, 2001]). Cluster analysis objectively groups samples based on how closely related they are to one another (percent similarity) such that samples within a cluster are more closely related compared to samples between clusters. Copepod species densities (no. m−3) from 2014 to 2016 were averaged monthly and transformed (log10 ×+1) to deemphasize the contribution of very abundant species. Hierarchical cluster analysis was performed with group average linking and 1000 permutations on a Bray-curtis similarity matrix constructed among species densities for each month. The resulting dendrogram was labeled by month and year and color-coded based on whether the winds were poleward (downwelling) or equatorward (upwelling). Data were not collected in December 2015 due to inclement weather and equipment failures.
3. Results
3.1. Time Series: PDO, ONI, SST, SSH, Winds, and Currents Off Central Oregon (44.625°N, 124.375°W)
Over interannual timescales, local sea surface properties, OISST (hereafter referred to as SST, unless otherwise noted) and SSH, tracked the cycles of two basin-scale oscillations, the PDO and ONI (Figures 3a–3c). For example, the highest correlation coefficient of SST and PDO was r = 0.59 with zero lag; the highest correlations of SST with ONI were r = 0.41 with lags of 1–3 months. Over the past 20 years, the PDO had four periods of warm phase, January 1996 to June 1998, August 2002 to September 2005, August 2009 to May 2010, and January 2014 to 2016. SSH and SST anomalies were also positive during most of these periods. During each of these warm periods, El Niño events occurred as indicated by positive values of the ONI.
Figure 3.
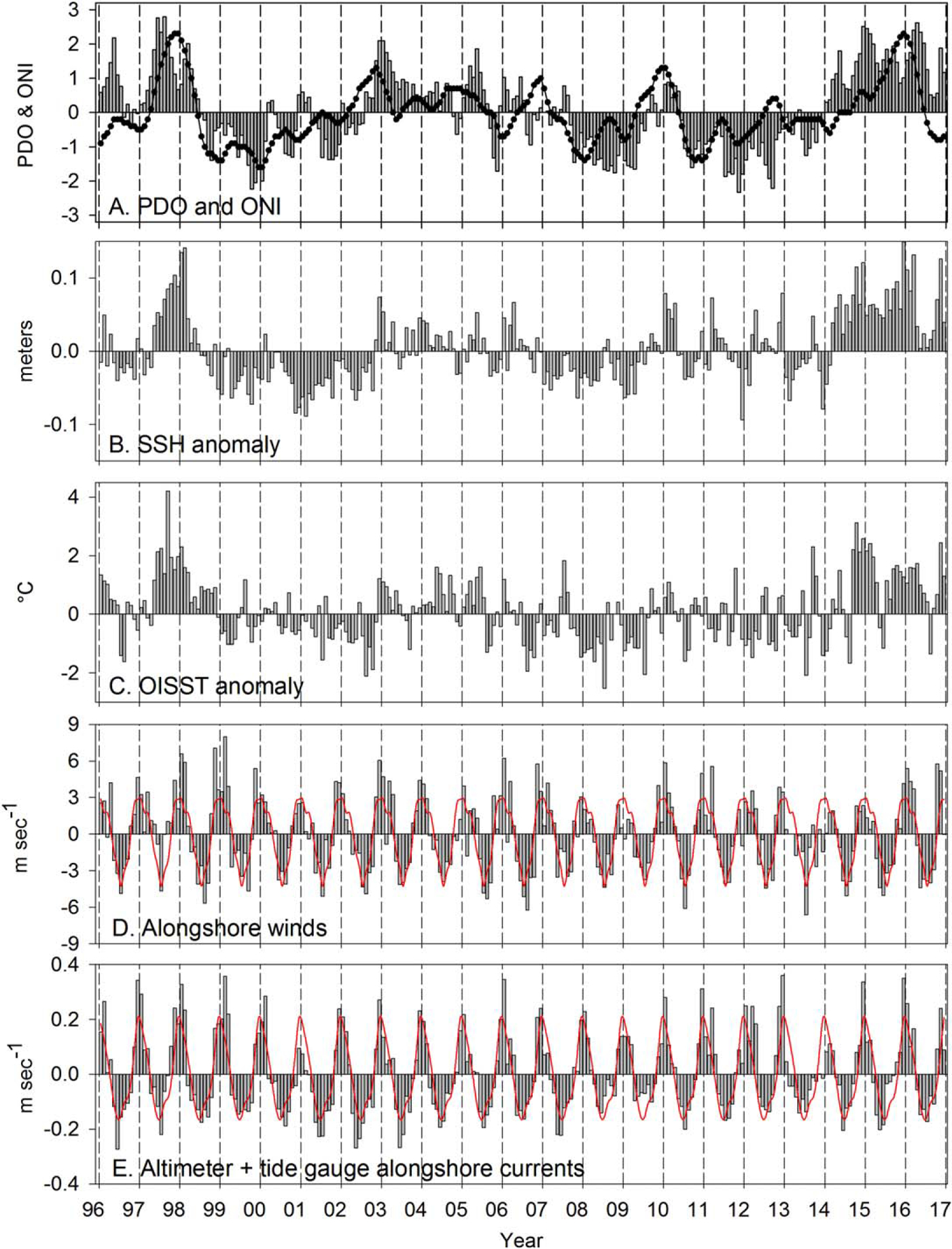
Pacific Decadal Oscillation (PDO; vertical bars), Oceanic Niño Index (ONI; line), Sea Surface Height (SSH), Optimum Interpolation Sea Surface Temperature (OISST), alongshore winds, and alongshore currents. The red lines in Figures 3d and 3e are the climatology of winds and currents respectively. Positive is poleward negative is equatorward.
The change in sign of the SSH anomalies generally occurred in the same month as the change in sign of the PDO with the highest correlation coefficients of the PDO with SSH at zero lag (r = 0.61) but with statistically significant correlations out to lags of at least 5 months (r = 0.47). Positive values of the PDO and ONI and positive anomalies of SSH and SST were present during the warm events of 2003–2005 and 2010. The strongest and longest period of positive anomalies were observed during the 1997–1998 El Niño and the extended warming in 2014–2016.
The alongshore component of the wind shows a pronounced seasonal cycle, poleward in winter and equatorward in summer (Figure 3d). During winter (November–March), poleward winds averaged 2.5 m s−1, were strongest during 1997–1998, 1998–1999, and 2015–2016, and weakest in 2013–2014 (Table 1). Equatorward winds in summer (May–September) averaged 2.8 m s−1, strongest during 2006, 2015, and 2016, and weakest in 1997, 2009, and 2013 (Table 1).
Table 1.
Current and Wind Speeds in Summer and Winter, Ranked From Strongest (Rank 1) to Weakest (Rank 20 or 21)a
| Summer (May–September) | Winter (November–March) | |||||||
|---|---|---|---|---|---|---|---|---|
| Currents | Winds | Currents | Winds | |||||
| Year | cm s−1 | Rank | m s−1 | Rank | cm s−1 | Rank | m s−1 | Rank |
| 1996 | −0.16 | 4 | −3.03 | 9 | 0.18 | 4 | 2.62 | 12 |
| 1997 | −0.09 | 18 | −0.73 | 21 | 0.19 | 3 | 3.92 | 2 |
| 1998 | −0.13 | 6 | −3.49 | 4 | 0.23 | 1 | 5.21 | 1 |
| 1999 | −0.13 | 7 | −2.50 | 16 | 0.14 | 9 | 2.94 | 7 |
| 2000 | −0.13 | 11 | −2.36 | 18 | 0.03 | 20 | 1.32 | 17 |
| 2001 | −0.18 | 1 | −2.78 | 12 | 0.15 | 8 | 2.65 | 10 |
| 2002 | −0.18 | 2 | −3.05 | 8 | 0.12 | 13 | 3.46 | 4 |
| 2003 | −0.17 | 3 | −3.09 | 5 | 0.11 | 17 | 2.63 | 11 |
| 2004 | −0.13 | 10 | −2.46 | 17 | 0.10 | 18 | 0.95 | 18 |
| 2005 | −0.12 | 14 | −2.97 | 10 | 0.15 | 6 | 2.68 | 9 |
| 2006 | −0.13 | 8 | −4.20 | 1 | 0.11 | 15 | 3.21 | 6 |
| 2007 | −0.15 | 5 | −2.55 | 15 | 0.12 | 14 | 1.38 | 16 |
| 2008 | −0.08 | 20 | −3.07 | 6 | 0.12 | 11 | 0.93 | 19 |
| 2009 | −0.07 | 21 | −2.27 | 19 | 0.12 | 12 | 3.28 | 5 |
| 2010 | −0.12 | 12 | −2.74 | 13 | 0.17 | 5 | 2.86 | 8 |
| 2011 | −0.12 | 13 | −2.60 | 14 | 0.15 | 7 | 1.80 | 13 |
| 2012 | −0.11 | 15 | −2.81 | 11 | 0.11 | 16 | 1.54 | 14 |
| 2013 | −0.09 | 19 | −1.70 | 20 | 0.05 | 19 | 0.54 | 20 |
| 2014 | −0.10 | 16 | −3.06 | 7 | 0.13 | 10 | 1.39 | 15 |
| 2015 | −0.09 | 17 | −3.53 | 3 | 0.19 | 2 | 3.59 | 3 |
| 2016 | −0.13 | 9 | −3.76 | 2 | ||||
Winter is defined as for example, “1996” are the winds averaged from November 1996 through March 1997. Data are not yet available for winter 2016. Negative values indicate equatorward flow.
Alongshore geostrophic currents over the shelf off Newport, Oregon (Figure 3e) show a seasonal cycle similar to the winds—poleward in winter and equatorward in summer. The mean value of poleward velocities averaged from November to March was 0.14 m s−1 and was strongest during winters beginning in November 1997, 1998, and 2015 (Table 1) and weakest during winters beginning in November 2000, 2004, and 2013 (the winter during which the Blob began to form in the Gulf of Alaska). During the summer upwelling season (May–September), velocities averaged −0.12 m s−1 (the negative sign indicates equatorward flow) and were strongest early in the time series (2001, 2002, and 2003), and weakest during the latter years of the time series (2008, 2009, and 2013–2015). Maximum poleward alongshore currents in winter are generally observed in December or January; maximum equatorward currents in May, June, or July.
3.2. Time Series: Hydrography, Nitrogen, Chlorophyll-a, and Copepod Biomass at Station NH-5
Temperature and salinity properties in waters 10 m above the sea floor at station NH-5 (Figure 4) tended to cycle between cold/salty and warm/fresh. For example, T-S anomalies were relatively warm/fresh during the El Niño of 1997–1998 and during the warm periods of 2014–2015 and 2015–2016. Waters were also warm/fresh during the fall/winter season of 2003–2004 and during winter 2010. T-S anomalies were cold/salty from 1999 to 2002, from 2007 to 2009, and during most of 2013. T-S properties at 150 m depth at NH-25 in the California Undercurrent were similar to the deep water on the shelf at NH-5 during the 1997–1998 El Niño event—extremely warm/fresh anomalies were observed, whereas during 2014–2016, anomalies at 150 m were similar to other warm events referenced above (Figure 4).
Figure 4.
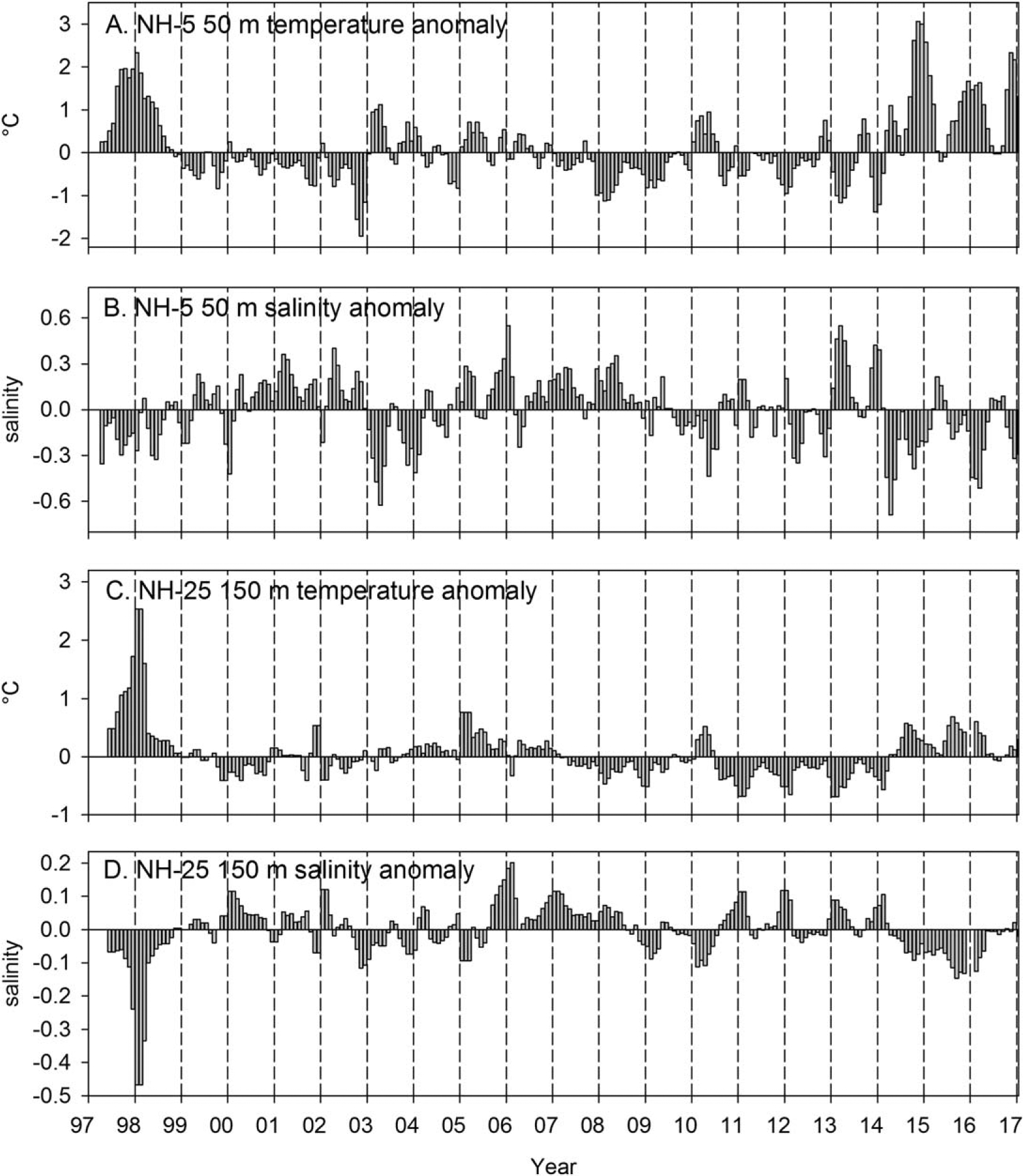
Temperature and salinity from near bottom 50 m at NH-5 (continental shelf; station depth 62 m) and at 150 m at NH-25 (continental slope; station depth 300 m). All data are smoothed with a 3 month running mean to remove high-frequency variability.
Temperature anomalies shown in Figure 5 illustrate the differences throughout the water column. At NH-5 (water depth 60 m), anomalies over the entire water column were frequently uniform. During the 1997–1998 El Niño (from September 1997 through spring 1998) and during the early phases of the Blob (September 2014 through February 2015) temperature anomalies of +4°C occurred to depths of at least 50 m. During fall-winter 2015–2016, the entire water column at NH-5 again experienced uniform positive anomalies of 2–3°C. At station NH-25 (water depth 300 m), during the 1997–1998 El Niño event, pronounced warm anomalies occurred to ~150 m whereas during the period when the Blob first came onshore off central Oregon (September 2014 through February 2015), strong anomalies occurred only in the upper 50–80 m. Deeper anomalies are seen earlier in 2014, while the surface waters remained cool/neutral due to upwelling. However, during fall-winter 2015–2016, weaker warm anomalies extended throughout the water column to 150 m. The weak but deep warming during 2015–2016 is similar to the observations during 2004–2005 and 2009–2010.
Figure 5.
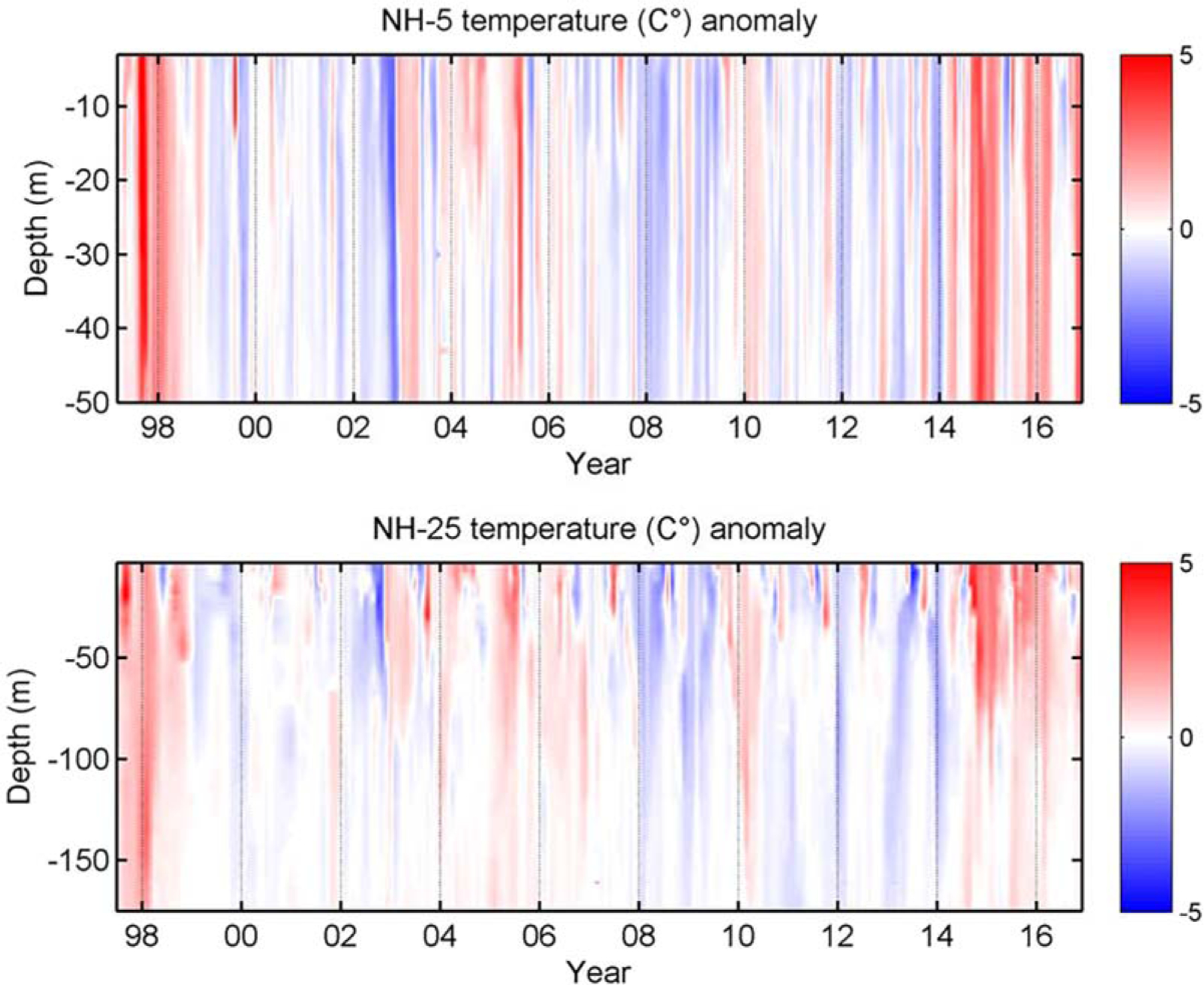
A Hovmöller diagram showing temperature anomalies (°C;) at NH-5 (top) and NH-25 (bottom), 1997 through December 2016.
Anomalies of nitrogen and chlorophyll-a concentration in surface waters at NH-5 tracked SST during many of the periods of extended warm ocean conditions (Figures 6a and 6b): in 1997–1998, 2003–mid-2005, mid-2014–2016, both nitrogen and chlorophyll-a concentrations were below average during most months (note the reversals of anomaly signs on the right-hand axes of Figures 6a and 6c). The largest negative anomalies of nitrogen (~10 μM) were observed at three points during the 20 year time series (Figure 6a): (i) during the summer of 1997, before the arrival of the main signal of the 1997–1998 El Niño along the USWC but coincident with the high SSH and SST (Figure 3), (ii) during summer (July 2014) before the arrival of the Blob at the surface (September 2014), and (iii) in July 2015, as 50 m anomalies at NH-5 return to warm/fresh after strong upwelling (Figures 4a and 4b). The strongest positive anomalies of nitrogen were observed when SST anomalies were weak or negative—2000, 2002, and mid-2005 through mid-2009, whereas chlorophyll-a anomalies were near zero. There were also periods with negative anomalies of both nitrogen and SST (mid-2000 to mid-2002) as well as periods (2009 and 2010) when nitrogen anomalies were negative but chlorophyll-a anomalies were the most positive in the 20 year record.
Figure 6.
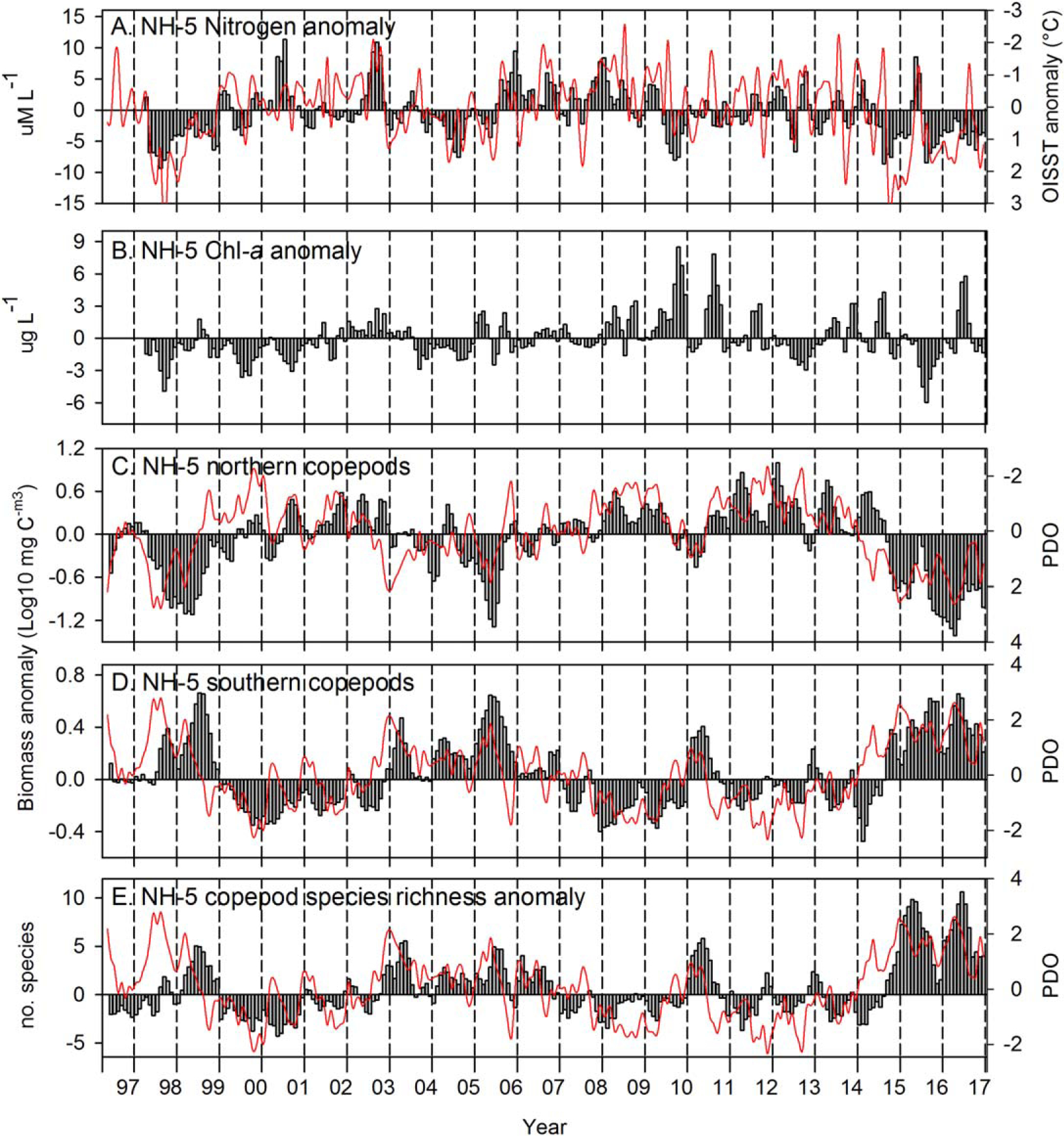
Nitrogen, chlorophyll-a, northern and southern copepod biomass and copepod species richness (vertical bars) anomalies at station NH-5, with OISST at the station and PDO overlaid as the red lines. All data are smoothed with a 3 month running mean to remove high-frequency variability. Note the reversals of anomaly signs on right axes of Figures 6a and 6c.
The copepod biomass tracked the basin-scale PDO and local SST (Figures 6a, 6c, and 6d), as well as the basin-scale ONI and local SSH (Figure 3). The highest correlations of the PDO with northern copepods occurred with a 1 month lag (r = −0.60); the lag with the southern copepods was 3 months (r = 0.60). The highest positive biomass anomalies of northern copepod species were observed when PDO and SST anomalies were negative whereas the highest positive anomalies of southern copepod species occurred during years when the PDO and SST anomalies were positive. The most negative anomalies of northern copepod biomass (and positive anomalies of southern copepods) were observed on four occasions: during the 1997–1998 El Niño, the summer of 2005 when the beginning of the upwelling season was delayed by several months, during the two warm periods of 2014–2016, with the most prolonged anomalies in the time series occurring during 2014–2016 (Figures 6c and 6d). Copepod species richness anomalies (Figure 6e) also tracked the PDO (r = 0.56 with a 2 month lag); positive anomalies were always observed in association with (during or just following) positive PDO warm-water events. The greatest copepod species richness was observed during the extended warming of 2014–2016, with weaker positive anomalies during other warm events (1997–1998 El Niño, 2003–2006, and in 2010).
3.3. Time Series: Biological Variables in Outer Shelf and Slope Waters
The biomass of the oceanic euphausiid, Euphausia pacifica, was consistently high (>10 mg C m−3) from 2001 through 2008, but has been more variable (low with occasional spikes that are smaller in magnitude) since then, with peaks in 2011, 2014, and 2016 and the lowest values (<5 mg C m−3) in 2013 and 2015 (Figure 7). The biomass of the coastal euphausiid, Thysanoessa spinifera, is always much lower than that of E. pacifica (Figure 7b). T. spinifera biomass was consistently higher (>2 mg C m−3) during the cool PDO period 2001–2003 and consistently lower (~1 mg C m−3) and more patchy during 2004–2009. Biomass showed an increase in variability from 2010 to 2014, with isolated spikes during 2010, 2012, and 2013 and very low biomass during 2011 and 2014. Throughout all of 2015 and 2016 very few (two individuals in 2015 and five individuals in 2016) adult T. spinifera were collected at NH-20.
Figure 7.
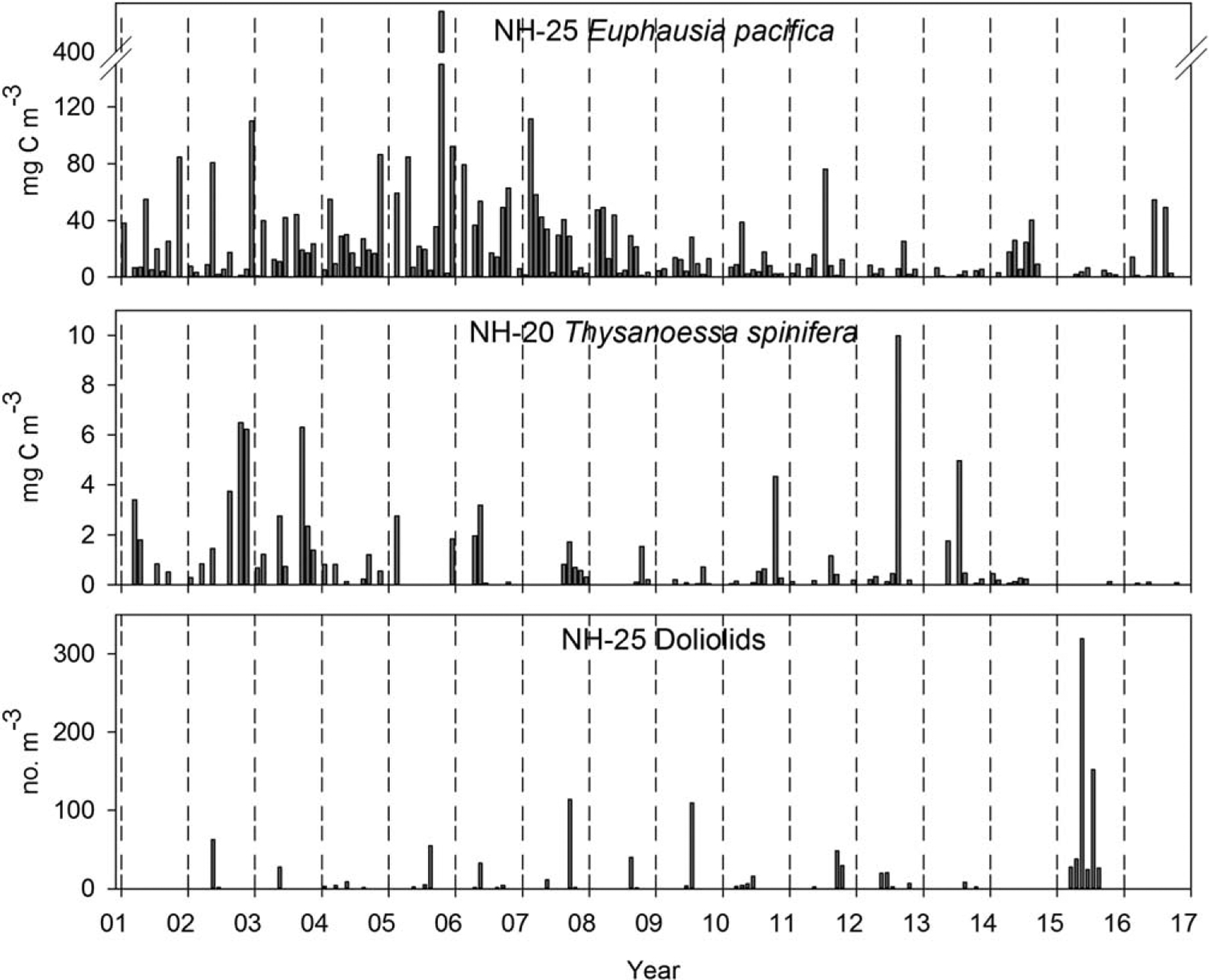
Biomass of adult E. pacifica from NH-25 and adult T. spinifera from NH-20 and density of doliolids from station NH-25. Doliolids have not been enumerated from samples in 2016.
The doliolid, Dolioletta gegenbauri, an indicator of blue water/oligotrophic ocean conditions, is typically found farther offshore than 40 km but was unusually common at station NH-25 during the summer of 2015 (Figure 7). The greatest abundance of doliolids in the NH-25 time series occurred during the summer of 2015 (98.8 individuals m−3, frequency of occurrence 6 out of 12 months) compared to 2005 (20.7 m−3, freq = 3 months), 2007 (31.8 m−3, freq = 4 months), 2009 (56.5 m−3, freq = 2 months), and 2011 (26.7 m−3, freq = 3 months).
3.4. Winds, Currents, SSH, and SST Off Central Oregon: 2013–2016
During winters, monthly poleward winds off central Oregon (Figure 8b) were calm in winter 2013 (January–March), and anomalously weak during the following winter 2013–2014 (November 2013 to January 2014). Winds were also below average in winter 2014–2015, but stronger during December 2015 to March 2016. Alongshore currents were similar: weak during January–March 2013 and September 2013 to January 2014, and average-to-strong from October 2014 to February 2015 and October 2015 to March 2016. The strongest currents were observed in December 2014 and December 2015 to January 2016. In terms of overall ranks, currents and winds during winter (November–March) 2013–2014 were 19th and 20th lowest, respectively, whereas winter 2015–2016 ranked 2nd strongest in currents and 3rd strongest in winds (Table 1). Currents and winds during winter 2014–2015 fell in the middle of the rankings.
Figure 8.
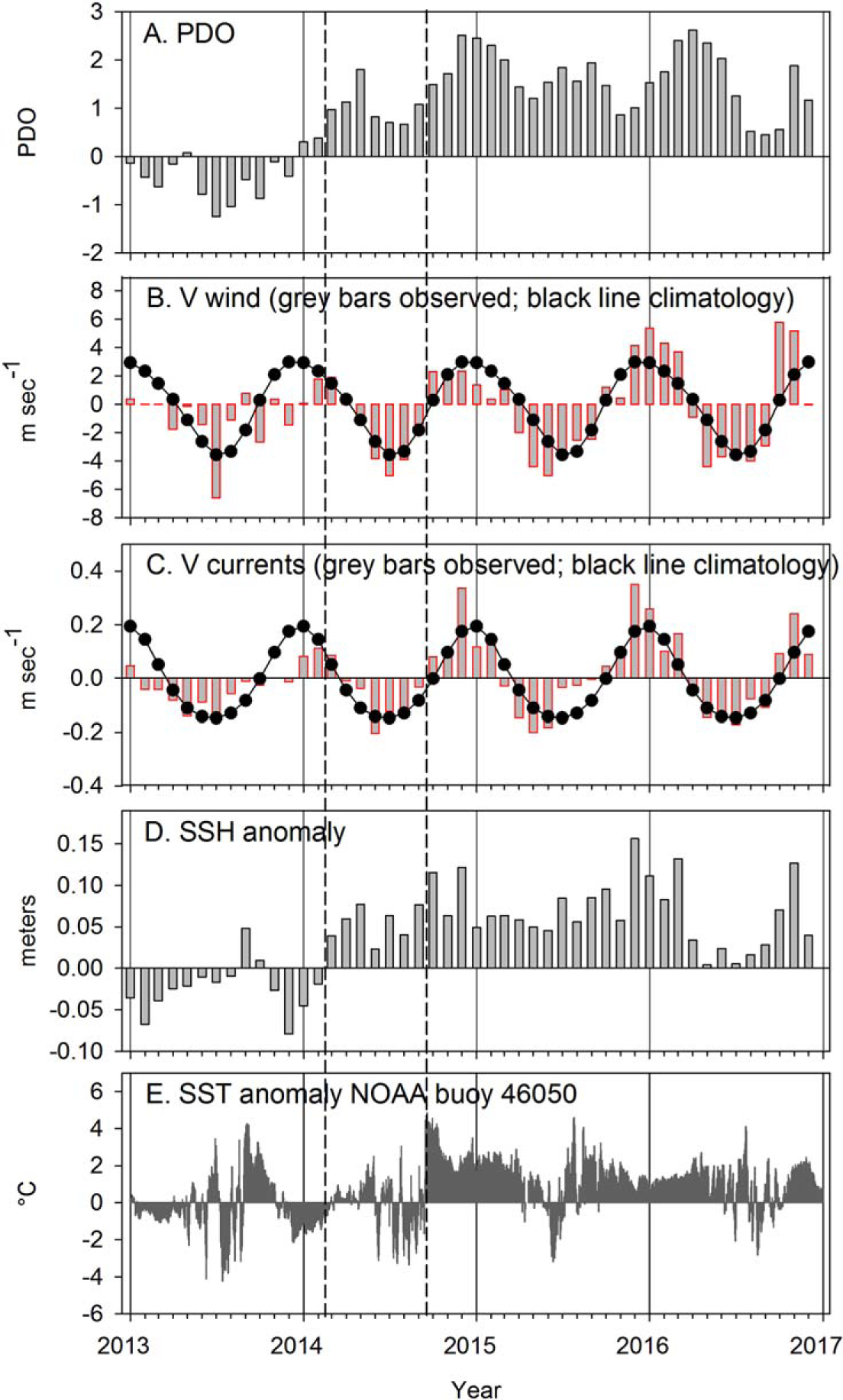
Monthly values of the PDO, alongshore winds and currents, monthly anomalies of SSH, and daily anomalies of SST from NOAA buoy 46050 from January 2013 through December 2016. The dashed vertical lines indicate (i) when SSH anomalies and the PDO became positive for an extended period (beginning in March 2014), (ii) when the Blob moved into continental shelf waters (mid-September 2014).
During summers, in 2013 equatorward winds were strong only in July (Figure 8b). Equatorward winds were stronger and more persistent during June–August 2014 and April–June 2015; they were slightly greater than average during May–September 2016 (Figure 8b and Table 1). Ranks during the longer summer period (May–September) were 20th in 2013, but 7th, 3rd, and 2nd in 2014, 2015, and 2016, respectively. The geostrophic currents (Figure 8c) responded to the wind forcing, with strong or average equatorward currents corresponding to the winds during June–August 2014 and April–June 2015. However, considering the longer summer period of May–September, average equatorward currents during the summers of 2014, 2015, and 2016 were among the weakest of the time series due to their shorter 3 month periods of strong upwelling, ranking 16th, 17th, and 9th, respectively. The decrease in equatorward currents beginning in July 2015 and continuing through September was much more abrupt than the decrease in equatorward winds, which continued at moderate strength during most of July–September 2015, as discussed below. Furthermore, despite the strong southward winds in summer, sea level anomalies over the shelf became positive in March 2014 and remained mostly positive and high throughout 2015 and into spring 2016. These SSH anomalies did decrease slightly during strong equatorward winds in June–August 2014 and May–June 2015. SSH anomalies did not return to near zero until May 2016 (Figure 8d).
Daily SST anomalies from NOAA Buoy 46050 (Figure 8e) were negative on most dates between November 2013 and mid-February 2014, and again from June to early September 2014. SST anomalies then increased overnight (on 14 September) from −2 to +4°C (to a maximum SST of 19.1°C) when the Blob came ashore in central Oregon. SST positive anomalies at the buoy remained high through 2016, apart from brief periods of cooling during May–June 2015, following strong equatorward winds and upwelling (Figure 8b and Table 1) and in summer 2016 during a 5 month period of equatorward winds that were similar to climatology.
3.5. Hydrography, Nitrogen, and Chlorophyll-a: 2013–2016
Throughout most of 2013 (excepting summer) and through February 2014, deep waters at NH-5 were among the coldest and saltiest of the 20 year time series (compare Figures 9a and 9b with Figures 4a and 4b). However, in March 2014, deep waters at both NH-5 and NH-25 began to warm and freshen and remained so throughout most of the remaining months of 2014, 2015, and 2016, the same time period over which SSH anomalies were positive. March 2014 is also the month when the Blob began moving closer to shore off the Pacific Northwest (Figure 1).
Figure 9.
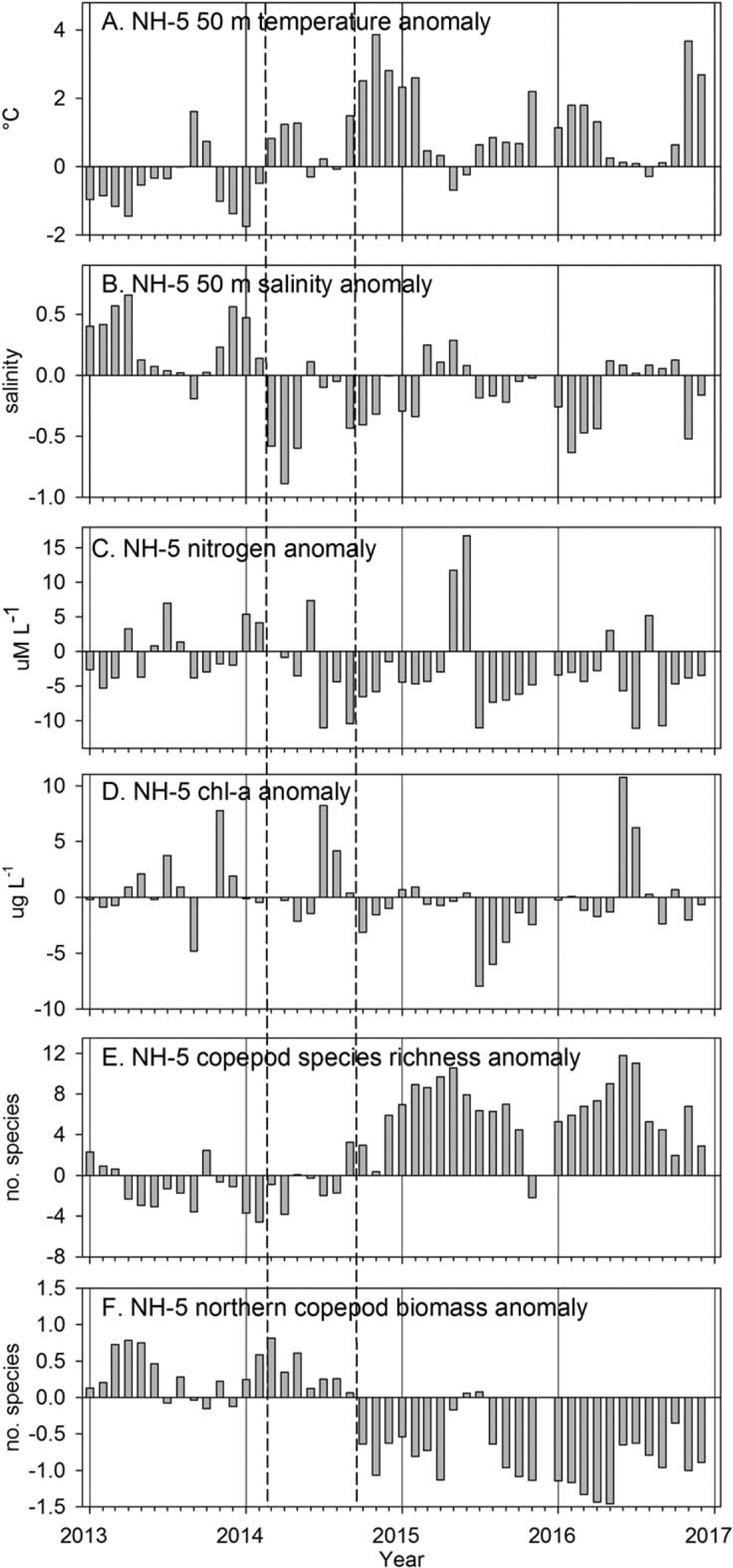
Monthly anomalies of temperature, salinity, nitrogen concentration, chlorophyll-a, copepod species richness, and northern copepod biomass at station NH-5, from January 2013 through December 2016. Dashed vertical lines as in Figure 8. Data were not collected in December 2015.
Monthly averaged 50 m temperature anomalies at NH-5 reached +4°C by November 2014. However, from March through June 2015, northerly winds were strong, resulting in coastal upwelling, during which T and S anomalies reversed at NH-5 and surface nitrogen increased dramatically (Figures 9a–9c). Although sea level dropped slightly during this period, SSH still exhibited positive anomalies. Following this period of moderate (but early) upwelling, warm/fresh anomalies resumed and nitrogen was below average at NH-5 from July 2015 through winter 2015–2016 (Figures 9a–9c). During the summer of 2016, average upwelling winds occurred and the temperature and salinity at NH-5 were average, but surface nitrogen remained mostly negative with the exception of 1 month of positive anomalies in August.
Monthly averaged surface chlorophyll-a anomalies became negative in October 2014 (Figure 9d) with the arrival of the Blob in shelf waters. During 2015, the most negative anomalies of chlorophyll-a concentration in the 20 year time series occurred in July—in association with somewhat weakened upwelling conditions, a return to deep warm/fresh anomalies at NH-5, and very low nitrogen concentrations (Figures 6b and 9c). Anomalies of surface chlorophyll-a concentration remained weakly negative through spring 2016 but increased dramatically in response to the average summer upwelling during summer 2016.
3.6. Copepod Species Composition: 2013–2016
Although the Blob began to influence deep waters of the shelf and slope off Newport as early as March 2014, there is no evidence of any response by the copepods during this period. Anomalies of northern (southern) copepod biomass were positive (negative), and anomalies of copepod species richness were slightly negative (Figures 9e and 9f). However, when the Blob moved into shelf waters in September 2014, anomalies of the three copepod indicators changed sign within 2 weeks after the arrival of the Blob: anomalies of northern copepod biomass became negative, anomalies of southern copepod biomass became positive, and positive anomalies of copepod species richness reached +10 species by winter of 2014–2015 (Figure 9e). Peak anomalies in species richness (+13 species) were observed in April and May 2015. Monthly averaged species richness declined to +6.5 species from July to September 2015, values similar to those observed during past El Niño events (1997–1998, 2003–2005, 2009–2010; see Figure 6). In early November 2015, species richness anomalies were negative on one cruise, but increased in January 2016, and again reached unprecedented values >10 species in June 2016, and continued to be high through 2016.
While copepod species richness remained anomalously high following the September arrival of the Blob in 2014, a key finding is that the copepod community composition shifted, and remained different from October 2014 through 2016, compared to the community that was present prior to the arrival of the Blob into shelf waters off Oregon. The cluster dendrogram (Figure 10) most strongly separates the copepod community during 2014–2016 into two groups found during two periods: (1) The period prior to the intrusion of the Blob into shelf waters (January–September 2014) along with the months in 2015 and 2016 with upwelling conditions (offshore Ekman transport and equatorward winds/currents); (2) October–December 2014 when the Blob had moved into shelf waters, along with the months in 2015–2016 with downwelling conditions (onshore Ekman transport and poleward winds/currents) (Figure 10). Exceptions to these patterns occurred during transition months when winds and currents were weak and/or changing direction. For example, during October 2015 and 2016, the copepod community clustered within the upwelling community, however those months were characterized by downwelling conditions following 6 months of upwelling. Similarly, the copepod community during April 2015 and 2016 clustered within the downwelling community; however, those months were characterized by upwelling conditions following 6 months of downwelling. The copepod community during the winter downwelling season from January–April 2014, prior to the arrival of the Blob, also did not cluster within the downwelling group, further showing how the copepod community that was present after the intrusion of the Blob was even different from a typical warm winter copepod community.
Figure 10.
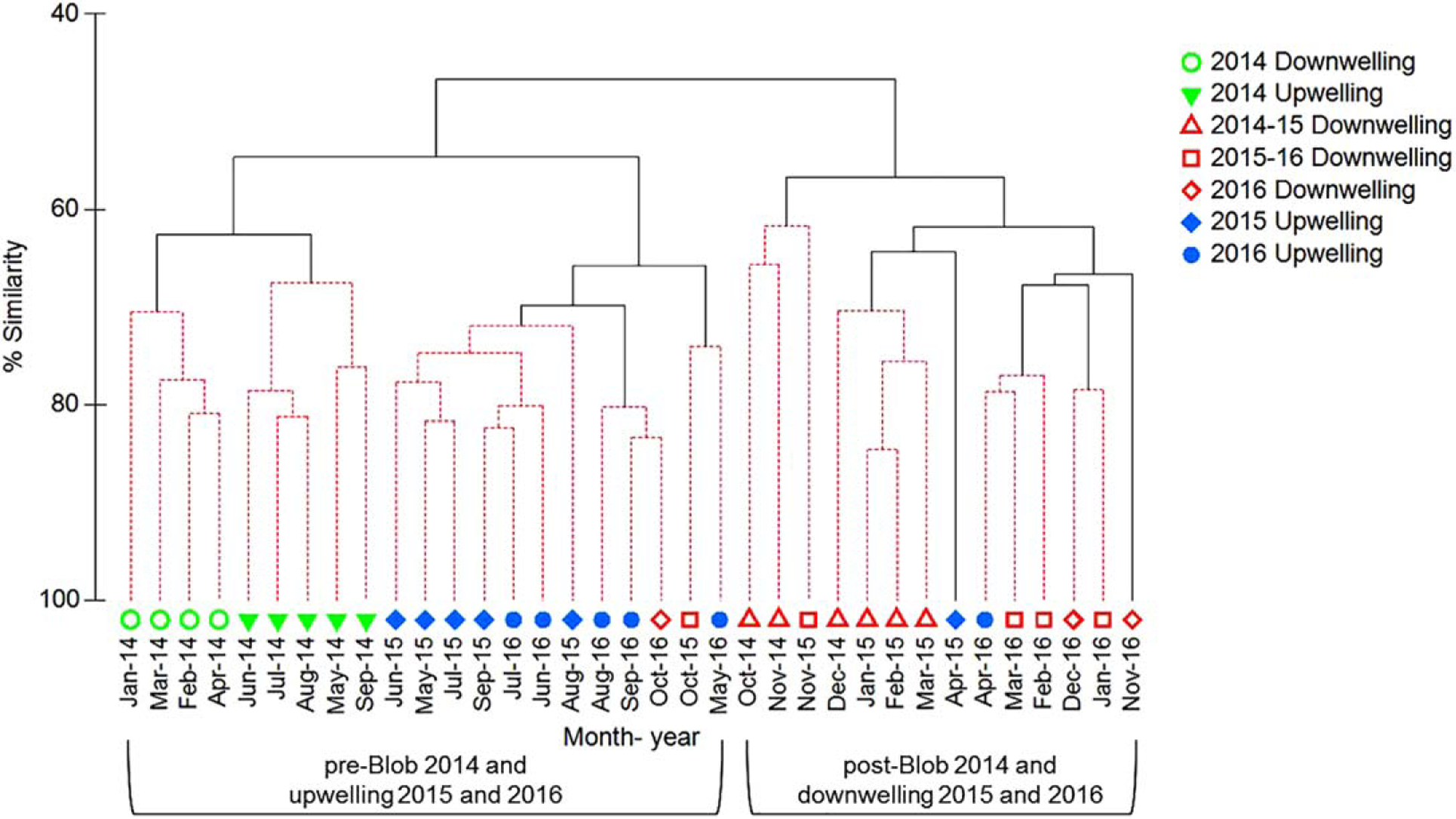
Cluster dendrogram representing changes in the copepod community monthly from 2014 to 2016. Open symbols represent downwelling and closed symbols represent upwelling. Colored symbols represent: 2014 downwelling (January–April) and upwelling (March–September) prior to the arrival of the Blob in Oregon shelf waters; downwelling from October 2014 to March 2015, October 2015 to March 2016, and October–December 2016; and upwelling during April–September 2015 and April–September 2016. Dashed lines depict no significant differences and solid lines represent significant differences (α=0.01). Data were not collected in December 2015.
The shifts in the copepod community that occurred during 2014–2016 were accompanied by the presence and absence of rare and unusual species with different water mass affinities. When the Blob moved into continental shelf waters in September 2014, accompanied by strong poleward flow until February 2015, many species arrived during that period that are rare, or to the best of our knowledge, have never occurred before in shelf/slope waters in the NCC (Table 2). With the exception of Clytemnestra rostrata that was collected January 2013, the species listed in Table 2 were not collected in 2013, or in 2014, prior to the arrival of the Blob. The new copepod species observed during this time are characteristic of water from the North Pacific Central Gyre with two species from unknown water mass affinities and one species from the coastal waters of Japan (Table 2). Examples include: Clausocalanus farrani, Cl. minor, Pleuromamma robusta, Subeucalanus crassus, Temora discaudata, Scolecithricella globulosa, Scaphocalanus echinatus, and Labidocera euchaeta. An additional six species were also new to the Oregon coast but these additional six species could not be identified because they are juvenile stages that lack the adult characteristics required for identification using taxonomic keys. Species that are not new occurrences but are rare in the shelf/slope waters in the NCC were also found during this period. Examples include Clausocalanus furcatus, Eucalanus hyalinus, Pleuromamma xiphias, Scolecithricella dentata, and Clytemnestra rostrata. During May–September 2015, the copepod community shifted and many of these rare/unusual species disappeared (Figure 10 and Table 2). During this period, upwelling winds and equatorward currents were strong to moderate, and shelf waters were slightly cold/salty (Figures 8b and 8c). This early upwelling and offshore Ekman transport likely shifted many of the unusual species offshore and away from our study region.
Table 2.
Timing of First Occurrence (*) or Rare Copepod Species Observed During the Warming of 2014–2016 and Their Water Mass Affinities [Mori, 1937; Fleminger, 1967; Frost and Fleminger, 1968; Hernandez-Trujillo, 1999; Jimenez-Perez and Lavaniegos, 2004]a

|
Fourteen new species were observed and at least six additional species were also collected, but remain unidentified because each was an immature stage that cannot be identified using taxonomic keys. Data were not collected in December 2015.
During fall 2015, the copepod community shifted once more (Figure 10), and species present during the Blob returned, along with an additional suite of rare/unusual species to the NCC that were not found during the earlier period. To the best of our knowledge, six of these species are first occurrences to the region and include Acartia negligens, Paracalanus indicus, Calocalanus pavoninus, Centropages spp., Acartia pacifica, and A. omorii. Other rare taxa only observed following the Blob period include Scolecithricella ovata, Mecynocera clausi, and several other species of Oncaea (species unidentified). Most of these species are known only as “coastal tropical” or “warm-water” species however two species are known from the coastal waters of Japan.
Other copepod species with known warm-water affinities that are characteristic of coastal waters to the south of Oregon, also appeared throughout the warm period from September 2014 through 2016. These warm-water species occur commonly off Oregon in winter months when the Davidson Current is strong and during periods of positive PDO and El Niño events. Examples include Calanus pacificus, Rhincalanus nasutus, Calocalanus styliremis, Clausocalanus lividus, Clauso. paululus, Clauso. jobei, Centropages bradyii, Acartia tonsa, and Corycaeus anglicus.
3.7. Phytoplankton Species Composition
Similar to changes in the copepod community, the phytoplankton community also shifted following the arrival of the Blob. Monthly anomalies of diatom species richness became persistently positive starting in July 2013 (Figure 11a) and remained mostly positive through mid-2016 (significant negative anomalies appeared only in February–March 2014, and August–September 2015). The highest anomalies occurred during March–April 2016. Dinoflagellate species richness anomalies did not become persistently positive until October 2014 (Figure 11b), coincident with the onshore intrusion of the Blob, remained positive throughout 2015 (with the exception of July), and increased markedly in March and April 2016, similar to the increases in diatom species richness.
Figure 11.
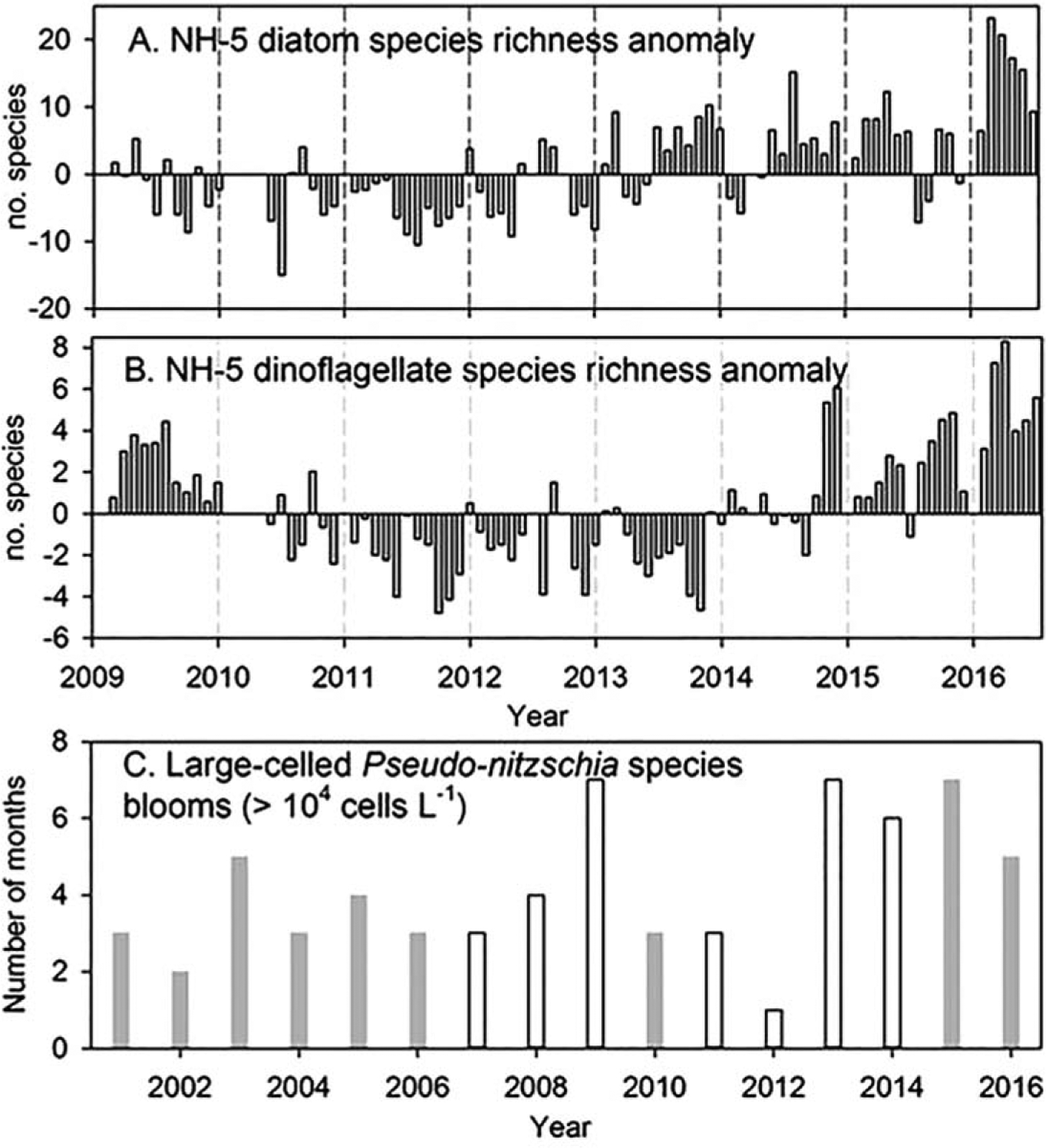
Diatom and dinoflagellate species richness (top) and number of months within a year when a bloom of the diatom Pseudo-nitzschia was observed since 2001 (bottom). Gray bars indicate when blooms were toxic within a year; white bars indicate that the blooms were not toxic. Data were not collected in December 2015.
While the species composition of the diatom community was dominated by a group of cold-water species (e.g., Thalassiosira spp.) in 2014–2016, there were decreases in the abundance (81%) and frequency of occurrence (9%) in the cold-water species Chaetoceros affinis, while there were increases in the abundance (73–92%) and frequency of occurrence (12–30%) for other cold-water taxa (Ch. similis, Ditylum brightwelli, Proboscia alata, and Rhizosolenia setigera).
A diverse set of warm-water phytoplankton species were also recorded in 2013–2016. As examples, warm-water diatoms Chaetoceros anastomosans and Stephanopixis palmeriana appeared in August 2013 and Hemiaulus hauckii in October and November 2013. Two neritic and southern temperate species, S. palmeriana and Chaetoceros didymus var. protuberans were observed in July 2014. After the Blob intruded onto the Oregon shelf, warm-water (oceanic or neritic) species were immediately observed: Ch. dadyi, Ch. densus, H. hauckii, Rhizosolenia calcar-avis, and Eucampia cornuta. The warmer conditions off Oregon may have favored growth of other diatom species that were either local (but uncommon) or that were advected from the west and south. These species include Chaetoceros curvisetus, Ch. didymus (and var. protuberans), Detonula pumilar, Rhizosolenia styliformis, and S. palmeriana, each of which became relatively more frequent and abundant in 2014–2016.
Unlike the increase in abundance of some warm-water diatom species, warm-water dinoflagellates were rarely observed in 2014–2016. However, there was an increase in frequency of occurrence of species commonly seen in the Northern California Current such as species in the genera Prorocentrum (minimum and gracile), Amphidinium (sphenoide), Ceratium (furca and lineatum), and Protoperidinium (e.g. depressum). Their frequent occurrence during 2014–2016 resulted in persistently high dinoflagellate species richness (Figure 11b).
Along with the increases in species richness of both diatoms and dinoflagellates from March–June 2016, a suite of new warm-water diatoms appeared: Chaetoceros atlanticus var. neapolitana (April), Ch. peruvianus (March–May), Odontella mobiliensis (March), Planktoniella sol (March), and Thalassionema frauenfeldii (April and June). In addition, three warm-water dinoflagellates were observed in March–April 2016: Ceratium azoricum, C. pentagonum, and Dinophysis tripos. All of the above warm-water species were defined based on published records showing historically higher abundance in waters off the Southern California coast [Cupp, 1943] in contrast to their rare appearance in our long-term observations off Newport.
The warm ocean conditions in 2014–2016 also contributed to prolonged blooms of the diatom Pseudo-nitzschia (PN). Blooms of this species have occurred in every year of our 16 year time series [e.g., McKibben et al., 2017] but PN were particularly dominant and persistent from 2013 through 2016 (Figure 10c). The toxic PN blooms off Newport in 2015 were the most prolonged (late-April through October 2015) and among the most toxic blooms observed [Du et al., 2016]. This bloom was also spatially extensive, from Southern California to Washington State [McCabe et al., 2016] and into the Gulf of Alaska [McClatchie et al., 2016].
4. Discussion
4.1. Ocean Conditions During the Past 20 Years
Over the past 20 years, temperature and salinity anomalies of bottom waters on the inner-middle shelf (at station NH-5) have followed the PDO and ONI indices on interannual and decadal timescales (Figures 3 and 4). SSH and SST also track these basin-scale indices, although with more high-frequency variability superimposed on the longer-period changes. The general pattern is for higher sea level and relatively warm and fresh water during positive phases of the PDO/ONI and lower sea level and cold/salty water during negative phases. Copepods also track the PDO/ONI, but with variable time lags [Fisher et al., 2015]. Nitrogen and chlorophyll-a do not track the PDO as closely, which is not surprising considering that their variability is controlled more strongly by local winds at the upwelling event timescale, as well as by rates of primary production and grazing. However, low nitrogen concentrations generally follow positive ONI values.
4.2. Ocean Conditions During 2014–2016: Characteristic Physical Features of the Blob and Previous Warm Events
Beginning in 2014, the Blob was widespread, occupying nearly the entire northern portion of the North Pacific. Past warm events associated with either the warm phase of PDO or with ENSO were neither as wide-spread nor as warm, and were generally seen only along the eastern boundary of the northeast Pacific Ocean. Moreover, the atmospheric conditions that gave rise to the Blob in the Gulf of Alaska were quite different from past warm events that are related to the PDO/ONI cycles [DiLorenzo and Mantua, 2016]: warm events are usually associated with a strong Aleutian Low in winter and strong poleward winds along the west coast of the United States [e.g., Amaya et al., 2016]. This was not the case in either the winter preceding the formation of the Blob (November 2012 to March 2013) or the following winter (November 2013 to March 2014). The Aleutian Low pressure index had a negative anomaly during both of these periods, indicating a weak Aleutian Low [Surry and King, 2015], and in fact during winter 2013–2014, the Aleutian Low had anomalously high pressure, on the order of 10 mbar above climatology [Bond et al., 2015].
Other physical characteristics observed during 2014–2016, in comparison to El Niño periods include the following: (i) After the upwelling in mid-2014 subsided, the strongest positive temperature and negative salinity anomalies off Newport associated with the Blob in 2014–2015 (Figure 5) were concentrated in the upper 50–80 m of the water column, whereas during the 1997–1998 El Niño event, waters as deep as 150 m show the “typical” warm/fresh signal; (ii) SST values in late 2014 were nearly the warmest on record (Figures 3 and 8); (iii) at the same time, temperatures at a depth of 50 m at the shelf station NH-5 were the warmest in at least 20 years (Figure 4); (iv) positive SSH anomalies persisted for at least 34 months (March 2014 through at least December 2016) as compared to only 14 months during the 1997–1998 El Niño event (Figure 3).
4.3. Hydrography and Nitrogen in 2014–2016
The large-scale processes in the North Pacific during 2014–2016 include the formation and eastward spread of the Blob during 2013–2016 and the occurrence of a strong (in SST) equatorial El Niño. These large-scale processes interact with the more local/regional processes involved with seasonal cycles of upwelling and downwelling (alongshore winds, cross-shelf and alongshore currents, etc.)
The offshore warm anomaly had the earliest and most direct effect on the Oregon coast. Having formed in the Gulf of Alaska during November 2013 to March 2014, the Blob spread eastward and came close to the coast of North America off Oregon and Washington in March–May 2014 (Figure 1). It clearly influenced sea level and hydrography of deep waters of the shelf and slope off central Oregon. Strong northerly winds in June–August 2014 created offshore Ekman transport that kept the surface expression of the Blob offshore. However, the deeper return flow associated with upwelling drew Blob water onshore beginning in March–April 2014, creating warm and fresh anomalies in the deep waters on the shelf at NH-5 and on the slope at NH-25. Although SSH over the shelf usually drops when cold/salty water replaces shelf water during upwelling, the SSH anomaly during March 2014 became positive, reflecting the replacement of the surface offshore transport by warm and fresh water. All of these indicators are consistent with the arrival of water from the Blob near the coast of Oregon and Washington during March 2014, but kept offshore at the surface by the relatively strong Ekman transport during June–August 2014, while being drawn onshore at depth. The Blob reached the continental shelf waters off Oregon at the sea surface in mid-September 2014, after equatorward winds weakened. Winds became anomalously poleward in October, enhancing onshore transport of the Blob to the continental shelf. At the same time, the warm anomalies off south/central California spread north to create a continuous band of warm water next to North America (Figure 1) [Gentemann et al., 2017]. As the warm anomalies coalesced along the USWC, cooling in the central North Pacific in late 2014 created an SST anomaly pattern that resembled the expression of the Pacific Decadal Oscillation (Figure 1). However, DiLorenzo and Mantua [2016] refer to this as the “ARC pattern” rather than “positive PDO pattern,” since it was not driven by a strong Aleutian Low in winter, which contributes to the “usual” positive PDO pattern.
During the summers of 2014–2016, the expression of upwelling (the presence of cold salty water with high nitrogen content in shelf waters) was weak during most months of the upwelling seasons off Oregon, despite some of the strongest northerly winds observed in 20 years. In both 2015 and 2016, apart from two or three upwelling events, the surface waters at the NOAA Buoy 46050, 20 miles from shore (44.6°N), seldom cooled. During summer of 2015, the SST fields (Figure 1) show the continued influence of the warm anomaly in the offshore waters along the USWC, moving back to the coast following the most intense upwelling during April–June [Gentemann et al., 2017]. This summer is also the period when a strong equatorial El Niño developed, raising the possibility of its influence on anomalous conditions along the USWC. Early and strong upwelling winds in April and June 2015 resulted in strong equatorward currents and cool SST anomalies by May–June on the central coast of Oregon (Figure 8). Bottom conditions at NH-5 became cool/salty, with extremely high nitrogen anomalies, typical of upwelling conditions. During July–September, equatorward winds decreased to normal summer values, while SST anomalies rose again to +2 to 3°C and bottom conditions at NH-5 returned to the anomalous warm-fresh, atypical of upwelling. Concentrations of nitrogen and chlorophyll-a also became extremely negative at this time. Like the winds, the alongshore currents decreased but remained equatorward, although their decrease was much more dramatic than the winds.
The two processes that could contribute to the hydrographic changes during July 2015 are an onshore return of the persistent offshore warm anomaly during the wind relaxation and/or a northward expansion of El Niño anomalies from the California coastal ocean. The onshore movement of surface water during upwelling relaxations is a well described process [Halpern, 1976; Kosro, 1987; Austin and Barth, 2002] along the USWC and is the explanation favored by Gentemann et al. [2017] for the large-scale California Current during 2015. The possible contribution of an El Niño signal off Oregon is less clear. Using a realistic basin-scale ocean circulation model, Frischknecht et al. [2017] attribute distant forcing (El Niño) as the cause of warm anomalies in a 50 km wide band along the central California coast during June–October 2015. Analyses of glider and model data along Southern and central California by Jacox et al. [2016] and Rudnick et al. [2016], show a depression of isopycnals; however, the strength of the depression is only approximately half that observed during the 1982–1983 and 1997–1998 El Niños. Jacox et al. [2016] and L’Heureux et al. [2017] find a similarly weak subsurface signal at the equator (depression of the 20°C isotherm) during 2015. Thus, any signal arriving from California and affecting central Oregon is expected to be weak.
We leave as an open question whether the increase in SSH, decrease in equatorward velocities, and return to warm/fresh bottom conditions and negative nitrogen and chlorophyll-a concentrations in 2015 were due to the onshore return of the Blob or northward progression of a weak El Niño. Both could have contributed, as is implied off California by the studies referenced above. What is more important for the discussion of zooplankton species distributions, however, is the direction of the alongshore and cross-shelf transports. Increases in SSH and depressions of the pycnocline/thermocline do not transport populations of plankton. Winds remained equatorward (offshore Ekman transport) through September 2015 and became strongly poleward (onshore Ekman transport) only in late November–December 2015. Thus, the earliest we might expect water masses and populations to arrive off Oregon from the south would be November 2015.
The hydrographic conditions that began in July 2015 continued during the 2015–2016 fall-winter downwelling period and were enhanced by stronger than usual poleward winds and currents from December 2015 through February 2016. Conditions continued to include warm SST, warm/fresh bottom conditions at NH-5, and negative nitrogen and chlorophyll-a concentrations at the same location. These returned to normal values in May 2016 and persisted during a 6 month period of upwelling-favorable winds. However, at the end of 2016 (October–December) conditions were similar to the downwelling period of fall-winter 2015–2016.
4.4. Phytoplankton and Copepods in 2014–2016
The movement of the Blob into continental shelf waters in autumn 2014 clearly transformed the lower trophic levels of the food chain. Species richness anomalies of dinoflagellates increased from +1 to +6 species, as did that of diatoms, a trend that continued in 2015, with further peaks in species richness of both dinoflagellates and diatoms from February through June 2016. New species of warm-water dinoflagellates and diatoms were seen at this time (winter 2016), following strong sea level anomalies and wind-driven poleward currents. Although these species are historically found in coastal waters off Southern California [Cupp, 1943], the water mass affinities of many diatom species are not well known. Based on descriptions of phytoplankton occurring throughout the California Current [Balech, 1960; Bollin and Abbott, 1963; Garrison, 1979; Horner, 2002], our best conclusion from the diatom species distributions is that many of the “warm-water” taxa found off Oregon during 2015–2016 could be from an offshore oceanic source, a southern (coastal) source, or a combination of both.
When the Blob intruded onto the Oregon shelf in the fall of 2014, the copepod community also shifted dramatically. Most of the cold-water copepod species, which dominated the shelf for the previous four years when the PDO was in negative phase (2010–2013), disappeared and were replaced by warm-water copepod species beginning in late September 2014. The process of replacement of one community with another off Oregon occurs whenever the PDO/ONI change sign, and is usually attributed to changes in alongshore transport [Keister et al., 2011; Bi et al., 2011], but with a variable time lag. However, the prolonged persistence of a warm-water copepod community for over 2 years is unprecedented, and greatly exceeds previous warm events such as the 1997–1998 El Niño when a warm copepod community persisted off Oregon for 17 months [Fisher et al., 2015]. These copepods lack lipid reserves used for overwintering, thus greatly reducing the transfer of energy to higher trophic levels. From studying the ecosystem recovery time from the past 10 warm events (El Niño), we know that the intensity and duration of the warm event linearly track the time it takes for the copepod community to transition back to the normal seasonal cycle [Fisher et al., 2015]. While the physical forcing leading to the Blob might be different than ENSO, the impacts on the pelagic ecosystem are similar, leading to a lipid deplete pelagic food chain. Therefore, we can infer that the recovery time from this prolonged warm event will exceed previous events we have studied over the past 20 years.
Changes in the copepod community during 2014–2016 tracked changes in the local forcing. Community shifts were especially sensitive to the direction of alongshore winds (noting that cross-shelf Ekman transports are associated with the alongshore winds) and alongshore currents, albeit with a 1 month lag. The copepod community exhibited a regular shift from a winter downwelling community to that of a summer upwelling community in May 2014, prior to the arrival of the Blob on the Oregon shelf. However, once the Blob arrived, the community shifted and remained different from October 2014 through December 2016, with a strongly different copepod community emerging when alongshore currents were poleward. While the copepod community did shift each summer during upwelling conditions, that community was not indicative of a normal summer community, as observed during 2014, with cold neritic lipid rich taxa [Hooff and Peterson, 2006]. Further, the copepod community that was present during the summer in 2015 and 2016 also differed from the 2014 winter community, signaling that the Blob altered the copepod community beyond the influence of previous basin scale warm events such as El Niño or positive PDO periods.
The shift in the local hydrography during late summer 2015 occurred 4 months prior to the shift in the copepod community. During the second half of the 2015 upwelling season (July–September), equatorward winds were average, alongshore currents were weak, and local conditions returned to warm/fresh while nitrogen and chlorophyll-a anomalies became strongly negative. However, the copepod community did not shift during this period. It was not until November 2015 when the copepod community changed significantly, coincident with strong poleward currents and increased SSH. While this timing falls within the lag of a biological community response to an equatorial El Niño signal (a lag of 0–8 months) [Fisher et al., 2015], the reported El Niño signal off California was weak [Jacox et al., 2016; Rudnick et al., 2016] and became even weaker during November 2015 due to anomalous equatorward winds [Frischknecht et al., 2017]. Lagrangian pathways that bring passive water parcels to the shelf are complex, involving both vertical and horizontal currents associated with transient, three-dimensional jets, meanders, eddies, and filaments. These may be poorly represented in monthly SSH, geostrophic and Ekman transports immediately adjacent to the coast. In addition, plankton species have evolved behaviors, such as vertical migration, that can maintain populations within their environments [Peterson et al., 1979]. Model studies with tracers and particle tracking would be useful to determine whether the shift in the copepod community that occurred during the fall of 2015 was from southern or offshore sources or both. The fact that many of the same species that appeared in fall-winter 2014–2015 returned during fall-winter 2015–2016 favors a less distant, offshore source.
The most notable difference in copepod species composition during the 2014–2016 period was the increased number of warm-water/tropical species (Figures 6e and 9e and Table 2), rather than any differences in the magnitude of biomass anomalies of the southern or northern copepod species or shifts in the copepod community (Figures 6c, 6d, and 10). The increase in copepod species richness is unusual in two ways. First, the number of species observed from plankton net samples was double compared to any past warm or El Niño event. Second, there were at least 14 species, that to the best of our knowledge, are first occurrences to the Oregon coast, an unparalleled observation given that the lead author (WTP) has enumerated copepods in several thousand plankton samples collected off Oregon over the course of nearly 50 years (1969–1973, summer 1974, 1977, 1978, 1983, 1990–1992, and 1996–present). Moreover, sampling has been conducted during a variety of warm and cold ocean conditions [Peterson and Miller, 1975; Peterson and Keister, 2003; Hooff and Peterson, 2006; Peterson, 2009] as well as 10 El Niño events [Fisher et al., 2015].
More first occurrence copepod species appeared during the 2014–2015 fall-winter downwelling period than during the downwelling period in fall-winter 2015–2016. The appearance of so many unusual copepods could only occur if the waters that flooded the shelf after the onshore arrival of the Blob had origins different from all past warm events. In the case of the Blob, we hypothesize that the waters which gave rise to the sudden increase in new copepod species were transported to the Oregon coast from a region farther west and/or from a more southwesterly region than any other past event, a hypothesis supported by what is known of the distribution and zoogeography of these unusual species as shown in Table 2 [Fleminger, 1967; Frost and Fleminger, 1968].
4.5. Other Changes to the Pelagic Ecosystem During 2014–2016
Krill.
The extended warming of 2014–2016 negatively affected the biomass of euphausiids off the Oregon coast. E. pacifica biomass at the shelf/slope station off Oregon has been low since 2009 (Figure 7), but biomass values dropped to near zero once Blob conditions began to affect the study area in the fall of 2014. Average lengths of adult E. pacifica during 2015 were several millimeters smaller than any other year in the time series (data not shown). The near absence of adult T. spinifera (a cold-water coastal species) off Oregon in both 2015 and 2016 is notable but not unexpected since throughout the euphausiid time series (2001–2016) T. spinifera biomass is generally low during warm-water conditions.
Doliolids.
The presence of doliolids in coastal Oregon waters is indicative of a blue-water subtropical and oligotrophic zooplankton community. These organisms are common in offshore waters (at station NH-65 located 65 nautical miles or 120 km offshore), occurring on 27 of 33 dates sampled during summer months (May–September) since 1998, with an average abundance of 115.5 m−3. They rarely occur in high abundances on the continental shelf, yet they were common over the shelf during 2015 and their abundance was similar to the long-term average at the offshore NH-65 station (99 individuals m−3 in 2015 and at NH-25 115.5 m−3 averaged over 1998–2014). These taxa feed entirely upon small (<5 μm) phytoplankton cells [Gibson and Paffenhöfer, 2000], suggesting that during this period a microbial loop-based food web was dominant in shelf waters, rather than the usual diatom-crustacean based food web typical of a coastal upwelling system.
Harmful algal blooms.
The Pseudo-nitzschia harmful algal bloom in spring 2015 was unique in that it was observed simultaneously throughout the California Current and extended into the Gulf of Alaska [McClatchie et al., 2016]. It was a prominent feature off central Oregon, recurring several times with negative economic impacts. Due to contamination by domoic acid from the bloom, the fishery for Dungeness crab (the most highly valued fishery in the Pacific Northwest) was closed off WA and OR for 1.5 months in December 2015 to January 2016. Throughout California, the crab fishery remained closed during most of 2016. Another PN toxic bloom developed in late 2016 and led to another closure of the Dungeness crab fishery off Oregon, delaying the typical opening time of 1 December 2016 to January 2017. The bloom is discussed in detail by Du et al. [2016]. The possibility that blooms will increase in frequency in a warmer ocean is further discussed by McKibben et al. [2017].
5. Conclusions
Observations made over the past 20 years along the Newport Hydrographic Line show clearly that warm ocean events disrupt the transfer of energy from lower to upper trophic levels. Warm ocean conditions observed from late September 2014 through 2016 resulted in a collapse of the usual food chain which subsequently became dominated by lipid-poor copepod species. A lack of lipids in copepods is transmitted up the food chain such that forage fishes (smelt, herring, sand lance) likely had a lower lipid content, resulting in poor feeding conditions for higher trophic level species (salmon, whiting, rockfish, some seabirds and humpback whales). Long-distance migratory fish and bird species (whiting, sardine, albatross, shearwaters, and humpback and gray whales) that migrate to the Northern California Current each summer to feed on a food chain with high fat content also encountered a lower quality food supply. Although the Blob resembled an El Niño event in terms of temperature and salinity properties, the water mass that arrived suddenly off Oregon in September 2014 came from a region different from past El Niño events, as evidenced by the species composition of the copepods. The Blob brought at least 14 species of copepods new to the shelf and slope waters off the NCC. An undetermined number of subtropical and tropical diatom species were also observed. The copepods had their origins in the subtropical North Pacific Gyre and one species is known to inhabit coastal Japanese waters. With the transition to a PDO-like SST pattern in spring 2015 [DiLorenzo and Mantua, 2016] and resumption of coastal upwelling in summer 2015, many of the unusual copepods disappeared. By fall 2015, some of these species reappeared and another suite of rare/unusual copepod species not observed during the initial 2014–2015 downwelling period arrived in the NCC. The source of the water mass during this period is unknown, but most likely included contributions from both offshore and southern sources.
The superposition of strong upwelling on a warmer coastal ocean environment in 2015 and 2016 may serve as a model for the future. The persistence of the warm-water planktonic species through the periods of strong upwelling in 2015 and 2016 suggests that even an increase in wind-forced upwelling in the future may not be able to replace a warm-water zooplankton ecosystem with a more lipid-rich cold-water species assemblage, especially during extended periods of warming. These prolonged events could result in even greater negative economic impacts on commercially important higher trophic levels than past warm events.
This paper demonstrates the power of the combination of physical data (in situ and remotely sensed) and plankton species identification. The physical data can be collected relatively routinely using electronic sensors deployed during brief cruises. However, the species identification requires the collection of water samples at sea under all conditions and careful identification of species (presently requiring significant effort by humans in the lab). Moreover, it needs to be done over a wide geographic region on a systematic sampling schedule. Thus, it requires coordinated monitoring programs by multiple institutions. If we are to understand how the marine ecosystem is responding to a changing ocean in the future, as is needed for ecosystem-based management of our fisheries, such monitoring systems must be maintained.
Key Points:
The unprecedented warm plankton community persisted for nearly three years altering the food chain for upper trophic levels
Persistent warm water events provide natural experiments that serve as proxies of potential future impacts on pelagic ecosystems
Long-term ecological observation programs must be maintained so that ecosystem impacts of future change can be understood
Acknowledgments
None of the work described here would have been possible without the generous support to WTP over the past 20 years from the U.S. GLOBEC and CAMEO programs, the NOAA FATE, SAIP, and BiOP programs, the Bonneville Power Administration, the NOAA/NCCOSS/MOCHA program, and the NSF/RISE project. P.T.S. and C.R. acknowledge funding from NASA grant NNX13AH22G. This publication is a contribution to the California Current Integrated Ecosystem Assessment. We thank Moira Galbraith, Fisheries and Oceans Canada (Sydney BC) for identifying some of the rare copepod species found in our samples. We are extraordinarily thankful for the website created by Claude Razouls (http://copepodes.obs-banyuls.fr/en/index.php) because species descriptions compiled here made efficient the identification of copepods new to our region. We thank Mike Jacox and an anonymous reviewer who provided detailed comments that greatly improved this manuscript. The data used are listed in the references, tables, figures, and URLs in the text.
References
- Amaya DJ, Bond NE, Miller AJ, and De Florio MJ (2016), The evolution and known atmospheric forcing mechanisms behind the 2013–2015 North Pacific warm anomalies, US CLIVAR Variations, 14(2), 1–6. [Google Scholar]
- Austin JA, and Barth JA (2002), Variation in the position of the upwelling front on the Oregon shelf, J. Geophys. Res, 107(C11), 3180, doi: 10.1029/2001JC000858. [DOI] [Google Scholar]
- Balech E (1960). The changes in the phytoplankton population off the California coast, Calif. Coop. Oceanic Fish. Invest. Rep, 7, 127–132. [Google Scholar]
- Bi H, Peterson W, and Strub P (2011), Transport and coastal zooplankton communities in the northern California Current system, Geophys. Res. Lett, 38, L12607, doi: 10.1029/2011GL047927. [DOI] [Google Scholar]
- Bollin R, and Abbott D (1963), Studies on the marine climate and phytoplankton of the central coastal area of California 1954–1960, Calif. Coop. Oceanic Fish. Invest. Rep, 9, 23–45. [Google Scholar]
- Bond NA, Cronin MF, Freeland H, and Mantua N (2015), Causes and impacts of the 2014 warm anomaly in the NE Pacific, Geophys. Res. Lett, 42, 3414–3420, doi: 10.1002/2015GL063306. [DOI] [Google Scholar]
- Brinton E, and Townsend A (2003), Decadal variability in abundance of the dominant euphausiid species in southern sectors of the California Current, Deep Sea Res., Part II, 50, 2449–2472. [Google Scholar]
- Clarke KR, and Warwick RM (2001), Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed, PRIMER-E, Plymouth, U. K. [Google Scholar]
- Cupp EE (1943), Marine Plankton Diatoms of the West Coast of North America, 237 pp., Univ. of Calif. Press, Berkeley. [Google Scholar]
- DiLorenzo E, and Mantua N (2016), Multi-year persistence of the 2014/15 North Pacific marine heatwave, Nat. Clim. Change, 6, 1042–1047, doi: 10.1038/NClimate3082. [DOI] [Google Scholar]
- Du X, Peterson W, and O’Higgins L (2015), Interannual variations in phytoplankton community structure in the northern California current during the upwelling seasons (2001–2010), Mar. Ecol. Prog. Ser, 519, 75–87. [Google Scholar]
- Du X, Peterson W, Fisher J, Hunter M, and Peterson J (2016), Initiation and development of a toxic and persistent Pseudo-nitzschia bloom off the Oregon coast in spring/summer 2015, PLoS One, 11(10), e163977, doi: 10.1371/journal.pone.0163977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fager E, and McGowan JA (1963), Biological species, water masses and currents, in The Sea II, edited by Hill MH, pp. 381–414. [Google Scholar]
- Fisher JL, Peterson WT, and Rykaczewski RR (2015), The impact of El Niño events on the pelagic food chain in the northern California Current, Global Change Biol., 21, 4401–4414. [DOI] [PubMed] [Google Scholar]
- Fleminger A (1967), Distributional atlas of calanoid copepods in the California Current region, Part II, Calif. Coop. Oceanic Fish. Invest. Atlas, 7, 1–213. [Google Scholar]
- Frischknecht M, Münnich M, and Gruber N (2017), Local atmospheric forcing driving an unexpected California Current System response during the 2015–2016 El Niño, Geophys. Res. Lett, 44, 304–311, doi: 10.1002/2016GL071316. [DOI] [Google Scholar]
- Frost B, and Fleminger A (1968), A Revision of the Genus Clausocalanus (Copepoda: Calanoida) With Remarks on Distributional Patterns in Diagnostic Features, Bull. Scripps Inst. Oceanogr, vol. 12, 235 p., Univ. of California Press, Berkeley. [Google Scholar]
- Garrison DL (1979), Monterey Bay phytoplankton I. Seasonal cycles of phytoplankton assemblages, J. Plankton Res, 1(3), 241–265, doi: 10.1093/plankt/1.3.241. [DOI] [Google Scholar]
- Gentemann CL, Fewings MR, and García-Reyes M (2017), Satellite sea surface temperatures along the West Coast of the United States during the 2014–2016 northeast Pacific marine heat wave, Geophys. Res. Lett, 44, 312–319, doi: 10.1002/2016GL071039. [DOI] [Google Scholar]
- Gibson G, and Paffenhöfer G-A (2000), Feeding and growth of the doliolid Dolioletta gegenbaurii Uljanin (Tunicata, Thaliacea),J. Plankton Res, 22(8), 1485–1500, doi: 10.1093/plankt/22.8.1485. [DOI] [Google Scholar]
- Halpern D (1976), Structure of a coastal upwelling event observed off Oregon during July 1973, Deep Sea Res. Oceanogr. Abstr, 23, 495–508. [Google Scholar]
- Hernandez-Trujillo S (1999), Variability of community structure of Copepoda related to El Niño 1982–83 and 1987–88 along the west coast of Baja California Peninsula, Mexico, FishZ. Oceanogr, 8, 284–295. [Google Scholar]
- Hooff R, and Peterson WT (2006), Recent increases in copepod biodiversity as an indicator of changes in ocean and climate conditions in the northern California current ecosystem, Limnol. Oceanogr, 51, 2042–2051. [Google Scholar]
- Horner R (2002), A Taxonomic Guide to Some Common Phytoplankton, Biopress Ltd, Bristol, U. K. [Google Scholar]
- Jacox MG, Hazen EL, Zaba KD, Rudnick DL, Edwards CA, Moore AM, and Bograd SJ (2016), Impacts of the 2015–2016 El Niño on the California Current System: Early assessment and comparison to past events, Geophys. Res. Lett, 43, 7072–7080, doi: 10.1002/2016GL069716. [DOI] [Google Scholar]
- Jiménez-Pérez L, and Lavaniegos BE (2004), Changes in dominance of copepods off Baja California curing the 1997–1999 El Niño and La Niña, Mar. Ecol. Prog. Ser, 277, 147–165. [Google Scholar]
- Keister JE, Johnson TB, Morgan CA, and Peterson WT (2005), Biological indicators of the timing and direction of warm-water advection during the 1997/98 El Niño off the central Oregon coast, USA,Mar. Ecol. Prog. Ser, 295, 43–48. [Google Scholar]
- Keister JE, DiLorenzo E, Morgan CA, Combes V, and Peterson WT (2011), Copepod species composition is linked to ocean transport in the northern California Current, Global Change Biol., 17, 2498–2511. [Google Scholar]
- Kosro PM (1987), Structure of the coastal current field off northern California during the Coastal Ocean Dynamics Experiment, J. Geophys. Res, 92(C2), 1637–1654. [Google Scholar]
- Kosro PM (2002), A poleward jet and an equatorward undercurrent observed off Oregon and northern California during the 1997–98 El Niño, Prog. Oceanogr, 54, 343–360. [Google Scholar]
- Lee RF, Hagen W, and Kattner G (2006), Lipid storage in marine zooplankton, Mar. Ecol. Prog. Ser, 307, 273–306. [Google Scholar]
- Levine AFZ, and McPhaden MJ (2016), How the July 2014 easterly wind burst gave the 2015–2016 El Niño a head start,Geophys. Res. Lett, 43, 6503–6510, doi: 10.1002/2016GL069204. [DOI] [Google Scholar]
- L’Heureux ML, Takahashi K, Watkins AB, Barnston AG, Becker EJ, Di Liberto TE, Gamble F, Gottschalck J, Halpert MS, Huang B, Mosquera-Vasquez K, and Wittenberg AT (2017), Observing and predicting the 2015/16 El Niño, Bull. Am. Meteorol. Soc, 98, 1363–1382, doi: 10.1175/BAMS-D-16-0009.1. [DOI] [Google Scholar]
- McClatchie S, et al. (2016), State of the California Current 2015–16: Comparison with the 1997–98 El Niño, Calif. Coop. Oceanic Fish. Invest. Rep, 57, 1–57. [Google Scholar]
- Mackas DL, Peterson WT, Ohman MD, and Lavaniegos BE (2006), Zooplankton anomalies in the California Current system before and during the warm ocean conditions of 2005, Geophys. Res. Lett, 33, L22S07, doi: 10.1029/2006GL027930. [DOI] [Google Scholar]
- McCabe RM, Hickey BM, Kudela RM, Lefebvre KA, Adams NG, Bill BD, Gulland F, Thomson RE, Cochlan WP, and Trainer VL (2016), An unprecedented coastwide toxic algal bloom linked to anomalous ocean conditions, Geophys. Res. Lett, 43, 10,366–10,376, doi: 10.1002/2016GL070023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKibben SM, Peterson W, Wood AM, Trainer VR, Hunter M, and White A (2017), Climate regulation of the neurotoxin domoic acid, Proc. Natl. Acad. Sci. U. S. A, 114(2), 239–244, doi: 10.1073/pnas.1606798114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McPhaden MJ (2015), Playing hide and seek with El Niño, Nat. Clim. Change, 5, 791–795. [Google Scholar]
- Mesinger F, et al. (2006), North American regional reanalysis, Bull. Am. Meteorol. Soc, 87, 343–360. [Google Scholar]
- Miller CB, Batchelder HP, Brodeur RD, and Pearcy WG (1985), Response of the zooplankton and ichthyoplankton off Oregon to the El Niño event of 1983 In El Niño North, edited by Wooster WSand Fluharty DL, pp. 185–187, Wash. Sea Grant Program, Univ. of Wash, Seattle. [Google Scholar]
- Miller JA, Peterson WT, Copeman LA, Du X, Morgan CA, and Litz MNC (2017), Temporal variation in the biochemical ecology of lower trophic levels in the Northern California Current, Prog. Oceanogr, 55, 1–12, doi: 10.1016/j.pocean.2017.05.003. [DOI] [Google Scholar]
- Mori T (1937), The Pelagic Copepoda from the Neighbouring Waters of Japan, 150 pp., Yokendo Company, Tokyo. [Google Scholar]
- Peterson W (2009), Copepod species richness as an indicator of long term changes in the coastal ecosystem of the northern California Current, Calif. Coop. Oceanic Fish. Invest. Rep, 50, 73–81. [Google Scholar]
- Peterson WT, and Keister JE (2003), Interannual variability in copepod community composition at a coastal station in the northern California Current: A multivariate approach, Deep Sea Res., Part II, 50, 2499–2517. [Google Scholar]
- Peterson WT, and Miller CB (1975), Year-to-year variations in the planktology of the Oregon upwelling zone, Fish. Bull. U.S, 73, 642–653. [Google Scholar]
- Peterson WT, and Miller CB (1977), The seasonal cycle of zooplankton abundance and species composition along the central Oregon coast, Fish. Bull. U.S, 75, 717–724. [Google Scholar]
- Peterson WT, and Schwing FB (2003), A new climate regime in Northeast Pacific ecosystems, Geophys. Res. Lett, 30(17), 1896, doi: 10.1029/2003GL017528. [DOI] [Google Scholar]
- Peterson WT, Miller CB, and Hutchinson A (1979), Zonation and maintenance of copepod populations in the Oregon upwelling zone, Deep Sea Res., Part A, 26(5), 467–494. [Google Scholar]
- Peterson WT, Keister JE, and Feinberg LR (2002), The effects of the 1997–98 El Niño/La Niña events on hydrography and zooplankton off the central Oregon coast, Prog. Oceanogr, 54(1), 381–398. [Google Scholar]
- Peterson WT, Fisher JL, Peterson JO, Morgan CA, Burke BJ, and Fresh KL (2014), Applied fisheries oceanography: Ecosystem indicators of ocean conditions inform fisheries management in the California Current, Oceanography, 27(4), 80–89. [Google Scholar]
- Peterson WT, Robert M, and Bond N (2015a), The warm Blob conditions in the Northeastern Pacific Ocean, PICES Press, 23(1), 36–38. [Google Scholar]
- Peterson WT, Robert M, and Bond N (2015b), The warm Blob continues to dominate the ecosystem of the northern California Current, PICES Press, 23(2), 44–46. [Google Scholar]
- Peterson WT, Bond N, and Robert M (2016), The Blob (Part three) Going, going, gone? PICES Press, 24(1), 46–48. [Google Scholar]
- Reynolds RW, Smith TM, Liu C, Chelton DB, Casey KS, and Schlax MG (2007), Daily high-resolution-blended analyses for sea surface temperature, J. Clim, 20, 5473–5496. [Google Scholar]
- Risien CM, and Chelton DB (2008), A global climatology of surface wind and wind stress fields from eight years of QuikSCAT scatterometer data, J. Phys. Oceanogr, 38, 2379–2413. [Google Scholar]
- Risien CM, and Strub PT (2016), Blended sea level anomaly fields with enhanced coastal coverage along the U.S. West Coast, Sci. Data, 3, doi: 10.1038/sdata.2016.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rudnick DL, Zaba KD, Todd RE, and Davis RE (2016), A climatology of the California Current System from a network of underwater gliders, Prog. Oceanogr, 154, 64–106, doi: 10.1016/j.pocean.2017.03.002. [DOI] [Google Scholar]
- Shaw CT, Peterson WT, and Feinberg LR (2010), Growth of Euphausia pacifica in the upwelling zone off the Oregon coast, Deep Sea Res., Part II, 57, 584–593, doi: 10.1016/j.dsr2.2009.10.008. [DOI] [Google Scholar]
- Strub PT, Allen JS, Huyer A, Smith RL, and Beardsley RC (1987), Seasonal cycles of currents, temperatures, winds and sea level over the NE Pacific continental shelf: 35°N to 48°N, J. Geophys. Res, 92, 1507–1526. [Google Scholar]
- Strub T, and James C (2002a), Altimeter-derived surface circulation in the large-scale NE Pacific Gyres. Part I. Seasonal variability, Prog. Oceanogr, 53, 163–183. [Google Scholar]
- Strub T, and James C (2002b), Altimeter-derived surface circulation in the large-scale NE Pacific Gyres. Part 2. 1997–1998 El Niño anomalies, Prog. Oceanogr, 53, 185–214. [Google Scholar]
- Surry AM, and King JR (2015), A new method for calculating the ALPI: The Aleutian Low pressure index, Can. Tech. Rep. Fish. Aquat. Sci, vol. 3135, v + 31 p., Pacific Region Pacific Biological Station, Columbia. [Google Scholar]
- Whitney FA (2014), Anomalous winter winds decrease 2014 transition zone productivity in the NE Pacific, Geophys. Res. Lett, 42, 428–431, doi: 10.1002/2014GL062634. [DOI] [Google Scholar]