

Summary
Cellular cheating leading to cancers exists in all branches of multicellular life, favoring the evolution of adaptations to avoid or suppress malignant progression, and/or to alleviate its fitness consequences. Ecologists have until recently largely neglected the importance of cancer cells for animal ecology, presumably because they did not consider either the potential ecological or evolutionary consequences of anticancer adaptations. Here, we review the diverse ways in which the evolution of anticancer adaptations has significantly constrained several aspects of the evolutionary ecology of multicellular organisms at the cell, individual, population, species, and ecosystem levels and suggest some avenues for future research.
Subject Areas: Biological Sciences, Evolutionary Biology, Evolutionary Ecology, Cancer Systems Biology
Graphical Abstract
Biological Sciences; Evolutionary Biology; Evolutionary Ecology; Cancer Systems Biology
Cancer defines a group of diseases, emerging when some individual cells in the body of metazoan organisms lose the normal cooperative behavior and become malignant, proliferating in an uncontrolled fashion, spreading from primary tumors to surrounding tissues and to distant organs, hence causing morbidity and sometimes death (Hanahan and Weinberg, 2011). This pathological process is due to clonal evolution and cell competition inside the body, being driven by Darwinian selection, that may also culminate in contagion across individuals and/or species (Ujvari et al., 2017). The story of cancer presumably begins about 1 billion years ago, at the time multicellularity appeared (Domazet-Lošo and Tautz, 2010; Athena Aktipis et al., 2015; Nunney, 2013). During this major evolutionary transition, the Darwinian unit of selection switched from individual cells to colonies of cooperative cells, yielding multicellular organisms (Szathmáry and Smith, 1995). Multicellularity, which arose independently multiple times in eukaryotes (i.e., convergent evolution) (Parfrey and Lahr, 2013), evolved because of the many advantages the colonies have gained from cooperating at the higher level of cellular organization, including, for example, increased size, functional specialization, and higher dispersal (Grosberg and Strathmann, 2007) (Bourke, 2013) (Biernaskie and West, 2015). However, the multicellular transition also came with a number of unprecedented challenges and costs. In addition to selecting for basic requirements essential for building functional multicellular bodies (e.g., cell adhesion, cell-cell communication and coordination), cells in multicellular organisms had to evolve the capacity to better propagate the shared genetic material. They achieved this at the sacrifices of giving up the individual reproductive interest (i.e., cell division), restraining resource consumption and committing “suicide” when necessary for the sake of the organism (i.e., programmed cell death) (Athena Aktipis et al., 2015). Even if the optimal evolutionary strategy for somatic cells is to cooperate for the benefit of the germline (see Hamilton, 1964), multi-level selection yields an inevitable tension between the social benefits of cellular cooperation and the benefits of the cheating individual cell. This conflict of interest between the cellular and the individual levels is the fundamental cause of cancer development in the multicellular body (Michod, 1996) (Nunney, 2013). Instead of being a distinct phenomenon, cancer remains a particular illustration of a universal problem faced by any cooperative systems (West et al., 2007): the vulnerability to exploitation by cheaters that have access to group benefits, but partially or completely withhold their fair share of contributions to those benefits (Ducasse et al., 2015).
Compared with the large number of natural cooperative systems that are routinely victims of antisocial cheats, relatively few are irreparably damaged or killed by cheating entities (cell/individual) (Gilbert et al., 2007) (Barron et al., 2001) (Van Dyken et al., 2011). This is largely because numerous protective mechanisms have evolved to suppress cheaters and/or to prevent them from exploiting the benefits of the collective (Frank, 1995) (Raihani et al., 2012). This also occurs with malignancies: since the dawn of multicellularity, cancer suppression must have been a strong selection force because individuals with uncontrolled cell proliferation would be at a severe selective disadvantage in competition with those capable of controlling cell division and eradicating malignancies to build functional multicellular bodies (Aktipis and Nesse, 2013) (Nunney, 2017). Not only were these potent tumor suppressive mechanisms initially necessary for the evolution of metazoans, they also became even more important as animals evolved large bodies and long lives (Casás-Selves and DeGregori, 2011).
As with other fitness-reducing diseases, evolutionary theory predicts that multicellular organisms are under selective pressure to: (1) avoid the source of cancer in the first instance unless it comes with significant compensatory fitness benefits, (2) prevent cancer progression once started, and finally (3) alleviate the fitness costs if further cancer development is not preventable (Ujvari et al., 2016). However, these strategies necessarily come with immediate and/or evolutionary costs that are traded against other functions (Jacqueline et al., 2017) and are thus likely to yield significant ecological and evolutionary consequences. Surprisingly, the potential for cancer defenses to constrain life history trade-offs and evolutionary trajectories has not been broadly considered by evolutionary ecologists (Muller, 2017, Aktipis et al., 2013, Brown and Aktipis, 2015). Here, we review the diverse ways in which the evolution of anticancer adaptations constrained multiple aspects of the ecology and evolution of multicellular organisms. We argue that oncogenic processes constitute a strong selective force on organisms not only because of their direct pathological consequences but also through the ongoing and persistent costs of the defenses that evolve against them.
Cancer Defenses
Selection during metazoan evolution has favored the evolution of an arsenal of mechanisms aimed at preventing the emergence of antisocial cells and/or detecting and suppressing them. These mechanisms operate at different levels, from genomes to cells and tissues, as well as the whole organism (DeGregori, 2011). These defense mechanisms (and hence their associated costs, see after) are sometimes a consequence of the different organization levels playing out and interacting, i.e., cell versus organism (Casás-Selves and DeGregori, 2011). For instance, the evolution of metazoans required the selection of mechanisms ensuring the proper development and function of complex tissues and organs, including cellular mechanisms that maintain appropriate numbers of cells within tissues. These organismal adaptations, expressed at the cells' level, also limit the chances that damaged and possibly malignant cells are propagated and therefore contribute to tumor suppression (Lowe et al., 2004). At the cell level, cell-intrinsic checks prevent cellular proliferation and invasion of surrounding tissues and organs. At the tissue and organism organization levels, other relevant interacting mechanisms exist including apoptosis, effective DNA repair, epigenetic modifications, cell cycle checkpoints, telomere shortening, tissue architecture, and immune surveillance.
These requirements to prevent inappropriate somatic cell proliferation arise with at least two observations that are relevant to consider in an evolutionary perspective. First, these protective mechanisms are most efficient before or during the reproductive age (Crespi and Summers, 2005). Second, even during these windows, these mechanisms do not systematically result in complete lesion or tumor suppression, suggesting that the costs of elimination are higher than the benefits (here tumor suppression) at ages when natural selection is most important (Hochberg et al., 2013). Theory predicts that the cost will inevitably generate indirect modifications (i.e., tradeoffs) of the host's life history traits as a result, especially between reproduction and maintenance, and/or via antagonistic pleiotropy (Ujvari et al., 2016). For example, cancer can induce substantial elevations in resting metabolism, which represents the largest component of total energy expenditure in humans and other animals. This additional cost is linked to increases in tumor size, immune response, and inflammation related to anti-cancer defenses, which in turn challenges individual energy budgets and leads to decreased activity levels (Nguyen et al., 2016) (Purcell et al., 2016).
Costs at Different Organizational Levels
Cell
Crucial in the evolution of multicellular organisms is the p53 protein, which has a vital role in controlling virtually all processes in the cell, ensuring, notably the genetic stability and uniformity of somatic cells (Chumakov, 2007) (Bargonetti and Manfredi, 2002). Indeed, p53 interferes with numerous normal cell activities including growth and division (Levine, 1997), being involved in cell cycle control (Shaw, 1996), DNA repair (Williams and Schumacher, 2016), angiogenesis regulation or other tissue functions (Farhang Ghahremani et al., 2013), and in induction of cell death (Fridman and Lowe, 2003). Depending on cell type, cell environment, and oncogenic alterations (see Vousden and Lu, 2002), p53 activation may indeed cause damaged cells in metazoans to commit suicide (i.e., apoptosis), rather than enter into competition with their healthy counterparts and/or initiating malignant tumors. The p53 protein is therefore considered a major cancer defense. Although still debated, it has been suggested that this capacity to interrupt cancer cell proliferation also interferes with the proliferation of normal cells, such as stem cells, which are needed for tissue renewal as the organism ages. Thus, the TP53 gene would exhibit antagonistic pleiotropy, having beneficial effects on tumor suppression but counterpart deleterious effects on aging, with all the fitness consequences associated with senescence (Campisi, 2002) (Ungewitter and Scrable, 2009).
A second example of antagonistic pleiotropy in cancer defense is cellular senescence. This phenomenon is protective because it limits cellular division either in case of oncogene activation or following telomere erosion (Campisi, 2001) (Wright and Shay, 2001). However, this tumor-preventive function would be beneficial primarily early in life, because it also contributes through time to a decrease of the regenerative capability of the tissues that might outweigh the initial benefit. Thus, although the number of senescent cells increases in an organ, there is a concomitant decrease in the number of cells that are available to replace damaged cells, yielding a lower regenerative capacity of body tissues and higher organismal frailty (Wright and Shay, 2001). These antagonistic pleiotropic effects can even favor cancer in old organisms (Giaimo and D’Adda di Fagagna, 2012) (Krtolica et al., 2001).
Another example of cancer defenses that paradoxically promote cancer comes from the fact that most cancers result from somatic mutations acquired during an organism's life, and only a few cancers have congenital origins. In this context, the defenses that prevent the formation of neoplasms due to somatic mutations are also preventing the inherited mutations from causing neoplasms. This limits their detrimental effect, hence allowing individuals to reproduce and those inherited mutations are passed to the offspring (Arnal et al., 2016). Natural defenses that evolve to prevent neoplasm formation and progression thus would also have concealed the effects of cancer-causing mutant alleles on fitness and concomitantly protect inherited ones from purging by purifying selection. This could be one of the reasons explaining why oncogenic congenital mutations are present in populations at a higher frequency than those predicted by the equilibrium between mutation/selection (Arnal et al., 2016).
Individual
Individuals vary both in their propensity to develop malignant cells and/or in their ability to manage them, for genetic and/or environmental reasons (Stensrud and Valberg, 2017) (Hochberg and Noble, 2017) (Doherty et al., 2017). Individuals within species typically show substantial variation in physiology, behavior, and life history traits that can affect risk of developing cancer and responses to it. For example, individuals with a tendency toward high activity rates and high metabolic rates also tend to have lower cancer risk both in humans (Biro et al., 2020) and in animals (see for example Thompson et al., 2017, Pedersen et al., 2016). Individuals and genotypes that allocate relatively more energy to immune function would also be expected to be less vulnerable (Finn, 2012), as would those that are able to maintain activity levels into later life periods (Duggal et al., 2019).
At the intraspecific level, different combinations of these two factors are likely to influence, all else being equal, inter-individual performances. For example, fitness-related traits such as reproductive investment, competitive ability, vulnerability to predators or infectious agents, and/or the ability to disperse may be differentially affected between individuals (Ujvari et al., 2016) (Vittecoq et al., 2013). For instance, individuals with a smaller energy budget due to internal or external energy limitation may be forced to reduce activity, similar to the familiar observation of lethargy in cancer sufferers (Purcell et al., 2016). As in the context of host-parasite relationships, the net result on the host phenotype will depend on the pathological costs themselves (due here to the malignant proliferation) and the costs of the defense responses (e.g., the immune response) that are known to be substantial in terms of energy consumption (Purcell et al., 2016). These interactions in the context of host-tumor interactions have been poorly studied to date but undoubtedly deserve more exploration given the omnipresence of oncogenic processes.
Vittecoq et al. (2015) reviewed evidence for the evolution of prophylactic behaviors against cancer through, for example, habitat selection in order to avoid areas polluted with natural or anthropogenic mutagens or sexual partners infected with oncogenic pathogens and/or transmissible cancers. Despite the relative high frequency of ecological contexts that are potentially associated with cancer risks, examples of behavioral adjustments involved in cancer prevention remain few. Although this could indicate a lack of suitable studies, it could also suggest that being selective with respect to habitat or sexual partners comes with costs (e.g., time, energy) that cannot be offset by the benefit of avoiding cancer initiation, unless the perception of cancer risk is strongly reliable and the resulting cancer entails significant fitness costs. The evolution of curative behaviors appears more probable than preventive behaviors, because the presence of malignancies is likely to be associated with internal cues that can be detected by the body and that accurately predict the fitness costs in the case of no response (Vittecoq et al., 2015).
Currently, the effects of subclinical cancers on the body and hence on fitness in wild animals remain unknown. If pathological costs exist, we could expect that tumor-bearing individuals will optimize their energy budget to fight the disease progression while also attempting to fuel other fitness-related activities (see, for instance, Aubert, 1999). That is, trade-offs are expected such that the most immediate “concerns,” e.g., cancer, are prioritized at the expense of reduced activity or allocation of resources to reproduction. Although self-medication has been documented in regard to infectious diseases (De Roode et al., 2013), there is actually no empirical evidence that animals specifically search for natural substances with potential effective cancer treatment properties. Also, because the thermal optimum of malignant cells is not necessarily the same as of healthy cells, animals (especially poikilotherm species) might display thermal preference as a way to limit cancer progression. If self-medication occurs, the time and the energy spent will be necessarily at the expense of other activities, and/or could potentially enhance exposure to predators and/or parasites. We could also expect strategies aimed at conserving energy (e.g., through resting and sleeping, investing less in partner choice) in order to allocate more resources into immune system efficiency and/or into parental care and/or to speed up their reproductive activities before dying or being unable to reproduce. To our knowledge, only three empirical studies have explored these issues to date. First, Arnal et al. (2017) found that experimentally induced Drosophila females harboring early stage gut malignancies alleviate the cancer fitness costs by modifying their reproductive schedule: flies with cancer reach their oviposition peak earlier than healthy females, and die earlier. In the same Drosophila system, Dawson et al. (2018) demonstrated that the social behavior of flies harboring early tumors was altered: when given the choice between a group of healthy and cancerous flies, flies with early tumors prefer to join other cancerous individuals with which they have less stressful interactions, and this significantly reduces the progression of their own tumor, thereby giving them more opportunity to mate before dying. Finally, by monitoring Tasmanian devils (Sarcophilus harrisii) harboring tumors of the transmissible cancer devil facial tumor disease (DFTD), Hamilton et al. (in press) demonstrated that DFTD negatively influences the devils' likelihood of interactions within their network, an effect that increases with increasing tumor load . The influence of oncogenic process dynamics on individual behavior and personalities remains poorly studied, primarily because reliably detecting cancer in wildlife is difficult (Madsen et al., 2017) and partly because ecologists have by tradition focused on other variables than cancer (e.g., “good genes,” parasitism) when studying inter-individual variability.
A promising direction to explore is undoubtedly sexual selection, since it is possible that the allocation of energy to activities that serve to suppress cancer (e.g., DNA repair) may trade off with the allocation of energy to early life reproductive activity (e.g., costly sexual ornaments or fighting in males) (e.g., Rubin, 2015, Wang et al., 2019). Sexual selection processes may also favor in certain cases cancer promoting oncogene alleles, as this is observed in the fish genus Xiphophorus developing melanoma (Fernandez and Morris, 2008) (Summers and Crespi, 2010). Despite significant deleterious cancer-induced effects (e.g., shorter lifespan), the oncogenic Xmrk allele persists in some natural populations of Xiphophorus, suggesting the presence of benefits early in life that outweigh late costs (i.e., antagonistic pleiotropy). Indeed, Xmrk allele is also associated to a larger body size, which increases an individual's reproductive success through mate choice and competition for mates.
Population
In addition to plastic adjustments at the individual level, the need for cancer defenses may in the long term influence the life history of populations and species (see next section). As a result of coevolution with oncogenic manifestations, individuals in certain populations may evolve a constitutive tumor-suppressive strategy in order to keep such cells at bay even in the absence of malignant cells at a given time.
Even if alternative explanations exist, a possible example of anticancer adaptations modifying life history traits in populations has been suggested by Jones et al. (2008), who showed that Tasmanian devils from populations heavily infected with the DFTD displayed an abrupt transition from iteroparity toward single breeding, with a significant proportion of individuals within the population exhibiting precocious sexual maturity. Although natural selection for less aggressive phenotypes could also be expected, it has, however, not been observed so far, presumably because aggressivity is a trait associated with increased mating and breeding success in this species (Hamede et al., 2013). Other biological models, such as sea turtles, are promising to explore within this perspective: although almost all populations worldwide are exposed to herpesviruses that cause cutaneous tumors, some populations are largely composed of tumor-bearing individuals, whereas in other populations, infected individuals are free of any clinical manifestation (Alfaro-Núñez et al., 2014). Costs and resulting trade-offs remain to be explored.
Relationships between environmental contaminants and tumor development in wildlife remain only partially understood at the moment, both because neoplasia detection in wild animals is often complex and also because several parameters intervene in the way pollution influences the development of oncogenic processes (ecological status, life history characteristics, variation in cancer defense mechanisms and/or tolerance, and also the combined effects of pollutants; see Sepp et al., 2019, Giraudeau et al., 2018). However, it remains, for instance, clearly established that effluents from aluminum smelting facilities are responsible for intestinal adenocarcinoma in beluga whales (Delphinapterus leucas) in the Saint Lawrence Estuary (Martineau et al., 1994) (Martel et al., 1986). Organochlorine contaminants are also associated to cancer in California sea lions (Zalophus californianus) (Randhawa et al., 2015). Polychlorinated biphenyls, in addition to be carcinogens themselves, also have immunosuppressive effects that in association with gammaherpesvirus enhance the risk of genital carcinomas in sea lions (McAloose and Newton, 2009) (Ylitalo et al., 2005). Increased levels of artificial radionuclides after the Chernobyl nuclear accident had a positive effect on the occurrence of external tumors in birds (Møller et al., 2013).
There are several other examples of local adaptations in animals to cope with anthropogenic pollution (Reid et al., 2016) (Whitehead et al., 2017) (Vittecoq et al., 2018). The fitness costs associated with the possession of adaptations that prevent associated deleterious effects can sometimes be indirectly assessed by examining how individuals from polluted habitats perform when placed in non-polluted conditions. For instance, in the killifish (Fundulus heteroclitus), individuals from populations living in polluted salt marshes experienced detectable costs, compared with those living in non-polluted waters, when placed in clean water (e.g., higher death rate following infection by parasites, higher rates of acute hypoxia) (Whitehead et al., 2017). Further studies are necessary to explore the extent to which species can, in polluted areas, evolve mechanisms that prevent the exposure to pollutants (e.g., birds in Chernobyl prefer to breed in sites with low radioactivity [Møller and Mousseau, 2007]), suppress the formation of malignant cells, and/or trigger their elimination.
Local selection of anticancer defenses could theoretically yield an evolved dependence situation, in which adapted individuals achieve a higher fitness in the presence, rather than in the absence, of oncogenic processes (see Thomas et al., 2018b). In the case of devils mentioned above, the fitness of individuals with precocious sexual maturity is indeed lower than other individuals in the absence of the disease, because precocial breeders usually breed much later in the year (once they reach the critical size) (Hamede, unpublished data). Offspring of precocial breeders are at disadvantage as they may not reach the minimum size to breed by the next mating season and thus miss the opportunity to generate offspring (e.g., a cumulative lag time from mother to offspring and so on). At the moment, these phenomena have been poorly considered by ecologists, despite the fact that several studies indicated that local adaptations to cope with pollution can sometimes be selected for in just a few generations in annelids, crustaceans, and fish (Klerks and Levinton, 1989) (Xie and Klerks, 2003) (Ward and Robinson, 2005) (Whitehead et al., 2012).
Species
At the species level, it has long been argued that the need to suppress somatic evolution should increase with larger body sizes and longer lifespans. Assuming that carcinogenesis occurs following mutations, and that every cell division has an identical probability to generate these mutations, large/long-lived animals that have an increased number of cell divisions should therefore have more cancers than smaller/shorter-lived ones (Nunney et al., 2015). However, evidence suggests that there is no correlation between body size, longevity, and cancer rates across species, an absence of relationship that is called “Peto's Paradox” (Peto et al., 1975) (Caulin and Maley, 2011) (Tollis et al., 2017) (Abegglen et al., 2015) (Vazquez et al., 2017). Recent studies have deciphered several genomic mechanisms at the origin of cancer suppression in species with very large body sizes. For instance, cells in African and Asian elephants (Loxodonta africana and Elephas maximus) display an enhanced DNA damage response because of 20 duplications of the tumor suppressor gene TP53 in elephant genomes (Abegglen et al., 2015) (Sulak et al., 2016). In the long-lived whale Balaena mysticetus, Keane et al. (2015) found multiple mechanisms for suppressing cancer, including positive selection on several aging and cancer-associated genes (e.g., ERCC1, which is involved in DNA repair pathway), as well as duplications of genes involved in DNA repair (e.g., PCNA) and cellular growth control (LAMTOR1) (see also Tollis et al., 2019 for an example on Megaptera novaeangliae). Although much interest has been devoted to exploring the adaptations retained by selection in large and/or long-lived species to cope with size-associated cancer problems, few studies have focused on the corollary consequences, that anti-cancer mechanism selection has always been a developmental and evolutionary constraint that limits the evolutionary trajectory of organisms (Galis and Metz, 2003). We can thus speculate that a larger size in these species potentially does not evolve because the selective advantages are outweighed by the disadvantage of exacerbated malignant problems for which there is no adapted cancer defense. The number of cervical vertebrae, generally seven in mammals, could be an illustration of this hypothesis (Galis, 1999a) (Galis, 1999b), since additional cervical vertebrae are apparently associated with increased susceptibility to pediatric cancers (Schumacher et al., 1992), whereas species such as manatees with only six cervical vertebrae appear to have extremely low cancer prevalence (Galis and Metz, 2003).
Although the mismatch concept in regard to health problems, notably cancer, has most often been discussed in regard to recent changes in human evolution (tobacco, food abundance, etc.) (Nesse, 2008), much less attention has been invested in considering the mismatches resulting from natural environmental changes. For most species, environmental biotic and/or abiotic conditions are not constant through time and/or in space. Organisms must therefore constantly evolve and may experience natural mismatches between cancer risks and cancer defenses. It has recently been hypothesized that very weak cancer defenses, instead of preventing the evolution of a given trait, may still favor the trait if (1) fitness benefits compensate the fitness costs due to associated cancers, and/or (2) if there is a concomitant selection for compensatory, potentially transient, adaptations to prevent or limit the detrimental fitness consequences of the associated cancers. For example, Leroi et al. (2003) suggested that the high rate of cancer in humans, compared with other primates could, at least partially, be explained by our recent evolutionary changes in anatomy, physiology, life history, and especially longevity (Hawks et al., 2007) (Caspari and Lee, 2004). However, despite the higher cancer risks, humans evolved these novel characteristics. At the scale of organs, a similar conclusion can be derived: although organ-specific cancer rates correlate negatively with an organ's evolutionary antiquity (i.e., cancer defenses are potentially not fixed) (Davies, 2004), they still may have evolved despite associated cancer risks. Recently, it has been argued (Thomas et al., 2019) that species experiencing such a mismatch could evolve traits that prevent or alleviate the negative cancer effects on fitness, at least until additional effective cancer defenses are selected for. This could, for instance, explain the evolution of menopause in humans and a few cetaceans. Parity, at least in women has a dual effect on breast cancer risk: early life full-term pregnancies decrease breast cancer risk in the long term, but in the short term, pregnancy also transiently increases cancer risk, because it boosts the development of oncogene-activated cells into tumors and/or promotes a metastatic cascade (Lambe et al., 2002). When, owing to recent human evolution, anticancer defenses are too weak given malignant risks, older females could not pursue reproduction without frequently initiating fatal invasive cancers nor maintain a normal reproductive physiology because pregnancy also promotes the growth of existing oncogenic processes (Figure 1). Therefore, a higher (inclusive) fitness can be achieved by ceasing their reproduction and/or going through menopause. This hypothesis implicitly assumes that these traits are easier to select than anticancer defenses, which also indirectly suggests that anticancer selection is an evolutionary constraint. Currently, in the animal kingdom, we ignore what diversity of traits may have evolved as compensatory fitness mechanisms, when other (cancer-associated) traits cannot evolve because of insufficient cancer defenses.
Figure 1.

Cancer Risk, Cancer Defense, and a Possible Scenario for the Evolution of Menopause
(A) In organisms possessing cancer defenses in alignment with cancer risks, malignant processes only slowly accumulate through time. Even if reproductive episodes, e.g., the pregnancy period in mammals, exacerbate the proliferation of existing malignant cells, they are not sufficient to trigger metastatic cancers: reproduction occurs across the lifespan and fitness is maximized.
(B–D) (B) Owing to ecological and/or evolutionary mismatches, cancer defenses are too weak given the cancer risks, oncogenic processes accumulate more rapidly, and reproductive episodes exacerbate metastatic cancer risks in aging females, resulting in a short lifespan and a low fitness. Here, natural selection can (1) favor the evolution of stronger cancer defenses in these species (i.e., as in elephants), yielding again to a situation comparable with (A), here (C), and also (2) favor females ceasing their reproduction prematurely to preserve their health (i.e., as in human females) (D). In this later situation, female's fitness is higher than in (B) because menopause permits grandparental care, which increases inclusive fitness. The (D) scenario can be just a transient situation until additional cancer defenses are selected and bring back the species to the (A)/(C) examples. Figure modified from Thomas et al., 2019.
Depending on the magnitude of the reproductive benefits associated with the acquisition of novel phenotypic traits (e.g., higher fecundity, higher size-related sexual competitiveness), the net fitness of individuals in species lacking cancer defenses may be high despite enhanced cancer risks, yielding a form of antagonistic pleiotropy that slows down the selection for greater cancer defenses. For example, early rapid growth and high fertility are often associated with higher risk of cancer (Boddy et al., 2020) (Aktipis, 2020). However, the benefits of reproduction may outweigh the cost of acquiring cancer later in life. A typical example of a species with a fast life history strategy is the African killifish, one of the shortest-lived and fastest-maturing vertebrates known that apparently does not invest into cancer defenses (Harel and Brunet, 2016). Conversely animals with slow life history strategies prioritize long-term growth and survival over short-term opportunities for reproduction and thus would be expected to invest in immune function to ensure survival to reproduction. These predictions have not been rigorously tested empirically and/or theoretically but appear promising to explain, at least in part, the differential vulnerabilities of species to cancer in relation to their life history and their recent evolutionary history. We could expect that animal species possessing high cancer rates in the wild will be species that have experienced rapid and recent evolutionary changes. Similarly, we could expect that species establishing in novel environments should be, at least transiently, exposed to a higher risk of cancer if the phenotypic changes favored in the novel habitat accentuate the mismatch between cancer risk and cancer defenses (e.g., a greater size and or longevity) (e.g., Sayol et al., 2018). We also encourage evolutionary ecologists to explore if successful invasive species have intrinsically low cancer vulnerability, and/or a good aptitude at selecting rapidly efficient anticancer defenses.
Ecosystem
Anticancer selection in natural conditions constrains evolutionary changes not only because “good mutation” appearance is a limiting factor but also largely because any potential solutions remain traded against other fitness-related functions. Immune system function is costly (Lochmiller and Deerenberg, 2000) (Purcell et al., 2016), leading to trade-offs with other demands, hence impacting life history traits (Norris and Evans, 2000) (Van Der Most et al., 2011). Therefore, to understand the evolution of anticancer selection, one must consider the complete ecological context in which animals evolve, including the community of organisms that inhabit the host (parasites, microbiota) and the ecosystem (predators, competitors). Only such an ecological approach will permit a true assessment of the selective pressures acting on anticancer selection. For example, since the Mesolithic, many domesticated species have lived in environmental conditions that are often oncogenic (inbreeding, mismatches with natural conditions) but are also not influenced by the traditional constraints of life in the wild (competition, predation, parasitism, food limitation). The relaxation of these constraints has sometimes opened the window for the selection of unprecedented anticancer defenses (Thomas et al., 2020), because domesticated animals, all things being equal, can in theory invest more resources to fuel their existing anti-cancer defenses. If costly anti-cancer defenses (resistance or tolerance) evolve, these defenses are more likely to be maintained in a domesticated setting, rather than a natural ecosystem harboring various natural enemies and competitors. A nice illustration of these phenomena is the case of melanoblastoma-bearing Libechov minipig. Although cutaneous melanoma is often a serious form of skin cancer, tumors in this pig spontaneously disappear without external influence (Vincent-Naulleau et al., 2004) (Bourneuf, 2017). More than 70% of these lesions appear on piglets before they are 3 months old, and the incidence of melanoma reaches 50% (Vincent-Naulleau et al., 2004). Spontaneous and complete tumor regression (even when at a metastatic stage) occurs on average 6 months after birth in 96% of animals and is followed by hair, skin, and iris depigmentation (Vincent-Naulleau et al., 2004). This example illustrates that, when we place a species in a novel ecosystem, with novel selective pressures, we may also alter the cost and trade-offs that normally apply in the wild and liberate the possible evolution of unprecedented solutions against malignancies (Thomas et al., 2020).
In natural ecosystems, species inhabit ecosystems alongside their cortege of predators, parasites, food sources, and competitors. The effect of the interaction between species and the abiotic environment within their ecosystem on the evolution of cancer avoidance is largely unexplored but should not be ignored since species are often exposed to those interactions over many generations. Below we summarize potential future research areas to explore and elucidate the link between ecosystem functioning and the evolution of cancer avoidance.
Interaction between species at the ecosystem level may change their exposure to cancer-causing risk factors such as UV light (UV-B) or physiological stress. During the late Precambrian, the combined production of dioxygen by blue-green algae at an ecosystemic spatial scale allowed the formation of an UV-absorbing ozone layer in the stratosphere, which paved the way for the emergence of life on emerged lands (Walker et al., 1976). Exposure to UV-B damages the DNA, compromises the immune function of animals and is recognized as a cause of skin cancers in humans (Paul and Gwynn-Jones, 2003) and other animals (e.g., fish [Sweet et al., 2012]). The amount of UV-B a species receives during its lifetime, and the associated likelihood of developing cancer, depends on the ecosystem type it inhabits. For example, engineer species such as trees or shrubs in the terrestrial ecosystems, or kelp forests in the marine ecosystem, offer shading that protects species relatively well from UV exposure (Brown et al., 1994). If a species primarily inhabits those habitats over many generations, we can then predict a lack of selection for cancer avoidance strategies associated with exposure to UV. The effect of UV light on ecosystems, and hence on cancer in terrestrial and aquatic species, is currently a developing and promising field (Paul and Gwynn-Jones, 2003).
In marine ecosystems, the respiration of organisms, the decomposition of organic matter by bacteria and the lack of water column mixing (for example, in bays and estuaries [Diaz, 2001]), or the effect of tides exposing intertidal species to air or confining them in rockpools or sediments can create chronic hypoxia (Mcmahon, 1988), associated with an increased production of DNA-damaging reactive oxygen species (ROS [Hermes-Lima and Zenteno-Savín, 2002]). Exposure to ROS is linked to an increase in cancer risk (Nechifor et al., 2012; Valko et al., 2006), which is mitigated by production of antioxidant molecules and enzymes, incurring an additional energetic cost compared with species inhabiting normoxic environments (Hermes-Lima and Zenteno-Savín, 2002). Although chronic hypoxia is currently exacerbated by anthropogenic factors (Diaz, 2001), sediment records and modeling indicates whole marine ecosystems had large areas exposed to climate-driven chronic hypoxia over geological-scale time periods (Praetorius et al., 2015; Zillén et al., 2008).
In an ecosystem, predators often have lasting non-consumptive (i.e., non-lethal) effects on their prey, altering their behavior and physiology in ways that may favor or reduce the emergence of cancer in those species (Sheriff and Thaler, 2014). For example, non-consumptive predator effect is associated with a reduction in antioxidant defenses and an increase in ROS-induced damages in damselfly larva (Enallagma cyathigerum) compared with predator-free situations (Janssens and Stoks, 2013). In the long-lived wild yellow-bellied marmot (Marmota flaviventris), exposure to high predation risk was linked to an upregulation of genes involved in cellular response to stress, DNA repair, and damage control (Armenta et al., 2019). The mitigation of predation risk as a trade-off between energy acquisition and risk of being predated upon also affect the physiology and body condition of species. For example, green sea turtles (Chelonya mydas) are known to use sub-optimal seagrass patches to avoid predation by sharks, only venturing into high-quality seagrass patch when their body condition is too low or the predators are absent (Heithaus et al., 2007). As consequence, green sea turtles experience variations in their body condition and may be less efficient at investing energy in cancer avoidance strategies (i.e., their immune system [Work et al., 2001]) when their body condition is low, for example, to prevent the emergence of life-threatening fibropapillomatosis (Jones et al., 2016; Work et al., 2001). As novel insights are obtained on the role of ecosystem functioning on cancer in wildlife species, we expect a body of evidence to accumulate on how species interactions contribute to the evolution of cancer avoidance strategies. Although the etiology of cancers in wildlife is not fully understood, a large number of oncogenic viruses have been reported in terrestrial and aquatic wildlife (McAloose and Newton, 2009) (Madsen et al., 2017). It is theoretically predicted that species that are the most vulnerable to these infectious agents should have experienced higher selective pressures to avoid them and/or to cope with their oncogenic consequences. Comparison of their biology with conspecifics in pathogen-free areas like zoos appears promising to evaluate, all things being equal, the cost of these adaptations.
Finally, because most, if not all, ecosystems on our planet are now polluted with mutagenic substances (Giraudeau et al., 2018), the differential vulnerability of species to cancer initiation/progression, coupled with their differential ability to evolve cancer defenses (e.g., Sepp et al., 2019), is likely to be responsible for alterations in biotic interaction equilibria (e.g., in a prey-predator system), yielding also to potential cascade effects. Exploring how the interplay between novel cancer risks experienced by wildlife species, cancer defense evolution, and biotic interactions can shape population dynamics and communities is an emerging topic in ecology (Perret et al., 2020; Dujon et al., 2020).
Could We Boost Cost-free Cancer Defenses in Organisms?
Natural defenses against cancer are costly primarily because the way natural selection shapes the evolution of adaptations in organisms imply trade-offs (Stearns, 2006). However, modern technologies, beyond classical therapies, may provide solutions that are not necessarily governed by the same constraints. Thus, it could be theoretically possible to alter, or cancel, some of these trade-offs in humans and/or domesticated species and provide a supplement to natural anticancer adaptations acquired through natural selection. Checkpoint inhibitors, by basically taking the “brakes” off the immune system, help to recognize and attack cancer cells. Boosting the immune system with a prophylactic cancer vaccination able to systematically eliminate precancerous/cancerous lesions are promising research directions (e.g., Yaddanapudi et al., 2012, Darvin et al., 2018, Ye et al., 2018). In parallel, there are several examples of cost-free cancer defenses in humans, involving vaccination against viral-induced cancers like hepatitis B and HPV (Schiller and Lowy, 2010). At the moment, all these perspectives are mostly limited to humans and domestic animals but could theoretically be extended in the future to other species, for instance, in conservation biology contexts (Pye et al., 2018). We cannot, however, exclude the possibility that there will be a collateral effect if this implies, for example, a constant activation of the immune system. Boosting the natural body tumor suppressor mechanisms could also unbalance system homeostasis and result in detrimental consequences such as collateral tissue damage, autoimmune complications, and nonspecific side effects. Further experiments are needed to explore this potential method for the future of cancer prevention and treatment (e.g., Thomas et al., 2018a). This area is still in its infancy concerning wildlife species, remaining the source of an important debate concerning its application and real efficiency (see Hamede et al., 2021).
Concluding Remarks
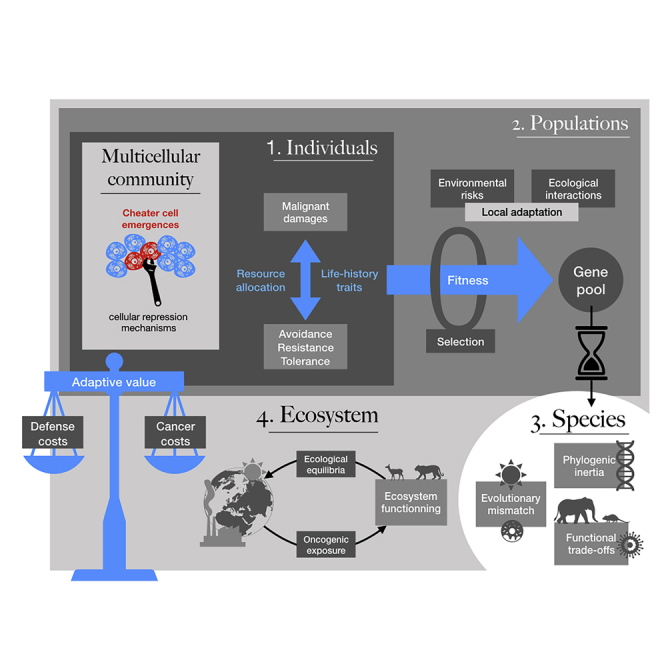
Cancer defenses are costly at multiple levels and have undoubtedly shaped the evolution of organisms at a level that is still underestimated (Figure 2). Scientists regularly try to imagine worlds “without” (e.g., microbes [Gilbert and Neufeld, 2014], parasites [Wood and Johnson, 2015], mosquitoes [Fang, 2010], mangroves [Duke et al., 2007], fires [Bond et al., 2005]) as a way of exploring the hidden ecological influence of certain species or phenomena. Although many uncertainties and speculations exist, the available evidence presented here suggests that a world without oncogenic processes would be quite different from the world we know (Aktipis, 2020), not only because of the suppression of the pathological effects of cancer but primarily because of the relaxation of anticancer selection, with effects extending from host individuals to populations, species, communities, and ecosystems. We can hope in the future that long-term experimental evolution experiments using animal populations and communities made of artificially cancer-free species (see above) will allow one to better assess how different could be a world without cancer constraints.
Figure 2.

Summary of the Different Levels at Which the Costs of Cancer Defenses Influence the Living World, from Cells to Individuals, Populations, Species, and Ecosystems
Acknowledgments
This work is supported by an ANR TRANSCAN (ANR-18-CE35-0009), the MAVA Foundation, the Rotary Club Les Sables d’Olonne, an ARC Linkage (LP170101105), Deakin SEBE_RGS_2019, and a CNRS International Associated Laboratory Grant. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. We also would like to thank two anonymous referees for their relevant and constructive suggestions on a previous version of this manuscript.
Author Contributions
J.B., A.D., B.U., and F.T. conceived the original idea. All authors participated in the discussions and in the writing and contributed to the final manuscript.
References
- Abegglen L.M., Caulin A.F., Chan A., Lee K., Robinson R., Campbell M.S., Kiso W.K., Schmitt D.L., Waddell P.J., Bhaskara S. Potential mechanisms for cancer resistance in elephants and comparative cellular response to DNA Damage in Humans. JAMA. 2015;314:1850–1860. doi: 10.1001/jama.2015.13134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aktipis A. The Cheating Cell. 2020. [DOI]
- Aktipis C.A., Nesse R.M. Evolutionary foundations for cancer biology. Evol. Appl. 2013;6:144–159. doi: 10.1111/eva.12034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aktipis C.A., Boddy A.M., Gatenby R.A., Brown J.S., Maley C.C. Life history trade-offs in cancer evolution. Nat. Rev. Cancer. 2013;13:883–892. doi: 10.1038/nrc3606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alfaro-Núñez A., Bertelsen M.F., Bojesen A.M., Rasmussen I., Zepeda-Mendoza L., Olsen M.T., Gilbert M.T.P. Global distribution of Chelonid fibropapilloma-associated herpesvirus among clinically healthy sea turtles. BMC Evol. Biol. 2014;14:206. doi: 10.1186/s12862-014-0206-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Armenta T.C., Cole S.W., Wayne R.K., Blumstein D.T. Adaptive gene regulation in wild mammals exposed to high predator abundance. Anim. Behav. 2019;152:53–61. [Google Scholar]
- Arnal A., Tissot T., Ujvari B., Nunney L., Solary E., Laplane L., Bonhomme F., Vittecoq M., Tasiemski A., Renaud F. The guardians of inherited oncogenic vulnerabilities. Evolution. 2016;70:1–6. doi: 10.1111/evo.12809. [DOI] [PubMed] [Google Scholar]
- Arnal A., Jacqueline C., Ujvari B., Leger L., Moreno C., Faugere D., Tasiemski A., Boidin-Wichlacz C., Misse D., Renaud F. Cancer brings forward oviposition in the fly Drosophila melanogaster. Ecol. Evol. 2017;7:272–276. doi: 10.1002/ece3.2571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Athena Aktipis C., Boddy A.M., Jansen G., Hibner U., Hochberg M.E., Maley C.C., Wilkinson G.S. Cancer across the tree of life: cooperation and cheating in multicellularity. Philos. Trans. R. Soc. B: Biol. Sci. 2015;370:20140219. doi: 10.1098/rstb.2014.0219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aubert A. Sickness and behaviour in animals: a motivational perspective. Neurosci. Biobehavioral Rev. 1999;23:1029–1036. doi: 10.1016/s0149-7634(99)00034-2. [DOI] [PubMed] [Google Scholar]
- Bargonetti J., Manfredi J.J. Multiple roles of the tumor suppressor p53. Curr. Opin. Oncol. 2002;14:86–91. doi: 10.1097/00001622-200201000-00015. [DOI] [PubMed] [Google Scholar]
- Barron A.B., Oldroyd B.P., Ratnieks F.L.W. Worker reproduction in honey-bees (Apis) and the anarchic syndrome: a review. Behav. Ecol. Sociobiol. 2001;50:199–208. [Google Scholar]
- Biernaskie J.M., West S.A. Cooperation, clumping and the evolution of multicellularity. Proc. R. Soc. B: Biol. Sci. 2015;282:20151075. doi: 10.1098/rspb.2015.1075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biro P.A., Thomas F., Ujvari B., Beckmann C. Can energetic capacity help explain why physical activity reduces cancer risk? Trends Cancer. 2020;6:829–837. doi: 10.1016/j.trecan.2020.06.001. [DOI] [PubMed] [Google Scholar]
- Boddy A.M., Abegglen L.M., Pessier A.P., Schiffman J.D., Maley C.C., Witte C. Lifetime cancer prevalence and life history traits in mammals. Evol. Med. Public Health. 2020 doi: 10.1093/emph/eoaa015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bond W.J., Woodward F.I., Midgley G.F. The global distribution of ecosystems in a world without fire. New Phytol. 2005;165:525–538. doi: 10.1111/j.1469-8137.2004.01252.x. [DOI] [PubMed] [Google Scholar]
- Bourke A.F.G. 2013. Principles of Social Evolution. [DOI] [Google Scholar]
- Bourneuf E. The MeLiM minipig: an original spontaneous model to explore cutaneous melanoma genetic basis. Front. Genet. 2017;13:146. doi: 10.3389/fgene.2017.00146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown J.S., Aktipis C.A. Inclusive fitness effects can select for cancer suppression into old age. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2015;370:20150160. doi: 10.1098/rstb.2015.0160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown M.J., Parker G.G., Posner N.E. A survey of ultraviolet-B radiation in forests. J. Ecol. 1994;82:843–854. [Google Scholar]
- Campisi J. Cellular senescence as a tumor-suppressor mechanism. Trends Cell Biol. 2001 doi: 10.1016/S0962-8924(01)02151-1. [DOI] [PubMed] [Google Scholar]
- Campisi J. Cancer and aging: yin, yang, and p53. Sci. Aging Knowledge Environ. 2002;2002:pe1. doi: 10.1126/sageke.2002.1.pe1. [DOI] [PubMed] [Google Scholar]
- Casás-Selves M., DeGregori J. How cancer shapes evolution and how evolution shapes cancer. Evol. Educ. Outreach. 2011 doi: 10.1007/s12052-011-0373-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caspari R., Lee S.H. Older age becomes common late in human evolution. Proc. Natl. Acad. Sci. U S A. 2004;101:10895–10900. doi: 10.1073/pnas.0402857101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caulin A.F., Maley C.C. Peto’s Paradox: evolution’s prescription for cancer prevention. Trends Ecol. Evol. 2011;26:175–182. doi: 10.1016/j.tree.2011.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chumakov P.M. Versatile functions of p53 protein in multicellular organisms. Biochemistry. 2007;72:1399–1421. doi: 10.1134/s0006297907130019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crespi B., Summers K. Evolutionary biology of cancer. Trends Ecol. Evol. 2005;20:545–552. doi: 10.1016/j.tree.2005.07.007. [DOI] [PubMed] [Google Scholar]
- Darvin P., Toor S.M., Sasidharan Nair V., Elkord E. Immune checkpoint inhibitors: recent progress and potential biomarkers. Exp. Mol. Med. 2018;50:1–11. doi: 10.1038/s12276-018-0191-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies J.A. Inverse correlation between an organ’s cancer rate and its evolutionary antiquity. Organogenesis. 2004;1:60–63. doi: 10.4161/org.1.2.1338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dawson E.H., Bailly T.P.M., Dos Santos J., Moreno C., Devilliers M., Maroni B., Sueur C., Casali A., Ujvari B., Thomas F. Social environment mediates cancer progression in Drosophila. Nat. Commun. 2018;9:3574. doi: 10.1038/s41467-018-05737-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeGregori J. Evolved tumor suppression: why are we so good at not getting cancer? Cancer Res. 2011;71:3739–3744. doi: 10.1158/0008-5472.CAN-11-0342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Der Most P.J., De Jong B., Parmentier H.K., Verhulst S. Trade-off between growth and immune function: a meta-analysis of selection experiments. Funct. Ecol. 2011;25:74–80. [Google Scholar]
- Diaz R.J. Overview of hypoxia around the world. J. Environ. Qual. 2001;30:275–281. doi: 10.2134/jeq2001.302275x. [DOI] [PubMed] [Google Scholar]
- Doherty A., Kernogitski Y., Kulminski A.M., de Magalhães J.P. Identification of polymorphisms in cancer patients that differentially affect survival with age. Aging. 2017 doi: 10.18632/aging.101305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Domazet-Lošo T., Tautz D. Phylostratigraphic tracking of cancer genes suggests a link to the emergence of multicellularity in metazoa. BMC Biol. 2010;8:66. doi: 10.1186/1741-7007-8-66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ducasse H., Arnal A., Vittecoq M., Daoust S.P., Ujvari B., Jacqueline C., Tissot T., Ewald P., Gatenby R.A., King K.C. Cancer: an emergent property of disturbed resource-rich environments? Ecology meets personalized medicine. Evol. Appl. 2015;8:527–540. doi: 10.1111/eva.12232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duggal N.A., Niemiro G., Harridge S.D.R., Simpson R.J., Lord J.M. Can physical activity ameliorate immunosenescence and thereby reduce age-related multi-morbidity? Nat. Rev. Immunol. 2019;19:563–572. doi: 10.1038/s41577-019-0177-9. [DOI] [PubMed] [Google Scholar]
- Dujon A.M., Ujvari B., Thomas F. Cancer risk landscapes: A framework to study cancer in ecosystems. Sci. Total Environ. 2020:142955. doi: 10.1016/j.scitotenv.2020.142955. Available online 14 October 2020. [DOI] [PubMed] [Google Scholar]
- Duke N.C., Meynecke J.-O., Dittmann S., Ellison A.M., Anger K., Berger U., Cannicci S., Diele K., Ewel K.C., Field C.D. A world without mangroves? Science. 2007;317:41–42. doi: 10.1126/science.317.5834.41b. [DOI] [PubMed] [Google Scholar]
- Van Dyken J.D., Linksvayer T.A., Wade M.J. Kin selection-mutation balance: a model for the origin, maintenance, and consequences of social cheating. Am. Naturalist. 2011;177:288–300. doi: 10.1086/658365. [DOI] [PubMed] [Google Scholar]
- Fang J. Ecology: a world without mosquitoes. Nature. 2010 doi: 10.1038/466432a. [DOI] [PubMed] [Google Scholar]
- Farhang Ghahremani M., Goossens S., Nittner D., Bisteau X., Bartunkova S., Zwolinska A., Hulpiau P., Haigh K., Haenebalcke L., Drogat B. P53 promotes VEGF expression and angiogenesis in the absence of an intact p21-Rb pathway. Cell Death Differ. 2013;20:888–897. doi: 10.1038/cdd.2013.12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandez A.a., Morris M.R. Mate choice for more melanin as a mechanism to maintain a functional oncogene. Proc. Natl. Acad. Sci. U S A. 2008;105:13503–13507. doi: 10.1073/pnas.0803851105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finn O.J. Immuno-oncology: understanding the function and dysfunction of the immune system in cancer. Ann. Oncol. 2012 doi: 10.1093/annonc/mds256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank S.A. Mutual policing and repression of competition in the evolution of cooperative groups. Nature. 1995;377:520–522. doi: 10.1038/377520a0. [DOI] [PubMed] [Google Scholar]
- Fridman J.S., Lowe S.W. Control of apoptosis by p53. Oncogene. 2003 doi: 10.1038/sj.onc.1207116. [DOI] [PubMed] [Google Scholar]
- Galis F. Why do almost all mammals have seven cervical vertebrae? Developmental constraints, Hox genes, and cancer. J. Exp. Zoolog. 1999;285:19–26. [PubMed] [Google Scholar]
- Galis F. On the homology of structures and Hox genes: the vertebral column. Novartis Found. Symp. 1999 doi: 10.1002/9780470515655.ch6. [DOI] [PubMed] [Google Scholar]
- Galis F., Metz J.A.J. Anti-cancer selection as a source of developmental and evolutionary constraints. BioEssays. 2003;25:1035–1039. doi: 10.1002/bies.10366. [DOI] [PubMed] [Google Scholar]
- Giaimo S., D’Adda di Fagagna F. Is cellular senescence an example of antagonistic pleiotropy? Aging Cell. 2012;11:378–383. doi: 10.1111/j.1474-9726.2012.00807.x. [DOI] [PubMed] [Google Scholar]
- Gilbert J.A., Neufeld J.D. Life in a world without microbes. PLoS Biol. 2014;12:e1002020. doi: 10.1371/journal.pbio.1002020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilbert O.M., Foster K.R., Mehdiabadi N.J., Strassmann J.E., Queller D.C. High relatedness maintains multicellular cooperation in a social amoeba by controlling cheater mutants. Proc. Natl. Acad. Sci. U S A. 2007;104:8913–8917. doi: 10.1073/pnas.0702723104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giraudeau M., Sepp T., Ujvari B., Ewald P.W., Thomas F. Human activities might influence oncogenic processes in wild animal populations. Nat. Ecol. Evol. 2018;2:1065–1070. doi: 10.1038/s41559-018-0558-7. [DOI] [PubMed] [Google Scholar]
- Grosberg R.K., Strathmann R.R. The evolution of multicellularity: a minor major transition? Annu. Rev. Ecol. Evol. Syst. 2007;38:621–654. [Google Scholar]
- Hamede R.K., Mccallum H., Jones M. Biting injuries and transmission of Tasmanian devil facial tumour disease. J. Anim. Ecol. 2013;82:182–190. doi: 10.1111/j.1365-2656.2012.02025.x. [DOI] [PubMed] [Google Scholar]
- Hamede R., Madsen T., McCallum H., Storfer A., Hohenlohe A.P., Siddle H., Kaufman J., Giraudeau, Jones M., Thomas F., Beata B. Darwin and the devil’s plight for managing transmissible cancers. Conservat. Biol. 2021 doi: 10.1111/cobi.13644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamilton W.D. The genetical evolution of social behaviour. I. J. Theor. Biol. 1964 doi: 10.1016/0022-5193(64)90038-4. [DOI] [PubMed] [Google Scholar]
- Hanahan D., Weinberg R.A. Review hallmarks of Cancer : the next generation. Cell. 2011;144:646–674. doi: 10.1016/j.cell.2011.02.013. [DOI] [PubMed] [Google Scholar]
- Harel I., Brunet A. The African turquoise killifish: a model for exploring vertebrate aging and diseases in the fast lane. Cold Spring Harbor Symposia Quantitative Biol. 2016 doi: 10.1101/sqb.2015.80.027524. [DOI] [PubMed] [Google Scholar]
- Hawks J., Wang E.T., Cochran G.M., Harpending H.C., Moyzis R.K. Recent acceleration of human adaptive evolution. Proc. Natl. Acad. Sci. U S A. 2007;104:20753–20758. doi: 10.1073/pnas.0707650104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heithaus M.R., Frid A., Wirsing A.J., Dill L.M., Fourqurean J.W., Burkholder D., Thomson J., Bejder L. State-dependent risk-taking by green sea turtles mediates top-down effects of tiger shark intimidation in a marine ecosystem. J. Anim. Ecol. 2007;76:837–844. doi: 10.1111/j.1365-2656.2007.01260.x. [DOI] [PubMed] [Google Scholar]
- Hermes-Lima M., Zenteno-Savín T. Animal response to drastic changes in oxygen availability and physiological oxidative stress. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2002;133:537–556. doi: 10.1016/s1532-0456(02)00080-7. [DOI] [PubMed] [Google Scholar]
- Hochberg M.E., Noble R.J. A framework for how environment contributes to cancer risk. Ecol. Lett. 2017;20:117–134. doi: 10.1111/ele.12726. [DOI] [PubMed] [Google Scholar]
- Hochberg M.E., Thomas F., Assenat E., Hibner U. Preventive evolutionary medicine of cancers. Evol. Appl. 2013 doi: 10.1111/eva.12033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacqueline C., Biro P.A., Beckmann C., Moller A.P., Renaud F., Sorci G., Tasiemski A., Ujvari B., Thomas F. Cancer: a disease at the crossroads of trade-offs. Evol. Appl. 2017;10:215–225. doi: 10.1111/eva.12444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janssens L., Stoks R. Predation risk causes oxidative damage in prey. Biol. Lett. 2013;9 doi: 10.1098/rsbl.2013.0350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones M.E., Cockburn A., Hamede R., Hawkins C., Hesterman H., Lachish S., Mann D., McCallum H., Pemberton D. Life-history change in disease-ravaged Tasmanian devil populations. Proc. Natl. Acad. Sci. U S A. 2008;105:10023–10027. doi: 10.1073/pnas.0711236105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones K., Ariel E., Burgess G., Read M. A review of fibropapillomatosis in Green turtles (Chelonia mydas) Vet. J. 2016;212:48–57. doi: 10.1016/j.tvjl.2015.10.041. [DOI] [PubMed] [Google Scholar]
- Keane M., Semeiks J., Webb A.E., Li Y.I., Quesada V., Craig T., Madsen L.B., van Dam S., Brawand D., Marques P.I. Insights into the evolution of longevity from the bowhead whale genome. Cell Rep. 2015;10:112–122. doi: 10.1016/j.celrep.2014.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klerks P.L., Levinton J.S. Rapid evolution of metal resistance in a benthic oligochaete inhabiting a metal-polluted site. Biol. Bull. 1989;176:135–141. [Google Scholar]
- Krtolica A., Parrinello S., Lockett S., Desprez P.Y., Campisi J. Senescent fibroblasts promote epithelial cell growth and tumorigenesis: a link between cancer and aging. Proc. Natl. Acad. Sci. U S A. 2001;98:12072–12077. doi: 10.1073/pnas.211053698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lambe M., Hsieh C., Trichopoulos D., Ekbom A., Pavia M., Adami H.-O. Transient increase in the risk of breast cancer after giving birth. N. Engl. J. Med. 2002;13:299–305. doi: 10.1056/NEJM199407073310102. [DOI] [PubMed] [Google Scholar]
- Leroi A.M., Koufopanou V., Burt A. Cancer selection. Nat. Rev. Cancer. 2003;3:226–231. doi: 10.1038/nrc1016. [DOI] [PubMed] [Google Scholar]
- Levine A.J. p53, the cellular gatekeeper for growth and division. Cell. 1997 doi: 10.1016/S0092-8674(00)81871-1. [DOI] [PubMed] [Google Scholar]
- Lochmiller R.L., Deerenberg C. Trade-offs in evolutionary immunology: just what is the cost of immunity? Oikos. 2000;88:87–98. [Google Scholar]
- Lowe S.W., Cepero E., Evan G. Intrinsic tumour suppression. Nature. 2004 doi: 10.1038/nature03098. [DOI] [PubMed] [Google Scholar]
- Madsen T., Arnal A., Vittecoq M., Bernex F., Abadie J., Labrut S., Garcia D., Faugère D., Lemberger K., Beckmann C. Cancer prevalence and etiology in wild and captive animals. Ecol. Evol. Cancer. 2017:11–46. doi: 10.1016/B978-0-12-804310-3.00002-8. [DOI] [Google Scholar]
- Martel L., Gagnon M.J., Massé R., Leclerc A., Tremblay L. Polycyclic aromatic hydrocarbons in sediments from the Saguenay Fjord, Canada. Bull. Environ. Contam. Toxicol. 1986;37:133–140. doi: 10.1007/BF01607740. [DOI] [PubMed] [Google Scholar]
- Martineau D., De Guise S., Fournier M., Shugart L., Girard C., Lagacé A., Béland P. Pathology and toxicology of beluga whales from the St. Lawrence Estuary, Quebec, Canada. Past, present and future. Sci. Total Environ. 1994;154:201–215. doi: 10.1016/0048-9697(94)90088-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McAloose D., Newton A.L. Wildlife cancer: a conservation perspective. Nat. Rev. Cancer. 2009 doi: 10.1038/nrc2665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mcmahon B.R. Physiological responses to oxygen depletion in intertidal animals. Integr. Comp. Biol. 1988 doi: 10.1093/icb/28.1.39. [DOI] [Google Scholar]
- Michod R.E. Cooperation and conflict in the evolution of individuality. II. Conflict mediation. Proc. R. Soc. B: Biol. Sci. 1996 doi: 10.1098/rspb.1996.0121. [DOI] [PubMed] [Google Scholar]
- Møller A.P., Mousseau T.A. Birds prefer to breed in sites with low radioactivity in Chernobyl. Proc. R. Soc. B: Biol. Sci. 2007;274:1443–1448. doi: 10.1098/rspb.2007.0005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Møller A.P., Bonisoli-Alquati A., Mousseau T.A. High frequency of albinism and tumours in free-living birds around Chernobyl. Mutat. Res. 2013;757:52–59. doi: 10.1016/j.mrgentox.2013.04.019. [DOI] [PubMed] [Google Scholar]
- Muller A.W.J. Cancer is an adaptation that selects in animals against energy dissipation. Med. Hypotheses. 2017;104:104–115. doi: 10.1016/j.mehy.2017.05.030. [DOI] [PubMed] [Google Scholar]
- Nechifor M., Neagu T.-M., Manda G. Reactive oxygen species, cancer and anti-cancer therapies. Curr. Chem. Biol. 2012;3:22–46. [Google Scholar]
- Nesse R.M. Evolution: medicine’s most basic science. Lancet. 2008 doi: 10.1016/s0140-6736(08)61877-2. [DOI] [Google Scholar]
- Nguyen T.Y.V., Batterham M.J., Edwards C. Comparison of resting energy expenditure between cancer subjects and healthy controls: a meta-analysis. Nutr. Cancer. 2016;68:374–387. doi: 10.1080/01635581.2016.1153667. [DOI] [PubMed] [Google Scholar]
- Norris K., Evans M.R. Ecological immunology: life history trade-offs and immune defense in birds. Behav. Ecol. 2000;11:19–26. [Google Scholar]
- Nunney L. The real war on cancer: the evolutionary dynamics of cancer suppression. Evol. Appl. 2013;6:11–19. doi: 10.1111/eva.12018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nunney L. The evolutionary origins of cancer and of its control by immune policing and genetic suppression. Ecol. Evol. Cancer. 2017 doi: 10.1016/B978-0-12-804310-3.00001-6. [DOI] [Google Scholar]
- Nunney L., Maley C.C., Breen M., Hochberg M.E., Schiffman J.D. Peto’s paradox and the promise of comparative oncology. Philos. Trans. R. Soc. B. 2015;370:20140177. doi: 10.1098/rstb.2014.0177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parfrey L.W., Lahr D.J.G. Multicellularity arose several times in the evolution of eukaryotes. BioEssays. 2013;35:339–347. doi: 10.1002/bies.201200143. [DOI] [PubMed] [Google Scholar]
- Paul N.D., Gwynn-Jones D. Ecological roles of solar UV radiation: towards an integrated approach. Trends Ecol. Evol. 2003;18:48–55. [Google Scholar]
- Pedersen L., Idorn M., Olofsson G.H., Lauenborg B., Nookaew I., Hansen R.H., Johannesen H.H., Becker J.C., Pedersen K.S., Dethlefsen C. Voluntary running suppresses tumor growth through epinephrine- and IL-6-dependent NK cell mobilization and redistribution. Cell Metab. 2016;23:554–562. doi: 10.1016/j.cmet.2016.01.011. [DOI] [PubMed] [Google Scholar]
- Perret C., Gidoin C., Ujvari B., Thomas F., Roche B. Predation shapes the impact of cancer on population dynamics and the evolution of cancer resistance. Evol. Appl. 2020;13:1733–1744. doi: 10.1111/eva.12951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peto H., Roe F.J.C., Lee P.N., Levy L., Clack J. Cancer and ageing in mice and men. Br. J. Cancer. 1975;32:411–426. doi: 10.1038/bjc.1975.242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Praetorius S.K., Mix A.C., Walczak M.H., Wolhowe M.D., Addison J.A., Prahl F.G. North Pacific deglacial hypoxic events linked to abrupt ocean warming. Nature. 2015;527:362–366. doi: 10.1038/nature15753. [DOI] [PubMed] [Google Scholar]
- Purcell S.A., Elliott S.A., Baracos V.E., Chu Q.S.C., Prado C.M. Key determinants of energy expenditure in cancer and implications for clinical practice. Eur. J. Clin. Nutr. 2016;70:1230–1238. doi: 10.1038/ejcn.2016.96. [DOI] [PubMed] [Google Scholar]
- Pye R., Patchett A., McLennan E., Thomson R., Carver S., Fox S., Pemberton D., Kreiss A., Morelli A.B., Silva A. Immunization strategies producing a humoral IgG immune response against devil facial tumor disease in the majority of Tasmanian devils destined for wild release. Front. Immunol. 2018;9:259. doi: 10.3389/fimmu.2018.00259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raihani N.J., Thornton A., Bshary R. Punishment and cooperation in nature. Trends Ecol. Evol. 2012;27:288–295. doi: 10.1016/j.tree.2011.12.004. [DOI] [PubMed] [Google Scholar]
- Randhawa N., Gulland F., Ylitalo G.M., DeLong R., Mazet J.A.K. Sentinel California sea lions provide insight into legacy organochlorine exposure trends and their association with cancer and infectious disease. One Health. 2015;1:37–43. doi: 10.1016/j.onehlt.2015.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reid N.M., Proestou D.A., Clark B.W., Warren W.C., Colbourne J.K., Shaw J.R., Karchner S.I., Hahn M.E., Nacci D., Oleksiak M.F. The genomic landscape of rapid repeated evolutionary adaptation to toxic pollution in wild fish. Science. 2016;354:1305–1308. doi: 10.1126/science.aah4993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Roode J.C., Lefevre T., Hunter M.D. Self-medication in animals. Science. 2013 doi: 10.1126/science.1235824. [DOI] [PubMed] [Google Scholar]
- Rubin J.B. Sexual selection and cancer biology. Oncotarget. 2015;6:15714–15715. doi: 10.18632/oncotarget.4592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sayol F., Downing P.A., Iwaniuk A.N., Maspons J., Sol D. Predictable evolution towards larger brains in birds colonizing oceanic islands. Nat. Commun. 2018;9:2820. doi: 10.1038/s41467-018-05280-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schiller J.T., Lowy D.R. Vaccines to prevent infections by oncoviruses. Annu. Rev. Microbiol. 2010 doi: 10.1146/annurev.micro.112408.134019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schumacher R., Mai A., Gutjahr P. Association of rib anomalies and malignancy in childhood. Eur. J. Pediatr. 1992;151:432–434. doi: 10.1007/BF01959357. [DOI] [PubMed] [Google Scholar]
- Sepp T., Ujvari B., Ewald P.W., Thomas F., Giraudeau M. Urban environment and cancer in wildlife: available evidence and future research avenues. Proc. Biol. Sci. 2019;286:20182434. doi: 10.1098/rspb.2018.2434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaw P.H. The role of p53 in cell cycle regulation. Pathol. Res. Pract. 1996 doi: 10.1016/S0344-0338(96)80088-4. [DOI] [PubMed] [Google Scholar]
- Sheriff M.J., Thaler J.S. Ecophysiological effects of predation risk; an integration across disciplines. Oecologia. 2014;176:607–611. doi: 10.1007/s00442-014-3105-5. [DOI] [PubMed] [Google Scholar]
- Stearns S.C. Trade-Offs in life-history evolution. Funct. Ecol. 2006 doi: 10.2307/2389364. [DOI] [Google Scholar]
- Stensrud M.J., Valberg M. Inequality in genetic cancer risk suggests bad genes rather than bad luck. Nat. Commun. 2017 doi: 10.1038/s41467-017-01284-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sulak M., Fong L., Mika K., Chigurupati S., Yon L., Mongan N.P., Emes R.D., Lynch V.J. TP53 copy number expansion is associated with the evolution of increased body size and an enhanced DNA damage response in elephants. eLife. 2016 doi: 10.7554/eLife.11994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Summers K., Crespi B.J. Xmrks the spot: life history tradeoffs, sexual selection and the evolutionary ecology of oncogenesis. Mol. Ecol. 2010;19:3022–3024. doi: 10.1111/j.1365-294x.2010.04739.x. [DOI] [PubMed] [Google Scholar]
- Sweet M., Kirkham N., Bendall M., Currey L., Bythell J., Heupel M. Evidence of melanoma in wild marine fish populations. PLoS One. 2012;7:e41989. doi: 10.1371/journal.pone.0041989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szathmáry E., Smith J.M. The major evolutionary transitions. Nature. 1995 doi: 10.1038/374227a0. [DOI] [PubMed] [Google Scholar]
- Thomas F., Donnadieu E., Charriere G.M., Jacqueline C., Tasiemski A., Pujol P., Renaud F., Roche B., Hamede R., Brown J. Is adaptive therapy natural? PLoS Biol. 2018;16:e2007066. doi: 10.1371/journal.pbio.2007066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas F., Kareva I., Raven N., Hamede R., Pujol P., Roche B., Ujvari B. Evolved dependence in response to cancer. Trends Ecol. Evol. 2018;33:269–276. doi: 10.1016/j.tree.2018.01.012. [DOI] [PubMed] [Google Scholar]
- Thomas F., Giraudeau M., Renaud F., Ujvari B., Roche B., Pujol P., Raymond M., Lemaitre J.F., Alvergne A. Can postfertile life stages evolve as an anticancer mechanism? PLoS Biol. 2019;17:e3000565. doi: 10.1371/journal.pbio.3000565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas F., Giraudeau M., Dheilly N.M., Gouzerh F., Boutry J., Beckmann C., Biro P.A., Hamede R., Abadie J., Labrut S. Rare and unique adaptations to cancer in domesticated species: an untapped resource? Evol. Appl. 2020 doi: 10.1111/eva.12920. [DOI] [Google Scholar]
- Thompson H.J., Jones L.W., Koch L.G., Britton S.L., Neil E.S., McGinley J.N. Inherent aerobic capacity-dependent differences in breast carcinogenesis. Carcinogenesis. 2017;38:920–928. doi: 10.1093/carcin/bgx066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tollis M., Boddy A.M., Maley C.C. Peto’s Paradox: how has evolution solved the problem of cancer prevention? BMC Biol. 2017;15:60. doi: 10.1186/s12915-017-0401-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ujvari B., Beckmann C., Biro P.A., Arnal A., Tasiemski A., Massol F., Salzet M., Mery F., Boidin-Wichlacz C., Misse D. Cancer and life-history traits: lessons from host-parasite interactions. Parasitology. 2016;143:533–541. doi: 10.1017/S0031182016000147. [DOI] [PubMed] [Google Scholar]
- Ujvari B., Roche B., Thomas F. 2017. Ecology and Evolution of Cancer. [Google Scholar]
- Ungewitter E., Scrable H. Antagonistic pleiotropy and p53. Mech. Ageing Dev. 2009;130:10–17. doi: 10.1016/j.mad.2008.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valko M., Rhodes C.J., Moncol J., Izakovic M., Mazur M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chemico-Biological Interactions. 2006;160:1–40. doi: 10.1016/j.cbi.2005.12.009. [DOI] [PubMed] [Google Scholar]
- Vazquez J.M., Sulak M., Chigurupati S., Lynch V. A zombie LIF gene in elephants is up-regulated by TP53 to induce apoptosis in response to DNA damage. Cell Rep. 2017;24:1765–1776. doi: 10.1016/j.celrep.2018.07.042. [DOI] [PubMed] [Google Scholar]
- Vincent-Naulleau S., Le Chalony C., Leplat J.J., Bouet S., Bailly C., Spatz A., Vielh P., Avril M.F., Tricaud Y., Gruand J. Clinical and histopathological characterization of cutaneous melanomas in the melanoblastoma-bearing libechov minipig model. Pigment Cell Res. 2004;17:24–35. doi: 10.1046/j.1600-0749.2003.00101.x. [DOI] [PubMed] [Google Scholar]
- Vittecoq M., Roche B., Daoust S.P., Ducasse H., Missé D., Abadie J., Labrut S., Renaud F., Gauthier-Clerc M., Thomas F. Cancer: a missing link in ecosystem functioning? Trends Ecol. Evol. 2013;28:628–635. doi: 10.1016/j.tree.2013.07.005. [DOI] [PubMed] [Google Scholar]
- Vittecoq M., Ducasse H., Arnal A., Møller A.P., Ujvari B., Jacqueline C.B., Tissot T., Missé D., Bernex F., Pirot N. Animal behaviour and cancer. Anim. Behav. 2015;101:19–26. [Google Scholar]
- Vittecoq M., Giraudeau M., Sepp T., Marcogliese D.J., Klaassen M., Renaud F., Ujvari B., Thomas F. Turning natural adaptations to oncogenic factors into an ally in the war against cancer. Evol. Appl. 2018;11:836–844. doi: 10.1111/eva.12608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vousden K.H., Lu X. Live or let die: the cell’s response to p53. Nat. Rev. Cancer. 2002;2:594–604. doi: 10.1038/nrc864. [DOI] [PubMed] [Google Scholar]
- Walker J.C.G., Margulis L., Rambler M. Reassessment of roles of oxygen and ultraviolet light in Precambrian evolution. Nature. 1976;264:620–624. [Google Scholar]
- Wang Y., Zhang C., Wang N., Li Z., Heller R., Liu R., Zhao Y., Han J., Pan X., Zheng Z. Genetic basis of ruminant headgear and rapid antler regeneration. Science. 2019;364:eaav6335. doi: 10.1126/science.aav6335. [DOI] [PubMed] [Google Scholar]
- Ward T.J., Robinson W.E. Evolution of cadmium resistance in Daphnia magna. Environ. Toxicol. Chem. 2005;24:2341. doi: 10.1897/04-429r.1. [DOI] [PubMed] [Google Scholar]
- West S.A., Griffin A.S., Gardner A. Evolutionary explanations for cooperation. Curr. Biol. 2007;17:R661–R672. doi: 10.1016/j.cub.2007.06.004. [DOI] [PubMed] [Google Scholar]
- Whitehead A., Pilcher W., Champlin D., Nacci D. Common mechanism underlies repeated evolution of extreme pollution tolerance. Proc. Biol. Sci. 2012;279:427–433. doi: 10.1098/rspb.2011.0847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitehead A., Clark B.W., Reid N.M., Hahn M.E., Nacci D. When evolution is the solution to pollution: key principles, and lessons from rapid repeated adaptation of killifish (Fundulus heteroclitus) populations. Evol. Appl. 2017;10:762–783. doi: 10.1111/eva.12470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams A.B., Schumacher B. p53 in the DNA-damage-repair process. Cold Spring Harbor Perspect. Med. 2016;6:a026070. doi: 10.1101/cshperspect.a026070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wood C.L., Johnson P.T.J. A world without parasites: exploring the hidden ecology of infection. Front. Ecol. Environ. 2015;13:403–456. doi: 10.1890/140368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Work T.M., Rameyer R.A., Balazs G.H., Cray C., Chang S.P. Immune status of free-ranging green turtles with fibropapillomatosis from Hawaii. J. Wildl. Dis. 2001;37:574–581. doi: 10.7589/0090-3558-37.3.574. [DOI] [PubMed] [Google Scholar]
- Wright W.E., Shay J.W. Cellular senescence as a tumor-protection mechanism: the essential role of counting. Curr. Opin. Genet. Dev. 2001 doi: 10.1016/S0959-437X(00)00163-5. [DOI] [PubMed] [Google Scholar]
- Xie L., Klerks P.L. Responses to selection for cadmium resistance in the least killifish, Heterandria formosa. Environ. Toxicol. Chem. 2003;22:313–320. [PubMed] [Google Scholar]
- Yaddanapudi K., Mitchell R.A., Putty K., Willer S., Sharma R.K., Yan J., Bodduluri H., Eaton J.W. Vaccination with embryonic stem cells protects against lung cancer: is a broad-spectrum prophylactic vaccine against cancer possible? PLoS One. 2012;7:e42289. doi: 10.1371/journal.pone.0042289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ye Z.L., Qian Q., Jin H.J., Qian Q.J. Cancer vaccine: learning lessons from immune checkpoint inhibitors. J. Cancer. 2018;9:263–268. doi: 10.7150/jca.20059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ylitalo G.M., Stein J.E., Hom T., Johnson L.L., Tilbury K.L., Hall A.J., Rowles T., Greig D., Lowenstine L.J., Gulland F.M.D. The role of organochlorines in cancer-associated mortality in California sea lions (Zalophus californianus) Mar. Pollut. Bull. 2005;50:30–39. doi: 10.1016/j.marpolbul.2004.08.005. [DOI] [PubMed] [Google Scholar]
- Zillén L., Conley D.J., Andrén T., Andrén E., Björck S. Past occurrences of hypoxia in the Baltic Sea and the role of climate variability, environmental change and human impact. Earth-Science Rev. 2008;91:77–92. [Google Scholar]