

ABSTRACT
No proven remedy is identified for COVID-19 yet. SARS-CoV-2, the viral agent, is recognized by some endosomal and cytosolic receptors following cell entry, entailing innate and adaptive immunity stimulation, notably through interferon induction. Impairment in immunity activation in some patients, mostly elderlies, leads to high mortalities; thus, promoting immune responses may help. BCG vaccine is under investigation to prevent COVID-19 due to its non-specific effects on the immune system. However, other complementary immune-induction methods at early stages of the disease may be needed. Here, the potentially preventive immunologic effects of BCG and influenza vaccination are compared with the immune response defects caused by aging and COVID-19. BCG co-administration with interferon-α/-β, or influenza vaccine is suggested to overcome its shortcomings in interferon signaling against COVID-19. However, further studies are highly recommended to assess the outcomes of such interventions considering their probable adverse effects especially augmented innate immune responses and overproduction of proinflammatory mediators.
KEYWORDS: COVID-19, BCG vaccine, influenza vaccine, immunity, pandemic, trained immunity, severe acute respiratory syndrome coronavirus 2
1. Introduction
The novel coronavirus disease 2019 (COVID-19) was announced as a pandemic by the World Health Organization (WHO) shortly after its first report in Dec. 2019 in Wuhan city, China.1 While worldwide enormous efforts are ongoing to find preventive and/or therapeutic approaches for COVID-19, its huge burden on countries and societies2 has prompted politicians to look for solutions as well. Donald Trump asked a while ago whether the flu vaccine combats the novel coronavirus, which was answered with a direct NO! Although this naïve question was raised because of a lack of knowledge about vaccination basics, it seems that the answer to his question could probably be yes!
Recently, it was suggested that the booster BCG (Bacillus Calmette–Guérin) vaccine, could have beneficial effects on preventing COVID-19 infection3 and reducing the incidence and severity of COVID-19 in previously BCG-vaccinated groups through its non-specific effects (NSEs). It was also proposed that the differences in COVID-19 severity amongst countries could be, to some extent, explained by various national policies on BCG children vaccination.4,5 However, this claim was questioned by some other studies.6,7
In this context, an open-label two-group phase III randomized controlled trial was first begun in up to 4170 healthcare workers in Australia. It is currently ongoing and aims to reveal the possible preventive effects of BCG vaccination against COVID-19.8 Presently, one observational and seven interventional clinical trials are also being conducted on this subject, which are briefed in Table 1.9
Table 1.
Clinical trials on BCG vaccine usage for prevention of COVID-19, Available from: https://clinicaltrials.gov/at 2020 September 1
| NCT Number | Title | Recruiting | Interventions | Outcome Measures | Age | Phase | Enrollment | Study Type | Study Designs | Start Date | Completion Date | Locations |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NCT04328441 | Reducing Health Care Workers Absenteeism in Covid-19 Pandemic Through BCG Vaccine | Yes | BCG Vaccine | Placebo | Health Care Workers absenteeism/the cumulative incidence of documented COVID-19/Hospital Admission due to documented COVID-19 and any reason, self-reported acute respiratory symptoms or fever/death due to documented COVID-19/ICU Admission due to documented COVID-19 and any reason/the cumulative incidence of self-reported fever/self-reported acute respiratory symptoms/death for any reason/the number of days of unplanned absenteeism, because of documented COVID-19, because of imposed quarantine as a result of exposure to COVID-19, because of imposed quarantine as a result of having acute respiratory symptoms/fever or documented COVID-19, because of self-reported acute respiratory symptoms/the number of days of self-reported fever [>38 deg C]/the number of days of self-reported acute respiratory symptoms/the cumulative incidence and magnitude of plasma-serum antibodies [IgA,M,G] and SARS-CoV-2-specific antibodies at 12 weeks after vaccination and at the end of the study period | 18< | 3 | 1500 | Interventional | Allocation: Randomized | Intervention Model: Parallel Assignment | Masking: Quadruple (Participant, Care Provider, Investigator, Outcomes Assessor) | Primary Purpose: Prevention | Mar-20 | Dec-20 | Netherlands |
| NCT04362124 | Performance Evaluation of BCG Vaccination in Healthcare Personnel to Reduce the Severity of SARS-COV-2 Infection | Not yet | BCG Vaccine | Placebo | Incidence of COVID-19 cases confirmed or probable in the study population/Incidence of severe or critical infection in COVID-19 cases, Lethality of the infection in both groups, Assess the safety (frequency, seriousness, and severity of adverse events) of BCG vaccination, Prevalence of SARS-Cov-2 infection | 18–65 | 3 | 1000 | Interventional | Allocation: Randomized | Intervention Model: Parallel Assignment | Masking: Quadruple (Participant, Care Provider, Investigator, Outcomes Assessor) | Primary Purpose: Supportive Care | Apr-20 | Nov-21 | Colombia |
| NCT04379336 | BCG Vaccination for Healthcare Workers in COVID-19 Pandemic | Yes | BCG Vaccine | Placebo | Incidence of HCWs hospitalized due to COVID-19 per arm | Incidence of SARS-CoV-2 infection per arm | Incidence of upper respiratory tract infections per arm | Days of unplanned absenteeism due to COVID-19 or any reason per arm | Incidence of hospitalization for any reason per arm | Incidence of ICU admission per arm | Incidence of death per arm | Prevalence of latent TB infection | Incidence of active TB per arm | Compare the effect of latent TB on morbidity and mortality due to COVID-19 per arm | Incidence of treatment related adverse events | 18< | 3 | 500 | Interventional | Allocation: Randomized | Intervention Model: Parallel Assignment | Masking: Quadruple (Participant, Care Provider, Investigator, Outcomes Assessor) | Primary Purpose: Prevention | May-20 | Apr-21 | South Africa |
| NCT04350931 | Application of BCG Vaccine for Immune-prophylaxis Among Egyptian Healthcare Workers During the Pandemic of COVID-19 | Not yet | BCG Vaccine | Placebo | incidence of confirmed COVID-19 | Effectiveness of BCG vaccine | 18< | 3 | 900 | Interventional | Allocation: Randomized | Intervention Model: Parallel Assignment | Masking: Single (Participant) | Primary Purpose: Prevention | Apr-20 | Dec-20 | Egypt |
| NCT04417335 | Reducing COVID-19 Related Hospital Admission in Elderly by BCG Vaccination | Active | BCG Vaccine | Placebo | SARS-CoV-2 related hospital admission | the duration of hospital admission due to documented COVID-19 | the cumulative incidence of documented SARS-CoV-2 infection, self-reported acute respiratory symptoms or fever, death due to documented SARS-CoV-2 infection, hospital admission for any reason, ICU Admission due to documented SARS-CoV-2 infection | 60< | 4 | 2014 | Interventional | Allocation: Randomized | Intervention Model: Parallel Assignment | Masking: Single (Participant) | Primary Purpose: Prevention | Apr-20 | May-21 | Netherlands |
| NCT04347876 | Outcome of COVID-19 Cases Based on Tuberculin Test: Can Previous BCG Alter the Prognosis? | Yes | Diagnostic Test: Tuberculin test | Pneumonia severity index | Need for ICU admission | COVID −19 test conversion | Mortality | 12–80 | 100 | Observational | Observational Model: Case-Control | Time Perspective: Prospective | Apr-20 | Jun-20 | Egypt | |
| NCT04475302 | BCG Vaccine in Reducing Morbidity and Mortality in Elderly Individuals in COVID-19 Hotspots | Yes | BCG Vaccine | Placebo | Mortality due to COVID-19 disease | Hospital admission and ICU admission due to Covid-19, Severity of Covid-19 (mild, moderate or severe), Hospital admission due to other respiratory febrile illness, immunological outcomes in the sub study | 60–80 | 3 | 2175 | Interventional | Allocation: Non-Randomized | Intervention Model: Single Group Assignment | Masking: None (Open Label) | Primary Purpose: Prevention | Jul-20 | May-21 | India |
| NCT04461379 | Prevention, Efficacy and Safety of BCG Vaccine in COVID-19 Among Healthcare Workers | Not yet | BCG Vaccine | Placebo | Demonstrate COVID- 19 disease incidence, cumulative hospitalization, specific Antibodies against SARS-CoV-2 at 3 and 6 months among Health care workers | Hospitalization, Oxygen supplementation in severe disease COVID-19 | Need for intubation or noninvasive ventilation for the patient. | Critical care admission with SARS-CoV2 | Mortality associated to progressive pulmonary disease | safety of the vaccine by measuring the incidence rates of local and systemic adverse effects that occur after one month its application. | Incidence of COVID-19 complications | Mean days of hospitalization and days in ICU by COIVD-19 | Cost associated with in-hospital medical care | Scores of the Clinical Prediction Rules associated with mortality using Sequential Organ Failure Assessment (SOFA score) | Scores of the Clinical Prediction Rules associated with mortality using Acute Physiology and Chronic Health disease Classification System (APACHE) | Alteration profile in laboratory studies | Registration of chronic medications | Need for vasopressors | 18< | 3 | 908 | Interventional | Allocation: Randomized | Intervention Model: Parallel Assignment | Masking: Triple (Participant, Care Provider, Outcomes Assessor) | Primary Purpose: Prevention | Jul-20 | Jan-21 | Mexico |
Quadruple masking: P = Participant, C = Care Provider, I = Investigator, O = Outcomes Assessor
The observed more efficient and improved immune responses against reinfections in the plants and invertebrate that lack adaptive immunity, and surprisingly in some mammals, encouraged researchers to investigate whether this process could also occur in humans.10 Thus, several epidemiological studies and clinical trials were performed to explore the post-effects of vaccination with live-attenuated vaccines (LAVs), which mimic a natural infection. Though, the capability of LAVs in generating a sort of cross-protection against some other non-related pathogens was established in several studies.11 In this regard, the NSEs of several vaccines (BCG, DTP, and measles vaccines) have been investigated.12 One primary mechanism behind this phenomenon was found to be the induction of long-lasting epigenetic changes in the innate immune cells, a process called “trained immunity”.11 Besides, heterologous lymphocyte responses may partly account for some NSEs of vaccines.13 On this point, antigen cross-reactivity and bystander activation of unrelated T and B cells are considered as possible mechanisms. The bystander lymphocytes could exert protective roles against unrelated heterologous pathogens through antigen-specific bystander responses or antigen-non-specific innate mechanisms, such as interferon (IFN)-γ production, which activates macrophages.11,13
The higher mortality rates in elderlies is a main concern in the COVID-19 pandemic, which is probably related to immunosenescence, or age-associated deterioration of immune functionality,14 leading to suboptimal immune responses. In view of this, boosting the immune responses against SARS-CoV-2 could potentially reduce the disease incidence and severity in this population as well.
Herein, a model on the immune responses against COVID-19 is proposed first. Then, the Yin and Yang of the NSEs of LAVs are described with a focus on BCG and influenza vaccines. Finally, the probable supportive roles of these vaccines on the protection against COVID-19 would be discussed.
2. Interaction of SARS-CoV-2 with the immune system
A brief overview of immune responses to SARS-CoV-2 is represented in Figure 1. SARS-CoV-2, similar to SARS-CoV, enters the cells through angiotensin-converting enzyme II (ACE2) receptor. It also uses transmembrane protease serine 2 (TMPRSS2) receptor to prime its spike (S) protein in target cells.15,16 TMPRSS2 plays a role in influenza virus pathogenicity as well by cleaving hemagglutinin.17 Other yet-unknown cellular entry modes might also be involved in SARS-CoV-2 infection, especially considering that only minimal percentages of leukocytes express ACE2.18–20
Figure 1.
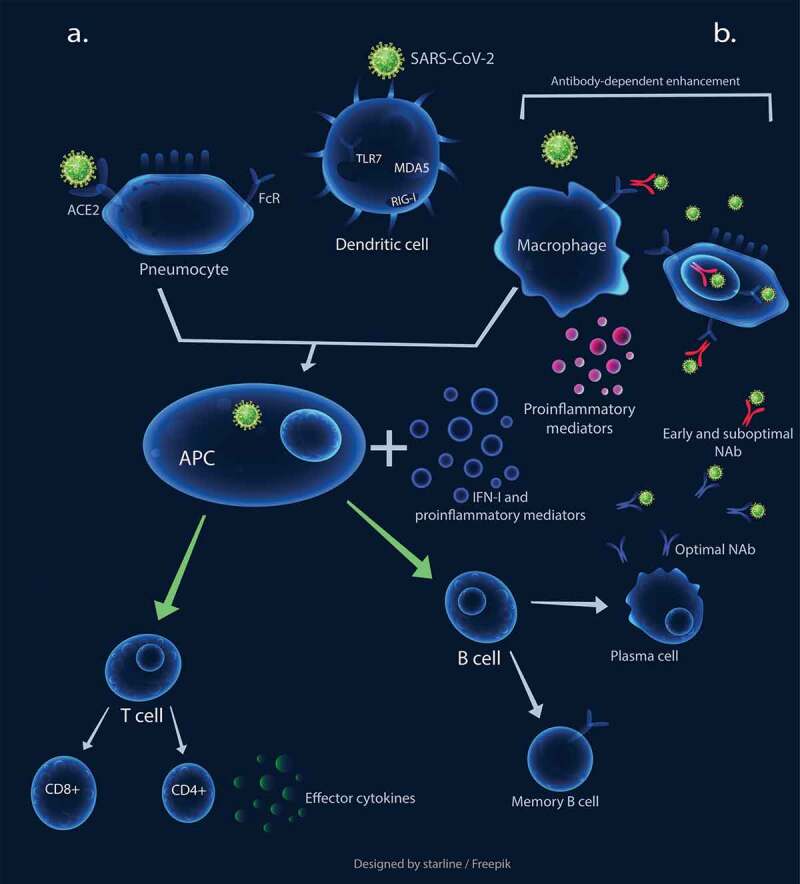
The schematic representation of immune responses against SARS-CoV-2. a. Optimal innate and adaptive immunity responses: The virus binds to ACE2 to infect cells. The engagement of pattern recognition receptors (PRRs) results in the production of interferons I and other proinflammatory mediators, which induces dendritic cells (DCs) to process antigens and present them to naïve circulating T cells, leading to T cell activation. The activated T cells migrate to the site of infection and secrete effector cytokines, such as IFN-γ. The APCs also induce B cells leading to optimal NAb production. b. Early and sub-optimal NAb activity results in antibody-dependent enhancement, leading macrophages to be exploited for virus replication. This process also causes elevated cytokine production and subsequent immunopathological overreactions
APC: Antigen Presenting Cell, NAb: Neutralizing Antibody
Host cells possibly recognize SARS-CoV-2, same as other RNA viruses, first by endosomal pattern recognition receptors, such as Toll-like receptor (TLR)3 and TLR7, as well as cytosolic RNA receptors, such as retinoic acid-inducible gene I (RIG-I) and melanoma differentiation-associated protein (MDA) 5. IFN-α or IFN-β (hereafter referred as IFN-I) and other proinflammatory mediators are induced following recognition of the virus,21 playing a crucial role in controlling CoV infections. Type-I IFNs could enhance the function of immune cells, including antigen-presenting cells (APCs), natural killer (NK) cells, T cells, and B cells.22,23
Lung resident respiratory dendritic cells (DCs) seem to process the acquired viral particles or antigens from the SARS-CoV-2-infected cells, then present it to the naïve circulating T cells in draining lymph nodes.21,24 The activated T cells, migrate to the infection site and secrete effector cytokines such as IFN-γ, which directly inhibits viral infection. Th1 type immune response seems to play a crucial role in the effective control of SARS-CoV infection. Higher rates of mortality was detected in patients with more serum Th2 cytokines.21 The exposure of naïve T cells to IFN-I, IFN-γ, and IL-12 is vital for Th1 cell polarization.25 Since neutralizing antibodies (NAbs) limit the viral infections, delayed and weak antibody production was also associated with poor clinical outcome in SARS.20 Moreover, the timing of IFN-I induction also seems crucial for the fine-tuning of B cell activation and consequent NAb production.22
3. Dysregulation of immune responses in COVID-19
Coronaviruses can circumvent the immune responses, particularly IFN-mediated antiviral responses, and replicate to reach a high peak load.26 Similar to SARS and MERS, substantial dysregulations of immunological responses occur in SARS-CoV-2 infection.20 Significant T cell lymphopenia, in particular CD4 + T cells, an elevated exhaustion level of T cells with reduced functionality, lower percentages of monocytes, eosinophils, and basophils, as well as higher neutrophil-lymphocyte-ratio (NLR), were observed in COVID-19 patients in different studies.27–29 The elevated levels of IL-1B, IFN-γ, IFN-γ-inducible protein 10 kDa (CXCL10; IP10), monocyte chemoattractant protein (MCP)1, as well as IL-4 and IL-10, which are related to Th1 and Th2 responses, respectively, were also identified.
The enhanced proinflammatory cytokines, to some extent, could contribute to the increased infiltration and activation of leukocytes in the lungs and subsequent lethal pneumonia.30,31 Additionally, the elevated levels of IL-6 is considered as an early biomarker of the aggravation of COVID-19 clinical course, which is associated with cytokine storm.32
Fu et al.20 categorized SARS-CoV-2-mediated inflammatory responses into two different stages. The primary response, which occurs due to viral replication and consequently mounts the host antiviral responses and viral-induced ACE2 downregulation and shedding. The primary inflammatory response is almost tolerated by patients and has a protective role against the infection through viral load reduction or viral clearance.
These possible mechanisms, in turn, cause elevated levels of proinflammatory cytokines/chemokine and cellular damage due to apoptosis and pyroptosis,33 shaping the second stage of immune responses. This stage is initiated with the generation of adaptive immunity and NAb secretion.23 The produced NAbs could trigger Fc receptor-mediated antibody-dependent enhancement (ADE) (Figure 1b), which possibly occurs due to early and sub-optimal antibody activity. This process could cause persistent viral replication, skewing of the macrophage responses, and further exuberant inflammatory responses, and subsequent cellular damage and lung injury.31 Noteworthy, the possible role of ADE in COVID-19 severity was also suggested by other researchers recently.32
Besides, given the crucial role of T cells, in particular CD4+ and CD8 + T cells, in modulating the over-activated inflammatory responses, some studies suggested that suboptimal and decreased T cell numbers generated by SARS-CoV-2 infection may also result in weak cellular immunity and undesired inflammatory responses.27,34
3.1. The possible role of IFNs in dysregulated immune responses
The relative timing of the IFN-I response and maximal viral replication was shown to contribute to the disease severity in SARS and MERS.30,35 Notably, IFN therapy before the virus titer peak decreased inflammatory cytokines production and generated protective roles in MERS-CoV infected mice. In contrast, at the later stages of the disease (two or 4-days post-infection), enhanced proinflammatory cytokines, increased infiltration, and the higher total number of highly activated monocytes, macrophages, and neutrophils may result in fatal pneumonia.35
The key role of dysregulated IFN-mediated immune responses in SARS-CoV-2 infection was uncovered as well. Despite heavier virus replication than SARS-CoV, SARS-CoV-2 does not significantly induce type-I, -II, or -III IFNs.36 Consistent with this finding, it was revealed that IFN-I deficiency could be considered as a hallmark of severe COVID-19.37,38 Moreover, impaired IFN-mediated immune responses, with a reduced dynamic range, in response to direct pattern recognition receptor (PRR) stimulation and viral infection were uncovered in older individuals.39 The impaired IFN antiviral responses could, to some extent, explain the poor clinical outcomes in elderlies.32 In congruence with these studies, Hadjadj et.al suggested that IFN administration could be used to overcome the IFN-I deficiency issue. On the other hand, applying anti-inflammatory agents that target IL-6 or TNFα, could, to some extent, dampen the inflammation and subsequent immunopathogenic over-reactions.38
4. Taking advantage of available vaccines against COVID-19
As mentioned before, using LAVs might help in reducing the risks of other infections through NSEs.11 In the following, the possible heterologous effects of BCG and influenza vaccines will be discussed. The impact of BCG, influenza vaccine, and IFN-I on the immune system in comparison with the immunity dysregulation caused by COVID-19 or aging are presented in Table 2.
Table 2.
The impact of BCG, influenza vaccine, and interferon (IFN)-I on the immune system compared with the dysregulation of immunity as a result of COVID-19 or aging
| BCG vaccine | Influenza vaccines | IFN-I | Aging | COVID-19 | |
|---|---|---|---|---|---|
| PRRs | Engaging TLR2, TLR4, TLR8, and C-type lectin receptors,40 NOD-like receptors, RIG-I41 | Engaging TLR7,42 LAIV: increasing expression of RIG-I and TLR-343 | Enhancing TLR responsiveness in macrophages44 | Decline in TLR expression and function45 | |
| Proinflammatory mediators | *Inducing the production of pro-inflammatory cytokines such as TNFα, IL-1β, and IL-646 | *Increasing the production of TNF-α and IL-6 and downregulation of IL-1b, IFN-γ, and IL-10 after stimulation of PBMCs with LPS, M. tuberculosis, C. albicans, and S. aureus.47 TIV/MF59: significant increase in IL-5 and IL-6, IFN-γ, IL-2, Th2 responses,48 AS03: STAT1 and MX1 upregulation49 | Cytokines/chemokine regulation such as IL-15 and IFN-γ50 | Increase in proinflammatory cytokines such as TNF, IL-1, and IL-651,52 | Elevated levels of IL1β, IFN-γ, IP10, MCP1, IL4, and IL10 at disease late stages, higher concentrations of GCSF, IP10, MCP1, MIP1A, and TNFα in ICU patients versus non-ICU patients,27 elevated levels of IL-6, which is associated with cytokine storm32 |
| IFNs [production and duration] | *Increasing IFN-γ production,53 inducing a negative regulator of JAK/STAT signaling pathway, SOCS154 | TIV: early induction of type-I and -III IFNs,55 LAIV: increased expression of IFN-related genes [mostly IFN-I] in three days, including STAT1, STAT2, TLR7, IRF3, and IRF7, slighter changes by unadjuvanted TIV,56 TIV and LAIV induce overexpression of IFN-related genes55 | Significant impairment of antiviral IFN responses, reduction in the magnitude of the inducible responses, lowering IFN-I production in DCs39 | Antagonizing the IFN-mediated immune responses by several strategies21,27 | |
| Neutrophils | *Increasing TNF-α, IL-6, and IL-12 production in lymph nodes41 | MF59: inducing CCL3 and CXCL8 chemoattractants,49 AS03: inducing neutrophil chemoattractants49 | Effect on recruitment by suppression of CXCL1 and CXCL2 production57 | Impairment of phagocytic and chemotactic abilities45 | Elevated levels of neutrophils58 |
| NK cells | *Inducing the production of IFN-γ,59 *inducing trained immunity in NK cells46 | *TIV: increased NK cell activity until 30 days, with a peak at day six,60 possibly due to increased IFN-α production,61 *enhancing the functionality of NK cells62 | Enhanced function and survival of NK cells,23 effects on NK cell recruitment by induction of CCL3, CCL4, and CCL557 | Decline in the functional capacity of NK cells45 | Reduced total number of NK cells58 |
| DCs | BCG sensing leads to DC maturation and migration, consequent co-stimulatory molecules expression, and pro-inflammatory mediators production41 |
MF59: activation of DCs49 | Potent induction of DC maturation and migration,24 enhanced expression of MHC and co-stimulatory molecules resulting in increasing their ability to induce T cells57 | Reduction of phagocytosis, pinocytosis, migratory capacity, and Ag presentation,45 dysregulation of inflammatory cytokine production, such as lower IFN-I production39 | |
| Macrophages | *Inducing GM-CSF production,40 *inducing trained immunity in macrophages, and shifting to an M1-like phenotype46 | MF59: activation of macrophages49 | Effects on cytokine production and antibody-dependent cytotoxicity,50 upregulation of IL-10 and PDL1 and down-regulation of IFN-γ receptor expression57 | Reduction of TLR expression and pro-inflammatory cytokines production, accumulation of alternatively activated [M2-like] macrophages45 | Macrophage infection, which subsequently leads to viral spread and excessive inflammation19 |
| T cells | *Inducing nonspecific lymphocyte responses through both cross-reactivity and bystander activation,11,13 *enhancing the responsiveness of Th1 and Th17 corresponding cytokine induction46 | *Mounting heterologous cellular immune responses against 2009 A[H1N1] pandemic influenza virus by 2007/2008 TIV/MF59,63 AS03: Inducing the upregulation of CD4 T cell responses,64 MF59: shifting toward Th2 responses49 | Direct activation of CD4+ and CD8 + T cells, enhancing ability of CD4 + T cells to help B cells,23 Th1 induction dependent on exposure to IL-12, IFN-I, and IFN-γ25 | Reduction in the number of naïve T cells and elevation in senescent or exhausted T cells, shift toward a Th2-like phenotype45 | Significant T cell lymphopenia,28,65 elevated exhaustion of T cells and reduced functionality,29 increased in the mortality rate of patients with more serum Th2 cytokines [17] |
| B cells | *Enhancing the Ab-mediated responses to nonspecific pathogen or vaccine11,13 | *Mounting heterologous humoral immune responses against nonspecific influenza strains.63 | Promoting B cells activation and antibody responses in the early stages of infection23 | Decrease in the ability of B cells to mount an optimal Ab response45 |
* represents relatedness to nonspecific effects.
4.1. BCG vaccine
BCG vaccine is a LAV used in many countries early after birth against tuberculosis (TB) disease. It has been injected to over four billion people so far.66 BCG vaccination is also recommended to reduce the mortalities of pandemics.67
Numerous studies have indicated the non-specific protective effects of BCG vaccination against viral infections caused by both RNA and DNA viruses. For instance, BCG vaccination could reduce the risk of respiratory tract infections in elderlies and adolescents and substantially enhance the responsiveness to influenza A and hepatitis B vaccines.67
The heterologous effects of BCG vaccination against viral infections may be exerted through several mechanisms. One fundamental mechanism is trained immunity induction. BCG vaccination could stimulate immunological memory in NK cells, monocytes, and macrophages.46,68 It also led to enhanced IFN-γ and monocyte-derived cytokines ‘four- to seven-fold’ and ‘two-fold’, respectively, in healthy volunteers.69
The importance of rapid and robust innate immune responses, as the primary SARS-CoV-2-mediated inflammatory response, in reducing viral load or even viral clearance is well-established.20 However, the raised production of proinflammatory cytokines after BCG vaccination and subsequent cellular damage due to apoptosis or proptosis should be considered.20,33
On the other hand, BCG vaccination could also induce non-specific lymphocyte responses through both cross-reactivity and bystander activation.11,13 For instance, BCG vaccination prior to pathogen insult or vaccination boosted the antibody-mediated responses.46 Moreover, it could enhance the responsiveness of Th1 to non-specific secondary infections as well as Th17 corresponding cytokine induction, such as IFN-γ, up to 1 year.46
The beneficial impacts of BCG vaccine on adaptive immunity could reasonably dampen the secondary inflammatory responses to SARS-CoV-2 and the consequent multi-organ failure.13,70
While there are multiple publications on BCG-induced NSEs for viral infections, there is a concern for its use in COVID-19 prophylaxis; as it induces the suppressor of cytokine signaling 1 (SOCS1), a crucial negative regulator in the JAK-STAT signaling pathway,54 which can lead to IFN signaling suppression. To minimize this obstacle, Mizuno et al.54 demonstrated that the administration of SOCS1 antagonist-expressing recombinant BCG enhanced the immune responses in a mouse model. Interestingly, BCG plus IFN-α, as a vaccine adjuvant,71 could promote Th1 type cytokines secretion and consequent effective immune responses.72 Moreover, it has been revealed that pretreatment of DCs with IFN-β results in the production of larger amount of IL-12p70 and IL-12, which could improve DC function and causes consequent enhancement in Th1 responses. The authors conclude that IFN-β could be used as an adjuvant and enhance the BCG immunogenicity.73,74 Another study revealed that the treatment of cells with IFN-β or IFN-γ created epigenetic memory resulting in faster and higher IFN-stimulated gene (ISG) induction after re-stimulation.75 Therefore, the administration of BCG plus IFN might rationally antagonize the adverse regulatory effects of SOCS1 and augment BCG-induced antiviral immune responses to some extent.
However, BCG vaccine-related pulmonary complications, such as hypersensitivity reactions and mycobacterial pneumonia, have to be considered.76 Thus, cautious studies should be performed to investigate the potential synergistic effects and safety profile of the proposed approach.
4.2. Influenza vaccines
Different types of influenza vaccines, including live-attenuated influenza vaccine (LAIVs) and inactivated influenza vaccines (IIVs), are marketed containing the antigens from certain viral strains. Some formulations also include adjuvants.77 Such diverse constituents cause variety in cytokine induction capabilities of influenza vaccines.78 For instance, TIV (trivalent subunit inactivated influenza vaccine) showed a stronger early induction of type-I and -III IFNs and higher amounts of activated DCs and proinflammatory cytokines, such as TNF, IL-6, −10, and −1β, than LAIV in an in vitro research on unadjuvanted influenza vaccines. Moreover, TIVs have shown some adverse events in children.79
On the other hand, LAIVs were found to protect against respiratory infections80 and induce innate immunity through various methods,56 while also triggering a broader spectrum of immunity against different serotypes of influenza virus than TIVs.81
Both TIV and LAIV induced the expression of IFN genes, which was observed on day one and seven after vaccination for TIV and LAIV, respectively.55 It was suggested that the trained immunity induced by influenza vaccine is, to some extent, related to induction of the pro-inflammatory mediators and IFN production.62,80
It is hypothesized that a respiratory virus infection confers immunity against the same and other respiratory viruses for a short time, perhaps a few weeks. Such protection is because of activation of the innate immune response mediated by the release of IFN-I and other cytokines that have broad protective effects against a range of viruses. This phenomenon is called viral interference.82 To inspect this concept, the short-term non-specific protection of cold-adapted, live attenuated influenza vaccine (CAIV) against subsequent RSV infection was investigated in a mouse infection model80 The author hypothesized that since influenza vaccine and RSV share common features in term of pathogenesis, influenza vaccine could attenuate the severity of RSV infection. Noteworthy, the protective effects of influenza vaccine were significantly diminished in TLR3-/-TLR7-/-mice, which suggests the importance of TLR3/7 signaling pathways in the beneficial protective effects of influenza vaccine. The author proposed that the heterologous effects of influenza vaccine against MERS infection need further investigations.80 An epidemiological article also validated this effect on RSV infection.82
The immune responses to CoV and influenza infections seem comparable as they share some common features, including stimulation of TLRs and RIG-I as well as subsequent antiviral-mediated immune responses.21 Therefore, the potential role of influenza vaccine in generating a non-specific, short term antiviral effect against SARS-CoV-2 has been suggested.80 In this regard, a few recent reports have displayed a hypothetical beneficial role of influenza vaccines against COVID-19 infection in high-risk groups.81,83 While Salem et al. described a flu-induced bystander effect of the generated immune responses as a probable protective mechanism, Kiseleva et al. opined that the LAIV might be more suitable than IIV because of its broad-spectrum potency.
On the other hand, there are concerns about the dampening effect of influenza vaccines on the immune responses to SARS-CoV-2, as well. Because influenza infection can result in TLR desensitization for several months, suppressing one of the main defense mechanisms of the innate immunity against COVID-19.84 Furthermore, influenza disease has shown some bizarre phenomena such as “vaccine-associated viral interference”, which implies that a viral infection would generate a non-specific short-time protection against other viruses normally,85 which could be impeded when a vaccine is administered.86 Another concern is “original antigenic sin” phenomenon, which denotes that in encountering with a similar or close pathogen, the immune system may fail to develop an effective immune response against the newer pathogen and would depend on the memory of the previous pathogen.87 Therefore, a successful vaccination should include all the subtypes of a pathogen.88 This phenomenon is highly important in vaccine design, expressing the importance of extensive clinical trials to validate the safety and side effects of vaccines.89 However, original antigenic sin has been rarely reported in a cross-disease manner and is mostly important for expected mutations in the same pathogen family. A similar phenomenon is “heterosubtypic non-specific temporary immunity”, in which the previous infection with a pathogen temporarily reduces the risk of future infections with another subtype. The phenomenon has been observed for influenza in the unvaccinated populations who had previous seasonal influenza infections and were less susceptible to get infected with pandemic influenza A (H1N1) 2009.90,91 It is also rarely observed among different families of pathogens.92,93 However, a substantial negative association between influenza vaccination and higher COVID-19 incidence or other infections has not been reported.94,95
All in all, the positive and negative sides of the influenza vaccines should be further investigated to evaluate their impact on COVID-19. On the other hand, BCG vaccination has been suggested as an adjuvant for influenza vaccination to increase its efficacy, especially in elderlies.96 It could be conferred from Table 2 that influenza vaccine could effectively induce IFN mediated immune responses. Hence, the co-administration of BCG and influenza vaccine might harness the beneficial heterologous effects of these vaccines and be a potentially effective approach to combat COVID-19. However, it needs to be validated through experiments.
5. Conclusion and future directions
Given the role of immune system overreactions in COVID-19-induced cytokine storm and the resultant serious harm to body organs and even death, the interventions that could reverse such hyperactivated immunological responses could be of high value. In this regard, LAVs, especially BCG vaccine, could hinder viral replication and the subsequent pathological inflammatory responses in COVID-19 via the arousal of innate and adaptive immunity, especially in the aged and immunocompromised groups. However, the probable adverse effects of BCG vaccination should be considered, including the induction of SOCS1 expression, which may cause the suppression of IFN signaling. In view of the importance of IFN signaling pathway in reducing viral replication early after the infection, the use of a SOCSI-antagonist expressing recombinant BCG or a combination of BCG vaccine with IFN-I might help to partly reverse this detrimental effect and boost the immune system successfully against SARS-CoV-2. These approaches deserve more investigation and experience to be validated in respect to protocol efficacy and safety.
The co-administration of BCG and influenza vaccine might also be potentially a candidate approach to combat COVID-19 and could probably exploit the positive impacts of these vaccines. However, the probability of undesired immunopathological overreactions and cumulative immune aberrations generated by the two vaccines should be taken seriously. In this respect, the risk of augmented innate immune responses, in particular, overproduction of proinflammatory mediators and overstimulation of immune responses, should also be regarded.
In a nutshell, if any vaccine is to be used to support against COVID-19, while compensation for the virus-induced attenuated immune response is the objective, maintaining a balance during both innate and adaptive immune responses should be considered as well. Further studies are highly recommended to assess the outcomes of such interventions.
Funding Statement
This study was supported by [Grant No. 99-01-106-22199] from the Research Council of Shiraz University of Medical Sciences, Shiraz, Iran.
Disclosure of potential conflicts of interest
The authors declare no conflict of interests.
References
- 1.Negahdaripour M. The battle against covid-19: where do we stand now? Iran J Med Sci. 2020;45:81–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Negahdaripour M. A world of changes: the inheritance of COVID-19. Iran J Med Sci. 2020;45:155–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Amirlak I, Haddad R, Hardy JD, Khaled NS, Chung MH, Amirlak B. Effectiveness of booster BCG vaccination in preventing Covid-19 infection. medRxiv 2020; 2020.08.10.20172288. doi: 10.1101/2020.08.10.20172288. [DOI] [PMC free article] [PubMed]
- 4.Dayal D, Gupta S. Connecting BCG vaccination and COVID-19: additional data. medRxiv 2020; 2020.04.07.20053272. doi: 10.1101/2020.04.07.2005327.2 [DOI]
- 5.Escobar LE, Molina-Cruz A, Barillas-Mury C. BCG vaccine protection from severe coronavirus disease 2019 (COVID-19). Proc Natl Acad Sci U S A. 2020;117:17720–26. doi: 10.1073/pnas.2008410117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hamiel U, Kozer E, Youngster I. SARS-CoV-2 rates in BCG-vaccinated and unvaccinated young adults. JAMA - J Am Med Assoc. 2020;323:2340–41. doi: 10.1001/jama.2020.8189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Riccò M, Gualerzi G, Ranzieri S, Luigi Bragazzi N. Stop playing with data: there is no sound evidence that bacille calmette-guérin may avoid SARS-CoV-2 infection for now. Acta Biomed. 2020;91:207–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.BCG vaccination to protect healthcare workers against COVID-19 - ClinicalTrials.gov [Internet]. [cited 2020. September 1]. Available from: https://clinicaltrials.gov/ct2/show/NCT04327206
- 9.ClinicalTrials.gov [Internet]. [cited 2020. September 1]. Available from: https://clinicaltrials.gov/.
- 10.Netea MG, Joosten LAB, Latz E, Mills KHG, Natoli G, Stunnenberg HG, O’Neill LAJ, Xavier RJ. Trained immunity: A program of innate immune memory in health and disease. Science. 2016;352(6284):427. doi: 10.1126/science.aaf1098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Goodridge HS, Ahmed SS, Curtis N, Kollmann TR, Levy O, Netea MG, Pollard AJ, Van Crevel R, Wilson CB. Harnessing the beneficial heterologous effects of vaccination. Nat Rev Immunol. 2016;16(6):392–400. doi: 10.1038/nri.2016.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Higgins J, Reingold A. Systematic review of the non-specific effects of BCG, DTP and measles containing vaccines. Wkly Epidemiol Rec. 2014;89:1–34.24466571 [Google Scholar]
- 13.Messina NL, Zimmermann P, Curtis N. The impact of vaccines on heterologous adaptive immunity. Clin Microbiol Infect. 2019;25(12):1484–93. doi: 10.1016/j.cmi.2019.02.016. [DOI] [PubMed] [Google Scholar]
- 14.To KKW, Tsang OTY, Leung WS, Tam AR, Wu TC, Lung DC, Yip CCY, Cai JP, Chan JMC, Chik TSH, et al. Temporal profiles of viral load in posterior oropharyngeal saliva samples and serum antibody responses during infection by SARS-CoV-2: an observational cohort study. Lancet Infect Dis. 2020;20(5):565–74. doi: 10.1016/S1473-3099(20)30196-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hoffmann M, Kleine-Weber H, Schroeder S, Krüger N, Herrler T, Erichsen S, Schiergens TS, Herrler G, Wu NH, Nitsche A, et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell. 2020;181(2):271–80. doi: 10.1016/j.cell.2020.02.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Owji H, Negahdaripour M, Hajighahramani N. Immunotherapeutic approaches to curtail COVID-19. Int Immunopharmacol. 2020;88:106924. doi: 10.1016/j.intimp.2020.106924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Antalis TM, Bugge TH, Wu Q. Membrane-anchored serine proteases in health and disease. Prog Mol Biol Transl Sci. 2011;99:1–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Yongwen C, Feng Z, Diao B, Wang R, Wang G, Wang C, Tan Y, Liu L, Wang C, Liu Y, et al. The novel severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) directly decimates human spleens and lymph nodes. medRxiv 2020; 2:2020.03.27.20045427; doi: 10.1101/2020.03.27.20045427 [DOI]
- 19.Park MD. Macrophages: a Trojan horse in COVID-19? Nat Rev Immunol. 2020;20(6):351. doi: 10.1038/s41577-020-0317-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Fu Y, Cheng Y, Wu Y. Understanding SARS-CoV-2-mediated inflammatory responses: from mechanisms to potential therapeutic tools. Virol Sin. 2020;35:266–71. doi: 10.1007/s12250-020-00207-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Prompetchara E, Ketloy C, Palaga T. Immune responses in COVID-19 and potential vaccines: lessons learned from SARS and MERS epidemic. Asian Pac J Allergy Immunol. 2020;38:1–9. [DOI] [PubMed] [Google Scholar]
- 22.Crouse J, Kalinke U, Oxenius A. Regulation of antiviral T cell responses by type i interferons. Nat Rev Immunol. 2015;15:231–42. doi: 10.1038/nri3806. [DOI] [PubMed] [Google Scholar]
- 23.McNab F, Mayer-Barber K, Sher A, Wack A, O’Garra A. Type I interferons in infectious disease. Nat Rev Immunol. 2015;15:87–103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Braciale TJ, Hahn YS. Immunity to viruses. Immunol Rev. 2013;255:5–12. doi: 10.1111/imr.12109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Swain SL, McKinstry KK, Strutt TM. Expanding roles for CD4 + T cells in immunity to viruses. Nat Rev Immunol. 2012;12:136–48. doi: 10.1038/nri3152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kindler E, Thiel V, Weber F. Interaction of SARS and MERS coronaviruses with the antiviral interferon response. Adv Virus Res. 2016;96:219–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Qin C, Zhou L, Hu Z, Zhang S, Yang S, Tao Y, Xie C, Ma K, Shang K, Wang W, et al. Dysregulation of immune response in patients with COVID-19 in Wuhan, China. SSRN Electron J. 2020. doi: 10.2139/ssrn.3541136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Diao B, Wang C, Tan Y, Chen X, Liu Y, Ning L, Chen L, Li M, Liu Y, Wang G, et al. Reduction and functional exhaustion of T cells in patients with coronavirus disease 2019 (COVID-19). Front Immunol. 2020. doi: 10.1016/fimmu.2020.00827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zheng HY, Zhang M, Yang CX, Zhang N, Wang XC, Yang XP, Dong XQ, Zheng YT. Elevated exhaustion levels and reduced functional diversity of T cells in peripheral blood may predict severe progression in COVID-19 patients. Cell Mol Immunol. 2020;17:541–43. doi: 10.1038/s41423-020-0401-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Channappanavar R, Perlman S. Pathogenic human coronavirus infections: causes and consequences of cytokine storm and immunopathology. Semin Immunopathol. 2017;39:529–39. doi: 10.1007/s00281-017-0629-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Cao X. COVID-19: immunopathology and its implications for therapy. Nat Rev Immunol. 2020;20:269–70. doi: 10.1038/s41577-020-0308-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Matricardi PM, Dal Negro RW, Nisini R. The first, holistic immunological model of COVID-19: implications for prevention, diagnosis, and public health measures. Pediatr Allergy Immunol. 2020;31:454–70. doi: 10.1111/pai.13271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Yang M. Cell pyroptosis, a potential pathogenic mechanism of 2019-nCoV infection. SSRN Electron J. 2020. doi: 10.2139/ssrn.3527420. [DOI] [Google Scholar]
- 34.Channappanavar R, Fehr AR, Vijay R, Mack M, Zhao J, Meyerholz DK, Perlman S. Dysregulated type I interferon and inflammatory monocyte-macrophage responses cause lethal pneumonia in SARS-CoV-infected mice. Cell Host Microbe. 2016;19:181–93. doi: 10.1016/j.chom.2016.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Channappanavar R, Fehr AR, Zheng J, Wohlford-Lenane C, Abrahante JE, Mack M, Sompallae R, McCray PB, Meyerholz DK, Perlman S. IFN-I response timing relative to virus replication determines MERS coronavirus infection outcomes. J Clin Invest. 2019;129:3625–39. doi: 10.1172/JCI126363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Chu H, Chan JFW, Wang Y, Yuen TTT, Chai Y, Hou Y, Shuai H, Yang D, Hu B, Huang X, et al. Comparative replication and immune activation profiles of SARS-CoV-2 and SARS-CoV in human lungs: an ex vivo study with implications for the pathogenesis of COVID-19. Clin Infect Dis. 2020;71:1400–09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wei L, Ming S, Zou B, Wu Y, Hong Z, Li Z, Zheng X, Huang M, Luo L, Liang J, et al. Viral invasion and type i interferon response characterize the immunophenotypes during COVID-19 infection. SSRN Electron J. 2020. doi: 10.2139/ssrn.3555695. [DOI] [Google Scholar]
- 38.Hadjadj J, Yatim N, Barnabei L, Corneau A, Boussier J, Pere H, Charbit B, Bondet V, Chenevier-Gobeaux C, Breillat P, et al. Impaired type I interferon activity and exacerbated inflammatory responses in severe Covid-19 patients. Science. 2020;369(6504):718–72461059. doi: 10.1126/science.abc6027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Molony RD, Malawista A, Montgomery RR. Reduced dynamic range of antiviral innate immune responses in aging. Exp Gerontol. 2018;107:130–35. doi: 10.1016/j.exger.2017.08.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Angelidou A, Diray-Arce J, Conti MG, Smolen KK, van Haren SD, Dowling DJ, Husson RN, Levy O. BCG as a case study for precision vaccine development: lessons from vaccine heterogeneity, trained immunity, and immune ontogeny. Front Microbiol. 2020;11:332. doi: 10.3389/fmicb.2020.00332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Covián C, Fernández-Fierro A, Retamal-Díaz A, Díaz FE, Vasquez AE, Lay MK, Riedel CA, González PA, Bueno SM, Kalergis AM. BCG-induced cross-protection and development of trained immunity: implication for vaccine design. Front Immunol. 2019;10:2806. doi: 10.3389/fimmu.2019.02806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Pulendran B, Maddur MS. Innate immune sensing and response to influenza. Curr Top Microbiol Immunol. 2015;386:23–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Fischer WA, Chason KD, Brighton M, Jaspers I. Live attenuated influenza vaccine strains elicit a greater innate immune response than antigenically-matched seasonal influenza viruses during infection of human nasal epithelial cell cultures. Vaccine. 2014;32:1761–67. doi: 10.1016/j.vaccine.2013.12.069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Sirén J, Pirhonen J, Julkunen I, Matikainen S. IFN-α regulates TLR-dependent gene expression of IFN-α, IFN-β, IL-28, and IL-29. J Immunol. 2005;174:1932–37. doi: 10.4049/jimmunol.174.4.1932. [DOI] [PubMed] [Google Scholar]
- 45.Oh SJ, Lee JK, Shin OS. Aging and the immune system: the impact of immunosenescence on viral infection, immunity and vaccine immunogenicity. Immune Netw. 2019;19(6):e37. doi: 10.4110/in.2019.19.e37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Uthayakumar D, Paris S, Chapat L, Freyburger L, Poulet H, De Luca K. Non-specific effects of vaccines illustrated through the BCG example: from observations to demonstrations. Front Immunol. 2018;9:2869. doi: 10.3389/fimmu.2018.02869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Bessler H, Djaldetti M. Research article global vaccines and immunology glob vaccines immunol. Glob Vaccines Immunol. 2016;2:1–4. [Google Scholar]
- 48.Valensi JP, Carlson JR, Van Nest GA. Systemic cytokine profiles in BALB/c mice immunized with trivalent influenza vaccine containing MF59 oil emulsion and other advanced adjuvants. J Immunol. 1994;153:4029–39. [PubMed] [Google Scholar]
- 49.Tregoning JS, Russell RF, Kinnear E. Adjuvanted influenza vaccines. Hum Vaccines Immunother. 2018;14:550–64. doi: 10.1080/21645515.2017.1415684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Hervas-Stubbs S, Perez-Gracia JL, Rouzaut A, Sanmamed MF, Le Bon A, Melero I. Direct effects of type I interferons on cells of the immune system. Clin Cancer Res. 2011;17:2619–27. doi: 10.1158/1078-0432.CCR-10-1114. [DOI] [PubMed] [Google Scholar]
- 51.Minciullo PL, Catalano A, Mandraffino G, Casciaro M, Crucitti A, Maltese G, Morabito N, Lasco A, Gangemi S, Basile G. Inflammaging and anti-inflammaging: the role of cytokines in extreme longevity. Arch Immunol Ther Exp (Warsz). 2016;64:111–26. doi: 10.1007/s00005-015-0377-3. [DOI] [PubMed] [Google Scholar]
- 52.Franceschi C, Garagnani P, Parini P, Giuliani C, Santoro A. Inflammaging: a new immune–metabolic viewpoint for age-related diseases. Nat Rev Endocrinol. 2018;14:576–90. doi: 10.1038/s41574-018-0059-4. [DOI] [PubMed] [Google Scholar]
- 53.Kandasamy R, Voysey M, McQuaid F, De Nie K, Ryan R, Orr O, Uhlig U, Sande C, O’Connor D, Pollard AJ. Non-specific immunological effects of selected routine childhood immunisations: systematic review. BMJ. 2016;355:i5225. doi: 10.1136/bmj.i5225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Mizuno S, Soma S, Inada H, Kanuma T, Matsuo K, Yasutomi Y. SOCS1 antagonist–expressing recombinant bacillus Calmette–Guérin enhances antituberculosis protection in a mouse model. J Immunol. 2019;203:188–97. doi: 10.4049/jimmunol.1800694. [DOI] [PubMed] [Google Scholar]
- 55.Cao RG, Suarez NM, Obermoser G, Lopez SMC, Flano E, Mertz SE, Albrecht RA, García-Sastre A, Mejias A, Xu H, et al. Differences in antibody responses between trivalent inactivated influenza vaccine and live attenuated influenza vaccine correlate with the kinetics and magnitude of interferon signaling in children. J Infect Dis. 2014;210:224–33. doi: 10.1093/infdis/jiu079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Nakaya HI, Wrammert J, Lee EK, Racioppi L, Marie-Kunze S, Haining WN, Means AR, Kasturi SP, Khan N, Li GM, et al. Systems biology of vaccination for seasonal influenza in humans. Nat Immunol. 2011;12:786–95. doi: 10.1038/ni.2067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lee AJ, Ashkar AA. The dual nature of type I and type II interferons. Front Immunol. 2018;9:2061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Chen N, Zhou M, Dong X, Qu J, Gong F, Han Y, Qiu Y, Wang J, Liu Y, Wei Y, et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: a descriptive study. Lancet. 2020;395:507–13. doi: 10.1016/S0140-6736(20)30211-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Bernatowska E, Skomska-Pawliszak M, Wolska-Kuśnierz B, Pac M, Heropolitanska-Pliszka E, Pietrucha B, Bernat-Sitarz K, Dąbrowska-Leonik N, Bohynikova N, Piątosa B, et al. BCG moreau vaccine safety profile and NK cells—double protection against disseminated BCG infection in retrospective study of BCG vaccination in 52 polish children with severe combined immunodeficiency. J Clin Immunol. 2020;40:138–46. doi: 10.1007/s10875-019-00709-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Schapiro JM, Segev Y, Rannon L, Alkan M, Rager‐Zisman B. Natural killer (NK) cell response after vaccination of volunteers with killed influenza vaccine. J Med Virol. 1990;30:196–200. doi: 10.1002/jmv.1890300310. [DOI] [PubMed] [Google Scholar]
- 61.Tai LH, Zhang J, Scott KJ, De Souza CT, Alkayyal AA, Ananth AA, Sahi S, Adair RA, Mahmoud AB, Sad S, et al. Perioperative influenza vaccination reduces postoperative metastatic disease by reversing surgery-induced dysfunction in natural killer cells. Clin Cancer Res. 2013;19:5104–15. doi: 10.1158/1078-0432.CCR-13-0246. [DOI] [PubMed] [Google Scholar]
- 62.Brillantes M, Beaulieu AM. Memory and memory-like NK cell responses to microbial pathogens. Front Cell Infect Microbiol. 2020;10:102. doi: 10.3389/fcimb.2020.00102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Iorio AM, Bistoni O, Galdiero M, Lepri E, Camilloni B, Russano AM, Neri M, Basileo M, Spinozzi F. Influenza viruses and cross-reactivity in healthy adults: humoral and cellular immunity induced by seasonal 2007/2008 influenza vaccination against vaccine antigens and 2009 A(H1N1) pandemic influenza virus. Vaccine. 2012;30:1617–23. doi: 10.1016/j.vaccine.2011.12.107. [DOI] [PubMed] [Google Scholar]
- 64.Moris P, Van Der Most R, Leroux-Roels I, Clement F, Dramé M, Hanon E, Leroux-Roels GG, Van Mechelen M. H5N1 influenza vaccine formulated with AS03 A induces strong Cross-reactive and polyfunctional CD4 T-Cell responses. J Clin Immunol. 2011;31:443–54. doi: 10.1007/s10875-010-9490-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Moon C. Fighting COVID-19 exhausts T cells. Nat Rev Immunol. 2020;20:277. doi: 10.1038/s41577-020-0304-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Dara M, Acosta CD, Rusovich V, Zellweger JP, Centis R, Migliori GB. Bacille Calmette-Guérin vaccination: the current situation in Europe. Eur Respir J. 2014;43:24–35. doi: 10.1183/09031936.00113413. [DOI] [PubMed] [Google Scholar]
- 67.Moorlag SJCFM, Arts RJW, van Crevel R, Netea MG. Non-specific effects of BCG vaccine on viral infections. Clin Microbiol Infect. 2019;25:1473–78. doi: 10.1016/j.cmi.2019.04.020. [DOI] [PubMed] [Google Scholar]
- 68.Netea MG, Domínguez-Andrés J, Barreiro LB, Chavakis T, Divangahi M, Fuchs E, Joosten LAB, van der Meer JWM, Mhlanga MM, Mulder WJM, et al. Defining trained immunity and its role in health and disease. Nat Rev Immunol. 2020;20:375–88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Kleinnijenhuis J, Quintin J, Preijers F, Joosten LAB, Ifrim DC, Saeed S, Jacobs C, Van Loenhout J, De Jong D, Hendrik S, et al. Bacille Calmette-Guérin induces NOD2-dependent nonspecific protection from reinfection via epigenetic reprogramming of monocytes. Proc Natl Acad Sci U S A. 2012;109:17537–42. doi: 10.1073/pnas.1202870109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Moore JB, June CH. Cytokine release syndrome in severe COVID-19. Science. 2020;368:473–74. doi: 10.1126/science.abb8925. [DOI] [PubMed] [Google Scholar]
- 71.Rizza P, Capone I, Moretti F, Proietti E, Belardelli F. IFN-α as a vaccine adjuvant: recent insights into the mechanisms and perspectives for its clinical use. Expert Rev Vaccines. 2011;10:487–98. doi: 10.1586/erv.11.9. [DOI] [PubMed] [Google Scholar]
- 72.Rivas-Santiago CE, Guerrero GG. IFN-α boosting of mycobacterium bovis Bacillus Calmette Güerin-vaccine promoted Th1 type cellular response and protection against M. tuberculosis Infection . Biomed Res Int. 2017;2017:8796760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Giacomini E, Remoli ME, Gafa V, Pardini M, Fattorini L, Coccia EM. IFN-β improves BCG immunogenicity by acting on DC maturation. J Leukoc Biol. 2009;85(3):462–68. doi: 10.1189/jlb.0908583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.El-Sahrigy SAF, Rahman AMOA, Samaha DY, Mohamed NA, Saber SM, Talkhan HA, Ismail GA, Ibraheem EM, Riad EM. The influence of interferon-β supplemented human dendritic cells on BCG immunogenicity. J Immunol Methods. 2018;457:15–21. doi: 10.1016/j.jim.2018.03.003. [DOI] [PubMed] [Google Scholar]
- 75.Kamada R, Yang W, Zhang Y, Patel MC, Yang Y, Ouda R, Dey A, Wakabayashi Y, Sakaguchi K, Fujita T, et al. Interferon stimulation creates chromatin marks and establishes transcriptional memory. Proc Natl Acad Sci U S A. 2018;115:E9162–71. doi: 10.1073/pnas.1720930115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Leeson CE, Ismail A, Hashad MM, Elmansy H, Shahrour W, Prowse O, Kotb A. Systematic review: safety of intravesical therapy for bladder cancer in the era of COVID-19. SN Compr Clin Med. 2020;18:1–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Fiore AE, Bridges CB, Katz JM, Cox NJ. Inactivated influenza vaccines. In: Plotkin S, Orenstein W, Offit P, editors. Vaccines. 6th Ed. Elsevier Saunders; 2012. p. 257–93. [Google Scholar]
- 78.van Essen GA, Beran J, Devaster JM, Durand C, Duval X, Esen M, Falsey AR, Feldman G, Gervais P, Innis BL, et al. Influenza symptoms and their impact on elderly adults: randomised trial of AS03-adjuvanted or non-adjuvanted inactivated trivalent seasonal influenza vaccines. Influenza Other Respi Viruses. 2014;8:452–62. doi: 10.1111/irv.12245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Cowling BJ, Fang VJ, Nishiura H, Chan KH, Ng S, Ip DKM, Chiu SS, Leung GM, Malik Peiris JS. Increased risk of noninfluenza respiratory virus infections associated with receipt of inactivated influenza vaccine. Clin Infect Dis. 2012;54:1778–83. doi: 10.1093/cid/cis307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Lee YJ, Lee JY, Jang YH, Seo SU, Chang J, Seong BL. Non-specific effect of vaccines: immediate protection against respiratory syncytial virus infection by a live attenuated influenza vaccine. Front Microbiol. 2018;9:83. doi: 10.3389/fmicb.2018.00083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Kiseleva I. New points of departure for more global influenza vaccine use. Vaccines. 2020;8:1–8. doi: 10.3390/vaccines8030410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Ånestad G. Interference between outbreaks of respiratory syncytial virus and influenza virus infection. Lancet. 1982;319:502. doi: 10.1016/S0140-6736(82)91466-0. [DOI] [PubMed] [Google Scholar]
- 83.Salem ML, El-Hennawy D. The possible beneficial adjuvant effect of influenza vaccine to minimize the severity of COVID-19. Med Hypotheses. 2020;140:109752. doi: 10.1016/j.mehy.2020.109752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Didierlaurent A, Goulding J, Patel S, Snelgrove R, Low L, Bebien M, Lawrence T, Van Rijt LS, Lambrecht BN, Sirard JC, et al. Sustained desensitization to bacterial Toll-like receptor ligands after resolution of respiratory influenza infection. J Exp Med. 2008;205:323–29. doi: 10.1084/jem.20070891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Linde A, Rotzén-Ostlund M, Zweygberg-Wirgart B, Rubinova S, Brytting M. Does viral interference affect spread of influenza? Euro Surveill. 2009;14:40. [PubMed] [Google Scholar]
- 86.Dyer O. What did we learn from Tamiflu? BMJ. 2020;368:m626. doi: 10.1136/bmj.m626. [DOI] [PubMed] [Google Scholar]
- 87.Vatti A, Monsalve DM, Pacheco Y, Chang C, Anaya JM, Gershwin ME. Original antigenic sin: A comprehensive review. J Autoimmun. 2017;83:12–21. doi: 10.1016/j.jaut.2017.04.008. [DOI] [PubMed] [Google Scholar]
- 88.Midgley CM, Bajwa-Joseph M, Vasanawathana S, Limpitikul W, Wills B, Flanagan A, Waiyaiya E, Tran HB, Cowper AE, Chotiyarnwon P, et al. An in-depth analysis of original antigenic sin in dengue virus infection. J Virol. 2011;85:410–21. doi: 10.1128/JVI.01826-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Suzuki M, Camacho A, Ariyoshi K. Potential effect of virus interference on influenza vaccine effectiveness estimates in test-negative designs. Epidemiol Infect. 2014;142:2642–46. doi: 10.1017/S0950268814000107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Kelly H, Barry S, Laurie K, Mercer G. Seasonal influenza vaccination and the risk of infection with pandemic influenza: A possible illustration of nonspecific temporary immunity following infection. Eurosurveillance. 2010;15:47. doi: 10.2807/ese.15.47.19722-en. [DOI] [PubMed] [Google Scholar]
- 91.Skowronski DM, de Serres G, Crowcroft NS, Janjua NZ, Boulianne N, Hottes TS, Rosella LC, Dickinson JA, Gilca R, Sethi P, et al. Association between the 2008-09 seasonal influenza vaccine and pandemic H1N1 illness during spring-summer 2009: four observational studies from Canada. PLoS Med. 2010;7:e1000258. doi: 10.1371/journal.pmed.1000258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Wolff GG. Influenza vaccination and respiratory virus interference among department of defense personnel during the 2017–2018 influenza season. Vaccine. 2020;38:350–54. doi: 10.1016/j.vaccine.2019.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Rynda-Apple A, Robinson KM, Alcorn JF. Influenza and bacterial superinfection: illuminating the immunologic mechanisms of disease. Infect Immun. 2015;83:3764–70. doi: 10.1128/IAI.00298-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Rikin S, Jia H, Vargas CY, Castellanos de Belliard Y, Reed C, LaRussa P, Larson EL, Saiman L, Stockwell MS. Assessment of temporally-related acute respiratory illness following influenza vaccination. Vaccine. 2018;36:1958–64. doi: 10.1016/j.vaccine.2018.02.105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.CDC . Key facts about seasonal flu vaccine [Internet]. Program2008 [cited 2020. August 17]; 1–3. Available from: http://www.cdc.gov/flu/protect/keyfacts.htm. [Google Scholar]
- 96.Leentjens J, Kox M, Stokman R, Gerretsen J, Diavatopoulos DA, Van Crevel R, Rimmelzwaan GF, Pickkers P, Netea MG. BCG vaccination enhances the immunogenicity of subsequent influenza vaccination in healthy volunteers: A randomized, placebo-controlled pilot study. J Infect Dis. 2015;212:1930–38. doi: 10.1093/infdis/jiv332. [DOI] [PubMed] [Google Scholar]