

Abstract
Clostridium sordellii is an emerging pathogen associated with highly-lethal female reproductive tract infections following childbirth, abortion, or cervical instrumentation. Gaps in our understanding of the pathogenesis of C. sordellii infections present major challenges to the development of better preventive and therapeutic strategies against this problem. We sought to determine the mechanisms whereby uterine decidual macrophages phagocytose this bacterium and tested the hypothesis that human decidual macrophages utilize class A scavenger receptors (CASR) to internalize unopsonized C. sordellii. In vitro phagocytosis assays with human decidual macrophages incubated with pharmacological inhibitors of CASRs (fucoidan, polyinosinic acid, and dextran sulfate) revealed a role for these receptors in C. sordellii phagocytosis. Soluble MARCO receptor prevented C. sordellii internalization, suggesting that MARCO is an important CASR in decidual macrophage phagocytosis of this microbe. Peritoneal macrophages from MARCO-deficient mice, but not wild-type or SR-AI/II-deficient mice, showed impaired C. sordellii phagocytosis. MARCO-null mice were more susceptible to death from C. sordellii uterine infection than wild-type mice and exhibited impaired clearance of this bacterium from the infected uterus. Thus, MARCO is an important phagocytic receptor utilized by human and mouse macrophages to clear C. sordellii from the infected uterus.
Introduction
The clostridia are anaerobic, spore-forming bacilli that cause a diverse array of toxin-mediated infections in humans, including botulism, tetanus, and antibiotic-associated diarrhea. Clostridium sordellii is an emerging pathogen associated with highly-lethal female reproductive tract infections, bacteremia, and soft tissue infections (1). The high mortality of C. sordellii infections is associated with a stereotypical toxic shock syndrome (2). A recent study estimated that nearly 1 in 200 deaths in women of reproductive age were associated with clostridial toxic shock, due to C. sordellii and/or the related C. perfringens (3). An increased number of severe C. sordellii infections has been reported over then past decade, following childbirth and abortion (4-6). Gaps in our understanding of the pathogenesis of C. sordellii infections present major challenges to the development of better preventive and therapeutic strategies against this emerging problem.
Macrophages are important in defending the host against invasive clostridial infections (7), although the role of uterine macrophages in innate immune defense against clostridia is undefined. During pregnancy, uterine decidual macrophages (DMs) participate in immunosurveillance, binding, ingesting, and clearing bacteria that ascend beyond the cervix (8). The receptor-mediated mechanisms by which DMs recognize and phagocytose unopsonized bacteria, including clostridia, have not been closely analyzed.
The macrophage scavenger receptors are a large family of immunosurveillance receptors utilized to recognize and internalize unopsonized pathogens, apoptotic host cells, and modified lipoproteins (9, 10). They are increasingly recognized to play a critical role in the clearance of unopsonized bacterial and parasitic microorganisms. Scavenger receptors are transmembrane proteins belonging to at least eight different subclasses (A-H) based on their tertiary structure (9, 10). The class A scavenger receptors (CASRs) are widely expressed on macrophages (9) and have been shown to bind Gram-positive bacteria through the recognition of lipoteichoic acid (11, 12). For example, the CASRs have been implicated in the phagocytosis of Listeria monocytogenes (13), various streptococci (14, 15), and the clostridial pathogen C. perfringens (16). We therefore speculated that CASRs would be expressed by DMs and capable of internalizing C. sordellii.
Two particular CASR subtypes, SR-AI/II and MARCO, participate in macrophage phagocytosis of Gram-positive bacteria. The SR-AI, II, and III isoforms are derived from the same gene (MSRI) on chromosome 8, with alternative splicing producing the different proteins (9). Of these splice variants, only SR-AI and II (referred herein as SR-AI/II) are phagocytic, while SR-AIII is unable to bind extracellular ligands because it is retained within the endoplasmic reticulum (9). The SR-AI/II receptors have been implicated in the clearance of Gram-positive pathogens (14) and SR-AI was reported to bind to C. perfringens in transfected non-phagocytic cells (16).
A separate gene (MARCO) on chromosome 2 encodes the MARCO receptor (9). Emerging data also support a major role of the MARCO receptor in macrophage phagocytosis of Gram-positive bacteria. The transcription and cell surface expression levels of MARCO are enhanced upon bacterial binding and provide evidence for its significance in bacterial clearance (9). Studies of alveolar macrophages have identified MARCO to be a major receptor for unopsonized bacteria (15, 17).
In this light, we questioned whether human DMs utilize CASRs to bind and internalize unopsonized C. sordellii and further asked whether SR-AI/II and/or MARCO would be important receptor subtypes in this process. Alternatively, because class B SRs (CBSRs), and the macrophage mannose receptor, have also been reported to be capable of phagocytosis of Gram-positive bacteria (16, 18), we questioned whether these receptors were also active in the clearance of C. sordellii.
To address these questions we studied the expression of class A and B scavenger receptor subtypes by human DMs and assessed their functional relevance in phagocytosis assays of C. sordellii in vitro. Macrophages derived from mice genetically lacking SR-AI/II or MARCO receptors were also used to examine the relevance of these receptors to immune clearance of C. sordellii. Lastly, a mouse model of C. sordellii uterine infection was utilized to define the importance of CASR subtypes in host defense. These studies shed new light onto the potential importance of CASRs in the innate immune defense against highly-lethal clostridial infections of the female reproductive tract.
Materials and Methods
Animals
Six-8 wk old, female SR-AI/II/II deficient (SR-AI/II−/−) mice on a 129/SvJ strain background were originally provided as a generous gift of Dr. Willem J. S. de Villiers (University of Kentucky Medical Center, Lexington, KY, USA). These were bred locally at our facility. Age- and sex-matched 129/SvJ wild-type (WT) mice were purchased from Jackson Laboratory (Bar Harbor, ME). Six-8 wk old, female mice genetically deficient in MARCO (MARCO−/−) on a BALB/c background were bred as previously published (19) and utilized with the generous permission of Dr. Karl Tryggvason (Karolinska Institute, Stockholm, Sweden), who initially provided the MARCO−/− mice. The genotypes of mouse strains were confirmed by tail-snip DNA PCR analyses performed by Transnetyx (Cordova, TN). Age- and sex-matched BALB/c WT mice were purchased from Jackson Laboratory. Animals were treated according to National Institutes of Health guidelines for the use of experimental animals with the approval of the University of Michigan Committee for the Use and Care of Animals.
Bacteria
C. sordellii strain ATCC9714 was obtained from the American Type Culture Collection (Manassas, VA) and grown anaerobically in broth culture overnight at 37°C in reinforced clostridial medium. Estimates of bacterial concentrations were derived from the optical density (OD) of bacterial cultures at 600 nm (OD600) and a previously-determined standard curve of colony forming units (CFU) vs. OD600. Estimated bacterial concentrations were confirmed by serial 10-fold dilutions on solid agar composed of reinforced clostridial media containing 1.5% (w/w) agar, incubated anaerobically. For phagocytosis experiments (below), heat-killed C. sordellii were prepared by heating to 121°C for 30 min.
Reagents
RPMI 1640, penicillin/streptomycin/amphotericin B solution, TRIzol®, and acetylated low-density lipoprotein (AcLDL) were from Life Technologies-Invitrogen (Carlsbad, CA). Fetal bovine serum (FBS) and charcoal-stripped FBS were from HyClone Laboratories (Waltham, MA). Reinforced clostridial medium, mouse anti-human CD14 IgG (clone MPHIP9), and monoclonal antibody CD11b (clone D12) were from BD Biosciences (San Jose, California). Cytochalasin D, hyaluronidase, deoxyribonuclease, percoll, fucoidan, polyinosinic acid (Poly(I)), polycytidylic acid (poly(C)), dextran sulfate, chondroitin sulfate, mannan, fluorescein isothiocyanate (FITC), and trypan blue were from Sigma-Aldrich (St. Louis, MO). Magnetic MACS® CD14 microbeads (human) were from Miltenyi Biotec (Auburn, CA). Recombinant mouse soluble MARCO (sMARCO) and monoclonal anti-human a monoCD163 (clone 215927) were purchased from R&D Systems (Minneapolis, MN). F4/80 (clone 6A545) monoclonal IgG was from Santa Cruz Biotechnology (Santa Cruz, California). Mouse monoclonal CD36 IgG (clone FA6-152) was purchased from Abcam (Cambridge, MA).
Human subjects
Following appropriate informed consent, human decidual tissue was obtained from healthy adult women aged 18-44 years undergoing elective surgical termination of pregnancy during the first trimester. This study was reviewed and approved by the University of Michigan Institutional Review Board.
Decidual macrophage isolation
This procedure was adapted from a previously described protocol (20). Decidual tissue was collected from surgical abortions under aseptic conditions. The tissue was weighed and minced into small pieces, then subjected to a tissue digest with a solution containing 1 mg/ml collagenase from C. histolyticum type I-A (Sigma), 1 mg/ml hyaluronidase from bovine testes type I-S (Sigma), and 150 μg/ml deoxyribonuclease I from bovine pancreas type IV (Sigma). 10 mls of digestion solution were used per gram of tissue and the samples were placed on a shaker at 37°C for 60 min. Sample were then washed using RPMI 1640 medium containing L-glutamine which was substituted with 1% antibiotic-antimycotic (Gibco) and centrifuged at 1,500 RPM at 4°C for 10 min. This was followed with serial 280 μm, 200 μm, and 100 μm nylon mesh filtrations to eliminate any remaining particulates. The cells were washed as previously described and the filtrate was re-suspended in 25% Percoll (Sigma) in RPMI (same as above) and overlaid onto 50% Percoll, with 2 mls PBS layered above the 25% Percoll. Decidual cells were recovered from the 25%/50% interface following a 4°C centrifugation for 45 min at 1,500 RPM. These cells were washed as above, followed by a red blood cell lysis using NH4Cl/Tris-HCl. After two washes, the cells were then passed through a 30 μm falcon filter (BD Biosciences).
Using antibodies for the typical macrophage markers CD14, F4/80, and CD11b, flow cytometry was employed to verify that macrophages make up ~10% of human decidual tissue (results not shown) (8). The isolated cells underwent a positive selection step for macrophages by passing through two successive large cell columns (Miltenyi Biotec) using MACS® CD14 microbeads (Miltenyi Biotec). Flow cytometry was used to confirm the purity of the isolation, with > 94% of cells CD14+ (results not shown).
Resident peritoneal macrophage isolation
Resident peritoneal macrophages from mice were obtained via peritoneal lavage as previously described (21, 22). A red blood cell lysis was performed and cells were resuspended in RPMI 1640 containing 10% FBS and 1% penicillin/streptomycin/amphotericin. Cells were plated in 384-well tissue culture-treated plates (Costar) at 2 x 105 cells/well and incubated overnight (37°C with 5% CO2). The following day cells were washed two times with warm medium to remove nonadherent cells.
RNA extraction, cDNA synthesis
TRIzol® (Invitrogen) was added to the CD14+ decidual cells to perform an RNA extraction. After complete dissociation, chloroform (Sigma) was added at 200 μl/ml TRIzol®. Samples were centrifuged at 13,000 RPM for 15 min at 4°C, the aqueous phase was saved and an equal volume of isopropanol was added. After 20 min incubation at 22°C, samples were centrifuged as before. Pellet was resuspended in 500 μl 80% ethanol and centrifuged as before. Diethylpyrocarbonate-treated water was added to pellet and RNA concentration was determined by ratiometric light absorbance at 260/280 nm. Then 1 μg RNA was transcribed into cDNA using the Transcriptor First Strand cDNA Synthesis Kit (Roche) per the manufacturer’s instructions. Samples were run on a Mastercycler Epgradient S (Invitrogen) with the following conditions; 65°C for 10 min, 50°C for 60 min, and 85°C for 5 min.
Semiquantitative real-time PCR
Primers and probes were designed using the Roche Universal Probe Library Assay Design Center (www.roche-applied-science.com). All primers (Table) were produced by Integrated DNA Technologies and all probes were from Roche. Universal ProbeLibrary Reference Gene Assays (Roche) for for human and mouse GAPDH were used as appropriate. To prepare the assay, 2 μl cDNA aliquots were used to prepare assays according to manufacturer’s instructions and run on the LightCycler 480 (Roche) with the following conditions; 95°C 10 min (preincubation), 95°C 10 sec, 60°c 30 sec, 72°C 1 sec (amplification, 45 cycles), 95°C 10 sec, 50°C 30 sec, 70°C (melting curve), 40°C 30 sec (cooling). Analysis was performed using Roche software; all samples were referenced to the expression of the housekeeping gene GAPDH.
Table I.
Primers and probes
| Gene | Forward Primer | Reverse Primer | UPL Probe # |
|---|---|---|---|
| MSR1 | tcccactggagaaagtggtc | ctccccgatcacctttaagac | 85 |
| MARCO | tgctgggttactccaaagga | cagccagatctgcccagt | 24 |
| SCARA5 | tgggcatcttcatcttagca | cattcagccggttcacatt | 24 |
| CD36 | tggaacagaggctgacaactt | ttgattttgatagatatgggatgc | 76 |
| CD163 | tggggaaagcataactgtga | gctcagatctgctccctttg | 67 |
| COLEC12 | cagagcgtgaaaatgaatgg | tgtccagctttccaatttttg | 42 |
| Msr1 (Mus musculus) | ctttaccagcaatgacaaaagaga | atttcacggattctgaactgc | 89 |
| Marco (Mus musculus) | ggcaccaagggagacaaa | tcccttcatgcccatgtc | 97 |
Phagocytosis Assays
CD14+ cells were cultured overnight (37°C) in 384-well tissue culture-treated plates with 2 x 105 cells/well. The cells were treated with compounds of interest and incubated for 30 min at 37°C. Heat-inactivated C. sordellii were surfaced-labeled with FITC per our previously published protocol (23). FITC-labeled C. sordellii (FITCC.sordellii) was applied at a multiple of infection (MOI) of 300 bacteria:1 cell and incubated for 3 hrs at 37°C. Phagocytosis was quantified according to our published method of measuring intracellular fluorescence as a surrogate marker of bacterial ingestion by macrophages (24). The fluorescence of intracellular FITCC.sordellii was determined using a microplate fluorometer (485ex/535em, SPECTRAMax GEMINI EM; Molecular Devices, Sunnyvale, CA) according to our previously published method (24). Briefly, fluorescence data were expressed in arbitrary relative fluorescence units (RFU), which were then converted into a phagocytic index (PI). The PI represented the fluorescence of intracellular (phagocytosed) bacteria (RFUi) and was calculated by subtracting the fluorescence of extracellular bacteria (RFUex) from the total fluorescence of the well (RFUtotal). The RFUex was estimated by treating some cells with the phagocytosis inhibitor, cytochalasin D (5 μl/ml; EMD Chemicals), for 30 min prior to exposure to FITCC. sordellii (25).The RFUex from the cytochalasin-treated wells was subtracted from the RFUtotal. Therefore, the PI = RFUi = RFUtotal - RFUex (24).
Select compounds tested included the nonspecific CASR blocking agents fucoidan, poly(I), and dextran sulfate; the negative control compounds chondroitin sulfate and poly(C); the mannose receptor blocker mannan; sMARCO, and the SR-AI/II blocking agent AcLDL. CBSR-blocking antibodies used included the mouse monoclonal anti-human a monoCD163 and CD36 IgGs (26). Phagocytosis assays with mouse macrophages were similarly performed, however a MOI of 150:1 was used. In addition, for assays comparing WT cells with either MARCO−/− or SR-AI/II−/−, the results were normalized to total intracellular lactate dehydrogenase activity levels to account for potential differences in cell number or adherence to tissue culture plates (27). Lactate dehydrogenase activity was determined by a commercially available assay according to the manufacturer’s instructions (Roche).
Intrauterine infection
C. sordellii was cultured anaerobically (24 hr, 37°C) in 10 ml liquid reinforced clostridial medium (Life Technologies-Invitrogen). The bacteria were then centrifuged (2,000 RPM x 10 min) and the pellet washed three times with sterile phosphate buffered saline (PBS) and resuspended in 1 ml total volume of PBS. Dilutions were made in PBS to allow the inoculation of ~1 x 104 CFU directly into one horn of the mouse uterus according to our previously published protocol (23), in a total volume of 35 μl. The actual inoculum was determined by plating serial 10-fold dilutions of the PBS suspension onto solid RCM agar and counting individual colonies the next day. For survival experiments, the mortality from infection was monitored for 8 d following inoculation.
Bacterial clearance and dissemination studies
For studies of bacterial clearance and dissemination, mice were infected intrauterine as above with 1 x 103-104 CFU C. sordellii strain ATCC9714. The uterus and spleen were harvested 15, 24, or 48 hrs after infection and mechanically homogenized in 1 ml of sterile PBS. Then, 50 μl of homogenized sample was cultured anaerobically on RCM agar and colonies were enumerated after 24 hrs. Results were expressed as CFU per ml of homogenate.
Statistical analyses
Mean values were compared using a one-way analysis of variance (ANOVA) followed by a Bonferroni correction or a Student t-test as indicated. Differences were considered significant if P ≥ 0.05. Comparison of survival curves for mice infected with C. sordellii was performed using a Mantel-Cox log-rank test. Experiments were performed on at least three separate occasions unless otherwise specified. Unless otherwise noted, data are presented as mean values ± standard error of the mean (SEM).
Results
Fucoidan inhibits the phagocytosis of C. sordellii by human decidual macrophages
Because C. perfringens was reported to be phagocytosed predominately through CASRs in non-reproductive tract macrophages (16), we hypothesized that human DMs utilized CASRs in phagocytosing C. sordellii. To test this, DMs were pretreated with fucoidan, an antagonist of CASRs that is not selective for the unique CASR subtypes (28). A 30-min preincubation with fucoidan at 100 and 500 μg/ml dose-dependently and significantly inhibited the phagocytosis of FITCC. sordellii by 75.8 ± 4.9% and 96.0 ± 2.0% respectively (Fig. 1A). Pilot experiments demonstrated that maximal inhibition was obtained using 500 μg/ml (data not shown), so this concentration was used as a positive, comparative control in other phagocytosis assays.
Figure 1. Evidence for functional scavenger receptors in human decidual macrophages (DMs).
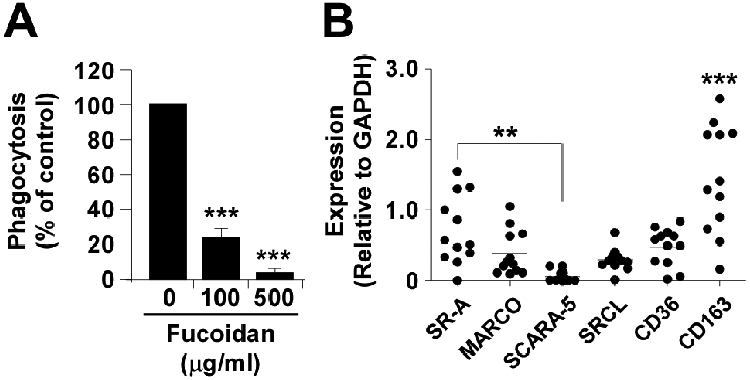
A, Human DMs were pretreated for 30 min with the scavenger receptor blocker fucoidan (in μg/ml) and phagocytosis of FITCC. sordellii quantified as detailed in Materials and Methods. Data are representative from three independent experiments performed in octuplet with similar results. Results are expressed as a percentage of relative to untreated cells. ***, P < 0.001 vs. untreated cells by ANOVA. B, Semi-quantitative real-time PCR performed to measure expression levels of scavenger receptors in human DMs, relative to GAPDH, described in Materials and Methods. Data points represent the mean result from 12 separate individuals performed in triplicate. **, P < 0.01; ***, P < 0.001 comparing CD163 against each of the other receptors (ANOVA).
Human DMs express CASR receptors
Having established a possible role for CASRs in immune recognition of C. sordellii, experiments were conducted to determine their expression levels in DMs using semi-quantitative real-time PCR. Although a functional mannose receptor has been identified on human DMs (29), to our knowledge other phagocytic receptors, including the CASRs and CBSRs, have not been well characterized. Thus, mRNA was isolated from unstimulated human DMs, and real-time PCR was performed for the CASR receptors SR-AI/II, MARCO, scavenger receptor with C-type lectin (SRCL), and SCARA-5. SRCL is an endothelial CASR that has also been identified on alveolar macrophages (30). SCARA5, was recently characterized in murine epithelial cells and was found to have similar binding properties to other CASR receptors (31). However, functional roles in microbial clearance for either SRCL or SCARA5 receptors have not been identified for human phagocytes. The expression of two CBSRs implicated in binding bacteria, CD36 and CD163, was also evaluated (32, 33). These experiments revealed expression of multiple SRs (Fig. 1B). Of the CASRs, the expression of SR-AI/II was greatest, being significantly higher than SCARA-5, which was only minimally expressed. SR-AI/II transcripts were not significantly greater than mRNA levels of MARCO. We also found a significantly higher expression of the CBSR, CD163, when compared with CD36.
Multiple pharmacological antagonists of CASRs impair phagocytosis of C. sordellii by human DMs
The above data suggested that human DMs express functional CASRs, but these experiments were limited by the use of a single, nonselective CASR antagonist, fucoidan. Thus, phagocytosis assays were performed using additional, standard, nonselective CASR blocking agents. When cells were pre-incubated with dextran sulfate (100 μg/ml), there was an 87.3 ± 6.1% decrease in phagocytic ability, while the negative control (but structurally similar) agent chondroitan sulfate (100 μg/ml) had no effect (Fig. 2A). Another classical CASR antagonist, Poly(I), suppressed phagocytosis by 48.5 ± 6.2% compared to vehicle treatment, while the negative control compound Poly(C), had no effect (Fig. 2B). The generalizability of these results for other types of macrophages was tested by conducting experiments using resident peritoneal macrophages isolated from female Wistar rats, which yielded similar results (data not shown). While these observations further implicated CASRs in the phagocytosis of C. sordellii by human DMs, they did not differentiate among the various subtypes.
Figure 2. Non-selective class A scavenger blockers inhibit phagocytosis of C. sordellii, while mannose receptor blockers have no effect.

A, human decidual macrophages (DMs) were pretreated for 30 min with the class A scavenger receptor (CASR) blocking agent dextran sulfate (DxSO4, 100 μg/ml) or the negative control agent chondroitan sulfate (ChdSO4, 100 μg/ml) and the phagocytosis of FITCC. sordellii was quantified by fluorometry after 180 min as detailed in Materials and Methods. B, human DMs were pretreated for 30 min with the CASR blocking agent Poly(I) (200 μg/ml) or the negative control Poly(C) (200 μg/ml) and the phagocytosis of FITCC. sordellii was quantified by fluorometry after 180 min as detailed in Materials and Methods. C, human DMs were pretreated with fucoidan (500 μg/ml) or the mannose receptor blocker, mannan (100 μg/ml ) for 30 min and the phagocytosis of FITCC. sordellii was quantified by fluorometry after 180 min as detailed in Materials and Methods. Data are representative from three independent experiments performed in octuplet with similar results. Results are expressed as a percentage of relative to untreated cells. ***, P < 0.001 vs. control as determined by ANOVA.
A previous study by O’Brien and Melville identified a minor role for the mannose receptor in the macrophage phagocytosis of C. perfringens (16). However, we found no role for this receptor in the phagocytosis of C. sordellii when human DMs were pre-incubated (30 min) with the antagonist mannan (100 μg/ml) (Fig. 2C). We also failed to identify a role for mannose receptors in C. sordellii phagocytosis using resident rat peritoneal macrophages (data not shown).
MARCO is the predominant phagocytic receptor for C. sordellii by human DMs
To determine which of the predominant CASRs expressed by human DMs (SR-AI/II and MARCO) are primarily responsible for internalizing C. sordellii, a previously published pharmacological approach to selectively antagonize these receptors was employed (34). Cells were preincubated (30 min) either with recombinant, mouse sMARCO or with the SR-AI/II-selective agent AcLDL (34). AcLDL had no effect on C. sordellii phagocytosis, while sMARCO inhibited phagocytosis by 79.5 ± 2.1% (Fig. 3A). These data implicated MARCO as the predominant phagocytic receptor for C. sordellii on human DMs.
Figure 3. Soluble MARCO and anti-CD36 IgG prevent the phagocytosis of C. sordellii by decidual macrophages (DMs).
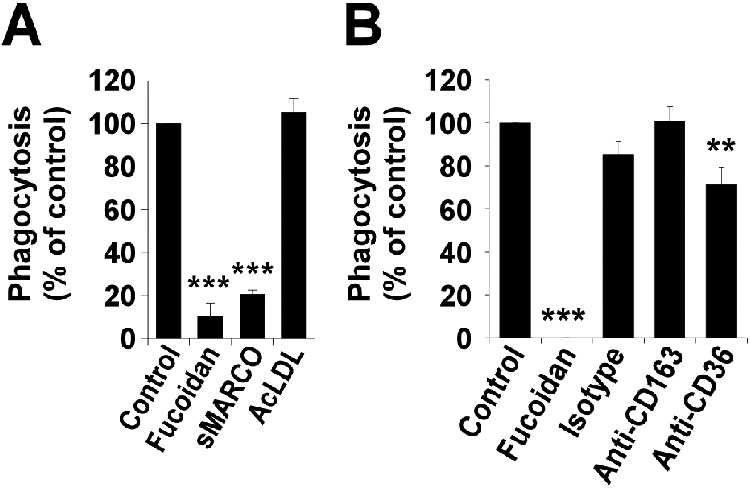
A, Human DMs were pretreated with sMARCO (100 μg/ml) or the SR-AI/II-binding agent acetylated low density lipoprotein (AcLDL; 100 μg/ml) for 30 min and the phagocytosis of FITCC. sordellii was quantified by fluorometry after 180 min as detailed in Materials and Methods. B, Human DMs were pretreated with mouse anti-human monoclonal IgG directed against CD36 or CD163 (or an isotype control) at 50 μg/ml for 30 min and the phagocytosis of FITCC. sordellii was quantified by fluorometry after 180 min as detailed in Materials and Methods. Fucoidan pretreatment for 30 min at 500 μg/ml used as positive control for each. Data are representative from three independent experiments performed in octuplet with similar results. Results are expressed as a percentage of relative to untreated cells. **, P < 0.01 and ***, P < 0.001 vs. untreated cells.
Because DMs express CBSRs (Fig. 1B) and macrophage CBSRs have been shown to bind and phagocytose bacteria (18, 32, 33), we preincubated DMs with monoclonal antibodies against human CD163 and CD36 (50 μg/ml) before measuring phagocytosis. Consistently, a small but statistically significant inhibitory effect (~30% inhibition) on phagocytosis was observed when cells were treated with anti-CD36 treatment, whereas the CD163 antibody had no effect (Fig. 3B). Thus, CD36 appears to play a minor role in phagocytosing C. sordellii.
MARCO but not SR-AI/II receptors are critical for mouse macrophage phagocytosis of C. sordellii
The above experiments were limited by their pharmacological approach. The availability of CASR-deficient mice allowed an alternative approach to test the importance of these receptors in the phagocytosis of C. sordellii. Resident peritoneal macrophages obtained from SR-AI/II−/− mice, MARCO−/− mice, or respective WT animals were challenged with FITCC. sordellii and their relative phagocytic capacity was determined. We observed that macrophages from SR-AI/II−/− mice exhibited ~3.18 ± 0.75-fold greater phagocytosis of C. sordellii compared to WT cells (P<0.05; Fig. 4A). This phagocytic ability was inhibited > 90% using sMARCO (data not shown). Real time PCR was performed to determine whether there were compensatory changes in MARCO expression in the SR-AI/II−/− macrophages correlating with their enhanced phagocytic capacity. As shown (Fig. 4B), MARCO mRNA levels were, on average, 5.3-fold higher in SR-AI/II−/− macrophages versus WT cells (P<0.05).
Figure 4. MARCO is more important than SR-AI/II in the phagocytosis of C. sordellii.
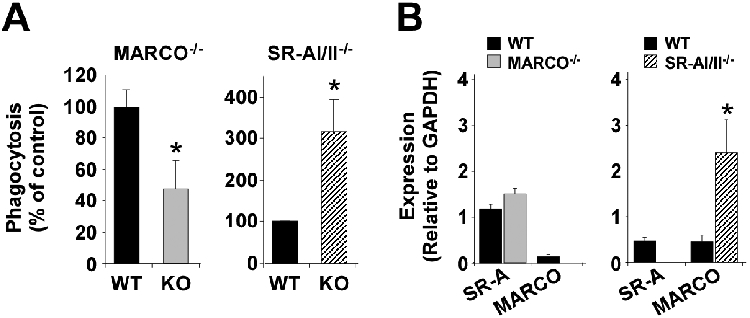
A, Left panel, peritoneal macrophages from either wild type (WT) BALB/c mice or MARCO−/− mice were allowed to phagocytose FITCC. sordellii for 3 hr. Right panel, peritoneal macrophages from either wild type (WT) 129/SvJ mice or SR-AI/II−/− mice were allowed to phagocytose FITCC. sordellii for 3 hr. The phagocytosis of FITCC. sordellii was quantified by fluorometry as detailed in Materials and Methods. KO, knockout cells. *P <0.05 vs. WT by t-test. Results are a mean ± SEM of 3-5 independent experiments performed in octuplet. B, mRNA expression of SR-AI/II or MARCO in mouse peritoneal macorphages was determined by semi-quantitative real-time PCR as detailed in Materials and Methods. Left panel, mRNA levels were compared between WT BALB/c peritoneal macrophages (black bars) or MARCO−/− cells (gray bars). Right panel, mRNA levels were compared between WT 129/SvJ peritoneal macrophages (black bars) or SR-AI/II−/− cells (striped bars). Data are normalized to GAPDH expression and represent the mean ± SEM of 8 mouse samples per assay. *P < 0.05 vs. WT cells by t-test.
These data suggested that in the absence of the SR-AI/II receptors, MARCO was able to internalize C. sordellii. To test this, we compared phagocytosis of unopsonized FITCC. sordellii using WT and MARCO−/− peritoneal macrophages. As shown (Fig. 4A), phagocytosis was significantly lower in macrophages lacking MARCO. Real-time PCR (Fig. 4B) did not detect any significant compensatory increase in mRNA expression of SRA-I/II.
MARCO deficient mice have enhanced susceptibility to C. sordellii infection
Because the above in vitro experiments with mouse peritoneal macrophages and human DMs suggested that MARCO is an important immunosurveillance receptor used by female reproductive tract macrophages to defend the host against C. sordellii, we tested its importance, in this context, in vivo. Wild type or MARCO−/− mice were inoculated intrauterine with the virulent C. sordellii strain ATCC9714. Using an inoculum of 1 x 104 CFU per mouse, only 15% of infected WT BALB/c mice died as a result of infection (Fig. 5A). In contrast, MARCO−/− mice were significantly more susceptible to death from infection, with a 3-fold higher mortality of 45% by 7 d post infection (Fig. 5A, n = 20 mice per group, P = 0.04 by Mantel-Cox log-rank test). In contrast, SR-AI/II−/− mice were not more susceptible to death from intrauterine C. sordellii infection compared to WT 129/SvJ mice (Fig. 5C).
Figure 5. MARCO is important to host defense against intrauterine C. sordellii infection.
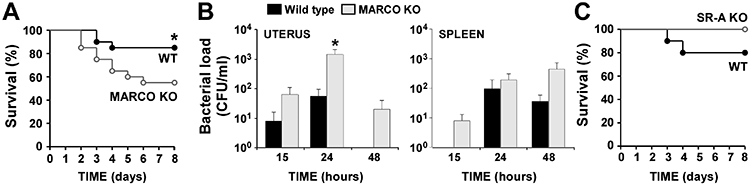
A, Wild type (WT) or MARCO knockout (KO) mice (n=20 per group) were infected by intrauterine injection with C. sordellii ATCC9714 (1x104 CFU per mouse) and survival was recorded. *P < 0.05 vs. WT survival. B, Clearance of C. sordellii from the uterus and dissemination to the spleen in WT or MARCO KO mice. Mice were inoculated intrauterine with 1 x 104 CFU of C. sordellii ATCC9714. After infection, uterine and splenic tissues were homogenized at the time points shown, and colony-forming units (CFU) determined as described in the Materials and Methods. Data are the mean ± SEM from 5 mice per strain at each time point, except at 24 hrs (n = 10 mice per strain). *P < 0.05 vs. WT by Student t-test. C, Wild type (WT) or SR-A knockout (KO) mice (n=10 per group) were infected by intrauterine injection with C. sordellii ATCC9714 (1x104 CFU per mouse) and survival was recorded. Differences were not significant.
MARCO is important for clearance of bacteria from the infected uterus
We questioned whether the increased mortality observed in MARCO−/− mice correlated with an impaired capacity to eliminate bacteria from the infected uterus or to prevent dissemination to distal organs such as the spleen. To address this question, mice were infected intrauterine with C. sordellii and the bacterial loads of the uterus and spleen were determined by anaerobic culture of tissue homogenates harvested at 15 hr, 24 hr, and 48 hr post infection. As illustrated (Fig. 5B), the numbers of bacteria recovered from uterine tissues of MARCO−/− mice were ~1 log higher than recovered from WT BALB/c mice at each of the three time points, although this only reached statistical significance at 24 hrs post infection. Notably, the WT mice cleared the local infection by 48 hrs.
Bacteria disseminated to the spleen in both WT and MARCO−/− mice (Fig. 5B). While the differences were not statistically significant, dissemination occurred sooner in the MARCO−/− mice (see the 15 hr time point). In addition, the splenic bacterial load appeared to peak by 24 hrs in WT mice but was continuing to rise by 48 hrs in the MARCO−/− mice. We have not detected differences in local clearance of C. sordellii from the uterus or dissemination to the spleen between WT or SR-AI/II−/− mice (data not shown).
Discussion
These studies newly define a potentially important role for MARCO receptors in the innate host defense against invasive C. sordellii infections of the female reproductive tract. Infections caused by toxigenic clostridia are emerging as important challenges to human health. While the problem of clostridial infections is growing, there has been little research into the mechanisms detailing how the innate immune system recognizes and attempts to eliminate these potential pathogens. The increase in published reports of highly lethal C. sordellii gynecological infections in women of childbearing age (1, 3, 6) prompted further study of the pathogenesis of C. sordellii infections. To our knowledge, these studies show for the first time the receptors through which human macrophages internalize these toxigenic bacteria.
Macrophages are key sentinels of innate immunity that respond to microbial invaders by elaborating immunoregulatory mediators (cytokines, chemokines, and lipids), phagocytosing and killing potential pathogens, and presenting antigens to cells of the adaptive immune system. Recently, the toll-like receptors (TLRs) 2 and 6 were found to participate in the immune response of macrophages to C. sordellii, through the activation of the transcription factor NFκB (35). However, these studies did not address how macrophages phagocytose this bacterium.
Previous investigations with the related pathogen, C. perfringens, revealed that CASRs, and to a lesser extent the mannose receptor, were involved in the macrophage phagocytosis of this pathogen (16). Phagocytosis of C. perfringens was impaired in those studies using the non-selective CASR blocking agent fucoidan (16). However, that study did not establish the relative roles played by individual CASRs in C. perfringens uptake. Although the authors used Chinese hamster ovarian cells transfected with the mouse SR-AI receptor to demonstrate binding of C. perfringens to SR-AI, the capacity of SR-AI to mediate the internalization of this bacterium was not reported (16).
Based largely on the findings of these important studies of C. perfringens, we questioned whether CASRs would also be important for the phagocytosis of C. sordellii by human macrophages. Similar to the approach of O’Brien and Melville (16), we incubated DMs with fucoidan and noted that this broad inhibitor of CASR phagocytosis was a potent blocker of C. sordellii internalization. We then used mRNA expression as a surrogate marker for phagocytic receptor expression by DMs. Using real-time PCR, we identified transcription of the MSR1 and the MARCO genes, encoding SR-AI/II and MARCO, respectively. These studies newly documented the expression of SRCL on DMs, a receptor expressed primarily on non-hematopoeitic cells (e.g., endothelial cells) (30, 36). We also investigated whether SCARA-5 was expressed by human DMs, as this receptor has not been well characterized in human cells and tissues (31). However, we did not observe significant expression of this receptor.
To confirm the results obtained with fucoidan, we utilized pairs of non-selective CASR blocking agents and structurally matched, negative control compounds that do not bind to CASRs (e.g., Poly(I)-Poly(C) and dextran sulfate-chondroitan sulfate). Indeed, phagocytosis was inhibited by both dextran sulfate and poly(I) but not by the negative control agents chondroitan sulfate or poly(C).
A study by Laskarin, G. et al. suggested the mannose receptor is a functional receptor on decidual mononuclear cells (29). The exact role of the mannose receptor in phagocytosis is uncertain with conflicting biochemical and genetic studies (37). Due to its decreased expression during inflammation (38) it has been suggested the mannose receptor may be more important for tissue homeostasis than for host defense (37). However, taking into consideration this information and the previously mentioned studies by O’Brien and Melville, this receptor was also tested to determine its importance in the binding and phagocytosis of C. sordellii. We found no major role for the mannose receptor in our experimental design.
Though controversial, data suggest that TLR2 receptor ligation is involved in the ingestion of Gram positive bacteria by phagocytic cells (39). It was also recently reported that TLR2 plays a role in bacterial phagocytosis by mouse trophoblast giant cells (40). Using monoclonal blocking antibodies against the TLR2 receptor on human DMs and rat peritoneal macrophages we have not observed an inhibitory effect on the phagocytosis of C. sordellii compared to isotype control immunoglobulins (data not shown). These data suggest that TLR2 does not play a significant role in the internalization of C. sordellii. However, alternative approaches, such as the use of macrophages from TLR2-deficient mice, or the use of genetic silencing in human cells, could provide additional evidence in this regard.
Given the potentially critical role for CASRs in the macrophage phagocytosis of C. sordellii we sought to decipher which specific subtypes were most important. There is precedent to suggest that different pathogens may bind unique CASRs with varying efficiency (41, 42). We initially incubated human DMs with either AcLDL, a specific antagonist of SR-AI/II receptors (42), or sMARCO, a specific inhibitor of MARCO-dependent phagocytosis (42, 43). These studies found that phagocytosis could be potently blocked by sMARCO but not by AcLDL, suggesting that MARCO is the more critical receptor for the phagocytosis of C. sordellii.
These pharmacological results were confirmed using receptor deficient mice. We were surprised to observe a significantly greater phagocytic capacity in SR-AI/II−/− macrophages compared with WT cells (Fig. 4) and speculated that this might be due to enhanced MARCO expression, which has been observed in SR-AI/II−/− mice, though on a different genetic background (44). The fact that sMARCO inhibited the phagocytic capacity of the SR-AI/II−/− macrophages suggested this might be the case (data not shown). Indeed, mRNA transcript levels for MARCO were significantly greater in SR-AI/II−/− macrophages compared with WT macrophages (Fig. 4), providing more evidence that MARCO expression is upregulated in mouse cells lacking SR-AI/II. These data were limited by the fact that mRNA transcripts were quantified, not surface expressed protein levels. However, a previous study of SR-AI/II−/− peritoneal macrophages (on a BALB/c background) showed higher surface expression of MARCO using an anti-MARCO antibody (44).
Notably, MARCO−/− peritoneal macrophages demonstrated an impaired capacity to phagocytose C. sordellii. These data supported our pharmacological results in the human cell. What is more, we observed, for the first time, a functional role for MARCO in immune defense against clostridia in vivo. Importantly, mice lacking functional MARCO receptors demonstrated increased susceptibility to lethal infection by C. sordellii and impaired clearance of these bacteria from the uterus (Fig. 5A and B). There was also a non-statistically significant trend toward greater dissemination to the spleen in the MARCO−/− mice. As expected, we did not see enhanced susceptibility to infection in SR-AI/II null mice (Fig. 5C). We have also not observed differences in bacterial clearance from the uterus comparing WT and SR-AI/II null mice (data not shown).
While our mouse uterine infection model data provide novel and correlative support for our in vitro results, suggesting that MARCO is an important component of the innate defense against C. sordellii infections, they do not prove that DMs per se are involved in host defense in vivo. The complex cellular defense mechanisms involved in innate immunity against C. sordellii remain to be fully defined. We previously reported that human β defensin antimicrobial peptides were rapidly bactericidal against C. sordellii (23), suggesting that epithelial cells might be important in controlling C. sordellii infection within the uterus. Though not presented here, we have examined whether C. sordellii can invade (or get phagocytosed by) uterine epithelial cells. Histological examination of mouse uterine tissues 48 hrs following intrauterine inoculation with C. sordellii did not reveal bacteria within uterine epithelial cells in situ, and we have not observed significant phagocytosis of C. sordellii by the human uterine epithelial RL95-2 cell line in vitro (data not shown). What is more, using real time, reverse transcriptase PCR we have found that RL95-2 cells do not express MARCO or SR-AI/II, though they do express SRCL, suggesting that SRCL is not an efficient phagocytic receptor for C. sordellii on these cells (data not shown).
Our data are in accord with previously published studies of the role of MARCO in immune defense against bacterial pathogens of the lung (45). In a murine model of pneumonia caused by the Gram-positive pathogen Streptococcus pneumoniae, MARCO-deficient mice displayed an impaired ability to clear bacteria from the lungs and significantly diminished survival (45). Our findings imply that genetic differences in MARCO expression or function might influence the risk of C. sordellii or other clostridial infections.
There are several limitations in our work. The bacteria used in our studies of phagocytosis were heat-killed, FITC-labeled C. sordellii, which would not be found in nature. Thus, these results may be different using live bacteria. In addition, the bacteria used were in a vegetative (non-sporulating) form, which models active invasion with replicating bacteria. However, future studies of the phagocytosis of spore forms will be of interest as well. Our study was also limited by the fact that we did not differentiate between the mRNA expression of SR-AI and SR-AII. These receptors are transcribed from the same gene and differ as a result of alternative mRNA splicing (46). An additional caveat is that the rodent cells used for these studies were peritoneal macrophages, which may differ significantly from reproductive tract cells. Thus, direct comparisons with human DMs are difficult.
These studies newly described expression of SRCL by human DMs. This receptor, to our knowledge, has not been shown to be a functional phagocytic receptor on macrophages. We did not examine its role in the phagocytosis of C. sordellii and this will be a subject of future research. We also did not identify a major role for CBSRs in the phagocytosis of C. sordellii using monoclonal antibodies to block CD36 and CD163. This approach has limitations and future studies using genetically-deficient, mouse-derived macrophages or gene-silencing approaches would be necessary to confirm our results. It is notable that human DMs express relatively high levels of mRNA for the CBSR CD163 (transcripts for CD163 were the highest of all scavenger receptors we examined by semi-quantitative real time PCR (Fig. 1B)). The CD163 receptor is involved primarily in the clearance of hemoglobin-haptoglobin complexes (47). It has been implicated in anti-inflammatory responses and is a surface marker of alternative (M2) macrophage activation (47). Thus, our finding of CD163 expression by human DMs provides further support to other studies demonstrating that DMs exhibit features of alternative (M2) activation (48, 49). The implications of this for bacterial infections in the postpartum or post-abortion period are unclear.
Studies of how DMs interact with pathogenic bacteria contribute to our understanding of these complex phagocytes. Mounting evidence suggests that DMs are important in host defense against bacterial infections during pregnancy, such as the Gram positive pathogen Listeria monocytogenes (50). While the clearance of bacteria may prevent overt intrauterine infection, the microbial activation of DMs can also drive preterm birth by stimulating the production of cytokines and prostaglandins that cause uterine contraction and cervical dilation (51). In addition, DMs suppress immune responses at the fetal/maternal interface to induce and maintain maternal tolerance to the developing fetus (52, 53). Thus, unlike other macrophages, DMs defend against microbial invasion while maintaining an immunosuppressive phenotype in the context of maternal-fetal development. Future studies are needed to determine whether the immunosuppressive (M2) phenotype of DMs contributes to the enhanced susceptibility of pregnant women to intrauterine infection.
In summary, we newly characterize scavenger receptors expressed by human DMs and their relative importance in the recognition and internalization of the toxigenic pathogen C. sordellii. Using a combination of in vitro and in vivo experiments, we demonstrate a role for MARCO in reproductive tract innate immunity. These results may prove to be important not only for understanding the pathogenesis of invasive clostridial infections, but also for developing better preventive and therapeutic measures against a range of both sexually and non-sexually transmitted infections.
Acknowledgments
This research was funded by National Institutes of Health Grants HD057176 (D.M.A.), ES11008 (L.K.), and HL056309 (J.L.C.). Funding for non-animal portions of this research was provided by a Doris Duke Charitable Fund Clinical Scientist Development Award (D.M.A.).
Abbreviations
- AcLDL
acetylated low density lipoprotein
- CASR
class A scavenger receptor
- CBSR
class B scavenger receptor
- DM
decidual macrophage
- FITC
fluoroscein isothiocyanate
- GAPDH
glyceraldehydes-3-phosphate dehydrogenase
- KO
knockout
- poly(I)
polyinosinic acid
- poly(C)
polycytidylic acid
- SR-AI/II
scavenger receptor AI and AII
- MARCO
macrophage receptor with collagenous structure
- SRCL
scavenger receptor with C-type lectin
- SCARA-5
scavenger receptor A5
- TLR
toll-like receptor
- WT
wild type
References
- 1.Aldape MJ, Bryant AE, and Stevens DL. 2006. Clostridium sordellii infection: epidemiology, clinical findings, and current perspectives on diagnosis and treatment. Clin Infect Dis 43:1436–1446. [DOI] [PubMed] [Google Scholar]
- 2.Aronoff DM, and Ballard JD. 2009. Clostridium sordellii toxic shock syndrome. The Lancet infectious diseases 9:725–726. [DOI] [PubMed] [Google Scholar]
- 3.Ho CS, Bhatnagar J, Cohen AL, Hacker JK, Zane SB, Reagan S, Fischer M, Shieh WJ, Guarner J, Ahmad S, Zaki SR, and McDonald LC. 2009. Undiagnosed cases of fatal Clostridium-associated toxic shock in Californian women of childbearing age. Am J Obstet Gynecol 201:459 e451–457. [DOI] [PubMed] [Google Scholar]
- 4.Fischer M, Bhatnagar J, Guarner J, Reagan S, Hacker JK, Van Meter SH, Poukens V, Whiteman DB, Iton A, Cheung M, Dassey DE, Shieh WJ, and Zaki SR. 2005. Fatal toxic shock syndrome associated with Clostridium sordellii after medical abortion. N Engl J Med 353:2352–2360. [DOI] [PubMed] [Google Scholar]
- 5.Sinave C, Le Templier G, Blouin D, Leveille F, and Deland E. 2002. Toxic shock syndrome due to Clostridium sordellii: a dramatic postpartum and postabortion disease. Clin Infect Dis 35:1441–1443. [DOI] [PubMed] [Google Scholar]
- 6.Hao Y, Senn T, J SO, Young VB, Thiele T, Srinivas G, Huang SK, and Aronoff DM. 2010. Lethal toxin is a critical determinant of rapid mortality in rodent models of Clostridium sordellii endometritis. Anaerobe 16:155–160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.O'Brien DK, and Melville SB. 2004. Effects of Clostridium perfringens alpha-toxin (PLC) and perfringolysin O (PFO) on cytotoxicity to macrophages, on escape from the phagosomes of macrophages, and on persistence of C. perfringens in host tissues. Infect Immun 72:5204–5215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Singh U, Nicholson G, Urban BC, Sargent IL, Kishore U, and Bernal AL. 2005. Immunological properties of human decidual macrophages--a possible role in intrauterine immunity. Reproduction 129:631–637. [DOI] [PubMed] [Google Scholar]
- 9.Murphy JE, Tedbury PR, Homer-Vanniasinkam S, Walker JH, and Ponnambalam S. 2005. Biochemistry and cell biology of mammalian scavenger receptors. Atherosclerosis 182:1–15. [DOI] [PubMed] [Google Scholar]
- 10.Peiser L, and Gordon S. 2001. The function of scavenger receptors expressed by macrophages and their role in the regulation of inflammation. Microbes Infect 3:149–159. [DOI] [PubMed] [Google Scholar]
- 11.Dunne DW, Resnick D, Greenberg J, Krieger M, and Joiner KA. 1994. The type I macrophage scavenger receptor binds to gram-positive bacteria and recognizes lipoteichoic acid. Proc Natl Acad Sci U S A 91:1863–1867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Greenberg JW, Fischer W, and Joiner KA. 1996. Influence of lipoteichoic acid structure on recognition by the macrophage scavenger receptor. Infect Immun 64:3318–3325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Suzuki H, Kurihara Y, Takeya M, Kamada N, Kataoka M, Jishage K, Ueda O, Sakaguchi H, Higashi T, Suzuki T, Takashima Y, Kawabe Y, Cynshi O, Wada Y, Honda M, Kurihara H, Aburatani H, Doi T, Matsumoto A, Azuma S, Noda T, Toyoda Y, Itakura H, Yazaki Y, Kodama T, and et al. 1997. A role for macrophage scavenger receptors in atherosclerosis and susceptibility to infection. Nature 386:292–296. [DOI] [PubMed] [Google Scholar]
- 14.Areschoug T, Waldemarsson J, and Gordon S. 2008. Evasion of macrophage scavenger receptor A-mediated recognition by pathogenic streptococci. Eur J Immunol 38:3068–3079. [DOI] [PubMed] [Google Scholar]
- 15.Arredouani MS, Palecanda A, Koziel H, Huang YC, Imrich A, Sulahian TH, Ning YY, Yang Z, Pikkarainen T, Sankala M, Vargas SO, Takeya M, Tryggvason K, and Kobzik L. 2005. MARCO is the major binding receptor for unopsonized particles and bacteria on human alveolar macrophages. J Immunol 175:6058–6064. [DOI] [PubMed] [Google Scholar]
- 16.O'Brien DK, and Melville SB. 2003. Multiple effects on Clostridium perfringens binding, uptake and trafficking to lysosomes by inhibitors of macrophage phagocytosis receptors. Microbiology 149:1377–1386. [DOI] [PubMed] [Google Scholar]
- 17.Palecanda A, Paulauskis J, Al-Mutairi E, Imrich A, Qin G, Suzuki H, Kodama T, Tryggvason K, Koziel H, and Kobzik L. 1999. Role of the scavenger receptor MARCO in alveolar macrophage binding of unopsonized environmental particles. J Exp Med 189:1497–1506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Stuart LM, Deng J, Silver JM, Takahashi K, Tseng AA, Hennessy EJ, Ezekowitz RA, and Moore KJ. 2005. Response to Staphylococcus aureus requires CD36-mediated phagocytosis triggered by the COOH-terminal cytoplasmic domain. J Cell Biol 170:477–485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Arredouani MS, Franco F, Imrich A, Fedulov A, Lu X, Perkins D, Soininen R, Tryggvason K, Shapiro SD, and Kobzik L. 2007. Scavenger Receptors SR-AI/II and MARCO limit pulmonary dendritic cell migration and allergic airway inflammation. J Immunol 178:5912–5920. [DOI] [PubMed] [Google Scholar]
- 20.Vince GS, Starkey PM, Jackson MC, Sargent IL, and Redman CW. 1990. Flow cytometric characterisation of cell populations in human pregnancy decidua and isolation of decidual macrophages. J Immunol Methods 132:181–189. [DOI] [PubMed] [Google Scholar]
- 21.Bailie MB, Standiford TJ, Laichalk LL, Coffey MJ, Strieter R, and Peters-Golden M. 1996. Leukotriene-deficient mice manifest enhanced lethality from Klebsiella pneumonia in association with decreased alveolar macrophage phagocytic and bactericidal activities. J Immunol 157:5221–5224. [PubMed] [Google Scholar]
- 22.Peters-Golden M, McNish RW, Hyzy R, Shelly C, and Toews GB. 1990. Alterations in the pattern of arachidonate metabolism accompany rat macrophage differentiation in the lung. J Immunol 144:263–270. [PubMed] [Google Scholar]
- 23.Aronoff DM, Hao Y, Chung J, Coleman N, Lewis C, Peres CM, Serezani CH, Chen GH, Flamand N, Brock TG, and Peters-Golden M. 2008. Misoprostol impairs female reproductive tract innate immunity against Clostridium sordellii. J Immunol 180:8222–8230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Aronoff DM, Canetti C, and Peters-Golden M. 2004. Prostaglandin E2 inhibits alveolar macrophage phagocytosis through an E-prostanoid 2 receptor-mediated increase in intracellular cyclic AMP. J Immunol 173:559–565. [DOI] [PubMed] [Google Scholar]
- 25.Thiele L, Rothen-Rutishauser B, Jilek S, Wunderli-Allenspach H, Merkle HP, and Walter E. 2001. Evaluation of particle uptake in human blood monocyte-derived cells in vitro. Does phagocytosis activity of dendritic cells measure up with macrophages? J Control Release 76:59–71. [DOI] [PubMed] [Google Scholar]
- 26.Mwaikambo BR, Sennlaub F, Ong H, Chemtob S, and Hardy P. 2006. Activation of CD36 inhibits and induces regression of inflammatory corneal neovascularization. Invest Ophthalmol Vis Sci 47:4356–4364. [DOI] [PubMed] [Google Scholar]
- 27.Ballinger MN, Paine R 3rd, Serezani CH, Aronoff DM, Choi ES, Standiford TJ, Toews GB, and Moore BB. 2006. Role of granulocyte macrophage colony-stimulating factor during gram-negative lung infection with Pseudomonas aeruginosa. Am J Respir Cell Mol Biol 34:766–774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bermudez LE, Parker A, and Goodman JR. 1997. Growth within macrophages increases the efficiency of Mycobacterium avium in invading other macrophages by a complement receptor-independent pathway. Infect Immun 65:1916–1925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Laskarin G, Cupurdija K, Tokmadzic VS, Dorcic D, Dupor J, Juretic K, Strbo N, Crncic TB, Marchezi F, Allavena P, Mantovani A, Randic L, and Rukavina D. 2005. The presence of functional mannose receptor on macrophages at the maternal-fetal interface. Hum Reprod 20:1057–1066. [DOI] [PubMed] [Google Scholar]
- 30.Selman L, Skjodt K, Nielsen O, Floridon C, Holmskov U, and Hansen S. 2008. Expression and tissue localization of collectin placenta 1 (CL-P1, SRCL) in human tissues. Mol Immunol 45:3278–3288. [DOI] [PubMed] [Google Scholar]
- 31.Jiang Y, Oliver P, Davies KE, and Platt N. 2006. Identification and characterization of murine SCARA5, a novel class A scavenger receptor that is expressed by populations of epithelial cells. J Biol Chem 281:11834–11845. [DOI] [PubMed] [Google Scholar]
- 32.Baranova IN, Kurlander R, Bocharov AV, Vishnyakova TG, Chen Z, Remaley AT, Csako G, Patterson AP, and Eggerman TL. 2008. Role of human CD36 in bacterial recognition, phagocytosis, and pathogen-induced JNK-mediated signaling. J Immunol 181:7147–7156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Fabriek BO, van Bruggen R, Deng DM, Ligtenberg AJ, Nazmi K, Schornagel K, Vloet RP, Dijkstra CD, and van den Berg TK. 2009. The macrophage scavenger receptor CD163 functions as an innate immune sensor for bacteria. Blood 113:887–892. [DOI] [PubMed] [Google Scholar]
- 34.Pluddemann A, Mukhopadhyay S, Sankala M, Savino S, Pizza M, Rappuoli R, Tryggvason K, and Gordon S. 2009. SR-A, MARCO and TLRs Differentially Recognise Selected Surface Proteins from Neisseria meningitidis: an Example of Fine Specificity in Microbial Ligand Recognition by Innate Immune Receptors. Journal of Innate Immunity 1:153–163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Aldape MJ, Bryant AE, Katahira EJ, Hajjar AM, Finegold SM, Ma Y, and Stevens DL. 2010. Innate immune recognition of, and response to, Clostridium sordellii. Anaerobe 16:125–130. [DOI] [PubMed] [Google Scholar]
- 36.Areschoug T, and Gordon S. 2009. Scavenger receptors: role in innate immunity and microbial pathogenesis. Cellular microbiology 11:1160–1169. [DOI] [PubMed] [Google Scholar]
- 37.Ernst JD, and Stendahl O, eds. 2006. Phagocytosis of Bacteria and Bacterial Pathogenicity. Cambridge University Press, New York, Cambridge. [Google Scholar]
- 38.Chroneos Z, and Shepherd VL. 1995. Differential regulation of the mannose and SP-A receptors on macrophages. Am J Physiol 269:L721–726. [DOI] [PubMed] [Google Scholar]
- 39.Blander JM, and Medzhitov R. 2004. Regulation of phagosome maturation by signals from toll-like receptors. Science 304:1014–1018. [DOI] [PubMed] [Google Scholar]
- 40.Watanabe K, Shin EK, Hashino M, Tachibana M, and Watarai M. Toll-like receptor 2 and class B scavenger receptor type I are required for bacterial uptake by trophoblast giant cells. Mol Immunol. [DOI] [PubMed] [Google Scholar]
- 41.Pluddemann A, Mukhopadhyay S, and Gordon S. 2006. The interaction of macrophage receptors with bacterial ligands. Expert reviews in molecular medicine 8:1–25. [DOI] [PubMed] [Google Scholar]
- 42.Pluddemann A, Mukhopadhyay S, Sankala M, Savino S, Pizza M, Rappuoli R, Tryggvason K, and Gordon S. 2009. SR-A, MARCO and TLRs Differentially Recognise Selected Surface Proteins from Neisseria meningitidis: an Example of Fine Specificity in Microbial Ligand Recognition by Innate Immune Receptors. Journal of Innate Immunity 1:153–163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Sankala M, Brannstrom A, Schulthess T, Bergmann U, Morgunova E, Engel J, Tryggvason K, and Pikkarainen T. 2002. Characterization of recombinant soluble macrophage scavenger receptor MARCO. The Journal of biological chemistry 277:33378–33385. [DOI] [PubMed] [Google Scholar]
- 44.Jozefowski S, Arredouani M, Sulahian T, and Kobzik L. 2005. Disparate regulation and function of the class A scavenger receptors SR-AI/II and MARCO. J Immunol 175:8032–8041. [DOI] [PubMed] [Google Scholar]
- 45.Arredouani M, Yang Z, Ning Y, Qin G, Soininen R, Tryggvason K, and Kobzik L. 2004. The scavenger receptor MARCO is required for lung defense against pneumococcal pneumonia and inhaled particles. J Exp Med 200:267–272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Matsumoto A, Naito M, Itakura H, Ikemoto S, Asaoka H, Hayakawa I, Kanamori H, Aburatani H, Takaku F, Suzuki H, and et al. 1990. Human macrophage scavenger receptors: primary structure, expression, and localization in atherosclerotic lesions. Proc Natl Acad Sci U S A 87:9133–9137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Moestrup SK, and Moller HJ. 2004. CD163: a regulated hemoglobin scavenger receptor with a role in the anti-inflammatory response. Ann Med 36:347–354. [DOI] [PubMed] [Google Scholar]
- 48.Cupurdija K, Azzola D, Hainz U, Gratchev A, Heitger A, Takikawa O, Goerdt S, Wintersteiger R, Dohr G, and Sedlmayr P. 2004. Macrophages of human first trimester decidua express markers associated to alternative activation. Am J Reprod Immunol 51:117–122. [DOI] [PubMed] [Google Scholar]
- 49.Gustafsson C, Mjosberg J, Matussek A, Geffers R, Matthiesen L, Berg G, Sharma S, Buer J, and Ernerudh J. 2008. Gene expression profiling of human decidual macrophages: evidence for immunosuppressive phenotype. PLoS One 3:e2078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Qiu X, Zhu L, and Pollard JW. 2009. Colony-stimulating factor-1-dependent macrophage functions regulate the maternal decidua immune responses against Listeria monocytogenes infections during early gestation in mice. Infect Immun 77:85–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Goldenberg RL, Hauth JC, and Andrews WW. 2000. Intrauterine infection and preterm delivery. N Engl J Med 342:1500–1507. [DOI] [PubMed] [Google Scholar]
- 52.Hunt JS 1990. Current topic: the role of macrophages in the uterine response to pregnancy. Placenta 11:467–475. [DOI] [PubMed] [Google Scholar]
- 53.Nagamatsu T, and Schust DJ. The immunomodulatory roles of macrophages at the maternal-fetal interface. Reprod Sci 17:209–218. [DOI] [PubMed] [Google Scholar]