

Abstract
The escalating rate at which coral communities are declining globally requires urgent intervention and new approaches to reef management to reduce and halt further coral loss. For reef systems with limited natural larval supply, the introduction of large numbers of competent coral larvae directly to natural reef substrata provides a potentially useful approach to replenish adult coral populations. While few experiments have tested this approach, only one experiment has demonstrated its long-term success to date. Given the differences in life-history traits among corals, and different sensitivities of larvae to abiotic and biotic factors, coupled with the dynamic nature of post-settlement survivorship and recruitment processes, trials of the larval enhancement technique with larvae of different coral species are needed to test the broader applicability and viability of this approach. Accordingly, in this paper we examine the applicability of the larval enhancement technique to restore a population of Acropora loripes in the Bolinao-Anda Reef Complex, Pangasinan, northwestern Philippines. Larvae were cultured ex situ following spawning of collected A. loripes colonies in June 2014. Competent larvae were transported to degraded reef areas and approximately 300,000 larvae were introduced in each of three 6 × 4 m plots directly on the reef. Fine mesh enclosures retained the larvae inside each treatment plot for five days. Three adjacent 6 × 4 m plots that served as controls were also covered with mesh enclosures, but no larvae were introduced. Each plot contained ten 10 × 10 cm conditioned settlement tiles cut from dead tabulate Acropora that were used to quantify initial larval settlement. After allowing larval settlement for five days, mean settlement on tiles from the larval enhancement plots that were monitored under stereomicroscopes was significantly higher (27.8 ± 6.7 spat per tile) than in control plots, in which not a single recruit was recorded. Post-settlement survivorship and growth of spat and coral recruits on tiles and reef substrata inside the experimental plots were monitored periodically for 35 months. After 35 months, the mean size of each of the remaining 47 A. loripes coral colonies surviving on the reef substrata was 438.1 ± 5.4 cm3, with a mean diameter of 7.9 ± 0.6 cm. The average production cost for each of the surviving A. loripes colonies at 35 months was USD 35.20. These colonies are expected to spawn and contribute to the natural larval pool when they become reproductively mature, thereby enhancing natural coral recovery in the area. This study demonstrates that mass coral larval enhancement can be successfully used for restoring populations of coral species with different life-history traits, and the techniques can rapidly increase larval recruitment rates on degraded reef areas, hence catalysing the regeneration of declining coral populations.
Introduction
The increasing rate at which coral communities are declining globally requires urgent intervention and new approaches to reef management to reduce and halt the loss, and increase coral cover and diversity on degraded reefs [1, 2]. Therefore, active management interventions and novel restoration approaches are needed to help in reef recovery at local, regional and global scales; recovery that is currently unlikely to occur based on natural recruitment processes alone. For these reasons, management and restoration tools are increasingly considered as essential to mitigate coral reef degradation caused by anthropogenic disturbances [3–7].
In the initial stages of coral restoration research, direct coral transplantation gained popularity as one approach to rapidly increase coral cover by re-attaching coral fragments or whole colonies to degraded reefs [5, 8, 9]. As an alternative, an intermediate step of rearing coral nubbins in different types of nurseries increased the number of transplant materials from a few source colonies [10–12]. While this grow-out nursery phase increases the size and robustness of coral transplants, which may lead to higher post-transplantation survival rates [13–15], it also increases production costs from hatchery and nursery facility construction, grow-out costs, and outplanting costs [5, 16].
More recently, sexual larval propagation has become more widely used in restoration studies. This approach aims to increase recruitment rates, coral cover [17], and genetic diversity that may improve coral adaptive and evolutionary potential, and increase resilience in depleted coral populations [18–20]. Using this approach, millions of sexually derived coral larvae can be sourced from sexually mature coral colonies from ex situ spawning in a controlled hatchery facility [21, 22], or in situ by using spawn collectors placed on top of individual corals [23], or from natural coral spawn slicks [24–26]. Typically, competent larvae are then settled on artificial substrata and kept in land- or ocean-based nurseries before they are outplanted on the reef [5]. For example, Villanueva et al. [21] cultured Acropora valida larvae that were settled on coral rubble. After a few months of hatchery rearing, the juvenile corals on substrata were directly attached to reef sites using adhesive. The same approach was employed by Baria et al. [27] where larvae of A. granulosa were settled on artificial substrata in the hatchery and subsequently transplanted to the reef. The cost of each juvenile in the nursery phase was 2.79 USD but increased to 20.01 USD each after transplantation because of additional outplanting costs and subsequent coral mortality [27].
To reduce these labor-intensive and costly grow-out and outplanting phases, coral larvae can be immediately outplanted shortly after the completion of metamorphosis and onset of skeleton formation [28]. For example, Acropora palmata settlers outplanted to the reef at the age of two weeks were 7 times more likely to survive and were 25 fold less costly to produce than conspecifics kept within a land-based nursery for 2.5 years [28, 29]. In addition, new tetrapod-shaped substrates have been developed that can be “seeded’ onto the reef in much less time and 5–18 fold lower costs compared with traditional outplanting techniques because their geometry allows them to be wedged within reef crevices, thus avoiding the need for adhesives and nails for attachment [28, 29]. Nonetheless, whether corals are derived from asexual or sexual propagation, the transplantation of corals remains costly, labor-intensive, time consuming and in many cases, has failed to restore self-sustaining coral populations and associated ecological functions at the restoration site [30, 31].
A less commonly used approach that avoids the need for manual transplantation of corals settled onto artificial substrates is the introduction of large numbers of competent larvae directly onto reef areas. This approach is however still underdeveloped and poorly explored, as most previous studies have either provided accounts of larval release in the sea without quantifying subsequent settlement and recruitment rates [32], or experiments were carried out in small areas (1 × 1–2 m) using artificial substrata, and lacked long-term monitoring or direct monitoring on the natural reef substrata [24, 33]. Overall, these early experiments did not demonstrate an increase in adult coral cover as a result of increasing larval supply.
Recent work by dela Cruz and Harrison [17] provided the only case study demonstrating a significant effect of enhancing larval supply on subsequent recruitment and increased adult coral cover on degraded reef areas, and re-established a breeding population within three years. About 400,000 A. tenuis larvae were released into four 4 × 6 m plots on degraded reef areas in the northwestern Philippines that were temporarily enclosed with fine mesh matting during the larval settlement period. Initial larval settlement was high and juvenile survivorship began stabilising after five months. At least two colonies per m2 survived on the available natural reef substrata and these grew rapidly and spawned successfully at the age of three years, thereby completing the coral life cycle.
Acknowledging the gap between the small scale of most current restoration attempts and current state of degraded coral reefs worldwide, there is increasing emphasis on the need to upscale restoration interventions that can create persistent, viable and ecologically functioning reef communities [5, 7, 34]. Many millions of larvae are produced after major coral spawning events [35, 36] that can be used for restoration, and there are well-established techniques for ex situ larval culture of different coral species [18, 34]. Therefore, there are good opportunities for improving and modifying the larval enhancement techniques for coral restoration interventions to cover larger areas of damaged but recoverable reefs, where larval supply is now limited.
Different coral species may respond differently to specific coral restoration methods. For example, Miller [37] had found that the survivorship of transplanted Acropora palmata recruits cultured from larvae was generally higher than for Orbicella faveolata recruits in the Caribbean. Given the differences in life-history traits among corals, and different sensitivities of larvae to abiotic and biotic factors (e.g., temperature, water quality, conditions on settlement substrata etc.), coupled with the dynamic nature of post-settlement survivorship and recruitment processes [34, 38–40], repeated trials of the larval enhancement technique with larvae of different coral species are needed to verify this approach.
Accordingly, in this paper we examine the broader applicability of the larval enhancement technique using larvae of Acropora loripes. This is a relatively common and widely distributed coral species in the Indo-Pacific region [38, 41]. As of 2014, it was listed as a “near threatened species” by the IUCN due to extensive population reduction [42]. Acropora loripes has a different life-history compared to A. tenuis, and grows more slowly and is likely to reach reproductive maturity later [38]. These corals are also phylogenetically divergent and belong in different clades [43], and A. tenuis is an “early spawner” that spawns just after sunset, and A. loripes is a “late spawner” that spawns a few hours after sunset [44]. Previous laboratory experiments have shown that both species exhibit high fertilization rates (>90%) and larval settlement rates (>50%) [45, 46]. However, A. loripes showed better survivorship than A. tenuis in laboratory experiments [45, 46]. Although it is one of the most common corals used for the aquarium trade [47], it is not a popular experimental species and very few researchers have used it for coral restoration intervention or ex situ coral culture [45, 48].
Materials and methods
Experimental design
This experiment was designed to test the effect of supplying large numbers of Acropora loripes coral larvae on replicate degraded reef areas and to quantify and compare initial larval settlement, and subsequent post-settlement survivorship and growth patterns of recruits for 35 months. The following methods were adapted from dela Cruz and Harrison [17].
Site selection
The A. loripes larval enhancement experiment was conducted on degraded reef areas at Magsaysay reef, Anda, Pangasinan (16°19’36” N, 120°02’01” E) at 4–5 m depth (Fig 1). The A. loripes plots were located between 5 to 15 m away from the plots used for the previous A. tenuis larval enhancement experiment a year earlier [17]. The same reef restoration area was selected to enable more direct comparison of the results from these two separate experiments using two different coral species. This 14.8 ha shallow reef area is an important fishing ground for people in the coastal community of Anda and nearby municipalities [49–52]. The major causes of destruction of the reef areas were blast fishing that no longer occurs, occasional typhoons and a large Crown-of-Thorns starfish infestation that was observed in 2007 [49].
Fig 1. Locations of the experimental larval-enhanced and control plots (Magsaysay reef) and the source of A. loripes colonies (Caniogan and Balingasay reefs) in the Bolinao-Anda Reef Complex, northwestern Philippines.

A total of six 6 × 4 m plots were haphazardly selected and demarcated using steel bars, and three plots were provided with larvae (larval enhancement) and the other three plots served as controls without cultured larvae. The plots were carefully searched for any A. loripes adult colonies and recruits (<5 cm) prior to the larval enhancement activity, and only one adult colony of A. loripes was present within one of the three control plots. Prior to the larval enhancement experiment, photographs of the plots were taken using a 1 × 1 m frame quadrat and photos were analyzed using CPCe [53] to quantify benthic cover and to determine the status of the benthic community. A total of 10 random points were generated and scored in each of 24 frames taken in each of the plots [54].
Ten 10 × 10 cm ‘natural’ settlement tiles (cut from dead table Acropora) with varying thickness (mean 3–4 cm) were deployed inside each plot just prior to the larval enhancement experiment, and each tile was identified with a coded tag (Fig 2A). The coral settlement tiles were designed to be easily removed and re-attached to the reef to allow more accurate, repeated monitoring of initial settlement and subsequent survival rates, especially during earlier stages of this study when recruits were very small and not visible with the naked eye. The total surface area of each tile was estimated to be about 360 ± 3.7 cm2 based on the 3D scanned tiles used in dela Cruz and Harrison [17]. The tiles were directly attached to the substrata using stainless base plates [55]. Dead tabular Acropora plates with naturally growing crustose coralline algae (CCA) used in the production of the tiles were collected at the intertidal zone beside Cory reef rubble bar (Fig 1). Recruitment tiles were biologically conditioned for a month in hatchery tanks at the Bolinao Marine Laboratory (BML) of The Marine Science Institute (University of the Philippines) with flow-through seawater and aeration to promote further CCA growth prior to deployment. This ex situ conditioning of tiles avoided the natural settlement of coral larvae onto the tiles prior to the experiment. This was confirmed by examining the tiles under stereomicroscopes before the experiment to ensure that no coral recruits were present.
Fig 2.

(a) Sample dead coral tile deployed in larval-enhanced and control plots used to determine initial larval settlement. (b) The mesh matting placed onto the larval enhancement plot.
Larval culture
Acropora loripes was used for this experiment. Colonies are usually semi-circular horizontal plates, and do not grow more than a metre across. Acropora loripes colonies have varying growth forms ranging from upright bushes to thick plates. This species is found in most reef environments from shallow reef habitats to 25 m deep, especially on reef slopes protected from wave action. In the Bolinao-Anda Reef Complex (BARC), northwestern Philippines, A. loripes can be found from shallow (2 m) to deeper reef areas (9 m).
A week before the full moon in June 2014, twenty-five gravid colonies of A. loripes (diameter of 7–15 cm) were collected from ~2–7 m deep reef areas on Caniogan reef and Balingasay reef, Bolinao and Caniogan, Anda, Pangasinan (Fig 1). The collections of corals were allowed under the Prior Informed Consent Certificate issued by the municipalities of Bolinao (document number 320:2013) and Anda (document number 165:2012). Colonies were confirmed to be gravid by carefully breaking a few branches to check for the presence of pigmented (pink to red) maturing oocytes [35]. Collected gravid colonies were carefully transported in polyethylene bins with seawater to the BML hatchery facility for ex situ observation of spawning and gamete collection.
Coral ex situ spawning, gamete collection and larval culture followed standard protocols [21, 56–59]. Colonies were held in concrete tanks with flow-through seawater and aeration. Spawning was checked nightly by periodic monitoring of colonies from 1930 to 2130 h. Seawater flow and aeration were turned off during each monitoring period to prevent disturbance to corals that may delay spawning. Spawning occurred 120 to 150 minutes after sunset on June 16 and 17, 2014 (3 and 4 nights after the full moon) and gametes from these spawning events were collected and used for the experiment.
Spawned egg-sperm bundles were skimmed off the water surface using 400 mL plastic cylindrical containers. The collecting container was slowly submerged into the water surface to allow the egg-sperm bundles and small amounts of seawater to flow in. Collected egg-sperm bundles were then transferred to a fertilization polyethylene tank containing 10 L of 1 μm filtered seawater. Gamete bundles were gently agitated to facilitate gamete separation and subsequent cross-fertilization. After 1 h, excess sperm were removed (sperm-washing) by slowly opening the valve located near the bottom of the fertilization tanks. The valve was closed before the water level with floating eggs reached the outflow, and new filtered seawater was slowly added. Washing was done thrice to remove excess sperm that may degrade water quality during larval culture [60]. Subsamples of embryos and eggs were collected using 15 mL tubes and examined under a stereomicroscope after a further hour to determine the percentage fertilization.
Developing embryos were transferred into 11 rearing tanks each containing 1,000 L of seawater. Fresh filtered seawater was added every day (~50 L) to maintain the developing larvae in a healthy condition. Aeration was supplied 24 h after fertilization. At 4 d post-fertilization, an estimated 939,000 ± 29,000 SEM competent larvae were collected using plankton net sieves (60 μm mesh pore) and transferred to a temporary holding tank. The total number of larvae was estimated by thoroughly mixing and dispersing larvae throughout the water column then taking three replicate 60 mL samples. The larvae were then distributed equally into 40 × 50 cm strong plastic bags. Oxygen was supplied to each bag before it was sealed for transport to the field for the larval enhancement experiment.
Coral larval enhancement
The in situ larval settlement enclosure system used by dela Cruz and Harrison [17] was used for this experiment (Fig 2B). Just prior to the experiment, corals inside the plots were temporarily covered with Amazon™ plastic mesh with pore size of 1 cm to avoid the matting from ripping or damaging coral tissues, and the mesh was removed after the 5-day settlement period. The larval mesh enclosures measured 6 × 4 m and were composed of a layer of organza cloth (100–150 μm mesh pore sizes) sewn onto a second layer of nylon net (1 mm openings). This matting assembly can effectively retain Acropora larvae whose diameters are 300–500 μm [56, 61]. To firmly hold the matting on the reef substrata and prevent the larvae from drifting out of the plot during the settlement period, cylindrical lead weights (1.75 × 4.00 cm; 20 g) were inserted along the matting edge. Additional steel bars, driven into the reef substrata, were added on each corner of the matting.
Approximately 300,000 A. loripes larvae were added to each of three replicate 4 × 6 m mesh enclosures on reef plots, with three replicate control plots that were not provisioned with cultured larvae. Larval mesh enclosures were removed from each of the six plots after the 5-day settlement period and the settlement tiles were carefully collected and transported to the BML facility while submerged in seawater, where the initial number of settled larvae on each tile were recorded under stereomicroscopes. Tiles were then returned to their correct location and orientation within each reef plot, and the survival and growth of settled spat on tiles was monitored at 2, 4, 6, 8, 10, 18, 21, 23, 25, 27, 31, 33 and 35 months after settlement.
Coral recruits on natural reef substrata
Surviving A. loripes recruits on natural reef substrata were visible for in situ monitoring at 8 months after settlement. At this time point, each juvenile colony was identified with a numbered aluminum tag placed nearby to facilitate repeated in situ monitoring of growth and survival at 8, 10, 18, 21, 23, 25, 27, 31, 33 and 35 months after settlement.
In situ growth monitoring commenced at this time, with the length (l), width (w) and height (h) of each of the juvenile corals on recruitment tiles and natural substrata measured using calipers. Mean planar diameter was calculated from the maximum and minimum diameters measured for each colony. The ecological volume (EV) was calculated using the volume formula: EV = πr2h, where r = (l+w)/4 [31]. Growth rates (ecological volume change per month) were also calculated, using the formula Gr = [EVf−EVi]/m, where Gr is standardized growth rate, EVf and EVi are final and initial mean ecological volumes, respectively, and m is the number of months elapsed [62].
The onset of sexual reproduction in A. loripes colonies that recruited onto coral tiles and natural reef substratum was monitored at age 23 and 35 months by carefully breaking small branches to check for the presence of gametes [35]. The broken branches were then gently wedged back between the fragments of the colony to avoid loss of branches and tissues. These gamete monitoring periods were completed just prior to the predicted potential spawning periods after 2 and 3 years of growth [38].
Coral production cost analysis
To estimate the cost of producing sexual coral recruits from this study, the costs were categorised and then summed for all materials and infrastructure, boat hire and fuel, diving and labour for gravid coral collection (using different wage rates for different skill levels as prescribed by the Department of Science and Technology Grant-in-Aid personnel), spawning and larval rearing, site preparation and capital costs for the larval mesh enclosures. To estimate the cost per coral colony produced at different ages, the total cost was divided by the total numbers of juvenile corals alive at 8 months and at three years of age in the three larval enhancement plots. Costs were in Philippine Pesos and were converted to US Dollars.
Statistical analyses
Data are reported as mean values ± standard error of the means. The three larval enhancement sites and the three control sites were used as statistical replicates (N = 3), with data from the ten tiles in each site averaged to quantify mean initial settlement rates, and subsequent growth and number of surviving recruits at age 35 months.
The variability in the benthic cover composition (e.g., sand, rubble, macroalgae, coral) between larval enhancement and control plots prior the larval enhancement experiment was analysed using Analysis of similarities (ANOSIM). Significant differences in the initial settlement patterns on tiles between larval-enhanced and control plots after five days of settlement was tested using One-way ANOVA. A post-hoc Tukey’s HSD test was conducted to determine any significant differences in settlement patterns among tile surface types (i.e., top, bottom, sides).
Survivorship of coral recruits on different surfaces of tiles was analysed using survival analysis, a non-parametric pairwise comparison test based on the Kaplan–Meier function [63]. The same analysis was used to determine any significant difference in survival patterns of juvenile corals on natural substrata and on tiles from nine to 35 months after the larval enhancement. Significant increases in growth (ecological volume and mean diameter) of juvenile corals through time were determined using Repeated Measures ANOVA. One-way ANOVA was used to compare growth rates (expressed as ecological volume and mean diameter) of juvenile corals on recruitment tiles versus growth rates on natural substrata. To determine if the assumptions of ANOVA were met, Shapiro-Wilk normality tests and Levene’s test of homoscedasticity were used on each independent variable. Sphericity tests were also conducted prior to running all repeated measure ANOVAs.
Results
Benthic cover and reef condition
Larval-enhanced and control plots had comparable benthic cover compositions before the larval enhancement experiment (ANOSIM, R: -0.11, p = 0.8). The benthic communities within the plots corresponded to the category “very poor coral cover” (sensu Wilkinson) [64] comprising of 12.3 ± 1.9% mean live coral cover (branching + non-branching hard corals; Fig 3). Other biotic benthic categories included low cover of soft coral, sponges, macroalgae, with dead coral covered with turf algae comprising 27.8 ± 4.5% mean cover. Plots also contained 36.7 ± 3.8% mean cover of abiotic dead coral rubble and dead hard coral substrata (Fig 3), with about 8 m2 (out of the 24 m2) of available substrata (dead hard coral + rubble) per plot for the larvae to settle.
Fig 3. Mean percentage cover of benthic categories in larval-enhanced (N = 3) and control (N = 3) plots before the experiment.

Total available area on each plot for coral settlement, which is about 8 m2, is a combination of dead hard coral and rubble. Error bars are ± SEM.
Larval development and initial settlement
A total of 834 A. loripes spat settled on the thirty 10 × 10 cm biologically conditioned natural Acropora skeleton tiles that were attached to reef surfaces in the larval enhancement treatment plots during the 5-day settlement period. Mean total settlement on the sets of ten tiles in each of the three larval enhancement treatment plots was 278 ± 67.3 spat, which was significantly higher than for control sites in which no A. loripes spat settled (Fig 4A).
Fig 4.
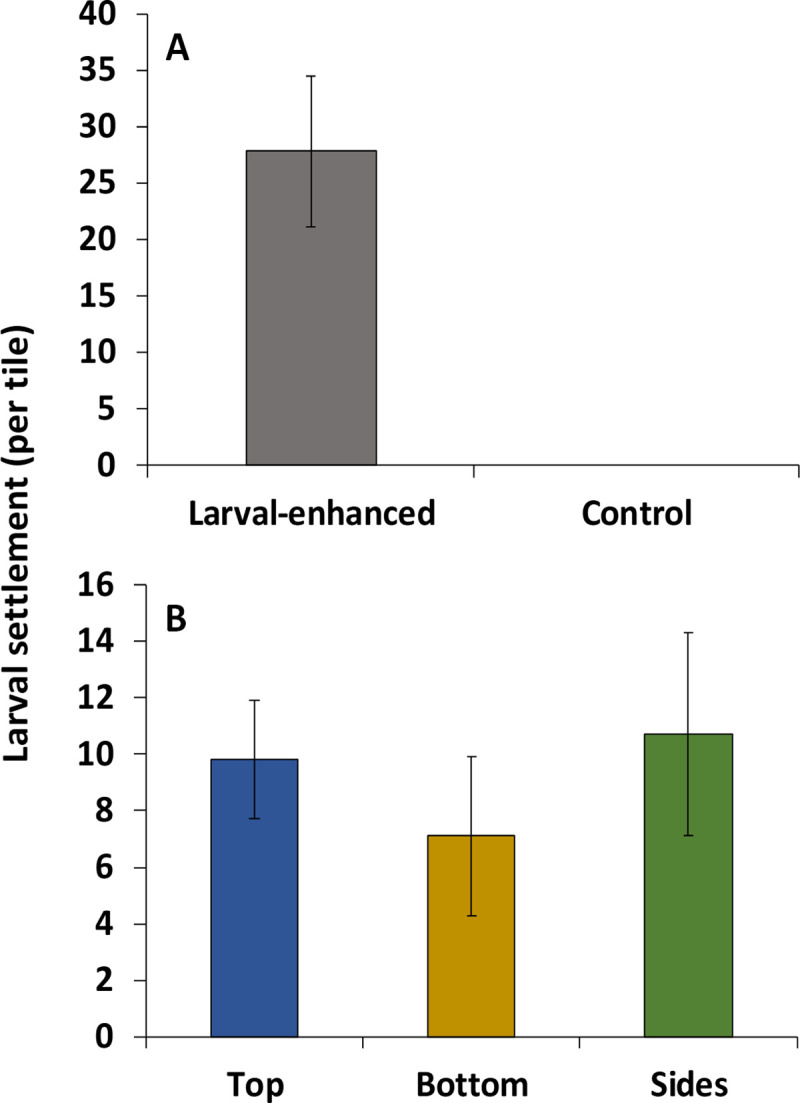
(a) Mean initial A. loripes larval settlement on all tile surfaces in the larval-enhanced (N = 3) and control (N = 3) plots after 5 days, and (b) mean settlement on the different surfaces of tiles in the larval-enhanced plots (N = 3). Means are for ten tiles per plot and are averaged among the three replicate plots. Error bars are ± SEM.
Mean larval settlement on top surfaces of the tiles was 98 ± 21.4, with 71.3 ± 28.3 and 108.7 ± 36.2 mean settlement on the bottom and side surfaces, respectively (Fig 4B). No significant differences were found between the mean number of settled spat among the surfaces of the settlement tiles (F7,1 = 1.67, p = 0.55).
Survivorship
Monitoring of coral spat on tiles showed the expected decline in survivorship after the larval settlement period. An almost 50% decline in survivorship on tiles was recorded after 2 months (Fig 5A). Survivorship continued to decline until 10 months, then few mortalities were recorded on tiles during subsequent monitoring periods up to 35 months (Fig 5A).
Fig 5.

Kaplan-Meier survivorship for 35 months for (a) A. loripes recruits settled on tiles (N = 834), and for visible recruits on natural reef substrata starting at 8 months post-settlement (N = 72) (b) A. loripes recruit survivorship on different tile surfaces (N = 834).
After eight months, the previously cryptic juveniles that had settled on the natural reef substrata became visible recruits at 1.2 ± 0.04 cm mean diameter (1.1 ± 0.2 cm3 mean ecological volume) for in situ growth and survivorship monitoring (Fig 5A). Eight months after settlement, a total of 72 A. loripes recruits were recorded on the natural reef substrata in the three larval enhancement plots, and a total of 23 recruits survived on the settlement tiles (95 juveniles in total).
For corals that settled on the natural reef substrata, 100% survivorship was recorded during the first 8 months after they became visible, and survivorship declined slowly and reached 59% survivorship on the last monitoring period in May 2017, 35 months after settlement (Fig 5A). Survivorship of recruits on tiles from June 2014 to May 2017 varied significantly among tile surfaces (χ2 = 93.3, p = <0.01, Log-rank test; top = sides > bottom; Fig 5B). During the first 9 months, survivorship of juvenile corals on the bottom of the tiles was higher compared to top and side surfaces. However, from 10 months onward most of the juveniles surviving were from the top and sides of tiles, and all the remaining juveniles at the last monitoring period were from the top surface. The mean total number of surviving recruits on the ten tiles in each settlement site after 35 months was 1.7 ± 0.3, which equates to 4.6 ± 0.9 colonies per m2 of tile surface. The total number of surviving A. loripes on the natural reef substrata after 35 months was 42 colonies, with five surviving colonies on the tiles (47 surviving colonies in total).
Recruit growth and onset of sexual reproduction
Initial mean volumes of juvenile corals on the natural substrata and on tiles 8 months after the larval enhancement activity were 1.1 ± 0.1 cm3 and 1.3 ± 0.9 cm3 respectively (Figs 6A, 6B, 7A and 7B). After 35 months, the mean volumes of corals on the tiles reached 438.1 ± 5.4 cm3 and 369.1 ± 8.4 cm3 for the corals on natural substrata (Figs 6A, 7C and 7D). The average volumes of corals on tiles and natural substrata did not differ significantly during any of the monitoring periods (Repeated Measures ANOVA; p = >0.05). Average growth rates of A. loripes recruits monitored on natural reef substrata from 8 months to 35 months reached 14 ± 3 cm3 mo-1 and were similar (F1,4 = 2.36, p = 0.71, ANOVA) to growth rates of recruits that settled on tiles (16 ± 5.7 cm3 mo-1). Thirty-five months after settlement, mean diameters of the colonies ranged from 2.2 cm to 13.9 cm with an average of 7.9 ± 0.6 cm (Fig 6B). Sampling of coral fragments for potential spawning periods after two and three years indicated that none of the colonies had become sexually reproductive.
Fig 6.

Mean volume (a) and mean diameter (b) of juvenile A. loripes on recruitment tiles and natural substrata inside the three larval-enhanced plots. * No data. Error bars are ± SEM.
Fig 7.

Representative A. loripes recruits at 8 months post-larval enhancement on (a) natural reef substrata and (b) on settlement tiles. Three-year-old A. loripes corals on (c) natural reef substrata and (d) on settlement tiles.
Production costs
The total production cost for the sexually derived A. loripes colonies in this study was USD 1654.00. The cost of producing the mesh mattings was divided into two because these mesh enclosures were re-used from the previous larval enhancement activity with A. tenuis [17]. This equates to a production cost of USD 17.41 for each of the 95 recruits alive at 8 months, and USD 35.20 for each of the 47 colonies surviving after 35 months (Table 1).
Table 1. Summary of costs for different activities related to larval enhancement and production of coral colonies.
| Activities | Days | Total (USD) |
|---|---|---|
| I. Collection of gravid A. loripes colonies | 1 | |
| A. Materials (including SCUBA equipment rental) | 74.32 | |
| B. Boat rental | 105.40 | |
| C. Hired labor (2 personnel) | 39.11 | |
| Sub-total | 218.83 | |
| II. Hatchery work | 10 | |
| A. Facility and support system | 9.10 | |
| B. Culture tanks and accessories* | 10.40 | |
| C. Hired labor (2 personnel) | 357.80 | |
| Sub-total | 377.30 | |
| III. Larval enclosures production** | ~15 | |
| A. Materials | 145.40 | |
| B. Hired labor (3 personnel) | 103.56 | |
| Sub-total | 248.96 | |
| IV. Site selection and preparation | 1 | |
| A. Materials (including SCUBA equipment rental) | 171.82 | |
| B. Boat rental | 105.40 | |
| C. Hired labor (3 personnel) | 54.67 | |
| Sub-total | 331.89 | |
| V. Larval enhancement activity | ||
| A. Materials (including SCUBA equipment rental) | 1 | 253.94 |
| B. Boat rental | 105.40 | |
| C. Hired labor (10 personnel) | 118.00 | |
| Sub-total | 477.34 | |
| TOTAL | 1654.32 | |
| Cost per colony 8 months post-larval enhancement | 17.41 | |
| Cost per colony 35 months post-larval enhancement | 35.20 |
SCUBA gear hire, air tanks and boat rental from BML to the study site were based on 2013 and 2014 rates. The BML outdoor hatchery facility and support system costs include equipment (seawater pump, air blower, pipe assemblies, sedimentation tank), maintenance and electricity. Costs were originally estimated in Philippine Pesos (PhP) and converted to USD using the conversion rate: PhP 45 = USD 1.
*total value divided over 10 years of use
**total value divided by two uses.
Discussion
Natural reef recovery following a disturbance depends on ample coral larval supply, successful settlement of larvae and survival to adult reproductive age [39, 65, 66]. Each of these coral life-history stages is a potential bottleneck that significantly affects coral population recovery. In previous decades, coral restoration interventions have primarily utilised juvenile and adult coral stages as transplant materials to immediately increase coral cover on degraded reefs [9]. Recent research has focused on sexual production of corals to increase genetic diversity and potential resilience [19, 26, 34], and the previous pilot study using A. tenuis larvae clearly demonstrated that supplying competent swimming coral larvae to settle directly on degraded reefs significantly increased coral recruitment [17]. The A. tenuis long-term reef experiment was the first to demonstrate that larval enhancement on degraded reef areas significantly increases settlement leading to increased recruitment and coral cover, and also re-established reproductively mature adult coral colonies within three years.
Over the past few decades, advances in ex situ larval culture research have improved the rearing and production of larvae for small scale laboratory assays and growing juvenile corals for transplantation [18, 22, 67, 68]. Larger scale ex situ larval culture of various coral species has also been used to rear millions of larvae for coral restoration experiments [17, 28, 33]. Ex situ larval culture may increase survivorship of settled larvae by carefully selecting stress tolerant, including heat-tolerant, breeding corals. Also, the ability to manipulate culture conditions using stress-hardening may produce larvae better adapted to warming environments, promoting better survivorship [6]. Furthermore, rearing large numbers of coral larvae from multiple coral colonies, whether in ex situ cultures or from natural coral spawn slicks, creates large pools of new genetic variants of corals. An advantage of the larval restoration approach is that these genetically diverse larvae are exposed to natural reef conditions during settlement, avoiding artificial selection pressures during settlement and early life stages compared with laboratory cultures that might prove to be maladapted to environmental conditions at the restoration sites. In the present study, 900,000 A. loripes larvae were successfully cultured ex situ in culture tanks and the larvae were readily transported and released into the reef plots where some A. loripes corals were observed to be growing nearby.
Unlike other Acropora species in Magsaysay reef (i.e. A. cytherea, A. hyacinthus, A. muricata, A. sarmentosa etc.) that recovered from stressors that these local reefs experienced during recent decades, only four colonies of A. loripes were recorded surviving in surveys on ~200 m2 of nearby reef area. With very few adult colonies to supply gametes or larvae to aid natural recruitment and reef recovery, A. loripes population recovery in Magsaysay reef will be very slow and may not occur without active intervention. In addition, the supply of larvae to surrounding reef areas in the Bolinao-Anda Reef Complex may be compromised, as computer model simulations suggest that Magsaysay reef is a source of coral larvae for the nearby reefs [69]. Therefore, the introduction of large numbers of competent A. loripes larvae to increase settlement and recruitment rates is an active management intervention designed to assist recovery of this coral population in Magsaysay reef. This intervention may also create a potential “spawning hot spot” or “spawning hub” in the future that may provide increased abundance of coral larvae with high genetic diversity for dispersal and restoration of adjacent degraded reef areas [66].
The initial settlement rates of A. loripes larvae on tiles in larval enhancement plots were relatively high, and comparable with those from the A. tenuis larval enhancement experiment, also conducted at the Magsaysay reef area a year earlier [17]. However, settlement patterns of A. loripes among tile surfaces was different from those recorded for A. tenuis. Initial settlement of A. tenuis larvae was significantly higher on tile sides with very few settlers on upper surfaces, whereas for A. loripes, larval settlement was not significantly different among tile surfaces. This suggests that larvae of these species may have different settlement orientation preferences, as recorded among other coral species [34, 70]. The initial settlement rates in the larval enhancement plots were about 70 times higher than the mean level of monitored natural acroporid recruitment in the study area over a two year period [17]. This pattern of enhanced larval settlement was similar to the ~100-fold increase in larval settlement recorded in the larval reseeding trial by Heyward et al. [24] at Ningaloo Reef in Western Australia, and much greater than the four-fold increase in settlement from the reseeding experiment of Edwards et al. [33] in Palau. These results suggest that larval density and benthic community composition can affect post-settlement survival and subsequent recruitment patterns. Therefore, to increase cost effectiveness, the optimal number of larvae introduced in a given reef area should be determined through density experiments to achieve maximum settlement and recruitment rates [71].
Survivorship of settled A. loripes spat on tiles showed a typical Type III survivorship curve. The proportion of surviving spat decreased significantly within the first month and continued to decline until 10 months when survivorship started to stabilise. That survivorship was similar to the survivorship pattern recorded for A. tenuis spat on tiles during the first 10 months after settlement [17], and in other laboratory cultured larvae settled on artificial substrata [72, 73]. These data show that the greatest bottleneck in coral post-settlement survivorship occurs when spat are small and most vulnerable during the first few weeks to months post-settlement [17, 19, 74, 75]. Possible causes for these declines in initial spat survivorship can be attributed to accidental or targetted grazing by fish and other macro-invertebrates, sediment smothering, and competitive interactions with other benthic organisms including microalgae and macroalgae, sponges and soft corals [75–79]. Post-settlement mortality is also intrinsically high among coral spat settled in controlled conditions in laboratory aquaria, even in the absence of predators, competitors, and other factors such as sediments [80]. Therefore, further investigations to determine the major drivers of post-settlement mortality are needed to increase the success of future direct larval enhancement interventions.
The survival rates of A. loripes corals that settled on the natural reef substrata and became visible at 10 months after settlement declined slowly over time but remained above 50% at the end of the 35 months monitoring period. This survivorship pattern contrasts with that recorded for A. tenuis, where there was no observed mortality of the faster growing A. tenuis recruits that settled on reef substrata from nine months to 35 months [17]. The different survivorship patterns of these two Acropora species may reflect intrinsic differences in their life-history characteristics, or may be related to size-escape thresholds [81, 82], with A. tenuis spat growing faster on average and attaining larger sizes more quickly than the A. loripes spat. Similar higher survivorship outcomes have been recorded when outplanting larger size sexually derived Acropora juvenile corals. In a study by Omori et al. [83], A. tenuis juveniles that had survived for 1.5 years in mid-water nurseries and reached 4 to 5 cm in diameter, had 89% survivorship after six months. In contrast, when smaller 1 cm diameter 6-month old A. valida juveniles were outplanted, a lower survivorship of 67% was recorded after six months [21].
After 35 months, the mean number of remaining A. loripes was 14 ± 2.9 per larval enhancement plot, which equates to an average of 1.8 coral colonies per m2 of available reef substrata. This density is slightly lower than the mean density of 2.3 A. tenuis colonies per m2 of available reef substrata recorded by dela Cruz and Harrison (2017), and partly reflects the slower growth and lower survivorship patterns for A. loripes colonies recorded in the present study. Edwards [9] suggested that for coral restoration interventions to be ecologically and economically cost-effective, there should be more than one coral surviving from every 104 settled larvae. The total number of larvae that settled directly on the reef in this experiment could not be quantified because the microscopic larvae are too small to census and tend to have cryptic settlement on the complex reef surfaces. A total of 47 colonies survived after 35 months from an estimated 939,000 larvae added to the larval enhancement plots, which equates to one three-year-old colony per 20,000 larvae supplied. Based on the numbers of larvae settling on the tiles, it is likely that less than 50% of the larvae successfully settled during the five-day settlement period, hence the production rate probably exceeds the threshold suggested by Edwards [9].
This study has also provided the first data on growth rates of sexually produced A. loripes colonies. The growth rates of A. loripes that survived on the tiles and natural substrata (mean diameter 2.6 cm yr-1) was considerably slower than the growth rates of A. tenuis colonies grown from settled larvae at Magsaysay reef (mean diameter 5.1 cm yr-1) [17] and other sexually produced Acropora corals. Cultured A. tenuis larvae that settled on artificial substrata in an outdoor hatchery, and were subsequently transplanted to reef areas in subtropical Okinawa, Japan reached a mean diameter of 20 cm (4 cm yr-1) after 4 years of transplantation [84]. Similar growth rates were recorded for A. millepora corals cultured and maintained in nurseries for 3 years (4 cm yr-1 mean diameter), or transplanted onto reef areas (4.6 cm yr-1 mean diameter) [19, 85].
In contrast to sexually derived A. tenuis colonies that reached sexual maturity at 35 months when they had grown to a colony size larger than 12.5 cm mean diameter [17], none of the A. loripes colonies were gravid at 35 months age. This indicates that the minimum age at first reproduction in A. loripes might be at least four or five years, as predicted by Wallace [38]. Furthermore, the sizes of gravid colonies collected at the start of this study (7–15 cm mean diameter) show that some colonies can be sexually reproductive at sizes less than 12 cm mean diameter, which confirms previous information on the minimum size of reproduction of A. loripes [38, 86]. The delay in the onset of sexual reproduction of A. loripes colonies surviving at the end of this study suggests that size-dependent reproduction may not strictly apply in A. loripes, rather sexual reproduction could be influenced by both age and size [34, 86]. These differences in life-history patterns among Acropora species highlight the importance of choosing the best coral species for achieving specific coral restoration goals.
Some of the key challenges for coral restoration are to lower the production cost and to be adaptable to developing nations where most of the world’s coral communities occur and which are subjected to rapidly increasing anthropogenic threats [9, 87]. The low-cost mesh matting larval enclosures used in this study were made using widely available materials including organza (wedding veil) cloth, and were constructed using a standard non-electric sewing machine by local people. The nets were sturdy enough to withstand the underwater environmental conditions during the 5-day larval settlement period. The average production cost per A. loripes colony after 8 months ($17.41) and 35 months ($35.20) was slightly higher than the production costs for A. tenuis colonies at similar ages ($14.77 and $20.94, respectively) [17]. Chamberland et al. [29] recorded a production cost of $13 USD per 2.5 year old Acropora palmata colony outplanted onto reef areas two weeks after settlement onto clay tripods, and a higher cost of $325 USD per colony maintained in a land-based nursery. The production costs for the larval enhancement method are significantly lower in large part because settlers are not maintained in ex situ or in situ grow-out nursery installations over long periods of time. The overall cost-effectiveness of the larval enhancement technique will also increase as restored colonies become sexually reproductive and contribute larvae to the natural larval pool that can re-populate and colonize other available reef areas in future.
One of the advantages of the larval enhancement method using temporary mesh enclosures is the applicability and adaptability of the technique to various corals and reef types with different environmental conditions. In the present study, flat mesh matting was used to contain larvae on reef areas with relatively low topographical relief, and the reef experiences occasional strong wave action. For reef systems with more complex substrata, the enclosures can be modified to adapt to the contours of the reef. Furthermore, the availability of plankton nets with various mesh size openings makes the larval enhancement technique applicable to a wide range of spawning and brooding coral species that produce larvae of different sizes. Future larval enhancement interventions could be done without mesh enclosures by releasing competent larvae at optimal densities during periods of low tidal flows and calm weather and low wave action. This technique should also be tested in different reef regions around the world with different environmental conditions and topography, for comparison with other existing coral restoration techniques.
While high post-settlement mortality remains a key challenge for coral restoration using sexual production [34], the results of this study demonstrate that settlement and recruitment can be significantly enhanced via the provision of large numbers of sexually produced larvae. The larval enhancement technique has the advantage of settling larvae in situ on the reef and therefore surviving corals are likely to be well-adapted to local environmental conditions. This technique also avoids the need for large scale production of artificial settlement substrata and construction of in situ or ex situ coral nurseries for coral husbandry, and the extra effort and expense of manually transporting and attaching the coral recruits or colonies onto the reef substrata [5, 9]. In addition, direct larval provision onto degraded reefs has strong potential for scaling up restoration efforts to larger reef areas in future.
Acknowledgments
We thank the staff and research assistants at the Bolinao Marine Laboratory of the Marine Science Institute, University of the Philippines for logistical support with this research especially E. Gomez, C. Ligson, R. Gomez and M. Ponce. We thank C. Barlow and A. Fleming from ACIAR for ongoing support. This publication is dedicated to our colleague at BML, Dr Ronald D. Villanueva who became terminally ill during 2015. This is MSI Contribution No. 476, and contribution 2 from the Southern Cross University ACIAR Coral Larval Restoration reef restoration trials.
Data Availability
The raw data of the study is deposited and can be accessed publicly on the following links: (https://doi.org/10.25918/data.41) (https://scu.primo.exlibrisgroup.com/permalink/61SCU_INST/v1pbpl/alma991012858600102368).
Funding Statement
This work was supported by the Australian Centre for International Agricultural Research (ACIAR) grants SRA FIS/2011/031 and FIS/2014/063 Coral Reef Restoration Project to P Harrison. The funders did not play any role in the study design, data collection, analysis and preparation of the manuscript.
References
- 1.De'ath G, Fabricius K, Lough J. Yes—Coral calcification rates have decreased in the last twenty-five years! Mar Geol. 2013;346: 400–2. [Google Scholar]
- 2.Hughes TP, Anderson KD, Connolly SR, Heron SF, Kerry JT, Lough JM, et al. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science. 2018;359: 80–3. 10.1126/science.aan8048 [DOI] [PubMed] [Google Scholar]
- 3.Briggs JC. Coral reefs: conserving the evolutionary sources. Biol Conserv. 2005;126: 297–305. [Google Scholar]
- 4.Anthony KR. Coral reefs under climate change and ocean acidification: challenges and opportunities for management and policy. Annu Rev Environ Resour. 2016;41: 59–81. [Google Scholar]
- 5.Boström-Einarsson L, Babcock RC, Bayraktarov E, Ceccarelli D, Cook N, Ferse SC, et al. Coral restoration–A systematic review of current methods, successes, failures and future directions. PLoS ONE. 2020;15: e0226631 10.1371/journal.pone.0226631 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.van Oppen MJ, Gates RD, Blackall LL, Cantin N, Chakravarti LJ, Chan WY, et al. Shifting paradigms in restoration of the world's coral reefs. Glob Change Biol. 2017;23: 3437–48. 10.1111/gcb.13647 [DOI] [PubMed] [Google Scholar]
- 7.Rinkevich B. Ecological engineering approaches in coral reef restoration. ICES J Mar Sci. 2020;fsaa022 10.1093/icesjms/fsaa022. [DOI] [Google Scholar]
- 8.Rinkevich B. Rebuilding coral reefs: does active reef restoration lead to sustainable reefs? Curr Opin Environ Sustain. 2014;7: 28–36. [Google Scholar]
- 9.Edwards AJ. Reef Restoration Manual. St Lucia, Australia: Coral Reef Targeted Research & Capacity Building for Management Program; 2010. 166 p. [Google Scholar]
- 10.dela Cruz DW, Rinkevich B, Gomez ED, Yap HT. Assessing an abridged nursery phase for slow growing corals used in coral restoration. Ecol Eng. 2015;84: 408–15. [Google Scholar]
- 11.Shaish L, Levy G, Gomez E, Rinkevich B. Fixed and suspended coral nurseries in the Philippines: Establishing the first step in the “gardening concept” of reef restoration. J Exp Mar Biol Ecol. 2008;358: 86–97. [Google Scholar]
- 12.Ligson CA, Tabalanza TD, Villanueva RD, Cabaitan PC. Feasibility of early outplanting of sexually propagated Acropora verweyi for coral reef restoration demonstrated in Philippines. Restor Ecol. 2019. 10.1111/rec.13054 [DOI] [Google Scholar]
- 13.Lirman D, Thyberg T, Herlan J, Hill C, Young-Lahiff C, Schopmeyer S, et al. Propagation of the threatened staghorn coral Acropora cervicornis: methods to minimize the impacts of fragment collection and maximize production. Coral Reefs. 2010;29: 729–35. [Google Scholar]
- 14.Mbije NE, Spanier E, Rinkevich B. A first endeavour in restoring denuded, post-bleached reefs in Tanzania. Estuar Coast Shelf Sci. 2013;128: 41–51. [Google Scholar]
- 15.Horoszowski-Fridman YB, Brêthes J-C, Rahmani N, Rinkevich B. Marine silviculture: Incorporating ecosystem engineering properties into reef restoration acts. Ecol Eng. 2015;82: 201–13. [Google Scholar]
- 16.Bayraktarov E, Stewart‐Sinclair PJ, Brisbane S, Boström‐Einarsson L, Saunders MI, Lovelock CE, et al. Motivations, success, and cost of coral reef restoration. Restor Ecol. 2019;27: 981–91. [Google Scholar]
- 17.dela Cruz DW, Harrison PL. Enhanced larval supply and recruitment can replenish reef corals on degraded reefs. Sci Rep. 2017;7: 13985 10.1038/s41598-017-14546-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Pollock FJ, Katz SM, van de Water JA, Davies SW, Hein M, Torda G, et al. Coral larvae for restoration and research: a large-scale method for rearing Acropora millepora larvae, inducing settlement, and establishing symbiosis. PeerJ. 2017;5: e3732 10.7717/peerj.3732 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Guest J, Baria M, Gomez E, Heyward A, Edwards A. Closing the circle: is it feasible to rehabilitate reefs with sexually propagated corals? Coral Reefs. 2014;33: 45–55. [Google Scholar]
- 20.van Oppen MJ, Oliver JK, Putnam HM, Gates RD. Building coral reef resilience through assisted evolution. Proc Natl Acad Sci U S A. 2015;112: 2307–13. 10.1073/pnas.1422301112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Villanueva RD, Baria MVB, dela Cruz DW. Growth and survivorship of juvenile corals outplanted to degraded reef areas in Bolinao-Anda Reef Complex, Philippines. Mar Biol Res. 2012;8: 877–84. [Google Scholar]
- 22.Omori M, Iwao K. Methods of farming sexually propagated corals and outplanting for coral reef rehabilitation; with list of references for coral reef rehabilitation through active restoration measure. Okinawa, Japan: Akajima Marine Science Laboratory, 2014. [Google Scholar]
- 23.Calle-Triviño J, Cortés-Useche C, Sellares-Blasco RI, Arias-González JE. Assisted fertilization of threatened Staghorn Coral to complement the restoration of nurseries in Southeastern Dominican Republic. Regional Studies in Marine Science. 2018;18: 129–34. [Google Scholar]
- 24.Heyward A, Smith L, Rees M, Field S. Enhancement of coral recruitment by in situ mass culture of coral larvae. Mar Ecol Prog Ser. 2002;230: 113–8. [Google Scholar]
- 25.Doropoulos C, Vons F, Elzinga J, ter Hofstede R, Salee K, van Koningsveld M, et al. Testing industrial-scale coral restoration techniques: harvesting and culturing wild coral-spawn slicks. Front Mar Sci. 2019;6: 658. [Google Scholar]
- 26.Harrison P, Villanueva R, dela Cruz D. Coral Reef Restoration Using Mass Coral Larval Reseeding. Australian Centre for International Agricultural Research, Canberra, Australia: 2016. [Google Scholar]
- 27.Baria‐Rodriguez MV, dela Cruz DW, Dizon RM, Yap HT, Villanueva RD. Performance and cost‐effectiveness of sexually produced Acropora granulosa juveniles compared with asexually generated coral fragments in restoring degraded reef areas. Aquatic Conservation: Marine and Freshwater Ecosystems. 2019;29: 891–900. [Google Scholar]
- 28.Chamberland VF, Petersen D, Guest JR, Petersen U, Brittsan M, Vermeij MJ. New seeding approach reduces costs and time to outplant sexually propagated corals for reef restoration. Sci Rep. 2017;7: 18076 10.1038/s41598-017-17555-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Chamberland VF, Vermeij MJ, Brittsan M, Carl M, Schick M, Snowden S, et al. Restoration of critically endangered elkhorn coral (Acropora palmata) populations using larvae reared from wild-caught gametes. Glob Ecol Conserv. 2015;4: 526–37. [Google Scholar]
- 30.Cabaitan PC, Gomez ED, Aliño PM. Effects of coral transplantation and giant clam restocking on the structure of fish communities on degraded patch reefs. J Exp Mar Biol Ecol. 2008;357: 85–98. [Google Scholar]
- 31.Shaish L, Levy G, Katzir G, Rinkevich B. Coral reef restoration (Bolinao, Philippines) in the face of frequent natural catastrophes. Restor Ecol. 2010;18: 285–99. [Google Scholar]
- 32.Nonaka M, Baird A, Kamiki T, Yamamoto H. Reseeding the reefs of Okinawa with the larvae of captive-bred corals. Coral Reefs. 2003;22: 34–. [Google Scholar]
- 33.Edwards AJ, Guest JR, Heyward AJ, Villanueva RD, Baria MV, Bollozos IS, et al. Direct seeding of mass-cultured coral larvae is not an effective option for reef rehabilitation. Mar Ecol Prog Ser. 2015;525: 105–16. [Google Scholar]
- 34.Randall CJ, Negri AP, Quigley KM, Foster T, Ricardo GF, Webster NS, et al. Sexual production of corals for reef restoration in the Anthropocene. Mar Ecol Prog Ser. 2020;635: 203–32. [Google Scholar]
- 35.Harrison PL, Babcock RC, Bull GD, Oliver JK, Wallace CC, Willis BL. Mass spawning in tropical reef corals. Science. 1984;223: 1186–9. 10.1126/science.223.4641.1186 [DOI] [PubMed] [Google Scholar]
- 36.Babcock R, Heyward A. Larval development of certain gamete-spawning scleractinian corals. Coral Reefs. 1986;5: 111–6. [Google Scholar]
- 37.Miller MW. Post-settlement survivorship in two Caribbean broadcasting corals. Coral Reefs. 2014;33: 1041–6. [Google Scholar]
- 38.Wallace CC. Reproduction, recruitment and fragmentation in nine sympatric species of the coral genus Acropora. Mar Biol. 1985;88: 217–33. [Google Scholar]
- 39.Gilmour JP, Smith LD, Heyward AJ, Baird AH, Pratchett MS. Recovery of an isolated coral reef system following severe disturbance. Science. 2013;340: 69–71. 10.1126/science.1232310 [DOI] [PubMed] [Google Scholar]
- 40.Gouezo M, Olsudong D, Fabricius K, Harrison P, Golbuu Y, Doropoulos C. Relative roles of biological and physical processes influencing coral recruitment during the lag phase of reef community recovery. Sci Rep. 2020;10: 1–12. 10.1038/s41598-019-56847-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Veron J. Corals of the World. Volumes 1–3. Australian Institute of Marine Science, Townsville, Australia: 2000. [Google Scholar]
- 42.Richards ZT, Delbeek JT, Lovell ER, Bass D, Aeby G, Reboton C. Acropora loripes The IUCN Red List of Threatened Species 2014: IUCN; 2014. Available from: 10.2305/IUCN.UK.2014-1.RLTS.T132948A54164687.en. [DOI] [Google Scholar]
- 43.Fukami H, Omori M, Hatta M. Phylogenetic relationships in the coral family Acroporidae, reassessed by inference from mitochondrial genes. Zool Sci. 2000;17: 689–96. 10.2108/zsj.17.689 [DOI] [PubMed] [Google Scholar]
- 44.van Oppen MJ, McDonald BJ, Willis B, Miller DJ. The evolutionary history of the coral genus Acropora (Scleractinia, Cnidaria) based on a mitochondrial and a nuclear marker: reticulation, incomplete lineage sorting, or morphological convergence? Mol Biol Evol. 2001;18: 1315–29. 10.1093/oxfordjournals.molbev.a003916 [DOI] [PubMed] [Google Scholar]
- 45.Chan WY, Peplow LM, Menéndez P, Hoffmann AA, van Oppen MJ. Interspecific hybridization may provide novel opportunities for coral reef restoration. Front Mar Sci. 2018;5: 160. [Google Scholar]
- 46.Conlan JA, Humphrey CA, Severati A, Francis DS. Influence of different feeding regimes on the survival, growth, and biochemical composition of Acropora coral recruits. PLoS ONE. 2017;12: e0188568 10.1371/journal.pone.0188568 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Borneman E. Aquarium Corals: Selection, Husbandry and Natural History. Neptune City, NJ, USA: TFH Publishing; 2001. [Google Scholar]
- 48.Taira D, Toh T, Sam S, Ng C, Chou L. Coral nurseries as habitats for juvenile corallivorous butterflyfish. Mar Biodivers. 2017;47: 787–8. [Google Scholar]
- 49.Cruz-Trinidad A, Geronimo RC, Cabral RB, Alino PM. How much are the Bolinao-Anda coral reefs worth? Ocean Coast Manag. 2011;54: 696–705. [Google Scholar]
- 50.Tamayo NCA, Anticamara JA, Acosta-Michlik L. National estimates of values of Philippine reefs' ecosystem services. Ecol Econ. 2018;146: 633–44. [Google Scholar]
- 51.Abrina TAS, Bennett J. Using choice modelling to estimate the non-market benefits of coral reef restoration in the Philippines. Ocean Coast Manag. 2019: 105039. [Google Scholar]
- 52.Abrina TAS, Bennett J. Using choice modelling to estimate the non-market benefits of coral reef restoration in the Philippines. Ocean Coast Manag. 2020;185: 105039. [Google Scholar]
- 53.Kohler KE, Gill SM. Coral Point Count with Excel extensions (CPCe): a visual basic program for the determination of coral and substrate coverage using random point count methodology. Comput Geosci. 2006;32: 1259–69. [Google Scholar]
- 54.Licuanan W, Robles R, Reyes M. Status and recent trends in coral reefs of the Philippines. Mar Pollut Bull. 2019;142: 544–50. 10.1016/j.marpolbul.2019.04.013 [DOI] [PubMed] [Google Scholar]
- 55.Mundy C. An appraisal of methods used in coral recruitment studies. Coral Reefs. 2000;19: 124–31. [Google Scholar]
- 56.Wilson J, Harrison P. Settlement-competency periods of larvae of three species of scleractinian corals. Mar Biol. 1998;131: 339–45. [Google Scholar]
- 57.Heyward A, Negri A. Natural inducers for coral larval metamorphosis. Coral Reefs. 1999;18: 273–9. [Google Scholar]
- 58.Nozawa Y, Harrison PL. Temporal settlement patterns of larvae of the broadcast spawning reef coral Favites chinensis and the broadcast spawning and brooding reef coral Goniastrea aspera from Okinawa, Japan. Coral Reefs. 2005;24: 274–82. [Google Scholar]
- 59.Harrison P. Settlement competency periods and dispersal potential of scleractinian reef coral larvae. Proc 10th Int Coral Reef Symp; 2006; Okinawa, Japan.
- 60.Willis B, Babcock R, Harrison P, Wallace C. Experimental hybridization and breeding incompatibilities within the mating systems of mass spawning reef corals. Coral Reefs. 1997;16: S53–S65. [Google Scholar]
- 61.Nishikawa A, Katoh M, Sakai K. Larval settlement rates and gene flow of broadcast-spawning (Acropora tenuis) and planula-brooding (Stylophora pistillata) corals. Mar Ecol Prog Ser. 2003;256: 87–97. [Google Scholar]
- 62.dela Cruz DW, Villanueva RD, Baria MVB. Community-based, low-tech method of restoring a lost thicket of Acropora corals. ICES J Mar Sci. 2014;71: 1866–75. [Google Scholar]
- 63.Lee ET, Wang J. Statistical Methods for Survival Data Analysis. 3rd ed: John Wiley & Sons; 2003. [Google Scholar]
- 64.Wilkinson C. Status of Coral Reefs of the World: 2008. Townsville, Australia: Global Coral Reef Monitoring Network and Reef and Rainforest Research Centre; 2008. 296 p. [Google Scholar]
- 65.Hughes T, Baird A, Dinsdale E, Moltschaniwskyj N, Pratchett M, Tanner J, et al. Supply-side ecology works both ways: the link between benthic adults, fecundity, and larval recruits. Ecology. 2000;81: 2241–9. [Google Scholar]
- 66.Gouezo M, Golbuu Y, Fabricius K, Olsudong D, Mereb G, Nestor V, et al. Drivers of recovery and reassembly of coral reef communities. Proceedings of the Royal Society B. 2019;286: 20182908 10.1098/rspb.2018.2908 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Ward S, Harrison P, editors. The effects of elevated nutrients on settlement of coral larvae during the ENCORE experiment. Proc 8th Int Coral Reef Symp; 1997; Panama.
- 68.Reichelt-Brushett AJ, Harrison PL. The effect of selected trace metals on the fertilization success of several scleractinian coral species. Coral Reefs. 2005;24: 524–34. [Google Scholar]
- 69.Vergara M, Geronimo R, Ticzon V, Dizon R, Villanueva R, Baria M, et al. Status of the coral reef in Bolinao-Anda reef complex, northwestern Philippines In: Deborah C, Jess M, Melanie K, Gabrielle S, editors. Building Capacity in Coral Reef Science: An Anthology of CRTR Scholars’ Research. St Lucia, Australia: The Coral Reef Targeted Research & Capacity Building for Management Program; 2010. p. 60–74. [Google Scholar]
- 70.Babcock R, Mundy C. Coral recruitment: consequences of settlement choice for early growth and survivorship in two scleractinians. J Exp Mar Biol Ecol. 1996;206: 179–201. [Google Scholar]
- 71.Cameron KA, Harrison PL. Density of coral larvae can influence settlement, post-settlement colony abundance and coral cover in larval restoration. Sci Rep. 2020;10: 1–11. 10.1038/s41598-019-56847-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Suzuki G, Kai S, Yamashita H, Suzuki K, Iehisa Y, Hayashibara T. Narrower grid structure of artificial reef enhances initial survival of in situ settled coral. Mar Pollut Bull. 2011;62: 2803–12. 10.1016/j.marpolbul.2011.08.050 [DOI] [PubMed] [Google Scholar]
- 73.Szmant AM, Miller MW, editors. Settlement preferences and post-settlement mortality of laboratory cultured and settled larvae of the Caribbean hermatypic corals Montastraea faveolata and Acropora palmata in the Florida Keys, USA. Proc 10th Int Coral Reef Symp; 2006; Okinawa, Japan.
- 74.Vermeij MJ, Sandin SA. Density‐dependent settlement and mortality structure the earliest life phases of a coral population. Ecology. 2008;89: 1994–2004. 10.1890/07-1296.1 [DOI] [PubMed] [Google Scholar]
- 75.Ritson-Williams R, Arnold SN, Fogarty ND, Steneck RS, Vermeij MJ, Paul VJ. New perspectives on ecological mechanisms affecting coral recruitment on reefs. Smithson Contrib Mar Sci. 2009;38: 437–57. [Google Scholar]
- 76.McCook L, Jompa J, Diaz-Pulido G. Competition between corals and algae on coral reefs: a review of evidence and mechanisms. Coral Reefs. 2001;19: 400–17. [Google Scholar]
- 77.Baria MVB, Guest JR, Edwards AJ, Aliño PM, Heyward AJ, Gomez ED. Caging enhances post-settlement survival of juveniles of the scleractinian coral Acropora tenuis. J Exp Mar Biol Ecol. 2010;394: 149–53. [Google Scholar]
- 78.Davies SW, Matz MV, Vize PD. Ecological complexity of coral recruitment processes: effects of invertebrate herbivores on coral recruitment and growth depends upon substratum properties and coral species. PLoS ONE. 2013;8: e72830 10.1371/journal.pone.0072830 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Tebben J, Guest JR, Sin TM, Steinberg PD, Harder T. Corals like it waxed: paraffin-based antifouling technology enhances coral spat survival. PLoS ONE. 2014;9: e87545 10.1371/journal.pone.0087545 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Doropoulos C, Evensen NR, Gómez-Lemos LA, Babcock RC. Density-dependent coral recruitment displays divergent responses during distinct early life-history stages. Open Science. 2017;4: 170082 10.1098/rsos.170082 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Raymundo LJ, Maypa AP. Getting bigger faster: mediation of size-specific mortality via fusion in junenile coral transplants. Ecol Appl. 2004;14: 281–95. [Google Scholar]
- 82.Doropoulos C, Ward S, Marshell A, Diaz-Pulido G, Mumby PJ. Interactions among chronic and acute impacts on coral recruits: the importance of size‐escape thresholds. Ecology. 2012;93: 2131–8. 10.1890/12-0495.1 [DOI] [PubMed] [Google Scholar]
- 83.Omori M, Iwao K, Tamura M. Growth of transplanted Acropora tenuis 2 years after egg culture. Coral Reefs. 2008;27: 165. [Google Scholar]
- 84.Iwao K, Omori M, Taniguchi H, Tamura M. Transplanted Acropora tenuis (Dana) spawned first in their life 4 years after culture from eggs. Galaxea. 2010;12: 47–. [Google Scholar]
- 85.Baria MVB, dela Cruz DW, Villanueva RD, Guest JR. Spawning of three-year-old Acropora millepora corals reared from larvae in northwestern Philippines. Bull Mar Sci. 2012;88: 61–2. [Google Scholar]
- 86.Harrison P, Wallace C. Reproduction, dispersal and recruitment of scleractinian corals In: Dubinsky Z, editor. Ecosystems of the World. 25 New York: Elsevier; 1990. p. 133–207. [Google Scholar]
- 87.Burke L, Reytar K, Spalding M, Perry A. Reefs at Risk Revisited. Washington, DC: World Resource Institute; 2011. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The raw data of the study is deposited and can be accessed publicly on the following links: (https://doi.org/10.25918/data.41) (https://scu.primo.exlibrisgroup.com/permalink/61SCU_INST/v1pbpl/alma991012858600102368).