

Summary
Vibrio cholerae isolates responsible for cholera pandemics represent only a small portion of the diverse strains belonging to this species. Indeed, most V. cholerae are encountered in aquatic environments. To better understand the emergence of pandemic lineages, it is crucial to discern what differentiates pandemic strains from their environmental relatives. Here, we studied the interaction of environmental V. cholerae with eukaryotic predators or competing bacteria and tested the contributions of the haemolysin and the type VI secretion system (T6SS) to those interactions. Both of these molecular weapons are constitutively active in environmental isolates but subject to tight regulation in the pandemic clade. We showed that several environmental isolates resist amoebal grazing and that this anti‐grazing defense relies on the strains' T6SS and its actincross‐linking domain (ACD)‐containing tip protein. Strains lacking the ACD were unable to defend themselves against grazing amoebae but maintained high levels of T6SS‐dependent interbacterial killing. We explored the latter phenotype through whole‐genome sequencing of 14 isolates, which unveiled a wide array of novel T6SS effector and (orphan) immunity proteins. By combining these in silico predictions with experimental validations, we showed that highly similar but non‐identical immunity proteins were insufficient to provide cross‐immunity among those wild strains.
Introduction
Diarrheal diseases can be caused by a variety of microorganisms, including the causative agent of cholera, Vibrio cholerae, which infects up to 4 million people every year (Ali et al., 2015). Cholera often spreads from its endemic area around the Ganges delta and has reached almost the entire world in the reported seven pandemics that have been witnessed since 1817. In addition to these pandemics, important localized outbreaks have occurred over the years, especially following natural disasters (Faruque et al., 1998; World Health Organization (WHO), 2006; Clemens et al., 2017).
V. cholerae strains can be classified into serogroups based on more than 200 different O‐antigens. The O1 serogroup is considered the primary cause of previous (e.g., classical serotype) and ongoing (e.g., El Tor serotype) cholera pandemics (Faruque et al., 1998; Cottingham et al., 2003). Isolates belonging to the O139 serogroup are genetically related to the seventh pandemic O1 El Tor strains (Johnson et al., 1994) but are rarely associated with disease outbreaks currently (Faruque et al., 1998; Clemens et al., 2017). Even though this species is best known due to the life‐threatening disease it causes, the vast majority of V. cholerae are common members of aquatic habitats. These mostly non‐O1/non‐O139 serogroup strains are thought to frequently associate with zooplankton and shellfish, and V. cholerae uses their chitinous exoskeletons as a source of carbon and nitrogen (Faruque et al., 1998; Cottingham et al., 2003; Kirn et al., 2005). Environmental isolates are typically considered harmless to humans despite reported associations with mild‐to‐severe forms of diarrhoea (Islam et al., 1992; Singh et al., 2001; Dziejman et al., 2002, 2005; Onifade et al., 2011; Deshayes et al., 2015; Hasan et al., 2015).
A major question, not only in the cholera field but also for infectious diseases in general, is how pathogenic isolates evolve from their non‐pathogenic environmental progenitors. In the case of V. cholerae, pandemic patient isolates are remarkably clonal, in sharp contrast to the high genomic variability encountered in environmental isolates (Faruque et al., 1998; Chun et al., 2009; Mutreja et al., 2011; Harris et al., 2012; Domman et al., 2017; Weill et al., 2017, 2019). Two major genetic features common to all pandemic strains are the CTXΦ prophage and the Vibrio pathogenicity island (VPI‐1 or TCP island). These genetic elements harbour genes encoding the main virulence factors, e.g., cholera toxin (CTX) and toxin‐coregulated pilus (TCP) (Taylor et al., 1987; Waldor and Mekalanos, 1996). The possession of these genomic regions, however, is not unique nor a ‘deterministic factor’ of pandemic strains. In fact, V. cholerae samples collected in several regions of the world have exposed environmental CTX‐ and/or TCP‐positive strains (Rivera et al., 2001; Faruque et al., 2003, 2004; Gennari et al., 2012; Bernardy et al., 2016; Shapiro et al., 2016). Additionally, V. cholerae strains that have caused localized cholera outbreaks without reaching pandemic levels have been reported, such as the O37 serogroup strains V52 and ATCC25872 (Aldova et al., 1968; Boyd and Waldor, 2002; Cottingham et al., 2003; Chun et al., 2009). Therefore, the terms ‘toxigenic’ (ability to cause cholera) and ‘pandemic’ (here, current seventh pandemic‐causing O1 El Tor strains) are not synonymous. What exactly determines whether strains become pandemic or not is still not fully understood and probably involves diverse aspects ranging from genetic content and strain‐specific phenotypes, along with the location of the initial outbreaks and ultimately towards social and sanitary factors, to name a few. Pertinent to the bacteria‐related aspects, previous studies hypothesized the existence of virulence adaptive polymorphisms (VAPs) circulating in environmental strains. These VAPs were suggested to be a prerequisite for pandemic transition before the horizontal acquisition of CTXΦ and VPI‐1 (Shapiro et al., 2016).
To better understand their emergence, the differences between pandemic strains and their environmental relatives need to be deciphered. Such differences most likely include phenotypic alterations that are not easily predictable by genomics. In the context of phenotypic variation, two minor virulence factors, namely, the type VI secretion system (T6SS) and the pore‐forming toxin haemolysin, are of special interest as they are differentially produced in pandemic compared with non‐pandemic toxigenic or environmental strains. The T6SS is a molecular killing device that resembles an inverted contractile bacteriophage tail and it is present in approximately 25% of all Gram‐negative bacteria (Ho et al., 2014; Cianfanelli et al., 2016; Galán and Waksman, 2018; Taylor et al., 2018). It is composed of a membrane‐spanning portion, a tube structure made of stacks of Hcp hexamer rings, and a tube‐surrounding sheath. The sheath is composed of the two proteins VipA and VipB which, upon contraction, propels the inner tube out of the cell together with its effector‐decorated tip proteins (VgrG and PAAR) (Zoued et al., 2014; Cherrak et al., 2019; Flaugnatti et al., 2020). Secreted Hcp therefore serves as an indicator of T6SS activity (Pukatzki et al. 2006; Basler et al., 2012; Bröms et al., 2013; Kube et al., 2014). The secreted effector proteins mostly target conserved cellular components, such as membranes, bacterial peptidoglycan, nucleic acids, or the eukaryotic cytoskeleton (Hood et al., 2010; Russell et al., 2014). T6SS effectors can be either secreted as (i) cargos that interact with T6SS structural proteins, such as Hcp or the tip protein VgrG; or as (ii) C‐terminal extensions of VgrG, Hcp, or PAAR (the so‐called ‘evolved’ proteins; Pukatzki et al., 2006, 2009; Hachani et al., 2016). Notably, T6SS‐producing bacteria protect themselves against their toxic effector repertoire by the production of effector‐cognate immunity proteins. These effector/immunity (E/I) pairs are usually encoded adjacent to each other (Russell et al., 2011; Brooks et al., 2013; Dong et al., 2013; Unterweger et al., 2014).
In current pandemic strains, the T6SS is encoded by four gene clusters: the large cluster and three auxiliary clusters. The large cluster primarily codes for structural proteins (recently reviewed by Crisan and Hammer, 2020), including the evolved tip protein VgrG3, which possesses a C‐terminal lysozyme‐like domain for peptidoglycan degradation (Zheng et al., 2011; Brooks et al., 2013). Auxiliary clusters 1 and 2 are both composed of genes encoding Hcp and VgrG (evolved in cluster 1 and structural in cluster 2), an adaptor protein (Tap1 or VasW; Liang et al., 2015; Unterweger et al., 2015) and an E/I module. The auxiliary cluster 1 effector TseL is a bifunctional lipase with anti‐bacterial and anti‐eukaryotic activity (Zheng et al., 2011; Dong et al., 2013; Russell et al., 2013). The auxiliary cluster 2 effector VasX acts as a pore‐forming toxin due to its bacterial/eukaryotic colicin‐like membrane‐disrupting activity (Miyata et al., 2011, 2013; Russell et al., 2014). Finally, the T6SS auxiliary cluster 3 is composed of genes encoding a second copy of PAAR (a tip‐sharpening protein that extends from VgrG; the first gene copy is at the start of the large cluster; Shneider et al., 2013) and a single E/I pair. When discovered, the aux 3 effector TseH was predicted to contain a hydrolase domain (Altindis et al., 2015), while the recently reported crystal structure supported its role as a papain‐like NlpC/P60 peptidase (Hersch et al., 2020) with structural similarity to the T6SS effector Tse1 of Pseudomonas aeruginosa that contains bacteriolytic peptidoglycan amidase activity (Chou et al., 2012).
Another toxic protein that shows differential activity in pandemic compared with non‐pandemic V. cholerae strains is the haemolysin protein (HlyA), which is likewise widespread among Vibrio species (Zhang and Austin, 2005). HlyA is considered a minor virulence factor as it contributes to toxicity in the context of intestinal infections (Ichinose et al., 1987; Olivier et al., 2007). We previously showed that the proper timing of HlyA activity is a prerequisite for pandemic V. cholerae to establish a replication niche within the aquatic amoeba Acanthamoeba castellanii, while constitutive activity kills this host prematurely (Van der Henst et al., 2018).
In this study, we deciphered phenotypic and genotypic differences between O1 El Tor pandemic strains and 15 environmental V. cholerae isolates. With respect to phenotypes, we focused primarily on T6SS‐ and haemolysin‐specific outcomes exerted on competing bacteria or amoebal predators. We show that clade‐specific anti‐amoebal toxicity is dependent on the actincross‐linking domain of a subset of T6SS effector proteins and that HlyA does not harm these predators under the tested conditions. Anti‐bacterial activity, on the other hand, is widespread among the environmental isolates and is based on their constitutive T6SS production, which is a major phenotypic difference compared with the exquisitely regulated T6SS of pandemic V. cholerae. We also performed long‐read PacBio‐based whole‐genome sequencing of the environmental isolates, which allowed us to perform basic comparative genomic analyses. Through this approach, we identified T6SS E/I modules as well as orphan immunity loci. Finally, we experimentally tested how the environmental isolates compete with one another and how this interbacterial competition correlates with their T6SS E/I repertoire.
Results and discussion
Genome sequencing of environmental V. cholerae strains
Apart from the major virulence factors, previous observations suggested important phenotypic differences between pandemic and environmental V. cholerae strains. We therefore decided to study 15 environmental isolates (Supporting Information Table S1) from diverse habitats along the central California coast, a region that is free of endemic or epidemic cholera. These strains were initially isolated in 2004 by Keymer and colleagues followed by basic characterization and comparative genome hybridization (CGH) analyses (Keymer et al., 2007; Miller et al., 2007). The latter approach was based on amplicon microarrays (representing 3357 of 3891 annotated open reading frames), which were designed using the first published V. cholerae genome sequence as a template, namely, pandemic O1 El Tor strain N16961 (Heidelberg et al., 2000). Based on the presence or absence of the microarray‐templated genes, the strains were classified into four clades (A–D) (Keymer et al., 2007; Miller et al., 2007), a classification we maintained throughout the current report. To better understand the accessory genome, including the T6SS E/I modules and to identify those genes that are novel when compared with strain N16961, we first whole‐genome sequenced these strains using a long‐read PacBio approach followed by the de novo assembly of their genomes. As a representative strain of the seventh cholera pandemic, we used strain O1 El Tor A1552 throughout this study (Yildiz and Schoolnik, 1998). This strain is connected to a cholera outbreak in Peru in the 1990s. We recently reported its genome sequence, including more than 1000 manual gene annotations, according to previous experimental validations (Matthey et al., 2018).
Supporting Information Table S2 shows the sequencing details and the features of the closed genomes of 15 environmental isolates. All genomes showed the dual chromosome architecture that is common for Vibrio species (Okada et al., 2005), a similar overall size of the two chromosomes, and average GC percentages within the same range as the one observed for the pandemic V. cholerae strains N16961 and A1552 (Heidelberg et al., 2000; Matthey et al., 2018). The assembly pipeline also predicted megaplasmids of ~300 kbp and 80 kbp for four strains belonging to clades C and D respectively. It should be noted that due to the size selection of the prepared sequencing libraries, putative smaller plasmids remained unidentified. Interestingly, we observed that the genomes of strains W6G and W7G were almost identical. This reflects the previous report by Keymer et al. In their original sampling study, the authors claimed that, based on CGH, 30 unique genotypes were identified within their collection of 41 environmental strains, while several genotypes were sampled multiple times from distinct sampling events (Keymer et al., 2007). Indeed, upon direct comparison of the W6G and W7G genomes, we observed pairwise identities of 99.98%, 99.997% and 99.998% for chromosome 1, chromosome 2, and the megaplasmid, respectively. Both strains possessed the same genes in all three replicons, and the few observed differences were primarily single nucleotides indels (which might, in part, reflect sequencing artifacts). These data suggest almost clonality between these two environmental samples (W6G and W7G). In addition, our whole‐genome sequencing data showed that strains E7G and SA7G of clade D also had high levels of identity (99.4%, 98.5%, and 99.9% pairwise identity of chromosomes 1, 2, and the megaplasmid, respectively), which again confirmed the previous CGH data (Keymer et al., 2007). Besides SNPs and indels, these strains differ by the presence/absence of small genomic islands on both chromosomes and an additional, second megaplasmid in strain SA7G, which is missing in E7G. The majority of other strains differed more significantly and therefore allowed us to test links between specific phenotypes and the corresponding genotypes.
Only a subset of environmental isolates block amoebal predation
We initially wondered how these environmental isolates would behave when confronted with predatory grazers, such as bacterivorous amoebae. We therefore tested the representative seventh pandemic strain A1552 and the above‐described collection of environmental strains for their ability to defend themselves against the grazing soil amoeba Dictyostelium discoideum. As shown in Fig. 1A, a clade‐specific behaviour was observable in which strains from clades A and B appeared as non‐toxic to D. discoideum and was efficiently grazed on while clades C and D strains completely resisted amoebal grazing. We concluded that anti‐amoebal defences significantly vary among the different environmental V. cholerae isolates.
Fig. 1.
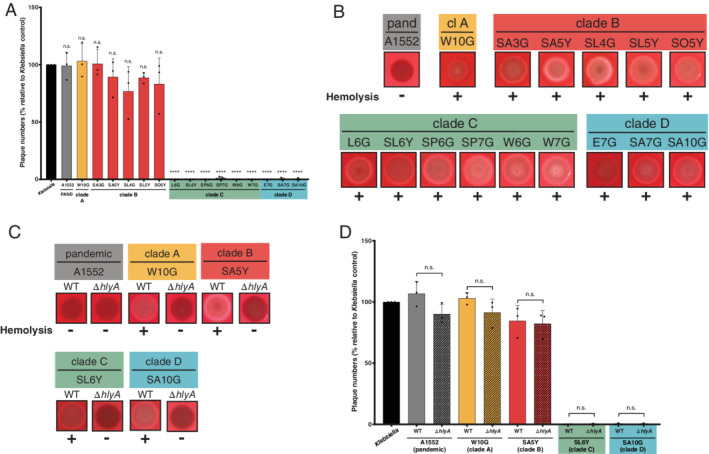
Defence against amoebal predation by a subset of V. cholerae strains.
A,D. Amoebal predation was scored using D. discoideum grazing assays in which formed plaques on bacterial lawns were enumerated. Plaque numbers are indicated relative to those formed on a lawn of K. pneumoniae, which served as a positive control. Bar plots represent the average of at least three independent biological replicates (±SD). Statistical significance is indicated (n.s., not significant; ****P < 0.0001).
B,C. Haemolytic activity was tested on blood agar plates. Pandemic V. cholerae strain A1552 as well as all environmental isolates (B) or a representative subset together with their respective hlyA‐minus derivatives (C) were assessed for haemolysis.
Enhanced haemolysin production in environmental V. cholerae does not contribute to their anti‐amoebal behaviour
As we observed that a subset of the environmental strains had a strong anti‐amoebal effect, we wondered whether this grazing inhibition was linked to toxic effectors of V. cholerae. Our group had previously demonstrated that the proper timing of the production or activity of the pore‐forming toxin haemolysin HlyA was essential for pandemic V. cholerae to form a replication niche inside the aquatic amoebae A. castellanii. Indeed, while pandemic V. cholerae exerted a tight regulation over this toxin and thereby successfully infected the amoebae's contractile vacuole, constitutive haemolysin activity by the environmental isolates killed the host prematurely, preventing the formation of this replication niche (Van der Henst et al., 2018). We speculated that haemolysin activity could also be involved in the resistance against D. discoideum grazing that we observed for the environmental strains from clades C and D. To follow‐up on this hypothesis, we first tested all environmental WT strains for haemolysis on blood agar plates. As shown in Fig. 1B, all isolates had strong haemolytic activity, especially when compared with the pandemic strain A1552. To ensure that the haemolysis was indeed caused by HlyA's activity, we interrupted the hlyA gene (loci comparable to locus tag VCA0219 in reference strain N16961) in a subset of the environmental strains through the integration of an antibiotic resistance cassette [the wild‐type (WT) versions of the environmental strains and their respective mutants are listed in the Supporting Information Table S1]. The selection of this subset of environmental isolates was thereby based on two criteria: (i) to represent each clade and (ii) to select those strains that showed efficient chitin‐induced natural transformability, which allowed for genetic manipulation of those strains. For these representative strains, we confirmed that the observed blood cell lysis was indeed caused by the specific haemolysin HlyA (Fig. 1C). After this confirmation, we tested the strains in the amoebal grazing assay. However, to our surprise, the haemolysin‐deficient mutants behaved the same way as their parental WT strains, indicating that HlyA is not responsible for the strong anti‐amoebal behaviour that we observed for the clade C and D strains (Fig. 1D). These results therefore suggest that, at least under the tested conditions, HlyA does not play an important role in anti‐amoebal grazing defence.
Environmental isolates constitutively produce T6SS
As HlyA activity was ruled out as a defence mechanism, we moved on to check the potential involvement of other molecular weapons. We next considered the T6SS, due to its widespread occurrence in Gram‐negative bacteria. Notably, V. cholerae's T6SS was initially discovered due to its anti‐eukaryotic activity that allowed the non‐pandemic V. cholerae strain V52 to avoid predation by D. discoideum (Pukatzki et al., 2006). Indeed, previous studies had indicated that, in contrast to the seventh pandemic strains, non‐pandemic toxigenic V. cholerae (such as the two O37 serogroup strains V52 and ATCC25872; Pukatzki et al., 2006; Basler et al., 2012; Van der Henst et al., 2018) as well as environmental isolates (Unterweger et al., 2012; Bernardy et al., 2016; Crisan and Hammer, 2020) maintain constitutive T6SS activity. This is in contrast to the silenced T6SS of pandemic strains, which are primarily induced by chitin or low c‐di‐GMP levels, concomitantly with natural competence and the production of extracellular enzymes, respectively (Borgeaud et al., 2015; Watve et al., 2015; Metzger and Blokesch, 2016; Metzger et al., 2016; Veening and Blokesch, 2017). To check whether the T6SS protects clade C and D strains, we first tested the general T6SS activity of the environmental isolates. As shown in Fig. 2A, we observed that the environmental strains efficiently eradicated Escherichia coli prey bacteria. Only strain SA3G of clade B reproducibly killed prey with a reduced efficiency, even though residual T6SS activity was still observed when compared with the non‐killing pandemic strain A1552 (Fig. 2A). However, these data alone do not unambiguously show whether the observed prey effacement was indeed T6SS‐dependent or was instead the result of any other modes of interbacterial competition, such as contact‐dependent inhibition, toxin secretion, bacteriocins, and so on (Hibbing et al., 2010; Stubbendieck and Straight, 2016; Granato et al., 2019). We therefore deleted the T6SS sheath protein‐encoding gene vipA in each of the clade‐representing strains and confirmed their lost T6SS activity by scoring for Hcp secretion. As illustrated in Fig. 2B, all strains were able to produce Hcp protein but only the T6SS‐active parental environmental strains were able to also secrete this protein into the supernatant. These data are therefore in agreement with the idea that the strains' T6SS is indeed constitutively active (at least under the tested conditions) and is the reason behind the observed interbacterial killing phenotype (Fig. 2A). We confirmed the latter idea by comparing the WT and vipA‐minus derivatives' killing ability in an interbacterial competition assay using E. coli as prey (Fig. 2C).
Fig. 2.
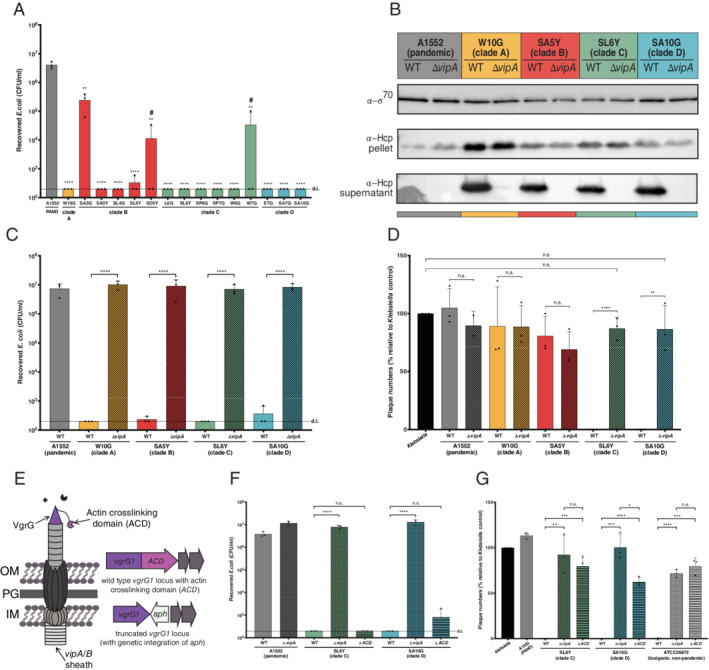
Constitutive T6SS activity linked to an ACD‐containing effector inhibits amoebal grazing.
A, C, F. Bacterial killing assays using E. coli as prey. Numbers of surviving prey are depicted on the Y‐axis (CFU ml−1). Statistical significance in panel A is shown above each strains' bar and calculated relative to the T6SS‐silent negative control strain A1552. #, for these strains, the killing activity was only reduced in one of the three independent experiments.
B. T6SS activity in representative environmental strains. Hcp detection in WT and ΔvipA mutants of representative environmental isolates. Intracellular (pellet) and secreted (supernatant) Hcp were assessed by immunoblotting using Hcp‐directed antibodies. Detection of σ70 served as a loading control.
D, G. T6SS‐ and ACD‐dependency of the anti‐amoebal defence. Plaque formation by D. discoideum on bacterial lawns formed by representative V. cholerae WT, ΔvipA derivatives (D and G) and ACD‐minus (G) strains. Details as in Fig. 1. The toxigenic non‐pandemic strain ATCC25872 and its site‐directed mutants served as control in panel G.
E. Simplified scheme of the T6SS. The actin cross‐linking domain (ACD) consists of a C‐terminal extension of the VgrG1 tip protein and this multidomain protein is encoded by the vgrG1 locus (shown on the right). Removal of the ACD‐encoding sequence was accomplished through site‐directed integration of a stop codon concomitantly with an aph selective marker. Bar plots in all panels represent the average of at least three independent biological replicates (±SD). d.l., detection limit. Statistical significance is indicated (n.s., not significant; *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001; for panel A, each sample was compared to the A1552 control).
Environmental strains use their VgrG‐linked ACD of the T6SS to fight amoebae
Having recognized that all environmental strains constitutively produce their T6SS, we moved on to assess the involvement of this machinery in the anti‐amoebal defence of clade C and D strains. Indeed, the observed inhibition of amoebal plaque formation by a subset of the environmental isolates was consistent with previous work by Unterweger and colleagues (Unterweger et al., 2012). These authors had studied four environmental isolates from estuaries of the Rio Grande delta for anti‐amoebal and anti‐bacterial activity and observed that two of these isolates could not resist amoebal predation. These strains were, however, also unable to kill E. coli prey, and the reason for this interbacterial non‐competitiveness was a frameshift mutation in the intermediate T6SS regulatory protein‐encoding gene vasH (Pukatzki et al., 2006; Unterweger et al., 2012). In contrast, apart from one exception (strain SA3G), all of the tested environmental isolates in our study efficiently eradicated E. coli prey (Fig. 2A), indicating that the T6SS was, in general, functional and active. We therefore reassessed the amoebal plaque formation against the genetically modified T6SS mutant strains. As shown in Fig. 2D, clade C and D isolates, whose parental WT strains completely blocked amoebal predation, became nontoxic when their T6SS was inactivated, indicating that their anti‐amoebal defence was indeed linked to the T6SS and uniquely caused by the latter.
Considering that all of the environmental isolates have a constitutively active T6SS under the tested conditions and efficiently killed other bacteria (Fig. 2A), we wondered why only the strains from clades C and D were able to use their T6SS as an anti‐eukaryotic defence tool. To answer this question and to also characterize the full E/I modules of these strains, we inspected the T6SS clusters in the new genomic sequencing data and observed a clear clade specificity. Only those strains belonging to clades C and D encoded evolved VgrG1 proteins with a C‐terminal actin cross‐linking domain (ACD) (Figs. 2 and 3; Supporting Information Table S3). Strains from clades A and B, on the other hand, encoded only structurally relevant VgrG1 proteins without an evolved effector domain. Importantly, pandemic strains also encoded such an ACD as part of vgrG1, but, as noted above, these strains do not produce functional T6SSs without specific environmental cues and therefore show neither anti‐bacterial (Fig. 2A) nor anti‐amoebal behaviour (Fig. 2D) under the tested conditions.
Fig. 3.
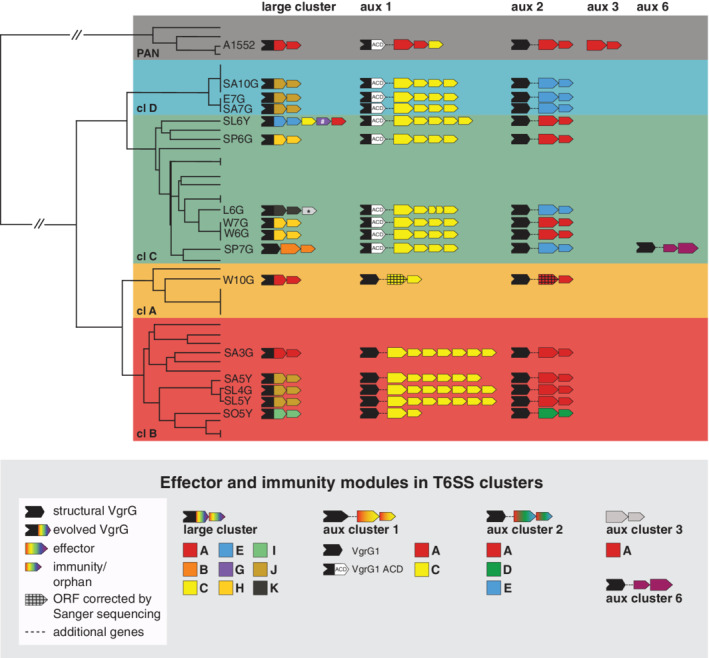
T6SS effector/immunity typing scheme of pandemic and environmental V. cholerae. The previously published phylogenetic tree was freely adapted from Keymer et al. 2007 (not fully to scale). The clade colour code is depicted in the background. The E/I type of each T6SS cluster (large cluster as well as auxiliary (aux) clusters 1, 2, 3 and 6) are schematized for each strain. Large black arrows symbolize vgrG genes, which were classified as structural (black) or evolved (coloured tips, to represent the different types of C‐terminal effector domains). Large and small coloured arrows represent effector and immunity genes, respectively, according to the colour code indicated in the legend below the scheme. Immunity genes that are not adjacent to a putative effector gene are considered as orphan immunity loci. * depicts an orphan immunity gene from strain L6G whose gene product was slightly below the amino acid identity threshold of 30% (20.3%) relative to C‐type immunity proteins. # depicts an orphan immunity gene that is a homolog to bona fide immunity genes in other genomes. Effector genes in auxiliary clusters 1 and 2 of strain W10G (marked in figure with a pattern) were wrongly annotated in the PacBio genome sequence (containing several stop codons) due to a frameshift sequencing artefact; this sequencing error was corrected by Sanger sequencing.
The ACD of VgrG1 in non‐pandemic but toxigenic O37 serogroup strain V52 (Chun et al., 2009), which produces its T6SS constitutively, was previously shown to be involved in V. cholerae's toxicity towards D. discoideum and macrophages (Pukatzki et al., 2006, 2007; Ma et al., 2009). Moreover, this VgrG1‐ACD was also responsible for intestinal inflammation and cholera toxin‐independent fluid accumulation in an infant mouse model of infection (Ma and Mekalanos, 2010). Furthermore, the VgrG1‐ACD of the seventh pandemic V. cholerae strain C6706 was implicated in alternating intestinal peristalsis of zebrafish larvae, leading to the expulsion of preinoculated commensal bacteria (Logan et al., 2018). However, this effect was only observable upon constitutive T6SS expression using a genetically engineered derivative of this pandemic strain in which T6SS production occurred based on artificial expression of the gene encoding the quorum sensing‐ and chitin‐linked transcription factor QstR (Lo Scrudato and Blokesch, 2013; Borgeaud et al., 2015; Watve et al., 2015; Jaskólska et al., 2018; Logan et al., 2018).
Given this previous work on toxigenic strains, we tested whether the ACD of the clades C and D environmental isolates was likewise causative of the observed anti‐amoebal response. To do so, we first generated truncated versions of VgrG1 that lacked the evolved ACD domain‐containing C‐terminus (Fig. 2E). Importantly, these vgrG1ΔACD strains maintained their full anti‐bacterial competitiveness (Fig. 2F), which indicates that the ACD deletion did not impact the general assembly and/or activity of the T6SS machinery. However, as shown in Fig. 2G, the amoebal grazing ability was restored on the lawns formed by the two vgrG1ΔACD environmental V. cholerae strains and resulted in equal (e.g., for strain SL6Y) or intermediate (e.g., strain SA10G) amoebal plaque numbers compared with those numbers observed for complete T6SS‐defective mutants. The non‐pandemic but toxigenic control strain ATCC25872 (Supporting Information Table S1; Aldova et al., 1968), which, like strain V52, is constitutive T6SS active (Van der Henst et al., 2018), also lost its anti‐amoebal activity in the absence of the T6SS core structure or when the ACD of VgrG1 was missing (Fig. 2G). We therefore concluded that some environmental V. cholerae might have evolved ACD‐extended VgrG‐encoding genes, as the ACD's anti‐eukaryotic activity protects them from environmental grazers. Whether pandemic strains subsequently horizontally acquired the ACD encoding region as previously suggested (Kirchberger et al., 2017), or whether the last common ancestor between the pandemic and environmental lineages already contained this specialty that was later then lost from some wild strains, is currently unclear. We did observe, however, that the VgrG1‐ACD protein is highly conserved between the pandemic strain A1552 and the environmental isolates, regardless of their clade (Supporting Information Table S4). Indeed, while the full length VgrG1‐ACD protein showed >97% identity when compared among these strains, the ACD part alone showed identity levels above 99% with several strains sharing 100% identical ACD domains (Supporting Information Table S4).
Diversity of T6SS effectors in environmental V. cholerae isolates
As briefly mentioned in the previous section, the ACD‐minus mutant of the clade D strain SA10G showed reduced but still significant residual T6SS‐dependent anti‐amoebal activity. We therefore wondered whether this could be explained by the presence of another anti‐eukaryotic effector in one or several of the T6SS clusters. Furthermore, we were also interested in characterizing the full set of E/I modules in these strains, which would allow us to speculate about the strains' competitive potential against one another. Using the newly assembled genomic data together with the previously reported genome sequence of clade B strain SA5Y (Matthey et al., 2018;Matthey et al., 2019) we determined the E/I modules of these 15 environmental strains and predicted their function based on BlastP analyses (Fig. 3 and Supporting Information Table S3). Moreover, to arrange the E/I pairs into putative compatible groups, we defined their modules based on a percentage amino‐acid identity of at least 30%, which is a typing approach previously applied (Unterweger et al., 2014; Kirchberger et al., 2017). Interestingly, we found evidence for several orphan immunity genes, meaning immunity genes that no longer coexisted with and were adjacent to a cognate effector‐encoding gene, as had been previously reported by Kirchberger and colleagues (Kirchberger et al., 2017). These orphan genes were located in the 3' regions of the T6SS clusters and were likewise classified according to their module type (Fig. 3 and Supporting Information Table S3). The characterization of the E/I modules and orphan immunity loci was restricted to the well‐characterized T6SS clusters of V. cholerae (e.g., the large cluster as well as auxiliary clusters 1 and 2), as the reported auxiliary cluster 3 (E/I pair TseH/TseI; Altindis et al., 2015) was absent from all environmental isolates. This finding is consistent with a preprinted study showing that this auxiliary cluster represents a pandemic strain‐associated mobile genetic element (Santoriello et al., 2019) (Fig. 3 and Supporting Information Table S3). Moreover, the recently characterized auxiliary clusters 4 and 5 were also absent from the environmental isolates (Labbate et al., 2016; Crisan et al., 2019). Instead, automatic annotations followed by a manual inspection identified the presence of a novel T6SS cluster in strain SP7G (clade C), which we defined as auxiliary cluster 6 (Figs. 3 and 4, and Supporting Information Table S3). This cluster resides in the small chromosome 2 of strain SP7G and the gene order resembles that of auxiliary clusters 1 and 2, namely: hcp, vgrG, a putative adaptor‐protein encoding gene (coding for a DUF4123 domain as described for Tap1 and VasW; Liang et al., 2015; Unterweger et al., 2015), a gene of unknown function and a noncanonically ordered putative immunity/effector (I/E) pair, located 250 bp downstream of the previous gene (Fig. 4A). BlastP analysis identified the predicted effector as a DUF2235‐containing protein, similar to the effector protein encoded in auxiliary cluster 1. However, an alignment of the sequences of these two effector proteins (encoded in aux 1 or aux 6 of strain SP7G) showed only a 18% identity. Importantly, this cluster is located inside a larger genomic island. Our search in the PATRIC nucleotide sequence database (Wattam et al., 2017) suggested that this aux 6 cluster (and the genomic island) is prominently represented in several V. cholerae strains, including 2013 environmental isolates from Bangladesh and an O35 strain (1311–69) isolated in 1969 from a patient in India (Bishop‐Lilly et al., 2014). Our experimental investigation showed that the auxiliary cluster 6 is active under laboratory conditions, as strain SP7G was able to kill its own kin when the latter lacked the region that included the putative I/E gene pair (Fig. 4B). Furthermore, SP7G lost some of its killing potential against E. coli when the I/E‐including region of the auxiliary cluster 6 was missing (Fig. 4C).
Fig. 4.
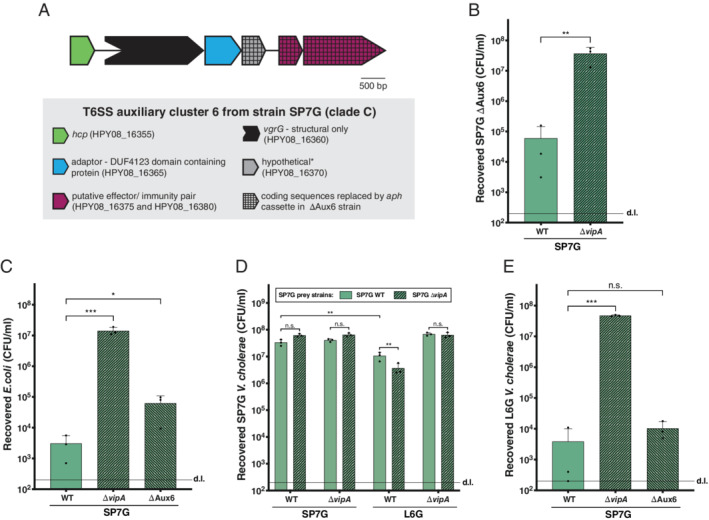
The T6SS auxiliary cluster 6 of strain SP7G is active.
A. The 6.4‐kb long auxiliary T6SS cluster 6 carried on the chromosome 2 of strain SP7G (locus tags HPY08_16355 to HPY08_16380 in CP053809) is depicted. Gene symbols are explained below the scheme. *Gene HPY08_16370 was annotated (PGAP method from NCBI) beyond internal stop codons, which were confirmed by Sanger sequencing (the initial Prokka annotation predicted two separate ORFs). The region spanning HPY08_16370 to HPY08_16380 was replaced by an aph cassette to test for aux 6 activity.
B–E. Bacterial killing assays using SP7GΔAux6 (B), E. coli (C), SP7G WT and ΔvipA (D), or L6G (E) as prey. Numbers of surviving prey are depicted on the Y‐axis (CFU ml−1). Bar plots represent the average of three independent biological replicates (±SD). Statistical significance is indicated (n.s., not significant; *P < 0.05; **P < 0.01; ***P < 0.001).
In the large T6SS cluster, pandemic V. cholerae strains, such as A1552 carry an A‐type E/I module (Fig. 3 and Supporting Information Table S3), for which, in fact, the peptidoglycan cell wall degradation effector corresponds to the C‐terminal domain of the evolved VgrG3 protein (Zheng et al., 2011; Brooks et al., 2013). When analysing this locus in the environmental V. cholerae isolates, we noticed that the evolved nature of VgrG3 was conserved among these strains, except for clade C strain SP7G in which VgrG3 is solely a structural T6SS component that is followed by a B‐type E/I module. How this effector is attached to the VgrG tip protein is, however, unclear, as no adaptor protein such as those encoded by tap1 and vasW could be identified. This B‐type effector is predicted to have a cellular adhesion function (Unterweger et al., 2014). Interestingly, strains W10G (clade A) and SA3G (clade B) contain a pandemic‐like A‐type E/I module (Fig. 3), while the other strains carried a wide variety of E/I pairs, as described in detail in the Supporting Information Table S3. Moreover, even though the amino acid identity of these effectors is below 30% when comparing different types, most of them have a common predicted function, namely, peptidoglycan degradation (Supporting Information Table S3). Therefore, with the exception of strain SP7G, all the environmental isolates have a dedicated anti‐bacterial E/I module in the T6SS large cluster (Fig. 3 and Supporting Information Table S3). Interestingly, pairwise comparisons of the effector and immunity protein sequences among different strains showed that strains harbouring E/I modules from the same family had 100% immunity identity (Supporting Information Table S5). Given the diversity of E/I modules in this cluster when comparing all strains, the complete identity of immunity proteins from the same family could indicate recent acquisition by horizontal gene transfer. Furthermore, L6G and SL6Y (clade C) are the only strains that also harbour one or several orphan immunity loci after the E/I module in this large T6SS cluster (Fig. 3).
These orphan immunity loci are distinct in their type when compared with the current resident E/I modules. When we searched the PATRIC translated nucleotide sequence database (Wattam et al., 2017) using the protein sequence from these orphan loci as the query, we noticed that these genes are only found as orphan loci in other genomes (i.e., they were located downstream of another E/I pair). The only exception was the second orphan locus in strain SL6Y, which encodes a G‐type immunity protein (marked by ‘#’ in Fig. 3). The predicted protein showed 98.4% identity to true immunity proteins (e.g., those encoded directly adjacent to an effector gene) from several V. cholerae strains. Among those was strain 2633‐78, an O1 CTX‐negative isolate collected from sewage in Brazil in 1978. Interestingly, this strain was experimentally tested in a previous study, where it was shown to have an active T6SS (Bernardy et al., 2016).
The auxiliary cluster 1 contains the structural or evolved (e.g., encoding C‐terminal ACD) vgrG1 gene, as mentioned above (Figs. 2 and 3). In addition, in the case of pandemic V. cholerae, this cluster harbours an A‐type E/I module encoding the lipase effector TseL with anti‐bacterial and anti‐eukaryotic activity (Zheng et al., 2011; Dong et al., 2013; Russell et al., 2013) followed by a C‐type orphan immunity gene (Kirchberger et al., 2017). Interestingly, all of the 15 environmental strains harbour C‐type E/I modules as part of this auxiliary cluster 1, even though there is considerable polymorphism in the effector and immunity proteins (Fig. 3 and Supporting Information Table S6). C‐type effectors have a predicted alpha/beta hydrolase domain (DUF2235), which has been previously associated with T6SS phospholipases from E. coli and Pseudomonas aeruginosa (Russell et al., 2013; Flaugnatti et al., 2016; Crisan et al., 2019). Furthermore, with the exception of strains W10G (clade A) and SO5Y (clade B), all environmental isolates also carry between two and five orphan immunity loci downstream of the E/I pair (Fig. 3). Importantly, even though these orphan loci are C‐type as the bona fide E/I pair, they do not appear as duplications of the resident immunity gene, as they only share 60%–85% identity with the latter. Considering the wide variety of E/I types in the large cluster as well as in auxiliary cluster 2 (see below), the apparent conservation of C‐type E/I modules in the auxiliary cluster 1 of these strains, in addition to the many C‐type orphan loci (as also observed in the in silico study by Kirchberger et al. 2017) is quite intriguing and supports the idea that the C‐type orphan immunity gene in the pandemic strains might be a remnant of a previous C‐type E/I module of the strains' progenitor.
In pandemic V. cholerae, the auxiliary cluster 2 carries an A‐type E/I module where the VasX effector is a pore‐forming toxin (Miyata et al., 2011; Miyata et al., 2013; Russell et al., 2014). Our comparative genomic analyses showed that several of the environmental isolates likewise encode VasX‐like effectors (A‐type) at the same locus (Fig. 3 and Supporting Information Tables S3 and S7), while other strains carry D‐ and E‐type E/I modules. While D‐type effectors have been predicted to foster peptidoglycan degradation, E‐type effectors are predicted to form pores, like VasX (Unterweger et al., 2014). We therefore reasoned that the residual ACD‐independent T6SS‐dependent anti‐amoebal impact observed for strain SA10G (clade D) might be caused by this E‐type effector from auxiliary cluster 2, especially as the latter is missing from clade C strain SL6Y in which removal of the ACD from VgrG1 was almost equivalent to a complete T6SS inactivation (Fig. 2G). Interestingly, our analyses suggest that these E‐type effectors also contain a common peptidoglycan‐binding domain (e.g., an N‐terminal Lysin Motif; Buist et al., 2008), which might render them bifunctional against bacteria and eukaryotes (Fig. 3 and Supporting Information Table S3). Future work will address the exact characteristics of the putative E/I modules in more detail.
Competition among environmental strains occurs in the presence of non‐identical E/I modules
Previous studies showed that a plethora of T6SS‐transported effectors have active anti‐bacterial purposes and that cognate immunity proteins are required to protect the producer or its siblings from intoxication (Dong et al., 2013; Durand et al., 2014; Russell et al., 2014; Unterweger et al., 2014). Strains with matching E/I modules could therefore coexist in the same environment (Unterweger et al., 2014), while competitive strains might clear a niche and propagate inside this niche in a clonal manner (McNally et al., 2017; Speare et al., 2018). Importantly, as these E/I modules seem to move horizontally in an as yet uncharacterized manner (Unterweger et al., 2014; Salomon et al., 2015), the level of compatibility between strains will not follow their phylogenetic relatedness. We therefore sought to experimentally probe the strains' compatibility or competitiveness within this collection of environmental isolates. Indeed, while we scored T6SS activity against a laboratory strain of E. coli (see Fig. 2), we wondered how the V. cholerae strains would behave when exposed to each other. Consistent with the fact that the E/I modules carried by the environmental strains are considerably different from those of pandemic V. cholerae (Fig. 3 and Supporting Information Table S3), we experimentally demonstrated that these bacteria efficiently outcompeted the T6SS‐silent pandemic strain A1552 (Fig. 5A). Next, we tested interbacterial competition between the clade‐representative strains in an assay in which the prey strains had their T6SS inactivated. As shown in Fig. 5B, reciprocal killing of the T6SS‐positive parental strains occurred and only kin strains were immune to the toxic assaults. However, as the E/I module types seemed more similar within clades than across clades, we extended our analyses and tested all pairwise combinations (this time, with both partners T6SS positive). The underlying rationale was that previous studies had identified different types of E/I modules in silico (Unterweger et al., 2014; Kirchberger et al., 2017; Crisan et al., 2019) but most pairwise competition assays were primarily linked to patient isolates and not to a larger collection of environmental isolates. Indeed, as described above and shown in Fig. 3, our in silico prediction showed that strains within clades often contained similar E/I module types (Supporting Information Tables S5–S7), even though the amino acid identity threshold for this categorization is rather low (30% as previously defined; Unterweger et al., 2014; Kirchberger et al., 2017).
Fig. 5.
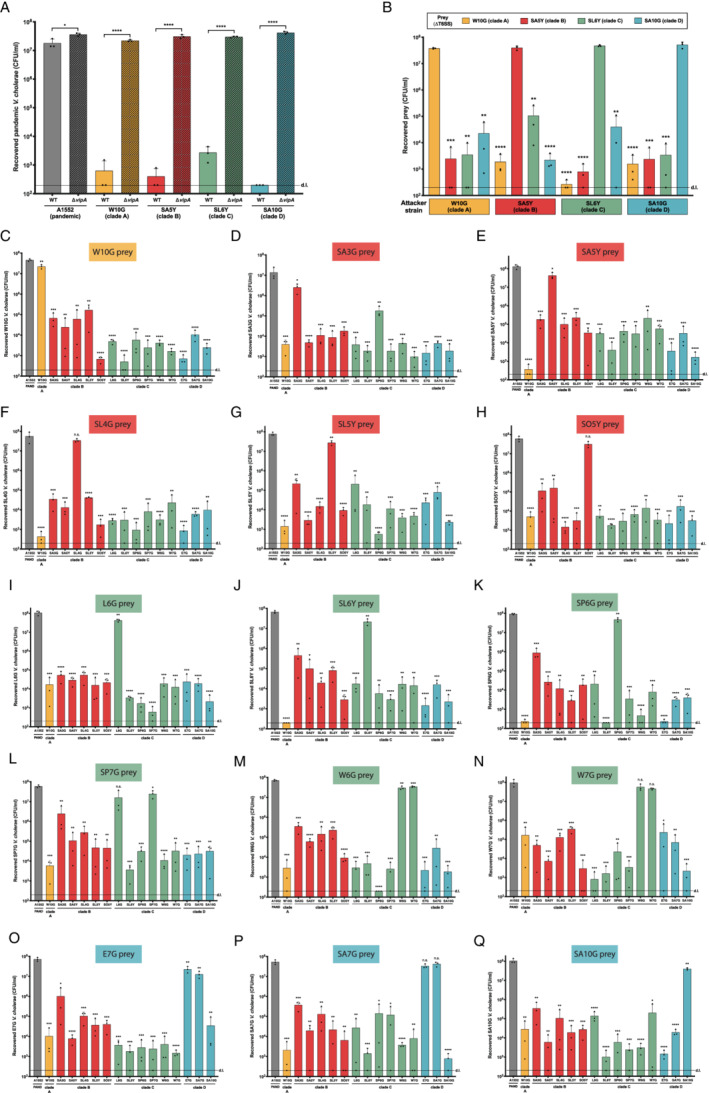
Competitiveness among environmental strains. Graphs in all panels represent bacterial killing assays as described in Fig. 2 using the predator strains as indicated on the X‐axes and as prey: (A) the pandemic V. cholerae strain A1552; (B) representative vipA‐minus (ΔT6SS) mutants of the environmental isolates as shown above the graph; or (C to Q) each environmental isolate as shown in the graph title for each panel and on the Y‐axis. Plots represent the average of three independent biological replicates (±SD). d.l., detection limit. Statistical significance is indicated (n.s., not significant; *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001).
As shown in Fig. 5, we observed interesting and complex phenotypes in these extensive pairwise killing experiments. For instance, strains W6G and W7G from clade C were fully protected against each other's attacks (Fig. 5 panels M and N). This finding is consistent with the 100% identity of all three T6SS clusters, including the orphan genes from aux cluster 1 (Supporting Information Tables S5–S7), and their overall near clonality as described above. Clade D strains have the same E/I module types in all T6SS clusters. However, upon closer inspection of their E/I protein sequences, we can observe a 100% sequence identity among the three strains only in the large and aux 1 cluster‐encoded proteins. For the aux 2 cluster, the encoded E/I proteins from strains E7G and SA7G are 100% identical, while the E/I pair carried by strain SA10G has only 95%/73% identity when compared with the other two strains' E/I pair (Supporting Information Tables S5–S7). Consequently, and as shown in Fig. 5 (panel O to Q), strains E7G and SA7G are fully compatible with each other, while strain SA10G can kill and be killed by the other two strains. We therefore concluded that the identity level in only one of the T6SS cluster‐encoded immunity proteins causes the competitiveness among these strains.
Surprisingly, clade B strains SA5Y and SL4G have 100% identity in all three T6SS cluster‐encoded immunity proteins (Supporting Information Tables S5–S7) but are still able to kill each other with considerable efficiency (Fig. 5, panels E and F). Why this is the case is currently unclear. However, we speculate that expression or immunity protein production might be impaired in those strains or that additional T6SS E/I modules are hidden in the strains' genomes, which were not easily identifiable based on a lack of the hallmark genes paar, hcp and vgrG in their vicinity.
Clade A strain W10G carries pandemic‐like A‐type E/I modules in the large and aux2 clusters (Fig. 3). Interestingly, this strain can kill many of the other environmental strains remarkably well, such as clade B strains SA5Y, SL4G and SL5Y, and clade C strains SL6Y and SP6G. Notably, the T6SS active toxigenic strain V52 (which harbours A‐type E/I modules in all T6SS clusters) was previously shown to outcompete strains carrying different combinations of E/I modules (Unterweger et al., 2014). Indeed, the AAA (A‐type in all clusters) E/I modules is conserved in pandemic V. cholerae, even in those strains that caused former pandemics (e.g., sixth pandemic O1 classical strains), as well as in non‐pandemic but toxigenic isolates. It was therefore speculated that this combination might be advantageous in a disease context (Unterweger et al., 2014). Clade B strain SA3G also contains A‐type E/I modules in the large and aux 2 clusters, but it does not kill other strains at the same level as strain W10G. However, as noted above, this strain is even less efficient against laboratory E. coli prey strains (Fig. 2A), which could mask its full effector toxicity potential.
Finally, a very interesting pairwise comparison is that of clade C strains SP7G and L6G. While L6G as a prey is very efficiently eliminated by strain SP7G (Fig. 5, panel I), SP7G prey seemed almost resistant to L6G intoxication (Fig. 5, panel L; please note, however, the commonly witnessed inter‐experimental variation). Importantly, the observed phenotype was only mildly dependent on SP7G's own T6SS activity. As shown in Fig. 4D, L6G is able to kill T6SS‐deficient SP7G as a prey roughly fivefold more efficiently compared with its T6SS‐positive parental strain. These two strains contain E/I sets of different families in the large cluster and show 78% and 99.7% identity in the immunity proteins encoded in aux cluster 1 and 2 respectively. Based on these differences, we would expect that these strains fully compete with each other; however, killing almost exclusively occurs with SP7G as the predator. We therefore speculate that the large cluster‐encoded K‐type effector domains of strain L6G's evolved VgrG3 as well as SP7G's noncanonical structural VgrG3 and its adjacently encoded putative effector protein might not be functional peptidoglycan destruction enzymes and therefore not necessarily active in interbacterial competition. Why a low level of protein identity in the aux cluster 2‐encoded immunity protein is sufficient to protect strain SP7G from L6G assaults is currently unclear. It is also tempting to speculate that the vice versa interaction leads to L6G killing due to the additional auxiliary cluster 6 that is carried by strain SP7G. Upon experimental testing of this idea, we only observed a mild role for this additional auxiliary cluster with the SP7GΔAux6 strain showing slightly lower L6G killing activity compared to the SP7G WT parental strain (Fig. 4E). Due to the overall inter‐experimental variation, this difference was, however, not statistically significant. Further work is therefore necessary in order to delve deeper into these observed phenotypes.
Overall, closer inspection of all of the pairwise killing data attests to the complexity behind the T6SS compatibility code. As mentioned previously, even though some strains might harbour the same E/I families in the T6SS clusters, pairwise comparisons of these proteins show that quite frequently their identity is not 100% (Supporting Information Tables S5–S7), which appears to be necessary to allow coexistence (also observed in the study by Speare et al., 2018). Our data therefore support what Unterweger and colleagues (Unterweger et al., 2014) initially speculated, namely that the compatibility between strains seems to follow the level of polymorphism of their immunity proteins and that diversity in only one cluster‐encoded protein is sufficient to drive competition.
Conclusion
In this study, we investigated two minor virulence factors, the pore forming haemolysin and the T6SS, in a set of 15 environmental V. cholerae strains. We assessed the relevance of these molecular weapons as defence mechanisms against amoebal predation (haemolysin and T6SS) and in the context of bacterial warfare (T6SS only). We showed that all of these environmental isolates possess a constitutively active T6SS and are able to use the machinery as a bacterial killing device. In contrast, only a subset of these strains was able to efficiently suppress grazing by D. discoideum amoebae, a phenotype that was dependent on the eukaryote‐specific ACD of the evolved VgrG1 T6SS effector. Careful in silico identification unveiled an extensive T6SS repertoire of E/I pairs and orphan immunity loci. Consistent with this finding, we observed extensive interbacterial competition under pairwise co‐culture conditions whereby mutual compatibility was rarely achieved. Importantly, our study also confirmed that both molecular weapons, the T6SS and the haemolysin toxin, are constitutively active in the environmental isolates. It is expected that these V. cholerae strains constantly compete with other microorganisms for nutrients and space within the marine environment besides being under persistent predation pressure from bacterivorous grazers. It seems therefore reasonable to assume that keeping their molecular weapons constitutively active might provide them with a competitive advantage in this natural habitat. Importantly, this is in sharp contrast to the tight regulation of these machineries in the well‐studied pandemic patient isolates. Future work is therefore required to decipher how this differential production pattern is achieved in pandemic versus non‐pandemic strains and whether this tight regulatory control might provide specific benefits to the former strains.
Experimental procedures
Bacterial strains and growth conditions
The bacterial strains (V. cholerae, E. coli and K. pneumoniae) used in this study are listed in the Supporting Information Table S1. Unless otherwise stated, all strains were grown aerobically in Lysogeny broth (LB; 10 g l−1 of tryptone, 5 g l−1 of yeast extract, 10 g l−1 of sodium chloride; Carl Roth) or on LB agar plates at 30°C. Half‐concentrated defined artificial seawater medium (0.5 × DASW) containing HEPES (4‐(2‐hydroxyethyl)‐1‐piperazineethanesulfonic acid) and vitamins (Meibom et al., 2005) was used for growth on chitinous surfaces for strain construction based on chitin‐induced natural transformation (see below).
D. discoideum amoebae (strain Ax2 Ka) were cultured in HL5 medium supplemented with glucose (Formedium, UK). For amoebal grazing assays (e.g., plaque formation assays; see below), SM/5 medium (final concentrations: 2 g l−1 of glucose, 2 g l−1 of bacto peptone, 2 g l−1 of yeast extract, 0.2 g l−1 of MgSO4 7H2O, 1.9 g l−1 of KH2PO4, 1 g l−1 of K2HPO4; pH 6.4), was mixed with 2% agar to prepare SM/5 plates (20 ml/plate) (Sussman, 1987). CaCl2 (50 μM)‐supplemented Sörensen's buffer (8 g/4 l of KH2PO4, 1.16 g/4 l of Na2HPO4; pH 6; Gerisch et al., 1967) was used as the washing and resuspension buffer for the amoebae and the bacteria that were used in the plaque formation assay.
The following antibiotics were added if required at the given concentration: kanamycin (75 μg ml−1), rifampicin (100 μg ml−1), streptomycin (100 μg ml−1) and chloramphenicol (2.5 μg ml−1).
Genetic engineering
V. cholerae strains were genetically modified using chitin‐induced transformation as previously described (Marvig and Blokesch, 2010; De Souza Silva and Blokesch, 2010; Blokesch, 2012; Borgeaud and Blokesch, 2013). This method relies on natural transformation triggered by growth on chitin followed by the addition of a PCR fragment that carried the desired genetic change. To achieve higher numbers of transformants, the protocol was slightly modified. The PCR fragments were added twice (24 h and 36 h after bacterial inoculation on the chitin flakes) and cells were enriched in 2 × YT medium (Carl Roth) before selective plating. PCR amplifications were conducted using Pwo (Roche) and GoTaq (Promega) polymerases according to the suppliers' recommendations. Following initial screening by PCR (using bacterial cells as the templates), genetically engineered loci were verified by Sanger sequencing (Microsynth, Switzerland).
The rifampicin‐sensitive V. cholerae strain A1552‐RifS was generated by a combination of natural cotransformation (Dalia et al., 2014) and our previously described counterselectable Trans2 approach (Van der Henst et al., 2018). To this end, a 4 kb PCR fragment was amplified, harbouring a mutation, which restored the native rpoB‐encoded protein (F531S substitution) from its mutated version in the parental strain A1552 (RpoB[S531F]; Matthey et al., 2018). V. cholerae A1552 was then cotransformed with this fragment and another 3924 bp fragment containing flanking regions matching lacZ and two selective markers (aph and pheS*; Supporting Information Table S1). Transformants were selected on kanamycin‐containing agar plates and the lost rifampicin resistance was scored based on replica plating on plates ±rifampicin using a velvet cloth. A second round of natural transformation followed to restore the lacZ gene by adding a WT lacZ PCR fragment to chitin‐grown cells followed by a counter selection of the pheS* allele on 4‐chloro‐phenylalanine (20 mM)‐containing agar plates as previously described (Van der Henst et al., 2018). To confirm the restoration of native rpoB, genomic DNA of A1552 RifS was isolated and the PCR‐amplified rpoB gene was Sanger‐sequenced.
Amoebal grazing assay
To determine the predatory capacity of D. discoideum on bacterial lawns of V. cholerae, plaque formation was scored following a previously described protocol (Pukatzki et al., 2006) with minor modifications. Briefly, bacteria were cultured overnight in LB medium at 30°C and harvested by centrifugation. The cell pellet was washed and resuspended in SorC buffer (Gerisch et al., 1967) to reach an optical density at 600 nm (OD600) of 5.5 in a final volume of 400 μl. Cultured D. discoideum amoebae (in HL5 with glucose; Formedium, UK) were detached from culture dishes using cell scraper (SPL Life Sciences) and collected by centrifugation (3 min 1000 rcf), resuspended in SorC buffer, and enumerated in a KOVA counting chamber (KOVA International, USA). The amoebal concentration was adjusted to 2 × 104 cells ml−1 and 20 μl of this suspension (corresponding to ~400 amoebal cells) was mixed with the 400 μl of bacterial suspension. The mixture was gently spread on two parallel SM/5 plates using a plastic rake (VWR), resulting in technical replicates. The plates were wrapped in aluminium foil and incubated at 24°C for 5 days. After this incubation period, D. discoideum plaque numbers were enumerated. As a positive control, we included a frequently used nonencapsulated Klebsiella strain (Benghezal et al., 2006) for which the resulting plaque numbers were set to 100%. Three biologically independent experiments were performed. The individual experimental data points (mean of technical replicates) as well as the overall average of the independent experiments (± standard deviation) are shown in each graph. A two‐tailed Student's t‐test was performed to determine statistical significance.
Haemolysin activity
The haemolytic activity of V. cholerae was assayed using trypticase soy agar containing 5% sheep blood (BD, Heidelberg, Germany). To do so, the respective overnight cultures were spotted (2 μl) onto the plates and incubated at 30°C for 24 h, after which pictures of the plates were taken.
Interbacterial killing assays
Bacterial killing was assessed following a previously established assay with minor modifications (Borgeaud et al., 2015). The prey cells (E. coli or V. cholerae, as indicated) and the respective predator bacteria were mixed at a ratio of 1:10 and spotted onto paper filters on prewarmed LB agar plates. After 4 h of incubation at 37°C, the bacteria were resuspended, serially diluted, and spotted onto antibiotic‐containing (rifampicin or streptomycin) LB agar plates to enumerate the colony‐forming units (shown as CFU ml−1). The majority of these killing experiments were performed using exponentially growing V. cholerae (OD600 ~ 1). For the pairwise killing experiments of all environmental strains, OD600‐adjusted overnight cultures were used, which resulted in biologically similar outcomes to the samples derived from exponentially growing cultures. Statistically significant differences were determined on log‐transformed data (Keene, 1995) by a two‐tailed Student's t‐test of three biologically independent replicates. If no prey bacteria were recovered, the value was set to the detection limit to allow for statistical analysis.
SDS‐PAGE and western blotting
To check the production of the Hcp protein, cell lysates were prepared as described previously (Metzger et al., 2016). In brief, exponentially growing bacteria (~3 h of growth after a 1:100 back dilution from overnight cultures) were pelleted and then resuspended in Laemmli buffer, adjusting for the total number of bacteria according to the cultures' OD600 values. To check for T6SS‐secreted Hcp, 1.5 ml of the culture supernatant was filter sterilized (0.2‐μm filter; VWR) and the proteins were precipitated using trichloroacetic acid (TCA). The precipitated proteins were washed with acetone before being resuspended in 30 μl of Laemmli buffer. All samples were heated at 95°C for 15 min.
Proteins were separated by sodium dodecyl sulfate (SDS)‐polyacrylamide gel electrophoresis (PAGE) using 15% gels and then western blotted as previously described (Lo Scrudato and Blokesch, 2012). Primary antibodies against Hcp (Eurogentec; Metzger et al., 2016) were used at 1:5000 dilution while anti‐Sigma70‐HRP antibodies (BioLegend, USA distributed via Brunschwig, Switzerland) were diluted 1:10 000 and served as a loading control. Goat anti‐rabbit horseradish peroxidase (HRP) (diluted at 1:20 000; Sigma‐Aldrich, Switzerland) was used as the secondary antibody against the anti‐Hcp primary antibody. Lumi‐LightPLUS western blotting substrate (Roche, Switzerland) served as the HRP substrate. The signals were detected using a ChemiDoc XRS+ station (BioRad).
Preparation of genomic DNA for whole‐genome sequencing
Genomic DNA (gDNA) was purified from 2 ml of an overnight culture of the respective strain. DNA extraction was performed using 100/G Genomic‐tips together with a Genomic DNA buffer set as described in the manufacturer's instructions (Qiagen). After precipitation, the DNA samples were washed twice with cold 70% ethanol and dissolved in Tris buffer (10 mM Tris–HCl, pH 8.0).
Long‐read PacBio‐based genome sequencing
Sample preparation and genome sequencing was performed by the Genomic Technology Facility of the University of Lausanne (Switzerland) using standard protocols. Briefly, DNA samples were sheared in Covaris g‐TUBEs to obtain fragments with a mean length of 20 kb. The sheared DNA was used to prepare each library with the PacBio SMRTbell template prep kit 1 (Pacific Biosciences) according to the manufacturer's recommendations. The resulting library was size selected on a BluePippin system (Sage Science) for molecules larger than 15 kb, which excluded smaller plasmids. Each library was sequenced on one single‐molecule real‐time (SMRT) cell with P6/C4 chemistry and MagBeads on a PacBio RS II system at a movie length of 360 min. Genome assembly was performed using the protocol RS_HGAP_Assembly.3 in SMRT Pipe 2.3.0, and circularization of the genomes was achieved using the Minimus assembler of the AMOS software package 3.1.0 using default parameters (Sommer et al., 2007). The assembled genomes were initially annotated using Prokka 1.12 (Seemann, 2014) but due to several incompatibilities with the NCBI database, they were reannotated with their own pipeline (PGAP annotation) during NCBI submission. The genomic data and NCBI accession numbers are summarized in the Supporting information Table S2.
Notably, we observed minor differences in the T6SS clusters of the strains when comparing Prokka and PGAP annotations. For instance, we noticed slight differences in the starting points of some of the predicted orphan loci from aux 1, even though the reading frames were comparable between the annotation methods. Additionally, the vgrG3 gene from the large cluster of some strains was shortened in the PGAP annotation, which most likely excluded the effector encoding part of the gene. In these cases (indicated in the Supporting Information Table S3), we considered the Prokka annotation for our analysis. Finally, some annotations were created beyond internal frameshifted stop codons. For instance, in the auxiliary cluster 1 of strain W10G, the effector gene (HPY12_07370) contains several predicted stop codons in the PGAP annotation, due to the insertion of a C in position 323. A similar case was observed in the putative effector gene (HPY12_14010) from auxiliary cluster 2, due to the insertion of a C in position 889. Both of these regions were Sanger sequenced after PCR‐amplification using the same genomic DNA samples as templates that were initially used for PacBio library preparation, which confirmed that the additional C bases were in both cases a sequencing artefact and the genes were properly maintained in strain W10G (as indicated in Fig. 3).
Characterization of E/I modules from environmental strains
T6SS clusters of the environmental strains were identified by searching the Prokka‐annotated genomes for conserved genes, such as paar, vgrG and hcp, as well as according to their location when aligned to the genome of the pandemic strain A1552 (Matthey et al., 2018; Matthey et al., 2019 ). All identified putative effectors were compared by BLAST against the NCBI database to identify conserved domains. Additional characterization was made based on the previous literature (Unterweger et al., 2014; Kirchberger et al., 2017). Furthermore, all E/I modules and orphan immunity loci were classified for their family type. Proteins with a sequence of less than 30% identity were considered as distinct incompatible types, as previously described (Unterweger et al., 2014; Kirchberger et al., 2017). For VgrG3 proteins, the typing was only based on the effector portion of the protein (Unterweger et al., 2014). To determine this part of the protein, the full VgrG3 sequence from all strains was aligned. The conserved region corresponding to the VgrG part of the protein was subsequently removed, which left only the variable C‐terminal effector domain. This part was then used for typing and pairwise comparisons. The putative orphan immunity proteins were queried against a translated nucleotide database (PATRIC; Wattam et al., 2017) to identify homologous bona fide immunity proteins encoded adjacent to an effector gene in the other T6SS clusters.
Data availability
PacBio raw reads of the 14 whole‐genome sequenced strains have been deposited in NCBI's Sequence Read Archive (SRA) under Bioproject accession number PRJNA633476. Details on the SRA accession numbers, BioSamples, and individual accession numbers of the de novo assembled and circularized genomes are provided in the Supporting Information Table S2.
Supporting information
Table S1 Vibrio cholerae, Escherichia coli and Klebsiella pneumoniae strains used in this study.
Table S2. Information of the long‐read whole genome sequencing data and assemblies of NCBI BioProject PRJNA633476.
Table S3. BlastP‐predicted T6SS effector and immunity proteins1 of the environmental V. cholerae isolates.
Table S4. Matrices of % identity (percentage of residues which are identical) among VgrG1‐ACD proteins in T6SS auxiliary cluster 1 from clade C and D environmental V. cholerae strains and pandemic strain A1552.
Table S5. Matrices of % identity (percentage of residues that are identical) among the effector and immunity proteins harboured in the T6SS large cluster from the environmental V. cholerae strains and the pandemic strain A1552.
Table S6. Matrices of % identity (percentage of residues that are identical) among the effector and immunity proteins harboured in the T6SS auxiliary cluster 1 from the environmental V. cholerae strains and the pandemic strain A1552.
Table S7. Matrices of % identity (percentage of residues that are identical) among the effector and immunity proteins harboured in the T6SS auxiliary cluster 2 from the environmental V. cholerae strains and the pandemic strain A1552.
Acknowledgements
The authors acknowledge Sandrine Stutzmann for technical assistance, A. Boehm for sharing the published environmental V. cholerae isolates, Lisa Metzger for provision of strain constructs, and Thierry Soldati for sharing the D. discoideum amoebae. The authors also thank Frédérique Le Roux and current and former members of the Blokesch group and of the Lausanne/Geneva amoeba club for fruitful discussions. The authors acknowledge the staff of the Lausanne Genomic Technologies Facility at the University of Lausanne for sample processing, sequencing, and genome assembly, the NCBI submission staff for help with data deposition, and Anne‐Catherine Portmann and Ivan Mateus for preliminary analysis of the genome data. The authors also note that the purchase of the Pacific Biosciences RS II instrument at the University of Lausanne was partially supported by the Loterie Romande through the Fondation pour la Recherche en Médecine Génétique. This work was supported by the Swiss National Science Foundation (310030_185022), the Novartis Foundation for medical‐biological research (#18C178), and a Consolidator grant by the European Research Council (724630). M.B. is a Howard Hughes Medical Institute (HHMI) International Research Scholar (grant 55008726).
References
- Aldova, E. , Laznickova, K. , Stepankova, E. , and Lietava, J. (1968) Isolation of nonagglutinable vibrios from an enteritis outbreak in Czechoslovakia. J Infect Dis 118: 25–31. [DOI] [PubMed] [Google Scholar]
- Ali, M. , Nelson, A.R. , Lopez, A.L. , and Sack, D.A. (2015) Updated global burden of cholera in endemic countries. PLoS Negl Trop Dis 9: 1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Altindis, E. , Dong, T. , Catalano, C. , and Mekalanos, J.J. (2015) Secretome analysis of Vibrio cholerae type VI secretion system reveals a new effector‐immunity pair. MBio 6: e00075‐15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Basler, M. , Pilhofer, M. , Henderson, G.P. , Jensen, G.J. , and Mekalanos, J.J. (2012) Type VI secretion requires a dynamic contractile phage tail‐like structure. Nature 483: 182–186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benghezal, M. , Fauvarque, M.O. , Tournebize, R. , Froquet, R. , Marchetti, A. , Bergeret, E. , et al (2006) Specific host genes required for the killing of Klebsiella bacteria by phagocytes. Cell Microbiol 8: 139–148. [DOI] [PubMed] [Google Scholar]
- Bernardy, E.E. , Turnsek, M.A. , Wilson, S.K. , Tarr, C.L. , and Hammer, B.K. (2016) Diversity of clinical and environmental isolates of Vibrio cholerae in natural transformation and contact‐dependent bacterial killing indicative of type VI secretion system activity. Appl Environ Microbiol 82: 2833–2842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bishop‐Lilly, K. , Johnson, S. , Verratti, K. , Truong, L. , Amy, K. , Joy, A. , et al (2014) Genome sequencing of 15 clinical isolates, including 13 non‐O1/non‐O139 serogroup strains. Genome Announc 2: e00893–e00814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blokesch, M. (2012) TransFLP ‐ a method to genetically modify Vibrio cholerae based on natural transformation and FLP‐recombination. J Vis Exp 68: e3761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borgeaud, S. , and Blokesch, M. (2013) Overexpression of the tcp gene cluster using the T7 RNA polymerase/promoter system and natural transformation‐mediated genetic engineering of Vibrio cholerae . PLoS One 8: e53952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borgeaud, S. , Metzger, L.C. , Scrignari, T. , and Blokesch, M. (2015) The type VI secretion system of Vibrio cholerae fosters horizontal gene transfer. Science 347: 63–67. [DOI] [PubMed] [Google Scholar]
- Boyd, E.F. , and Waldor, M.K. (2002) Evolutionary and functional analyses of variants of the toxin‐coregulated pilus protein TcpA from toxigenic Vibrio cholerae non‐O1/nonO139 serogroup isolates. Microbiology 148: 1655–1666. [DOI] [PubMed] [Google Scholar]
- Bröms, J.E. , Ishikawa, T. , Wai, S.N. , and Sjöstedt, A. (2013) A functional VipA‐VipB interaction is required for the type VI secretion system activity of Vibrio cholerae O1 strain A1552. BMC Microbiol 13: 96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brooks, T.M. , Unterweger, D. , Bachmann, V. , Kostiuk, B. , and Pukatzki, S. (2013) Lytic activity of the Vibrio cholerae type VI secretion toxin VgrG‐3 is inhibited by the antitoxin TsaB. J Biol Chem 288: 7618–7625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buist, G. , Steen, A. , Kok, J. , and Kuipers, O.P. (2008) LysM, a widely distributed protein motif for binding to (peptido)glycans. Mol Microbiol 68: 838–847. [DOI] [PubMed] [Google Scholar]
- Cherrak, Y. , Flaugnatti, N. , Durand, E. , Journet, L. , and Cascales, E. (2019) Structure and regulation of the type VI secretion system. Microbiol Spectr 7: PSIB‐0031‐2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou, S. , Bui, N.K. , Russell, A.B. , Lexa, K.W. , Gardiner, T.E. , LeRoux, M. , et al (2012) Structure of a peptidoglycan amidase effector targeted to gram‐negative bacteria by the type VI secretion system. Cell Rep 1: 656–664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chun, J. , Grim, C.J. , Hasan, N.A. , Lee, J.H. , Choi, S.Y. , Haley, B.J. , et al (2009) Comparative genomics reveals mechanism for short‐term and long‐term clonal transitions in pandemic Vibrio cholerae . Proc Natl Acad Sci U S A 106: 15442–15447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cianfanelli, F.R. , Monlezun, L. , and Coulthurst, S.J. (2016) Aim, load, fire: the type VI secretion system, a bacterial nanoweapon. Trends Microbiol 24: 51–62. [DOI] [PubMed] [Google Scholar]
- Clemens, J.D. , Nair, G.B. , Ahmed, T. , Qadri, F. , and Holmgren, J. (2017) Cholera. Lancet 390: 1539–1549. [DOI] [PubMed] [Google Scholar]
- Cottingham, K.L. , Chiavelli, D.A. , and Taylor, R.K. (2003) Environmental microbe and human pathogen: the ecology and microbiology of Vibrio cholerae . Front Ecol Environm 1: 80–86. [Google Scholar]
- Crisan, C.V. , Chande, A.T. , Williams, K. , Raghuram, V. , Rishishwar, L. , Steinbach, G. , et al (2019) Analysis of Vibrio cholerae genomes identifies new type VI secretion system gene clusters. Genome Biol 20: 163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crisan, C.V. , and Hammer, B.K. (2020) The Vibrio cholerae type VI secretion system: toxins, regulators and consequences. Environ Microbiol (Online Ahead of Print). [DOI] [PubMed] [Google Scholar]
- Dalia, A.B. , McDonough, E.K. , and Camilli, A. (2014) Multiplex genome editing by natural transformation. Proc Natl Acad Sci U S A 111: 8937–8942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deshayes, S. , Daurel, C. , Cattoir, V. , Parienti, J.J. , Quilici, M.L. , and de La Blanchardière, A. (2015) Non‐O1, non‐O139 Vibrio cholerae bacteraemia: case report and literature review. Springerplus 4: 575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Domman, D. , Quilici, M.‐L. , Dorman, M.J. , Njamkepo, E. , Mutreja, A. , Mather, A.E. , et al (2017) Integrated view of Vibrio cholerae in the Americas. Science 358: 789–793. [DOI] [PubMed] [Google Scholar]
- Dong, T.G. , Ho, B.T. , Yoder‐Himes, D.R. , and Mekalanos, J.J. (2013) Identification of T6SS‐dependent effector and immunity proteins by Tn‐seq in Vibrio cholerae . Proc Natl Acad Sci U S A 110: 2623–2628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Durand, E. , Cambillau, C. , Cascales, E. , and Journet, L. (2014) VgrG, tae, Tle, and beyond: the versatile arsenal of type VI secretion effectors. Trends Microbiol 22: 498–507. [DOI] [PubMed] [Google Scholar]
- Dziejman, M. , Balon, E. , Boyd, D. , Fraser, C.M. , Heidelberg, J.F. , and Mekalanos, J.J. (2002) Comparative genomic analysis of Vibrio cholerae: genes that correlate with cholera endemic and pandemic disease. Proc Natl Acad Sci U S A 99: 1556–1561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dziejman, M. , Serruto, D. , Tam, V.C. , Sturtevant, D. , Diraphat, P. , Faruque, S.M. , et al (2005) Genomic characterization of non‐O1, non‐O139 Vibrio cholerae reveals genes for a type III secretion system. Proc Natl Acad Sci 102: 3465–3470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Faruque, S.M. , Albert, M.J. , and Mekalanos, J.J. (1998) Epidemiology, genetics, and ecology of toxigenic Vibrio cholerae . Microbiol Mol Biol Rev 62: 1301–1314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Faruque, S.M. , Chowdhury, N. , Kamruzzaman, M. , Dziejman, M. , Rahman, M.H. , Sack, D.A. , et al (2004) Genetic diversity and virulence potential of environmental Vibrio cholerae population in a cholera‐endemic area. Proc Natl Acad Sci U S A 101: 2123–2128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Faruque, S.M. , Kamruzzaman, M. , Meraj, I.M. , Chowdhury, N. , Nair, G.B. , Sack, R.B. , et al (2003) Pathogenic potential of environmental Vibrio cholerae strains carrying genetic variants of the toxin‐coregulated pilus pathogenicity Island. Infect Immun 71: 1020–1025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flaugnatti, N. , Le, T.T.H. , Canaan, S. , Aschtgen, M.S. , Nguyen, V.S. , Blangy, S. , et al (2016) A phospholipase A1 antibacterial type VI secretion effector interacts directly with the C‐terminal domain of the VgrG spike protein for delivery. Mol Microbiol 99: 1099–1118. [DOI] [PubMed] [Google Scholar]
- Flaugnatti, N. , Rapisarda, C. , Rey, M. , Beauvois, S.G. , Nguyen, V.A. , Canaan, S. , et al (2020) Structural basis for loading and inhibition of a bacterial T6SS phospholipase effector by the VgrG spike. EMBO J 39: e104129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galán, J.E. , and Waksman, G. (2018) Protein‐injection machines in bacteria. Cell 172: 1306–1318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gennari, M. , Ghidini, V. , Caburlotto, G. , and Lleo, M.M. (2012) Virulence genes and pathogenicity islands in environmental Vibrio strains nonpathogenic to humans. FEMS Microbiol Ecol 82: 563–573. [DOI] [PubMed] [Google Scholar]
- Gerisch, G. , Lüderitz, O. , and Ruschmann, E. (1967) Antikörper fördern die Phagozytose von Bakterien durch Amöben. Zsch Naturforsch 22b: 109. [PubMed] [Google Scholar]
- Granato, E.T. , Meiller‐Legrand, T.A. , and Foster, K.R. (2019) The evolution and ecology of bacterial warfare. Curr Biol 29: R521–R537. [DOI] [PubMed] [Google Scholar]
- Hachani, A. , Wood, T.E. , and Filloux, A. (2016) Type VI secretion and anti‐host effectors. Curr Opin Microbiol 29: 81–93. [DOI] [PubMed] [Google Scholar]
- Harris, J.B. , LaRocque, R.C. , Qadri, F. , Ryan, E.T. , and Calderwood, S.B. (2012) Cholera. Lancet 379: 2466–2476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hasan, N.A. , Rezayat, T. , Blatz, P.J. , Choi, S.Y. , Griffitt, K.J. , Rashed, S.M. , et al (2015) Nontoxigenic Vibrio cholerae non‐O1/O139 isolate from a case of human gastroenteritis in the U.S. Gulf Coast. J Clin Microbiol 53: 9–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heidelberg, J.F. , Elsen, J.A. , Nelson, W.C. , Clayton, R.A. , Gwinn, M.L. , Dodson, R.J. , et al (2000) DNA sequence of both chromosomes of the cholera pathogen Vibrio cholerae . Nature 406: 477–483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van der Henst, C. , Vanhove, A.S. , Drebes Dörr, N.C. , Stutzmann, S. , Stoudmann, C. , Clerc, S. , et al (2018) Molecular insights into Vibrio cholerae's intra‐amoebal host‐pathogen interactions. Nat Commun 9: 3420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hersch, S.J. , Watanabe, N. , Stietz, M.S. , Manera, K. , Kamal, F. , Burkinshaw, B. , et al (2020) Envelope stress responses defend against type six secretion system attacks independently of immunity proteins. Nat Microbiol 5: 706–714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hibbing, M.E. , Fuqua, C. , Parsek, M.R. , and Peterson, S.B. (2010) Bacterial competition: surviving and thriving in the microbial jungle. Nat Rev Microbiol 8: 15–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ho, B.T. , Dong, T.G. , and Mekalanos, J.J. (2014) A view to a kill: the bacterial type VI secretion system. Cell Host Microbe 15: 9–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hood, R.D. , Singh, P. , Hsu, F.S. , Güvener, T. , Carl, M.A. , Trinidad, R.R.S. , et al (2010) A type VI secretion system of Pseudomonas aeruginosa targets a toxin to bacteria. Cell Host Microbe 7: 25–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ichinose, Y. , Yamamoto, K. , Nakasone, N. , Tanabe, M.J. , Takeda, T. , Miwatani, T. , and Iwanaga, M. (1987) Enterotoxicity of El Tor‐like hemolysin of non‐O1 Vibrio cholerae . Infect Immun 55: 1090–1093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Islam, M.S. , Alam, M.J. , and Neogi, P.K.B. (1992) Seasonality and toxigenicity of Vibrio cholerae non‐O1 isolated from different components of pond ecosystems of Dhaka City, Bangladesh. World J Microbiol Biotechnol 8: 160–163. [DOI] [PubMed] [Google Scholar]
- Jaskólska, M. , Stutzmann, S. , Stoudmann, C. , and Blokesch, M. (2018) QstR‐dependent regulation of natural competence and type VI secretion in Vibrio cholerae . Nucleic Acids Res 46: 10619–10634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson, J.A. , Salles, C.A. , Panigrahi, P. , Albert, M.J. , Wright, A.C. , Johnson, R.J. , and Morris, J.G. (1994) Vibrio cholerae O139 synonym Bengal is closely related to Vibrio cholerae El Tor but has important differences. Infect Immun 62: 2108–2110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keene, O.N. (1995) The log transformation is special. Stat Med 14: 811–819. [DOI] [PubMed] [Google Scholar]
- Keymer, D.P. , Miller, M.C. , Schoolnik, G.K. , and Boehm, A.B. (2007) Genomic and phenotypic diversity of coastal Vibrio cholerae strains is linked to environmental factors. Appl Environ Microbiol 73: 3705–3714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirchberger, P.C. , Unterweger, D. , Provenzano, D. , Pukatzki, S. , and Boucher, Y. (2017) Sequential displacement of type VI secretion system effector genes leads to evolution of diverse immunity gene arrays in Vibrio cholerae . Sci Rep 7: 45133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirn, T.J. , Jude, B.A. , and Taylor, R.K. (2005) A colonization factor links Vibrio cholerae environmental survival and human infection. Nature 438: 863–866. [DOI] [PubMed] [Google Scholar]
- Kube, S. , Kapitein, N. , Zimniak, T. , Herzog, F. , Mogk, A. , and Wendler, P. (2014) Structure of the VipA/B type VI secretion complex suggests a contraction‐state‐specific recycling mechanism. Cell Rep 8: 20–30. [DOI] [PubMed] [Google Scholar]
- Labbate, M. , Orata, F.D. , Petty, N.K. , Jayatilleke, N.D. , King, W.L. , Kirchberger, P.C. , et al (2016) A genomic Island in Vibrio cholerae with VPI‐1 site‐specific recombination characteristics contains CRISPR‐Cas and type VI secretion modules. Sci Rep 6: 36891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liang, X. , Moore, R. , Wilton, M. , Wong, M.J.Q. , Lam, L. , and Dong, T.G. (2015) Identification of divergent type VI secretion effectors using a conserved chaperone domain. Proc Natl Acad Sci U S A 112: 9106–9111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Logan, S.L. , Thomas, J. , Yan, J. , Baker, R.P. , Shields, D.S. , Xavier, J.B. , et al (2018) The Vibrio cholerae type VI secretion system can modulate host intestinal mechanics to displace gut bacterial symbionts. Proc Natl Acad Sci U S A 115: E3779–E3787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma, A.T. , McAuley, S. , Pukatzki, S. , and Mekalanos, J.J. (2009) Translocation of a Vibrio cholerae type VI secretion effector requires bacterial endocytosis by host cells. Cell Host Microbe 5: 234–243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma, A.T. , and Mekalanos, J.J. (2010) In vivo actin cross‐linking induced by Vibrio cholerae type VI secretion system is associated with intestinal inflammation. Proc Natl Acad Sci U S A 107: 4365–4370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marvig, R.L. , and Blokesch, B. (2010). Natural transformation of Vibrio cholerae as a tool ‐ Optimizing the procedure. BMC Microbiol 10: 115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matthey, N. , Drebes Dörr, N.C. , and Blokesch, M. (2018) Long‐read‐based genome sequences of pandemic and environmental Vibrio cholerae strains. Microbiol Resour Announc 7: e01574–e01518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matthey, N. , Stutzmann, S. , Stoudmann, C. , Guex, N. , Iseli, C. , and Blokesch, M. (2019). Neighbor predation linked to natural competence fosters the transfer of large genomic regions in Vibrio cholerae . eLife 8: e48212 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McNally, L. , Bernardy, E. , Thomas, J. , Kalziqi, A. , Pentz, J. , Brown, S.P. , et al (2017) Killing by type VI secretion drives genetic phase separation and correlates with increased cooperation. Nat Commun 8: 14371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meibom, K.L. , Blokesch, M. , Dolganov, N.A. , Wu, C.Y. , and Schoolnik, G.K. (2005) Chitin induces natural competence in Vibrio cholerae . Science 310: 1824–1827. [DOI] [PubMed] [Google Scholar]
- Metzger, L.C. , and Blokesch, M. (2016) Regulation of competence‐mediated horizontal gene transfer in the natural habitat of Vibrio cholerae . Curr Opin Microbiol 30: 1–7. [DOI] [PubMed] [Google Scholar]
- Metzger, L.C. , Stutzmann, S. , Scrignari, T. , Van der Henst, C. , Matthey, N. , and Blokesch, M. (2016) Independent regulation of type VI secretion in Vibrio cholerae by TfoX and TfoY. Cell Rep 15: 951–958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller, M.C. , Keymer, D.P. , Avelar, A. , Boehm, A.B. , and Schoolnik, G.K. (2007) Detection and transformation of genome segments that differ within a coastal population of Vibrio cholerae strains. Appl Environ Microbiol 73: 3695–3704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyata, S.T. , Kitaoka, M. , Brooks, T.M. , McAuley, S.B. , and Pukatzki, S. (2011) Vibrio cholerae requires the type VI secretion system virulence factor vasx to kill Dictyostelium discoideum . Infect Immun 79: 2941–2949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyata, S.T. , Unterweger, D. , Rudko, S.P. , and Pukatzki, S. (2013) Dual expression profile of type VI secretion system immunity genes protects pandemic Vibrio cholerae . PLoS Pathog 9: e1003752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mutreja, A. , Kim, D.W. , Thomson, N.R. , Connor, T.R. , Lee, J.H. , Kariuki, S. , et al (2011) Evidence for several waves of global transmission in the seventh cholera pandemic. Nature 477: 462–465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okada, K. , Iida, T. , Kita‐Tsukamoto, K. , and Honda, T. (2005) Vibrios commonly possess two chromosomes. J Bacteriol 187: 752–757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olivier, V. , Haines, G.K. , Tan, Y. , and Fullner Satchell, K.J. (2007) Hemolysin and the multifunctional autoprocessing RTX toxin are virulence factors during intestinal infection of mice with Vibrio cholerae El tor O1 strains. Infect Immun 75: 5035–5042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Onifade, T.J.M. , Hutchinson, R. , van Zile, K. , Bodager, D. , Baker, R. , and Blackmore, C. (2011) Toxin producing Vibrio cholerae O75 outbreak, United States, march to April 2011. Eurosurveillance 16: 20–22. [PubMed] [Google Scholar]
- Pukatzki, S. , Ma, A.T. , Revel, A.T. , Sturtevant, D. , and Mekalanos, J.J. (2007) Type VI secretion system translocates a phage tail spike‐like protein into target cells where it cross‐links Actin. Proc Natl Acad Sci U S A 104: 15508–15513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pukatzki, S. , Ma, A.T. , Sturtevant, D. , Krastins, B. , Sarracino, D. , Nelson, W.C. , et al (2006) Identification of a conserved bacterial protein secretion system in Vibrio cholerae using the Dictyostelium host model system. Proc Natl Acad Sci U S A 103: 1528–1533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pukatzki, S. , McAuley, S.B. , and Miyata, S.T. (2009) The type VI secretion system: translocation of effectors and effector‐domains. Curr Opin Microbiol 12: 11–17. [DOI] [PubMed] [Google Scholar]
- Rivera, I.N.G. , Chun, J. , Huq, A. , Sack, R.B. , and Colwell, R.R. (2001) Genotypes associated with virulence in environmental isolates of Vibrio cholerae . Appl Environ Microbiol 67: 2421–2429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russell, A.B. , Hood, R.D. , Bui, N.K. , Leroux, M. , Vollmer, W. , and Mougous, J.D. (2011) Type VI secretion delivers bacteriolytic effectors to target cells. Nature 475: 343–349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russell, A.B. , Leroux, M. , Hathazi, K. , Agnello, D.M. , Ishikawa, T. , Wiggins, P.A. , et al (2013) Diverse type VI secretion phospholipases are functionally plastic antibacterial effectors. Nature 496: 508–512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russell, A.B. , Peterson, S.B. , and Mougous, J.D. (2014) Type VI secretion system effectors: poisons with a purpose. Nat Rev Microbiol 12: 137–148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salomon, D. , Klimko, J.A. , Trudgian, D.C. , Kinch, L.N. , Grishin, N.V. , Mirzaei, H. , and Orth, K. (2015) Type VI secretion system toxins horizontally shared between marine bacteria. PLoS Pathog 11: e1005128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santoriello, F.J. , Michel, L. , Unterweger, D. , and Pukatzki, S. (2019) Vibrio cholerae type VI secretion system auxiliary cluster 3 is a pandemic‐associated mobile genetic element. bioRxiv. 10.1101/868539. [DOI] [Google Scholar]
- Lo Scrudato, M. , and Blokesch, M. (2013) A transcriptional regulator linking quorum sensing and chitin induction to render Vibrio cholerae naturally transformable. Nucleic Acids Res 41: 3644–3658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lo Scrudato, M. , and Blokesch, M. (2012) The regulatory network of natural competence and transformation of vibrio cholerae. PLoS Genet 8: e1002778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seemann, T. (2014) Prokka: rapid prokaryotic genome annotation. Bioinformatics 30: 2068–2069. [DOI] [PubMed] [Google Scholar]
- Shapiro, B.J. , Levade, I. , Kovacikova, G. , Taylor, R.K. , and Almagro‐Moreno, S. (2016) Origins of pandemic Vibrio cholerae from environmental gene pools. Nat Microbiol 2: 16240. [DOI] [PubMed] [Google Scholar]
- Shneider, M.M. , Buth, S.A. , Ho, B.T. , Basler, M. , Mekalanos, J.J. , and Leiman, P.G. (2013) PAAR‐repeat proteins sharpen and diversify the type VI secretion system spike. Nature 500: 350–353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Souza Silva, O. , and Blokesch, M. (2010) Genetic manipulation of Vibrio cholerae by combining natural transformation with FLP recombination. Plasmid 64: 186–195. [DOI] [PubMed] [Google Scholar]
- Singh, D.V. , Matte, M.H. , Matte, G.R. , Jiang, S. , Sabeena, F. , Shukla, B.N. , et al (2001) Molecular analysis of Vibrio cholerae O1, O139, non‐O1, and non‐O139 strains: clonal relationships between clinical and environmental isolates. Appl Environ Microbiol 67: 910–921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sommer, D.D. , Delcher, A.L. , Salzberg, S.L. , and Pop, M. (2007) Minimus: a fast, lightweight genome assembler. BMC Bioinformatics 8: 64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Speare, L. , Cecere, A.G. , Guckes, K.R. , Smith, S. , Wollenberg, M.S. , Mandel, M.J. , et al (2018) Bacterial symbionts use a type VI secretion system to eliminate competitors in their natural host. Proc Natl Acad Sci U S A 115: E8528–E8537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stubbendieck, R.M. , and Straight, P.D. (2016) Multifaceted interfaces of bacterial competition. J Bacteriol 198: 2145–2155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sussuman, M. (1987) Cultivation and synchronous morphogenesis of Dictyostelium under controlled experimental conditions In Spudich J.A. (ed). Methods in Cell Biology, vol. 28 ‐ Dictyostelium discoideum: Molecular Approaches to Cell Biology. New York: Academic Press, Inc., 9–29. [DOI] [PubMed] [Google Scholar]
- Taylor, N.M.I. , van Raaij, M.J. , and Leiman, P.G. (2018) Contractile injection systems of bacteriophages and related systems. Mol Microbiol 108: 6–15. [DOI] [PubMed] [Google Scholar]
- Taylor, R.K. , Miller, V.L. , Furlong, D.B. , and Mekalanos, J.J. (1987) Use of phoA gene fusions to identify a pilus colonization factor coordinately regulated with cholera toxin. Proc Natl Acad Sci U S A 84: 2833–2837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Unterweger, D. , Kitaoka, M. , Miyata, S.T. , Bachmann, V. , Brooks, T.M. , Moloney, J. , et al (2012) Constitutive type VI secretion system expression gives Vibrio cholerae intra‐ and interspecific competitive advantages. PLoS One 7: e48320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Unterweger, D. , Kostiuk, B. , Otjengerdes, R. , Wilton, A. , Diaz‐Satizabal, L. , and Pukatzki, S. (2015) Chimeric adaptor proteins translocate diverse type VI secretion system effectors in Vibrio cholerae . EMBO J 34: 2198–2210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Unterweger, D. , Miyata, S.T. , Bachmann, V. , Brooks, T.M. , Mullins, T. , Kostiuk, B. , et al (2014) The Vibrio cholerae type VI secretion system employs diverse effector modules for intraspecific competition. Nat Commun 5: 3549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Veening, J.W. , and Blokesch, M. (2017) Interbacterial predation as a strategy for DNA acquisition in naturally competent bacteria. Nat Rev Microbiol 15: 621–629. [DOI] [PubMed] [Google Scholar]
- Waldor, M.K. , and Mekalanos, J.J. (1996) Lysogenic conversion by a filamentous phage encoding cholera toxin. Science 272: 1910–1914. [DOI] [PubMed] [Google Scholar]
- Wattam, A.R. , Davis, J.J. , Assaf, R. , Boisvert, S. , Brettin, T. , Bun, C. , et al (2017) Improvements to PATRIC, the all‐bacterial bioinformatics database and analysis resource center. Nucleic Acids Res 45: D535–D542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watve, S.S. , Thomas, J. , and Hammer, B.K. (2015) CytR is a global positive regulator of competence, type VI secretion, and chitinases in Vibrio cholerae . PLoS One 10: e0138834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weill, F.‐X. , Domman, D. , Njamkepo, E. , Tarr, C. , Rauzier, J. , Fawal, N. , et al (2017) Genomic history of the seventh pandemic of cholera in Africa. Science 358: 785–789. [DOI] [PubMed] [Google Scholar]
- Weill, F.X. , Domman, D. , Njamkepo, E. , Almesbahi, A.A. , Naji, M. , Nasher, S.S. , et al (2019) Genomic insights into the 2016–2017 cholera epidemic in Yemen. Nature 565: 230–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- World Health Organization (WHO) (2006) Communicable diseases following natural disasters: risk assessment and priority interventions.
- Yildiz, F.H. , and Schoolnik, G.K. (1998) Role of rpoS in stress survival and virulence of Vibrio cholerae . J Bacteriol 180: 773–784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, X.H. , and Austin, B. (2005) Haemolysins in Vibrio species. J Appl Microbiol 98: 1011–1019. [DOI] [PubMed] [Google Scholar]
- Zheng, J. , Ho, B. , and Mekalanos, J.J. (2011) Genetic analysis of anti‐amoebae and anti‐bacterial activities of the type vi secretion system in Vibrio cholerae . PLoS One 6: e23876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zoued, A. , Brunet, Y.R. , Durand, E. , Aschtgen, M.S. , Logger, L. , Douzi, B. , et al (2014) Architecture and assembly of the type VI secretion system. Biochim Biophys Acta ‐ Mol Cell Res 1843: 1664–1673. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1 Vibrio cholerae, Escherichia coli and Klebsiella pneumoniae strains used in this study.
Table S2. Information of the long‐read whole genome sequencing data and assemblies of NCBI BioProject PRJNA633476.
Table S3. BlastP‐predicted T6SS effector and immunity proteins1 of the environmental V. cholerae isolates.
Table S4. Matrices of % identity (percentage of residues which are identical) among VgrG1‐ACD proteins in T6SS auxiliary cluster 1 from clade C and D environmental V. cholerae strains and pandemic strain A1552.
Table S5. Matrices of % identity (percentage of residues that are identical) among the effector and immunity proteins harboured in the T6SS large cluster from the environmental V. cholerae strains and the pandemic strain A1552.
Table S6. Matrices of % identity (percentage of residues that are identical) among the effector and immunity proteins harboured in the T6SS auxiliary cluster 1 from the environmental V. cholerae strains and the pandemic strain A1552.
Table S7. Matrices of % identity (percentage of residues that are identical) among the effector and immunity proteins harboured in the T6SS auxiliary cluster 2 from the environmental V. cholerae strains and the pandemic strain A1552.
Data Availability Statement
PacBio raw reads of the 14 whole‐genome sequenced strains have been deposited in NCBI's Sequence Read Archive (SRA) under Bioproject accession number PRJNA633476. Details on the SRA accession numbers, BioSamples, and individual accession numbers of the de novo assembled and circularized genomes are provided in the Supporting Information Table S2.