

Abstract
Diacylglycerol kinases (DGKs) play important roles in restraining diacylglycerol (DAG)-mediated signaling. Within the DGK family, the ζ isoform appears to be the most important isoform in T cells for controlling their development and function. DGKζ has been demonstrated to regulate T cell maturation, activation, anergy, effector/memory differentiation, defense against microbial infection, and anti-tumor immunity. Given its critical functions, DGKζ function should be tightly regulated to ensure proper signal transduction; however, mechanisms that control DGKζ function are still poorly understood. We report here that DGKζ dynamically translocates from the cytosol into the nuclei in T cells after TCR stimulation. In mice, DGKζ mutant defective in nuclear localization displayed enhanced ability to inhibit TCR-induced DAG-mediated signaling in primary T cells, maturation of conventional αβT and iNKT cells, and activation of peripheral T cells compared with wild-type DGKζ. Our study reveals for the first time nuclear sequestration of DGKζ as a negative control mechanism to spatially restrain it from terminating DAG mediated signaling in T cells. Our data suggest that manipulation of DGKζ nucleus-cytosol shuttling as a novel strategy to modulate DGKζ activity and immune responses for treatment of autoimmune diseases and cancer.
Keywords: TCR, diacylglycerol kinases, T cell development, invariant NKT cell, Ras/MAPK
INTRODUCTION
Diacylglycerol (DAG) is an important second messenger that mediates signal transduction downstream of numerous receptors. It associates with and regulates the subcellular localization and activities of multiple effector molecules. DAG-mediated signal pathways play important roles in numerous cellular processes and functions such as cell growth, differentiation, proliferation, metabolism, survival, and effector function [1, 2]. In T lymphocytes, DAG activates the RasGRP1-Ras-Erk1/2 and PKCθ-IKK-NFκB pathways to promote T cell development and activation [3–7]. Evidence has also illustrated equal importance of tight control of DAG-mediated signaling. Dysregulation of DAG signal can cause developmental defects, abnormal cellular activation and function, and even tumorigenesis [8, 9].
One pathway involved in terminating DAG is mediated by DAG kinases (DAG) via phosphorylation and conversion to phosphatidic acid (PA) [1, 2]. PA itself is also an important second messenger that regulates the activities of multiple effector molecules such as PI5K and SHP1 [10, 11]. Thus, DGKs can function as a switch by terminating DAG signal and simultaneously initiating PA mediated signal. Several studies have suggested that such a switch could exert important roles in the immune system [8, 12, 13]. In mammals, ten DGKs encoded by different genes have been identified. Some DGKs contain splicing isoforms, adding complexity of this family of enzymes [1, 2]. DGKα, ζ, and δ are the predominant isoforms of the DGK family expressed in T cells [14–17]. While the physiological function of DGKδ in the immune system remains unclear, DGKα and ζ have been demonstrated to be important regulators in multiple immune cell lineages, particularly in T lymphocytes, by functioning synergistically or redundantly in these cells. Both DGKα and ζ, inhibit the RasGRP1-Ras-Erk1/2 pathway, the PKCθ-IKK-NFκB pathway, PI3K-AKT and mTOR signaling [14, 18–21]. They regulate the development of conventional αβT cells, iNKT cells, mucosal associated invariant T cells, and regulatory T cells, inhibit T cell activation but promote T cell anergy, regulate activation induced T cell death, control T helper cell differentiation and effector and memory CD8 T cell responses to viral and bacterial pathogens, and inhibit CD8 T cell- and chimeric antigen receptor T cell-mediated anti-tumor responses[8, 15, 16, 19, 20, 22–36]. In T cells, B cells, and NK cells, DGKζ appears to exert greater importance than DGKα [15, 31, 37, 38].
All DGK isoforms contain a catalytic domain and 2 – 3 cystine rich C1-like domains. They also contain other structural features, based on which DGKs have been divided into five types. Such type-specific structural features may mediate protein-protein and protein-lipid interaction to control DGK activity, subcellular localization, and function [39–43]. Many in vitro studies with various cell line models have suggested important roles of some DGK structural features in regulating DGK function [14, 40, 41, 44]. However, in vivo importance of specific structural features of the DGK family under physiological and pathological conditions has not been illustrated thus far.
DGKζ, belonging to type IV of the DGK family, contains several type-specific structural motifs/domains besides the catalytic and C1-like domains. A nuclear localization sequences (NLS) overlaps with a MARCKS motif and is localized between the C1 domain and the catalytic domain; a nuclear export sequence is located within the catalytic domain; a PDZ-binding motif sits at the carboxyl terminus; and several ankyrin repeats are localized close to the C-terminus [45–47]. DGKζ can be detected in both cytosolic and nuclear compartments [44, 48, 49]. Cytosolic location of DGKζ allows it to quickly translate to the cytoplasmic membrane and immune synapse to assess it substrate [50]. However, the functional significance of DGKζ nuclear localization in immune cells is still unclear.
In this report, we generated and examined multiple mouse models that allow conditional expression of wild-type (WT), kinase dead (KD) mutant, or nuclear localization mutant (ΔNLS) DGKζ after Cre-mediated deletion of a transcription stop cassette flanked by two LoxP sites. We demonstrated that WT DGKζ dynamically translocated from the cytosolic compartment into the nuclei in primary T cells after TCR engagement and that mutation of its NLS prevented it from entering the nucleus. Compared with DGKζWT, DGKζΔNLS mutant displayed enhanced ability to inhibit DAG-mediated signaling, intrathymic T cell and iNKT cell development, and T cell activation. Our data reveal that nuclear localization is a mechanism to negative control of DGKζ function by preventing its access to substrate. Additionally, we provide evidence that iNKT cell development was subjected to stricter requirement of DAG signaling than conventional αβT cells.
RESULTS
Generation of mice carrying inducible WT and mutant DGKζ
To investigate whether DGKζ function is regulated by nuclear localization, we generated three lines of mice that carry either a GFP-DGKζWT, GFP-DGKζΔNLS (NLS mutant with lysines in the NLS mutated to alanine), or GFP-DGKζKD (kinase dead with the glycine residue in the ATP binding site mutated to alanine) transgene knocked into the Rosa26 locus (Figure 1A, 1B). These transgenes contained the 104 kDa DGKζ1 isoform fused to the carboxyl terminus of EGFP to allow for identification of DGKζ expressing cells and DGKζ localization by GFP. A CAG promoter was inserted 5’ to the transgene to increase its expression and a transcription STOP cassette flanked by two loxP sites was located between the promoter and the GFP-DGKζ knockin (GFP-DGKζKI) transgene so that transgenic GFP-DGKζ could only be expressed after Cre-mediated deletion of the transcription STOP. Successful generation of these mice was confirmed by PCR amplification of the transgenes (Figure 1C).
Figure 1. Generation of WT and mutant DGKζ transgenic mice.
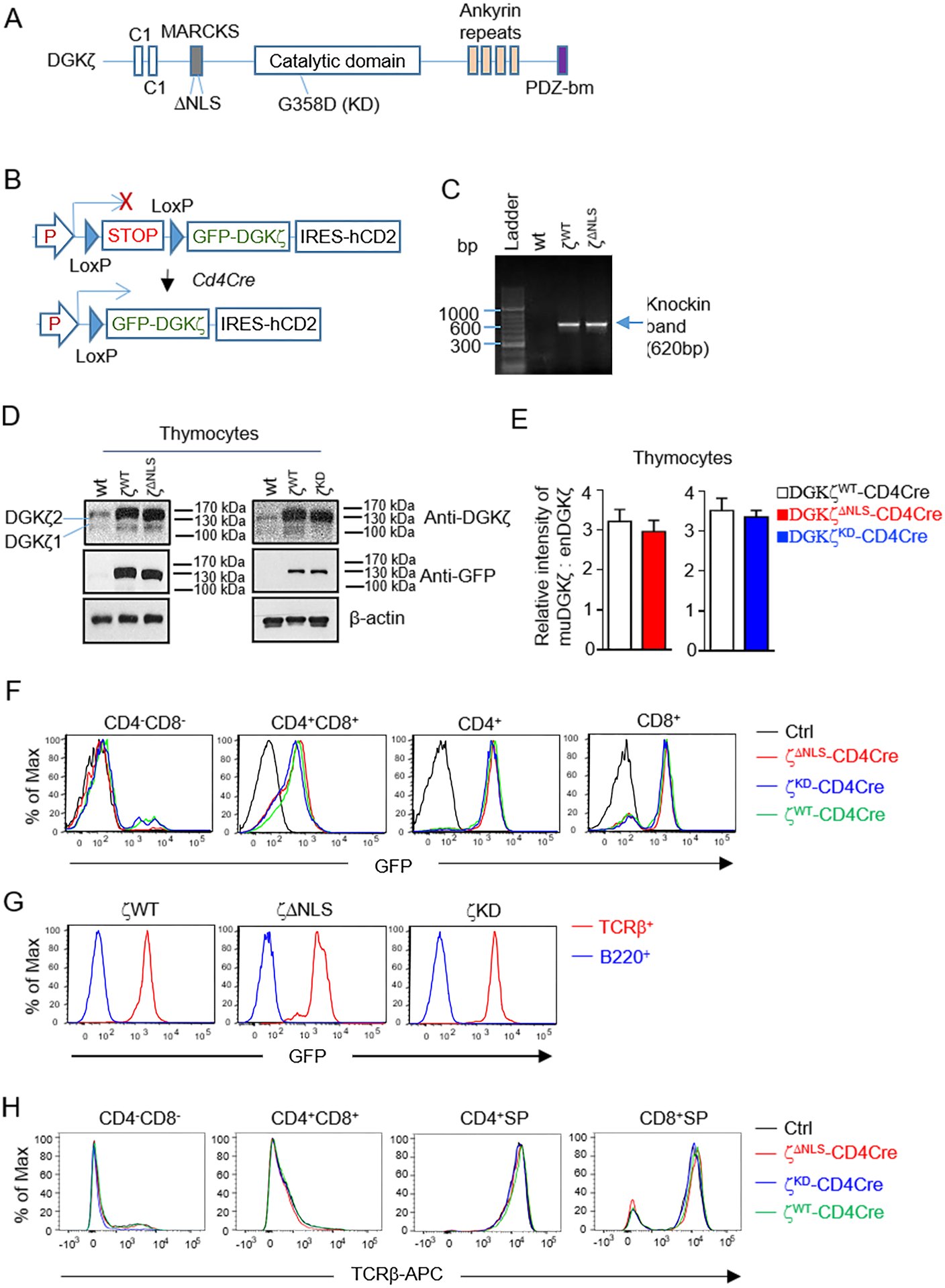
A. Schematic structure of DGKζ. B. GFP-DGKζ knock-in mice. Rosa: Rosa26 locus; P: CAG promoter; STOP, transcription stop cassette. C. Genotyping of GFP-DGKζKI mice by PCR. D. Detection of DGKζ by western blot with an anti-DGKζ antibody and anti-GFP antibody. Endogenous DGKζ1 and ζ2 are indicated with horizontal lines. Red arrow indicates GFP-DGKζ function proteins, which overlap with the endogenous DGKζ2. Loading control is detected with an anti-TCRβ antibody. E. Quantification the GFP-DGKζ fusion protein levels relative to endogenous DGKζ2 in WT thymocytes using Image J. F. Detection of GFP expression in thymocytes. G. Detection of GFP expression in B220+ B cells. H. TCRβ expression in different thymocyte subsets. Data in F and H were measured by flow cytometry. Each histogram represents 100% of the indicated cell population. Data shown are representative of or pooled from three experiments and error bars represent mean ± SEM, N = 3 mice for WT (Ctrl), GFP-DGKζWT-CD4Cre, GFP-DGKζΔNLS-CD4Cre, and GFP-DGKζKD-CD4Cre mice in D – H.
After breeding these mice with the CD4Cre transgenic mice to delete the STOP cassette starting at the CD4+CD8+ double positive (DP) thymocytes, we could detect GFP-DGKζ fusion protein in GFP-DGKζWT-, GFP-DGKζKD-, and GFP-DGKζΔNLS-CD4Cre thymocytes with an anti-GFP antibody. Both endogenous DGKζ1 and ζ2 could be detected in control total thymocyte lysates using an anti-DGKζ antibody and the GFP-DGKζ fusion proteins overlapped with the endogenous DGKζ2 (124kDa). The expression levels of these fusion proteins appeared similar in thymocytes detected by Western blot analysis (Figure 1D, 1E) and in live CD4+CD8+ double positive (DP), CD4+CD8− single positive (SP), and CD4−CD8+ SP thymocytes based on GFP expression (Figure 1F). GFP-DGKζWT, GFP-DGKζKD, and GFP-DGKζΔNLS fusion protein levels in thymocytes were about 3.4, 3, and 3.3 folds of the endogenous WT DGKζ2 in thymocytes (Figure 1D, 1E). Different from T cells, B220+ B cells did not show obvious GFP expression (Figure 1G). Expression of TCRβ in DN, DP, and SP thymocytes were not obviously affected by these DGKζ transgenes (Figure 1H). Together, these results demonstrated successful generation of these conditional GFP-DGKζ transgenic mice and that NLS and KD mutations did not obviously affect DGKζ stability.
Inhibition of T cell maturation by DGKζ ΔNLS mutant
DAG-mediated signaling and its tight regulation by DGKζ play important roles in T cell maturation in the thymus [8, 51, 52]. Among DAG effector pathways, the RasGRP1-Ras-Erk1/2 pathway mediates positive selection of DP thymocytes and maturation to CD4+ or CD8+ single positive (SP) stage [51, 52]. In GFP-DGKζWT-CD4Cre mice, total thymocyte numbers were similar to WT controls (Figure 2A). TCRβ+ thymocyte percentages and numbers were also comparable to WT control mice (Figure 2B, 2C). Similar to GFP-DGKζWT-CD4Cre mice, GFP-DGKζKD-CD4Cre mice did not displayed obvious differences to WT control mice in total thymic cellularity and TCRβ+ thymocyte numbers. Interestingly, GFP-DGKζΔNLS-CD4Cre mice contained slightly increased total thymocyte numbers with 40% increases compared to WT control mice. However, the percentages and numbers of TCRβ+ thymocytes were decreased, suggesting impaired T cell generation in these mice (Figure 2B, 2C).
Figure 2. Impaired T cell generation by DGKζΔNLS.
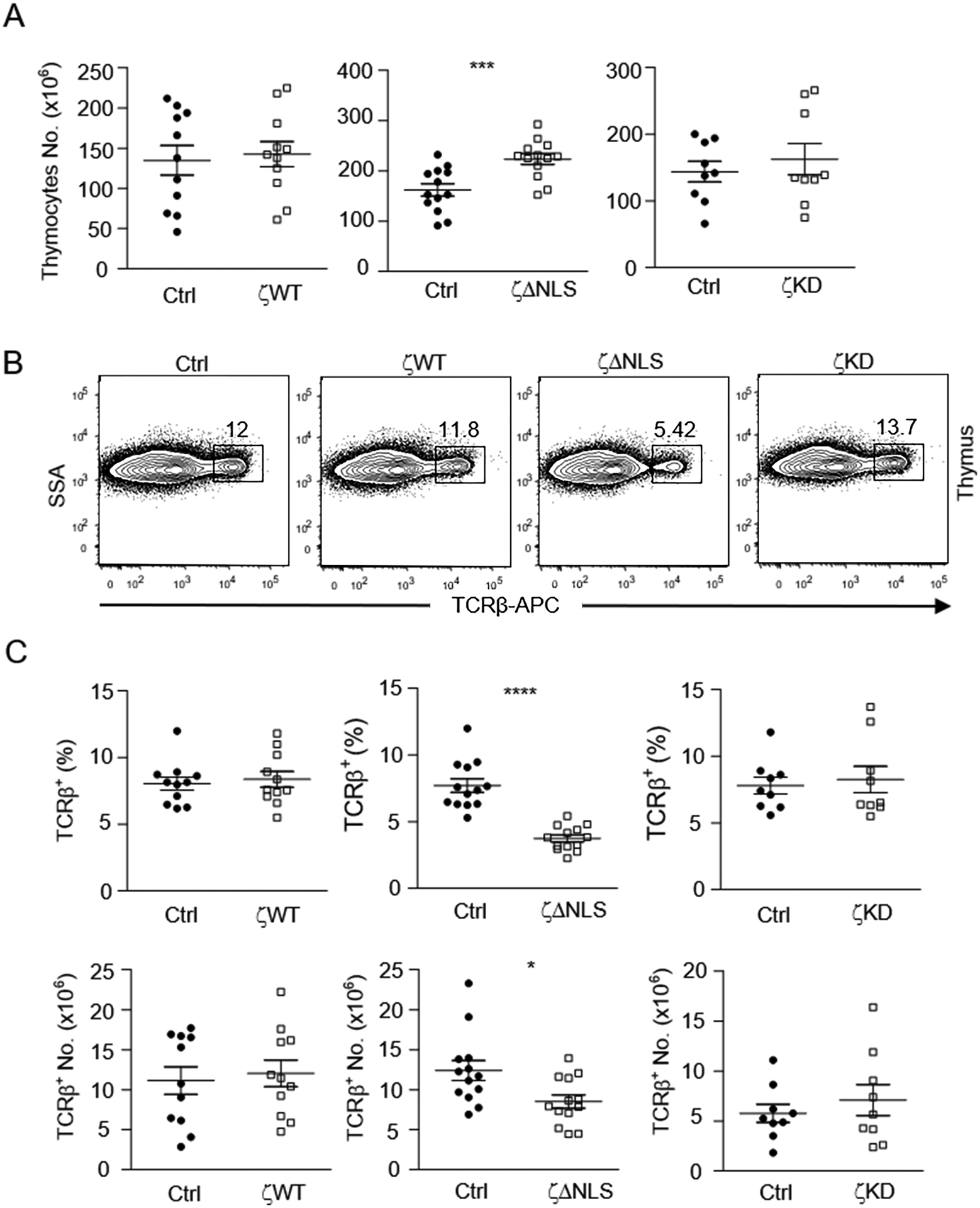
Thymocytes from 6 – 10 weeks old GFP-DGKζWT-CD4Cre, GFP-DGKζKD-CD4Cre, GFP-DGKζΔNLS-CD4Cre, and control mice were examined. A. Total thymocyte numbers. B. Representative flow cytometry plots showing TCRβ staining in thymocytes. C. Percentages and numbers of TCRβ+ thymocytes. Data shown are representative of (B) or pooled (A, C) from at least eight experiments and are shown as mean ± SEM. Each circle or square represents one mouse of the indicated genotypes. *, p<0.05; ***, p<0.001; ****, p<0.0001 determined by Student t-test. (N=11 for both DGKζWT-CD4Cre and control mice, N=13 for both DGKζΔNLS-CD4Cre and control mice, N=9 for both DGKζKD-CD4Cre and control mice)
T cells mature in the thymocytes sequentially through the CD4−CD8− DN, CD4+CD8+ DP, to the CD4+ or CD8+ SP stages. In GFP-DGKζWT-CD4Cre mice and GFP-DGKζKD-CD4Cre mice, the percentages and numbers of DN, DP, and SP thymocytes were not obviously different from control mice, indicating no gross blockade of T cell maturation (Figure 3A, 3B, 3D). However, in GFP-DGKζΔNLS-CD4Cre mice, CD4+CD8−TCRβ+ and CD4−CD8+TCRβ+ SP mature T cells were decreased in both percentages and numbers (Figure 3A, 3C). Within DP thymocytes, the percentages and numbers of the post-selection CD69+TCRβ+ cells were decreased in GFP-DGKζΔNLS-CD4Cre mice (Figure 3E), suggesting impairment of positive-selection of developing thymocytes. The decreases of CD4+ SP thymocytes (69.5%) appeared more severe than the decreases of CD8+ SP thymocytes (32.4%), suggesting differentiation and/or positive selection of CD4+ T cell lineage was more sensitive to GFP-DGKζΔNLS than CD8+ T cell lineage. In contrast to decreases of SP thymocytes, DP thymocytes in GFP-DGKζΔNLS-CD4Cre mice were slightly increased about 39% (Figure 3D). Consistent with impaired maturation, both percentages and numbers of CD4 T cells in the spleen and lymph nodes (LNs) from GFP-DGKζΔNLS-CD4Cre mice but not from GFP-DGKζWT-CD4Cre mice and GFP-DGKζKD-CD4Cre mice were decreased compared with control mice (Figure 3F–3H). The decreases of peripheral CD8 T cells were subtler than CD4 T cells. Together, these data revealed that GFP-DGKζΔNLS but not GFP-DGKζWT or GFP-DGKζKD were capable of inhibiting T cell maturation from DP to SP stages.
Figure 3. Inhibition of T cell maturation by DGKζΔNLS.
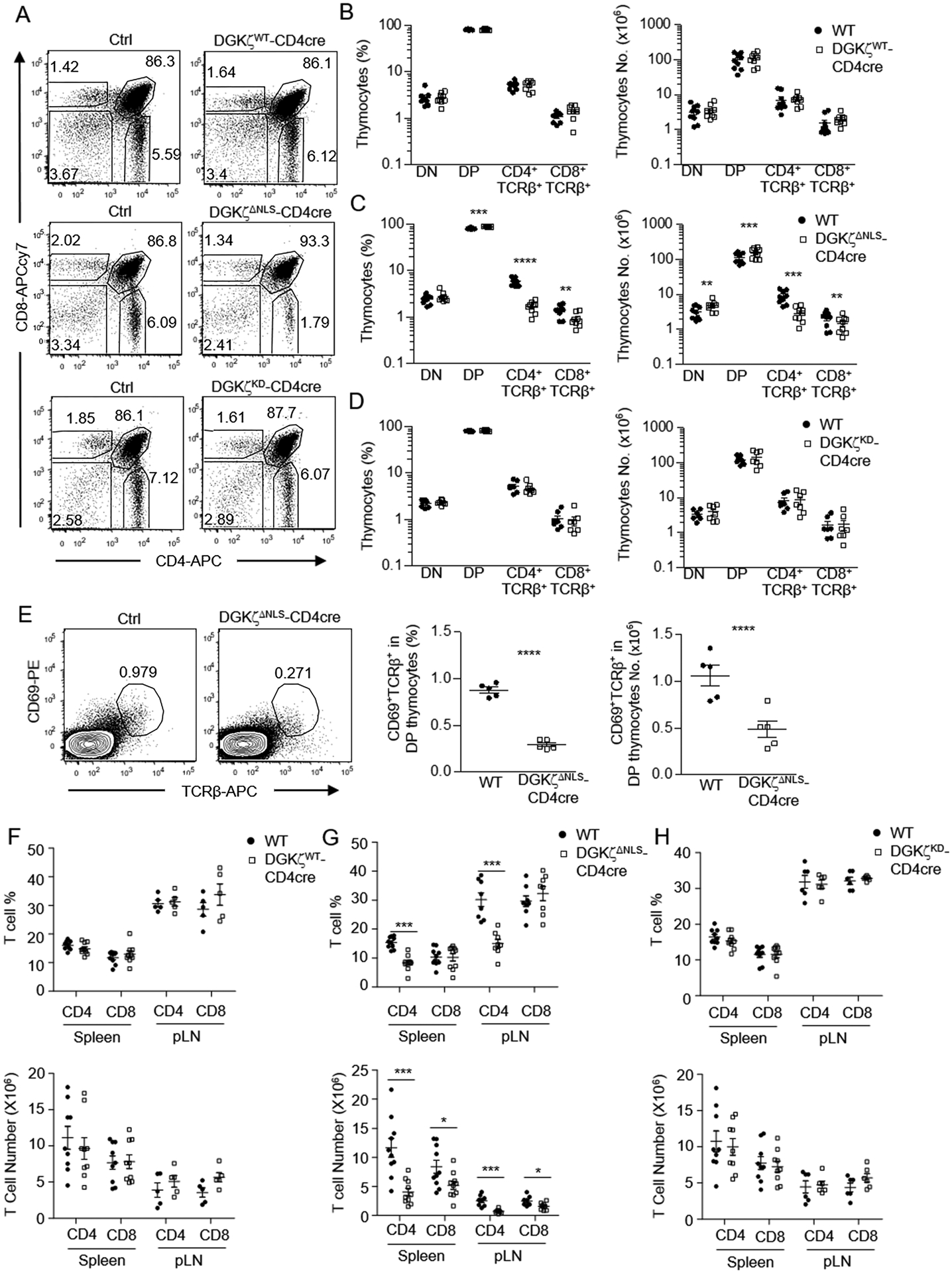
Thymocytes (A – D) and splenocyte (E – G) from 6 – 10 weeks old GFP-DGKζWT-CD4Cre, GFP-DGKζKD-CD4Cre, GFP-DGKζΔNLS-CD4Cre, and control mice were examined by flow cytometry. A. Representative flow cytometry plots showing CD4 and CD8 staining of thymocytes. B. Percentages and numbers of indicated thymocyte populations in GFP-DGKζWT-CD4Cre and control mice. C. Percentages and numbers of indicated thymocyte populations in GFP-DGKζΔNLS-CD4Cre and control mice. D. Percentages and numbers of indicated thymocyte populations in GFP-DGKζKD-CD4Cre and control mice. Data shown are representative of (A) and pooled from (B–D) at least seven experiments (N=9 in both B and C, N=7 in D) and error bars represent mean ± SEM. E. Representative flow cytometry plots showing CD69 and TCRβ staining of DP thymocytes. Scatter plots show percentages and numbers of CD69+TCRβ+ cells in DP thymocytes. Data shown are representative of (plot) or pooled from five experiments, N=5 for both WT and DGKζΔNLS-CD4Cre mice and error bars represent mean ± SEM. F. Percentages and numbers of CD4+ and CD8+ splenic and LN T cells in GFP-DGKζWT-CD4Cre and control mice. Data shown are pooled from at least five experiments, N=9 for both WT and GFP-DGKζWT-CD4Cre mice in spleen, and N=5 for both WT and GFP-DGKζWT-CD4Cre mice in pLN. G. Percentages and numbers of CD4+ and CD8+ splenic and LN T cells in GFP-DGKζΔNLS-CD4Cre and control mice. Data shown are pooled from at least eight experiments. N=10 for both WT and GFP-DGKζΔNLS-CD4Cre mice in spleen, and N=8 for both WT and GFP-DGKζΔNLS-CD4Cre mice in pLN. H. Percentages and numbers of CD4+ and CD8+ splenic and LN T cells in GFP-DGKζKD-CD4Cre and control mice. Data shown are pooled from at least six experiments. N=9 for both WT and GFP-DGKζKD-CD4Cre mice in spleen, and N=6 for both WT and GFP-DGKζKD-CD4Cre mice in pLN. Each circle or square represents one mouse of the indicated genotypes. (F–H) error bars represent mean ± SEM. *, p<0.05; **, p<0.01; ***, p<0.001; ****, p<0.0001, determined by Student t-test (Fig B, E, F, G, and H) or by Mann-Whitney test (Fig C and D).
Enhanced inhibition of TCR induced DAG signaling and Egr1 expression by DGKζΔNLS
Activation of RasGRP1 by DAG leads to Erk1/2 activation in T cells. Following anti-CD3 stimulation, thymocytes from GFP-DGKζWT-CD4Cre mice displayed decreased Erk1/2 phosphorylation (Figure 4A, 4B), suggesting that increased DGKζ expression inhibited activation of the RasGRP1-Ras-Erk1/2 pathway. Compared with GFP-DGKζWT-CD4Cre thymocytes, anti-CD3-induced Erk1/2 phosphorylation in GFP-DGKζΔNLS-CD4Cre thymocytes was further reduced. DAG-mediated RasGRP1-Ras-Erk1/2 functions upstream of both mTORC1 and mTORC2 activation after TCR engagement [21] and the DAG-PKCθ-CARMA1 pathway promoted mTORC1 activation [53]. DGKα and ζ deficiency led to enhanced TCR-induced mTORC1 and mTORC2 activation [21]. TCR induced S6 phosphorylation and Akt S473 phosphorylation, indicators of mTORC1 and mTORC2 activation respectively, were decreased slightly in GFP-DGKζWT-CD4Cre thymocytes but decreased more obviously in GFP-DGKζΔNLS-CD4Cre thymocytes. Together, these observations demonstrated that increased expression of DGKζ in developing thymocytes inhibited TCR-induced Erk1/2 and mTOR activation and that GFP-DGKζΔNLS mutant was a gain-of-function mutant with enhanced ability to terminate DAG signaling compared with WT DGKζ.
Figure 4. Enhanced inhibition of DAG-mediated signaling and downregulation of Egr1 expression by DGKζΔNLS.
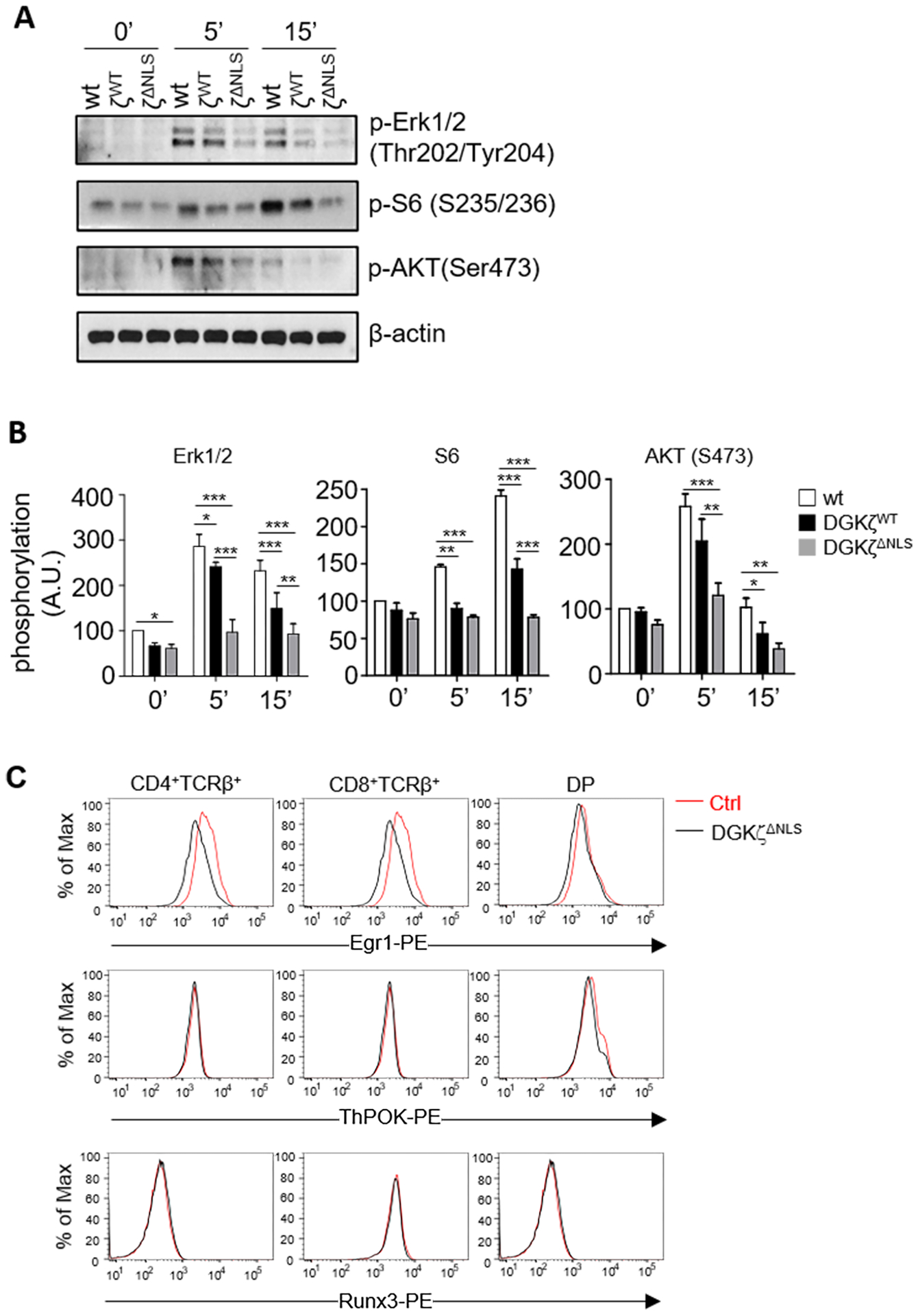
A, B. Thymocytes from WT, GFP-DGKζWT-CD4Cre, and GFP-DGKζΔNLS-CD4Cre mice were left unstimulated or stimulated with an anti-CD3 antibody (500A2) at 37°C for 5 or 15 minutes. Cell lysates were subjected to immunoblotting analyses using the indicated antibodies. β-actin was detected as loading controls (A). Quantification of relative intensity of western blot bands (B). Data shown are representative of (A) or pooled from (B) three experiments (N=3 for WT, GFP-DGKζWT-CD4Cre, and GFP-DGKζΔNLS-CD4Cre mice). Error bars represent mean ± SEM. C. Representative histograms showing Egr1, ThPOK, and Runx3 expression in DP, CD4SP, and CD8SP thymocytes from WT and GFP-DGKζΔNLS-CD4Cre mice. Each histogram represents 100% of the indicated cell population. Data shown are representative of four experiments. N = 4 mice for Ctrl and GFP-DGKζΔNLS-CD4Cre mice. *, p<0.05; **, p<0.01; ***, p<0.001; ****, p<0.0001, determined by Two-way ANOVA test.
During T cell development, the transcription factor Egr1 promotes positive selection of conventional T cells and iNKT cells downstream of the Ras/Erk1/2 pathway [54–56]. Both transcription factor ThPOK and Runx3 are critical for the development of CD4 and CD8 lineages, respectively [57–59]. Consistent with decreased Erk1/2 activation, Egr1 expression in the DGKζΔNLS-CD4Cre DP, CD4+, and CD8+ thymocytes was decreased compared with WT mice. However, ThPOK and Runx3 levels in these cells were similar to WT controls (Figure 4C). These results suggest that decreased Egr1 expression may contribute to impaired positive selection of both conventional T cells and iNKT cells as will be described later and that different sensitivities of CD4 and CD8 SP thymocytes to GFP-DGKζΔNLS might not be caused altered expression of ThPOK or Runx3.
DGKζΔNLS displays enhanced ability to inhibit T cell activation
DAG mediated signaling pathways are important for T cell activation. CD69 upregulation is one of the early T cell activation events that is dependent on activation of the DAG-RasGRP1-Ras-Erk1/2-AP1 (activator protein 1) pathway. T cells from GFP-DGKζWT mice showed reduced CD69 upregulation following anti-CD3 stimulation (Figure 5A, 5B). CD69 upregulation was further reduced in T cells from GFP-DGKζΔNLS-CD4Cre mice compared with GFP-DGKζWT-CD4Cre mice. In addition to CD69, CD25 is also upregulated during T cell activation. GFP-DGKζWT was able to weakly inhibit CD25 upregulation but GFP-DGKζΔNLS demonstrated stronger inhibition than GFP-DGKζWT (Figure 5A, 5B). Using a Celltrace violet proliferation dye dilution assay, we also observed weak inhibition of TCR-induced T cell proliferation by GFP-DGKζWT but much stronger inhibition of proliferation of both CD4 and CD8 T cells by GFP-DGKζΔNLS (Figure 5C). Moreover, GFP-DGKζΔNLS inhibited T cell production of IL2 after TCR stimulation (Figure 5D). Such inhibitory effects of GFP-DGKζΔNLS displayed diminished magnitudes when stimulated with both anti-CD3 and anti-CD28 antibodies and were abolished. when stimulated with PMA and ionomycin (Figure 5D, Supplemental Figure S2A–2C), which was consistent with the ability of PMA to resistant to DGK activity. Different from GFP-DGKζΔNLS and GFP-DGKζWT, GFP-DGKζKD did not obviously affected TCR induced Erk1/2 phosphorylation, CD69 or CD25 upregulation, and proliferation of T cells (Supplemental Figure S3A – 3D), indicating that the inhibitory activity of DGKζ was dependent of its kinase activity. Together, these observations indicate that GFP-DGKζΔNLS contained enhanced capability to inhibit T cell activation compared with GFP-DGKζWT.
Figure 5. Enhanced inhibition of T cell activation by DGKζΔNLS.
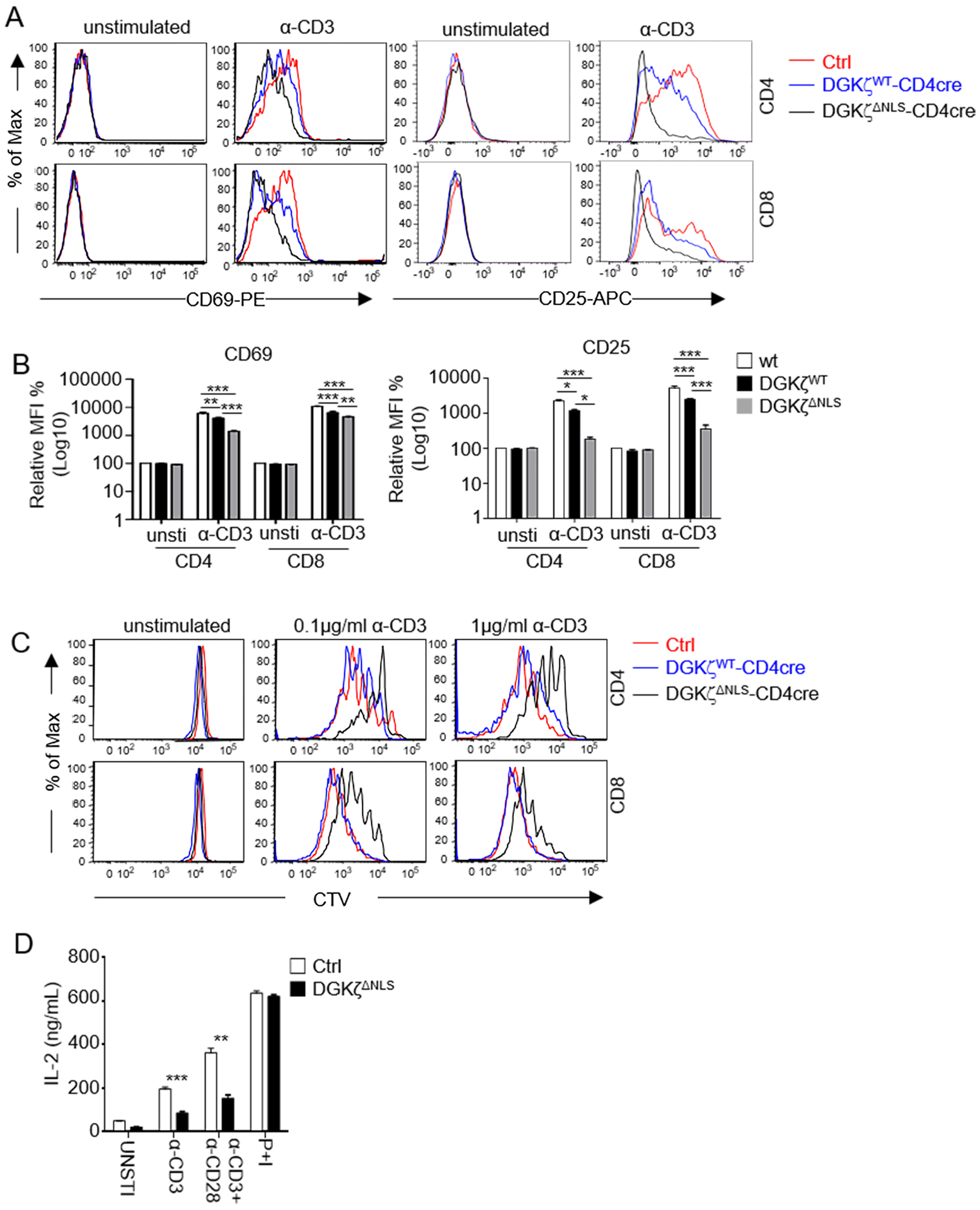
A. Representative flow cytometry of overlaid histograms of CD69 and CD25 expression in WT, DGKζWT, and DGKζΔNLS CD4 and CD8 T cells after overnight incubation of splenocytes in the presence or absence of an anti-CD3 antibody (0.1 μg/ml). Each histogram represents 100% of the indicated cell population. B. Relative MFI of CD69 and CD25 in the indicated T cells. N = 3 for Ctrl, DGKζWT-CD4Cre and GFP-DGKζΔNLS-CD4Cre mice. Data shown are representative of (A) or pooled from (B) three experiments, N = 3 for Ctrl, GFP-DGKζWT-CD4Cre, and GFP-DGKζΔNLS-CD4Cre mice. Error bars represent mean ± SEM. C. Representative flow cytometry overlaid histograms showing WT, DGKζWT, and DGKζΔNLS CD4 and CD8 T cell proliferation after in vitro stimulation for 72 hours with indicated concentrations of anti-CD3 antibodies. Data shown are representative of three experiments, N = 3 for Ctrl, GFP-DGKζWT-CD4Cre, and GFP-DGKζΔNLS-CD4Cre mice. Each histogram represents 100% of the indicated cell population. D. ELISA for IL-2 concentrations in the supernatant from LN T cells with indicated-treatments for 48 hours. Data are pooled from three experiments. N = 3 for both WT and GFP-DGKζΔNLS-CD4Cre mice. Error bars represent mean ± SEM. *, p<0.05; **, p<0.01; ***, p<0.001; ****, p<0.0001, determined by Two-way ANOVA test (Fig B) or by Student t-test (Fig D).
Inhibition iNKT cell maturation by GFP-DGKζWT and GFP-DGKζΔNLS
RasGRP1-Ras-Erk1/2 and PKCθ-IKK-NFκB pathways as well as mTOR signaling are important for iNKT cell maturation [33, 34, 56, 60–68]. Because DGKα and ζ inhibit activation of these signal events [6, 21, 53], we further examined the effects of DGKζ mutants on iNKT cell development. We observed about 50% decreases of TCRβ+PBS57 loaded CD1d tetramer (CD1Dtet)+ iNKT cells in the thymus, spleen, and liver in GFP-DGKζWT-CD4Cre mice (Figure 6A, 6B) but not in GFP-DGKζKD-CD4Cre mice (Figure 6C, 6D) compared with their respectively WT controls, indicating kinase activity dependent inhibition of iNKT cell development by GFP-DGKζWT. The decreases of iNKT cells were much severer in GFP-DGKζΔNLS-CD4Cre mice with 92.4, 92.2, and 87.5% reduction of iNKT cells in the thymus, spleen, and liver, respectively, compared with WT controls (Figure 6E, 6F). Together, these data indicated that iNKT cell development was sensitive to increased DGK activity. The much severer iNKT cell developmental blockade in GFP-DGKζΔNLS-CD4Cre mice than in GFP-DGKζWT-CD4Cre further supported that DGKζΔNLS is a gain-of-function mutation.
Figure 6. Effects of DGKζWT and DGKζΔNLS on iNKT cell development.
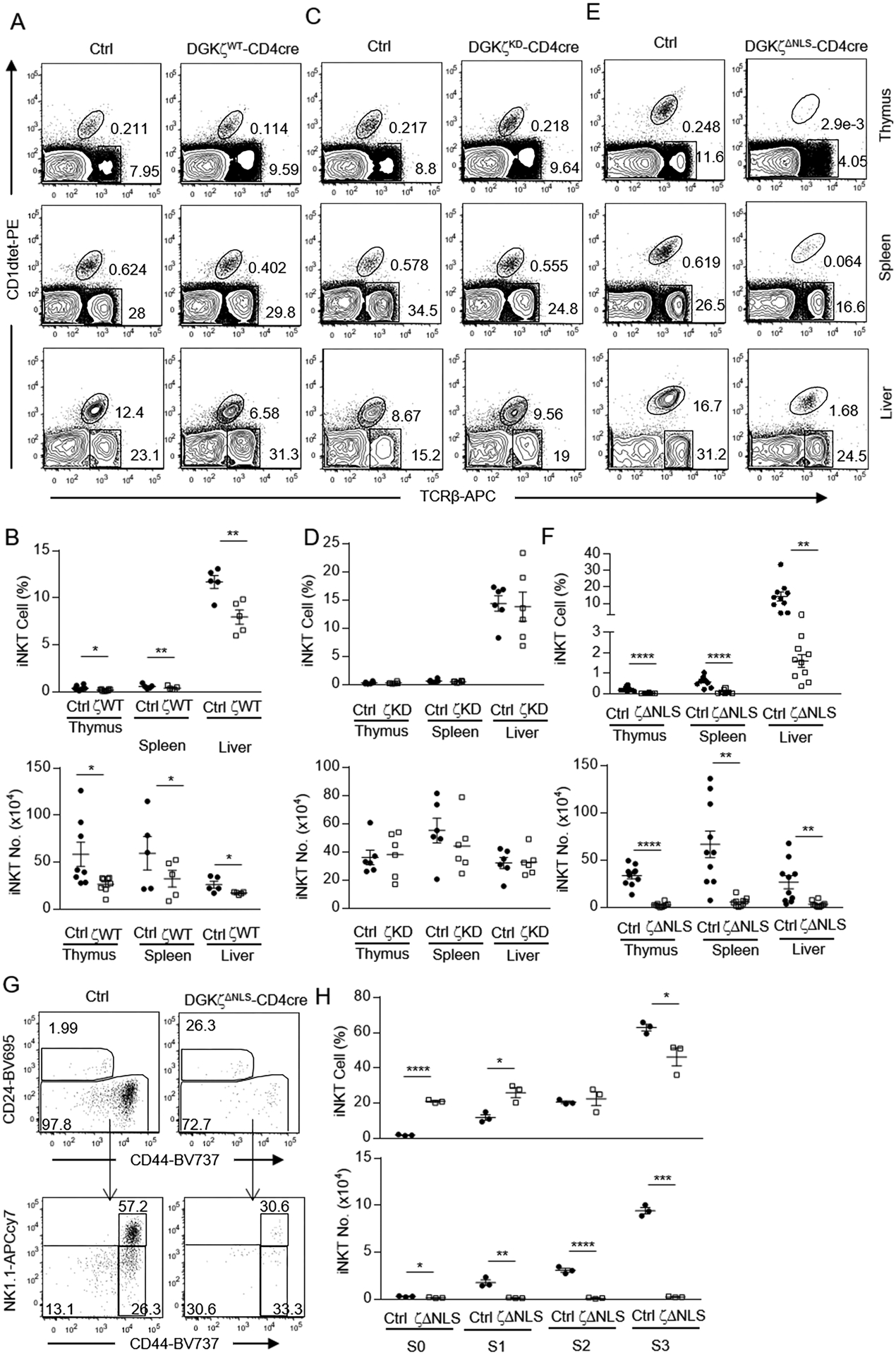
Thymocytes, splenocytes, and liver mononuclear cells from 6 – 10 weeks old GFP-DGKζWT-CD4Cre, GFP-DGKζKD-CD4Cre, GFP-DGKζΔNLS-CD4Cre, and control mice were examined. A, C, E. Representative flow cytometry plots showing TCRβ and PBS57-loaded CD1d tetramer (CD1dTet) staining in live gated cells from thymocytes, splenocytes, and liver mononuclear cells from GFP-DGKζWT-CD4Cre and control mice (A), GFP-DGKζKD-CD4Cre and control mice (C), GFP-DGKζΔNLS-CD4Cre and control mice (E). B, D, F. Percentages and numbers of iNKT cells in the indicated organs and mice. Data shown are representative of (A) and pooled from (B) at least five experiments and are shown as mean ± SEM. For the experiment of iNKT cell percentages and numbers in thymus, N=8 for both Ctrl and DGKζWT-CD4Cre mice. For the experiment of iNKT percentages and numbers in both spleen and liver, N=5 for both Ctrl and DGKζWT-CD4Cre mice. Data shown are representative of six (C) or ten (E) and pooled from six (D) or ten (F) experiments and are shown as mean ± SEM. Each experiment included one ctrl and one test mice. G. Representative flow cytometry plots showing CD24 and CD44 expression in live gated Lin (CD11b, CD11c, B220, Ter119, F4/80)− iNKT cells in the thymus (top panels) and CD44 and NK1.1 expression in CD24− iNKT cells (bottom panels). H. Percentages and numbers of indicated iNKT cell stages. CD24+CD44− stage 0 iNKT cells were calculated after exclusion of NK1.1+ cells. Data shown in G are representative of at least three experiments. Data shown in H are calculated from one experiment with three pairs of test and control mice and are shown as mean ± SEM. N = 3 for both Ctrl and GFP-DGKζΔNLS-CD4Cre mice. Each circle or square represents one mouse of the indicated genotypes. *, p<0.05; **, p<0.01; ***, p<0.001; ****, p<0.0001 determined by Student t-test.
iNKT cell development in the thymus can be defined into CD24+CD44−NK1.1− stage 0, and CD24−CD44−NK1.1− stage 1, CD24−CD44+NK1.1− stage 2, and CD24−CD44+NK1.1+ stage 3 [69, 70]. We observed decreases of iNKT cells in the thymus of GFP-DGKζΔNLS-CD4Cre mice began at stage 0 and became more obvious at stages 1 – 3 (Figure 6G, 6H), suggesting that DGKζΔNLS might inhibit not only iNKT cell development at the earliest stage but also the accumulation/expansion of late stage iNKT cells.
Dynamic nucleus localization of DGKζ in the activated-T cells during TCR stimulation
To explore the relationship between DGKζ function and subcellular localization, we examined GFP-DGKζ localization in peripheral T cells from GFP-DGKζWT-CD4Cre, GFP-DGKζΔNLS-CD4Cre, and GFP-DGKζKD-CD4Cre mice. In unstimulated T cells, GFP-DGKζWT and GFP-DGKζKD were detected in both the cytosol and the nucleus, with stronger fluorescence intensity in the cytosol than in the nucleus (Figure 7A, 7B). During anti-CD3 stimulation, GFP-DGKζWT and GFP-DGKζKD began to accumulate in the nucleus at 10 minutes after stimulation and such nuclear accumulation of DGKζ appeared intensified 30’ and 4 hours after stimulation and maximized after overnight stimulation, suggesting that the kinase activity did not contribute to TCR-induced DGKζ nuclear translocation. However, GFP-DGKζΔNLS was predominantly localized in the cytoplasmic compartment with little nuclear localization in unstimulated T cells. Moreover, it did not translocate into the nucleus after anti-CD3 stimulation. To rule out the possibility that DGKζ localization was influenced by GFP, we examined endogenous DGKζ localization in WT T cells. We observed that endogenous DGKζ mainly localized in the cytosol before stimulation and translocated to the nucleus after overnight TCR stimulation (Figure 7C, 7D). Thus, both endogenous DGKζ and GFP-DGKζ localized similarly in the cytosol in unstimulated state and translocated into the nucleus after TCR stimulation.
Figure 7. TCR-induced nuclear localization of DGKζ.

A. LN T cells from GFP-DGKζWT-CD4Cre, GFP-DGKζΔNLS-CD4Cre, and GFP-DGKζKD-CD4Cre mice were stimulated with or without anti-CD3 (145–2C11) for the indicated times, followed in fixation/permeabilization, staining with an Alexa 488 conjugated anti-GFP antibody, and confocal microscopy. Objective magnification: 63×; Zoom: 6×; Scale bar: 8 μM. Representative images are shown. B. Quantification of anti-GFP intensity in the cytosol and nucleus. Each dot shows nuclear/cytoplasm ratio of MFI of GFP in a T cell. C. WT LN CD4+ T cells were unstimulated or stimulated with a plate-bound anti-CD3 antibody (145–2C11) for 22 hours, followed sequential by fixation/permeabilization, staining with a rabbit anti-DGKζ antibody and then a FITC conjugated goat anti-rabbit IgG antibody, and visualization by confocal microscopy. Objective magnification: 63×; Zoom: 6×; Scale bar: 8 μM. Representative images are shown. D. Quantification of FITC intensity in the cytosol and nucleus. Each dot shows nuclear/cytoplasm ratio of MFI of FITC in a T cell. Figure A and B are representative of or pooled from four experiments for GFP-DGKζWT-CD4Cre and GFP-DGKζΔNLS-CD4Cre mice and three experiments for GFP-DGKζKD-CD4Cre mice. N = 1 for each genotype in each experiment. Figure C and D are representative of or pooled from three experiments. One WT mouse was used for each experiment. Error bars represent mean ± SEM. *P < 0.05; **P < 0.01; ***P < 0.001; **** P < 0.0001, determined by Mann-Whitney test (Fig B) or by the Student t test (Fig D).
Because primary T cells had very small cytosolic compartment and were difficult to distinguish cytosol and cytoplasm membrane, we established 2B4 T cell lines that expressed either GFP-DGKζWT or GFP-DGKζΔNLS to examine cytosolic and plasma membrane distribution of DGKζ during TCR engagement. As shown in figure S4A – S4C, both DGKζWT and DGKζΔNLS were detected in the cytosol and cytoplasm membrane before TCR stimulation. After overnight anti-CD3 stimulation, DGKζWT translocated into the nucleus or presented at the nucleus envelope. However, the DGKζΔNLS mostly presented at the cytoplasm membrane.
Taken together, our results indicated that WT DGKζ localization was dynamically regulated. It was predominantly localized in the cytosolic compartment in resting state and dynamically translocated into the nucleus after TCR stimulation and the NLS was crucial for DGKζ nuclear localization to occur. When NLS was mutated, DGKζ displayed increased tendency to localize in the cytosol and at the cytoplasmic membrane. Collectively, these observations together with enhanced inhibition of DAG-mediated signaling, T cell activation, and T cell development by GFP-DGKζΔNLS indicated that DGKζ nuclear translocation from the cytosol may sequester it from access to its substrates in the cytoplasmic compartment and, especially, at the cytoplasm membrane and thus functions as a negative control mechanism of DGKζ function during T cell development and activation.
DISCUSSION
DGK isoforms are differentially expressed in various tissues to regulate diverse cellular processes. Given their important physiological functions, DGKs should be tightly regulated. Although both transcriptional and post-transcriptional mechanisms have been reported to control the expression and function of specific isoforms in multiple cell line models [17, 71, 72], mechanisms that regulate DGKs and the physiological importance of such regulation, especially in vivo settings, are still poorly understood.
In this study, we have focused on DGKζ because its predominant roles in several immune cell lineages. Previous studies have found that DGKζ is dynamically regulated in T cells with high expression in naïve resting T cells, anergic T cells, and tumor infiltration T cells but downregulated expression during T cell activation [15, 16, 72]. High levels of DGKζ in naïve T cells might be important to prevent DAG accumulation in these cells so that tonic TCR signal would not trigger spontaneous T cell activation; while downregulation of DGKζ expression during T cell activation may allow full activation of these cells. Such dynamic expression of DGKζ is in part controlled by miR34a that directly targets DGKζ [72]. Elevated DGKζ in anergic and tumor-infiltrating T cells may contribute to their impaired anti-tumor immunity [23, 73]. To explore additional mechanisms that regulate DGKζ function, we have generated three lines of mice with conditional expression of GFP-DGKζWT, GFP-DGKζKD, and GFP-DGKζΔNLS in developing thymocytes and peripheral T cells. We have demonstrated that expression of GFP-DGKζWT but not GFP-DGKζKD weakly inhibits TCR-induced Erk1/2 activation, CD69 and CD25 upregulation, and proliferation of T cells and causes moderate reduction of iNKT cells but not conventional αβT cells. These data not only extend previous studies in cell line models that overexpression of DGKζ inhibits TCR signaling and upregulation of activation markers to primary cells but also reveals important consequences when DGKζ is overexpressed in vivo and revealed that iNKT cell development is more sensitive to altered DGKζ activity than cαβT cells. More importantly, we show that GFP-DGKζΔNLS displays much stronger effects on T cells than GFP-DGKζWT in inhibiting DAG-mediated signaling, T cell activation, and iNKT cell development. Moreover, GFP-DGKζΔNLS but not GFP-DGKζWT is able to impede conventional αβT cell maturation. Based on these observations, we conclude that DGKζΔNLS is a gain-of-function mutant in developing thymocytes and peripheral T cells with enhanced ability to suppress DAG-mediated signaling.
Subcellular localization of DGKζ varies dramatically among different cell types. It has been shown to localize predominantly in the nucleus, in the cytosolic compartment, or distribute in both nucleus and cytosol dependent on different cell types and tissues examined [41, 44, 47, 74–77]. Our data indicate that DGKζ function is dynamically regulated in T cells via shuttling between the cytosol and nucleus and that nuclear localization of DGKζ serves as a negative control of DGKζ function by preventing it from accessing its substrate in the cytoplasmic membrane. In primary T cells, GFP-DGKζWT as well as endogenous DGKζ preferentially localizes in the cytosolic compartment with a small portion of the protein residing in the nucleus in unstimulated primary T cells. TCR stimulation induces rapid DGKζ translocation and accumulation in the nucleus accompanying concordant depletion in the cytosol. An event critical for TCR-induced T cell activation is the activation of PLCγ1 and subsequent production of DAG. Newly generated DAG localizes in the cytoplasmic membrane, particularly in the immune synapse [78]. Nuclear translocation of DGKζ during T cell activation may prevent it from accessing the newly generated DAG to ensure DAG-mediated activation of downstream signal cascades for T cell activation. Failure of nuclear translocation of the DGKζΔNLS mutant allows for its persistence in the cytosol, leading to accelerated termination of DAG generated after TCR engagement.
It is interesting to note that DGKζ is mainly localized in the nucleus in several other cell types such as neurons and myoblasts. Nuclear localization of DGKζ in these cells is important for its stability as cytosolic DGKζ is subjected to ubiquitination and proteasomal degradation and for its ability to negative control of cell cycle [44, 48, 49, 71, 79, 80]. Different from these cells, both endogenous DGKζ and GFP-DGKζ are primarily localized in the cytosol in primary T cells in the quiescent state but translocate into the nucleus during T cell activation. In addition, we do not observe decreased expression of GFP-DGKζΔNLS compared with GFP-DGKζWT. Such differences between T cells and neurons as well as several other cell types suggest that T cells utilize different mechanisms to control DGKζ localization and stability.
Activation of the RasGRP1-Ras-Erk1/2 pathway is critical for positive selection of DP thymocytes to develop into SP mature T cells as well as iNKT cells [34, 51, 56]. Decreases of CD69+TCRβ+ post-selection DP thymocytes and SP mature T cells by GFP-DGKζΔNLS suggests that positive selection is impaired. These data indicate that certain levels of DAG are needed in DP thymocytes for successful execution of positive selection and DGKζΔNLS may deplete the DAG pool necessitated for activation of the RasGRP1-Ras-Erk1/2 pathway for positive selection. Of note, although GFP-DGKζWT weakly inhibits DAG-mediated signaling and T cell activation, it does not cause obvious reduction of SP thymocytes, which could be resulted from alterations of T cell repertoire to compensate for reduced DAG levels. Different from conventional αβT cells, iNKT cells express an invariant TCR-Vα14 chain and limited TCRVβ chains. Their semi-fixed TCR repertoire may limit their ability to accommodate slightly weakened DAG signaling caused by GFP-DGKζWT, leading to their hypersensitivity to altered DAG signaling during development.
Although we consider the impact of DGKζΔNLS is mainly caused by enhanced terminating DAG due to its failure to shuttle to the nucleus, our data do not rule out that DGKζΔNLS may act through other mechanisms. DGKζΔNLS may overproduce PA in the cytoplasmic membrane, which may itself have important biological consequences. It has also been reported that cytoplasmic DGKζ reduces p53-mediated cytotoxicity against doxorubicin-induced DNA damage by facilitating cytoplasmic anchoring and degradation of p53 through an ubiquitin-proteasome system [41]. DGKζ also interacts with the adaptor molecules γ1-syntrophin, which links ion channels and signaling proteins to dystrophin and its homologues, and such interaction is important for γ1-syntrophin translocation to the nucleus and for anchoring DGKζ to the cytoplasm membrane [75, 81]. Accumulation of DGKζΔNLS in the cytosol during T cell activation could potentially affect these molecules and exert other functions besides regulating DAG signaling. Additionally, DGKζ may perform important functions inside the nucleus. The nucleus contains different pools of DAG and nuclear DGKζ has been shown to reduce nuclear DAG concentration in HEK293 cells [44]. In the nucleus of skeletal muscle, nuclear DGKζ interacts with the class O of forkhead box transcription factor (FoxO) to mitigate FoxO-mediated ubiquitin-proteasome system dependent proteolysis to prevent muscle atrophy, which is independent on its kinase activity but dependent on its nuclear localization [77]. Whether nuclear DGKζ performs similar functions in T cells remains to be determined. Because the endogenous DGKζ, which co-expresses with GFP-DGKζΔNLS, could still translocate to the nucleus, it is unlikely that nuclear DGK activity would be defective in GFP-DGKζΔNLS expressing T cells to cause the phenotypes we have observed.
An important question remains to be addressed is how DGKζ shuttling between the cytosol and nucleus is regulated. In HEK293 cells, DGKζ nuclear localization is regulated by PKC-mediated phosphorylation in its MARCKS motif [44]. Because the MARCKS motif and NLS overlap within DGKζ, phosphorylation of the serine residues within MARCKS motif may control DGKζ cytosol/nucleus shuttling directly or via binding to yet to be identified proteins. Additionally, the K to A mutations in DGKζΔNLS could also influence MARCKS motif phosphorylation, which needs to be determined in the future. It has also been reported that nucleosome assembly protein 1-like 1 (NAP1L1) and NAP1L4 shuttle between the nucleus and the cytoplasm in HEK293 cells and they interact with DGKζ to prohibit its nuclear import via blocking its association with import carrier proteins, Qip1 (importin α3) and NPI1 (importin α5) [82]. Future studies should determine whether these mechanisms are involved regulating DGKζ subcellular localization in T cells.
In summary, our data demonstrate that DGKζ nuclear localization serves as a mechanism to negatively control DGKζ function during T cell development and activation. We propose that manipulating cytosolic and nuclear DGKζ levels as a novel strategy to enhance or inhibit T cell activation for cancer immunotherapy or for treatment of autoimmune diseases. To achieve this, future studies should illustrate mechanisms that control DGKζ nuclear localization.
MATERIALS AND METHODS
Mice
Cd4Cre mice were acquired from Charles River Laboratory. Coding sequences for GFP-DGKζWT, GFP-DGKζKD, and GFP-DGKζΔNLS fusion proteins were generated by attaching WT and mutant DGKζ1 coding sequence to the carboxyl terminus of EGFP and subsequently cloned into the Asc I site of the Rosa26-hCD2 targeting vector using standard molecular cloning approaches. DGKζKD contains a G355D mutation in the ATP binding site within the catalytic domain. DGKζΔNLS contains L255KASKKKKRASFKRKSSKKG274 to LAASAAAARASFAAASSAAG in its NLS that overlaps with the MARCKS motif. Linearized targeting vectors were transfected into the C57BL/6N ES cells. Successfully targeted clones, identified by PCR screen, were used to generate chimeric founder mice. Founders were bred with Cd4Cre (B6.Cg-Tg(CD4-cre)1Cwi N9) mice [83] to induce GFP-DGKζ expression in T cells. All animal experiments were performed according to protocols approved by the Institutional Animal Care and Use Committee of Duke University.
Antibodies, tetramers, and flow cytometry
Fluorochrome-conjugated TCR-β (clone H57–597), NK1.1 (clone PK136), CD62L (clone MEL-14), CD44 (clone IM7), CD24 (clone M1/69), CD3e (clone 145–2C11), CD69 (clone H1.2F3), CD25 (clone 3C7), CD4 (clone GK1.5), CD8a (clone 53–6.7), ThPOK (clone 11H11A14), and CD73 (clone TY/11.8), Ultra-LEAF™ purified anti-mouse CD3 antibody (clone 145–2C11), and anti-mouse CD28 antibody (clone 37.51) were purchased from Biolegend. Fluorochrome-conjugated Egr1 (clone 44D5) antibody was purchased from Cell Signaling Technology. PE- or APC-conjugated PBS57 loaded CD1d tetramer was kindly provided by the NIH tetramer facility. Cell death was identified using the Live/Dead Fixable Violet Dead Cell Stain (ThermoFisher) or 7-AAD. Rabbit anti-DGKζ monoclonal antibody (ab239080) was purchased from Abcam. FITC-labeled goat anti-rabbit IgG (31635) and Alexa 488 conjugated anti-GFP antibody (A21311) was purchased from ThermoFisher. Anti–phospho-Erk1/2 T202/Y204 (D13.14.4E), phospho-S6 S235/236 (2F9), and phospho-Akt S473 (193H12) were from Cell Signaling Technology. Anti–β-actin was from Sigma-Aldrich. Single cell suspensions of the thymus, spleen, lymph nodes (LNs), and liver mononuclear cells (MNCs) in complete IMDM media (IMDM-10, 10% FBS, penicillin/streptomycin, 50 μM 2-mercaptoethanol) were prepared as previous described [34]. Cells were stained with fluorescence conjugated antibodies and tetramers in 2% (vol/vol) FBS in PBS at 4°C for 30 min. Data were collected using BD FACSCanto II and BD LSRFortessa X-20 flow cytometer and analyzed with Flowjo. Flow cytometry analyses were adhered to the ‘Guidelines for the use of flow cytometry and cell sorting in immunological studies’ [84]. Gating strategies are shown in supplemental Figure S1.
Establishing GFP-DGKζWT and GFP-DGKζΔNLS T cell lines.
Retrovirus-mediated gene transfer was used to express GFP-DGKζWT and GFP-DGKζΔNLS proteins in the 2B4 cell line. GFP-DGKζWT and GFP-DGKζΔNLS coding sequences were cloned in a retroviral vector, Migr1. Retroviral vectors were transfected into the Phoenix-Eco packaging cell line using the calcium phosphate method. Viral supernatants were collected 48 hours after transfection. For infection, 2B4 cells were mixed with viral supernatants and 5 μg/ml of polybrene (Sigma-Aldrich). Cells were then centrifuged at 2500g for 90 minutes at room temperature. The culture medium was changed with fresh RPMI 1640 (containing 10% FBS) 7 and 24 hours after infection. GFP+ cells were sorted using SONY SH800 sorter 48 hours after infection.
Immunoblotting
Thymocytes were rested in Dulbecco’s PBS containing calcium and magnesium at 37°C for 30 minutes, followed by stimulation with 5μg/mL of anti-CD3ε (500A2) or PMA (10 ng/ml) plus ionomycin (1 μg/ml) for the indicated times. After stimulation, cells were lysed in RIPA lysis buffer with freshly added protease and phosphatase inhibitors and were then subjected to SDS/PAGE and Western blot analysis with the indicated antibodies.
T cell stimulation in vitro
Splenocytes or LN cells were either labeled or unlabeled with CellTrace Violet (CTV) cell proliferation kit (C34571, ThermoFisher) or eFluor 670 cell proliferation dye (65-0840-85, ThermoFisher) following the manufacturer’s protocols. Cells were plated in a 96-well U bottom plate (0.5× 106), left unstimulated or stimulated with anti-CD3ε (145–2C11) at 37 °C. Unlabeled cells were stained for CD4, CD8, CD69, CD25, and 7AAD or live/dead dye for analysis of activation marker expression after overnight stimulation. CTV- or eFluor™ 670-labeled cells were stained for CD4 and CD8 to assess proliferation seventy-two hours after stimulation.
ELISA
LN T cells from WT and GFP-DGKζΔNLS mice were stimulated in the indicated conditions for 48 hours. IL-2 levels in the supernatants were measured using ELISA following the manufacture’s instruction (Biolegend, 431001).
Immunofluorescence and confocal microscopy
LN T cells in IMDM-10 from GFP-DGKζWT-Cd4Cre and GFP-DGKζΔNLS-Cd4Cre mice were stimulated with an anti-CD3ε antibody (145–2C11, 1μg/ml) in a U-bottom 96-well plate for 10 minutes, 30 minutes, 4 hours, or 22 hours. 2B4 cells that overexpressed GFP-DGKζWT or GFP-DGKζΔNLS were stimulated with plate bound anti-CD3ε antibody (145–2C11, 1μg/ml) for 22 hours. Cells were immediately subjected to fixation using cold methanol, blocked in 5% bovine serum albumin with 0.2% Triton X-100/PBS (30 min, on ice), then stained with Alexa 488 conjugated anti-GFP antibody (1:100 in PBS with 0.2% Triton X-100 and 2% FBS) overnight at 4°C. The nucleus was stained with DAPI (Biolegend). After staining, cells were washed in PBS, followed by cytospin and mounting. Images were captured using a Leica SP5 confocal microscope. GFP-DGKζWT, GFP-DGKζKD, and GFP-DGKζΔNLS intensity in the cytosol, cytoplasmic membrane, and nucleus was quantified using the ImageJ software. Mean fluorescence intensity (MFI) of GFP in the nuclear and cytoplasmic regions were measured and subtracted from the background. The localization index of GFP-DGKζ fusion proteins was calculated as the ratio of nuclear/cytoplasmic MFI in individual cells [85]. For endogenous DGKζ localization, WT LN CD4+ T cells purified using CD4 microbeads (130-117-043, Miltenyi Biotec) were stimulated with plate bound anti-CD3ε antibodies (145–2C11, 1μg/ml) in a flat-bottom 96-well plate for 22 hours. After fixation, blockage, and permeabilization, cells were stained with a rabbit anti-DGKζ primary antibody (ab239080) at 4°C overnight, cells were washed with 0.2% Triton X-100/PBS buffer twice, and then subjected to staining with FITC-labeled a goat anti-rabbit IgG secondary antibody at 4°C for 4 hours. Stained cells were washed in PBS, followed by cytospin and mounting and microscopic visualization.
Statistical analysis
Data were presented as mean ± SEM and analyzed for statistical differences using the Prism 6.01/GraphPad software. Scatter plots in each figure were pooled from multiple experiments. Data distribution was analyzed with Shapiro-Wilk test (if n⩾3) using SPSS Statistics 23 software, Kolmogorov-Smirnov test (if n⩾5) and D’Agostino & Pearson omnibus normality test (if n⩾8) using Prism 6.01/GraphPad software. Data with normal distribution were compared using two-tailed Student t-test or Two-Way ANOVA test. Other data were compared using a Mann-Whitney non-parametric test. P-values less than 0.05 were considered significant.
Supplementary Material
Acknowledgement
We thank Cheryl Bock, Meilang Flowers, and Julie Kent in the Transgenic and Knockout Mice facility at Duke Cancer Institute for generating DGKζ knocking-in mice, Dr. Matthew Topham for DGKζ plasmids, Dr. Jörn Coers for the Rosa26-hCD2 targeting vector, Dr. Warren Pear for the Migr1 vector, and the Flow Cytometry Core Facility and the Light Microscopy Core Facility at Duke Cancer Institute for the service. This study is supported in part by NIH R01AI079088 and R56AG060984 (XZ).
Abbreviation list:
- DAG
Diacylglycerol
- PA
phosphatidic acid
- DGK
Diacylglycerol kinase
- DP
double positive
- SP
signal positive
- pLN
peripheral lymph node
- BM
bone marrow
- cαβT cell
conventional αβT cells
- MNCs
mononuclear cells
- CTV
CellTrace Violet
- DKO
DGKα and ζ double deficient
- MARCKS
myristoylated alanine-rich C-kinase substrate
- NLS
nuclear localization sequences
- EGFP
enhanced green fluorescent protein
- iNKT
invariant NKT
- mTORC
mammalian target of rapamycin complex
- PI3K
Phosphoinositide 3-kinases
- RasGRP1
RAS guanyl nucleotide-releasing protein1
- Erk1/2
extracellular signal-regulated protein kinases 1 and 2
- NFκB
nuclear factor kappa-light-chain-enhancer of activated B cells
- IKK
IκB kinase
- PKC
Protein kinase C
- TCR
T-cell receptor
- CARMA1
Caspase recruitment domain-containing membrane-associated guanylate kinase protein-1
- PLCγ1
Phospholipase C, gamma 1
- FBS
fetal bovine serum
- DAPI
4’,6-diamidino-2-phenylindole
- MFI
Mean fluorescence intensity
- Ctrl
control
- ATP
adenosine triphosphate
- PCR
polymerase chain reaction
Footnotes
Conflict of Interest Disclosures
The authors declare no commercial or financial conflict of interests.
REFERENCES
- 1.Krishna S and Zhong X, Role of diacylglycerol kinases in T cell development and function. Crit Rev Immunol 2013. 33: 97–118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Merida I, Arranz-Nicolas J, Rodriguez-Rodriguez C and Avila-Flores A, Diacylglycerol kinase control of protein kinase C. Biochem J 2019. 476: 1205–1219. [DOI] [PubMed] [Google Scholar]
- 3.Mariathasan S, Zakarian A, Bouchard D, Michie AM, Zuniga-Pflucker JC and Ohashi PS, Duration and strength of extracellular signal-regulated kinase signals are altered during positive versus negative thymocyte selection. J Immunol 2001. 167: 4966–4973. [DOI] [PubMed] [Google Scholar]
- 4.Alberola-Ila J and Hernandez-Hoyos G, The Ras/MAPK cascade and the control of positive selection. Immunol Rev 2003. 191: 79–96. [DOI] [PubMed] [Google Scholar]
- 5.Goplen N, Karim Z, Guo L, Zhuang Y, Huang H, Gorska MM, Gelfand E, et al. , ERK1 is important for Th2 differentiation and development of experimental asthma. FASEB J 2012. 26: 1934–1945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Chen SS, Hu Z and Zhong XP, Diacylglycerol Kinases in T Cell Tolerance and Effector Function. Front Cell Dev Biol 2016. 4: 130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sun Z, Arendt CW, Ellmeier W, Schaeffer EM, Sunshine MJ, Gandhi L, Annes J, et al. , PKC-theta is required for TCR-induced NF-kappaB activation in mature but not immature T lymphocytes. Nature 2000. 404: 402–407. [DOI] [PubMed] [Google Scholar]
- 8.Guo R, Wan CK, Carpenter JH, Mousallem T, Boustany RM, Kuan CT, Burks AW, et al. , Synergistic control of T cell development and tumor suppression by diacylglycerol kinase alpha and zeta. Proc Natl Acad Sci U S A 2008. 105: 11909–11914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Merida I, Torres-Ayuso P, Avila-Flores A, Arranz-Nicolas J, Andrada E, Tello-Lafoz M, Liebana R, et al. , Diacylglycerol kinases in cancer. Adv Biol Regul 2017. 63: 22–31. [DOI] [PubMed] [Google Scholar]
- 10.Jenkins GH, Fisette PL and Anderson RA, Type I phosphatidylinositol 4-phosphate 5-kinase isoforms are specifically stimulated by phosphatidic acid. J Biol Chem 1994. 269: 11547–11554. [PubMed] [Google Scholar]
- 11.Frank C, Keilhack H, Opitz F, Zschornig O and Bohmer FD, Binding of phosphatidic acid to the protein-tyrosine phosphatase SHP-1 as a basis for activity modulation. Biochemistry 1999. 38: 11993–12002. [DOI] [PubMed] [Google Scholar]
- 12.Liu CH, Machado FS, Guo R, Nichols KE, Burks AW, Aliberti JC and Zhong XP, Diacylglycerol kinase zeta regulates microbial recognition and host resistance to Toxoplasma gondii. J Exp Med 2007. 204: 781–792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Avila-Flores A, Santos T, Rincon E and Merida I, Modulation of the mammalian target of rapamycin pathway by diacylglycerol kinase-produced phosphatidic acid. J Biol Chem 2005. 280: 10091–10099. [DOI] [PubMed] [Google Scholar]
- 14.Zhong XP, Hainey EA, Olenchock BA, Zhao H, Topham MK and Koretzky GA, Regulation of T cell receptor-induced activation of the Ras-ERK pathway by diacylglycerol kinase zeta. J Biol Chem 2002. 277: 31089–31098. [DOI] [PubMed] [Google Scholar]
- 15.Olenchock BA, Guo R, Carpenter JH, Jordan M, Topham MK, Koretzky GA and Zhong XP, Disruption of diacylglycerol metabolism impairs the induction of T cell anergy. Nat Immunol 2006. 7: 1174–1181. [DOI] [PubMed] [Google Scholar]
- 16.Zha Y, Marks R, Ho AW, Peterson AC, Janardhan S, Brown I, Praveen K, et al. , T cell anergy is reversed by active Ras and is regulated by diacylglycerol kinase-alpha. Nat Immunol 2006. 7: 1166–1173. [DOI] [PubMed] [Google Scholar]
- 17.Martinez-Moreno M, Garcia-Lievana J, Soutar D, Torres-Ayuso P, Andrada E, Zhong XP, Koretzky GA, et al. , FoxO-dependent regulation of diacylglycerol kinase alpha gene expression. Mol Cell Biol 2012. 32: 4168–4180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sanjuan MA, Pradet-Balade B, Jones DR, Martinez AC, Stone JC, Garcia-Sanz JA and Merida I, T cell activation in vivo targets diacylglycerol kinase alpha to the membrane: a novel mechanism for Ras attenuation. J Immunol 2003. 170: 2877–2883. [DOI] [PubMed] [Google Scholar]
- 19.Zhong XP, Hainey EA, Olenchock BA, Jordan MS, Maltzman JS, Nichols KE, Shen H, et al. , Enhanced T cell responses due to diacylglycerol kinase zeta deficiency. Nat Immunol 2003. 4: 882–890. [DOI] [PubMed] [Google Scholar]
- 20.Baldanzi G, Pighini A, Bettio V, Rainero E, Traini S, Chianale F, Porporato PE, et al. , SAP-mediated inhibition of diacylglycerol kinase alpha regulates TCR-induced diacylglycerol signaling. J Immunol 2011. 187: 5941–5951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gorentla BK, Wan CK and Zhong XP, Negative regulation of mTOR activation by diacylglycerol kinases. Blood 2011. 117: 4022–4031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Shin J, O’Brien TF, Grayson JM and Zhong XP, Differential regulation of primary and memory CD8 T cell immune responses by diacylglycerol kinases. J Immunol 2012. 188: 2111–2117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Jing W, Gershan JA, Holzhauer S, Weber J, Palen K, McOlash L, Pulakanti K, et al. , T Cells Deficient in Diacylglycerol Kinase zeta Are Resistant to PD-1 Inhibition and Help Create Persistent Host Immunity to Leukemia. Cancer Res 2017. 77: 5676–5686. [DOI] [PubMed] [Google Scholar]
- 24.Riese MJ, Wang LC, Moon EK, Joshi RP, Ranganathan A, June CH, Koretzky GA, et al. , Enhanced effector responses in activated CD8+ T cells deficient in diacylglycerol kinases. Cancer Res 2013. 73: 3566–3577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ruffo E, Malacarne V, Larsen SE, Das R, Patrussi L, Wulfing C, Biskup C, et al. , Inhibition of diacylglycerol kinase alpha restores restimulation-induced cell death and reduces immunopathology in XLP-1. Sci Transl Med 2016. 8: 321ra327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Schmidt AM, Lu W, Sindhava VJ, Huang Y, Burkhardt JK, Yang E, Riese MJ, et al. , Regulatory T cells require TCR signaling for their suppressive function. J Immunol 2015. 194: 4362–4370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Arumugam V, Bluemn T, Wesley E, Schmidt AM, Kambayashi T, Malarkannan S and Riese MJ, TCR signaling intensity controls CD8+ T cell responsiveness to TGF-beta. J Leukoc Biol 2015. 98: 703–712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Schmidt AM, Zou T, Joshi RP, Leichner TM, Pimentel MA, Sommers CL and Kambayashi T, Diacylglycerol kinase zeta limits the generation of natural regulatory T cells. Sci Signal 2013. 6: ra101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Joshi RP, Schmidt AM, Das J, Pytel D, Riese MJ, Lester M, Diehl JA, et al. , The zeta isoform of diacylglycerol kinase plays a predominant role in regulatory T cell development and TCR-mediated ras signaling. Sci Signal 2013. 6: ra102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Arranz-Nicolas J, Ogando J, Soutar D, Arcos-Perez R, Meraviglia-Crivelli D, Manes S, Merida I, et al. , Diacylglycerol kinase alpha inactivation is an integral component of the costimulatory pathway that amplifies TCR signals. Cancer Immunol Immunother 2018. 67: 965–980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Avila-Flores A, Arranz-Nicolas J, Andrada E, Soutar D and Merida I, Predominant contribution of DGKzeta over DGKalpha in the control of PKC/PDK-1-regulated functions in T cells. Immunol Cell Biol 2017. 95: 549–563. [DOI] [PubMed] [Google Scholar]
- 32.Almena M, Andrada E, Liebana R and Merida I, Diacylglycerol metabolism attenuates T-cell receptor signaling and alters thymocyte differentiation. Cell Death Dis 2013. 4: e912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Shen S, Wu J, Srivatsan S, Gorentla BK, Shin J, Xu L and Zhong XP, Tight regulation of diacylglycerol-mediated signaling is critical for proper invariant NKT cell development. J Immunol 2011. 187: 2122–2129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Shen S, Chen Y, Gorentla BK, Lu J, Stone JC and Zhong XP, Critical roles of RasGRP1 for invariant NKT cell development. J Immunol 2011. 187: 4467–4473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Pan Y, Deng W, Xie J, Zhang S, Wan ECK, Li L, Tao H, et al. , Graded diacylglycerol kinases alpha and zeta activities ensure mucosal-associated invariant T-cell development in mice. Eur J Immunol 2020. 50: 192–204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Yang J, Zhang P, Krishna S, Wang J, Lin X, Huang H, Xie D, et al. , Unexpected positive control of NFkappaB and miR-155 by DGKalpha and zeta ensures effector and memory CD8+ T cell differentiation. Oncotarget 2016. 7: 33744–33764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wheeler ML, Dong MB, Brink R, Zhong XP and DeFranco AL, Diacylglycerol kinase zeta limits B cell antigen receptor-dependent activation of ERK signaling to inhibit early antibody responses. Sci Signal 2013. 6: ra91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Yang E, Singh BK, Paustian AM and Kambayashi T, Diacylglycerol Kinase zeta Is a Target To Enhance NK Cell Function. J Immunol 2016. 197: 934–941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Evangelisti C, Riccio M, Faenza I, Zini N, Hozumi Y, Goto K, Cocco L, et al. , Subnuclear localization and differentiation-dependent increased expression of DGKzeta in C2C12 mouse myoblasts. J Cell Physiol 2006. 209: 370–378. [DOI] [PubMed] [Google Scholar]
- 40.Santos T, Carrasco S, Jones DR, Merida I and Eguinoa A, Dynamics of diacylglycerol kinase zeta translocation in living T-cells. Study of the structural domain requirements for translocation and activity. J Biol Chem 2002. 277: 30300–30309. [DOI] [PubMed] [Google Scholar]
- 41.Tanaka T, Okada M, Hozumi Y, Tachibana K, Kitanaka C, Hamamoto Y, Martelli AM, et al. , Cytoplasmic localization of DGKzeta exerts a protective effect against p53-mediated cytotoxicity. J Cell Sci 2013. 126: 2785–2797. [DOI] [PubMed] [Google Scholar]
- 42.Topham MK and Epand RM, Mammalian diacylglycerol kinases: molecular interactions and biological functions of selected isoforms. Biochim Biophys Acta 2009. 1790: 416–424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Walker AJ, Draeger A, Houssa B, van Blitterswijk WJ, Ohanian V and Ohanian J, Diacylglycerol kinase theta is translocated and phosphoinositide 3-kinase-dependently activated by noradrenaline but not angiotensin II in intact small arteries. Biochemical Journal 2001. 353: 129–137. [PMC free article] [PubMed] [Google Scholar]
- 44.Topham MK, Bunting M, Zimmerman GA, McIntyre TM, Blackshear PJ and Prescott SM, Protein kinase C regulates the nuclear localization of diacylglycerol kinase-zeta. Nature 1998. 394: 697–700. [DOI] [PubMed] [Google Scholar]
- 45.Raben DM and Tu-Sekine B, Nuclear diacylglycerol kinases: regulation and roles. Front Biosci 2008. 13: 590–597. [DOI] [PubMed] [Google Scholar]
- 46.Bunting M, Tang W, Zimmerman GA, McIntyre TM and Prescott SM, Molecular cloning and characterization of a novel human diacylglycerol kinase zeta. J Biol Chem 1996. 271: 10230–10236. [PubMed] [Google Scholar]
- 47.Evangelisti C, Gaboardi GC, Billi AM, Ognibene A, Goto K, Tazzari PL, McCubrey JA, et al. , Identification of a functional nuclear export sequence in diacylglycerol kinase-zeta. Cell Cycle 2010. 9: 384–388. [DOI] [PubMed] [Google Scholar]
- 48.Ali H, Nakano T, Saino-Saito S, Hozumi Y, Katagiri Y, Kamii H, Sato S, et al. , Selective translocation of diacylglycerol kinase zeta in hippocampal neurons under transient forebrain ischemia. Neurosci Lett 2004. 372: 190–195. [DOI] [PubMed] [Google Scholar]
- 49.Saino-Saito S, Hozumi Y and Goto K, Excitotoxicity by kainate-induced seizure causes diacylglycerol kinase zeta to shuttle from the nucleus to the cytoplasm in hippocampal neurons. Neurosci Lett 2011. 494: 185–189. [DOI] [PubMed] [Google Scholar]
- 50.Gharbi SI, Rincon E, Avila-Flores A, Torres-Ayuso P, Almena M, Cobos MA, Albar JP, et al. , Diacylglycerol kinase zeta controls diacylglycerol metabolism at the immunological synapse. Mol Biol Cell 2011. 22: 4406–4414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Dower NA, Stang SL, Bottorff DA, Ebinu JO, Dickie P, Ostergaard HL and Stone JC, RasGRP is essential for mouse thymocyte differentiation and TCR signaling. Nat Immunol 2000. 1: 317–321. [DOI] [PubMed] [Google Scholar]
- 52.Alberola-Ila J, Hogquist KA, Swan KA, Bevan MJ and Perlmutter RM, Positive and negative selection invoke distinct signaling pathways. J Exp Med 1996. 184: 9–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Hamilton KS, Phong B, Corey C, Cheng J, Gorentla B, Zhong X, Shiva S, et al. , T cell receptor-dependent activation of mTOR signaling in T cells is mediated by Carma1 and MALT1, but not Bcl10. Sci Signal 2014. 7: ra55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Bettini M, Xi H, Milbrandt J and Kersh GJ, Thymocyte development in early growth response gene 1-deficient mice. J Immunol 2002. 169: 1713–1720. [DOI] [PubMed] [Google Scholar]
- 55.Seiler MP, Mathew R, Liszewski MK, Spooner CJ, Barr K, Meng F, Singh H, et al. , Elevated and sustained expression of the transcription factors Egr1 and Egr2 controls NKT lineage differentiation in response to TCR signaling. Nat Immunol 2012. 13: 264–271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hu T, Gimferrer I, Simmons A, Wiest D and Alberola-Ila J, The Ras/MAPK pathway is required for generation of iNKT cells. PLoS One 2011. 6: e19890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Egawa T, Runx and ThPOK: a balancing act to regulate thymocyte lineage commitment. J Cell Biochem 2009. 107: 1037–1045. [DOI] [PubMed] [Google Scholar]
- 58.Sinclair C and Seddon B, Overlapping and asymmetric functions of TCR signaling during thymic selection of CD4 and CD8 lineages. J Immunol 2014. 192: 5151–5159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Woolf E, Xiao C, Fainaru O, Lotem J, Rosen D, Negreanu V, Bernstein Y, et al. , Runx3 and Runx1 are required for CD8 T cell development during thymopoiesis. Proc Natl Acad Sci U S A 2003. 100: 7731–7736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Wu J, Shin J, Xie D, Wang H, Gao J and Zhong XP, Tuberous sclerosis 1 promotes invariant NKT cell anergy and inhibits invariant NKT cell-mediated antitumor immunity. J Immunol 2014. 192: 2643–2650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Wu J, Shen S, Yang J, Xia Z and Zhong XP, Diacylglycerol kinase zeta positively controls the development of iNKT-17 cells. PLoS One 2013. 8: e75202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Shin J, Wang S, Deng W, Wu J, Gao J and Zhong XP, Mechanistic target of rapamycin complex 1 is critical for invariant natural killer T-cell development and effector function. Proc Natl Acad Sci U S A 2014. 111: E776–783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Wu J, Yang J, Yang K, Wang H, Gorentla B, Shin J, Qiu Y, et al. , iNKT cells require TSC1 for terminal maturation and effector lineage fate decisions. J Clin Invest 2014. 124: 1685–1698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Fang X, Wang R, Ma J, Ding Y, Shang W and Sun Z, Ameliorated ConA-induced hepatitis in the absence of PKC-theta. PloS one 2012. 7: e31174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Wei J, Yang K and Chi H, Cutting edge: Discrete functions of mTOR signaling in invariant NKT cell development and NKT17 fate decision. J Immunol 2014. 193: 4297–4301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Zhang L, Tschumi BO, Corgnac S, Ruegg MA, Hall MN, Mach JP, Romero P, et al. , Mammalian target of rapamycin complex 1 orchestrates invariant NKT cell differentiation and effector function. J Immunol 2014. 193: 1759–1765. [DOI] [PubMed] [Google Scholar]
- 67.Prevot N, Pyaram K, Bischoff E, Sen JM, Powell JD and Chang CH, Mammalian target of rapamycin complex 2 regulates invariant NKT cell development and function independent of promyelocytic leukemia zinc-finger. J Immunol 2015. 194: 223–230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Yang W, Gorentla B, Zhong XP and Shin J, mTOR and its tight regulation for iNKT cell development and effector function. Mol Immunol 2015. 68: 536–545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Bendelac A, Savage PB and Teyton L, The biology of NKT cells. Annu Rev Immunol 2007. 25: 297–336. [DOI] [PubMed] [Google Scholar]
- 70.Godfrey DI, Stankovic S and Baxter AG, Raising the NKT cell family. Nat Immunol 2010. 11: 197–206. [DOI] [PubMed] [Google Scholar]
- 71.Okada M, Hozumi Y, Tanaka T, Suzuki Y, Yanagida M, Araki Y, Evangelisti C, et al. , DGKzeta is degraded through the cytoplasmic ubiquitin-proteasome system under excitotoxic conditions, which causes neuronal apoptosis because of aberrant cell cycle reentry. Cell Signal 2012. 24: 1573–1582. [DOI] [PubMed] [Google Scholar]
- 72.Shin J, Xie D and Zhong XP, MicroRNA-34a enhances T cell activation by targeting diacylglycerol kinase zeta. PLoS One 2013. 8: e77983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Jung IY, Kim YY, Yu HS, Lee M, Kim S and Lee J, CRISPR/Cas9-Mediated Knockout of DGK Improves Antitumor Activities of Human T Cells. Cancer Res 2018. 78: 4692–4703. [DOI] [PubMed] [Google Scholar]
- 74.Akiyama H, Hozumi Y, Nakano T, Kubota I and Goto K, Nuclear relocation of DGKzeta in cardiomyocytes under conditions of ischemia/reperfusion. Histol Histopathol 2011. 26: 1383–1390. [DOI] [PubMed] [Google Scholar]
- 75.Hogan A, Shepherd L, Chabot J, Quenneville S, Prescott SM, Topham MK and Gee SH, Interaction of gamma 1-syntrophin with diacylglycerol kinase-zeta. Regulation of nuclear localization by PDZ interactions. J Biol Chem 2001. 276: 26526–26533. [DOI] [PubMed] [Google Scholar]
- 76.Hasegawa H, Nakano T, Hozumi Y, Takagi M, Ogino T, Okada M, Iseki K, et al. , Diacylglycerol kinase zeta is associated with chromatin, but dissociates from condensed chromatin during mitotic phase in NIH3T3 cells. J Cell Biochem 2008. 105: 756–765. [DOI] [PubMed] [Google Scholar]
- 77.You JS, Dooley MS, Kim CR, Kim EJ, Xu W, Goodman CA and Hornberger TA, A DGKzeta-FoxO-ubiquitin proteolytic axis controls fiber size during skeletal muscle remodeling. Sci Signal 2018. 11 eaao6847 DOI: 10.1126/scisignal.aao6847 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Spitaler M, Emslie E, Wood CD and Cantrell D, Diacylglycerol and protein kinase D localization during T lymphocyte activation. Immunity 2006. 24: 535–546. [DOI] [PubMed] [Google Scholar]
- 79.Goto K, Tanaka T, Nakano T, Okada M, Hozumi Y, Topham MK and Martelli AM, DGKzeta under stress conditions: “to be nuclear or cytoplasmic, that is the question”. Adv Biol Regul 2014. 54: 242–253. [DOI] [PubMed] [Google Scholar]
- 80.Evangelisti C, Tazzari PL, Riccio M, Fiume R, Hozumi Y, Fala F, Goto K, et al. , Nuclear diacylglycerol kinase-zeta is a negative regulator of cell cycle progression in C2C12 mouse myoblasts. FASEB J 2007. 21: 3297–3307. [DOI] [PubMed] [Google Scholar]
- 81.Abramovici H, Mojtabaie P, Parks RJ, Zhong XP, Koretzky GA, Topham MK and Gee SH, Diacylglycerol kinase zeta regulates actin cytoskeleton reorganization through dissociation of Rac1 from RhoGDI. Mol Biol Cell 2009. 20: 2049–2059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Okada M, Hozumi Y, Ichimura T, Tanaka T, Hasegawa H, Yamamoto M, Takahashi N, et al. , Interaction of nucleosome assembly proteins abolishes nuclear localization of DGKzeta by attenuating its association with importins. Exp Cell Res 2011. 317: 2853–2863. [DOI] [PubMed] [Google Scholar]
- 83.Lee PP, Fitzpatrick DR, Beard C, Jessup HK, Lehar S, Makar KW, Perez-Melgosa M, et al. , A critical role for Dnmt1 and DNA methylation in T cell development, function, and survival. Immunity 2001. 15: 763–774. [DOI] [PubMed] [Google Scholar]
- 84.Cossarizza A, Chang HD, Radbruch A, Akdis M, Andra I, Annunziato F, Bacher P, et al. , Guidelines for the use of flow cytometry and cell sorting in immunological studies. Eur J Immunol 2017. 47: 1584–1797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Grove LM, Mohan ML, Abraham S, Scheraga RG, Southern BD, Crish JF, Naga Prasad SV, et al. , Translocation of TRPV4-PI3Kgamma complexes to the plasma membrane drives myofibroblast transdifferentiation. Sci Signal 2019. 12 eaau1533 DOI: 10.1126/scisignal.aau1533 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.