

Abstract
Eucalyptus oil has been used since ancient times for its bactericidal, anti-inflammatory, analgesic and sedative effects. In recent years, the action of Eucalyptus oil has been scientifically proven, and there have been reports that Eucalyptus oil suppresses the production of chemokines, cytokines and lipid mediators in basophils, alveolar macrophages and monocytes. Based on this information, we aimed to verify whether Eucalyptus oil can be used for allergic dermatitis, the incidence of which has been increasing among human skin diseases. This effect was verified using a mouse IgE-mediated local allergic model. In conclusion, topical application of Eucalyptus oil suppressed oedema and vascular permeability enhancement due to IgE-mediated allergic on the skin. In addition, we also verified the degranuration of mast cells, which is a part of its action, and examined whether 1,8-cineole, which is the main component of Eucalyptus oil, suppresses the phosphorylation of PLCγ and p38 directly or indirectly. 1,8-cineole was found to suppress degranulation of mast cells.
Subject terms: Molecular biology, Health care, Cell biology, Cell signalling, Immunology, Cytokines, Chemokines, Interleukins, Medical research, Drug development, Diseases, Skin diseases
Introduction
Eucalyptus oil has been used by Australian natives as a remedy for wounds and inflammation1,2. Essential oils obtained from Eucalyptus leaves are said to have anti-inflammatory, analgesic, sedative and bactericidal effects, and they are used in medicines and aromatherapy3. In the UK, oil extracted from Eucalyptus globulus is used to treat airway inflammation and various symptoms of rheumatism. In Japan, the essential oil obtained via steam distillation of the leaves of Eucalyptus globulus Labillardière or other closely related plants (Myrtaceae) is used as an anti-inflammatory and analgesic. Clinical studies confirmed the anti-inflammatory effects of Eucalyptus oil in patients with asthma, such as alleviating sinusitis symptoms and preventing the worsening of chronic obstructive pulmonary disease4–7. A comprehensive report on plant oil using basophils has reported the inhibitory effect of Eucalyptus oil on IgE-mediated basophil activation8. It is suggested that Eucalyptus oil suppresses inflammation by directly acting on immune cells such as macrophages and monocytes9, as supported by several studies. Specifically, Eucalyptus oil was demonstrated to inhibit lipopolysaccharide (LPS)-induced nitric oxide production in mouse macrophage cell lines10 improve the acute inflammatory response of the lungs in a mouse acute lung injury model11 and inhibit active oxygen release by cultured alveolar macrophages from patients with chronic obstructive pulmonary disease12. Additionally, 1,8-cineole, the main component of Eucalyptus oil, blocked arachidonic acid metabolism in blood monocytes from patients with asthma13 and inhibited LPS-induced IL-1β production by human monocytes14. In recent years, it has also been reported that eucalyptol exerts anti-inflammatory effects in mice through effects on TRPM8 channels15.
Eucalyptus oil was shown to have an analgesic effect in the formalin and carrageenan paw oedema test despite being applied topically16,17. As a bath agent, it was reported to improve the skin symptoms of patients with atopic dermatitis18.
Therefore, based on this information, we aimed to verify whether or not Eucalyptus oil can be used for allergic dermatitis, the incidence of which has been increasing among human skin diseases.
In this study, we first evaluated whether Eucalyptus oil suppressed inflammation in an IgE-mediated local allergic model, a model of inflammatory allergic disease caused by mast cell activation19–21. We confirmed that the anti-inflammatory effect of Eucalyptus oil in wild-type mice was attributable to the suppression of mast cell activation based on the findings in mast cell deficient mice. Because Eucalyptus oil suppressed mast cell degranulation, the mechanism of action was also examined using an in vitro cell assay system and bone-marrow-derived mast cells (BMMCs), which revealed that Eucalyptus oil and its main component 1,8-cineole suppressed mast cell degranulation and alleviated allergic dermatitis, supporting their potential as crude medicines for relieving allergic dermatitis.
Results
Eucalyptus oil suppresses IgE-mediated local allergic inflammation in mice
To determine whether Eucalyptus oil can suppress mast cell degranulation, we tested its effects using passive cutaneous anaphylaxis (PCA) responses in mice. PCA is a local skin allergic reaction resulting from hyperpermeability and plasma extravasation following allergen exposure, and it has been used as an animal model for IgE-mediated allergic reactions22,23. We evaluated whether Eucalyptus oil can suppress IgE-mediated allergic reactions by vascular hyperpermeability and local ear oedema by assessing the leakage of Evans blue dye (Fig. 1).
Figure 1.


Eucalyptus oil (Eu) suppresses passive cutaneous anaphylaxis (PCA) in a mouse model. (a) Schedule of treatment and experimental procedures for the PCA and ear swelling response tests. (b) Anti-DNP-IgE was injected intravenously 24 h before DNFB application. One hour before DNFB application, Eucalyptus oil or vehicle (3:1 acetone/olive oil) was applied to the inside of each ear of all animals. DNFB was applied to the outside of the ear, followed by an immediate injection of Evans blue dye. Thirty minutes later, the ears were excised, and the absorbance of the dye was measured. Data are presented as the mean ± SEM (N = 8/group). The statistical significance of the differences was assessed using Dunnett’s test (**P < 0.01 versus vehicle-DNFB). (c) The time course of ear oedema was investigated. Ear thickness was measured using a thickness gauge. Data are presented as the mean ± SEM (N = 8/group). (d) Haematoxylin and eosin staining of ear tissue 6 h after DNFB application. (e,f) Toluidine blue staining of ear tissue 1 or 6 h after DNFB application. (e) Tissue sections were mast cells obtained from ear tissue collected 6 h after the application of DNFB. Mast cells were identified in tissue sections by their characteristic granular, deep blue-purple metachromatic appearance against blue orthochromatic background tissue. ND no degranulation, D degranulation. (f) Left panel shows the number of cutaneous mast cells at the dermal/epidermal (D/E) junction. Right panel shows the percentage of degranulated mast cells. Each bar is mean ± SEM (N = 6/group) of 12 fields of views. Statistical significance of the differences was assessed using the Tukey–Kramer test (**P < 0.01 versus vehicle, ##P < 0.01 versus vehicle-DNFB).
In addition, prior to the Eucalyptus oil test, it was verified whether the inhibitory effect can be detected, using sodium cromoglycate, which has an inhibitory effect on mast cell degranulation (Supplementary Fig. 2).
Eucalyptus oil significantly suppressed the amount of dye leaking into the tissue compared with the effects of the vehicle-DNFB. Eucalyptus oil also inhibited dye leakage in a dose-dependent manner (Fig. 1b). Meanwhile, oedema was induced in the ears of mice following DNFB application, peaking at 6 h, and Eucalyptus oil inhibited this oedema in a dose-dependent manner (Fig. 1c).
Haematoxylin and eosin (H&E) staining of the auricle was performed 6 h after DNFB application. As a result, swelling of the dermal tissue, thickening of the stratum corneum and infiltration of inflammatory cells into the dermal layer via DNFB application were noticed, and these findings were suppressed by Eucalyptus oil (Fig. 1d).
Toluidine blue staining was performed to confirm that Eucalyptus oil suppressed mast cell degranulation after DNFB application. The number of degranulated mast cells was quantified by counting the mast cells in the image of skin tissue24,25.
Almost no degranulation of mast cells was observed in the vehicle. Vehicle-DNFB degranulated 50% of mast cells after 1 h and 68% after 6 h. Furthermore, after 6 h, the number of mast cells confirmed by toluidine blue staining reduced.
In the group to which the Eucalyptus oil was applied, a significant suppression of mast cell degranulation rate was observed 1 h after the application of DNFB as compared with that observed after the application of vehicle-DNFB. A significant inhibitory effect was observed even 6 h after DNFB application; however, the inhibitory effect observed 1 h after DNFB application was weaker than that observed 6 h after DNFB application (Fig. 1e,f).
These results revealed that Eucalyptus oil suppressed PCA induced by anti-DNP-IgE antibody exposure in mice. It is known that mast cell degranulation is strongly involved in the very early phase of ear oedema 1 h after reaction caused by intravenous administration of anti-DNP IgE antibody and subsequent application of DNFB, and that migration of inflammatory cells is involved in the early phase of ear oedema26,27. The ear oedema suppression effect of Eucalyptus oil has been observed from a relatively early stage and this effect gradually weakens after 3 h. As a result of the inhibition of mast cell degranulation by toluidine blue staining, it was clarified that Eucalyptus oil plays a role in mast cell degranulation inhibition.
However, the inflammation caused by anti-DNP-IgE antibodies also involves inflammatory cells other than mast cells and therefore we aimed to clarify how Eucalyptus oil affects mast cell deficient mice27,28.
The inhibitory effect of Eucalyptus oil was not observed in mice lacking mast cells
W/Wv mice are known to lack mast cells because of the lack of mast cell progenitor cells, and they are widely used to confirm the physiological roles of mast cells28–30. We measured the ear thickness by PCA reaction and analysed the inhibitory effect of Eucalyptus oil on the increase of vascular permeability. In wild-type mice, Eucalyptus oil significantly suppressed dye leakage into the tissue compared with the effects of the vehicle. In W/Wv mice deficient in mast cells, both vehicle and Eucalyptus oil had significantly less tissue dye leakage than wild-type vehicle application. Also, there was no difference in the dye leakage by vehicle and Eucalyptus oil (Fig. 2a).
Figure 2.

Inhibitory effect of Eucalyptus oil (Eu) on passive cutaneous anaphylaxis (PCA) in mast celldeficient mice. (a) Effect of Eucalyptus oil on Evans blue dye leakage in wild-type and W/Wv mice. One hour before DNFB application, Eucalyptus oil or vehicle (3:1 acetone/olive oil) was applied to the inside of the ears of wild-type and W/Wv mice sensitised with anti-DNP-IgE. DNFB was applied to the outside of the ear, which was immediately injected intravenously with Evans blue dye. Thirty minutes later, the ears were excised, and the absorbance of the dye was measured. Data are presented as the mean ± SEM (N = 8). The statistical significance of the differences was assessed using Tukey–Kramer test (**P < 0.01, *P < 0.05 versus vehicle). (b) Time-dependent changes in the suppression of pinna oedema by Eucalyptus oil in wild-type and W/Wv mice. Ear thickness was measured using a thickness gauge. Data are presented as the mean ± SEM (N = 8). (c) Haematoxylin and eosin-stained and toluidine blue-stained images of ear tissue from Eucalyptus-oil-treated wild-type and W/Wv mice 6 h after DNFB application.
DNFB application induced oedema in the ears of wild-type mice, peaking at 6 h in the vehicle (Fig. 2b), and these effects were suppressed by Eucalyptus oil. The vehicle- and Eu-treated W/Wv mice showed increased ear thickness with the application of DNFB, peaking at 3 h, but the increase was obviously smaller in the vehicle-treated wild-type mice (Fig. 2b).
H&E and toluidine blue staining of the auricle 6 h after DNFB application revealed swelling of dermal tissue, thickening of the stratum corneum and infiltration of inflammatory cells into the dermal layer in vehicle-treated wild-type mice, and these effects were suppressed by Eucalyptus oil treatment (Fig. 2c, Supplementary Fig. 3). Vehicle-treated W/Wv mice exhibited less swelling of dermal tissue, thickening of the stratum corneum and infiltration of inflammatory cells into the dermis layer compared with the findings in vehicle-treated wild-type mice, but there was no difference in the findings between vehicle- and Eucalyptus-oil-treated W/Wv mice (Fig. 2c).
Toluidine blue staining of the ear 6 h after DNFB application confirmed mast cell degranulation in vehicle-treated wild-type mice. Fewer mast cells were degranulated in Eucalyptus-oil-treated wild-type mice. Meanwhile, no mast cells were identified in either vehicle- or Eucalyptus-oil-treated W/Wv mice (Fig. 2c).
Predictably, treatment with vehicle and Eucalyptus oil in mast cell deficient W/Wv mice did not differ in terms of the inhibitory effect on PCA responses. However, W/Wv and wild-type mice are known to have no differences in T lymphocytes, B lymphocytes and NK cells31, but have low anaemia and red blood cell count due to lack of skin pigment cells27. Therefore, detailed thorough examination is needed in the future.
Mast cells play an important role in IgE-mediated hypersensitivity reactions, making them an important target for alleviating allergic symptoms32. When mast cells are exposed to antigens, IgE-binding high-affinity IgE receptors (FcεRI) on the plasma membrane are cross-linked, triggering mast cell activation and degranulation. Therefore, an in vitro cell-based assay using BMMCs was performed to investigate the mechanism by which Eucalyptus oil suppresses mast cell degranulation.
Eucalyptus oil and 1,8-cineole inhibit the degranulation of BMMCs
The effects of Eucalyptus oil and its main component 1,8-cineole on BMMC degranulation were investigated. Anti-DNP-IgE antibody-sensitised BMMCs were treated with Eucalyptus oil or 1,8-cineole and stimulated with DNP-human serum albumin (HSA) as an antigen. The amount of β-hexosaminidase released from the cells was used as a marker of enzymatic activity to evaluate the effects of Eucalyptus oil and 1,8-cineole on mast cell degranulation. β-Hexosaminidase is released together with histamine during mast cell degranulation, and the amount released is generally used as an indicator of degranulation33,34.
First, the viability of BMMCs following exposure to various concentrations of Eucalyptus oil or 1,8-cineole was examined. Both Eucalyptus oil and 1,8-cineole had no effect on cell viability at concentrations below 1 µg/mL (Fig. 3a). When Eucalyptus oil or 1,8-cineole was applied to BMMCs at a concentration of 0.1, 0.5 or 1 µg/mL, β-hexosaminidase release was significantly suppressed. These results indicated that Eucalyptus oil and 1,8-cineole suppressed mast cell degranulation (Fig. 3b).
Figure 3.

Effect of Eucalyptus oil (Eu) and 1,8-cineole on IgE-induced degranulation in bone-marrow-derived mast cells (BMMCs). (a) Effect of Eucalyptus oil and 1,8-cineole on BMMCs viability. Cells were treated with various concentrations of Eucalyptus oil or 1,8-cineole for 24 h. Next, cell viability was measured using the MTT assay. Relative cell viability was calculated by comparing the absorbance in cells treated with Eucalyptus oil or 1,8-cineole with that in cells treated with vehicle (0.1% DMSO). Data are expressed as the mean ± SEM (N = 3). (b) Eucalyptus oil or 1,8-cineole was applied to anti-DNP-IgE-sensitised BMMCs immediately after stimulation with DNP-human serum albumin (HSA), and after 1 h, β-hexosaminidase release was determined by comparing the enzymatic activity in BMMCs between treatment with vehicle (0.1% DMSO) and DNP-HSA. Data are expressed as the mean ± SEM (N = 4). Dunnett’s test was used to assess the statistical significance (**P < 0.01versus vehicle-DNP-HSA).
Eucalyptus oil and 1,8-cineole suppress inflammatory chemokine, cytokine and lipid mediator production by BMMCs
Mast cells activated locally by inflammation produce histamine as well as cytokines such as IL-4 and IL-13 and lipid mediators such as prostaglandin and leukotriene C4, thereby promoting allergic inflammation32,35. The effects of Eucalyptus oil and 1,8-cineole on the production of histamine, IL-4, IL-13, prostaglandin and leukotriene C4 were investigated via an in vitro assay using BMMCs. When BMMCs sensitised with anti-DNP-IgE were stimulated with the antigen, histamine, IL-4, IL-13, prostaglandin and leukotriene C4 were detected in the supernatant (Fig. 4a–e). When Eucalyptus oil or 1,8-cineole was applied at 0.1, 0.5 or 1 µg/mL, histamine production was significantly inhibited (Fig. 4a), and IL-4 and IL-13 production was also significantly suppressed (Fig. 4b,c). Neither Eucalyptus oil nor 1,8-cineole inhibited the production of prostaglandin and leukotriene C4 at 0.1 g/mL. At a concentration 0.5 µg/mL, their production was suppressed, albeit without significance. At 1 µg/mL, significant suppression was observed (Fig. 4d,e). There was no difference between the inhibitory effects of Eucalyptus oil and 1,8-cineole. Regarding mast cell degranulation, the effects of Eucalyptus oil could be attributable to 1,8-cineole. The inhibitory effects on prostaglandin D2 and leukotriene C4 were weaker than those on histamine, IL-4 and IL-13.
Figure 4.

Effects of Eucalyptus oil (Eu) and 1,8-cineole on inflammatory chemokine and cytokine production by bone-marrow-derived mast cells (BMMCs). (a) Histamine release was determined after treating anti-DNP-IgE-sensitised BMMCs with Eucalyptus oil and 1,8-cineole. Histamine release in the supernatant was specifically after 30 min of stimulation with DNP-human serum albumin (HSA) via ELISA. Histamine release was calculated relative to that in cells treated with vehicle (0.1% DMSO) and stimulated with DNP-HSA. Data are expressed as the mean ± SEM (N = 4). Dunnett’s test was used to assess the statistical significance (**P < 0.01 versus vehicle-DNP-HSA). (b,c) Production of IL-4 and IL-13 by anti-DNP-IgE-sensitised BMMCs was determined after treatment with Eucalyptus oil or 1,8-cineole. ELISA was performed after culturing cells with DNP-HSA for 3 h. IL-4 and IL-13 production was calculated relative to that in cells treated with vehicle (0.1% DMSO) and stimulated with DNP-HSA. Data are expressed as the mean ± SEM (N = 4). Dunnett’s test was used to assess the statistical significance (**P < 0.01 versus vehicle-DNP-HSA). (d,e) Production of prostaglandin D2 and leukotriene C4 by anti-DNP-IgE-sensitised BMMCs was determined after treatment with Eucalyptus oil or 1,8-cineole. ELISA was performed after culturing cells with DNP-HSA for 1 h. Prostaglandin D2 and leukotriene C4 production was calculated relative to that in cells treated with vehicle (0.1% DMSO) and stimulated with DNP-HSA. Data are expressed as the mean ± SEM (N = 4). Dunnett’s test was used to assess the statistical significance (**P < 0.01) of differences between vehicle- and DNP-HSA-treated cells. Dunnett’s test was used to assess the statistical significance (*P < 0.05, **P < 0.01 versus vehicle-DNP-HSA).
Eucalyptus oil and 1,8-cineole suppress the increase in intracellular Ca2+ concentration in BMMCs
When IgE is cross-linked by the antigen, FcεRI aggregation initiates signalling cascades, leading to the increase in intracellular Ca2+ concentration is one of the major signalling pathways for mast cell degranulation36. We next investigated the effect of 1,8-cineole on the intracellular Ca2+ increase in BMMCs.
Stimulation with DNP-HSA was performed for 60 s after the start of Ca2+ measurement. As depicted in Fig. 5a, the fluorescence intensity of Fluo 4 increased immediately after the DNP-HSA stimulation of vehicle-treated cells, whereas the fluorescence intensity of Fluo 4 remained unchanged without DNP-HSA. Fluo 4 fluorescence intensity was significantly suppressed when cells treated with 1,8-cineole were stimulated with DNP-HSA.
Figure 5.

1,8-cineole suppresses the elevation of intracellular Ca2+ concentration. BMMCs sensitised with anti-DNP IgE were incubated in medium containing Fluo 4-AM. BMMCs were treated with various concentrations of 1,8-cineole and stimulated with DNP-HSA 60 s after initiating the monitoring of fluorescence intensity. (a) Time course of intracellular Ca2 + concentration in BMMCs. Data are displayed as mean ± SEM (N = 5). (b) Relative fluorescence intensity 60 s after DNP-HSA stimulation. Data are displayed as mean ± SEM (N = 5). Statistical significance was assessed using Dunnett’s test (** P < 0.01 vs vehicle-DNP-HSA).
Figure 5b shows the results 60 s after DNP-HSA stimulation. Compared with the fluorescence intensity of DNP-HSA-stimulated cells treated with vehicle, there was significant suppression at 0.1, 0.5 and 1.0 μg/mL every concentration of 1,8-cineole, and although no significant difference was detected, a concentration-dependent suppression was observed.
It was found that 1,8-cineole suppressed the intracellular Ca2+ increase stimulated by DNP-HSA. Therefore, we next examined the effect of 1,8-cineole on the intracellular signalling cascades of BMMCs other than Ca2+.
1,8-cineole does not affect Lyn and Syk phosphorylation
Mast cells express FcεRI on their surface, and when IgE is cross-linked by an antigen, the Src family tyrosine kinase Lyn, which is related to FcεRI, is phosphorylated, followed by a series of signal transductions. Then, signs of mast cell activation such as degranulation, cytokine production and lipid mediator production are observed29,32,37,38. Therefore, the effects of 1,8-cineole on the phosphorylation status of intracellular signalling proteins were examined via immunoblot analysis.
Anti-DNP-IgE antibody-sensitised BMMCs were treated with 0.5 µg/mL 1,8-cineole, stimulated with DNP-HSA as an antigen, lysed and immunoblotted. As a result, the phosphorylation of Syk and Lyn was not suppressed by 1,8-cineole, even when the concentration was increased to as much as 2.0 µg/mL (Fig. 6a,b).
Figure 6.

Effect of 1,8-cineole on the phosphorylation of Syk and Lyn. (a,b) Eucalyptus oil or 1,8-cineole was applied to BMMCs sensitised with anti-DNP-IgE 24 h before stimulation with DNP-human serum albumin, and then after 10 min, cells were lysed, separated by SDS-PAGE and immunoblotted to analyse p-Syk, Syk, p-Lyn and Lyn expression. The left side presents representative examples of blots for each protein, and the right bar graphs present the phosphorylation relative ratios for Syk and Lyn.Phosphorylation ratios were calculated relative to Syk and Lyn expression (each set as 1), and data are presented as the mean ± SEM of 4–6 independent experiments. Dunnett’s test was used to assess the statistical significance (**P < 0.01 versus DNP-HSA). The original blots are shown in the supplementary Fig. 5.
1,8-cineole suppresses PLCγ and p38 phosphorylation
Lyn and Syk phosphorylation was not inhibited by 1,8-cineole. Therefore, we investigated the intracellular signalling cascade downstream of Lyn and Syk. Anti-DNP-IgE antibody-sensitised BMMCs were treated with Eucalyptus oil or 1,8-cineole, stimulated with DNP-HSA as an antigen, lysed and immunoblotted. As a result, PLCγ phosphorylation was significantly suppressed by 0.5 μg/mL 1,8-cineole (Fig. 7a), as was p38 phosphorylation (Fig. 7b). Phosphorylation of phospholipase A2 (PLA2) was not suppressed by 0.5 µg/mL 1,8-cineole(Fig. 7c). The results confirm that 1,8-cineole suppressed PLCγ and p38 phosphorylation.
Figure 7.

Effect of 1,8-cineole on the phosphorylation of PLCγ, p38 and phospholipase A2 (PLA2). (a–c) 1,8-cineole was applied to bone-marrow-derived mast cells (BMMCs) after sensitisation with anti-DNP-IgE for 24 h, and cells were subsequently stimulated with DNP-human serum albumin (HSA). After 10 min, the cells were lysed, and SDS-PAGE was performed. Proteins were separated, and p-PLCγ, PLCγ, p-p38, p38, p-PLA2 and PLA2 expression was examined via immunoblotting. The left side presents representative blots for each protein, and the right bar graphs show the relative ratios of PLCγ, p38 and PLA2 phosphorylation. Phosphorylation ratios were calculated relative to PLCγ, p38 and PLA2 expression (each set as 1), and data are presented as the mean ± SEM of 4–6 independent experiments. Dunnett’s test was used to assess the statistical significance (**P < 0.01 versus DNP-HSA). The original blots are shown in the supplementary Figs. 6, 7.
Discussion
To confirm the usefulness of Eucalyptus oil before starting this test, we conducted a comparative test with Orange oil,Hinokitiol,L-menthol,dl-camphol, γ-oryzanol, etc., which are plant oils known to have degranulation inhibitory effects on basophils8,39,40, and confirmed the potential of Eucalyptus oil (Supplementary Fig. 1). In this study, we aimed to investigate the suppression of the degranulation effect of Eucalyptus oil by topical application and its mechanism of action.
Using an IgE-sensitised allergy mouse model26,27, this study revealed that Eucalyptus oil may be effective against IgE-mediated allergic dermatitis. The therapeutic effect was demonstrated even when applied externally to the skin.
In a test in which IgE-sensitised allergy was induced in mast cell deficient mice, Eucalyptus oil had a suppressive effect with a result same as that of the vehicle, and it was speculated that Eucalyptus oil is involved in suppressing the degranulation of mast cells.
Moreover, the release of histamine, which is considered to be caused by degranulation, the production of cytokines such as IL-4 and IL-13, and the production of lipid mediators such as prostaglandin D2 and leukotriene C4 were suppressed in the test using BMMCs.
The incidence of allergic diseases caused by mast cell activation such as bronchial asthma, atopic dermatitis and hay fever has increased in recent years32,33. Antihistamines and mast cell stabilisers are commonly used to treat IgE-mediated allergic diseases. Although antihistamines are the most commonly prescribed medications to improve allergic symptoms, they can cause adverse effects such as drowsiness and headaches41.
Eucalyptus oil has been reported to induce oedema and peritoneal mast cell degranulation in rats at high concentrations when administered intra-dermally42, but no side effects such as sleepiness have been reported. In the present IgE-sensitised allergy model, the Eucalyptus oil was applied transdermally and there were no risks of adverse effects; thus, it can be used as a topical treatment separately or in combination with antihistamines.
In a topical study using 10% Eucalyptus oil, no irritation or sensitisation was noted in 25 subjects41. The recommended concentration for external 1,8-cineole application is 20–25% or less according to the Commission E Monograph of Germany and the Canadian Department of Health. Thus, 1,8-cineole is considered a useful treatment for allergic diseases if used topically at a concentration not exceeding 20%.
On the other hand, to clarify the degranulation inhibitory mechanism of Eucalyptus oil in BMMCs, we investigated the increase of intracellular Ca2+ concentration in BMMCs using 1,8-cineole, which accounts for 80% of Eucalyptus oil.
Our results showed that 1,8-cineole suppressed the increase of Ca2+ in BMMCs. It is known that the increase of Ca2+ in BMMCs by the FcεRI aggregation signal is associated with the phosphorylation signal immediately below FcεRI37,38. Therefore, we explored whether 1,8-cineole suppresses the phosphorylation of Lyn and syk43,44, which play important roles in the signalling cascade immediately below FcεRI. However, 1,8-cineole did not suppress the phosphorylation of Lyn and Syk. This is consistent with the result that PLA2 phosphorylation is not suppressed.
Next, we confirmed the phosphorylation of PLCγ, p38, and PLA2, which are the downstream signals of Syk. 1,8-cineole suppressed the phosphorylation of PLCγ and p38. PLCγ is known as an upstream proteins of the degranulation-related molecule DAG, PKC and IP319,20,38. 1,8-cineole directly or indirectly suppressed the phosphorylation of PLCγ. Therefore, it was believed that it suppressed the increase of intracellular Ca2+ and the release of histamine through degranulation (Fig. 8).
Figure 8.
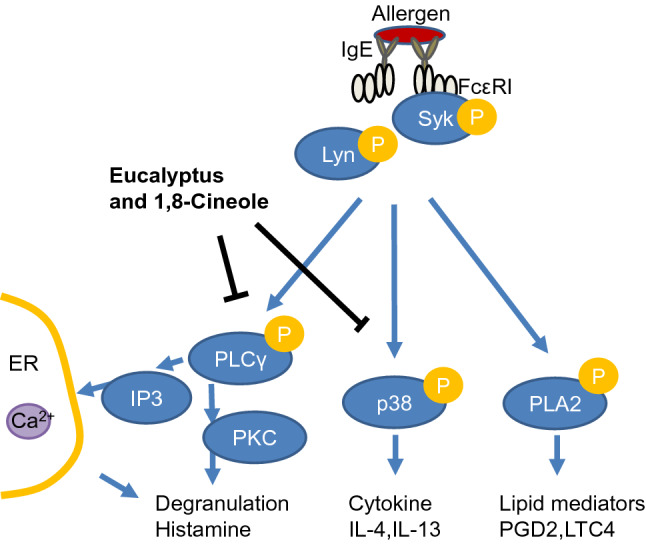
Eucalyptus and 1,8-cineole are putative sites of inducible inhibition of mast cell degranulation.
Studies have reported that p38 is located in the upstream of the production of cytokines and chemokines20,35,38. We considered that the production of cytokines such as IL-4 and IL-13 was suppressed by directly or indirectly suppressing the phosphorylation of p38 (Fig. 8). However, it is known that the production of cytokines and chemokines involves several factors such as ERK, JNK and AKT35,38, and therefore, a more detailed analysis was considered to be necessary.
Lastly, no phosphorylation of PLA2 by 1,8-cineole could be confirmed. This result is inconsistent with the result of PGD2 and LTC4 suppression by ELISA. From the knowledge that PLA2 affects the production of lipid mediators PGD2 and LTC434, we assume that 1,8-cineole inhibits PGD2 and LTC4 production by pathways other than inhibition of PLA2 phosphorylation. However, the details could not be revealed in our survey.
In conclusion, we demonstrated that application of Eucalyptus oil to mice with IgE-mediated allergic dermatitis suppresses vascular hyperpermeability and oedema of the skin. Moreover, focusing on one of these action points, the degranulation of mast cells, 1,8-cineole acts directly or indirectly on the phosphorylation of PLCγ and p-38 to suppress degranulation.
Materials and methods
Mice
Female BALB/c mice (Japan SLC, Shizuoka, Japan) and male WBB6F1 + / + (wild-type) and WBB6Ft/J-W/Wv (W/Wv) mice (Japan SLC, Shizuoka, Japan) were used at 6–12 weeks old. The mice were housed in a room under controlled temperature (22 °C ± 2 °C), humidity (50% ± 10%) and light (lights on from 07:00 to 19:00). Food and water were freely available. The study was approved by the Animal Research Committee of Ikedamohando Research Center in accordance with the National Research Council Guidelines.
Reagents
DNCB, anti-DNP-IgE and DNP-HSA were purchased from Sigma-Aldrich (St. Louis, MO, USA), Eucalyptus oil, procured from Nippon Terpene Chemicals (Tokyo, Japan), conforms with the Japanese Pharmacopoeia, and its main component, 1,8-cineole, accounts for 82.5% of the oil. 1,8-cineole was obtained from Nacalai Tesque (Kyoto, Japan), and dimethyl sulfoxide (DMSO) was acquired from Wako (Osaka, Japan). The following antibodies for immunoblotting were purchased from Cell Signalling Technology (Beverly, MA, USA): anti-Syk, anti-phospho-Syk, anti-Lyn, anti-phospho-Lyn, anti-PLA2, anti-phospho-PLA2, anti-PLCγ, anti-phospho-PLCγ, anti-p38 and anti-phospho-p38. Goat anti-rabbit IgG horseradish peroxidase was purchased from Cell Signaling Technology (Danvers, MA, USA).
Passive cutaneous anaphylaxis (PCA)
The mice were sensitised via the intravenous administration of 20 μg of anti-DNP-IgE. In the increased vascular permeability test, after 24 h, 0.6% DNFB was applied to the outside ear of each mouse, and 1% Evans blue (Sigma-Aldrich, St. Louis, MO, USA) was injected into the tail vein. One hour before DNFB application, 0.5%, 2% or 8% Eucalyptus oil was applied to the inside of the ear. After 30 min, the mice were sacrificed, and their ears were collected. Skin was excised and incubated with KOH at 55 °C for 24 h, and then, the extravasated Evans blue dye was extracted. The absorbance of the dye at 620 nm was measured using a multi-well spectrophotometer (Bio-Rad, Hercules, CA, USA).
In the ear swelling response test, no dye was administered, and Eucalyptus oil was applied to the inside of the ear. After 1 h, DNFB was applied to the outside of the ear, and the thickness of the ear was measured with a digital thickness gauge (Mitutoyo, Kanagawa, Japan).
Preparation and culture of BMMCs
Bone marrow was collected from the femurs and tibias of wild-type BALB/c mice (female, 6–10 weeks old; Japan SLC, Shizuoka, Japan) after sacrifice via cervical dislocation. Their bone marrow cells were cultured in RPMI 1640 medium supplemented with 10% heat-inactivated FCS, 100 U/mL penicillin, 0.1 mg/mL streptomycin, 50 µM 2-ME and 5 ng/mL IL-3 (PeproTech, Cranbury, NJ, USA). The medium was changed every 3 days during culture. Mature BMMCs were obtained after 4 weeks of culture and stained with toluidine blue. Most (> 90%) of these cells expressed both c-Kit and FcεRI. All experiments were performed with BMMCs cultured for 4–7 weeks.
β-Hexosaminidase release assay
BMMCs (1 × 105 cells/mL) were incubated for 24 h with 1 μg/mL anti-DNP-IgE. Surface-bound IgE was cross-linked for 30 min at 37 °C using DNP-HSA. Reactions were stopped by placing the cells on ice. β-Hexosaminidase in the supernatant and cell lysate was incubated with 26 mM citrate buffer (pH 4.5) and 2.3 mg/mL p-nitrophenyl-N-acetyl-α-d-glucosaminide for 60 min at 37 °C. The reaction was developed by adding 0.4 M glycine (pH 10.7), and the absorbance was measured at 570 nm using a multi-plate reader (Bio-Rad). The percentage of β-hexosaminidase release was calculated relative to the total β-hexosaminidase content22,40.
Histology
Ear specimens were fixed with 10% neutral buffered formalin and embedded in paraffin. Four-micrometre-thick sections cut from each paraffin block were stained routinely with H&E and toluidine blue. Extrusion of toluidine blue-stained granules was the evidence indicating mast cell degranulation. The number of mast cells and the percentage of degranulated mast cells at the dermal/epidermal junction were analysed in tissue sections, as described previously24,25.
Cell viability assay
BMMCs (1 × 104 cells/mL) were grown in 96-well micro-titre plates for 24 h. Eucalyptus oil or 1,8-cineole were added to the cells at a concentration of 0.5, 1.0, 5.0, 10.0 or 50.0 μg/mL, followed by incubation for 24 h. The cytotoxic effects of Eucalyptus oil and 1,8-cineole were evaluated using the conventional 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Specifically, 20 μL of MTT (5 mg/mL) was added to each well, followed by incubation at 37 °C for 4 h. The medium was removed, and 200 μL of DMSO was added to each well. The optical density was measured at 490 nm using a multi-well spectrophotometer (Bio-Rad).
Measurement of intracellular calcium levels
Intracellular calcium levels were measured using a Calcium kit Fluo 4 (Dojindo Laboratories, Kumamoto, Japan) according to the manufacturer’s instructions45,46. BMMCs were seeded at 4.0 × 104 cells/well in a 96-well black culture plate and sensitised with anti-DNP IgE (50 ng/mL) at 37 °C overnight. After washing with PBS, the cells were incubated with Fluo 4-AM for 1 h at 37 °C. Cells were again washed with PBS, treated with the recording buffer containing Eucalyptus oil and 1,8-cineole, and then incubated at 37 °C for 30 min. Next, the cells were stimulated with DNP-HSA (50 ng/mL), and the fluorescence intensity was immediately monitored with an excitation wavelength of 485 nm and an emission wavelength of 535 nm using a microplate reader (BioTeh, Winooski, St, USA).
ELISA for histamine, IL-4, IL-13, prostaglandin D2 and leukotriene C4 production
BMMCs were stimulated as described previously. The cell culture supernatants were harvested 3 h after stimulation, and IL-6 and IL-13 concentrations were measured using ELISA kits (R&D Systems, Minneapolis, MN, USA). The prostaglandin D2 and leukotriene C4 concentrations in cell culture supernatants were measured at 1 h after stimulation using ELISA kits (Cayman Chemical, Ann Arbor, MI, USA) according to the manufacturer’s instructions. Histamine concentrations in cell culture supernatants were measured at 30 min after stimulation using an ELISA kit (Cayman Chemical) according to the manufacturer’s instructions.
Immunoblot analysis
BMMCs were washed and lysed with iced lysis buffer (1% Triton X-100, 50 mM Tris–HCl [pH 7.5], 150 mM NaCl, protease inhibitor cocktail and phosphatase inhibitor cocktail; Nacalai Tesque)47. The lysates were subjected to SDS-PAGE and immunoblotting. The reactive bands were visualised using a 5-bromo-4-chloro-3-indolyl phosphate/NBT colour development substrate (Promega, Madison, WI, USA) or an ECL chemiluminescence detection kit. Densitometric analysis of the data obtained from at least three independent experiments was performed using a cooled CCD camera system EZ-Capture II and CS analyser ver. 3.00 software (ATTO, Tokyo, Japan).
Statistical analysis
All data are expressed as the mean ± SEM. Each experiment was performed independently at least three times. For statistical significance, two-tailed Student’s t-test was performed when the results were between two groups. Comparisons between other groups were performed by one-way analysis of variance using Tukey–Kramer or Dunnett’s multiple comparison test.
Ethical approval
All animal testing protocols were approved by the Animal Experiment Committee of Ikeda Mohando Co., Ltd. The Animal Experiment Committee of Ikeda Mohando Co., Ltd., has been certified by the Japan Health Science Foundation, including on-site surveys.
Supplementary information
Acknowledgements
The authors declare that they have no conflicts of interest related to this study. The authors would like to thank Enago (www.enago.jp) for the English language review.
Author contributions
T.N. and Y.K. designed research; A.H. H.K. and H.T. performed experiments; T.N. and A.H. analysed data T.N., Y.Y. and N.Y. wrote the manuscript.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
is available for this paper at 10.1038/s41598-020-77039-5.
References
- 1.Hart PH, et al. Terpinen-4-ol, the main component of the essential oil of Melaleuca alternifolia (tea tree oil), suppresses inflammatory mediator production by activated human monocytes. Inflamm. Res. 2000;49:619–626. doi: 10.1007/s000110050639. [DOI] [PubMed] [Google Scholar]
- 2.Byard R. Traditional medicine of aboriginal Australia. CMAJ. 1988;139:792–794. [PMC free article] [PubMed] [Google Scholar]
- 3.Akhtar MA, Raju R, Beattie KD, Bodkin F, Munch G. Medicinal plants of the australian aboriginal Dharawal people exhibiting anti-inflammatory activity. Evid. Based Complement. Alternat. Med. 2016;2016:2935403. doi: 10.1155/2016/2935403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Worth H, Schacher C, Dethlefsen U. Concomitant therapy with Cineole (Eucalyptole) reduces exacerbations in COPD: A placebo-controlled double-blind trial. Respir. Res. 2009;10:69. doi: 10.1186/1465-9921-10-69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kehrl W, Sonnemann U, Dethlefsen U. Therapy for acute nonpurulent rhinosinusitis with cineole: Results of a double-blind, randomized, placebo-controlled trial. Laryngoscope. 2004;114:738–742. doi: 10.1097/00005537-200404000-00027. [DOI] [PubMed] [Google Scholar]
- 6.Juergens UR, et al. Anti-inflammatory activity of 18-cineol (eucalyptol) in bronchial asthma: A double-blind placebo-controlled trial. Respir Med. 2003;97:250–256. doi: 10.1053/rmed.2003.1432. [DOI] [PubMed] [Google Scholar]
- 7.Meister R, Wittig T, Beuscher N, de Mey C. Efficacy and tolerability of myrtol standardized in long-term treatment of chronic bronchitis. A double-blind, placebo-controlled study. Study Group Investigators. Arzneimittelforschung. 1999;49:351–358. doi: 10.1055/s-0031-1300426. [DOI] [PubMed] [Google Scholar]
- 8.Ikawati Z, Wahyuono S, Maeyama K. Screening of several Indonesian medicinal plants for their inhibitory effect on histamine release from RBL-2H3 cells. J. Ethnopharmacol. 2001;75:249–256. doi: 10.1016/s0378-8741(01)00201-x. [DOI] [PubMed] [Google Scholar]
- 9.Juergens UR. Anti-inflammatory properties of the monoterpene 1.8-cineole: Current evidence for co-medication in inflammatory airway diseases. Drug Res. (Stuttg) 2014;64:638–646. doi: 10.1055/s-0034-1372609. [DOI] [PubMed] [Google Scholar]
- 10.Zhao C, Sun J, Fang C, Tang F. 1,8-cineol attenuates LPS-induced acute pulmonary inflammation in mice. Inflammation. 2014;37:566–572. doi: 10.1007/s10753-013-9770-4. [DOI] [PubMed] [Google Scholar]
- 11.Kim KY, Lee HS, Seol GH. Eucalyptol suppresses matrix metalloproteinase-9 expression through an extracellular signal-regulated kinase-dependent nuclear factor-kappa B pathway to exert anti-inflammatory effects in an acute lung inflammation model. J. Pharm. Pharmacol. 2015;67:1066–1074. doi: 10.1111/jphp.12407. [DOI] [PubMed] [Google Scholar]
- 12.Rantzsch U, Vacca G, Duck R, Gillissen A. Anti-inflammatory effects of Myrtol standardized and other essential oils on alveolar macrophages from patients with chronic obstructive pulmonary disease. Eur. J. Med. Res. 2009;14(Suppl 4):205–209. doi: 10.1186/2047-783x-14-s4-205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Juergens UR, Stober M, Schmidt-Schilling L, Kleuver T, Vetter H. Antiinflammatory effects of euclyptol (1.8-cineole) in bronchial asthma: Inhibition of arachidonic acid metabolism in human blood monocytes ex vivo. Eur. J. Med. Res. 1998;3:407–412. [PubMed] [Google Scholar]
- 14.Juergens UR, Stober M, Vetter H. Inhibition of cytokine production and arachidonic acid metabolism by eucalyptol (1.8-cineole) in human blood monocytes in vitro. Eur. J. Med. Res. 1998;3:508–510. [PubMed] [Google Scholar]
- 15.Caceres AI, et al. Transient receptor potential cation channel subfamily M member 8 channels mediate the anti-inflammatory effects of eucalyptol. Br. J. Pharmacol. 2017;174:867–879. doi: 10.1111/bph.13760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Gbenou JD, et al. Phytochemical composition of Cymbopogon citratus and Eucalyptus citriodora essential oils and their anti-inflammatory and analgesic properties on Wistar rats. Mol. Biol. Rep. 2013;40:1127–1134. doi: 10.1007/s11033-012-2155-1. [DOI] [PubMed] [Google Scholar]
- 17.Santos FA, Rao VS. Antiinflammatory and antinociceptive effects of 1,8-cineole a terpenoid oxide present in many plant essential oils. Phytother. Res. 2000;14:240–244. doi: 10.1002/1099-1573(200006)14:4<240::aid-ptr573>3.0.co;2-x. [DOI] [PubMed] [Google Scholar]
- 18.Tukamoto K, et al. The examination of the usefulness for atopic dermatitis patient of Eucalyptus extract mixture bathing medicine. Nishinihon J. Dermatol. 1999;61:515–519. doi: 10.2336/nishinihonhifu.61.515. [DOI] [Google Scholar]
- 19.Marino A, et al. The novel H2 S donor 4-carboxy-phenyl isothiocyanate inhibits mast cell degranulation and renin release by decreasing intracellular calcium. Br. J. Pharmacol. 2016;173:3222–3234. doi: 10.1111/bph.13583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ogawa K, et al. DOCK5 functions as a key signaling adaptor that links FcepsilonRI signals to microtubule dynamics during mast cell degranulation. J. Exp. Med. 2014;211:1407–1419. doi: 10.1084/jem.20131926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kawakami T, Galli SJ. Regulation of mast-cell and basophil function and survival by IgE. Nat. Rev. Immunol. 2002;2:773–786. doi: 10.1038/nri914. [DOI] [PubMed] [Google Scholar]
- 22.Nishi K, et al. Docosahexaenoyl ethanolamide mitigates IgE-mediated allergic reactions by inhibiting mast cell degranulation and regulating allergy-related immune cells. Sci. Rep. 2019;9:16213. doi: 10.1038/s41598-019-52317-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Finkelman FD. Anaphylaxis: Lessons from mouse models. J. Allergy Clin. Immunol. 2007;120:506–515. doi: 10.1016/j.jaci.2007.07.033. [DOI] [PubMed] [Google Scholar]
- 24.Joseph LB, et al. Sulfur mustard induced mast cell degranulation in mouse skin is inhibited by a novel anti-inflammatory and anticholinergic bifunctional prodrug. Toxicol. Lett. 2018;293:77–81. doi: 10.1016/j.toxlet.2017.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Composto GM, et al. Mitigation of nitrogen mustard mediated skin injury by a novel indomethacin bifunctional prodrug. Exp. Mol. Pathol. 2016;100:522–531. doi: 10.1016/j.yexmp.2016.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Katayama I, Tanei R, Yokozeki H, Nishioka K, Dohi Y. Induction of eczematous skin reaction in experimentally induced hyperplastic skin of Balb/C mice by monoclonal anti-DNP IgE antibody: possible implications for skin lesion formation in atopic dermatitis. Int. Arch. Allergy Appl. Immunol. 1990;93:148–154. doi: 10.1159/000235294. [DOI] [PubMed] [Google Scholar]
- 27.Yokozeki H, et al. In vivo transfection of a cis element 'decoy' against signal transducers and activators of transcription 6 (STAT6)-binding site ameliorates IgE-mediated late-phase reaction in an atopic dermatitis mouse model. Gene Ther. 2004;11:1753–1762. doi: 10.1038/sj.gt.3302341. [DOI] [PubMed] [Google Scholar]
- 28.Kitamura Y, Matsuda H, Hatanaka K. Clonal nature of mast-cell clusters formed in W/Wv mice after bone marrow transplantation. Nature. 1979;281:154–155. doi: 10.1038/281154a0. [DOI] [PubMed] [Google Scholar]
- 29.Patricio ES, et al. Mechanisms underlying the scratching behavior induced by the activation of proteinase-activated receptor-4 in mice. J. Invest. Dermatol. 2015;135:2484–2491. doi: 10.1038/jid.2015.183. [DOI] [PubMed] [Google Scholar]
- 30.Norman MU, et al. Mast cells regulate the magnitude and the cytokine microenvironment of the contact hypersensitivity response. Am. J. Pathol. 2008;172:1638–1649. doi: 10.2353/ajpath.2008.070559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Tanooka H, et al. Evidence for involvement of mast cells in tumor suppression in mice. J. Natl. Cancer Inst. 1982;69:1305–1309. [PubMed] [Google Scholar]
- 32.Galli SJ, Tsai M. IgE and mast cells in allergic disease. Nat. Med. 2012;18:693–704. doi: 10.1038/nm.2755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Schwartz LB, Lewis RA, Seldin D, Austen KF. Acid hydrolases and tryptase from secretory granules of dispersed human lung mast cells. J. Immunol. 1981;126:1290–1294. [PubMed] [Google Scholar]
- 34.MacDonald AJ, et al. IgE-mediated release of rat mast cell protease II, beta-hexosaminidase and leukotriene C4 from cultured bone marrow-derived rat mast cells. Immunology. 1989;67:414–418. [PMC free article] [PubMed] [Google Scholar]
- 35.Pawankar R, Okuda M, Yssel H, Okumura K, Ra C. Nasal mast cells in perennial allergic rhinitics exhibit increased expression of the Fc epsilonRI, CD40L, IL-4, and IL-13, and can induce IgE synthesis in B cells. J. Clin. Invest. 1997;99:1492–1499. doi: 10.1172/JCI119311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Pearce FL. Calcium and mast cell activation. Br. J. Clin. Pharmacol. 1985;20(Suppl 2):267S–274S. doi: 10.1111/j.1365-2125.1985.tb02812.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Siraganian RP, Zhang J, Suzuki K, Sada K. Protein tyrosine kinase Syk in mast cell signaling. Mol. Immunol. 2002;38:1229–1233. doi: 10.1016/s0161-5890(02)00068-8. [DOI] [PubMed] [Google Scholar]
- 38.Metcalfe DD, Peavy RD, Gilfillan AM. Mechanisms of mast cell signaling in anaphylaxis. J. Allergy Clin. Immunol. 2009;124:639–646. doi: 10.1016/j.jaci.2009.08.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Persia FA, Mariani ML, Fogal TH, Penissi AB. Hydroxytyrosol and oleuropein of olive oil inhibit mast cell degranulation induced by immune and non-immune pathways. Phytomedicine. 2014;21:1400–1405. doi: 10.1016/j.phymed.2014.05.010. [DOI] [PubMed] [Google Scholar]
- 40.Cariddi L, et al. Monoterpenes isolated from Minthostachys verticillata (Griseb.) Epling essential oil modulates immediate-type hypersensitivity responses in vitro and in vivo. Planta Med. 2011;77:1687–1694. doi: 10.1055/s-0030-1271090. [DOI] [PubMed] [Google Scholar]
- 41.Wallace DV, et al. The diagnosis and management of rhinitis: An updated practice parameter. J. Allergy Clin. Immunol. 2008;122:S1–84. doi: 10.1016/j.jaci.2008.06.003. [DOI] [PubMed] [Google Scholar]
- 42.Santos FA, Rao VS. Mast cell involvement in the rat paw oedema response to 1,8-cineole, the main constituent of eucalyptus and rosemary oils. Eur. J. Pharmacol. 1997;331:253–258. doi: 10.1016/s0014-2999(97)01013-3. [DOI] [PubMed] [Google Scholar]
- 43.Meurer SK, et al. Isolation of mature (peritoneum-derived) mast cells and immature (bone marrow-derived) mast cell precursors from mice. PLoS ONE. 2016;11:e0158104. doi: 10.1371/journal.pone.0158104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Takano H, et al. Establishment of the culture model system that reflects the process of terminal differentiation of connective tissue-type mast cells. FEBS Lett. 2008;582:1444–1450. doi: 10.1016/j.febslet.2008.03.033. [DOI] [PubMed] [Google Scholar]
- 45.Levi-Schaffer F, Austen KF, Gravallese PM, Stevens RL. Coculture of interleukin 3-dependent mouse mast cells with fibroblasts results in a phenotypic change of the mast cells. Proc. Natl. Acad. Sci. U.S.A. 1986;83:6485–6488. doi: 10.1073/pnas.83.17.6485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Gong L, et al. The antinociception of oxytocin on colonic hypersensitivity in rats was mediated by inhibition of mast cell degranulation via Ca(2+)-NOS pathway. Sci. Rep. 2016;6:31452. doi: 10.1038/srep31452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Nakamura T, et al. Glycyrrhetinic acid inhibits contact hypersensitivity induced by trichophytin via dectin-1. Exp. Dermatol. 2016;25:299–304. doi: 10.1111/exd.12931. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.