

Abstract
The study of extracellular vesicles (EVs) has the potential to identify unknown cellular and molecular mechanisms in intercellular communication and in organ homeostasis and disease. Exosomes, with an average diameter of ~100 nanometers, are a subset of EVs. The biogenesis of exosomes involves their origin in endosomes, and subsequent interactions with other intracellular vesicles and organelles generate the final content of the exosomes. Their diverse constituents include nucleic acids, proteins, lipids, amino acids, and metabolites, which can reflect their cell of origin. In various diseases, exosomes offer a window into altered cellular or tissue states, and their detection in biological fluids potentially offers a multicomponent diagnostic readout. The efficient exchange of cellular components through exosomes can inform their applied use in designing exosome-based therapeutics.
BACKGROUND:
All cells, prokaryotes and eu-karyotes, release extracellular vesicles (EVs) as part of their normal physiology and during acquired abnormalities. EVs can be broadly divided into two categories, ectosomes and exosomes. Ectosomes are vesicles that pinch off the surface of the plasma membrane via outward budding, and include microvesicles, microparticles, and large vesicles in the size range of ~50 nm to 1 μm in diameter. Exosomes are EVs with a size range of ~40 to 160 nm (average ~100 nm) in diameter with an endosomal origin. Sequential invagination of the plasma membrane ultimately results in the formation of multivesicular bodies, which can intersect with other intracellular vesicles and organelles, contributing to diversity in the constituents of exosomes. Depending on the cell of origin, EVs, including exosomes, can contain many constituents of a cell, including DNA, RNA, lipids, metabolites, and cytosolic and cell-surface proteins. The physiological purpose of generating exosomes remains largely unknown and needs investigation. One speculated role is that exosomes likely remove excess and/or unnecessary constituents from cells to maintain cellular homeostasis. Recent studies reviewed here also indicate a functional, targeted, mechanism-driven accumulation of specific cellular components in exosomes, suggesting that they have a role in regulating intercellular communication.
ADVANCES:
Exosomes are associated with immune responses, viral pathogenicity, pregnancy, cardiovascular diseases, central nervous system-related diseases, and cancer progression. Proteins, metabolites, and nucleic acids delivered by exosomes into recipient cells effectively alter their biological response. Such exosome-mediated responses can be disease promoting or restraining. The intrinsic properties of exosomes in regulating complex intracellular pathways has advanced their potential utility in the therapeutic control of many diseases, including neurodegenerative conditions and cancer. Exosomes can be engineered to deliver diverse therapeutic payloads, including short interfering RNAs, antisense oligonucleotides, chemotherapeutic agents, and immune modulators, with an ability to direct their delivery to a desired target. The lipid and protein composition of exosomes can affect their pharmacokinetic properties, and their natural constituents may play a role in enhanced bioavailability and in minimizing adverse reactions. In addition to their therapeutic potential, exosomes also have the potential to aid in disease diagnosis. They have been reported in all biological fluids, and the composition of the complex cargo of exosomes is readily accessible via sampling of biological fluids (liquid biopsies). Exosome-based liquid biopsy highlights their potential utility in diagnosis and determining the prognosis of patients with cancer and other diseases. Disease progression and response to therapy may also be ascertained by a multicomponent analysis of exosomes.
OUTLOOK:
The study of exosomes is an active area of research. Ongoing technological and experimental advances are likely to yield valuable information regarding their heterogeneity and biological function(s), as well as enhance our ability to harness their therapeutic and diagnostic potential. As we develop more standardized purification and analytical procedures for the study of exosomes, this will likely reveal their functional heterogeneity. Nonetheless, functional readouts using EVs enriched for exosomes have already provided new insights into their contribution to various diseases. New genetic mouse models with the ability for de novo or induced generation of cell-specific exosomes in health and disease will likely show the causal role of exosomes in cell-to-cell communication locally and between organs. Whether exosome generation and content change with age needs investigation, and such information could offer new insights into tissue senescence, organ deterioration, and programmed or premature aging. Whether EVs and/or exosomes preceded the first emergence of the single-cell organism on the planet is tempting to speculate, and focused bioelectric and biochemical experiments in the future could reveal their cell-independent biological functions. Single-exosome identification and isolation and cryoelectron microscopy analyses have the potential to substantially improve our understanding of the basic biology of exosomes and their use in applied science and technology. Such knowledge will inform the therapeutic potential of exosomes for various diseases, including cancer and neurodegenerative diseases.
Graphical Abstract
Exosomes: A cell-to-cell transit system in the human body with pleiotropic functions. Exosomes are extracellular vesicles generated by all cells and they carry nucleic acids, proteins, lipids, and metabolites. They are mediators of near and long-distance intercellular communication in health and disease and affect various aspects of cell biology.
The study of extracellular vesicles (EVs) and the mechanisms that govern their generation and function(s) in multicellular organisms spans from physiological tissue regulation to pathogenic injury and organ remodeling. Research in this field is stimulated by the potential of EVs as diagnostic and therapeutic tools for the treatment of various diseases, including neurodegeneration, cardiovascular dysfunction, and cancer. Increasingly, EV research is aimed at classification of EVs, isolation methods, and cataloging their putative functions in disease progression and therapy (1–5). Current characterization of biological activities of EVs has largely relied on tissue culture generated (and possibly amplified), nonphysiological readouts, as well as diverse EV isolation methods, which require further refinement (6, 7). Therefore, it remains unclear whether some of the purported properties of EVs are physiologically relevant in whole organisms in health or disease. Nonetheless, the production of EVs by cells appears to extend beyond a simple protein-recycling function, as initially reported for the transferrin receptor in reticulocyte maturation (8, 9), and varies according to cellular origin, metabolic status, and environment of the cells. EV research remains restricted by current experimental limitations in single-particle detection and isolation, and the inability to image and track exosomes in vivo at a reliable resolution. Despite such experimental caveats, exciting discoveries have emerged. The utility of EVs as liquid biopsies is particularly promising because of their presence in all biological fluids and their potential for multicomponent analyses (2).
Although the classification of EVs is continuously evolving (1), they generally fall into two major categories, ectosomes and exosomes (10) (Fig. 1). Ectosomes are vesicles generated by the direct outward budding of the plasma membrane, which produces microvesicles, microparticles, and large vesicles in the size range of ~50 nm to 1 μm in diameter. By contrast, exosomes are of endosomal origin and in a size range of ~40 to 160 nm in diameter (~100 nm on average). In this review, we focus on exosomes and discuss other EVs to offer contrast and comparison when relevant. Critically, challenges remain when establishing purification and analytical procedures for the study of exosomes, possibly resulting in a heterogeneous population of EVs that include exosomes. As such, some of the findings discussed may reflect those of exosomes mixed with other EVs. Exosomes are of particular interest in biology because their creation involves a distinct intracellular regulatory process that likely determines their composition, and possibly their function(s), once secreted into the extracellular space (2, 6, 11). It is important to recognize that exosome isolation methods are constantly evolving, and current biological markers may only recognize a subpopulation of exosomes with specific contents (1, 7, 12, 13). Therefore, some findings will need to be refined as new technology is embraced.
Fig. 1. Identity and the heterogeneity of extracellular vesicles and exosomes.
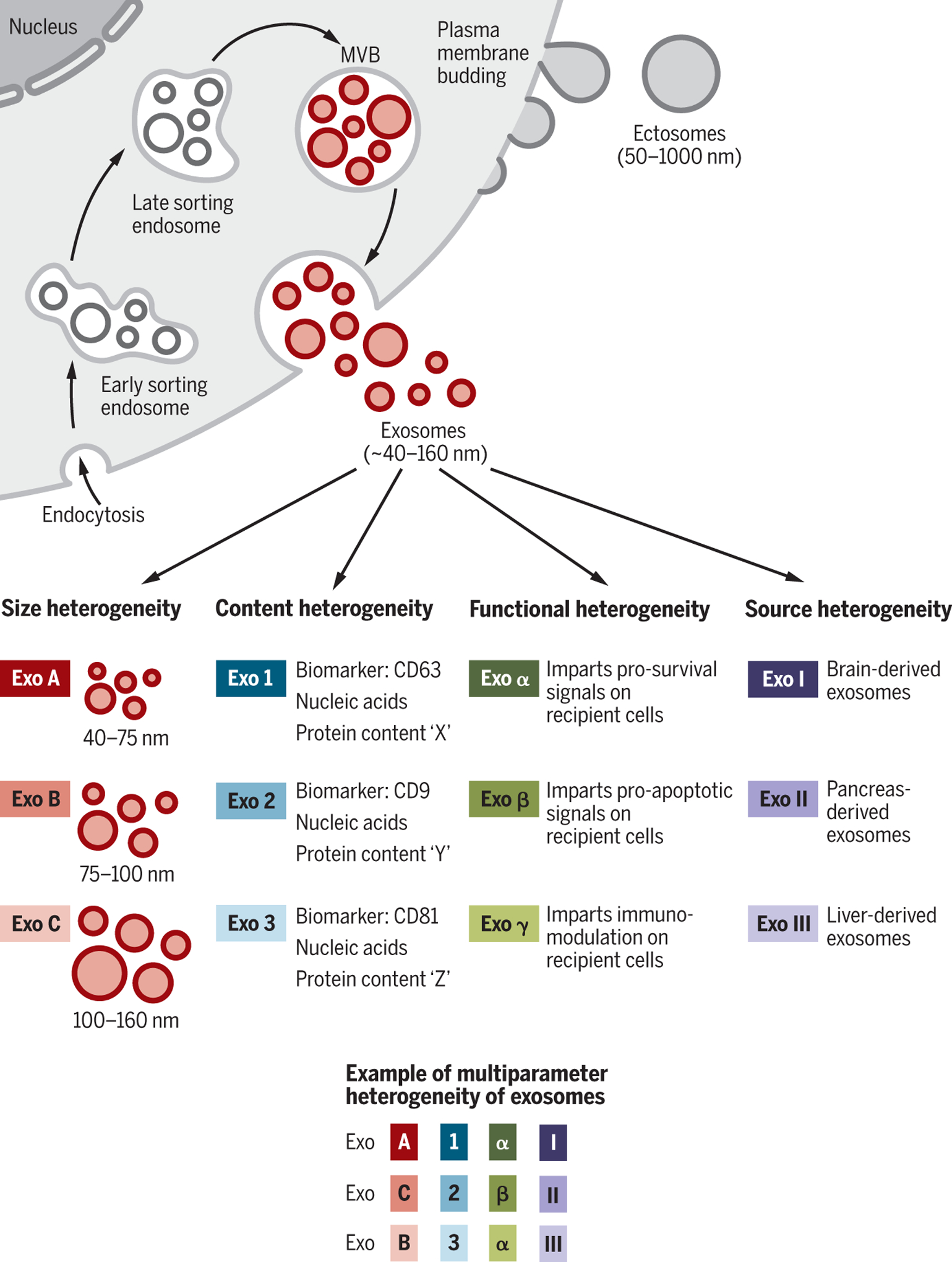
The two major categories of EVs are ectosomes and exosomes. Ectosomes are released through plasma membrane budding and are in the size range of ~50 nm to 1 μm. Exosomes originate from the endosomal pathway by the formation of the ESEs, LSEs, and ultimately MVBs, which contain ILVs. When MVBs fuse with the plasma membrane, exosomes are released (size range ~40 to 160 nm). Exosomes can be a highly heterogeneous population and have distinct abilities to induce a complex biological response. The heterogeneity of exosomes may be conceptualized on the basis of their size, content (cargo), functional impact on recipient cells, and cell of origin (source). Distinct combinations of these characteristics give rise to a complex heterogeneity of exosomes.
The biogenesis of exosomes
Exosomes are generated in a process that involves double invagination of the plasma membrane and the formation of intracellular multivesicular bodies (MVBs) containing intraluminal vesicles (ILVs). ILVs are ultimately secreted as exosomes with a size range of ~40 to 160 nm in diameter through MVB fusion to the plasma membrane and exocytosis (Fig. 1). The first invagination of the plasma membrane forms a cup-shaped structure that includes cell-surface proteins and soluble proteins associated with the extracellular milieu (Fig. 2). This leads to the de novo formation of an early-sorting endosome (ESE) and in some cases may directly merge with a preexisting ESE. The trans-Golgi network and endoplasmic reticulum can also contribute to the formation and the content of the ESE (2, 4–6, 13, 14). ESEs can mature into late-sorting endosomes (LSEs) and eventually generate MVBs, which are also called multivesicular endosomes. MVBs form by inward invagination of the endosomal limiting membrane (that is, double invagination of the plasma membrane). This process results in MVBs containing several ILVs (future exosomes). The MVB can either fuse with lysosomes or autophagosomes to be degraded or fuse with the plasma membrane to release the contained ILVs as exosomes (3, 4).
Fig. 2. Biogenesis and identification of exosomes.
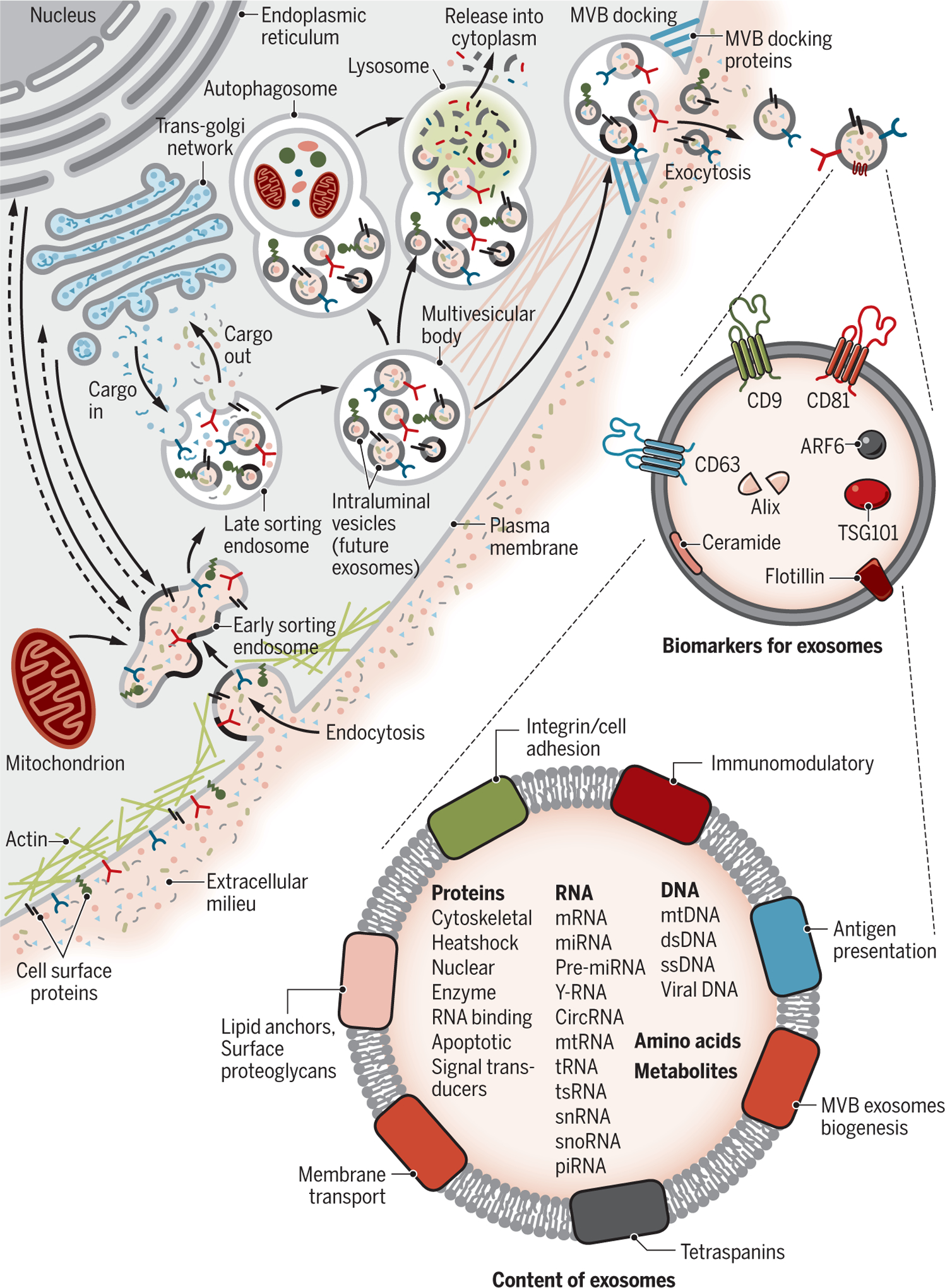
Fluid and extracellular constituents such as proteins, lipids, metabolites, small molecules, and ions can enter cells, along with cell surface proteins, through endocytosis and plasma membrane invagination. The resulting plasma membrane bud formation in the luminal side of the cell presents with outside-in plasma membrane orientation. This budding process leads to the formation of ESEs or possible fusion of the bud with ESEs preformed by the constituents of the endoplasmic reticulum (ER), trans-Golgi network (TGN), and mitochondria. The ESEs could also fuse with the ER and TGN, possibly explaining how the endocytic cargo reaches them. Some of the ESEs can therefore contain membrane and luminal constituents that can represent diverse origins. ESEs give rise to LSEs. Second invagination in the LSE leads to the generation of ILVs, and this step can lead to further modification of the cargo of the future exosomes, with cytoplasmic constituents entering the newly forming ILV. As part of the formation of ILVs, proteins (that were originally on the cell surface) could be distinctly distributed among ILVs. Depending on the invagination volume, the process could give rise to ILVs of different sizes with distinct content. LSEs give rise to MVBs with defined collection of ILVs (future exosomes). MVBs can fuse with autophagosomes, and ultimately the contents can undergo degradation in the lysosomes. The degradation products could be recycled by the cells. MVBs can also directly fuse with lysosomes for degradation. MVBs that do not follow this trajectory can be transported to the plasma membrane through the cytoskeletal and microtubule network of the cell and dock on the luminal side of the plasma membrane with the help of MVB-docking proteins. Exocytosis follows and results in the release of the exosomes with a similar lipid bilayer orientation as the plasma membrane. Several proteins are implicated in exosome biogenesis and include Rab GTPases, ESCRT proteins (see text), as well as others that are also used as markers for exosomes (CD9, CD81, CD63, flotillin, TSG101, ceramide, and Alix). Exosome surface proteins include tetraspanins, integrins, immunomodulatory proteins, and more. Exosomes can contain different types of cell surface proteins, intracellular protein, RNA, DNA, amino acids, and metabolites.
The Ras-related protein GTPase Rab, Sytenin-1, TSG101 (tumor susceptibility gene 101), ALIX (apoptosis-linked gene 2-interacting protein X), syndecan-1, ESCRT (endosomal sorting complexes required for transport) proteins, phospholipids, tetraspanins, ceramides, sphingomyelinases, and SNARE [soluble N-ethylmaleimide-sensitive factor (NSF) attachment protein receptor] complex proteins are involved in the origin and biogenesis process of exosomes, although their precise rate-limiting actions and functions in exosome biogenesis require further in-depth exploration, especially in vivo (6, 11, 15). An intersection of the exosome biogenesis pathway with other molecular pathways associated with the trafficking of intracellular vesicles has confounded the interpretation of functional studies. Specifically, loss- or gain-of-function experiments involving Rab and ESCRT proteins likely interfere with other distinct vesicular activities within cells, such as autophagy and lysosomal pathways, and Golgi apparatus-derived vesicle trafficking, which may exert indirect effects on exosome biogenesis. Distinct cell types, culture conditions, and genomic health of the cells may also favor or dispense some of the putative key regulators of exosome biogenesis in vivo (6). The potential inconsistencies in identifying regulatory elements associated with exosome biogenesis could also result from different methods for exosome production, enrichment, and concentration (13).
Computing the rate of exosome production is challenged by the dynamic process associated with the de novo production and uptake of external exosomes by any given cell type. A study using time-lapse monitoring of single human cells cultured in a platform that enabled tetraspanin antibody capture of shed exosomes indicated distinct rates of net exosome production by noncancerous versus cancerous breast epithelial cells. The breast cancer cells shed lower numbers of exosomes (~60 to 65 per cell per hour) compared with tissue-matched, nontumorigenic cell line-derived exosomes (16). In other studies, it has been suggested that cancer cells secrete more exosomes compared with normal cells from the same or other tissues [reviewed in (2, 11)], but such studies relied on different isolation methods that may measure both exosomes and ectosomes of similar size.
Exosome heterogeneity
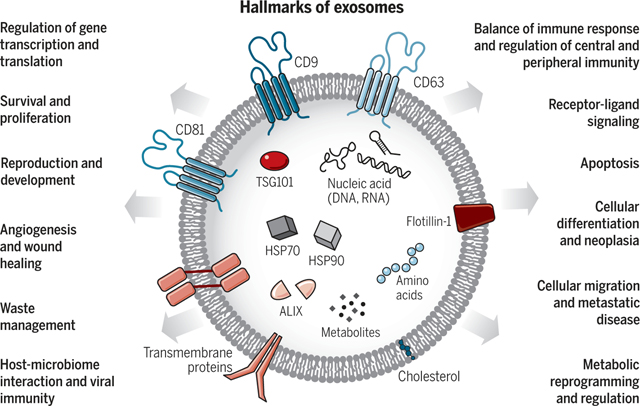
The heterogeneity of exosomes is likely reflective of their size, content, functional impact on recipient cells, and cellular origin (Fig. 1). Size inequality could be due to uneven invagination of the limiting membrane of the MVB, resulting in distinct total content of fluid and solids, or isolation methods that include other EVs (6, 11, 15). Refined fractionation methods involving EVs revealed that exosomes may contain subpopulations defined by a distinct size range (17). Size heterogeneity can also result in different amounts of exosomal content. The microenvironment and the inherent biology of the cells may influence the content of the exosomes and their biological markers. Exosomes can contain membrane proteins, cytosolic and nuclear proteins, extracellular matrix proteins, metabolites, and nucleic acids, namely mRNA, noncoding RNA species, and DNA (18–21) (Fig. 2). Although exosomal cargo analyses require large pools of purified exosomes, not all exosomes contain a similar abundance of a given cargo, as observed, for example, with miRNA exosomal cargo (22). Proteomic analyses of EVs have revealed the marker heterogeneity of exosomes, cautioning their utility in experimental design using marker-determined purification methodologies (23). Nonetheless, the proteome of breast cancer cells and their exosomes can show whether the cell of origin was epithelial like or mesenchymal like (24), and distinct proteins and nucleic acids are enriched in exosomes compared with their cell of origin, suggesting a specific protein-sorting mechanism associated with exosome biogenesis and/or content loading.
The effects of exosomes on recipient cells can be different because of their varied expression of cell surface receptors, and such functional heterogeneity can result in one set of exosomes inducing cell survival, another set inducing apoptosis, and a different set inducing immunomodulation, etc., in different target cell types (Fig. 1). Heterogeneity can also be based on the organ and tissue of origin of the exosomes, including whether they are from cancer cells (24), giving them distinct properties such as tropism to certain organs and uptake by specific cell types. A combination of all of these features would have the potential to give rise to a higher order of complexity and heterogeneity of exosomes.
Intercellular communication
The questions surrounding the function of exosomes are largely focused on understanding the fate of their constituents and the phenotypic and molecular alterations that they induce on recipient cells in cell-culture systems. Exosome uptake and secretion pathways may intersect, resulting in net production of a mixed population of exosomes over time for any given cell that is composed of both endogenously produced and recycled exosomes (Fig. 3). The distinct mechanisms and pathways associated with exosome uptake (6, 25, 26), and the putative specificity of exosomes for certain cell types, add complexity to the function of exosomes in intercellular communication. For example, oncogenic signals induced by mutant KRAS expression promote exosome uptake by macropinocytosis in human pancreatic cancer cells (27, 28). Human melanoma cells uptake exosomal cargo through their fusion with the plasma membrane (29), and the neurosecretory PC12 cells (derived from rat adrenal medullary tumor) more readily rely on clathrin-dependent endocytosis for uptake (30). It is unknown whether a different mode of exosome uptake by recipient cells results in distinct localization, degradation, and/or functional outcomes of the exosomal constituents. Moreover, it remains poorly understood whether administration of externally generated exosomes from different cell types into mice results in different organ tropism and/or retention compared with physiological tropism by de novo-produced exosomes (28, 31–35). It is possible that the “turnover rate” for internalized exosomal cargo varies depending on the nature of the cargo and the recipient cell’s metabolic status that regulates uptake of extracellular molecules and vesicles.
Fig. 3. Cellular journey of internalized exosomes and endogenously produced exosomes.
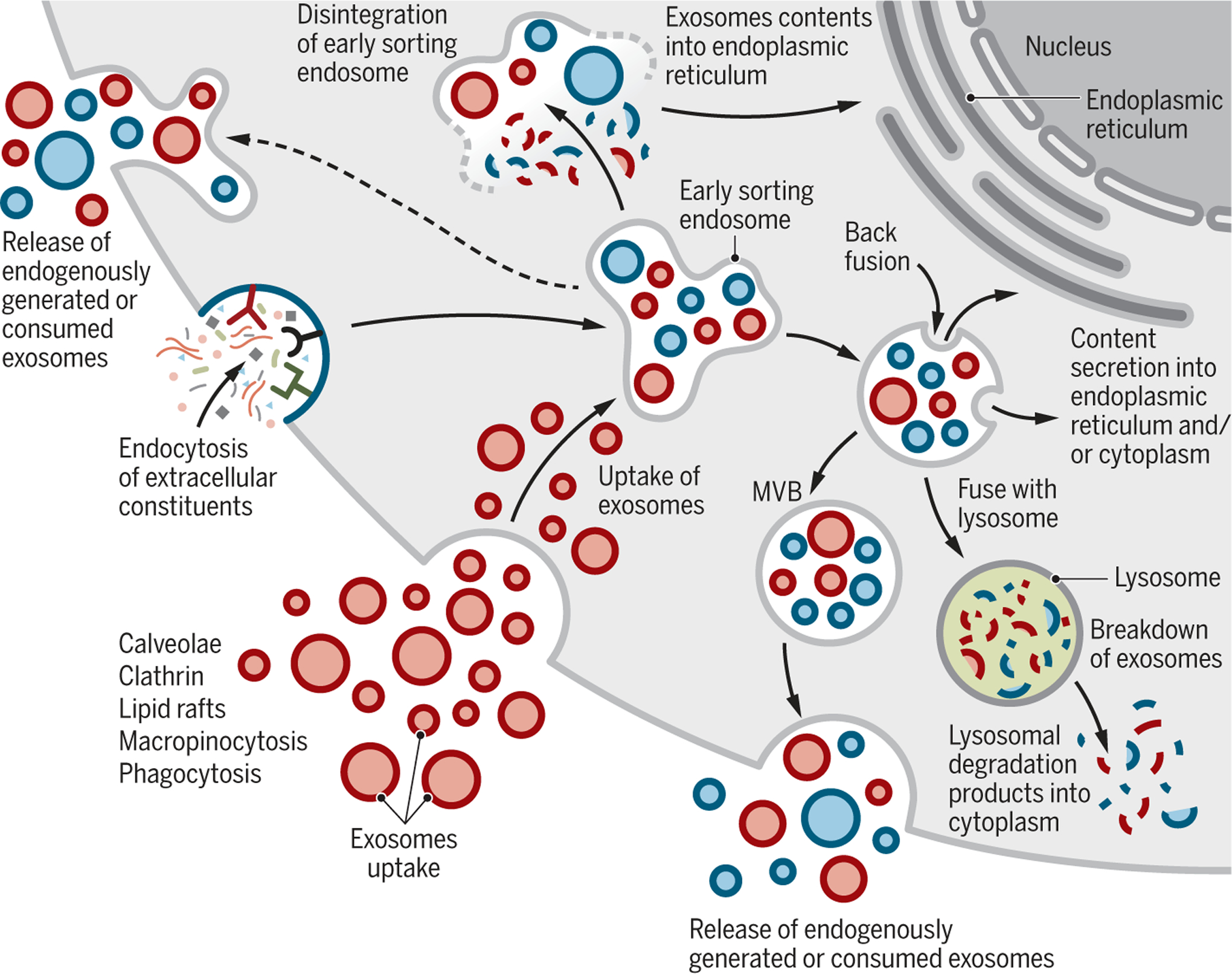
Exosomes may directly enter cells by different mechanisms (red). Exosomes are generated de novo by cells through the endocytosis process (blue). Exosomes are continuously being generated by and taken up by cells. It is likely that they can be secreted as a mixture of the de novo-generated and consumed exosomes (red and blue). It is unknown if the release of endogenously generated or consumed exosomes occurs together or separately. Exosomes that are taken up can get degraded by lysosomes. Exosomes that enter cells may enter or fuse with preexisting ESEs and subsequently disintegrate and release their contents into the cytoplasm. Alternatively, endosomes could fuse back with the plasma membrane and release exosomes outside the cells.
To track intercellular exchange of exosomes under physiological conditions, in vivo experiments involving various genetic strategies in mice were explored (36–38). These studies demonstrate that exosomes can deliver mRNA to a recipient cell on rare occasions. Such rare events were enhanced by the activation and expansion of exosome-producing immune cells in mouse models of acute inflammation (peritonitis) or chronic inflammation (subcutaneous tumor) (36). Therapeutic interventions such as chemotherapy could also influence exosome uptake and subsequent biological responses of tumors. For example, inhibiting a proton pump or altering cellular pH in melanoma cells limits exosome uptake (29).
Although it is not surprising that the proteome of the exosomes reflects the proteome of the originating cell, exosome protein cargo from cancer cells can be altered. For example, proteomic studies have revealed that oncogenic alteration, such as constitutively active expression of EGFRvIII (epidermal growth factor receptor variant III) in glioblastoma cells, yields exosomes with a protein cargo specifically enriched in many proinvasive molecules (39). Neural stem cells challenged with inflammatory cytokines produce exosomes with IFNγ (interferon gamma)-bound-IFNGR1 (interferon gamma receptor 1), and these specifically activate STAT1 (signal transducer and activator of transcription 1) signaling in recipient cells that also express IFNGR1 (40). These results support the idea that proteins are sorted into exosomes and can selectively induce specific signals in recipient cells to regulate processes such as those seen in development, immune responses, and disease.
Mammalian reproduction and development
Human reproduction, pregnancy, and embryonic development require precise, finely tuned, and dynamic intercellular communication. Semen, amniotic fluid, blood, and breast milk all contain exosomes with putative functions. Seminal plasma exosomes have been implicated in sperm maturation (41). Molecular profiling indicates that the microRNAs let-7a, let-7b, miR-148a, miR-375, and miR-99a are enriched in seminal plasma-derived exosomes from multiple human donors (42). These miRNAs are implicated in the expression of interleukins (IL-10 and IL-13), raising the possibility that exosomes play a role in genitalia-resident immunity (42). Seminal plasma-derived exosomes also inhibit HIV-1 infection (43, 44), possibly by blocking HIV early protein transcriptional activator (Tat) recruitment and subsequent transcription of HIV-1 (45) (Fig. 4).
Fig. 4. Exosomes in viral infection.
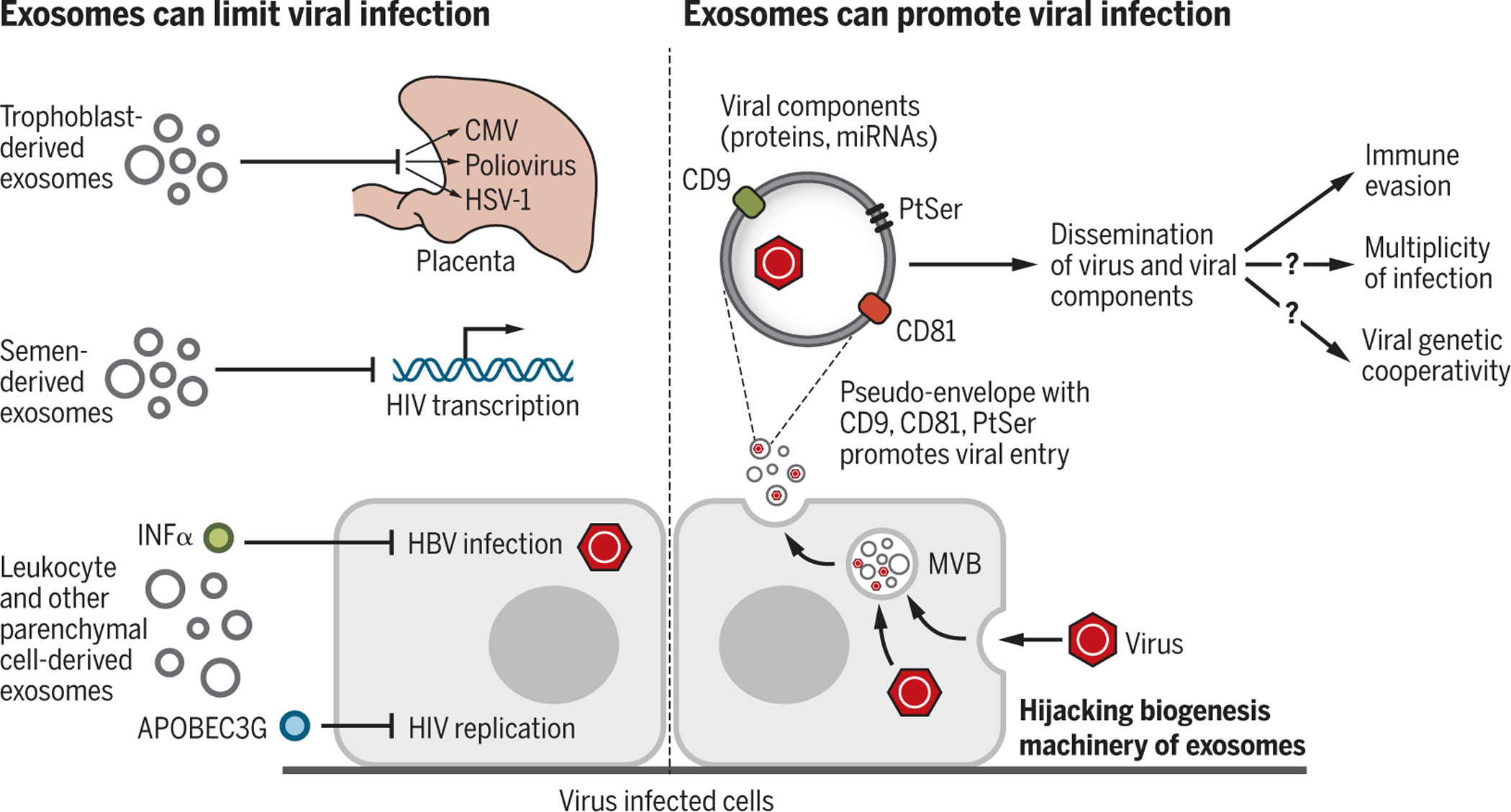
Exosomes can limit or promote viral infection. Exosomal cargo such as IFNα or APOBEC3G can suppress infection by limiting viral replication or enhancing antiviral immunity. Viruses can also highjack the exosome biogenesis machinery to promote viral dissemination. Exosomes may serve as a pseudoenvelope that enhances viral entry by tetraspanins (CD81, CD9) and PtSer interaction and uptake into recipient cells and aid in evading antiviral immunity. Cotransport of a viral component (proteins and miRNA) may also enhance infectivity. Exosome-mediated transfer of viruses may participate in viral genetic cooperativity and multiplicity of infection. CMV, cytomegalovirus; HSV-1, herpes simplex virus 1.
Exosomes may also help to prevent infection of the placenta by delivery of exosomal miRNA (chromosome 19 miRNA cluster, C19MC) from specialized cells of the placenta (trophoblasts) to nonplacental cells to induce autophagy and defense against viral infections such as poliovirus, human cytomegalovirus, and herpes simplex virus 1 infection (46). In the blood plasma of pregnant women, the exosomal miRNA and protein cargo change with respect to gestational age and when compared with preterm birth (47, 48). Plasma-derived exosomes dynamically evolve during pregnancy in mice as well, and gestationstage specific exosomes are functionally linked to labor and delivery (49). Specifically, late-gestation plasma-derived exosomes induce preterm birth in near-term pregnancies in mice but do not affect pregnancies at an earlier stage of gestation (49).
In breast milk, exosomes seem to promote postnatal health and growth. Breast milk-derived exosomes contain miRNAs with immune-related functions (50) and enhance the number of peripheral blood-derived T-regulatory cells ex vivo, possibly to regulate immune tolerance (51). Although breast milk-derived exosomes have been shown to promote the proliferation of porcine intestinal epithelial cells in culture conditions and the mouse intestinal tract in vivo (52), it remains unclear to what extent, if any, the transfer of nucleic acids and other exosomal cargos is preserved after ingestion, having to overcome digestive enzymes and uptake in the intestinal epithelium. The impact of different routes of administration on tissue uptake of exosomes will likely influence potential therapeutic strategies (53).
Immune responses and infection
The role of exosomes in immune responses has been widely documented (54–56), although it should be noted that no severe immune reaction was observed in mice repeatedly administered with a relatively low dose of mouse or human cell-derived exosomes for extended periods of time (28, 35, 57). Whole-blood and plasma transfusions, which have been performed for >50 years, do not appear to be associated with potential EV-mediated immune reactions despite no effort to match HLA (human leukocyte antigen) types and the infusion of trillions of EVs including exosomes. Exogenous administration and endogenous secretion of exosomes may thus elicit immune responses in a context- and dose-dependent manner and this remains to be elucidated.
Recent experiments with engineered exosomes have nonetheless indicated a function of exosomes in eliciting adaptive and innate immune reactions, supporting their utility for therapy development and a potential role in coordinating immune reactions in response to infectious agents or cancer (Fig. 5). The function of exosomes in immune regulation is likely due to the transfer and presentation of antigenic peptides, delivery of DNA-inducing cGAS-STING (cyclic GMP-AMP synthase stimulator of interferon genes) signaling in recipient cells (an immune pathway wherein sensing of cytosolic DNA triggers the expression of inflammatory genes and a type I IFN response), gene-expression manipulation by exosomal miRNA, and induction of different signaling pathways by surface ligands present on the exosomes.
Fig. 5. Regulation of immune response by exosomes.
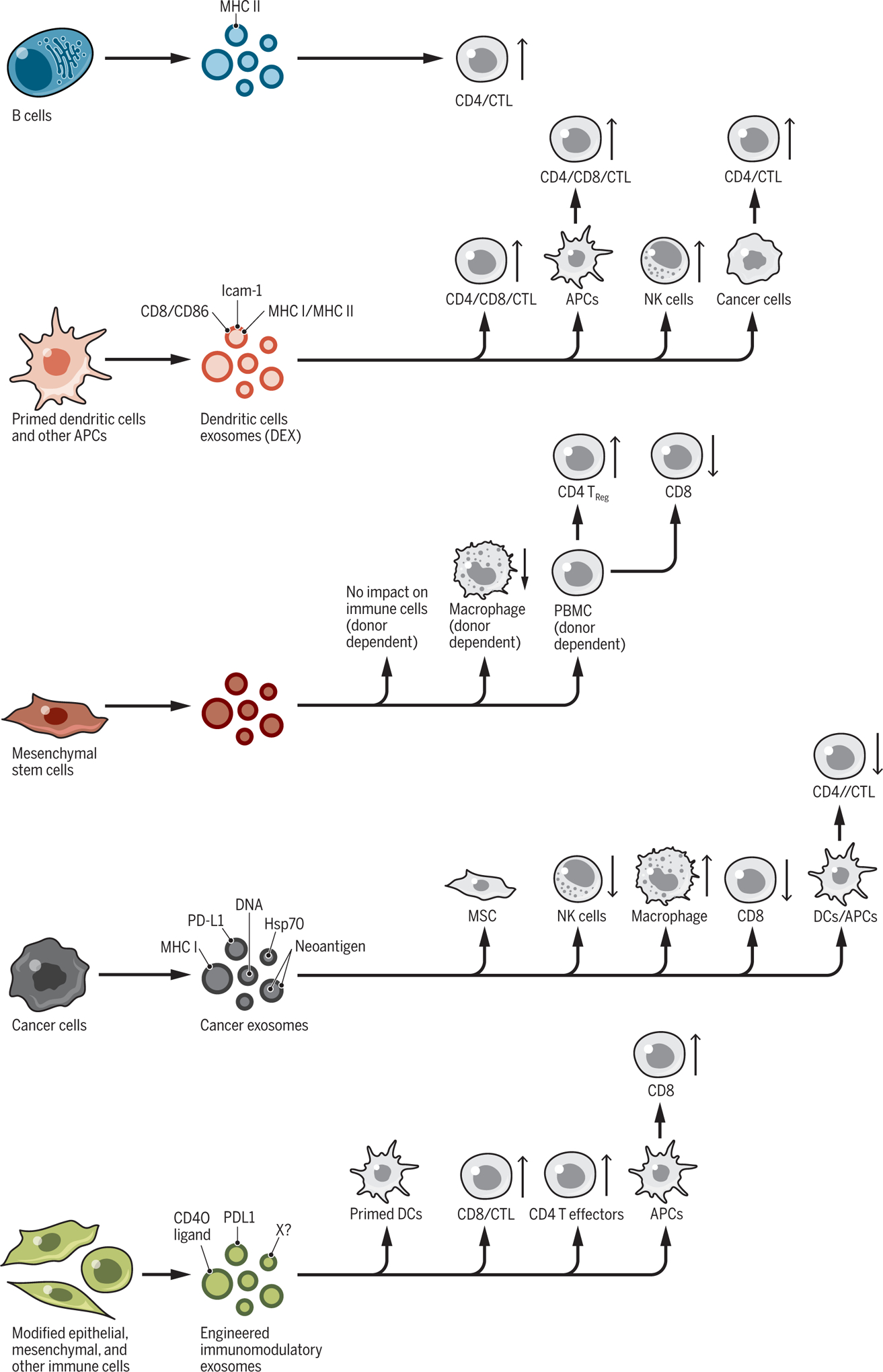
Exosomes from distinct cellular sources, including immune cells (B cells and dendritic cells), cancer cells, epithelial, and mesenchymal cells, shed exosomes with cargos that can influence the proliferation and respective activity of recipient cells of both the innate and adaptive immune system. CD4+ and CD8+ T cells [cytotoxic T cells (CTL)] can be directly or indirectly influenced by exosomes, stimulating or suppressing their proliferation and function(s). PBMC, peripheral blood mononuclear cell; X?, other potential immunomodulatory proteins.
Exosomesfromantigen-presenting cells(APCs) carry p-MHC-II [major histocompatibility complex II with antigenic peptide (p)] and costimulatory signals, and directly present the peptide antigen to specific T cells to induce their activation. However, for reasons that remain to be elucidated, T cell stimulation by exosomes is less effective than that by APCs (58–60). In mice, tumor eradication and growth delay were observed with a single intradermal injection of APC-derived exosomes with MHC-II loaded with tumor peptide (60). The potency and durability of the observed CD8+ cytotoxic T cell-mediated antitumor response also implied indirect antigen presentation because of the transfer of the antigenic peptide on exosomes to the APCs, which in turn prime naïve T and/or B cells for activation. Immature mouse dendritic cells activated by exosome-derived immunogenic peptides indirectly activate APCs and induce specific CD4+ T cell proliferation (61). Exosomes shed by human dendritic cells, regardless of their maturity, promote a T helper 1 response (IFNγ production) in culture (62). Exosomes from ovalbumin (OVA)-pulsed dendritic cells were more efficient in eliciting antigen (OVA)-specific CD8+ T cell activation than were microvesicles (63), supporting a potential molecular intersection between exosome biogenesis (which is distinct from microvesicle biogenesis, as discussed above) and antigen presentation. The role of exosomes in antigen presentation was also explored in the context of bacterial infection (with a focus on Mycobacterium tuberculosis and Helicobacter pylori), wherein exosomes may enhance antibacterial immune responses by promoting bacterial antigen presentation from macrophage-derived exosomes (64). This could subsequently influence the adaptive immune response (64); the production of IFNα and IFNγ, tumor necrosis factor α (TNFα), and IL-containing exosomes from macrophages to promote dendritic cell maturation and CD4+ and CD8+ T cell activation (65); and the regulation of macrophage IL expression (66). Bacteria-derived EVs have also been identified in humans and have implications in health and disease (67, 68), and given the emerging role of exosomes in antigen presentation in the context of bacterial infection, it is plausible that exosomes would play a role in microbiota-associated inflammation.
The nucleic acid exosomal cargo, namely DNA and miRNA, has been implicated in regulating innate and adaptive immune responses. The DNA of intracellular bacteria (e.g., Listeria, Legionella pneumophila, and Franciscella tularensis) are sorted into exosomes with the capacity to stimulate cGAS-STING signaling in nearby cells, effectively activating innate immune responses. However, in the case of Listeria, this happens at the expense of suppressing T cells and thus lowering antibacterial defense (69). By contrast, in the context of M. tuberculosis infection, bacterial RNA shed from infected macrophages enhances host immunity by eliciting the RNA-sensing pathway and promoting phagosome maturation in recipient macrophages (70). Although the functional role of exosomes in immune responses against fungal and parasitic infections is largely unknown, some studies related to parasite-derived exosomes have indicated that exosomes may participate in disease virulence (71, 72). Specifically, the malaria-causing parasite Plasmodium falciparum was shown to shed its DNA and small RNAs into the exosomes from the red blood cells that it infects (73). Instead of enhancing the STING-dependent antipathogen immune response, human monocytes that take up exosomes containing the parasitic DNA may elicit STING-dependent DNA sensing as a decoy strategy to enhance parasite survival (73).
The role of exosomal DNA in the immune response was also shown to be functionally relevant to cancer progression. Adaptive immune responses elicited by exosomes include the activation of dendritic cells with the uptake of breast cancer cell-derived exosomal genomic DNA and activation of cGAS-STING signaling and antitumor response in mice (74). In vitro, after T cell contact, the priming of dendritic cells (immune instruction that changes the activity of dendritic cells to enhance their response to future stimulation) is also associated with the uptake of exosomal genomic and mitochondrial DNA (mtDNA) from T cells, inducing type I IFN production by cGAS-STING signaling (75). Although exosomal DNA uptake by recipient cells alters their signaling, exosome biogenesis may also play a role in clearing cytoplasmic DNA and in preventing activation of the cytosolic DNA-sensing machinery and reactive oxygen species-dependent DNA damage response (76). In the context of cancer, this exosomal DNA shedding may be beneficial, such that inhibition of EGFR in cancer cells leads to increased DNA in the exosomes from those cells and could induce cGAS-STING signaling in dendritic cells and contribute to overall suppression of tumor growth (77). By contrast, the impact of tumor exosomal DNA on inflammatory responses can indirectly worsen cancer, and uptake of tumor-derived exosomal DNA by circulating neutrophils was shown to enhance the production of tissue factor and IL-8, which play a role in promoting tumor inflammation and paraneoplastic events (thrombosis) (78).
Exosomes may also regulate the immune response by influencing gene expression and signaling pathways in recipient cells, principally by the transfer of miRNAs. Exosomal miRNA can exchange between dendritic cells and repress gene expression (79), and such exosome-mediated intercellular communication may influence dendritic cell maturation. Tumor-derived exosomal miR-212–3p down-regulates the MHC-II transcription factor RFXAP (regulatory factor X associated protein) in dendritic cells, possibly promoting immune evasion by cancer cells (80). Tumor-derived exosomal miR-222–3p down-regulates SOCS3 (suppressor of cytokine signaling 3) in monocytes, which promotes STAT3-mediated M2 polarization of macrophages (81), possibly generating an immunosuppressive microenvironment.
Modulation of immune responses by exosomes might also involve presentation of immunoregulatory molecules such as PD-L1 (programmed cell death ligand 1) and FasL (Fas cell surface death receptor ligand) on their surface. PD-L1 on melanoma-derived exosomes suppresses CD8+ T cell antitumor function in vivo (82), and cancer cell-derived exosomes block dendritic cell maturation and migration in a PD-L1-dependent manner (83). Further, cancer cell-derived PD-L1+ exosomes promote T cell exhaustion in the draining lymph nodes of tumor-bearing mice, promoting tumor growth (84). FasL on melanoma or prostate cancer-derived exosomes induces Fas-dependent apoptosis of T cells (85, 86). In addition to exosomes, ligand-mediated signaling to T cells and enzymatic activities associated with exosomes derived from multiple human cancer cell types that express with CD39 (ectonucleoside triphosphate diphosphohydrolase 1) and CD73 (5′ nuleotidase) result in 5′AMP-to-adenosine conversion and adenosine signaling in T cells, effectively limiting their activation in vitro (87). Such actions could ultimately suppress the adaptive immune response. By contrast, mast cell-derived exosomes express MHC-II, CD86, LFA-1 (lymphocyte function-associated antigen 1), and ICAM-1 (intercellular adhesion molecule 1) and induce the proliferation of B and T cells in vitro and in vivo (88). Finally, cancer cell-derived exosomes engineered to overexpress CD40L (CD40 ligand or CD154, a costimulatory molecule on T cells that binds to CD40 on APCs) promotes dendritic cell maturation, resulting in increased proliferation of T cells and antitumor activity in vivo (89).
The role of exosomes in the innate immune response in cancer has also been reported. Exosomes from pancreatic cancer cells and plasma of pancreatic cancer patients were shown to limit complement-mediated lysis by acting as decoys, thus decreasing cytotoxicity against cancer cells (90). Exosomal HSP72 (heat shock protein 70 kDa protein 1) can trigger myeloid-derived suppressor cell activation by STAT3 (91), and exosomes derived from glioblastoma stem cells induce a STAT3-mediated immunosuppressive (M2 type) switch of macrophage phenotype (92), which would limit antitumoral immune response in the tumor microenvironment. Exosomal miR-21 and miR-29a from cancer cells trigger human TLR8 and mouse TLR7-mediated NF-κB (nuclear factor-κB) activation in macrophages and the production of IL-6 and TNF-α to promote melanoma lung metastasis and lung cancer in mice (93).
Exosomes not only play a role in immune responses related to cancer cells, but also those associated with infectious agents (bacteria, viruses, fungi, and parasites) (71, 94). Exosomes might promote viral infection by enabling the dissemination of viral components, and viruses may highjack the exosome biogenesis pathway for their survival [reviewed in (95)] (Fig. 4). Viral infection associated with both enveloped and nonenveloped virus is regulated by exosomes. The prototypic nonenveloped hepatitis A and hepatitis E viruses (HAV and HEV, respectively) can exist in a pseudoenveloped form within exosomes (96, 97). The use of the exosome biogenesis machinery by viruses and exosomes as a pseudoviral envelope evokes a “Trojan horse” ploy to favor viral entry into the cell, thereby enhancing infectivity (98). The similarities—in size, density, molecular cargo, and use of common components to harness the cellular proteins and vesicle-trafficking machinery—between enveloped retroviruses (in particular, HIV-1/2) and exosomes support this idea (98, 99). It has been proposed that multiple viruses may package within exosomes, a process that would promote multiplicities of infection and viral genetic cooperativity (99). Although it remains unclear whether exosomes participate in viral immune evasion by limiting detection by neutralizing antibodies (96), they take part in augmenting viral entry into cells through the tetraspanins (transmembrane proteins) CD81 and CD9 present on exosomes, possibly by stabilizing the interaction of exosomes containing virus particles with the cellular plasma membrane and delivery of viral constituents (100–103). Similarly, the phosphatidylserine (PtdSer) receptor TIM-4 (T cell immunoglobulin and mucin domain containing 4) on exosomes may facilitate the cellular entry of HIV-1 because of its PtdSer-rich envelope (104).
A postulated advantage of viruses using exosomes to exit cells could be to evade inflammation and prevent virus-induced cell lysis. Tumor-derived transfer of EGFR-associated exosomes to macrophages weakens their antiviral response in a MEKK2 (mitogen-activated protein kinase kinase 2)- and IRF3 (interferon regulatory factor 3)-dependent manner, suggesting that cancer may enhance viral infection (105). Although exosomes shed from virus-infected cells can promote infection (see above), exosomes also participate in antiviral immunity. For example, IFNα-stimulated human macrophages shed exosomes that deliver antiviral mediators, including the single-stranded DNA cytidine deaminase APOBEC3G (apolipoprotein B mRNA editing enzyme, catalytic polypeptide-like 3G), protecting human hepatocytes from HBV (hepatitis B virus) infection (106). Exosomal APOBEC3G from exosomes also impairs HIV-1 infection of T cells (107). The HIV-1 receptor CD4 on exosomes from CD4+ T cells was shown to reduce HIV-1 infection in vitro, and the HIV-1 accessory protein Nef in infected T cells reduced the expression of exosomal CD4, effectively enhancing HIV-1 infection (108). Future studies will hopefully further clarify the opposing roles of exosomes in HIV-1 infection in vivo.
Metabolic and cardiovascular diseases
Exosomes may play a role in the emergence of metabolic diseases as well as in cardiovascular fitness. They have been found to transfer metabolites and to facilitate intercellular communication through exosomal miRNA exchange among pancreatic β-cells, adipose tissue, skeletal muscles, and the liver of mice and humans (109). Reciprocal signaling between adipocytes and macrophages mediated by exosomes in the Leptin gene-knockout spontaneous mouse model of obesity implicates RBP4 (retinol binding protein 4) in stimulation of macrophages and insulin resistance (110). Obese mice fed a high-fat diet display distinct circulating exosomal miRNAs, which are sufficient to promote insulin resistance in lean mice, possibly through down-regulation of Ppara (peroxisome proliferator-activated receptor alpha) expression in white adipose tissue (111). Cachexia, a condition of severe weight loss and muscle wasting associated with chronic disease such as cancer, as well as other metabolic paraneoplastic syndromes (e.g., new-onset diabetes in pancreatic cancer), may be exacerbated by cancer cell-derived exosomal cargo acting on mouse and human adipocytes and muscle cells (112). Adrenomedullin, a peptide hormone inducing lipolysis, was found in exosomes generated by human pancreatic cancer cells and induced lipolysis in mouse and human adipocytes (113) and inhibited insulin secretion in rat and human islet cells (114). Mouse and human cancer cell derived-exosomes, which are rich in heat shock proteins (HSP70 and HSP90), are also functionally implicated in muscle wasting in mice (115). These findings support that cancer cell-derived exosomes can change the metabolism of noncancer cells, including adipocytes and pancreatic islet cells, thus functionally contributing to the development of cachexia and paraneoplastic syndrome.
Exosomes from mouse and human cell culture supernatant (endothelial cells, cardiac progenitor cells, cardiac fibroblasts, cardiomyocytes) have been associated with metabolic disease, including atherosclerosis, diabetes-related cardiovascular disease (CVD), and metabolic adaptation associated with heart failure (116). The functions of exosomes in preventing atherosclerosis was demonstrated in mice, where platelet-derived exosomes reduced macrophage scavenger receptor CD36 expression and consequently reduced the uptake of harmful cholesterol [oxidized low-density lipoprotein (LDL)] (117). By contrast, human smooth muscle cell-derived exosomes may promote thrombogenesis, as shown by in vitro assays (118). The use of stem cell (bone marrow-derived stem cells and embryonic stem cells)-derived exosomes in cardiovascular protection (119) has emerged as a potential therapeutic approach in mice and rats despite limited knowledge on how exosomes potentiate these effects. Exosomal microRNAs, including miR-19a, miR-21 [from murine cardiac progenitor cells targeting PDCD4 (programmed cell death 4) in rat myoblasts in vitro (120)], miR-22 [from mouse bone marrow-derived mesenchymal stromal cells (MSCs) targeting MECP2 (methyl CpG binding protein 2) in ischemic mouse heart (121)], and miR-21–5p [from human bone marrow-derived MSCs targeting SERCA2a (sarcoendoplasmic reticulum Ca2+) adenosine triphosphatase (ATPase) and L-type calcium channels in human cardiac myocytes derived from pluripotent stem cells in vitro (122)], mediate cardiovascular protective effects, possibly by limiting cardiomyocyte apoptosis, promoting mitochondrial function, and preserving cardiac contractility.
Neurodegeneration
The intersection between exosomal biogenesis and the regulation of secretory vesicles in neuronal cells offered new insight into the putative connection between exosomes and the pathogenesis of neurodegenerative diseases. Exosomes may promote or limit aggregation of unfolded and abnormally folded proteins in the brain (123–128). Exosomes could participate in the clearing of misfolded proteins, thereby exerting detoxifying and neuroprotective functions, or participate in the propagation and aggregation of misfolded proteins, effectively promoting the “infectivity” of protein aggregates and contributing to disease progression. Such opposing functions of exosomes might not be mutually exclusive and are described below.
Pharmacological blocking using GW4869 (which inhibits inward budding of MVBs) or enhancement of exosome production using monensin (which increases intracellular Ca2+ and MVB generation) results in a decrease or increase, respectively, of the transmission of the infectious prion protein PrPsc, which is associated with Creutzfeldt-Jakob disease in vitro (129). Both Tau and Aβ (β-amyloid generated by the cleavage of amyloid precursor protein [APP]), implicated in Alzheimer’s disease, are found in exosomes, including patients’ cerebrospinal fluid-derived exosomes (Tau), mouse microglial cell culture supernatant-derived exosomes (Tau), and exosomes of supernatant from the culture of mouse and human cell lines (Aβ). Pathological propagation of Tau aggregation by exosomes was noted in vitro and in vivo (130, 131). Using a simple circuit of neurons in a microfluidic device, exosomal transfer of Tau between neurons was proposed to include takeover of the endosomal pathway (131). The cleavage of APP was observed in early endosomes, and Aβ accumulated in MVBs of N2a (mouse neuroblastoma) and HeLa cells modified to express fluorescent APP (132); however, whether exosomes promote neurotoxic Aβ oligomerization in vivo is unknown.
The exosome biogenesis machinery may also be neuroprotective. Exosomes may impair neurotoxic oligomer formation (133) or exosomes may carry them out of cells (134). More recently, exosomal secretion of Aβ from the brains of mice engineered to overexpress APP was implicated in the initiation and propagation of toxic amyloid. This process involves the deregulation of ECE1/2 (endothelin-converting enzyme 1/2), effectively resulting in an increase in oligomerized Aβ in exosomes from the brains of APP-transgenic mice (135).
Similar observations were made in distinct proteinopathies such as Parkinson’s disease (PD) and amyotrophic lateral sclerosis (ALS). The pathological protein α-synuclein is found in cerebrospinal fluid-derived exosomes of patients with PD or dementia with Lewy bodies (136), and the exosome biogenesis machinery is implicated in the accumulation α-synuclein, with α-synuclein downregulating ESCRT and limiting its intracellular degradation (137). SOD1 (superoxide dismutase 1) and TDP-43 (transactive response DNA binding protein 43 kDa), two misfolded proteins associated with ALS, have been identified in exosomes (138–140). Exosomes containing SOD1 from mouse astrocytes resulted in the death of mouse spinal cord-derived motor neurons in culture (138), mutant SOD1 could be transferred between human mesenchymal cells in vitro (139), and TDP-43 was found in exosomes from the culture supernatant of mouse neuroblast cells (140). However, in vivo suppression of exosome secretion using GW4869 in TDP-43A325T-transgenic mice was detrimental because this appeared to limit the clearance of pathological TDP-43 from neurons (140). Although exosomes containing neurotoxic proteins could be transferred to distinct cell types in vitro (see above), possibly promoting disease progression, it remains unknown whether exosome-mediated exchange of such proteins affects—either positively or negatively—disease progression in vivo.
Although the function of exosomes in neurodegenerative disorders has focused on exosome control of misfolded protein accumulation, nucleic acids and other constituents may be implicated in worsening or ameliorating other neurological disorders. In a study evaluating the serum-derived exosomes of children with autism spectrum disorder (ASD), mtDNA exosomal cargo was proposed to illicit microglia IL-1β secretion, possibly contributing to the inflammation associated with ASD (141). The role of exosomes in the pathophysiology of neurodegenerative disorder and ASD requires more study, but this has not hindered efforts to use them in therapy development. Such effort is largely encouraged by the intrinsic properties of exosomes to efficiently pass through the blood-brain barrier, a vascular network functioning as a selective filter to keep drugs or toxins from reaching the brain (28, 142–144).
Cancer
The study of exosomes in cancer has progressed at a rapid pace compared with research into their role in other diseases (2, 145), and exosomes have been associated with several hallmark features of cancer (146). Exosomes influence neoplasia, tumor growth and metastasis, paraneoplastic syndromes, and resistance to therapy. The role of exosomes in cancer progression is likely dynamic and specific to cancer type, genetics, and stage.
Exosomes may induce or promote neoplasia. Exosomes from pancreatic cancer cells were shown to initiate cell transformation by inducing mutations in NIH/3T3 recipient cells (147). Exosomes derived from breast cancer and prostate cancer cells induce neoplasia through transfer of their miRNA cargo (148, 149). miR-125b, miR-130, miR-155, as well as HRas and Kras mRNAs in exosomes from prostate cancer cells, participate in neoplastic reprogramming and tumor formation of adipose stem cells (149). The plasticity of cancer cells may also be attributed in part to exosomes, with exosomal miR-200 from metastatic breast cancer cells enhancing the epithelial to mesenchymal transition (EMT) and metastasis of otherwise weakly metastatic breast cancer cells (150). Although more work is needed to decipher the rate-limiting role of exosomes in neoplasia and EMT, research has focused on the exchange of exosomal cargo between cancer cells and stromal cells in the tumor microenvironment and on defining the functional outcome of such exchange on tumor growth and metastasis. These studies have explored cancer in mouse models and often rely on exogenously administered exosomes in mice.
In most studies, the stromal cell recipients of cancer cell-derived exosomes are cancer-associated fibroblasts (CAFs) and immune cells, which dynamically regulate one another in the tumor microenvironment. Distinct cancer cell-derived exosomal cargo, such as nucleic acids, signaling proteins, and metabolites, can exert protumorigenic effects on stromal cells. For example, breast cancer exosome-derived miR-122 suppresses pyruvate kinase and subsequent glucose uptake in the lungs, which promotes metastasis (151). Although RNA shielded by proteins prevents their recognition as pathological RNAs that would otherwise elicit inflammatory responses, breast cancer cells induce the accumulation of unshielded RN7SL1 (RNA component of signal recognition particle 7SL1) RNA in exosomes from CAFs, which ultimately produces a proinflammatory response when delivered to immune cells and results in increased tumor growth and metastasis in mice (152).
Examples of exosomes from cancer cells eliciting a parenchymal signaling response at metastatic sites, effectively remodeling distant microenvironments to enhance metastasis, have been reported in multiple cancer types. For example, TGFβ (transforming growth factor-β) expressed on the surface of cancer cell-derived exosomes induces fibroblast activation by gain of αSMA (α-smooth muscle actin) and FGF2 (fibroblast growth factor 2) expression (153). The recruitment of bone marrow progenitor cells and macrophages to metastatic sites by cancer cell-derived exosomes has been reported in melanoma (154) and pancreatic cancer (155) and implicated in metastasis. PEDF (pigment epithelium-derived factor) on the surface of exosomes from mouse and human nonmetastatic melanoma cells elicits the expansion of patrolling monocytes by NR4A1 (nuclear receptor subfamily 4 group A member 1) induction, which suppresses metastasis in the lungs of mice (156). Cancer cell-derived exosomes are also proposed to play a role in organotropic metastasis of breast and pancreatic cancers, in part through integrin expression on exosomes and organ-specific proinflammatory responses (157), and the delivery of exosomal EGFR from gastric cancer cells to Kupffer cells and hepatic stellate cells promotes liver-specific metastasis through enhanced HGF (hepatocyte growth factor) signaling in the liver (158). These results are among the growing body of evidence that support a complex exosome-mediated cell-to-cell communication in the tumor microenvironment.
A reciprocal exosome exchange from the stroma to cancer cells also modulates cancer progression and metastasis. For example, mtDNA in exosomes from CAFs induces oxidative phosphorylation (with expression of mtRNA) in breast cancer cells, promoting their survival and exit from metabolic dormancy in mice (159). Another example of stromal exosomal cargo promoting cancer cell progression includes astrocyte-derived miR-19a delivered to breast cancer cells, which results in PTEN (phosphatase and tensin homolog) suppression and contributed to metastasis (160). Fibroblast-derived exosomes also stimulate the migration of breast cancer cells by inducing Wnt-PCP (planar cell polarity) autocrine signaling (161). In addition, exosomes encapsulate metabolites, including lactate, glutamate, acetate, stearate, palmitate, and amino acids (162, 163). 13C-labeled CAF-derived exosomes fuel the tricarboxylic acid cycle of recipient cancer cells through metabolite transfer, and exosomes from prostate and pancreatic CAFs also replenish lipids in cancer cells, enhancing their fitness during tumor growth (163). Plasma-derived exosomes contain metabolic enzymes, including hexokinase 1, pyruvate kinase, lactate dehydrogenase, enolase, and glyceraldehyde 3-phospate dehydrogenase, and these enzymes mediate the production of adenosine 5′-triphosphate (ATP) in exosomes (164). CAF-derived exosomes suppress oxidative phosphorylation in prostate and pancreas cancer cells by transferring miR-22, let7a, and miR-125b, and promote glycolysis and glutamine-dependent reductive carboxylation by metabolite transfer (163). The cancer stroma-derived exosomes thus promote the metabolic fitness of cancer cells growing as tumors in mice.
Exosomes have also been implicated in the angiogenic and extracellular matrix remodeling of the tumor microenvironment, a critical step in tumor growth and metastatic dissemination. miR-105 from breast cancer cell-derived exosomes suppresses endothelial tight junction ZO-1 (zonular occludens 1) expression, resulting in increased metastasis by impairing the integrity of blood vessels and enhancing vascular permeability (165). Exosomes from hypoxic glioblastoma (GBM) cells induce proangiogenic programming of endothelial cells and GBM cell proliferation (166). Recent findings implicate neutrophil-derived exosomes in proteolytic degradation of the lung extracellular matrix associated with chronic obstructive pulmonary disease (167). In the context of cancer, MMP1 (matrix metalloprotease 1) in exosomes from ovarian cancer cells may play a role in compromising the mesothelium and promoting peritoneal dissemination of cancer cells (168).
Exosomes shed by cancer cells are reported to promote resistance to various chemotherapeutic agents and antibodies. CD20+ exosomes from B cell lymphoma act as a decoy for the binding of anti-CD20 to B cells (169), and HER2 (human epidermal growth factor receptor 2)-positive exosomes from breast cancer cells act as a decoy for anti-HER2 therapy (170), thus limiting their activity toward cancer cells. CAF-derived exosomes promote colorectal cancer chemoresistance by enhancing the growth of cancer stem cells (171) and aid in the spread of drug-resistance properties between cancer cell populations. This process may be mediated by horizontal transfer of exosomal miRNAs (observed in breast cancer cells) (172). Specifically, an exosomal long noncoding RNA (lncRNA) called lncARSR [lncRNA activated in renal cell carcinoma (RCC) with sunitinib resistance] binds competitively to miR-34 and miR-449 and enhances expression of the tyrosine kinases AXL and MET, overcoming the effect of sunitinib (173). When tumors are treated with radiation therapy or gamma secretase inhibitor, the expansion of tumor-initiating cells resistant to radiation therapy emerges through CAF-derived exosomal RNA and transposable elements transfer to cancer cells (174). CAF-derived exosomal miR-21 binding to APAF1 (apoptotic protease activating factor 1) in ovarian cancer cells confers resistance to paclitaxel (175), and macrophage-derived exosomal miR-385 induces cytidine deaminase activity in pancreatic cancer cells and confers resistance to gemcitabine (176). Chemotherapy and radiation therapy could also directly affect exosome biogenesis and the content of exosomes with potential implications on therapy outcome (177). Radiation therapy enhances exosomal miR-7–5p production by cancer cells, which induces bystander cell autophagy (178). These findings capture the distinct role of exosomes in promoting resistance to therapy, which results from exosomes directly interacting with therapeutic agents and decreasing their efficacy against cancer cells, or from exosomes (mainly from CAFs) changing the transcriptome of cancer cells to promote their survival.
Clinical applications of exosomes
The biology of exosomes in disease is still emerging, and the number of studies addressing their utility in the diagnosis and treatment of various pathologies has increased substantially. This takes advantage of the complex cargo of exosomes, allowing for a multicomponent diagnostic window into disease detection and monitoring. The characteristic properties of exosomes in delivering functional cargos to diseased cells also favor their use as therapeutic vehicles, both at the basic and applied levels.
Diagnostic potential of exosomes
Exosomes are found in all biological fluids and are secreted by all cells, rendering them attractive as minimally invasive liquid biopsies with the potential for longitudinal sampling to follow disease progression. Exosome biogenesis enables the capture of a complex extracellular and intracellular molecular cargo for comprehensive, multiparameter diagnostic testing (Fig. 2). Surface proteins on exosomes also facilitate their immune capture and enrichment. Diseases that have been the focus of diagnostic application of exosomes include CVDs (116, 179), diseases affecting the central nervous system (CNS) (180), and cancer (2, 181). This effort is rapidly expanding to other diseases involving the liver (182), kidney (183), and lung (184).
Some studies have suggested that small amounts of DNA can be found in exosomes and that this DNA can be of value in detecting cancer-associated mutations in serum exosomes (185–188). Although some studies suggest that exosomes from human cell lines and serum do not contain DNA, this remains contentious and quantitative studies are required. One study did not specify the quantity of exosomes used in its analytical assays, leading to ambitious conclusions (12). Should exosomal DNA reflect larger fragments of DNA than circulating free DNA, this may be beneficial in detecting mutations, including in KRAS and TP53, in the circulating exosomes of patients with pancreatic cancer (186–192). Specific miRNAs or groups of miRNAs in exosomes may provide diagnostic or prognostic potential in the detection of cancer (193). Oncogenic and tumor-suppressor miRNAs in exosomes may be of high diagnostic value because of their differential expression between cancer cells and normal cells, possibly enhancing their usefulness in early diagnosis (193). This may be in part driven by the genomic land-scape of cancer cells, with oncogenic Kras reported to differentially enrich for miR-100 in exosomes (194). Elevated circulating exosomal miR-21 has been associated with glioblastomas and pancreatic, colorectal, colon, liver, breast, ovarian, and esophageal cancers, and elevated urine-derived exosomal miR-21 has been associated with bladder and prostate cancer [reviewed in (193, 195)]. Other exosomal oncogenic microRNAs associated with multiple cancer types include miR-155, the miR-17–92 cluster, and miR-1246 (196–199). These are noted to be up-regulated in cancers of the brain, pancreas, colorectum, colon, liver, breast, prostate, and esophagus; in oral squamous cell cancer; as well as in lymphoma and leukemia (193). Tumor-suppressor miRNAs, including miR-146a and miR-34a, are associated with liver, breast, colon, pancreatic, and hematologic malignancies (193). The combination of multiple microRNAs may enhance the diagnostic and prognostic potential of exosomal miRNA, and exosomal miR signatures are continuously emerging in association with cancer diagnosis and prognosis (195, 200–204). The diagnostic potential of phosphoprotein in circulating exosomes from breast cancer patients has also been reported (205), as well as exosome surface protein analyses (206). Several independent laboratories have reported the utility of GPC1 (glypican 1)-positive exosomes in the diagnosis of pancreatic, breast, and colon cancer, with GPC1 being enriched in cancer cell-derived exosomes, thus enabling the detection of cancer and possibly response to therapy (decrease in exosome numbers and thus tumor burden) (207–216). Immunocapture strategies are also under investigation to detect circulating cancer exosomes using surface CD147 expression in patients with colorectal cancer (217). The relative PtdSer composition of exosomes may also prove useful for the early detection of cancer in mice, as evaluated from the serum of mice bearing breast or pancreatic tumors (218). The possibility of combining protein, lipid, RNA, and miRNA exosomal cargos in cancer diagnosis and prognostic evaluation is currently being considered. A multicomponent, combinatorial approach using a combination of markers that reflect distinct aspects of disease-generating exosomes (e.g., metabolite, RNA, and protein content) could potentially enhance the specificity and sensitivity of an exosome-based diagnostic. Such efforts would be more likely to identify collective disease-specific changes, and lipid bilayer encapsulation could preserve enzyme-sensitive molecular cargos.
Therapeutic potential of exosomes
Exosomes by themselves or as vehicles for the delivery of drug payload(s) are being actively explored as therapeutic agents (Fig. 6). In contrast to liposomes, injected exosomes are efficient at entering other cells and can deliver a functional cargo with minimal immune clearance upon exogenous administration in mice (2, 181, 219–221). In addition, the therapeutic application of exosomes is promising because they have been demonstrated to be well tolerated. Exosomes from mesenchymal cells and epithelial cells do not induce toxicity when repeatedly injected in mice (35, 57). MSC-derived exosomes have been proposed to be therapeutic by themselves (222), and the use of MSC-derived exosomes in the treatment of a patient with graft versus host disease showed that repeated injections were well tolerated, were not associated with substantial side effects, and resulted in patient response (223).
Fig. 6. Cellular uptake of therapeutic exosomes.
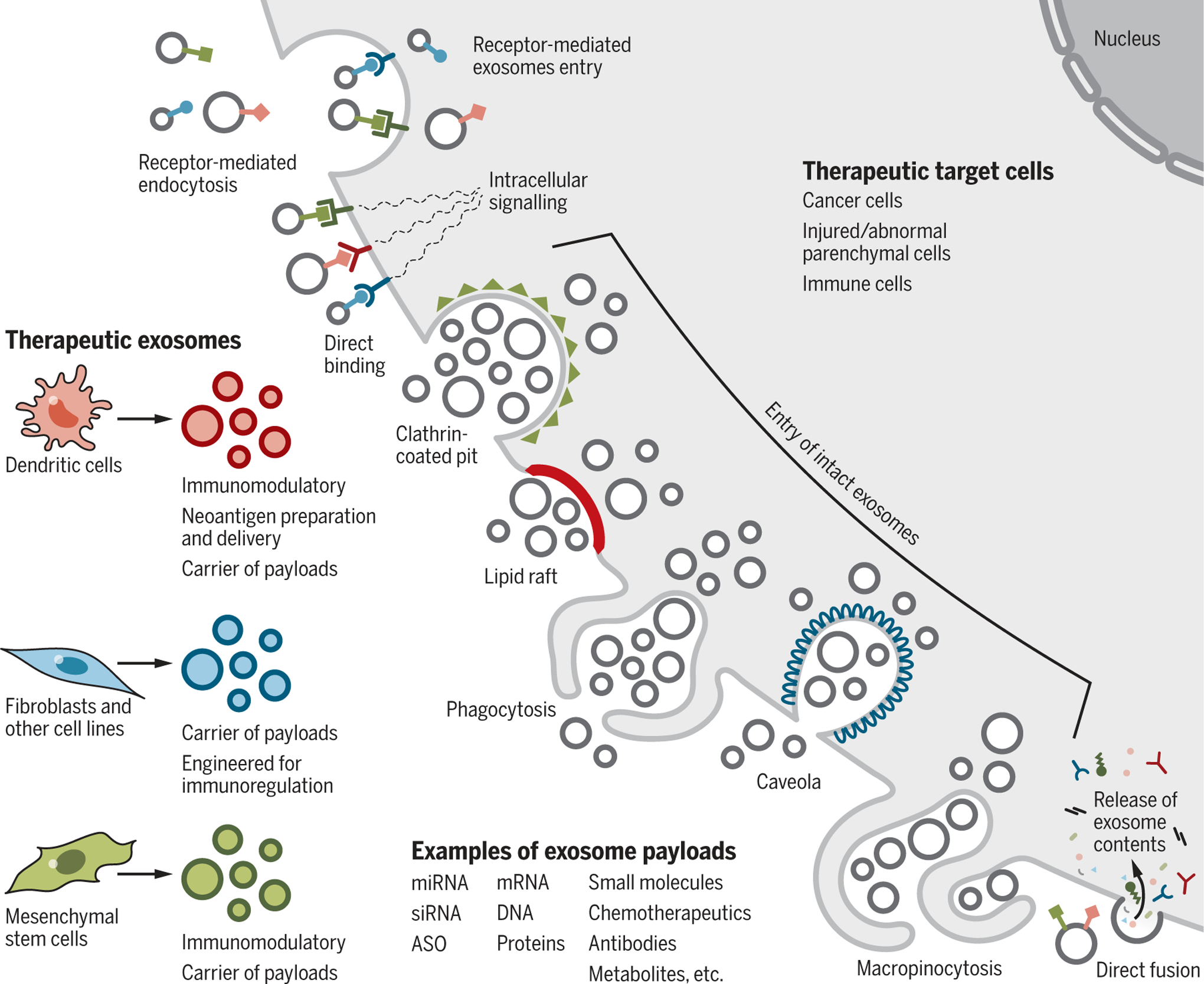
Therapeutic exosomes isolated from dendritic cells, fibroblasts, and mesenchymal cells can impart specific effects on the target cells, including neoantigen presentation, immunomodulation, and drug payload delivery. The impact of therapeutic exosomes on target cells may be controlled by the different mechanisms of entry or interaction. Entry of intact exosomes can involve receptor-mediated endocytosis, clathrin-coated pits, lipid rafts, phagocytosis, caveolae, and macropinocytosis. Entry of the content of the exosomes, or induction of signals by exosomes, can involve ligand-receptor-induced intracellular signaling or fusion to deposit the contents of the exosomes into the cytoplasm. Examples of therapeutic payload are listed. Target cells include cancer cells, injured parenchymal cells, and immune cells. ASO, antisense oligonucleotide (a DNA oligo-binding RNA target).
Enrichment of exosomes on the basis of their surface ligand presentation may also enable the development of receptor-mediated tissue targeting. Ligand enrichment on engineered exosomes may also be used to induce or inhibit signaling events in recipient cells or to target exosomes to specific cell types. For example, αv integrin-specific RGD (R, arginine; G, glycine; D, aspartic acid)-modified peptide (a modified tumor-homing peptide sequence that acts as a recognition sequence for integrins) on immature dendritic cell-derived exosomes loaded with doxorubicin showed therapeutic response in mammary tumor-bearing mice (224). Other chemotherapeutic compounds have also been loaded into exosomes for cancer therapy and tested in mice, and antitumor efficacy and reduced toxicity were reported. For example, macrophage-derived exosomes loaded with paclitaxel induced lung tumor responses in mice (225).
Building on the observation that exosomal microRNAs effectively engage target mRNA and suppress gene expression in recipient cells, engineering of exosomes to deliver a specific miRNA or small interfering RNA (siRNA) payload has been developed for CNS diseases and cancer. The exosomal RNAs may be protected from degradation by blood-derived ribonu-cleases (226) and, combined with superior systemic retention compared with liposomes, this could allow exosomes to exert their function at distant sites. Preclinical testing with the delivery of miRNA or siRNA payload using exosomes has focused on anticancer treatment in rodents with mammary carcinoma (227), glioma (228), and pancreatic cancer (28, 35), as well as exploratory brain targeting. EGFR+ breast cancer cell targeting using exosomes modified with GE11 synthetic peptide and delivery of microRNA let-7a to the cancer cells limited their growth in vivo (227). MSC-derived exosomes enabled miR-146b delivery and EGFR targeting in glioma in rats (228). Clinical-grade MSC-derived exosomes with KrasG12D siRNA payload (iExosomes) have been used to treat pancreatic cancer in multiple animal models (28, 35). These studies demonstrated that iExosomes, administered as a single agent, yield a robust increase in overall survival of mice and enable specific target engagement without any obvious toxicity (28, 35). It was shown that CD47 on exosomes results in a “don’t eat me” signal, protecting them from phagocytosis and limiting their clearance form circulation (28). Further, macropinocytosis associated with cancer cells enhanced the entry of exogenously administered iExosomes (28). Further development of iExosome-based therapy has led to a phase I clinical trial for the treatment of patients with KrasG12D mutation-associated pancreatic cancer (ClinicalTrials.gov identifier: NCT03608631).
In the context of neurological diseases, intranasal administration of human MSC-derived exosomes resulted in amelioration of autistic-like behavior in mice (BTBR mouse model), although the precise mechanisms are unknown (229). Intravenous administration of human MSC-derived exosomes supports neuroprotection, as shown by a swine model of traumatic brain injury (230). RVG (rabies virus glycoprotein)-modified dendritic cell-derived exosomes with therapeutic Bace1-targeting siRNA were intravenously administrated to mice and the results showed suppression of BACE1 expression in the brain, a potential target for the treatment of Alzheimer’s disease (142). RVG-modified exosomes with siRNA targeting α-synuclein reduced aggregate formation in the brains of S129D α-synuclein mice and improved brain pathology (143). Macrophage-derived exosomes show the capacity to effectively negotiate the blood-brain barrier and deliver protein cargo (231), supporting the idea that minimal modification of exosomes is required to reach the brain parenchyma. Macrophage-derived exosomes loaded with catalase showed therapeutic benefit (neuroprotective effect) when administered intranasally in a mouse model of PD (232). Finally, blood-derived exosomes loaded with dopamine reached the brain after intravenous injection and, compared with free dopamine, exhibited improved therapeutic efficacy with decreased toxicity in a PD mouse model (233). These findings support the potential of therapeutic cargo in exosomes reaching clinically challenging targets in the brain, in part because of engineered exosomal cargo (siRNA)-targeting genes for which there are no effective pharmacological agents, and in part because of their ability to pass through the blood-brain barrier.
The role of exosomes in polarizing the tumor immune microenvironment (discussed above) has also prompted the design of therapeutic exosomes aimed at enhancing antitumor immune responses (54). The antitumor action of exosomes from dendritic cells potentially caused by antigen presentation was tested in a clinical setting (234). The engineered exosomes, called “dexosomes,” were obtained from IFN-γ-matured dendritic cells and loaded with MART1 (melanoma antigen recognized by T cells 1) peptides. Although the approach did not yield a measurable cancer-specific T cell response, dexosomes induced increased cytolytic activity associated with natural killer cells in patients with stage IIIB/IV non-small-cell lung cancer (234). Only one among the 22 patients treated with dexosomes showed substantial liver toxicity, and 7 out of 22 patients exhibited disease stabilization exceeding 4 months (234), although the response could not be attributed specifically to the dexosomes. Together, these early clinical data and the numerous preclinical studies offer encouragement for the development of exosomes as therapeutic agents.
Conclusions
Although interesting exosome biology is being unraveled largely using cell-culture systems, there is a need for experiments using mouse models and physiologically relevant experimental conditions. Exosomes are reported to induce molecular alteration in cells but the question remains whether such observations are of relevance because of the use of supraphysiological numbers of cell culture-derived exosomes, which often also need more precise isolation and characterization procedures (1). The need for precise and accurate characterization of exosomes will continue to grow as our knowledge of the heterogeneity of EVs, their cargo, and functions evolve. Exogenous bolus doses of supraphysiological levels of exosomes into mice were associated with a penetrant cellular phenotype, including modulation of cancer progression (144, 151, 165, 235), induction of neoplasia (148), and regeneration of tissue (236). It remains unclear whether un-manipulated, physiological levels of exosomes exert regulatory homeostatic or pathological functions (or neither) in vivo. The field is in urgent need of animal models with which to study biogenesis, trafficking, and cellular entry of exosomes. Drosophila, C. elegans, Xenopus, and zebrafish models may offer additional insights (237–239).
Exosomes are generated by cells, but it is tempting to wonder whether they are reminiscent of early primordial particles that contributed to the generation of the first protocell (240, 241). It remains to be determined whether exosomes can grow and divide and, given the right environment, participate in signaling events and autonomous biochemical reactions. The similarities between exosomes and retrovirus (242) also raise the possibility that exosomes may have functioned as primordial particles that preceded single-cell organisms.
ACKNOWLEDGMENTS
The authors apologize to the investigators and research laboratories whose original studies were not cited because of space limitations.
Funding: The authors acknowledge support by research funds from MD Anderson Cancer Center. The exosome-related research in the Kalluri laboratory is funded by NCI RO1 CA213233, NCI RO1 CA195733, and NCI CA 231465.
Footnotes
Competing interests: MD Anderson Cancer Center and R.K. hold patents in the area of exosome biology and are licensed to Codiak Biosciences, Inc. MD Anderson Cancer Center and R.K. are stock equity holders in Codiak Biosciences, Inc. R.K. is a consultant and scientific adviser for Codiak Biosciences, Inc. V.S.L. is a paid consultant for Codiak Biosciences, Inc.
REFERENCES AND NOTES
- 1.Théry C et al. , Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 7, 1535750 (2018). doi: 10.1080/20013078.2018.1535750; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kalluri R, The biology and function of exosomes in cancer. J. Clin. Invest 126, 1208–1215 (2016). doi: 10.1172/JCI81135; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kahlert C, Kalluri R, Exosomes in tumor microenvironment influence cancer progression and metastasis. J. Mol. Med 91, 431–437 (2013). doi: 10.1007/s00109-013-1020-6; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.van Niel G, D’Angelo G, Raposo G, Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol 19, 213–228 (2018). doi: 10.1038/nrm.2017.125; [DOI] [PubMed] [Google Scholar]
- 5.McAndrews KM, Kalluri R, Mechanisms associated with biogenesis of exosomes in cancer. Mol. Cancer 18, 52 (2019). doi: 10.1186/s12943-019-0963-9; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Mathieu M, Martin-Jaular L, Lavieu G, Théry C, Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol 21, 9–17 (2019). doi: 10.1038/s41556-018-0250-9; [DOI] [PubMed] [Google Scholar]
- 7.Shurtleff MJ, Temoche-Diaz MM, Schekman R, Extracellular vesicles and cancer: Caveat lector. Annu. Rev. Cancer Biol 2, 395–411 (2018). doi: 10.1146/annurev-cancerbio-030617-050519 [DOI] [Google Scholar]
- 8.Harding C, Heuser J, Stahl P, Receptor-mediated endocytosis of transferrin and recycling of the transferrin receptor in rat reticulocytes. J. Cell Biol 97, 329–339 (1983). doi: 10.1083/jcb.97.2.329; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Pan BT, Teng K, Wu C, Adam M, Johnstone RM, Electron microscopic evidence for externalization of the transferrin receptor in vesicular form in sheep reticulocytes. J. Cell Biol 101, 942–948 (1985). doi: 10.1083/jcb.101.3.942; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cocucci E, Meldolesi J, Ectosomes and exosomes: Shedding the confusion between extracellular vesicles. Trends Cell Biol 25, 364–372 (2015). doi: 10.1016/j.tcb.2015.01.004; [DOI] [PubMed] [Google Scholar]
- 11.Bebelman MP, Smit MJ, Pegtel DM, Baglio SR, Biogenesis and function of extracellular vesicles in cancer. Pharmacol. Ther 188, 1–11 (2018). doi: 10.1016/j.pharmthera.2018.02.013; [DOI] [PubMed] [Google Scholar]
- 12.Jeppesen DK et al. , Reassessment of exosome composition. Cell 177, 428–445.e18 (2019). doi: 10.1016/j.cell.2019.02.029; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Willms E, Cabañas C, Mäger I, Wood MJA, Vader P, Extracellular vesicle heterogeneity: subpopulations, isolation techniques, and diverse functions in cancer progression. Front. Immunol 9, 738 (2018). doi: 10.3389/fimmu.2018.00738; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hessvik NP, Llorente A, Current knowledge on exosome biogenesis and release. Cell. Mol. Life Sci 75, 193–208 (2018). doi: 10.1007/s00018-017-2595-9; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ciardiello C et al. , Focus on extracellular vesicles: New frontiers of cell-to-cell communication in cancer. Int. J. Mol. Sci 17, 175 (2016). doi: 10.3390/ijms17020175; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chiu YJ, Cai W, Shih YR, Lian I, Lo YH, A single-cell assay for time lapse studies of exosome secretion and cell behaviors. Small 12, 3658–3666 (2016). doi: 10.1002/smll.201600725; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Zhang H et al. , Identification of distinct nanoparticles and subsets of extracellular vesicles by asymmetric flow field-flow fractionation. Nat. Cell Biol 20, 332–343 (2018). doi: 10.1038/s41556-018-0040-4; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Keerthikumar S et al. , ExoCarta: A web-based compendium of exosomal cargo. J. Mol. Biol 428, 688–692 (2016). doi: 10.1016/j.jmb.2015.09.019; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Pathan M et al. , Vesiclepedia 2019: A compendium of RNA, proteins, lipids and metabolites in extracellular vesicles. Nucleic Acids Res 47, D516–D519 (2019). doi: 10.1093/nar/gky1029; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.van Balkom BW, Eisele AS, Pegtel DM, Bervoets S, Verhaar MC, Quantitative and qualitative analysis of small RNAs in human endothelial cells and exosomes provides insights into localized RNA processing, degradation and sorting. J. Extracell. Vesicles 4, 26760 (2015). doi: 10.3402/jev.v4.26760; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lasda E, Parker R, Circular RNAs co-precipitate with extracellular vesicles: A possible mechanism for circRNA clearance. PLOS ONE 11, e0148407 (2016). doi: 10.1371/journal.pone.0148407; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Chevillet JR et al. , Quantitative and stoichiometric analysis of the microRNA content of exosomes. Proc. Natl. Acad. Sci. U.S.A 111, 14888–14893 (2014). doi: 10.1073/pnas.1408301111; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kowal J et al. , Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc. Natl. Acad. Sci. U.S.A 113, E968–E977 (2016). doi: 10.1073/pnas.1521230113; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wen SW et al. , Breast cancer-derived exosomes reflect the cell-of-origin phenotype. Proteomics 19, e1800180 (2019). doi: 10.1002/pmic.201800180; [DOI] [PubMed] [Google Scholar]
- 25.Mulcahy LA, Pink RC, Carter DR, Routes and mechanisms of extracellular vesicle uptake. J. Extracell. Vesicles 3, 24641 (2014). doi: 10.3402/jev.v3.24641; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.McKelvey KJ, Powell KL, Ashton AW, Morris JM, McCracken SA, Exosomes: Mechanisms of uptake. J Circ Biomark 4, 7 (2015). doi: 10.5772/61186; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Commisso C et al. , Macropinocytosis of protein is an amino acid supply route in Ras-transformed cells. Nature 497, 633–637 (2013). doi: 10.1038/nature12138; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kamerkar S et al. , Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer. Nature 546, 498–503 (2017). doi: 10.1038/nature22341; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Parolini I et al. , Microenvironmental pH is a key factor for exosome traffic in tumor cells. J. Biol. Chem 284, 34211–34222 (2009). doi: 10.1074/jbc.M109.041152; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Tian T et al. , Exosome uptake through clathrin-mediated endocytosis and macropinocytosis and mediating miR-21 delivery. J. Biol. Chem 289, 22258–22267 (2014). doi: 10.1074/jbc.M114.588046; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Gangadaran P, Hong CM, Ahn BC, An update on in vivo imaging of extracellular vesicles as drug delivery vehicles. Front. Pharmacol 9, 169 (2018). doi: 10.3389/fphar.2018.00169; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Gangadaran P et al. , A new bioluminescent reporter system to study the biodistribution of systematically injected tumor-derived bioluminescent extracellular vesicles in mice. Oncotarget 8, 109894–109914 (2017). doi: 10.18632/oncotarget.22493; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lai CP et al. , Dynamic biodistribution of extracellular vesicles in vivo using a multimodal imaging reporter. ACS Nano 8, 483–494 (2014). doi: 10.1021/nn404945r; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Morishita M, Takahashi Y, Nishikawa M, Takakura Y, Pharmacokinetics of exosomes: An important factor for elucidating the biological roles of exosomes and for the development of exosome-based therapeutics. J. Pharm. Sci 106, 2265–2269 (2017). doi: 10.1016/j.xphs.2017.02.030; [DOI] [PubMed] [Google Scholar]
- 35.Mendt M et al. , Generation and testing of clinical-grade exosomes for pancreatic cancer. JCI Insight 3, e99263 (2018). doi: 10.1172/jci.insight.99263; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ridder K et al. , Extracellular vesicle-mediated transfer of genetic information between the hematopoietic system and the brain in response to inflammation. PLOS Biol 12, e1001874 (2014). doi: 10.1371/journal.pbio.1001874; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Zomer A et al. , In Vivo imaging reveals extracellular vesicle-mediated phenocopying of metastatic behavior. Cell 161, 1046–1057 (2015). doi: 10.1016/j.cell.2015.04.042; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Tkach M, Théry C, Communication by extracellular vesicles: Where we are and where we need to go. Cell 164, 1226–1232 (2016). doi: 10.1016/j.cell.2016.01.043; [DOI] [PubMed] [Google Scholar]
- 39.Choi D et al. , The impact of oncogenic EGFRvIII on the proteome of extracellular vesicles released from glioblastoma cells. Mol. Cell. Proteomics 17, 1948–1964 (2018). doi: 10.1074/mcp.RA118.000644; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Cossetti C et al. , Extracellular vesicles from neural stem cells transfer IFN-γ via Ifngr1 to activate Stat1 signaling in target cells. Mol. Cell 56, 193–204 (2014). doi: 10.1016/j.molcel.2014.08.020; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Sullivan R, Saez F, Girouard J, Frenette G, Role of exosomes in sperm maturation during the transit along the male reproductive tract. Blood Cells Mol. Dis 35, 1–10 (2005). doi: 10.1016/j.bcmd.2005.03.005; [DOI] [PubMed] [Google Scholar]
- 42.Vojtech L et al. , Exosomes in human semen carry a distinctive repertoire of small non-coding RNAs with potential regulatory functions. Nucleic Acids Res 42, 7290–7304 (2014). doi: 10.1093/nar/gku347; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Madison MN, Jones PH, Okeoma CM, Exosomes in human semen restrict HIV-1 transmission by vaginal cells and block intravaginal replication of LP-BM5 murine AIDS virus complex. Virology 482, 189–201 (2015). doi: 10.1016/j.virol.2015.03.040; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Welch JL et al. , Effect of prolonged freezing of semen on exosome recovery and biologic activity. Sci. Rep 7, 45034 (2017). doi: 10.1038/srep45034; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Welch JL, Kaddour H, Schlievert PM, Stapleton JT, Okeoma CM, Semen exosomes promote transcriptional silencing of HIV-1 by disrupting NF-kB/Sp1/Tat circuitry. J. Virol 92, e00731–18 (2018). doi: 10.1128/JVI.00731-18; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Delorme-Axford E et al. , Human placental trophoblasts confer viral resistance to recipient cells. Proc. Natl. Acad. Sci. U.S.A 110, 12048–12053 (2013). doi: 10.1073/pnas.1304718110; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Menon R et al. , Circulating exosomal miRNA profile during term and preterm birth pregnancies: A longitudinal study. Endocrinology 160, 249–275 (2019). doi: 10.1210/en.2018-00836; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Menon R et al. , Quantitative proteomics by SWATH-MS of maternal plasma exosomes determine pathways associated with term and preterm birth. Endocrinology 160, 639–650 (2019). doi: 10.1210/en.2018-00820; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Sheller-Miller S, Trivedi J, Yellon SM, Menon R, Exosomes cause preterm birth in mice: Evidence for paracrine signaling in pregnancy. Sci. Rep 9, 608 (2019). doi: 10.1038/s41598-018-37002-x; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Foster BP et al. , Extracellular vesicles in blood, milk and body fluids of the female and male urogenital tract and with special regard to reproduction. Crit. Rev. Clin. Lab. Sci 53, 379–395 (2016). doi: 10.1080/10408363.2016.1190682; [DOI] [PubMed] [Google Scholar]
- 51.Admyre C et al. , Exosomes with immune modulatory features are present in human breast milk. J. Immunol 179, 1969–1978 (2007). doi: 10.4049/jimmunol.179.3.1969; [DOI] [PubMed] [Google Scholar]
- 52.Chen T et al. , Porcine milk-derived exosomes promote proliferation of intestinal epithelial cells. Sci. Rep 6, 33862 (2016). doi: 10.1038/srep33862; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Wiklander OP et al. , Extracellular vesicle in vivo biodistribution is determined by cell source, route of administration and targeting. J. Extracell. Vesicles 4, 26316 (2015). doi: 10.3402/jev.v4.26316; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Gehrmann U, Näslund TI, Hiltbrunner S, Larssen P, Gabrielsson S, Harnessing the exosome-induced immune response for cancer immunotherapy. Semin. Cancer Biol 28, 58–67 (2014). doi: 10.1016/j.semcancer.2014.05.003; [DOI] [PubMed] [Google Scholar]
- 55.Kurywchak P, Tavormina J, Kalluri R, The emerging roles of exosomes in the modulation of immune responses in cancer. Genome Med 10, 23 (2018). doi: 10.1186/s13073-018-0535-4; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Robbins PD, Morelli AE, Regulation of immune responses by extracellular vesicles. Nat. Rev. Immunol 14, 195–208 (2014). doi: 10.1038/nri3622; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Zhu X et al. , Comprehensive toxicity and immunogenicity studies reveal minimal effects in mice following sustained dosing of extracellular vesicles derived from HEK293T cells. J. Extracell. Vesicles 6, 1324730 (2017). doi: 10.1080/20013078.2017.1324730; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Raposo G et al. , B lymphocytes secrete antigen-presenting vesicles. J. Exp. Med 183, 1161–1172 (1996). doi: 10.1084/jem.183.3.1161; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Vincent-Schneider H et al. , Exosomes bearing HLA-DR1 molecules need dendritic cells to efficiently stimulate specific T cells. Int. Immunol 14, 713–722 (2002). doi: 10.1093/intimm/dxf048; [DOI] [PubMed] [Google Scholar]
- 60.Zitvogel L et al. , Eradication of established murine tumors using a novel cell-free vaccine: Dendritic cell-derived exosomes. Nat. Med 4, 594–600 (1998). doi: 10.1038/nm0598-594; [DOI] [PubMed] [Google Scholar]
- 61.Montecalvo A et al. , Exosomes as a short-range mechanism to spread alloantigen between dendritic cells during T cell allorecognition. J. Immunol 180, 3081–3090 (2008). doi: 10.4049/jimmunol.180.5.3081; [DOI] [PubMed] [Google Scholar]
- 62.Tkach M et al. , Qualitative differences in T-cell activation by dendritic cell-derived extracellular vesicle subtypes. EMBO J 36, 3012–3028 (2017). doi: 10.15252/embj.201696003; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Wahlund CJE et al. , Exosomes from antigen-pulsed dendritic cells induce stronger antigen-specific immune responses than microvesicles in vivo. Sci. Rep 7, 17095 (2017). doi: 10.1038/s41598-017-16609-6; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Cheng Y, Schorey JS, Exosomes carrying mycobacterial antigens can protect mice against Mycobacterium tuberculosis infection. Eur. J. Immunol 43, 3279–3290 (2013). doi: 10.1002/eji.201343727; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Giri PK, Schorey JS, Exosomes derived from M. bovis BCG infected macrophages activate antigen-specific CD4+ and CD8+ T cells in vitro and in vivo. PLOS ONE 3, e2461 (2008). doi: 10.1371/journal.pone.0002461; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Wang J et al. , MicroRNA-155 in exosomes secreted from Helicobacter pylori infection macrophages immunomodulates inflammatory response. Am. J. Transl. Res 8, 3700–3709 (2016). [PMC free article] [PubMed] [Google Scholar]
- 67.Ahmadi Badi S et al. , Microbiota-derived extracellular vesicles as new systemic regulators. Front. Microbiol 8, 1610 (2017). doi: 10.3389/fmicb.2017.01610; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Smythies LE, Smythies JR, Exosomes in the gut. Front. Immunol 5, 104 (2014). doi: 10.3389/fimmu.2014.00104; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Nandakumar R et al. , Intracellular bacteria engage a STING-TBK1-MVB12b pathway to enable paracrine cGAS-STING signalling. Nat. Microbiol 4, 701–713 (2019). doi: 10.1038/s41564-019-0367-z; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Cheng Y, Schorey JS, Extracellular vesicles deliver Mycobacterium RNA to promote host immunity and bacterial killing. EMBO Rep 20, e46613 (2019). doi: 10.15252/embr.201846613; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Wang J et al. , Host derived exosomes-pathogens interactions: Potential functions of exosomes in pathogen infection. Biomed. Pharmacother 108, 1451–1459 (2018). doi: 10.1016/j.biopha.2018.09.174; [DOI] [PubMed] [Google Scholar]
- 72.Marcilla A et al. , Extracellular vesicles in parasitic diseases. J. Extracell. Vesicles 3, 25040 (2014). doi: 10.3402/jev.v3.25040; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Sisquella X et al. , Malaria parasite DNA-harbouring vesicles activate cytosolic immune sensors. Nat. Commun 8, 1985 (2017). doi: 10.1038/s41467-017-02083-1; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Kitai Y et al. , DNA-containing exosomes derived from cancer cells treated with topotecan activate a STING-dependent pathway and reinforce antitumor immunity. J. Immunol 198, 1649–1659 (2017). doi: 10.4049/jimmunol.1601694; [DOI] [PubMed] [Google Scholar]
- 75.Torralba D et al. , Priming of dendritic cells by DNA-containing extracellular vesicles from activated T cells through antigen-driven contacts. Nat. Commun 9, 2658 (2018). doi: 10.1038/s41467-018-05077-9; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Takahashi A et al. , Exosomes maintain cellular homeostasis by excreting harmful DNA from cells. Nat. Commun 8, 15287 (2017). doi: 10.1038/ncomms15287; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Montermini L et al. , Inhibition of oncogenic epidermal growth factor receptor kinase triggers release of exosome-like extracellular vesicles and impacts their phosphoprotein and DNA content. J. Biol. Chem 290, 24534–24546 (2015). doi: 10.1074/jbc.M115.679217; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Chennakrishnaiah S et al. , Leukocytes as a reservoir of circulating oncogenic DNA and regulatory targets of tumor-derived extracellular vesicles. J. Thromb. Haemost 16, 1800–1813 (2018). doi: 10.1111/jth.14222; [DOI] [PubMed] [Google Scholar]
- 79.Montecalvo A et al. , Mechanism of transfer of functional microRNAs between mouse dendritic cells via exosomes. Blood 119, 756–766 (2012). doi: 10.1182/blood-2011-02-338004; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Ding G et al. , Pancreatic cancer-derived exosomes transfer miRNAs to dendritic cells and inhibit RFXAP expression via miR-212–3p. Oncotarget 6, 29877–29888 (2015). doi: 10.18632/oncotarget.4924; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Ying X et al. , Epithelial ovarian cancer-secreted exosomal miR-222–3p induces polarization of tumor-associated macrophages. Oncotarget 7, 43076–43087 (2016). doi: 10.18632/oncotarget.9246; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Chen G et al. , Exosomal PD-L1 contributes to immunosuppression and is associated with anti-PD-1 response. Nature 560, 382–386 (2018). doi: 10.1038/s41586-018-0392-8; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Ning Y et al. , Tumor exosomes block dendritic cells maturation to decrease the T cell immune response. Immunol. Lett 199, 36–43 (2018). doi: 10.1016/j.imlet.2018.05.002; [DOI] [PubMed] [Google Scholar]
- 84.Poggio M et al. , Suppression of exosomal PD-L1 induces systemic anti-tumor immunity and memory. Cell 177, 414–427. e13 (2019). doi: 10.1016/j.cell.2019.02.016; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Andreola G et al. , Induction of lymphocyte apoptosis by tumor cell secretion of FasL-bearing microvesicles. J. Exp. Med 195, 1303–1316 (2002). doi: 10.1084/jem.20011624; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Abusamra AJ et al. , Tumor exosomes expressing Fas ligand mediate CD8+ T-cell apoptosis. Blood Cells Mol. Dis 35, 169–173 (2005). doi: 10.1016/j.bcmd.2005.07.001; [DOI] [PubMed] [Google Scholar]
- 87.Clayton A, Al-Taei S, Webber J, Mason MD, Tabi Z, Cancer exosomes express CD39 and CD73, which suppress T cells through adenosine production. J. Immunol 187, 676–683 (2011). doi: 10.4049/jimmunol.1003884; [DOI] [PubMed] [Google Scholar]
- 88.Skokos D et al. , Mast cell-dependent B and T lymphocyte activation is mediated by the secretion of immunologically active exosomes. J. Immunol 166, 868–876 (2001). doi: 10.4049/jimmunol.166.2.868; [DOI] [PubMed] [Google Scholar]
- 89.Wang J, Wang L, Lin Z, Tao L, Chen M, More efficient induction of antitumor T cell immunity by exosomes from CD40L gene-modified lung tumor cells. Mol. Med. Rep 9, 125–131 (2014). doi: 10.3892/mmr.2013.1759; [DOI] [PubMed] [Google Scholar]
- 90.Capello M et al. , Exosomes harbor B cell targets in pancreatic adenocarcinoma and exert decoy function against complement-mediated cytotoxicity. Nat. Commun 10, 254 (2019). doi: 10.1038/s41467-018-08109-6; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Chalmin F et al. , Membrane-associated Hsp72 from tumor-derived exosomes mediates STAT3-dependent immunosuppressive function of mouse and human myeloid-derived suppressor cells. J. Clin. Invest 120, 457–471 (2010). doi: 10.1172/JCI40483; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Gabrusiewicz K et al. , Glioblastoma stem cell-derived exosomes induce M2 macrophages and PD-L1 expression on human monocytes. OncoImmunology 7, e1412909 (2018). doi: 10.1080/2162402X.2017.1412909; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Fabbri M et al. , MicroRNAs bind to Toll-like receptors to induce prometastatic inflammatory response. Proc. Natl. Acad. Sci. U.S.A 109, E2110–E2116 (2012). doi: 10.1073/pnas.1209414109; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.van der Grein SG, Defourny KAY, Slot EFJ, Nolte-’t Hoen ENM, Intricate relationships between naked viruses and extracellular vesicles in the crosstalk between pathogen and host. Semin. Immunopathol 40, 491–504 (2018). doi: 10.1007/s00281-018-0678-9; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Crenshaw BJ, Gu L, Sims B, Matthews QL, Exosome biogenesis and biological function in response to viral infections. Open Virol. J 12, 134–148 (2018). doi: 10.2174/1874357901812010134; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Feng Z et al. , A pathogenic picornavirus acquires an envelope by hijacking cellular membranes. Nature 496, 367–371 (2013). doi: 10.1038/nature12029; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Nagashima S et al. , Hepatitis E virus egress depends on the exosomal pathway, with secretory exosomes derived from multivesicular bodies. J. Gen. Virol 95, 2166–2175 (2014). doi: 10.1099/vir.0.066910-0; [DOI] [PubMed] [Google Scholar]
- 98.Gould SJ, Booth AM, Hildreth JE, The Trojan exosome hypothesis. Proc. Natl. Acad. Sci. U.S.A 100, 10592–10597 (2003). doi: 10.1073/pnas.1831413100; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Altan-Bonnet N, Extracellular vesicles are the Trojan horses of viral infection. Curr. Opin. Microbiol 32, 77–81 (2016). doi: 10.1016/j.mib.2016.05.004; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Ramakrishnaiah V et al. , Exosome-mediated transmission of hepatitis C virus between human hepatoma Huh7.5 cells. Proc. Natl. Acad. Sci. U.S.A 110, 13109–13113 (2013). doi: 10.1073/pnas.1221899110; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Sims B et al. , Tetraspanin blockage reduces exosome-mediated HIV-1 entry. Arch. Virol 163, 1683–1689 (2018). doi: 10.1007/s00705-018-3737-6; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Lenassi M et al. , HIV Nef is secreted in exosomes and triggers apoptosis in bystander CD4+ T cells. Traffic 11, 110–122 (2010). doi: 10.1111/j.1600-0854.2009.01006.x; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Pegtel DM et al. , Functional delivery of viral miRNAs via exosomes. Proc. Natl. Acad. Sci. U.S.A 107, 6328–6333 (2010). doi: 10.1073/pnas.0914843107; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Sims B et al. , Role of TIM-4 in exosome-dependent entry of HIV-1 into human immune cells. Int. J. Nanomedicine 12, 4823–4833 (2017). doi: 10.2147/IJN.S132762; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Gao L et al. , Tumor-derived exosomes antagonize innate antiviral immunity. Nat. Immunol 19, 233–245 (2018). doi: 10.1038/s41590-017-0043-5; [DOI] [PubMed] [Google Scholar]
- 106.Li J et al. , Exosomes mediate the cell-to-cell transmission of IFN-α-induced antiviral activity. Nat. Immunol 14, 793–803 (2013). doi: 10.1038/ni.2647; [DOI] [PubMed] [Google Scholar]
- 107.Khatua AK, Taylor HE, Hildreth JE, Popik W, Exosomes packaging APOBEC3G confer human immunodeficiency virus resistance to recipient cells. J. Virol 83, 512–521 (2009). doi: 10.1128/JVI.01658-08; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.de Carvalho JV et al. , Nef neutralizes the ability of exosomes from CD4+ T cells to act as decoys during HIV-1 infection. PLOS ONE 9, e113691 (2014). doi: 10.1371/journal.pone.0113691; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Guay C, Regazzi R, Exosomes as new players in metabolic organ cross-talk. Diabetes Obes. Metab 19 (Suppl 1), 137–146 (2017). doi: 10.1111/dom.13027; [DOI] [PubMed] [Google Scholar]
- 110.Deng ZB et al. , Adipose tissue exosome-like vesicles mediate activation of macrophage-induced insulin resistance. Diabetes 58, 2498–2505 (2009). doi: 10.2337/db09-0216; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Castaño C, Kalko S, Novials A, Párrizas M, Obesity-associated exosomal miRNAs modulate glucose and lipid metabolism in mice. Proc. Natl. Acad. Sci. U.S.A 115, 12158–12163 (2018). doi: 10.1073/pnas.1808855115; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Chitti SV, Fonseka P, Mathivanan S, Emerging role of extracellular vesicles in mediating cancer cachexia. Biochem. Soc. Trans 46, 1129–1136 (2018). doi: 10.1042/BST20180213; [DOI] [PubMed] [Google Scholar]
- 113.Sagar G et al. , Pathogenesis of pancreatic cancer exosome-induced lipolysis in adipose tissue. Gut 65, 1165–1174 (2016). doi: 10.1136/gutjnl-2014-308350; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Javeed N et al. , Pancreatic cancer-derived exosomes cause paraneoplastic β-cell dysfunction. Clin. Cancer Res 21, 1722–1733 (2015). doi: 10.1158/1078-0432.CCR-14-2022; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Zhang G et al. , Tumor induces muscle wasting in mice through releasing extracellular Hsp70 and Hsp90. Nat. Commun 8, 589 (2017). doi: 10.1038/s41467-017-00726-x; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Zhang Y, Hu YW, Zheng L, Wang Q, Characteristics and roles of exosomes in cardiovascular disease. DNA Cell Biol 36, 202–211 (2017). doi: 10.1089/dna.2016.3496; [DOI] [PubMed] [Google Scholar]
- 117.Srikanthan S, Li W, Silverstein RL, McIntyre TM, Exosome poly-ubiquitin inhibits platelet activation, downregulates CD36 and inhibits pro-atherothombotic cellular functions. J. Thromb. Haemost 12, 1906–1917 (2014). doi: 10.1111/jth.12712; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Kapustin AN et al. , Prothrombin loading of vascular smooth muscle cell-derived exosomes regulates coagulation and calcification. Arterioscler. Thromb. Vasc. Biol 37, e22–e32 (2017). doi: 10.1161/ATVBAHA.116.308886; [DOI] [PubMed] [Google Scholar]
- 119.Yuan Y et al. , Stem cell-derived exosome in cardiovascular diseases: Macro roles of micro particles. Front. Pharmacol 9, 547 (2018). doi: 10.3389/fphar.2018.00547; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Xiao J et al. , Cardiac progenitor cell-derived exosomes prevent cardiomyocytes apoptosis through exosomal miR-21 by targeting PDCD4. Cell Death Dis 7, e2277 (2016). doi: 10.1038/cddis.2016.181; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Feng Y, Huang W, Wani M, Yu X, Ashraf M, Ischemic preconditioning potentiates the protective effect of stem cells through secretion of exosomes by targeting Mecp2 via miR-22. PLOS ONE 9, e88685 (2014). doi: 10.1371/journal.pone.0088685; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Mayourian J et al. , Exosomal microRNA-21–5p mediates mesenchymal stem cell paracrine effects on human cardiac tissue contractility. Circ. Res 122, 933–944 (2018). doi: 10.1161/CIRCRESAHA.118.312420; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Budnik V, Ruiz-Cañada C, Wendler F, Extracellular vesicles round off communication in the nervous system. Nat. Rev. Neurosci 17, 160–172 (2016). doi: 10.1038/nrn.2015.29; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Quek C, Hill AF, The role of extracellular vesicles in neurodegenerative diseases. Biochem. Biophys. Res. Commun 483, 1178–1186 (2017). doi: 10.1016/j.bbrc.2016.09.090; [DOI] [PubMed] [Google Scholar]
- 125.Yuan L, Li JY, Exosomes in Parkinson’s disease: Current perspectives and future challenges. ACS Chem. Neurosci 10, 964–972 (2019). doi: 10.1021/acschemneuro.8b00469; [DOI] [PubMed] [Google Scholar]
- 126.Howitt J, Hill AF, Exosomes in the pathology of neurodegenerative diseases. J. Biol. Chem 291, 26589–26597 (2016). doi: 10.1074/jbc.R116.757955; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Soria FN et al. , Exosomes, an unmasked culprit in neurodegenerative diseases. Front. Neurosci 11, 26 (2017). doi: 10.3389/fnins.2017.00026; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Levy E, Exosomes in the diseased brain: First insights from in vivo studies. Front. Neurosci 11, 142 (2017). doi: 10.3389/fnins.2017.00142; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Guo BB, Bellingham SA, Hill AF, Stimulating the release of exosomes increases the intercellular transfer of prions. J. Biol. Chem 291, 5128–5137 (2016). doi: 10.1074/jbc.M115.684258; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Asai H et al. , Depletion of microglia and inhibition of exosome synthesis halt tau propagation. Nat. Neurosci 18, 1584–1593 (2015). doi: 10.1038/nn.4132; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Baker S, Polanco JC, Götz J, Extracellular vesicles containing P301L mutant tau accelerate pathological tau phosphorylation and oligomer formation but do not seed mature neurofibrillary tangles in ALZ17 mice. J. Alzheimers Dis 54, 1207–1217 (2016). doi: 10.3233/JAD-160371; [DOI] [PubMed] [Google Scholar]
- 132.Rajendran L et al. , Alzheimer’s disease beta-amyloid peptides are released in association with exosomes. Proc. Natl. Acad. Sci. U.S.A 103, 11172–11177 (2006). doi: 10.1073/pnas.0603838103; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Falker C et al. , Exosomal cellular prion protein drives fibrillization of amyloid beta and counteracts amyloid beta-mediated neurotoxicity. J. Neurochem 137, 88–100 (2016). doi: 10.1111/jnc.13514; [DOI] [PubMed] [Google Scholar]
- 134.Yuyama K et al. , A potential function for neuronal exosomes: Sequestering intracerebral amyloid-β peptide. FEBS Lett 589, 84–88 (2015). doi: 10.1016/j.febslet.2014.11.027; [DOI] [PubMed] [Google Scholar]
- 135.Pacheco-Quinto J et al. , Intracellular metalloprotease activity controls intraneuronal Abeta aggregation and limits secretion of Abeta via exosomes. FASEB J 33, 3758–3771 (2019). doi: 10.1096/fj.201801319R; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Stuendl A et al. , Induction of α-synuclein aggregate formation by CSF exosomes from patients with Parkinson’s disease and dementia with Lewy bodies. Brain 139, 481–494 (2016). doi: 10.1093/brain/awv346; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Spencer B et al. , α-Synuclein interferes with the ESCRT-III complex contributing to the pathogenesis of Lewy body disease. Hum. Mol. Genet 25, 1100–1115 (2016). doi: 10.1093/hmg/ddv633; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Basso M et al. , Mutant copper-zinc superoxide dismutase (SOD1) induces protein secretion pathway alterations and exosome release in astrocytes: Implications for disease spreading and motor neuron pathology in amyotrophic lateral sclerosis. J. Biol. Chem 288, 15699–15711 (2013). doi: 10.1074/jbc.M112.425066; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Grad LI et al. , Intercellular propagated misfolding of wild-type Cu/Zn superoxide dismutase occurs via exosome-dependent and -independent mechanisms. Proc. Natl. Acad. Sci. U.S.A 111, 3620–3625 (2014). doi: 10.1073/pnas.1312245111; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Iguchi Y et al. , Exosome secretion is a key pathway for clearance of pathological TDP-43. Brain 139, 3187–3201 (2016). doi: 10.1093/brain/aww237; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Tsilioni I, Theoharides TC, Extracellular vesicles are increased in the serum of children with autism spectrum disorder, contain mitochondrial DNA, and stimulate human microglia to secrete IL-1β. J. Neuroinflammation 15, 239 (2018). doi: 10.1186/s12974-018-1275-5; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Alvarez-Erviti L et al. , Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat. Biotechnol 29, 341–345 (2011). doi: 10.1038/nbt.1807; [DOI] [PubMed] [Google Scholar]
- 143.Cooper JM et al. , Systemic exosomal siRNA delivery reduced alpha-synuclein aggregates in brains of transgenic mice. Mov. Disord 29, 1476–1485 (2014). doi: 10.1002/mds.25978; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Skog J et al. , Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat. Cell Biol 10, 1470–1476 (2008). doi: 10.1038/ncb1800; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Rajagopal C, Harikumar KB, The origin and functions of exosomes in cancer. Front. Oncol 8, 66 (2018). doi: 10.3389/fonc.2018.00066; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Hanahan D, Weinberg RA, Hallmarks of cancer: The next generation. Cell 144, 646–674 (2011). doi: 10.1016/j.cell.2011.02.013; [DOI] [PubMed] [Google Scholar]
- 147.Stefanius K et al. , Human pancreatic cancer cell exosomes, but not human normal cell exosomes, act as an initiator in cell transformation. eLife 8, e40226 (2019). doi: 10.7554/eLife.40226; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Melo SA et al. , Cancer exosomes perform cell-independent microRNA biogenesis and promote tumorigenesis. Cancer Cell 26, 707–721 (2014). doi: 10.1016/j.ccell.2014.09.005; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Abd Elmageed ZY et al. , Neoplastic reprogramming of patient-derived adipose stem cells by prostate cancer cell-associated exosomes. Stem Cells 32, 983–997 (2014). doi: 10.1002/stem.1619; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Le MT et al. , miR-200-containing extracellular vesicles promote breast cancer cell metastasis. J. Clin. Invest 124, 5109–5128 (2014). doi: 10.1172/JCI75695; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Fong MY et al. , Breast-cancer-secreted miR-122 reprograms glucose metabolism in premetastatic niche to promote metastasis. Nat. Cell Biol 17, 183–194 (2015). doi: 10.1038/ncb3094; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Nabet BY et al. , Exosome RNA unshielding couples stromal activation to pattern recognition receptor signaling in cancer. Cell 170, 352–366.e13 (2017). doi: 10.1016/j.cell.2017.06.031; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Webber J, Steadman R, Mason MD, Tabi Z, Clayton A, Cancer exosomes trigger fibroblast to myofibroblast differentiation. Cancer Res 70, 9621–9630 (2010). doi: 10.1158/0008-5472.CAN-10-1722; [DOI] [PubMed] [Google Scholar]
- 154.Peinado H et al. , Melanoma exosomes educate bone marrow progenitor cells toward a pro-metastatic phenotype through MET. Nat. Med 18, 883–891 (2012). doi: 10.1038/nm.2753; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Costa-Silva B et al. , Pancreatic cancer exosomes initiate pre-metastatic niche formation in the liver. Nat. Cell Biol 17, 816–826 (2015). doi: 10.1038/ncb3169; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Plebanek MP et al. , Pre-metastatic cancer exosomes induce immune surveillance by patrolling monocytes at the metastatic niche. Nat. Commun 8, 1319 (2017). doi: 10.1038/s41467-017-01433-3; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Hoshino A et al. , Tumour exosome integrins determine organotropic metastasis. Nature 527, 329–335 (2015). doi: 10.1038/nature15756; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Zhang H et al. , Exosome-delivered EGFR regulates liver microenvironment to promote gastric cancer liver metastasis. Nat. Commun 8, 15016 (2017). doi: 10.1038/ncomms15016; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Sansone P et al. , Packaging and transfer of mitochondrial DNA via exosomes regulate escape from dormancy in hormonal therapy-resistant breast cancer. Proc. Natl. Acad. Sci. U.S.A 114, E9066–E9075 (2017). doi: 10.1073/pnas.1704862114; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Zhang L et al. , Microenvironment-induced PTEN loss by exosomal microRNA primes brain metastasis outgrowth. Nature 527, 100–104 (2015). doi: 10.1038/nature15376; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Luga V et al. , Exosomes mediate stromal mobilization of autocrine Wnt-PCP signaling in breast cancer cell migration. Cell 151, 1542–1556 (2012). doi: 10.1016/j.cell.2012.11.024; [DOI] [PubMed] [Google Scholar]
- 162.Vallabhaneni KC et al. , Extracellular vesicles from bone marrow mesenchymal stem/stromal cells transport tumor regulatory microRNA, proteins, and metabolites. Oncotarget 6, 4953–4967 (2015). doi: 10.18632/oncotarget.3211; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Zhao H et al. , Tumor microenvironment derived exosomes pleiotropically modulate cancer cell metabolism. eLife 5, e10250 (2016). doi: 10.7554/eLife.10250; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Ronquist KG, Ek B, Stavreus-Evers A, Larsson A, Ronquist G, Human prostasomes express glycolytic enzymes with capacity for ATP production. Am. J. Physiol. Endocrinol. Metab 304, E576–E582 (2013). doi: 10.1152/ajpendo.00511.2012; [DOI] [PubMed] [Google Scholar]
- 165.Zhou W et al. , Cancer-secreted miR-105 destroys vascular endothelial barriers to promote metastasis. Cancer Cell 25, 501–515 (2014). doi: 10.1016/j.ccr.2014.03.007; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Kucharzewska P et al. , Exosomes reflect the hypoxic status of glioma cells and mediate hypoxia-dependent activation of vascular cells during tumor development. Proc. Natl. Acad. Sci. U.S.A 110, 7312–7317 (2013). doi: 10.1073/pnas.1220998110; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Genschmer KR et al. , Activated PMN exosomes: Pathogenic entities causing matrix destruction and disease in the lung. Cell 176, 113–126.e15 (2019). doi: 10.1016/j.cell.2018.12.002; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Yokoi A et al. , Malignant extracellular vesicles carrying MMP1 mRNA facilitate peritoneal dissemination in ovarian cancer. Nat. Commun 8, 14470 (2017). doi: 10.1038/ncomms14470; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Aung T et al. , Exosomal evasion of humoral immunotherapy in aggressive B-cell lymphoma modulated by ATP-binding cassette transporter A3. Proc. Natl. Acad. Sci. U.S.A 108, 15336–15341 (2011). doi: 10.1073/pnas.1102855108; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Ciravolo V et al. , Potential role of HER2-overexpressing exosomes in countering trastuzumab-based therapy. J. Cell. Physiol 227, 658–667 (2012). doi: 10.1002/jcp.22773; [DOI] [PubMed] [Google Scholar]
- 171.Hu Y et al. , Fibroblast-derived exosomes contribute to chemoresistance through priming cancer stem cells in colorectal cancer. PLOS ONE 10, e0125625 (2015). doi: 10.1371/journal.pone.0125625; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Chen WX et al. , Exosomes from drug-resistant breast cancer cells transmit chemoresistance by a horizontal transfer of microRNAs. PLOS ONE 9, e95240 (2014). doi: 10.1371/journal.pone.0095240; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Qu L et al. , Exosome-transmitted lncARSR promotes sunitinib resistance in renal cancer by acting as a competing endogenous RNA. Cancer Cell 29, 653–668 (2016). doi: 10.1016/j.ccell.2016.03.004; [DOI] [PubMed] [Google Scholar]
- 174.Boelens MC et al. , Exosome transfer from stromal to breast cancer cells regulates therapy resistance pathways. Cell 159, 499–513 (2014). doi: 10.1016/j.cell.2014.09.051; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Au Yeung CL et al. , Exosomal transfer of stroma-derived miR21 confers paclitaxel resistance in ovarian cancer cells through targeting APAF1. Nat. Commun 7, 11150 (2016). doi: 10.1038/ncomms11150; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Binenbaum Y et al. , Transfer of miRNA in macrophage-derived exosomes induces drug resistance in pancreatic adenocarcinoma. Cancer Res 78, 5287–5299 (2018). doi: 10.1158/0008-5472.CAN-18-0124; [DOI] [PubMed] [Google Scholar]
- 177.Cesselli D et al. , Extracellular vesicles: How drug and pathology interfere with their biogenesis and function. Front. Physiol 9, 1394 (2018). doi: 10.3389/fphys.2018.01394; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Song M et al. , Bystander autophagy mediated by radiation-induced exosomal miR-7–5p in non-targeted human bronchial epithelial cells. Sci. Rep 6, 30165 (2016). doi: 10.1038/srep30165; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Jansen F, Li Q, Exosomes as diagnostic biomarkers in cardiovascular diseases. Adv. Exp. Med. Biol 998, 61–70 (2017). doi: 10.1007/978-981-10-4397-0_4; [DOI] [PubMed] [Google Scholar]
- 180.Kanninen KM, Bister N, Koistinaho J, Malm T, Exosomes as new diagnostic tools in CNS diseases. Biochim. Biophys. Acta 1862, 403–410 (2016). doi: 10.1016/j.bbadis.2015.09.020; [DOI] [PubMed] [Google Scholar]
- 181.Fitts CA, Ji N, Li Y, Tan C, Exploiting exosomes in cancer liquid biopsies and drug delivery. Adv. Healthc. Mater 8, e1801268 (2019). doi: 10.1002/adhm.201801268; [DOI] [PubMed] [Google Scholar]
- 182.Masyuk AI, Masyuk TV, Larusso NF, Exosomes in the pathogenesis, diagnostics and therapeutics of liver diseases. J. Hepatol 59, 621–625 (2013). doi: 10.1016/j.jhep.2013.03.028; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Zhang W et al. , Extracellular vesicles in diagnosis and therapy of kidney diseases. Am. J. Physiol. Renal Physiol 311, F844–F851 (2016). doi: 10.1152/ajprenal.00429.2016; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Alipoor SD et al. , Exosomes and exosomal miRNA in respiratory diseases. Mediators Inflamm 2016, 5628404 (2016). doi: 10.1155/2016/5628404; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Cai J et al. , Extracellular vesicle-mediated transfer of donor genomic DNA to recipient cells is a novel mechanism for genetic influence between cells. J. Mol. Cell Biol 5, 227–238 (2013). doi: 10.1093/jmcb/mjt011; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Kahlert C et al. , Identification of double-stranded genomic DNA spanning all chromosomes with mutated KRAS and p53 DNA in the serum exosomes of patients with pancreatic cancer. J. Biol. Chem 289, 3869–3875 (2014). doi: 10.1074/jbc.C113.532267; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Kalluri R, LeBleu VS, Discovery of double-stranded genomic DNA in circulating exosomes. Cold Spring Harb. Symp. Quant. Biol 81, 275–280 (2016). doi: 10.1101/sqb.2016.81.030932; [DOI] [PubMed] [Google Scholar]
- 188.Thakur BK et al. , Double-stranded DNA in exosomes: A novel biomarker in cancer detection. Cell Res 24, 766–769 (2014). doi: 10.1038/cr.2014.44; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Allenson K et al. , High prevalence of mutant KRAS in circulating exosome-derived DNA from early-stage pancreatic cancer patients. Ann. Oncol 28, 741–747 (2017). doi: 10.1093/annonc/mdx004; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Bernard V et al. , Circulating nucleic acids are associated with outcomes of patients with pancreatic cancer. Gastroenterology 156, 108–118.e4 (2019). doi: 10.1053/j.gastro.2018.09.022; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Fernando MR, Jiang C, Krzyzanowski GD, Ryan WL, New evidence that a large proportion of human blood plasma cell-free DNA is localized in exosomes. PLOS ONE 12, e0183915 (2017). doi: 10.1371/journal.pone.0183915; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Yang S et al. , Detection of mutant KRAS and TP53 DNA in circulating exosomes from healthy individuals and patients with pancreatic cancer. Cancer Biol. Ther 18, 158–165 (2017). doi: 10.1080/15384047.2017.1281499; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Salehi M, Sharifi M, Exosomal miRNAs as novel cancer biomarkers: Challenges and opportunities. J. Cell. Physiol 233, 6370–6380 (2018). doi: 10.1002/jcp.26481; [DOI] [PubMed] [Google Scholar]
- 194.Cha DJ et al. , KRAS-dependent sorting of miRNA to exosomes. eLife 4, e07197 (2015). doi: 10.7554/eLife.07197; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Thind A, Wilson C, Exosomal miRNAs as cancer biomarkers and therapeutic targets. J. Extracell. Vesicles 5, 31292 (2016). doi: 10.3402/jev.v5.31292; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Bhagirath D et al. , microRNA-1246 is an exosomal biomarker for aggressive prostate cancer. Cancer Res 78, 1833–1844 (2018). doi: 10.1158/0008-5472.CAN-17-2069; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Li XJ, Ren ZJ, Tang JH, Yu Q, Exosomal Micro RNA, Exosomal microRNA MiR-1246 promotes cell proliferation, invasion and drug resistance by targeting CCNG2 in breast cancer. Cell. Physiol. Biochem 44, 1741–1748 (2017). doi: 10.1159/000485780; [DOI] [PubMed] [Google Scholar]
- 198.Pigati L et al. , Selective release of microRNA species from normal and malignant mammary epithelial cells. PLOS ONE 5, e13515 (2010). doi: 10.1371/journal.pone.0013515; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Sakha S, Muramatsu T, Ueda K, Inazawa J, Exosomal microRNA miR-1246 induces cell motility and invasion through the regulation of DENND2D in oral squamous cell carcinoma. Sci. Rep 6, 38750 (2016). doi: 10.1038/srep38750; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Alhasan AH et al. , Circulating microRNA signature for the diagnosis of very high-risk prostate cancer. Proc. Natl. Acad. Sci. U.S.A 113, 10655–10660 (2016). doi: 10.1073/pnas.1611596113; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Ebrahimkhani S et al. , Deep sequencing of circulating exosomal microRNA allows non-invasive glioblastoma diagnosis. NPJ Precis Oncol 2, 28 (2018). doi: 10.1038/s41698-018-0071-0; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Halvaei S et al. , Exosomes in Cancer Liquid Biopsy: A Focus on Breast Cancer. Mol. Ther. Nucleic Acids 10, 131–141 (2018). doi: 10.1016/j.omtn.2017.11.014; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Stevic I et al. , Specific microRNA signatures in exosomes of triple-negative and HER2-positive breast cancer patients undergoing neoadjuvant therapy within the GeparSixto trial. BMC Med 16, 179 (2018). doi: 10.1186/s12916-018-1163-y; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Zhou X et al. , Diagnostic value of a plasma microRNA signature in gastric cancer: A microRNA expression analysis. Sci. Rep 5, 11251 (2015). doi: 10.1038/srep11251; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Chen IH et al. , Phosphoproteins in extracellular vesicles as candidate markers for breast cancer. Proc. Natl. Acad. Sci. U.S.A 114, 3175–3180 (2017). doi: 10.1073/pnas.1618088114; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Castillo J et al. , Surfaceome profiling enables isolation of cancer-specific exosomal cargo in liquid biopsies from pancreatic cancer patients. Ann. Oncol 29, 223–229 (2018). doi: 10.1093/annonc/mdx542; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Etayash H, McGee AR, Kaur K, Thundat T, Nanomechanical sandwich assay for multiple cancer biomarkers in breast cancer cell-derived exosomes. Nanoscale 8, 15137–15141 (2016). doi: 10.1039/C6NR03478K; [DOI] [PubMed] [Google Scholar]
- 208.Frampton AE et al. , Glypican-1 is enriched in circulating-exosomes in pancreatic cancer and correlates with tumor burden. Oncotarget 9, 19006–19013 (2018). doi: 10.18632/oncotarget.24873; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Hu J et al. , A signal-amplifiable biochip quantifies extracellular vesicle-associated RNAs for early cancer detection. Nat. Commun 8, 1683 (2017). doi: 10.1038/s41467-017-01942-1; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Lai X et al. , A microRNA signature in circulating exosomes is superior to exosomal glypican-1 levels for diagnosing pancreatic cancer. Cancer Lett 393, 86–93 (2017). doi: 10.1016/j.canlet.2017.02.019; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Lewis JM et al. , Integrated analysis of exosomal protein biomarkers on alternating current electrokinetic chips enables rapid detection of pancreatic cancer in patient blood. ACS Nano 12, 3311–3320 (2018). doi: 10.1021/acsnano.7b08199; [DOI] [PubMed] [Google Scholar]
- 212.Li J et al. , GPC1 exosome and its regulatory miRNAs are specific markers for the detection and target therapy of colorectal cancer. J. Cell. Mol. Med 21, 838–847 (2017). doi: 10.1111/jcmm.12941; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Li J et al. , The clinical significance of circulating GPC1 positive exosomes and its regulative miRNAs in colon cancer patients. Oncotarget 8, 101189–101202 (2017). doi: 10.18632/oncotarget.20516; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Melo SA et al. , Glypican-1 identifies cancer exosomes and detects early pancreatic cancer. Nature 523, 177–182 (2015). doi: 10.1038/nature14581; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Qian JY, Tan YL, Zhang Y, Yang YF, Li XQ, Prognostic value of glypican-1 for patients with advanced pancreatic cancer following regional intra-arterial chemotherapy. Oncol. Lett 16, 1253–1258 (2018). doi: 10.3892/ol.2018.8701; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Yang KS et al. , Multiparametric plasma EV profiling facilitates diagnosis of pancreatic malignancy. Sci. Transl. Med 9, eaal3226 (2017). doi: 10.1126/scitranslmed.aal3226; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Yoshioka Y et al. , Ultra-sensitive liquid biopsy of circulating extracellular vesicles using ExoScreen. Nat. Commun 5, 3591 (2014). doi: 10.1038/ncomms4591; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Sharma R, Huang X, Brekken RA, Schroit AJ, Detection of phosphatidylserine-positive exosomes for the diagnosis of early-stage malignancies. Br. J. Cancer 117, 545–552 (2017). doi: 10.1038/bjc.2017.183; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Barile L, Vassalli G, Exosomes: Therapy delivery tools and biomarkers of diseases. Pharmacol. Ther 174, 63–78 (2017). doi: 10.1016/j.pharmthera.2017.02.020; [DOI] [PubMed] [Google Scholar]
- 220.Ferguson SW, Nguyen J, Exosomes as therapeutics: The implications of molecular composition and exosomal heterogeneity. J. Control. Release 228, 179–190 (2016). doi: 10.1016/j.jconrel.2016.02.037; [DOI] [PubMed] [Google Scholar]
- 221.Liao W et al. , Exosomes: The next generation of endogenous nanomaterials for advanced drug delivery and therapy. Acta Biomater (2018). [DOI] [PubMed] [Google Scholar]
- 222.Yeo RW et al. , Mesenchymal stem cell: An efficient mass producer of exosomes for drug delivery. Adv. Drug Deliv. Rev 65, 336–341 (2013). doi: 10.1016/j.addr.2012.07.001; [DOI] [PubMed] [Google Scholar]
- 223.Kordelas L et al. , MSC-derived exosomes: A novel tool to treat therapy-refractory graft-versus-host disease. Leukemia 28, 970–973 (2014). doi: 10.1038/leu.2014.41; [DOI] [PubMed] [Google Scholar]
- 224.Tian Y et al. , A doxorubicin delivery platform using engineered natural membrane vesicle exosomes for targeted tumor therapy. Biomaterials 35, 2383–2390 (2014). doi: 10.1016/j.biomaterials.2013.11.083; [DOI] [PubMed] [Google Scholar]
- 225.Kim MS et al. , Development of exosome-encapsulated paclitaxel to overcome MDR in cancer cells. Nanomedicine (Lond.) 12, 655–664 (2016). doi: 10.1016/j.nano.2015.10.012; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Cheng L, Sharples RA, Scicluna BJ, Hill AF, Exosomes provide a protective and enriched source of miRNA for biomarker profiling compared to intracellular and cell-free blood. J. Extracell. Vesicles 3, 23743 (2014). doi: 10.3402/jev.v3.23743; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Ohno S et al. , Systemically injected exosomes targeted to EGFR deliver antitumor microRNA to breast cancer cells. Mol. Ther 21, 185–191 (2013). doi: 10.1038/mt.2012.180; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Katakowski M et al. , Exosomes from marrow stromal cells expressing miR-146b inhibit glioma growth. Cancer Lett 335, 201–204 (2013). doi: 10.1016/j.canlet.2013.02.019; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229.Perets N, Hertz S, London M, Offen D, Intranasal administration of exosomes derived from mesenchymal stem cells ameliorates autistic-like behaviors of BTBR mice. Mol. Autism 9, 57 (2018). doi: 10.1186/s13229-018-0240-6; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Williams AM et al. , Mesenchymal stem cell-derived exosomes provide neuroprotection and improve long-term neurologic outcomes in a swine model of traumatic brain injury and hemorrhagic shock. J. Neurotrauma 36, 54–60 (2019). doi: 10.1089/neu.2018.5711; [DOI] [PubMed] [Google Scholar]
- 231.Yuan D et al. , Macrophage exosomes as natural nanocarriers for protein delivery to inflamed brain. Biomaterials 142, 1–12 (2017). doi: 10.1016/j.biomaterials.2017.07.011; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Haney MJ et al. , Exosomes as drug delivery vehicles for Parkinson’s disease therapy. J. Control. Release 207, 18–30 (2015). doi: 10.1016/j.jconrel.2015.03.033; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Qu M et al. , Dopamine-loaded blood exosomes targeted to brain for better treatment of Parkinson’s disease. J. Control. Release 287, 156–166 (2018). doi: 10.1016/j.jconrel.2018.08.035; [DOI] [PubMed] [Google Scholar]
- 234.Besse B et al. , Dendritic cell-derived exosomes as maintenance immunotherapy after first line chemotherapy in NSCLC. OncoImmunology 5, e1071008 (2015). doi: 10.1080/2162402X.2015.1071008; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Liu Y et al. , Tumor exosomal RNAs promote lung pre-metastatic niche formation by activating alveolar epithelial TLR3 to recruit neutrophils. Cancer Cell 30, 243–256 (2016). doi: 10.1016/j.ccell.2016.06.021; [DOI] [PubMed] [Google Scholar]
- 236.Mao G et al. , Exosomes derived from miR-92a-3p-overexpressing human mesenchymal stem cells enhance chondrogenesis and suppress cartilage degradation via targeting WNT5A. Stem Cell Res. Ther 9, 247 (2018). doi: 10.1186/s13287-018-1004-0; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Beer KB, Wehman AM, Mechanisms and functions of extracellular vesicle release in vivo: What we can learn from flies and worms. Cell Adhes. Migr 11, 135–150 (2017). doi: 10.1080/19336918.2016.1236899; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Danilchik M, Tumarkin T, Exosomal trafficking in Xenopus development. Genesis 55, e23011 (2017). doi: 10.1002/dvg.23011; [DOI] [PubMed] [Google Scholar]
- 239.Verweij FJ et al. , Live tracking of inter-organ communication by endogenous exosomes in vivo. Dev. Cell 48, 573–589.e4 (2019). doi: 10.1016/j.devcel.2019.01.004; [DOI] [PubMed] [Google Scholar]
- 240.Budin I, Debnath A, Szostak JW, Concentration-driven growth of model protocell membranes. J. Am. Chem. Soc 134, 20812–20819 (2012). doi: 10.1021/ja310382d; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Joyce GF, Szostak JW, Protocells and RNA self-replication. Cold Spring Harb. Perspect. Biol 10, a034801 (2018). doi: 10.1101/cshperspect.a034801; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.Nolte-’t Hoen E, Cremer T, Gallo RC, Margolis LB, Extracellular vesicles and viruses: Are they close relatives? Proc. Natl. Acad. Sci. U.S.A 113, 9155–9161 (2016). doi: 10.1073/pnas.1605146113; [DOI] [PMC free article] [PubMed] [Google Scholar]