

Abstract
SRY-box transcription factor 2 (SOX2) overlapping transcript (SOX2-OT) is an evolutionarily conserved long noncoding RNA. Its intronic region contains the SOX2 gene, the major regulator of the pluripotency of embryonic stem cells. The human SOX2-OT gene comprises multiple exons and has multiple transcription start sites and generates hundreds of transcripts. Transcription factors (IRF4, AR, and SOX3), transcriptional inhibitors (NSPc1, MTA3, and YY1), and miRNAs (miR-211 and miR-375) have been demonstrated to control certain SOX2-OT transcript level at the transcriptional or posttranscriptional levels. Accumulated evidence indicates its crucial roles in the regulation of the SOX2 gene, miRNAs, and transcriptional process. Restricted expression of SOX2-OT transcripts in the brain results in the association between SOX2-OT single nucleotide polymorphisms and mental illnesses such as schizophrenia and anorexia nervosa. SOX2-OT is notably elevated in tumor tissues, and a high level of SOX2-OT is well correlated with poor clinical outcomes in cancer patients, leading to the establishment of its role as an oncogene and a prognostic or diagnostic biomarker for cancers. The emerging evidence supports that SOX2-OT mediates diabetic complications. In summary, SOX2-OT has diversified functions and could be a therapeutic target for various diseases.
1. Introductions
SRY-box transcription factor 2 (SOX2) overlapping transcript (official symbol SOX2-OT according to the HUGO Gene Nomenclature Committee) is an evolutionarily conserved long noncoding RNA (lncRNA). The SOX2-OT gene is mapped to human chromosomal locus 3q26.33 and is located in a highly conserved region of more than 750 kb in humans and other vertebrates [1]. The SOX2-OT gene contains the key regulator of embryonic stem cell pluripotency, i.e., the SOX2 gene, within its intronic region, and both SOX2-OT and SOX2 are transcribed in the same orientation [2]. The human SOX2-OT gene comprises multiple exons and has multiple transcription start sites with complicated transcriptional features [1, 2]. Initially, Amaral et al. identified several variants of the SOX2-OT gene in mice and humans, including transcripts with multiple transcription start sites [2]. This group also identified SOX2-OT variants in chickens, frogs, and zebrafishes, and some transcripts appear to be species-specific [2]. As deep DNA sequencing technology has advanced, researchers have found that the SOX2-OT gene is expressed as 104 mRNA-like transcripts, the longest of which is approximately 4.3 kb in humans (according to the Ensembl genome database project) [3]. The comprehensive noncoding RNA sequence database RNA Central, which is maintained by the European Molecular Biology Laboratory-European Bioinformatics Institute (EMBL-EBI), includes information for 161 transcripts of the human SOX2-OT gene [4]. In mice, the SOX2 overlapping transcript (official symbol Sox2ot according to Mouse Genome Informatics) is mapped to chromosome 3qA3, and the transcript length is shorter than that of the human homolog. The Ensembl genome database includes 18 information for transcripts of the mouse Sox2ot gene [5].
We searched studies regarding the SOX2-OT gene on PubMed and found that the SOX2-OT gene has received unprecedented attention within the last five years. The PubMed records indicated that only 17 articles exploring the functions of the SOX2-OT gene were published before 2015, whereas nearly 80 articles investigating the SOX2-OT gene were published from 2015 to date (May 2020). The earliest study regarding the SOX2-OT gene was published in 2003 [6]. In this study, the SOX2 gene was discovered by genomic analysis to be located in an intron of another gene, which they named SOX2-OT [6]. The researchers demonstrated that SOX2-OT contains at least five exons (current studies have shown that it contains dozens of exons) and produces a mRNA-like transcript from the same strand that SOX2 is located on [6]. This transcript is evolutionarily conserved; the human SOX2-OT transcript and available mouse expressed sequence tags share 80% nucleotide identity [6]. In addition, the genomic region (approximately 40 kb) encompassing the SOX2-OT transcription unit is highly conserved across vertebrates [6]. Subsequently, accumulating evidence has indicated that the SOX2-OT gene is associated with mental illnesses, cancers, and diabetic complications. SOX2-OT expression is upregulated during the central nervous system development and is restricted to the brain in adult humans and other vertebrates [2, 7]. Therefore, single-nucleotide polymorphisms (SNPs) in the SOX2-OT gene are associated with mental illnesses [8, 9]. Moreover, an increased expression of SOX2-OT is observed in tissues from various cancers; SOX2-OT typically functions as an oncogene to influence cancer progression and can serve as a prognostic or diagnostic biomarker for cancers [10]. In addition, studies have demonstrated that the SOX2-OT gene is involved in diabetic complications and other diseases [11–13]. In this review, we comprehensively summarize the most recent research progress in the regulation and function of SOX2-OT and the association of this lncRNA with various diseases. Moreover, we discuss the potential opportunities and challenges revealed by these findings.
2. SOX2-OT Regulates SOX2 Expression
The SOX2 gene is a key regulator of stem cell pluripotency and is embedded in an intron of SOX2-OT [1, 2]. lncRNAs can regulate the expression of adjacent overlapping genes via specific mechanisms [14]. Various studies have investigated the regulatory relationship between SOX2-OT and SOX2 (Table 1). Almost all cancer studies involving SOX2-OT and SOX2 have indicated that upregulation of SOX2-OT promotes SOX2 expression in cancer cells (Table 1); however, one study showed that SOX2-OT overexpression did not affect SOX2 expression [15]. Studies on septic cardiomyopathy demonstrated that the level of SOX2-OT is inversely correlated with that of SOX2 [16]. Furthermore, the levels of SOX2-OT and SOX2 are negatively correlated during neural differentiation of mouse embryonic stem cells [7] (Figure 1).
Table 1.
SOX2-OT regulates SOX2 expression.
| Regulation | Intermediator | Cell model | Cellular function | Reference |
|---|---|---|---|---|
| Increase | miR-200c | Bladder cancer | Metastasis and stemness | Zhan et al. [17] |
| Increase | Unknown | Esophageal squamous cell carcinoma | Metastasis and stemness | Du et al. [56] |
| Decrease | Unknown | Septic cardiomyopathy | Mitochondrial dysfunction | Chen et al. [16] |
| No effect | Unknown | Esophageal squamous cell carcinoma | Cell proliferation | Wu et al. [15] |
| Increase | Unknown | Cholangiocarcinoma | Proliferation and metastasis | Wei et al. [57] |
| Decrease | Promoter-enhancer loop | Embryonic development | Neural differentiation | Messemaker et al. [7] |
| Increase | miR-200 family | Pancreatic ductal adenocarcinoma | EMT, stemness, invasion, and metastasis | Li et al. [18] |
| Decrease | YY1 | Embryonic development | Represses neural progenitor proliferation and promotes neuronal differentiation | Knauss et al. [20] |
| Increase | Unknown | Pancreatic ductal adenocarcinoma | Proliferation and tumor growth | Zhang et al. [19] |
| Increase | Unknown | Lung cancer | Proliferation, migration, invasion, and stemness | Wang et al. [38] |
| Increase | Unknown | Breast cancer | Proliferation | Askarian-Amiri et al. [58] |
Note: EMT: epithelial-mesenchymal transition; YY1: Yin Yang-1.
Figure 1.
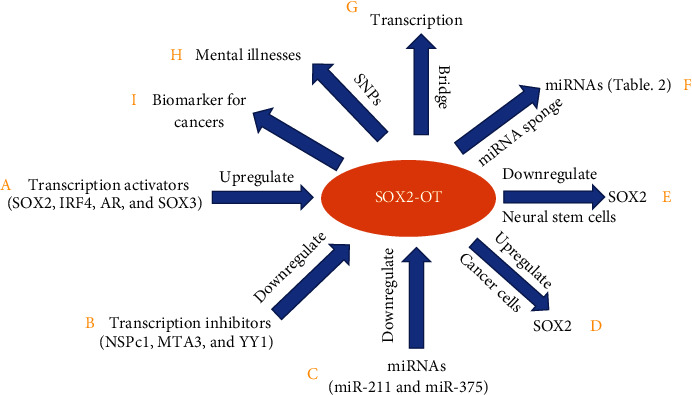
Long noncoding RNA SOX2-OT's regulations, functions, and roles on mental illnesses, cancers, and diabetic complications. Transcription activators ((A) SOX2, IRF4, AR, and SOX3) can upregulate SOX2-OT expression, but transcription inhibitors ((B) NSPc1, MTA3, and YY1) and miRNAs ((C) miR-211 and miR-375) downregulate SOX2-OT expression. D. SOX2-OT upregulates SOX2 expression in cancer cells. (E) SOX2-OT downregulates SOX2 expression in neural stem cells. SOX2-OT can control miRNA levels via serving as a miRNA sponge (F) and affect transcription via serving as a bridge between epigenetic factors and DNA (G). (H) SOX2-OT SNPs are associated with mental illnesses. (I) SOX2-OT is a biomarker for cancers.
Mechanistic investigations have revealed that SOX2-OT upregulates or downregulates the SOX2 expression through diverse pathways. Two studies demonstrated that SOX2-OT upregulates the SOX2 expression via the miR-200 family members in cancer cells [17, 18]. SOX2-OT acts as a miRNA sponge that competitively binds to miR-200 family members in order to upregulate the expression of SOX2 in cancer cells [17, 18]. One study revealed that the luciferase activity of the SOX2 promoter is significantly increased when SOX2-OT is overexpressed in pancreatic ductal adenocarcinoma cells, suggesting that SOX2-OT is a transcriptional activator of the SOX2 gene [19]. However, a study on central nervous system development showed that SOX2-OT physically interacts with the multifunctional transcriptional regulator YY1, which binds to several CpG islands in the SOX2 locus in a SOX2-OT-dependent manner and downregulates SOX2 expression in neural stem cells [20]. Another study showed that SOX2-OT impairs the formation of the chromatin promoter-enhancer loop upstream of the SOX2 gene and disrupts SOX2 transcription in neural stem cells [7]. Although few studies have investigated the mechanism by which SOX2-OT regulates the SOX2 expression in cancer cells or neural stem cells (Table 1), the regulation of the SOX2 expression by SOX2-OT in tumor cells follows a pattern opposite to that in neural stem cells (Table 1).
3. SOX2-OT Is a miRNA Sponge and a Regulator of Transcription
Research has suggested that some lncRNAs are involved in the competitive binding of miRNAs [21]. The members of this major subset of lncRNAs are called competing endogenous RNAs (ceRNAs), or miRNA sponges, and they form a regulatory network that controls the expression of protein-coding genes [22]. In this network, lncRNAs positively regulate the expression of protein-coding genes by competitively binding to their miRNAs [22]. SOX2-OT has been identified as an important ceRNA that affects cancer progression (Table 2, Figure 2(a)). An omics study revealed that SOX2-OT interacted with 6 differentially expressed miRNAs (hsa-mir-192-5p, hsa-mir-215-5p, hsa-mir-204-5p, hsa-mir-205-5p, hsa-mir-338-3p, hsa-mir-375) among 96 esophageal squamous cell carcinoma samples and 13 normal tissue samples [23]. In addition, numerous studies have demonstrated that SOX2-OT can bind to unique miRNAs in various cancers, and almost no overlapping miRNAs have been identified among those cancers (Table 2). miR-200c is the only exception, as SOX2-OT can target miR-200c in both bladder cancer and pancreatic ductal adenocarcinoma [17, 18]. Although SOX2-OT can target various miRNAs, it regulates similar cellular functions and behaviors, such as cancer cell proliferation, migration, invasion, metastasis, epithelial-mesenchymal transition (EMT), and stemness maintenance (Table 2).
Table 2.
SOX2-OT is a miRNA sponge.
| miRNA | Cancer | Target | Cellular function | Reference |
|---|---|---|---|---|
| hsa-mir-192-5p, hsa-mir-215-5p, hsa-mir-204-5p, hsa-mir-205-5p, hsa-mir-338-3p, hsa-mir-375 | Esophageal squamous cell carcinoma | Unknown | Unknown | Tian et al. [23] |
| miR-200c | Bladder cancer | SOX2 | Increases bladder cancer cell stemness and metastasis | Zhan et al. [17] |
| miR-146b-5p | Nasopharyngeal carcinoma | HNRNPA2B1 | Increases proliferation and metastasis; decreases apoptosis | Zhang and Li [59] |
| miR-369-3p | Prostate cancer | CFL2 | Increases proliferation and migration | Wo et al. [60] |
| miR-363 | Ewing's sarcoma | FOXP4 | Increases proliferation and invasion; decreases apoptosis | Ma et al. [61] |
| miR-132 | Non-small-cell lung cancer | ZEB2 | Increases proliferation, migration, invasion, and EMT | Zhang et al. [62] |
| miR-211 | Pheochromocytoma | MCL-1 isoform 2 | Increases cell viability, migration, and invasion; decreases apoptosis and autophagy | Yin et al. [49] |
| miR-194-5p | Gastric cancer | AKT2 | Promotes proliferation, metastasis, invasion, migration, and EMT | Wei et al. [63], Qu et al. [64] |
| miR-200 family | Pancreatic ductal adenocarcinoma | SOX2 | Promotes EMT and stem cell-like properties | Li et al. [18] |
| miR-194-5p, miR-122 | Glioma | SOX3 | Increases proliferation, migration, and invasion; decreases apoptosis | Su et al. [65] |
Note: EMT: epithelial-mesenchymal transition; Sox2: SRY-box transcription factor 2; HNRNPA2B1: heterogeneous nuclear ribonucleoprotein A2/B1; CFL2: cofilin 2; FOXP4: forkhead box P4; ZEB2: zinc finger E-box binding homeobox 2; MCL1 Isoform 2: myeloid cell leukemia sequence 1 isoform 2; AKT2: AKT serine/threonine kinase 2; SOX3: SRY-box transcription factor 3.
Figure 2.
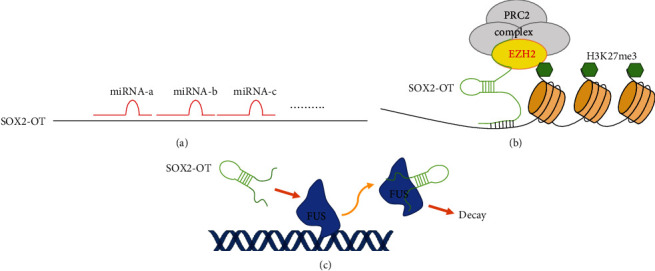
SOX2-OT is a miRNA sponge and a regulator of transcription. (a) SOX2-OT is a miRNA sponge. (b) SOX2-OT acts as a bridge between epigenetic factors and DNA to affect gene expression. (c) SOX2-OT acts as a destabilizer of transcription factors to control gene expression. PRC: polycomb repressive complex; EZH2: enhancer of zeste 2; FUS: FUS RNA binding protein.
In addition to acting as a miRNA sponge, SOX2-OT acts as a regulator of transcription by serving as a bridge between epigenetic factors and DNA to affect gene expression (Figure 2(b)). A recent study revealed that SOX2-OT interacts with EZH2, recruits EZH2 to DNA to form the polycomb repressive complex 2 (PRC2), induces H3K27me3, and epigenetically inhibits PTEN expression in laryngeal squamous cell carcinoma cells [24]. Studies have demonstrated that SOX2-OT binds to nervous system polycomb 1 (NSPc1), a key component of polycomb repressive complex 1 (PRC1), in H4 glioma cells [25] and U87 glioma cells [26], and regulates cancer cell proliferation and apoptosis. SOX2-OT can also act as a destabilizer of transcription factors to control gene expression (Figure 2(c)). A study suggested that SOX2-OT directly binds to the transcription factor FUS and that FUS protein stability is altered by this binding [27]. Thus, SOX2-OT acts as a tumor promoter in pancreatic ductal adenocarcinoma by physically binding to FUS to regulate its downstream cell cycle-associated factors CCND1 and p27 [27] (Figure 1).
4. SOX2-OT Is Regulated at the Transcriptional and Posttranscriptional Levels
Most relevant studies have shown that SOX2-OT levels are increased in various cancers and have described the SOX2-OT gene as an oncogene [10]. An increasing number of studies have investigated the mechanism underlying SOX2-OT upregulation in cancer cells (Table 3). These studies have focused on transcriptional and posttranscriptional regulation. Four transcription factors (SOX2, IRF4, AR, and SOX3) were identified to be able to bind directly to the SOX2-OT promoter and promote its transcription (Table 3). Other studies identified three transcriptional inhibitors (NSPc1, MTA3, and YY1) that recruit the repressive complex to the SOX2-OT promoter to repress its expression (Table 3). Interestingly, Shafiee et al. revealed that two miRNAs (miR-211 and miR-375) are responsible for SOX2-OT downregulation in a model of Helicobacter pylori-induced carcinogenesis (Table 3, Figure 1).
Table 3.
SOX2-OT is regulated at the transcriptional and posttranscriptional levels.
| Factor | Regulatory effect | Cell model | Reference |
|---|---|---|---|
| NSPc1 | Represses transcription | Glioma cells | Liang et al. [26] |
| MTA3 | Represses transcription | Esophageal squamous cell carcinoma cells | Du et al. [56] |
| SOX2 | Promotes transcription | Esophageal squamous cell carcinoma cells | Wu et al. [15] |
| IRF4 | Promotes transcription | Cholangiocarcinoma cells | Wei et al. [57] |
| YY1 | Represses transcription | Pancreatic ductal adenocarcinoma cells | Zhang et al. [19] |
| AR | Promotes transcription | Embryonic neural stem cells | Tosetti et al. [66] |
| SOX3 | Promotes transcription | Glioblastoma stem cells | Su et al. [65] |
| miR-211 | Downregulates expression by directly binding to SOX2-OT | Embryonal carcinoma stem cells (NT-2) | Shafiee et al. [67] |
| miR-375 | Downregulates expression by directly binding to SOX2-OT | Embryonal carcinoma stem cells (NT-2) | Shafiee et al. [68] |
Note: NSPc1: nervous system polycomb 1; MTA3: metastasis-associated protein 3; IRF4: interferon regulatory factor 4; YY1L Yin Yang-1; ARL androgen receptor; Sox3: SRY-box transcription factor 3.
5. SOX2-OT Is Upregulated during Central Nervous System Development, and Its Expression Is Restricted to the Brain
Studies have reported that a striking 40% of lncRNAs are expressed specifically in the brain, indicating the importance of lncRNAs in central nervous system development [28]. Numerous lncRNAs have been identified as regulators of the central nervous system development. Early studies showed that SOX2-OT is highly expressed in mouse embryonic stem cells and is downregulated during the differentiation of embryoid bodies into mesoderm [2]. However, Messemaker et al. demonstrated strong upregulation of SOX2-OT upon the differentiation of embryoid bodies into neuroectoderm, and upregulation of SOX2-OT was found to coincide with neural progenitor/stem cell formation as assessed via the induction of the SOX1 expression, which is a very early and specific marker of the neuroectodermal lineage [7]. Furthermore, SOX2-OT expressed sequence tags have been found in differentiated mouse neural stem cells, and its expression is confirmed in mouse primary neuronal cells [2]. RNA whole-mount in situ hybridization showed that in mice, SOX2-OT expression is limited to the developing brain, the ventral part of the neural tube, and the optic vesicle in mice [7]. Another study indicated that SOX2-OT is expressed in the developing cerebral cortex of mice, where it represses neural progenitor cell proliferation and promotes neuronal differentiation [20].
To investigate the possible involvement of SOX2-OT in neural differentiation processes, Amaral et al. examined the dynamic change in the SOX2-OT expression via a neurosphere assay, an in vitro model of neurogenesis with cultures of neurospheres originating from neural stem cells and undifferentiated precursors in the subventricular zone of adult mice [2]. The differentiated population of neurons and glial cells from neurospheres cultured for 7 days in differentiation medium exhibited increased expression of SOX2-OT [2].
Similar results have also been found in developing zebrafish embryos [7]. Studies have revealed that SOX2-OT is expressed in neuroectodermal tissue in zebrafish embryos at the tailbud stage [2]. Subsequently, SOX2-OT is highly expressed throughout the developing brain and eyes and is expressed at lower levels in the posterior neural tube at 28 hours postfertilization (hpf). In situ hybridization indicated specific expression of SOX2-OT in the retina and central nervous system in 48 hpf embryos, and this expression was maintained in the brain throughout the embryonic development until at least 6 days postfertilization (dpf) [2].
Importantly, data from the Genotype-Tissue Expression (GTEx) project show that in adult humans, the SOX2-OT expression is almost completely restricted to the brain, including regions such as the cortex, hippocampus, hypothalamus, cerebellum, and spinal cord [29]. Single-cell RNA-seq data in the Human Cell Landscape (HCL) project indicate that SOX2-OT expression is concentrated in oligodendrocytes and FGF13+ or CXCL14+ neurons in adult humans [30].
In summary, SOX2-OT is upregulated during central nervous system development (neurogenesis), and its expression is ultimately restricted to the brain in adult vertebrates.
6. SOX2-OT SNPs Are Associated with Mental Illnesses
Because SOX2-OT expression is restricted to the brain in adult humans, SOX2-OT SNPs are correlated with various mental illnesses, as identified by various studies. Genome-wide association studies (GWAS) indicate that the SNPs mapped to the SOX2-OT gene are associated with mental illnesses such as schizophrenia, general cognitive disorders, insomnia, eating disorders, night sleep phenotypes, and anorexia nervosa (Table 4). More than 50% of SOX2-OT-associated diseases are mental illnesses (Table 4). Interestingly, almost all SOX2-OT SNPs are located in the intronic region of the SOX2-OT gene, possibly because the SOX2-OT gene encompasses a genomic region of more than 750 kb. However, one mutation (rs75380963) is located in the exonic region of the SOX2-OT gene (Table 4). Some of the mutations, for example, rs2567646 (general cognitive disorders), rs2216428 (general cognitive disorders), rs4854912 (eating disorders in patients with bipolar disorder), and rs13086738 (eating disorders in patients with bipolar disorder), are strongly correlated with mental illnesses, with odds ratios (ORs) of greater than 1.5 (Table 4, Figure 1).
Table 4.
The SNPs of SOX2-OT are associated with various diseases.
| SNP | Mapped gene | Context | Disease/abnormality | PubMed ID |
|---|---|---|---|---|
| rs13096176 | SOX2-OT | intron_variant | Schizophrenia | 31740837 [69] |
| rs4855019 | SOX2-OT | intron_variant | Schizophrenia | 31740837 [69] |
| rs9841616 | SOX2-OT | intron_variant | Schizophrenia | 31740837 [69] |
| rs35788479 | SOX2-OT | intron_variant | General risk tolerance | 30643258 [70] |
| rs114600294 | SOX2-OT | intron_variant | General risk tolerance | 30643258 [70] |
| rs833268 | SOX2-OT | intron_variant | Male-pattern baldness | 30573740 [71] |
| rs12632136 | SOX2-OT | intron_variant | Reaction time | 29844566 [72] |
| rs2216428 | SOX2-OT | intron_variant | General cognitive disorder | 29844566 [72] |
| rs1345417 | SOX2-OT | intron_variant | Excessive hairiness | 29895819 [73] |
| rs60733335 | SOX2-OT | intron_variant | Hair color | 30595370 [74] |
| rs2216427 | SOX2-OT | intron_variant | Insomnia | 30804565 [75] |
| rs12485391 | SOX2-OT | intron_variant | Smoking status | 30595370 [74] |
| rs2567646 | SOX2-OT | intron_variant | General cognitive disorder | 29844566 [72] |
| rs9841616 | SOX2-OT | intron_variant | Schizophrenia | 25056061 [72] |
| rs1345417 | SOX2-OT | intron_variant | Eyebrow thickness | 30248107 [76] |
| rs9841616 | SOX2-OT | intron_variant | Schizophrenia | 28991256 [77] |
| rs9859557 | SOX2-OT | intron_variant | Schizophrenia | 28991256 [77] |
| rs833270 | SOX2-OT | intron_variant | Balding type 1 | 30595370 [74] |
| rs77025239 | SOX2-OT | intron_variant | Educational attainment | 30595370 [74] |
| rs1805207 | SOX2-OT | intron_variant | Body mass index | 30595370 [74] |
| rs9841616 | SOX2-OT | intron_variant | Schizophrenia | 26198764 [78] |
| rs1805203 | SOX2-OT | intron_variant | Schizophrenia | 26198764 [78] |
| rs1878874 | SOX2-OT | intron_variant | Schizophrenia | 26198764 [78] |
| rs13086738 | SOX2-OT | intron_variant | Eating disorder in individuals with bipolar disorder | 26433762 [79] |
| rs4854912 | SOX2-OT | intron_variant | Bipolar disorder and eating disorder | 26433762 [79] |
| rs1345417 | SOX2-OT | intron_variant | Monobrow | 27182965 [80] |
| rs2718791 | SOX2-OT | intron_variant | Smoking initiation | 30643251 [81] |
| rs9859557 | SOX2-OT | intron_variant | Schizophrenia | 30285260 [82] |
| rs9859557 | SOX2-OT | intron_variant | Schizophrenia | 30285260 [82] |
| rs9841616 | SOX2-OT | intron_variant | Schizophrenia | 30285260 [82] |
| rs9841616 | SOX2-OT | intron_variant | Schizophrenia | 30285260 [82] |
| rs75380963 | SOX2-OT | exon_variant | Corneal astigmatism | 30306274 [83] |
| rs77025239 | SOX2-OT | intron_variant | Educational attainment | 30038396 [84] |
| rs2718791 | SOX2-OT | intron_variant | Educational attainment | 30038396 [84] |
| rs77025239 | SOX2-OT | intron_variant | Educational attainment | 30038396 [84] |
| rs9841382 | SOX2-OT | intron_variant | Self-reported risk-taking behavior | 30271922 [85] |
| rs9841382 | SOX2-OT | intron_variant | Self-reported risk-taking behavior | 30181555 [86] |
| rs9841382 | SOX2-OT | intron_variant | Self-reported risk-taking behavior | 30181555 [86] |
| rs4133078 | SOX2-OT | intron_variant | Height | 30595370 [74] |
| rs7631379 | SOX2-OT | intron_variant | Smoking initiation | 30643251 [81] |
| rs34308817 | SOX2-OT | intron_variant | Ankle injury | 28957384 [87] |
| rs6443750 | SOX2-OT | intron_variant | Body mass index | 30595370 [74] |
| rs6443750 | SOX2-OT | intron_variant | Body mass index | 30239722 [88] |
| rs186834402 | SOX2-OT | intron_variant | Interferon gamma levels | 27989323 [89] |
| rs10937060 | SOX2-OT | intron_variant | Night sleep phenotypes | 27126917 [90] |
| rs9839776 | SOX2-OT | intron_variant | Anorexia nervosa | 24514567 [8] |
| rs4510419 | SOX2-OT | intron_variant | Smoking initiation | 30643251 [81] |
| rs9839776 | SOX2-OT | intron_variant | Breast cancer | 28240100 [31] |
| rs9839776 | SOX2-OT | intron_variant | Recurrent miscarriage | 31827385 [48] |
Note: SNPs: single-nucleotide polymorphisms; OR: odds ratio.
In contrast to the evidence supporting the relationship between SOX2-OT and mental illnesses, evidence for the association between SOX2-OT SNPs and cancers is scarce. We found no data regarding the association between SOX2-OT SNPs and cancers in the Catalogue of Somatic Mutations in Cancer (COSMIC) or The Cancer Genome Atlas (TCGA) Program database. However, one study demonstrated that a SOX2-OT SNP (rs9839776) is strongly associated with increased expression of SOX2-OT in breast cancer tissues and that this SNP increases the risk of breast cancer in Chinese women (OR: 1.42; 95% CI: 1.06-1.90; p = 0.018) [31]. In addition, another study revealed that copy number alteration (CNA) in the SOX2-OT locus is associated with esophageal squamous cell carcinoma [32].
7. SOX2-OT Is an Oncogene and a Biomarker for Cancers
lncRNAs have been demonstrated to be upregulated or downregulated during tumorigenesis and to function as oncogenes, suppressors, clinically useful diagnostic/prognostic biomarkers, or therapeutic targets in cancers because of their high sensitivity and specificity [33]. Accumulating evidence indicates that SOX2-OT is a key regulator of cancer stem cells and participates in cancer progression [10]. SOX2-OT is notably upregulated in numerous tumor tissues and cells (Table 5) and plays a vital role as an oncogene to promote the proliferation, invasion, migration, and growth of cancer cells and to suppress their apoptosis [10]. Depletion of SOX2-OT inhibits tumor cell proliferation, migration, invasion, and EMT [10]. However, a study showed that SOX2-OT is downregulated in gastric cancer, which contradicts the findings of the other four studies (Table 4). This contradictory result may have occurred because SOX2-OT has multiple splice variants. Indeed, Wang et al. thoroughly summarized recent studies regarding SOX2-OT expression, function, regulatory mechanisms, and clinical utility in human cancers [10].
Table 5.
Expression status of SOX2-OT in various cancers.
| Expression status | Cancer | Reference |
|---|---|---|
| Increased | Lung cancer | Hou et al. [34], Zhang et al. [62], Jazi et al. [91] |
| Decreased | Gastric cancer | Farhangian et al. [92] |
| Increased | Gastric cancer | Zou et al. [37], Zhang et al. [36], Wei et al. [63], Qu et al. [64] |
| Increased | Esophageal cancer | Aliereza et al. [93], Tian et al. [23], Wu et al. [15] |
| Increased | Breast cancer | Iranpour et al. [94], Tang et al. [31] |
| Increased | Hepatocellular carcinoma | Sun et al. [42], Shi et al. [35] |
| Increased | Ovarian cancer | Han et al. [95] |
| Increased | Pancreatic ductal adenocarcinoma | Li et al. [18], Zhang et al. [19] |
| Increased | Cholangiocarcinoma | Li et al. [40], Wei et al. [57] |
| Increased | Osteosarcoma | Wang et al. [38] |
| Increased | Laryngeal squamous cell carcinoma | Tai et al. [24], Feng et al. [96] |
| Increased | Nasopharyngeal carcinoma | Zhang et al. [59] |
| Increased | Glioblastoma | Wang et al. [25] |
| Increased | Bladder cancer | Zhan et al. [17] |
| Increased | Prostate cancer | Wo et al. [60] |
| Increased | Ewing's sarcoma | Ma et al. [61] |
| Increased | Colorectal cancer | Liu et al. [97] |
SOX2-OT has been identified as a novel lncRNA that can serve as a prognostic biomarker for cancers. A high level of SOX2-OT correlates well with poor clinical outcomes in cancers [34–45]. Li et al. performed a meta-analysis of 13 selected studies by a comprehensive search of PubMed, EMBASE, Cochrane Library, and TCGA and found that the elevated SOX2-OT expression is significantly related to shorter overall and disease-free survival times in cancer patients [45]. Cancer patients with high SOX2-OT expression are more likely to have an advanced clinical stage, earlier lymphatic metastasis, earlier distant metastasis, a larger tumor size, and more extreme tumor invasion than those with low SOX2-OT expression [45]. In addition, two other meta-analyses consistently demonstrated that high SOX2-OT expression is significantly associated with worse overall survival, advanced clinical stage, worse tumor differentiation, earlier distant metastasis, and earlier lymph node metastasis in various cancers [39, 41, 46]. SOX2-OT expression could thus be a promising prognostic biomarker for poor survival in a variety of cancers.
In addition to its prognostic value, circulating or exosome-derived SOX2-OT exhibits diagnostic value in non-small-cell lung cancer and lung squamous cell carcinoma [43, 44, 47]. Kamel et al. demonstrated that circulating SOX2-OT can distinguish non-small-cell lung cancer patients from control individuals, with an area under the curve of 0.73 (76.3% sensitivity and 78.6% specificity) [44]. Moreover, the combination of GAS5 expression and SOX2-OT expression can differentiate non-small-cell lung cancer patients from control individuals with increased sensitivity (83.8) and specificity (81.4) compared with those of SOX2-OT expression alone [44]. Teng et al. analyzed the level of exosomal SOX2-OT in plasma and concluded that the level of exosomal SOX2-OT is significantly increased in lung squamous cell carcinoma patients compared to normal control individuals, indicating the strong power of exosomal SOX2-OT for detecting lung squamous cell carcinoma. In that analysis, the area under the curve was 0.815, and the sensitivity and specificity were 76% and 73.17%, respectively [47]. Thus, SOX2-OT may serve as a promising noninvasive plasma-based diagnostic biomarker for cancers (Figure 1).
8. SOX2-OT Mediates Diabetic Complications
A few studies have investigated the possible association of SOX2-OT with diabetic complications, including diabetic nephropathy [12, 13] and diabetic retinopathy [11]. Microarray and bioinformatics analyses indicated that SOX2-OT is significantly downregulated in mice with diabetic nephropathy compared to control mice, and this result was confirmed in cultured human podocytes and mesangial cells [12]. SOX2-OT overexpression significantly alleviates high glucose-induced injury to human podocytes via autophagy induction through the miR-9/SIRT1 axis [13]. Conversely, although the SOX2-OT expression is significantly downregulated in the retinas of mice with streptozocin-induced diabetes, SOX2-OT knockdown protects retinal ganglion cells against high glucose-induced injury in vitro [11].
9. SOX2-OT and Other Diseases
In addition to the evidence supporting its involvement in cancers, mental illnesses, and diabetic complications, emerging evidence indicates the association of SOX2-OT with other diseases and events, such as miscarriage [48], septic cardiomyopathy [16], spinal cord injury [49], multiple sclerosis [50], and myopia [51]. An SNP (rs9839776 C>T) in the intronic region of the SOX2-OT gene is associated with increased risk for recurrent miscarriage (CT vs. CC: adjusted OR = 1.357, 95%CI = 1.065 − 1.728, p = 0.0134) [48]. In addition, Chen et al. found that SOX2-OT was overexpressed and mitochondrial dysfunction occurred in a mouse model of lipopolysaccharide-induced septic cardiomyopathy; moreover, cardiac-specific knockdown of SOX2-OT via adeno-associated virus 9 (AAV9) harboring SOX2-OT siRNA ameliorated mitochondrial dysfunction in septic cardiomyopathy [16]. A lncRNA PCR array containing 90 common lncRNAs in peripheral blood mononuclear cells from patients with multiple sclerosis revealed a group of dysregulated lncRNAs in multiple sclerosis patients, and SOX2-OT was one of the most strongly downregulated lncRNAs with p < 0.001 [50]. However, the SOX2-OT level is not associated with clinical variables such as the disease duration and expanded disability status scale score [50].
10. Conclusions and Future Directions
SOX2-OT is upregulated in many cancers and plays an oncogenic role in most tumors. In addition, SOX2-OT is upregulated during central nervous system development and is ultimately restricted to the brain in adult vertebrates. Emerging evidence indicates that multiple factors, including transcriptional activators (SOX2, IRF4, AR, and SOX3) and transcriptional inhibitors (NSPc1, MTA3, and YY1), as well as miRNAs (miR-211 and miR-375), can control the SOX2-OT expression transcriptionally or posttranscriptionally. However, rigorous investigations of the cause and effect mechanism underlying its upregulation in cancers and the central nervous system remain limited.
The downstream targets of SOX2-OT have been elucidated. SOX2-OT performs various molecular and cellular functions via regulation of SOX2 (direct or indirect interactions), regulation of miRNAs (acting as a miRNA sponge), or regulation of transcriptional process (acting as a bridge between epigenetic factors and DNA). However, the precise role of the SOX2-OT gene in neurogenesis, cancers, mental illnesses, and diabetic complications must be systematically investigated and confirmed in a knockout animal model. Currently, no SOX2-OT knockout model is available to demonstrate the essential role of the SOX2-OT gene in neurogenesis and various diseases, because genetic depletion of a lncRNA—especially a lncRNA with multiple exons and transcription start sites, such as SOX2-OT—is difficult. Fortunately, strategies have been applied to generate lncRNA knockout mice, i.e., transcription start site disruption through the insertion of a transcription termination signal and deletion of important gene segments/exons via CRISPR/Cas9 genome editing [52, 53].
Due to the complexity of transcriptional characteristics, including multiple transcription start sites and numerous transcripts in humans and other vertebrates, each transcript may play a unique role in different tissues, embryonic developmental stages, and disease conditions. There is an urgent demand to develop a method to systemically study each transcript under specific conditions. The most recently developed pooled CRISPR screening platform may constitute a good approach for studying the function of each SOX2-OT transcript [54, 55].
SOX2-OT SNPs are associated with mental illnesses, but the precise functions of these SNPs are still obscure. We may need to investigate whether these SNPs alter SOX2-OT expression. In addition, the upregulation of SOX2-OT is correlated with poor outcomes in cancer patients, suggesting its potential function as a diagnostic and prognostic marker in tumors. However, the expression and chemical stability of SOX2-OT in body fluids remain unclear.
The SOX2-OT gene has been widely studied in the past five years, and many important accomplishments have been achieved. However, studies on the SOX2-OT gene are still rare; less than one hundred papers on the SOX2-OT gene have been to date, despite an increasing trend. We still face many challenges, and many aspects of the SOX2-OT gene need to be investigated to provide a foundation for understanding its functions.
Acknowledgments
We sincerely thank Springer Nature Author Services for language editing. This research was funded by the National Natural Science Foundation of China (grant numbers 81901485, 81472234, and 81972571).
Conflicts of Interest
The authors declare no conflicts of interest.
Authors' Contributions
D.Y.D did the conceptualization. P.Y.L wrote and prepared the original draft. S.G.G and P.W. wrote, reviewed, and edited the manuscript. D.Y.D supervised the study. D.Y.D. and S.G.G acquired funding. All authors have read and agreed to the published version of the manuscript.
References
- 1.Shahryari A., Jazi M. S., Samaei N. M., Mowla S. J. Long non-coding RNA SOX2OT: expression signature, splicing patterns, and emerging roles in pluripotency and tumorigenesis. Frontiers in Genetics. 2015;6 doi: 10.3389/fgene.2015.00196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Amaral P. P., Neyt C., Wilkins S. J., et al. Complex architecture and regulated expression of the Sox2ot locus during vertebrate development. RNA. 2009;15(11):2013–2027. doi: 10.1261/rna.1705309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ensembl genome database (human) 2020, http://useast.ensembl.org/Homo_sapiens/Gene/Summary?db=core;g=ENSG00000242808;r=3:180989762-181836880.
- 4.RNACentral. 2020, https://rnacentral.org/search?q=sox2-ot%20AND%20TAXONOMY:%229606%22%20AND%20rna_type:%22lncRNA%22.
- 5.Ensembl genome database (mouse) 2020, http://useast.ensembl.org/Mus_musculus/Gene/Summary?db=core;g=ENSMUSG00000105265;r=3:34104270-34682619.
- 6.Fantes J., Ragge N. K., Lynch S. A., et al. Mutations in SOX2 cause anophthalmia. Nature Genetics. 2003;33(4):461–463. doi: 10.1038/ng1120. [DOI] [PubMed] [Google Scholar]
- 7.Messemaker T. C., van Leeuwen S. M., van den Berg P. R., et al. Allele-specific repression of Sox2 through the long non-coding RNA Sox2ot. Scientific Reports. 2018;8(1):p. 386. doi: 10.1038/s41598-017-18649-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Boraska V., Franklin C. S., Floyd J. A. B., et al. A genome-wide association study of anorexia nervosa. Molecular Psychiatry. 2014;19(10):1085–1094. doi: 10.1038/mp.2013.187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Arisi I., D'Onofrio M., Brandi R., et al. Gene expression biomarkers in the brain of a mouse model for Alzheimer's disease: mining of microarray data by logic classification and feature selection. Journal of Alzheimer's Disease. 2011;24(4):721–738. doi: 10.3233/JAD-2011-101881. [DOI] [PubMed] [Google Scholar]
- 10.Wang Y., Wu N., Luo X., Zhang X., Liao Q., Wang J. SOX2OT, a novel tumor-related long non-coding RNA. Biomedicine & Pharmacotherapy. 2020;123, article 109725 doi: 10.1016/j.biopha.2019.109725. [DOI] [PubMed] [Google Scholar]
- 11.Li C. P., Wang S. H., Wang W. Q., Song S. G., Liu X. M. Long noncoding RNA-Sox2OT knockdown alleviates diabetes mellitus-induced retinal ganglion cell (RGC) injury. Cellular and Molecular Neurobiology. 2017;37(2):361–369. doi: 10.1007/s10571-016-0380-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zhang X., Shang J., Wang X., et al. Microarray analysis reveals long non‑coding RNA SOX2OT as a novel candidate regulator in diabetic nephropathy. Molecular Medicine Reports. 2018;18(6):5058–5068. doi: 10.3892/mmr.2018.9534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zhang Y., Chang B., Zhang J., Wu X. LncRNA SOX2OT alleviates the high glucose-induced podocytes injury through autophagy induction by the miR-9/SIRT1 axis. Experimental and Molecular Pathology. 2019;110, article 104283 doi: 10.1016/j.yexmp.2019.104283. [DOI] [PubMed] [Google Scholar]
- 14.Marchese F. P., Raimondi I., Huarte M. The multidimensional mechanisms of long noncoding RNA function. Genome Biology. 2017;18(1):p. 206. doi: 10.1186/s13059-017-1348-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wu Y., Liang Y., Li J., et al. Overexpression of long non-coding RNA SOX2OT promotes esophageal squamous cell carcinoma growth. Cancer Cell International. 2018;18(1) doi: 10.1186/s12935-018-0570-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chen M., Guan Y., Li A., et al. LncRNA SOX2OT mediates mitochondrial dysfunction in septic cardiomyopathy. DNA and Cell Biology. 2019;38(11):1197–1206. doi: 10.1089/dna.2019.4839. [DOI] [PubMed] [Google Scholar]
- 17.Zhan Y., Chen Z., He S., et al. Long non-coding RNA SOX2OT promotes the stemness phenotype of bladder cancer cells by modulating SOX2. Molecular Cancer. 2020;19(1):p. 25. doi: 10.1186/s12943-020-1143-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Li Z., Jiang P., Li J., et al. Tumor-derived exosomal lnc-Sox2ot promotes EMT and stemness by acting as a ceRNA in pancreatic ductal adenocarcinoma. Oncogene. 2018;37(28):3822–3838. doi: 10.1038/s41388-018-0237-9. [DOI] [PubMed] [Google Scholar]
- 19.Zhang J. J., Zhu Y., Zhang X. F., et al. Yin Yang-1 suppresses pancreatic ductal adenocarcinoma cell proliferation and tumor growth by regulating SOX2OT-SOX2 axis. Cancer Letters. 2017;408:144–154. doi: 10.1016/j.canlet.2017.08.032. [DOI] [PubMed] [Google Scholar]
- 20.Knauss J. L., Miao N., Kim S. N., et al. Long noncoding RNA Sox2ot and transcription factor YY1 co-regulate the differentiation of cortical neural progenitors by repressing Sox2. Cell Death & Disease. 2018;9(8):p. 799. doi: 10.1038/s41419-018-0840-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Yoon J. H., Abdelmohsen K., Gorospe M. Functional interactions among microRNAs and long noncoding RNAs. Seminars in Cell & Developmental Biology. 2014;34:9–14. doi: 10.1016/j.semcdb.2014.05.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bak R. O., Mikkelsen J. G. miRNA sponges: soaking up miRNAs for regulation of gene expression. Wiley Interdisciplinary Reviews: RNA. 2014;5(3):317–333. doi: 10.1002/wrna.1213. [DOI] [PubMed] [Google Scholar]
- 23.Tian W., Jiang C., Huang Z., Xu D., Zheng S. Comprehensive analysis of dysregulated lncRNAs, miRNAs and mRNAs with associated ceRNA network in esophageal squamous cell carcinoma. Gene. 2019;696:206–218. doi: 10.1016/j.gene.2019.02.051. [DOI] [PubMed] [Google Scholar]
- 24.Tai Y., Ji Y., Liu F., et al. Long noncoding RNA SOX2-OT facilitates laryngeal squamous cell carcinoma development by epigenetically inhibiting PTEN via methyltransferase EZH2. IUBMB Life. 2019;71(9):1230–1239. doi: 10.1002/iub.2026. [DOI] [PubMed] [Google Scholar]
- 25.Wang Y., Liang Z., Li H., Tao J., Sun Y., Gong Y. NSPc1 polycomb protein complex binds and cross‑talks to lncRNAs in glioma H4 cells. Oncology Reports. 2019;41(4):2575–2584. doi: 10.3892/or.2019.7000. [DOI] [PubMed] [Google Scholar]
- 26.Liang Z., Wang Y., Li H., Sun Y., Gong Y. lncRNAs combine and crosstalk with NSPc1 in ATRA-induced differentiation of U87 glioma cells. Oncology Letters. 2019;17:5821–5829. doi: 10.3892/ol.2019.10254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Chen L., Zhang J., Chen Q., et al. Long Noncoding RNA SOX2OT Promotes the Proliferation of Pancreatic Cancer by Binding to FUS. International Journal of Cancer. 2020;147(1):175–188. doi: 10.1002/ijc.32827. [DOI] [PubMed] [Google Scholar]
- 28.Briggs J. A., Wolvetang E. J., Mattick J. S., Rinn J. L., Barry G. Mechanisms of long non-coding RNAs in mammalian nervous system development, plasticity, disease, and evolution. Neuron. 2015;88(5):861–877. doi: 10.1016/j.neuron.2015.09.045. [DOI] [PubMed] [Google Scholar]
- 29.Genotype-tissue expression (GTEx) project. 2020, https://www.gtexportal.org/home/gene/SOX2-OT.
- 30.Human cell landscape (HCL) project. 2020, https://db.cngb.org/HCL/index.html.
- 31.Tang X., Gao Y., Yu L., et al. Correlations between lncRNA-SOX2OT polymorphism and susceptibility to breast cancer in a Chinese population. Biomarkers in Medicine. 2017;11(3):277–284. doi: 10.2217/bmm-2016-0238. [DOI] [PubMed] [Google Scholar]
- 32.Wang P., Shan L., Xue L., Zheng B., Ying J., Lu N. Genome wide copy number analyses of superficial esophageal squamous cell carcinoma with and without metastasis. Oncotarget. 2017;8(3):5069–5080. doi: 10.18632/oncotarget.13847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Huarte M. The emerging role of lncRNAs in cancer. Nature Medicine. 2015;21(11):1253–1261. doi: 10.1038/nm.3981. [DOI] [PubMed] [Google Scholar]
- 34.Hou Z., Zhao W., Zhou J., et al. A long noncoding RNA Sox2ot regulates lung cancer cell proliferation and is a prognostic indicator of poor survival. The International Journal of Biochemistry & Cell Biology. 2014;53:380–388. doi: 10.1016/j.biocel.2014.06.004. [DOI] [PubMed] [Google Scholar]
- 35.Shi X. M., Teng F. Up-regulation of long non-coding RNA Sox2ot promotes hepatocellular carcinoma cell metastasis and correlates with poor prognosis. International Journal of Clinical and Experimental Pathology. 2015;8(4):4008–4014. [PMC free article] [PubMed] [Google Scholar]
- 36.Zhang Y., Yang R., Lian J., Xu H. LncRNA Sox2ot overexpression serves as a poor prognostic biomarker in gastric cancer. American Journal of Translational Research. 2016;8(11):5035–5043. [PMC free article] [PubMed] [Google Scholar]
- 37.Zou J. H., Li C. Y., Bao J., Zheng G. Q. High expression of long noncoding RNA Sox2ot is associated with the aggressive progression and poor outcome of gastric cancer. European Review for Medical and Pharmacological Sciences. 2016;20(21):4482–4486. [PubMed] [Google Scholar]
- 38.Wang Z., Tan M., Chen G., Li Z., Lu X. LncRNA SOX2-OT is a novel prognostic biomarker for osteosarcoma patients and regulates osteosarcoma cells proliferation and motility through modulating SOX2. IUBMB Life. 2017;69(11):867–876. doi: 10.1002/iub.1681. [DOI] [PubMed] [Google Scholar]
- 39.Gao S., Zhao Z. Y., Wu R., Zhang Y., Zhang Z. Y. Prognostic value of long noncoding RNAs in gastric cancer: a meta-analysis. Oncotargets and Therapy. 2018;11:4877–4891. doi: 10.2147/OTT.S169823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Li Z., Li J., Ji D., et al. Overexpressed long noncoding RNA Sox2ot predicts poor prognosis for cholangiocarcinoma and promotes cell proliferation and invasion. Gene. 2018;645:131–136. doi: 10.1016/j.gene.2017.12.017. [DOI] [PubMed] [Google Scholar]
- 41.Song X., Yao H., Liu J., Wang Q. The prognostic value of long noncoding RNA Sox2ot expression in various cancers: a systematic review and meta-analysis. Clinica Chimica Acta. 2018;484:52–59. doi: 10.1016/j.cca.2018.05.038. [DOI] [PubMed] [Google Scholar]
- 42.Sun J., Wei X., Xu L. Upregulation of lncRNA Sox2ot indicates a poor prognosis for patients with hepatocellular carcinoma and promotes cell invasion. Oncology Letters. 2018;16(1):1189–1195. doi: 10.3892/ol.2018.8725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Xie Y., Zhang Y., du L., et al. Circulating long noncoding RNA act as potential novel biomarkers for diagnosis and prognosis of non-small cell lung cancer. Molecular Oncology. 2018;12(5):648–658. doi: 10.1002/1878-0261.12188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kamel L. M., Atef D. M., Mackawy A. M. H., Shalaby S. M., Abdelraheim N. Circulating long non-coding RNA GAS5 and SOX2OT as potential biomarkers for diagnosis and prognosis of non-small cell lung cancer. Biotechnology and Applied Biochemistry. 2019;66(4):634–642. doi: 10.1002/bab.1764. [DOI] [PubMed] [Google Scholar]
- 45.Li Y., Du M., Wang S., et al. Clinicopathological implication of long non-coding RNAs SOX2 overlapping transcript and its potential target gene network in various cancers. Frontiers in Genetics. 2020;10 doi: 10.3389/fgene.2019.01375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Jing X., Lin J., Wang H., et al. Prognostic value of lncRNA SOX2OT for Chinese cancer patients: a meta-analysis. PLoS One. 2017;12(5, article e0176889) doi: 10.1371/journal.pone.0176889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Teng Y., Kang H., Chu Y. Identification of an exosomal long noncoding RNA SOX2-OT in plasma as a promising biomarker for lung squamous cell carcinoma. Genetic Testing and Molecular Biomarkers. 2019;23(4):235–240. doi: 10.1089/gtmb.2018.0103. [DOI] [PubMed] [Google Scholar]
- 48.Fang Z., Che D., Qing S., et al. The lncRNA SOX2OT rs9839776 C>T polymorphism indicates recurrent miscarriage susceptibility in a southern Chinese population. Mediators of Inflammation. 2019;2019:6. doi: 10.1155/2019/9684703.9684703 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Yin D., Zheng X., Zhuang J., Wang L., Liu B., Chang Y. Downregulation of long noncoding RNA Sox2ot protects PC-12 cells from hydrogen peroxide-induced injury in spinal cord injury via regulating the miR-211-myeloid cell leukemia-1 isoform2 axis. Journal of Cellular Biochemistry. 2018;119(12):9675–9684. doi: 10.1002/jcb.27280. [DOI] [PubMed] [Google Scholar]
- 50.Fenoglio C., Oldoni E., Serpente M., et al. LncRNAs expression profile in peripheral blood mononuclear cells from multiple sclerosis patients. Journal of Neuroimmunology. 2018;324:129–135. doi: 10.1016/j.jneuroim.2018.08.008. [DOI] [PubMed] [Google Scholar]
- 51.Andrew T., Maniatis N., Carbonaro F., et al. Identification and replication of three novel myopia common susceptibility gene loci on chromosome 3q26 using linkage and linkage disequilibrium mapping. PLoS Genetics. 2008;4(10, article e1000220) doi: 10.1371/journal.pgen.1000220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Hansmeier N. R., Widdershooven P. J. M., Khani S., Kornfeld J. W. Rapid generation of long noncoding RNA knockout mice using CRISPR/Cas9 technology. Non-Coding RNA. 2019;5(1):p. 12. doi: 10.3390/ncrna5010012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Zare K., Shademan M., Ghahramani Seno M. M., Dehghani H. CRISPR/Cas9 knockout strategies to ablate CCAT1 lncRNA gene in cancer cells. Biological Procedures Online. 2018;20(1):p. 21. doi: 10.1186/s12575-018-0086-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Wang T., Wei J. J., Sabatini D. M., Lander E. S. Genetic screens in human cells using the CRISPR-Cas9 system. Science. 2014;343(6166):80–84. doi: 10.1126/science.1246981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Shalem O., Sanjana N. E., Hartenian E., et al. Genome-scale CRISPR-Cas9 knockout screening in human cells. Science. 2014;343(6166):84–87. doi: 10.1126/science.1247005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Du L., Wang L., Gan J., et al. MTA3 represses cancer stemness by targeting the SOX2OT/SOX2 Axis. iScience. 2019;22:353–368. doi: 10.1016/j.isci.2019.11.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Wei C. X., Wong H., Xu F., Liu Z., Ran L., Jiang R. D. IRF4-induced upregulation of lncRNA SOX2-OT promotes cell proliferation and metastasis in cholangiocarcinoma by regulating SOX2 and PI3K/AKT signaling. European Review for Medical and Pharmacological Sciences. 2018;22(23):8169–8178. doi: 10.26355/eurrev_201812_16509. [DOI] [PubMed] [Google Scholar]
- 58.Askarian-Amiri M. E., Seyfoddin V., Smart C. E., et al. Emerging role of long non-coding RNA SOX2OT in SOX2 regulation in breast cancer. PLoS One. 2014;9(7, article e102140) doi: 10.1371/journal.pone.0102140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Zhang E., Li X. LncRNA SOX2-OT regulates proliferation and metastasis of nasopharyngeal carcinoma cells through miR-146b-5p/HNRNPA2B1 pathway. Journal of Cellular Biochemistry. 2019;120(10):16575–16588. doi: 10.1002/jcb.28917. [DOI] [PubMed] [Google Scholar]
- 60.Wo Q., Zhang D., Hu L., et al. Long noncoding RNA SOX2-OT facilitates prostate cancer cell proliferation and migration via miR-369-3p/CFL2 axis. Biochemical and Biophysical Research Communications. 2019;520(3):586–593. doi: 10.1016/j.bbrc.2019.09.108. [DOI] [PubMed] [Google Scholar]
- 61.Ma L., Sun X., Kuai W., et al. LncRNA SOX2 overlapping transcript acts as a miRNA sponge to promote the proliferation and invasion of Ewing's sarcoma. American Journal of Translational Research. 2019;11(6):3841–3849. [PMC free article] [PubMed] [Google Scholar]
- 62.Zhang K., Li Y., Qu L., Ma X., Zhao H., Tang Y. Long noncoding RNA Sox2 overlapping transcript (SOX2OT) promotes non-small-cell lung cancer migration and invasion via sponging microRNA 132 (miR-132) Oncotargets and Therapy. 2018;11:5269–5278. doi: 10.2147/OTT.S168654. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 63.Wei R., Ding C., Rodriguez R. A., del Mar Requena Mullor M. The SOX2OT/miR-194-5p axis regulates cell proliferation and mobility of gastric cancer through suppressing epithelial-mesenchymal transition. Oncology Letters. 2018;16:6361–6368. doi: 10.3892/ol.2018.9433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Qu F., Cao P. Long noncoding RNA SOX2OT contributes to gastric cancer progression by sponging miR-194-5p from AKT2. Experimental Cell Research. 2018;369(2):187–196. doi: 10.1016/j.yexcr.2018.05.017. [DOI] [PubMed] [Google Scholar]
- 65.Su R., Cao S., Ma J., et al. Knockdown of SOX2OT inhibits the malignant biological behaviors of glioblastoma stem cells via up-regulating the expression of miR-194-5p and miR-122. Molecular Cancer. 2017;16(1):p. 171. doi: 10.1186/s12943-017-0737-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Tosetti V., Sassone J., Ferri A. L. M., et al. Transcriptional role of androgen receptor in the expression of long non-coding RNA Sox2OT in neurogenesis. PLoS One. 2017;12(7, article e0180579) doi: 10.1371/journal.pone.0180579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Shafiee M., Aleyasin S. A., Vasei M., Semnani S. S., Mowla S. J. Down-regulatory effects of miR-211 on long non-coding RNA SOX2OT and SOX2 genes in esophageal squamous cell carcinoma. Cell Journal. 2016;17(4):593–600. doi: 10.22074/cellj.2016.3811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Shafiee M., Aleyasin S. A., Mowla S. J., Vasei M., Yazdanparast S. A. The effect of microRNA-375 overexpression, an inhibitor of helicobacter pylori-induced carcinogenesis, on lncRNA SOX2OT. Jundishapur Journal of Microbiology. 2016;9(9, article e23464) doi: 10.5812/jjm.23464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Lam M., Chen C.-Y., Li Z., et al. Comparative genetic architectures of schizophrenia in East Asian and European populations. Nature Genetics. 2019;51(12):1670–1678. doi: 10.1038/s41588-019-0512-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Linnér R. K., Biroli P., Kong E., et al. Genome-wide association analyses of risk tolerance and risky behaviors in over 1 million individuals identify hundreds of loci and shared genetic influences. Nature Genetics. 2019;51(2):245–257. doi: 10.1038/s41588-018-0309-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Yap C. X., Sidorenko J., Wu Y., et al. Dissection of genetic variation and evidence for pleiotropy in male pattern baldness. Nature Communications. 2018;9(1):p. 5407. doi: 10.1038/s41467-018-07862-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Davies G., Lam M., Harris S. E., et al. Study of 300,486 individuals identifies 148 independent genetic loci influencing general cognitive function. Nature Communications. 2018;9(1):p. 2098. doi: 10.1038/s41467-018-04362-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Endo C., Johnson T. A., Morino R., et al. Genome-wide association study in Japanese females identifies fifteen novel skin-related trait associations. Scientific Reports. 2018;8(1):p. 8974. doi: 10.1038/s41598-018-27145-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Kichaev G., Bhatia G., Loh P. R., et al. Leveraging polygenic functional enrichment to improve GWAS power. American Journal of Human Genetics. 2019;104(1):65–75. doi: 10.1016/j.ajhg.2018.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Jansen P. R., Watanabe K., Stringer S., et al. Genome-wide analysis of insomnia in 1,331,010 individuals identifies new risk loci and functional pathways. Nature Genetics. 2019;51(3):394–403. doi: 10.1038/s41588-018-0333-3. [DOI] [PubMed] [Google Scholar]
- 76.Wu S., Zhang M., Yang X., et al. Genome-wide association studies and CRISPR/Cas9-mediated gene editing identify regulatory variants influencing eyebrow thickness in humans. PLoS Genetics. 2018;14(9, article e1007640) doi: 10.1371/journal.pgen.1007640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Li Z., Chen J., Yu H., et al. Genome-wide association analysis identifies 30 new susceptibility loci for schizophrenia. Nature Genetics. 2017;49(11):1576–1583. doi: 10.1038/ng.3973. [DOI] [PubMed] [Google Scholar]
- 78.Goes F. S., McGrath J., Avramopoulos D., et al. Genome-wide association study of schizophrenia in Ashkenazi Jews. American Journal of Medical Genetics. Part B, Neuropsychiatric Genetics. 2015;168(8):649–659. doi: 10.1002/ajmg.b.32349. [DOI] [PubMed] [Google Scholar]
- 79.Liu X., Bipolar Genome S., Kelsoe J. R., Greenwood T. A. A genome-wide association study of bipolar disorder with comorbid eating disorder replicates the SOX2-OT region. Journal of Affective Disorders. 2016;189:141–149. doi: 10.1016/j.jad.2015.09.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Pickrell J. K., Berisa T., Liu J. Z., Ségurel L., Tung J. Y., Hinds D. A. Detection and interpretation of shared genetic influences on 42 human traits. Nature Genetics. 2016;48(7):709–717. doi: 10.1038/ng.3570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Liu M., Jiang Y., Wedow R., et al. Association studies of up to 1.2 million individuals yield new insights into the genetic etiology of tobacco and alcohol use. Nature Genetics. 2019;51(2):237–244. doi: 10.1038/s41588-018-0307-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Ikeda M., Takahashi A., Kamatani Y., et al. Genome-wide association study detected novel susceptibility genes for schizophrenia and shared trans-populations/diseases genetic effect. Schizophrenia Bulletin. 2019;45(4):824–834. doi: 10.1093/schbul/sby140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Shah R. L., Guggenheim J. A., Eye U. K. B., Vision C. Genome-wide association studies for corneal and refractive astigmatism in UK Biobank demonstrate a shared role for myopia susceptibility loci. Human Genetics. 2018;137(11-12):881–896. doi: 10.1007/s00439-018-1942-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Lee J. J., Wedow R., Okbay A., et al. Gene discovery and polygenic prediction from a genome-wide association study of educational attainment in 1.1 million individuals. Nature Genetics. 2018;50(8):1112–1121. doi: 10.1038/s41588-018-0147-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Clifton E. A. D., Perry J. R. B., Imamura F., et al. Genome-wide association study for risk taking propensity indicates shared pathways with body mass index. Communications Biology. 2018;1(1):p. 36. doi: 10.1038/s42003-018-0042-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Strawbridge R. J., Ward J., Lyall L. M., et al. Genetics of self-reported risk-taking behaviour, trans-ethnic consistency and relevance to brain gene expression. Translational Psychiatry. 2018;8(1):p. 178. doi: 10.1038/s41398-018-0236-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Kim S. K., Kleimeyer J. P., Ahmed M. A., et al. Two genetic loci associated with ankle injury. PLoS One. 2017;12(9, article e0185355) doi: 10.1371/journal.pone.0185355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Pulit S. L., Stoneman C., Morris A. P., et al. Meta-analysis of genome-wide association studies for body fat distribution in 694 649 individuals of European ancestry. Human Molecular Genetics. 2019;28(1):166–174. doi: 10.1093/hmg/ddy327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Ahola-Olli A. V., Würtz P., Havulinna A. S., et al. Genome-wide association study identifies 27 loci influencing concentrations of circulating cytokines and growth factors. American Journal of Human Genetics. 2017;100(1):40–50. doi: 10.1016/j.ajhg.2016.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Spada J., Scholz M., Kirsten H., et al. Genome-wide association analysis of actigraphic sleep phenotypes in the LIFE Adult Study. Journal of Sleep Research. 2016;25(6):690–701. doi: 10.1111/jsr.12421. [DOI] [PubMed] [Google Scholar]
- 91.Saghaeian Jazi M., Samaei N. M., Ghanei M., Shadmehr M. B., Mowla S. J. Overexpression of the non-coding SOX2OT variants 4 and 7 in lung tumors suggests an oncogenic role in lung cancer. Tumour biology : the journal of the International Society for Oncodevelopmental Biology and Medicine. 2016;37(8):10329–10338. doi: 10.1007/s13277-016-4901-9. [DOI] [PubMed] [Google Scholar]
- 92.Farhangian P., Jahandoost S., Mowla S. J., Khalili M. Differential expression of long non-coding RNA SOX2OT in gastric adenocarcinoma. Cancer Biomarkers : section A of Disease markers. 2018;23(2):221–225. doi: 10.3233/CBM-181325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Shahryari A., Rafiee M. R., Fouani Y., et al. Two novel splice variants of SOX2OT, SOX2OT-S1, and SOX2OT-S2 are coupregulated with SOX2 and OCT4 in esophageal squamous cell carcinoma. Stem Cells (Dayton, Ohio) 2014;32(1):126–134. doi: 10.1002/stem.1542. [DOI] [PubMed] [Google Scholar]
- 94.Iranpour M., Soudyab M., Geranpayeh L., et al. Expression analysis of four long noncoding RNAs in breast cancer. Tumour biology : the journal of the International Society for Oncodevelopmental Biology and Medicine. 2016;37(3):2933–2940. doi: 10.1007/s13277-015-4135-2. [DOI] [PubMed] [Google Scholar]
- 95.Han L., Zhang W., Zhang B., Zhan L. Long non-coding RNA SOX2OT promotes cell proliferation and motility in human ovarian cancer. Experimental and Therapeutic Medicine. 2018;15(2):2182–2188. doi: 10.3892/etm.2017.5618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Feng L., Wang R., Lian M., et al. Integrated analysis of long noncoding RNA and mRNA expression profile in advanced laryngeal squamous cell carcinoma. PLoS One. 2016;11(12, article e0169232) doi: 10.1371/journal.pone.0169232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Liu S., Xu B., Yan D. Enhanced expression of long non-coding RNA Sox2ot promoted cell proliferation and motility in colorectal cancer. Minerva Medica. 2016;107(5):279–286. [PubMed] [Google Scholar]