

Abstract
Operant renewal is a form of relapse in which a previously extinguished response recurs due to a change in context. We designed two experiments to examine the impact of differential reinforcement of alternative behavior on ABA renewal in a translational model of relapse with 12 children. We compared levels of renewal in 2, 3-phase arrangements. In one arrangement, we reinforced target responding in Context A, extinguished responding in Context B, and returned to Context A while continuing to implement extinction. In a second arrangement, an alternative response produced reinforcement in Context B and during the return to Context A. Results across the two experiments indicated 3 general findings. First, extinction plus differential reinforcement disrupted target behavior more consistently in Context B relative to extinction alone. Second, renewal tended to be greater and more persistent during extinction alone relative to extinction plus differential reinforcement. Third, the renewal effect appeared to depend on whether the alternative response had a history of extinction in Context A. We discuss methodological implications for the treatment of severe destructive behavior.
Keywords: differential reinforcement, renewal, translational research, treatment relapse
Treatment relapse refers to the recurrence of a previously suppressed response when treatment conditions change (Pritchard, Hoerger, & Mace, 2014). One type of treatment relapse, called renewal, occurs following a change in stimulus context (see McConnell & Miller, 2014; Podlesnik, Kelley, Jimenez-Gomez, & Bouton, 2017, for recent reviews). The term “context” can describe any aspect of the environment that influences behavior, such as physical stimuli, time, deprivation states, and recent experiences (Bouton, 1993; 2002; 2014). Experimenters typically investigate renewal in a three-phase test with respondent or operant behavior (Podlesnik et al., 2017). For instance, Bouton, Todd, Vurbic, and Winterbauer (2011) investigated operant renewal of rats’ lever pressing maintained by food in three renewal tests. In the ABA test, experimenters first reinforced lever presses according to a variable-interval (VI) 30-s schedule in Context A. Then, lever presses were extinguished in Context B. Finally, the rats were returned to Context A while extinction continued. The experimental procedures in the ABC test were like those in the ABA test, except the experimenters tested for renewal in a new context (Context C) after extinction in Context B. In the third test, Bouton et al. reinforced and then extinguished lever pressing in Context A before testing for renewal in a new context (Context B), producing an AAB test. Renewal occurred in all three tests, despite the continuation of extinction for target responding in each.
Several variables impact renewal, including (a) rate of reinforcement for the target response in baseline (Berry, Sweeney, & Odum, 2014; Podlesnik & Shahan, 2009), (b) length of reinforcement history for the target response (Todd, Winterbauer, & Bouton, 2012), (c) rate of target responding in baseline (see discussion of Bouton et al., 2011 and Podlesnik & Shahan, 2009, in Podlesnik et al., 2017), and (d) similarity of reinforcement and extinction contexts (Todd et al., 2012). Little is known, however, about the conditions under which renewal occurs in applied settings and with socially significant behavior.
Drawing on the findings from basic research conducted on operant renewal with nonhuman animals (e.g., Berry et al., 2014; Bouton et al., 2011; Nakajima, Tanaka, Urushihara, & Imada, 2000; Podlesnik & Shahan, 2009), Kelley, Liddon, Ribeiro, Greif, and Podlesnik (2015) demonstrated the generality of the renewal effect in a translational model of treatment relapse with two children diagnosed with autism spectrum disorder (ASD). Kelley et al. reinforced target responding (i.e., tracing or matching) in Context A, defined by a unique set of colored t-shirts, poster boards, and task stimuli. After responding stabilized in Context A, target responding was extinguished in Context B. However, Kelley et al. later observed renewal when they returned participants to Context A while extinction of target responding continued. These findings underscore the necessity of continued research on the variables that contribute to operant renewal in applied settings (see also Liddon, Kelley, Rey, Liggett, & Ribeiro, 2018).
As an example of renewal in an applied setting, a parent might reinforce destructive behavior during a caregiver-implemented functional analysis (Context A; see Kurtz, Fodstad, Huete, & Hagopian, 2013) before behavior analysts then initiate treatment (Context B). After treating destructive behavior, often with a combination of extinction and differential reinforcement of alternative behavior (DRA), behavior analysts typically reintroduce the caregiver into therapy sessions following caregiver training. Destructive behavior often returns upon reintroducing the caregiver (i.e., reinstating Context A), even though the caregiver may implement the treatment protocol with high fidelity (e.g., Ibañez, Piazza, & Peterson, 2019). That is, destructive behavior is susceptible to operant renewal, even when caregivers implement treatment procedures perfectly. Thus, it seems prudent to investigate the conditions under which operant ABA renewal occurs, specifically under conditions that mimic those found in practice (Sullivan, Saini, & Roane, 2018).
Despite the generality of renewal (Bernal-Gamboa, Nieto, & Uengoer, 2017; Kelley et al., 2015; Podlesnik & Miranda-Dukoski, 2015), the above hypothetical example highlights an apparent disconnect between current research and practice. Unlike most renewal tests evaluated to date, practitioners do not typically program extinction alone for destructive behavior; instead they either provide reinforcement on a time-based schedule (i.e., noncontingent reinforcement [NCR]; Phillips, Iannaccone, Rooker, & Hagopian, 2017; Vollmer, Iwata, Zarcone, Smith, & Mazaleski, 1993), or they differentially reinforce an alternative response (DRA, Petscher, Rey, & Bailey, 2009). To better understand common stimulus and reinforcement conditions that occasion the renewal of undesirable behavior, it is necessary to explore extensions of the standard, three-phase renewal tests (i.e., those that rely on extinction alone).
Two recent studies demonstrated the utility of investigating renewal when alternative reinforcement was available for clinically significant behavior. Kelley, Jimenez-Gomez, Podlesnik, and Morgan (2018) investigated the generality of a behavioral treatment for severe aggression exhibited by a child diagnosed with ASD. First, in a classroom setting (Context A), one therapist provided 20 s of escape from academic instructions following aggression. Next, in a padded treatment room (Context B), a second therapist placed aggression on extinction and differentially reinforced compliance by presenting a 20-s break. The researchers then returned to Context A while continuing to implement the treatment procedures. Kelley et al. observed renewal of aggression and decreases in compliance despite continued reinforcement of compliance. These findings suggest that simply changing the intervening therapist may engender renewal despite the continued availability of reinforcement for an alternative response.
Similarly, Saini, Sullivan, Baxter, DeRosa, and Roane (2018) recently demonstrated ABA renewal of severe destructive behavior despite differential reinforcement of functional communication responses (i.e., the alternative response in functional communication training). Functional communication training is a common DRA-based treatment that involves placing destructive behavior on extinction and differentially reinforcing a functionally equivalent communication response (Carr & Durand, 1985; Tiger, Hanley, & Bruzek, 2008). First, in the home context (Context A), caregivers reinforced destructive behavior on an fixed ratio (FR) 1 schedule. Next, in the clinic context (Context B), therapists implemented functional communication training. Finally, caregivers implemented treatment in the home context (Context A) after training on the correct implementation of the treatment procedures. Despite perfect treatment integrity from all caregivers, Saini et al. observed renewal of destructive behavior with three of the four participants. The collective findings from Kelley et al. (2018) and Saini et al. (2018) suggest that DRA may not weaken operant ABA renewal.
It is currently unclear how renewal during extinction alone differs from renewal during extinction plus DRA in regards to magnitude and persistence of the renewal effect. Also unclear is whether any potential differences between the two renewal preparations are the result of (a) the availability of an alternative response, (b) the alternative reinforcement contingency, (c) the availability of alternative reinforcers that may mitigate the effects of a change in other contextual variables, or (d) some combination of the above. Furthermore, the recent findings of Kelley et al. (2018) and Saini et al. (2018) suggest that further research is necessary to test the generality of renewal during extinction with and without DRA. The purpose of the present research was twofold. First, we wished to compare the magnitude of ABA renewal across two distinct arrangements characterized by the presence or absence of DRA in Context B and during the renewal test in Context A. The second purpose was to determine what role (if any) response competition played in our findings of Experiment 1 by including an inactive alternative (i.e., control) response in the renewal test for the extinction-alone arrangement in Experiments 2a and 2b. The primary distinction between Experiments 2a and 2b was whether the inactive alternative response was also present in baseline.
General Method
Participants, Setting, and Materials
Twelve individuals1, ranging in age from 2 to 8 years old (M = 4.25 years old), participated. Angel, James, Robert, David, Julian, and Eric participated in Experiment 1. Trevor, Peter, and Jean participated in Experiment 2a. Teon, Magnus, and Alison participated in Experiment 2b. All of the participants were diagnosed with ASD except for Alison and Angel. Alison was neurotypical, and Angel was neurotypical but had a language delay. David, Jean, and Magnus communicated using gestures, by pointing, or by using augmentative communication devices. James, Robert, Eric, Peter, and Teon communicated using two-to-four word phrases (e.g., “Toy, please”, “I want crayons, please”). Angel, Julian, Trevor, and Alison communicated using full sentences with occasional prompting. All individuals met our inclusion criteria by demonstrating (a) the ability to sit on the floor or in a chair at a table for at least 3 min while not engaging in destructive behavior (e.g., aggression or self-injury), (b) preference for one of several available toys, and (c) lack of response persistence in a response-maintenance test (see Preexperimental Assessments).
Experimental sessions occurred at a center-based treatment facility in padded therapy rooms and in cubicle work areas. During each phase of the study, we arranged additional colored stimuli in each room. Context-specific colored materials included either red or blue (a) shirts worn by the experimenter, (b) wristbands, (c) poster boards (0.9 m by 1.2 m), (d) placemats centered under the response materials, and (e) task materials. Room type and color of materials in the room constituted our programmed Contexts A and B across experiments. We counterbalanced room type and color of materials across participants. The experimenter remained the same across all phases for a given participant.
Session materials included (a) Montessori object permanence box(es) (see Liggett, Nastri, & Podlesnik, 2018, for description of similar operanda), (b) a single plastic ball that measured 3.2 cm in diameter, (c) furniture (e.g., table, chairs), (d) equipment for data collection, (e) context-specific colored stimuli, and (f) a highly preferred toy. The rooms did not include any other materials. Box 1 consisted of a square box and a return tray that measured 27.9 × 13.2 × 12.5 cm with a hole in the top of the box that measured 3.8 cm in diameter. Box 2 consisted of a rectangular box and a return tray that measured 29.0 × 14.0 × 12.0 cm with a hole in the top of the box that also measured 3.8 cm in diameter. We counterbalanced box assignment across participants such that Box 1 was the target for some participants but the alternative for other participants, and vice versa for Box 2.
Response Measurement and Reliability
We measured target and alternative responses (when applicable) when the participant released the ball in the hole of the designated object permanence box. After placing the ball through the hole, the ball returned to the tray automatically, creating a free-operant arrangement. Observers used DataPal (a beta version of BDataPro; Bullock, Fisher, & Hagopian, 2017) to record the frequency of responding.
We defined renewal as a rate of target responding during any session of the renewal test in Context A that was (a) higher than the rate of target responding during the last session in Context B and that (b) failed to maintain at least an 85% reduction in the mean response rate during the last five baseline sessions in Context A. For example, if target responding during the last session of Context B occurred at a rate of 0 responses per min but the mean response rate during the last five sessions of baseline in Context A was 4.0 responses per min, we scored an instance of renewal if target responding in any session of the renewal test in Context A exceeded 0.6 responses per min.
A second independent observer collected data with the primary data collector either simultaneously or through video recordings for at least 30% of sessions for all participants. We calculated interobserver agreement using an exact agreement method. That is, we divided each data collector’s response-measurement record into 10-s intervals, and we compared the number of responses scored during each interval. Next, we scored an agreement if both data collectors recorded the same frequency of responses during an interval. Finally, we divided the number of agreement intervals by the total number of intervals in the session and converted this quotient to a percentage. Table 1 displays interobserver agreement coefficients across participants.
Table 1.
Interobserver-Agreement Coefficients
| Participant | Target Responding | Alternative Responding |
|---|---|---|
| M (range) | M (range) | |
| Alison | 96% (88%–100%) | 94% (77%–100%) |
| Angel | 94% (80%–100%) | 97% (93%–100%) |
| David | 94% (80%–100%) | 97% (90%–100%) |
| Eric | 98% (93%–100%) | 96% (67%–100%) |
| James | 91% (67%–100%) | 92% (73%–100%) |
| Jean | 96% (93%–100%) | 94% (83%–100%) |
| Julian | 87% (76%–100%) | 98% (90%–100%) |
| Magnus | 97% (88%–100%) | 96% (83%–100%) |
| Peter | 97% (90%–100%) | 98% (90%–100%) |
| Robert | 93% (73%–100%) | 96% (73%–100%) |
| Teon | 97% (90%–100%) | 95% (86%–100%) |
| Trevor | 93% (83%–100%) | 94% (83%–100%) |
Preexperimental Assessments2
Preference assessment.
Experimenters conducted multiple-stimulus without replacement preference assessments (DeLeon & Iwata, 1996) to identify highly preferred stimuli to deliver as reinforcers for responding in experimental sessions. Each assessment included four to five toys, which we selected based on caregiver or therapist nomination. The highest-preference item was an iPad for David, Angel, James, Julian, Peter, Magnus, and Alison; a toy car set for Robert and Jean; a toy train set for Trevor; a dinosaur toy for Eric; and crayons for Teon. The experimenter also delivered attention with the high-preference item for James, Robert, Julian, Trevor, Jean, and Alison, as they preferred to play with the therapist.
Response-maintenance test.
We used analog tasks (i.e., Montessori object permanence boxes) with each participant to control for histories of reinforcement for target and alternative responding (Baron & Perone, 1998). We first tested for the maintenance of responding in the absence of programmed consequences to rule out the possibility that responding with the object permanence box was automatically reinforced (see Querim et al., 2013, for a similar screening tool). Experimenters briefly modeled the response (i.e., placing the ball in one of the two object permanence boxes, randomly assigned across participants for this test) and required the participant to imitate the response. Following the first independent response, the experimenter initiated a 30-min session in which he or she provided no programmed consequences for responding or any prompts. We terminated the response-maintenance test after 5 min with five or fewer responses. Responding averaged 17.1 responses (range, 1–82 responses) across participants, and the test lasted an average of 8.1 min (range, 5–16 min). Thus, responding did not maintain for any participant, suggesting that the analog task was not automatically reinforcing.
Experimental Design
Table 2 shows the stimulus and reinforcement conditions across phases of Experiment 1, Experiment 2a, and Experiment 2b. We used a within-subject, six-phase sequential design that incorporated two, three-phase ABA renewal tests (see Liddon et al., 2018, for a similar design). The three-phase renewal test is widely used to test for renewal (e.g., Bouton, 2002; Kelley et al., 2015; Nakajima et al., 2000; Podlesnik et al., 2017), and we supplemented this approach by conducting two independent renewal tests per participant. In the renewal test with extinction alone, we reinforced target responding in Context A, placed target responding on extinction in Context B, and tested for renewal by returning to Context A with extinction still in place. In the renewal test with extinction plus DRA, we reinforced target responding in Context A, placed target responding on extinction and differentially reinforced an alternative response in Context B, and we tested for renewal by returning to Context A without altering the contingencies previously in effect. That is, extinction continued for target responding, and differential reinforcement continued for the alternative response in this third phase. In both tests, we programmed ABA stimulus context changes but ABB reinforcement contingency changes. This design allowed us to isolate the effects of context change while the reinforcement contingencies remained the same (see Kelley et al., 2015, for discussion). We counterbalanced the order of the renewal tests across participants. We first exposed James, David, Eric, Trevor, Jean, Teon, and Alison to the renewal test with extinction alone and Angel, Robert, Julian, Peter, and Magnus to the renewal test with extinction plus DRA.
Table 2.
Stimulus and Reinforcement Conditions Across Phases of Experiments 1, 2a, 2b
| Experiment | Design | Context A | Context B | Context A (Renewal Test) |
|---|---|---|---|---|
| 1 | Extinction | Target – VR 2 | Target – EXT | Target – EXT |
| Extinction + DRA | Target – VR 2 | Target – EXT Alternative – VR 2 | Target – EXT Alternative – VR 2 | |
| 2a | Extinction | Target – VR 2 Alternative – EXT | Target – EXT Alternative – EXT | Target – EXT Alternative – EXT |
| Extinction + DRA | Target – VR 2 Alternative – EXT | Target – EXT Alternative – VR 2 | Target – EXT Alternative – VR 2 | |
| 2b | Extinction | Target – VR 2 | Target – EXT Alternative – EXT | Target – EXT Alternative – EXT |
| Extinction + DRA | Target – VR 2 | Target – EXT Alternative – VR 2 | Target – EXT Alternative – VR 2 |
Additionally, we introduced all phase changes within days so that spontaneous recovery was not interpreted as renewal. All sessions of a given renewal test occurred on the same day for all participants except Julian. For Julian, we conducted the renewal test with extinction alone across multiple days due to persistent target responding. Within-participants, we yoked the number of sessions in Context B in the second renewal test to the number of sessions conducted in Context B in the first renewal test, thereby equating time in Context B across the two renewal tests for a given participant. We made similar attempts to yoke the number of sessions in Context A. We typically conducted four to five sessions per day.
Experiment 1
Procedure
Before each session, experimenters provided 30-s access to the highly preferred toy from the stimulus preference assessment. Experimenters presented one or two object permanence boxes, depending on the condition, plus a single plastic ball within arm’s reach of the participant. Sessions began when the therapist removed the highly preferred toy. Experimenters did not provide instructions to the participants in this or in any other experiment. Engagement with the toy during reinforcement intervals did not prevent the participant from responding. However, participants rarely responded during reinforcement intervals, and when they did, the experimenter ignored the response. During all phases with two response alternatives, the target and alternative boxes were separated by 0.46 m. After each reinforcer delivery, the experimenter placed the ball in the middle of the two object permanence boxes. Sessions lasted 5 min.
Baseline (Context A).
To establish the target response, we used a progressive prompt delay (0 s, 2 s, 5 s, 10 s; Charlop, Shreibman, & Thibodeau, 1985) to teach the participants to place the ball in the target box. Target responding produced 20-s access to the highly preferred toy on an FR 1 schedule. Experimenters terminated this training procedure after one 5-min session in which all target responses occurred independently. Once the target response was established for a given participant, these training procedures were not replicated upon subsequent returns to the baseline context. All prompts were removed from subsequent sessions.
Experimenters then initiated a variable-ratio (VR) 2 schedule for target responses. The VR 2 schedule included values of 1, 2, and 3 selected randomly and with replacement (Liggett et al., 2018). We used a VR 2 schedule for three reasons. First, descriptive data suggest that destructive behavior is likely maintained by variable schedules of reinforcement in the natural environment (Mace & Lalli, 1991; Thompson & Iwata, 2001). Second, dense schedules of reinforcement often produce more persistent behavior during extinction relative to lean schedules of reinforcement (Berry et al., 2014; Podlesnik & Shahan, 2009). Third, we wished to maintain target responding at relatively high response rates, which may influence levels of renewal (see discussion in Podlesnik et al., 2017). Baseline lasted at least 10 sessions to increase the likelihood of renewal (Todd et al., 2012).
Extinction (Context B).
Experimenters provided no programmed consequences for target responding. This phase continued until we observed (a) three consecutive sessions with target-response rates at zero or (b) the number of sessions in this phase matched the number of sessions conducted in extinction plus DRA in Context B when participants experienced that renewal test first.
Extinction (Context A).
Experimenters returned to Context A and continued to provide no programmed consequences for target responding for a minimum of three sessions.
Extinction plus DRA (Context B).
Experimenters provided no programmed consequences for target responding but provided reinforcement for alternative responding with access to the highly preferred toy on a VR 2 schedule to promote similar rates of reinforcement for the target and alternative responses (Trask & Bouton, 2016). This phase continued until we observed (a) three consecutive sessions with target-response rates at zero or (b) the number of sessions in this phase matched the number of sessions conducted in extinction of target responding in Context B when participants experienced that renewal test first.
Extinction plus DRA (Context A).
Experimenters provided no programmed consequences for target responding but provided reinforcement for alternative responding with access to the highly preferred toy on a VR 2 schedule. This phase lasted for at least three sessions for all participants except Teon in Experiment 2b, whose final phase ended prematurely due to scheduling conflicts.
Results and Discussion
Figure 1 depicts the results for Angel, James, and Robert. Figure 2 depicts the results for David, Julian, and Eric. We plotted the data in Figures 1 and 2 to show the counterbalanced order of renewal tests across participants. Target responding in baseline stabilized in Context A for all participants in the extinction alone renewal arrangement. Extinction alone in Context B decreased target responding to zero or near-zero rates for all participants. During the renewal test in the return to Context A with extinction alone, five of six participants (Angel, Robert, James, David, and Julian) displayed a recurrence of target behavior despite the continuation of extinction. Renewal of target responding occurred for only a single session for James during the test with extinction alone. Alternatively, renewal of target responding during extinction alone persisted for multiple sessions for Angel, Robert, David, and Julian. Julian’s target responding did not extinguish during the renewal test with extinction alone.
Figure 1.
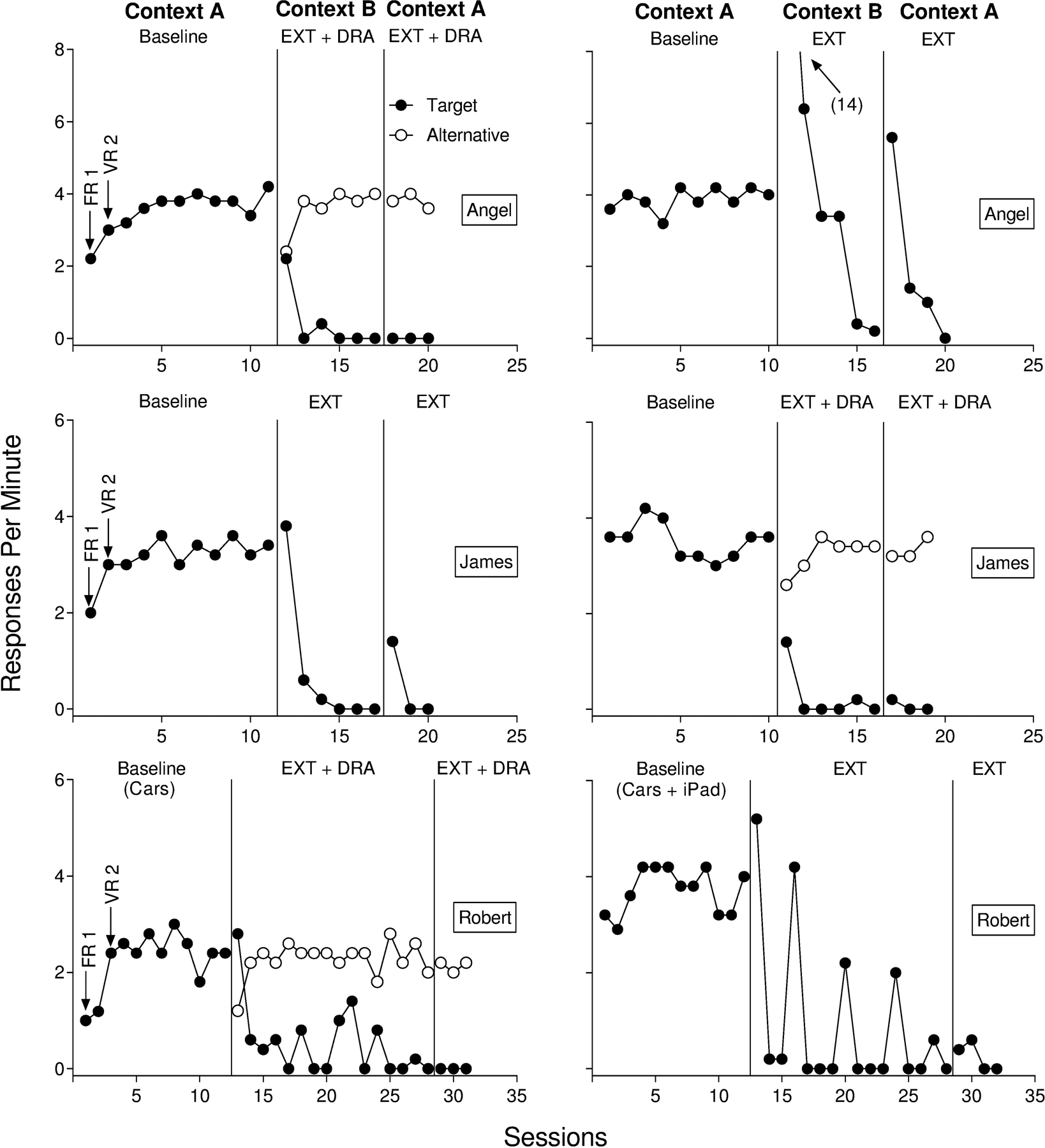
Target and alternative responding for Angel (top), James (middle), and Robert (bottom) from Experiment 1. Sessions 13, 16, 20, 24, and 27 for Robert in the ABA renewal test with extinction alone (bottom, right) were the first sessions of each day in Context B. Robert’s subsequent renewal test occurred on the third session of the day.
Figure 2.
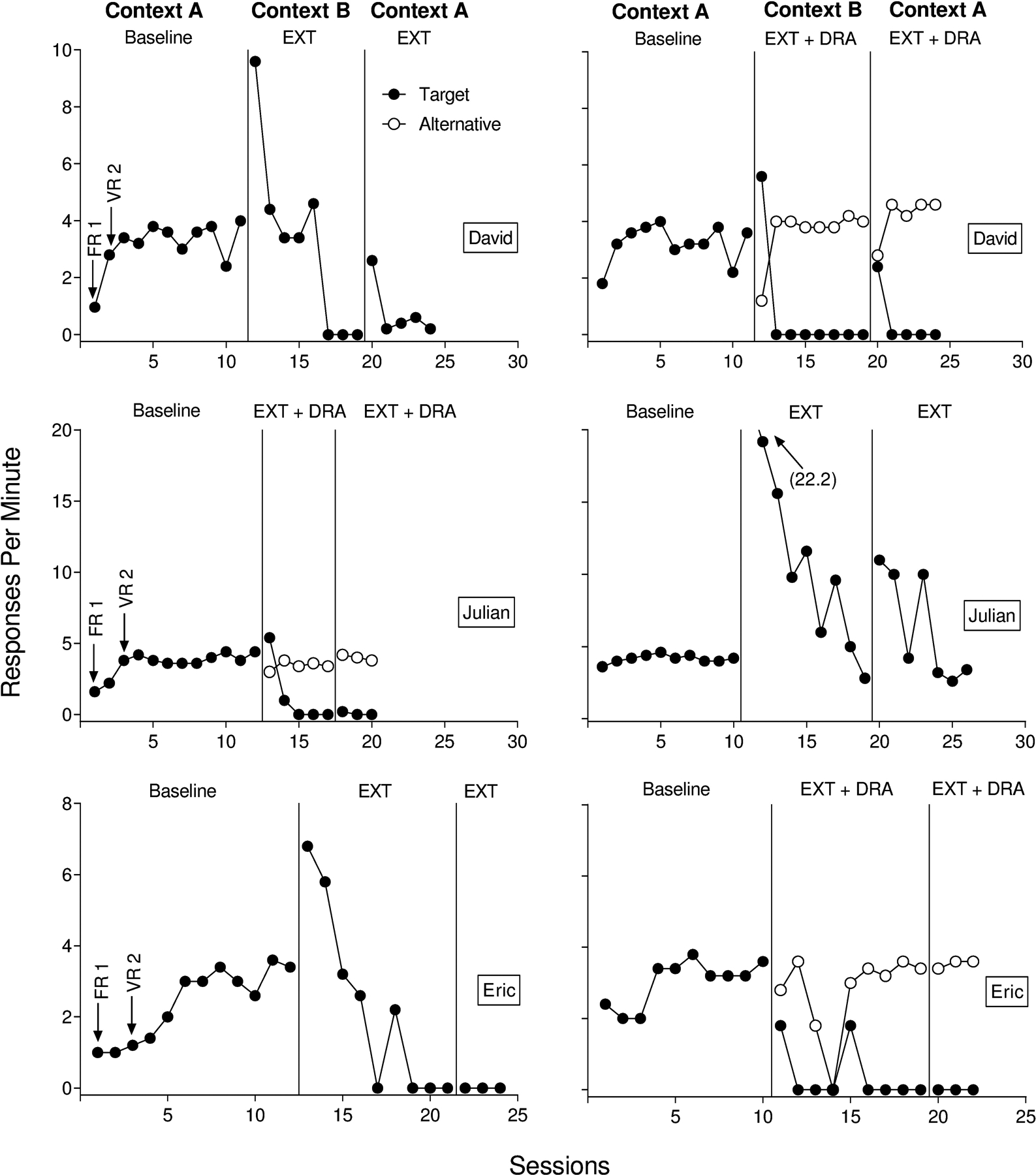
Target and alternative responding for David (top), Julian (middle), and Eric (bottom) from Experiment 1.
Target responding in baseline stabilized in Context A for all participants in the renewal test with extinction plus DRA. Rates of alternative responding were high and stable when we reinforced the alternative response in Context B, and target responding decreased to zero or near-zero rates. Unlike the renewal test with extinction alone in Context A, we observed renewal of target responding for only one of six participants (David) during the renewal test with extinction plus DRA in Context A. For David, renewal occurred in a single session (2.4 responses per min) before decreasing to zero.
In Experiment 1, we examined renewal when extinction alone was present during the renewal test in Context A (Kelley et al., 2015) and when extinction plus DRA was present during the renewal test in Context A. By and large, the findings from the present study are consistent with past research on ABA renewal. That is, a target response that was previously reinforced in Context A, and extinguished in Context B, recurred during the renewal test in Context A, despite the continuation of extinction (Bouton et al., 2011; Nakajima et al., 2000; Todd et al., 2012). These data replicate and extend the results of Kelley et al. (2015) and clarifythe conditions under which renewal occurs with human behavior (Bandarian Balooch & Neamann, 2011; Cohenour, Volkert, & Allen, 2018; Ibañez et al., 2019; Kelley et al., 2018; Nelson, Sanjuan, Vadillo-Ruiz, Perez, & Leon, 2011; Saini et al., 2018; Shiban, Pauli, & Muhlberger, 2013). Saini et al. (2018) found robust renewal effects when testing for renewal during extinction plus DRA. We did not replicate these findings. The data from Experiment 1 suggest that renewal may not be as robust when testing under extinction plus DRA relative to extinction alone.
Perhaps the most interesting finding from Experiment 1 was the difference in magnitudes of renewal between the two, three-phase renewal tests. We observed more renewal during extinction alone relative to tests with extinction plus DRA. One parsimonious interpretation is that the mere presence of a second response mitigated renewal during the renewal test with extinction plus DRA. That is, the availability of two response alternatives may have allowed the alternative response to compete with the target response (Kimball, Kelley, Podlesnik, Forton, & Hinkle, 2018; Podlesnik & Kelley, 2014), thus decreasing target responding relative to a condition in which only the target was available (i.e., extinction alone). In Experiment 1, however, response availability was inherently confounded with the presence of reinforcement for the alternative response (i.e., DRA). Countering this interpretation, however, we observed little to no decrement in alternative responding for Angel, James, Robert, Julian, and Eric when transitioning from Context B to Context A during the renewal test with extinction plus DRA, suggesting that alternative reinforcement and not the alternative response per se was more likely responsible for renewal mitigation in Experiment 1.
Experiment 2
We designed Experiment 2 to clarify the results from Experiment 1 concerning the number of responses available during each renewal test and the history of reinforcement for the alternative response in Context A.
Experiment 2a
In this experiment, we included an inactive alternative response that did not produce reinforcement across all phases of the renewal test with extinction alone. We also introduced the inactive alternative response during baseline in Context A of the renewal test with extinction plus DRA (see Table 2). An added benefit of this arrangement was that returning to Context A in both renewal tests reinstated the identical stimulus conditions present in Context A during baseline, as alternative response availability did not differ across phases within Context A as it had for extinction plus DRA in Experiment 1.
Procedure.
All experimental procedures were identical to those from Experiment 1, except as noted.
Baseline (Context A).
We delivered 20-s access to a highly preferred toy on an FR 1 schedule to establish target responding. An alternative response box was present, but placing the ball in the hole of the alternative response box did not produce reinforcement. Once the target response was established, experimenters provided 20-s access to a highly preferred toy on a VR 2 schedule for target responding. Alternative responding continued to result in extinction.
Extinction (Context B).
Experimenters provided no programmed consequences for target or alternative responding.
Extinction (Context A).
Experimenters returned to Context A and continued to provide no programmed consequences for target or alternative responding.
Extinction plus DRA (Context B).
Experimenters provided reinforcement for alternative responding with access to a highly preferred toy on a VR 2 schedule and placed target responding on extinction.
Extinction plus DRA (Context A).
Experimenters provided reinforcement for alternative responding with access to a highly preferred toy on a VR 2 schedule, and extinction continued for target responding.
Results and Discussion.
Responses per min for Trevor, Peter, and Jean from Experiment 2a are displayed in Figure 3. In the renewal test with extinction alone, reinforcement of the target response in Context A during baseline increased target responding to high and steady rates, and each participant engaged in only minimal alternative responses. Extinction of target responding in Context B decreased target responding to zero for all participants. Alternative responding remained low or decreased to zero across participants in Context B. We observed renewal for all three participants in the renewal test with extinction alone. For Trevor and Peter, target responding in the renewal test persisted for more than one session, but we observed renewal for only a single session with Jean in the extinction-alone test. Peter and Jean did not engage in alternative responding during the renewal test with extinction alone. In contrast, Trevor’s alternative responding increased during the renewal test with extinction alone but decreased to near zero rates across sessions.
Figure 3.
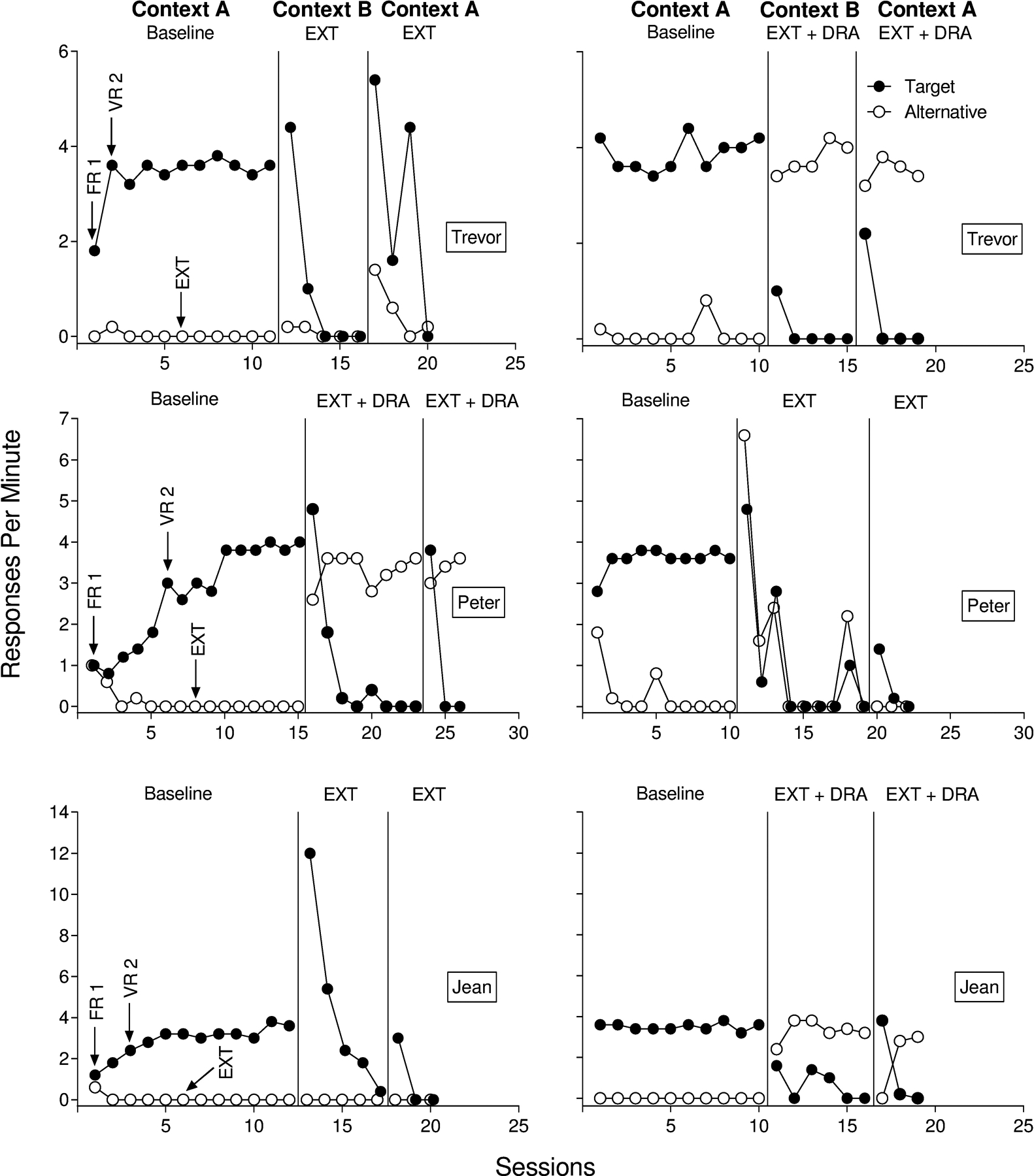
Target and alternative responding for Trevor (top), Peter (middle), and Jean (bottom) from Experiment 2a.
Baseline in Context A increased target responding to steady rates, and extinction for the inactive alternative response resulted in minimal alternative responding in the renewal test with extinction plus DRA. Differential reinforcement of alternative responding in Context B produced high and steady rates of alternative responding and low rates of target responding for all participants. Interestingly, renewal occurred for all three participants in the renewal test with extinction plus DRA in Experiment 2a, in sharp contrast to the results for Experiment 1. Recall that only one of six participants showed renewal in the renewal test with extinction plus DRA in Experiment 1. Moreover, Peter and Jean showed more pronounced renewal in the renewal test with extinction plus DRA than in the renewal test with extinction alone. Trevor and Peter showed renewal during only a single session before target responding decreased to zero responses per minute in the extinction plus DRA test. In contrast, renewal for Jean persisted for two sessions before decreasing to zero responses per minute in the same test condition.
One possible explanation for the difference in magnitudes of renewal in the renewal test with extinction plus DRA across Experiments 1 and 2a involves the different histories with the alternative response in baseline across the two experiments. In Experiment 1, the alternative response was unavailable in baseline in Context A, whereas in Experiment 2a, the alternative response was available but placed on extinction in this same condition. Thus, for participants in Experiment 2a, it is possible that Context A signaled not only the availability of reinforcement for the target response but the unavailability of reinforcement for the alternative response. This dual discrimination in the presence of Context A may have increased the likelihood of renewal when returning to Context A during the renewal test with extinction plus DRA in Experiment 2a. Such a discrimination would have been unlikely to occur and affect renewal in Experiment 1. This hypothesis may explain why Jean’s rate of alternative responding (3.2 responses per min) during the last session in Context B dropped to a rate of 0 during the first session of the renewal test with extinction plus DRA despite the continued availability of differential reinforcement.
Experiment 2b
In this experiment, we replicated the procedures of Experiment 2a, except the baseline condition of both renewal tests did not include the alternative response. We did this to more closely replicate the baseline procedures in Experiment 1 while hoping to address the question of whether response competition mitigated renewal in that experiment.
Procedure.
The inactive alternative response was present only in Context B and during the return to Context A for the renewal test with extinction alone. The alternative response and its associated reinforcement contingency remained in place in Context B and during the return to Context A for the renewal test with extinction plus DRA. Sessions lasted 3 min for Magnus and Alison to accommodate shortened appointment times for Magnus and Alison’s younger age of 2 years old.
Results and Discussion.
Responses per min for Teon, Magnus, and Alison from Experiment 2b are displayed in Figure 4. Similar to Experiments 1 and 2a, baseline in Context A produced steady response rates for all participants. Extinction plus DRA in Context B produced more rapid decreases in target responding relative to extinction alone in Context B. Alternative responding increased to a steady state for all three participants in Context B during extinction plus DRA. Alternative responding decreased or remained low in Context B during extinction alone. We observed robust renewal for all three participants in the renewal test with extinction alone, and renewal persisted for more than one session for each participant. On the contrary, we only observed minimal renewal for one of three participants in the renewal test with extinction plus DRA. Alison engaged in only two target responses during the renewal test with extinction plus DRA. All participants engaged in few alternative responses during the renewal test with extinction alone.
Figure 4.
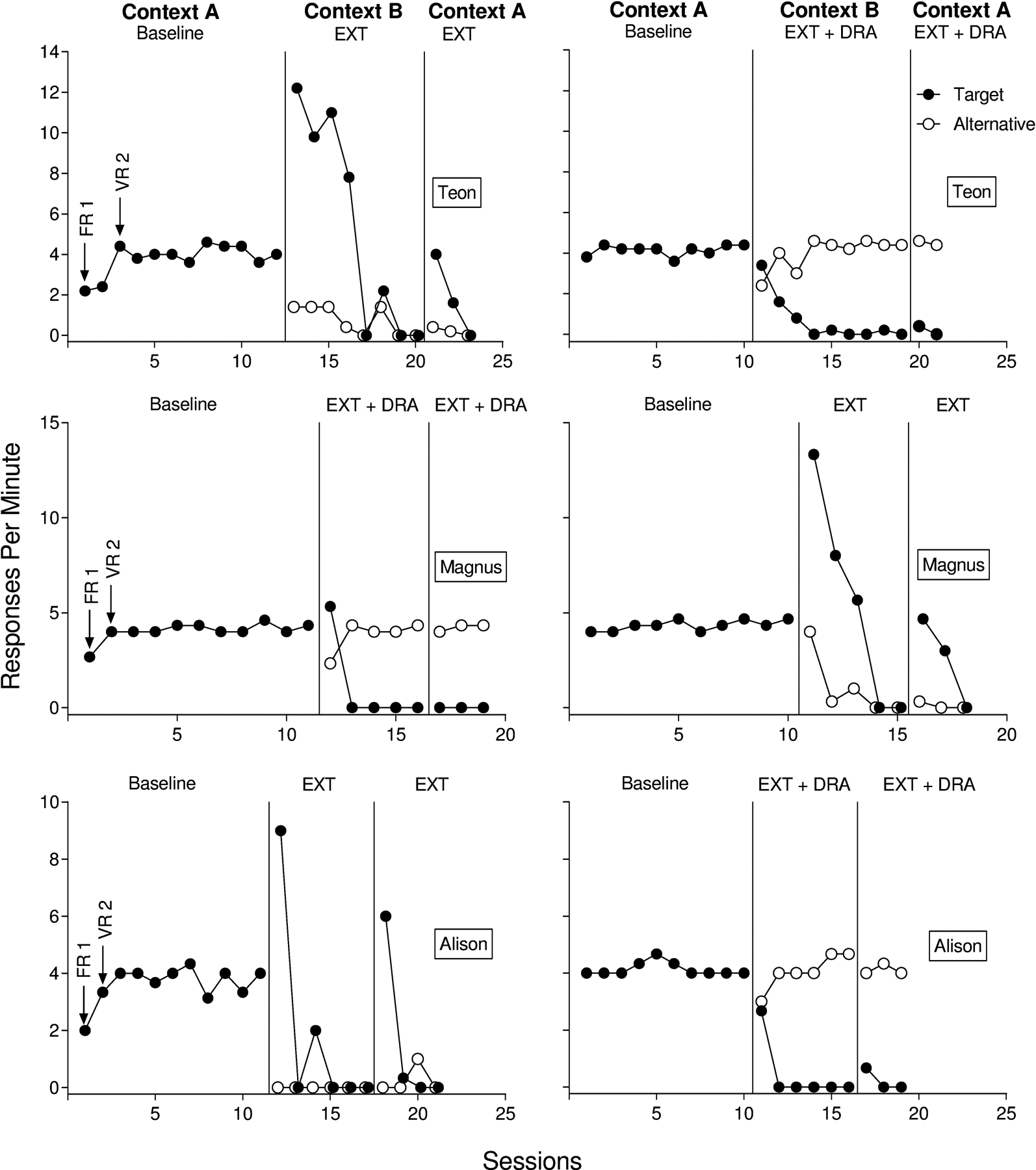
Target and alternative responding for Teon (top), Magnus (middle), and Alison (bottom) from Experiment 2b.
To compare our findings from Experiment 2b to those of Experiments 1 and 2a, we calculated summary data for each experiment. Figure 5 displays mean target responses in Context B during both extinction alone and extinction plus DRA for all participants across Experiments 1, 2a, and 2b. For Julian and other participants for whom we conducted an unequal number of sessions in Context B across the two renewal tests, we analyzed the final sessions in Context B common to both renewal tests, excluding earlier sessions when necessary. Mean target response rates in Context B tended to be lower with extinction plus DRA than with extinction alone across the three experiments. These data are consistent with past findings demonstrating superior suppression of target responding with extinction plus DRA relative to extinction alone (Lowry & Lachter, 1977; Rescorla & Skucy, 1969).
Figure 5.
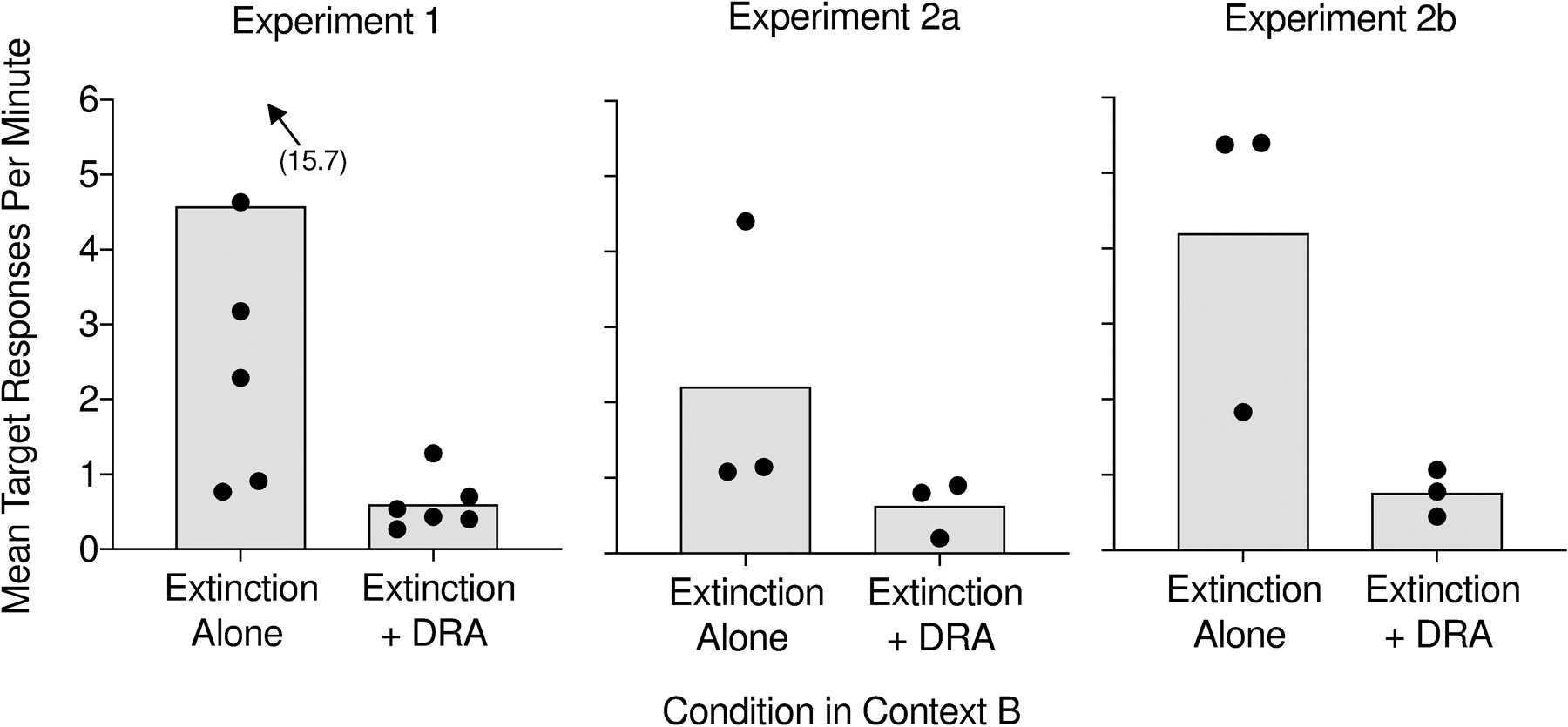
Bars depict mean rates of target responding across participants during Context B in Experiments 1, 2a, and 2b. The data points represent mean response rates for each participant.
Figure 6 displays mean target response rates during renewal tests with extinction alone and renewal tests with extinction plus DRA across participants in Experiments 1, 2a, and 2b. Similar to Figure 5, we analyzed the first sessions during the renewal tests common to both renewal tests, excluding later sessions when necessary. Mean target response rates during renewal tests tended to be lower with extinction plus DRA than with extinction alone in Experiments 1 and 2b, but this difference was much less pronouced in Experiment 2a.
Figure 6.
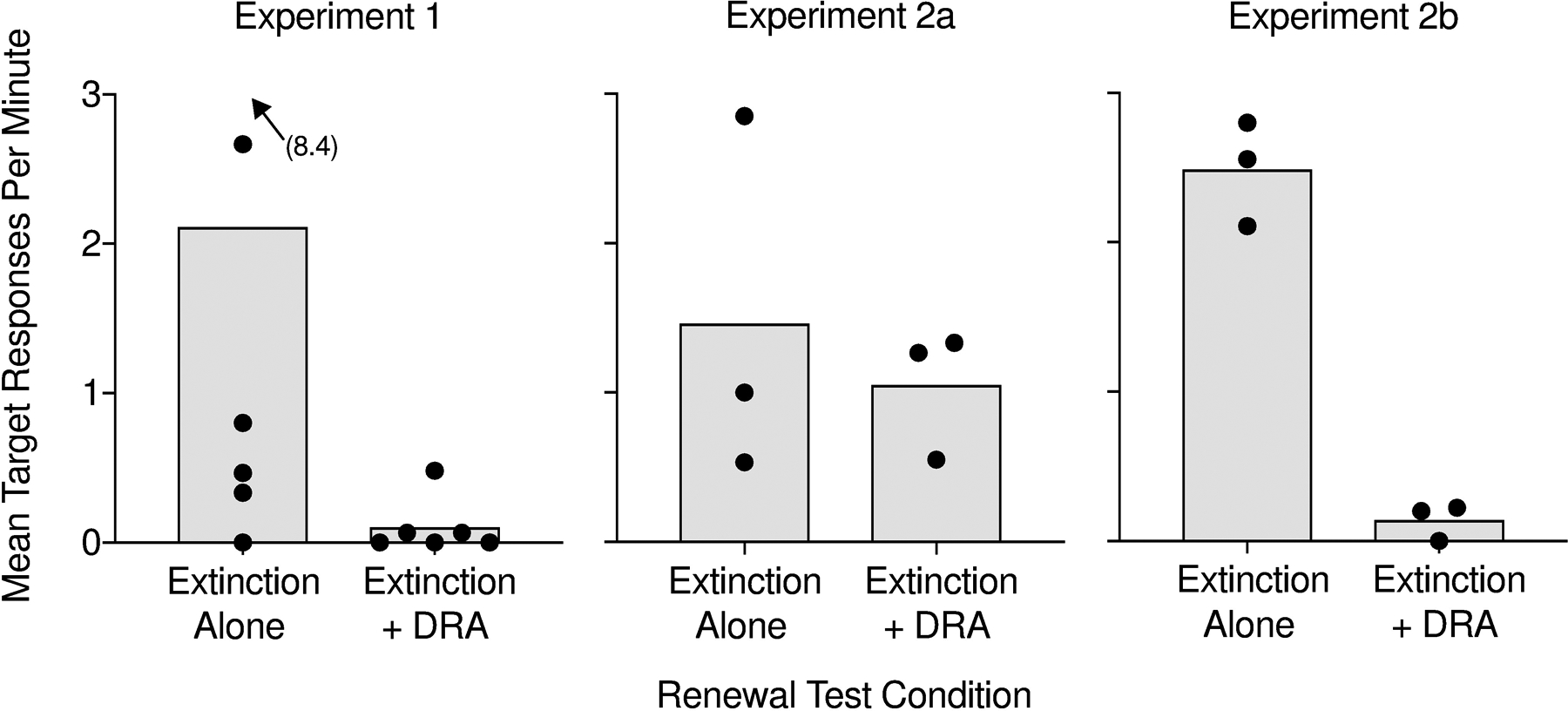
Bars depict mean rates of target responding across participants during the renewal tests in Experiments 1, 2a, and 2b. The data points represent mean response rates for each participant.
To provide a more formal analysis of the present outcomes, we also compared renewal between extinction alone and extinction plus DRA on a session-by-session level. Specifically, we paired consecutive sessions of the renewal tests within participant (e.g., Angel’s first session of the extinction alone renewal test was paired with his first session of the extinction plus DRA renewal test). We then calculated the proportion of such session pairs in which renewal-test response rates were higher in the extinction alone condition than in the extinction plus DRA condition. Pairs of sessions that did not include target responding in either condition were not included in this analysis. Renewal was greater during extinction alone in 92% (13/14) of paired sessions relative to extinction plus DRA in Experiment 1, 71% (5/7) of paired sessions in Experiment 2a, and 100% (6/6) of paired sessions in Experiment 2b. We conducted paired-sign tests using the binomial distribution to compare these empirical proportions against the null hypothesis that prenewal = .5 (i.e., a 50% chance of one session of the pair producing a higher response rate than the other session of the pair). In Experiments 1 (p < .001) and 2b (p = .015), target response rates during the renewal test were reliably greater with extinction alone relative to extinction plus DRA. In Experiment 2a, response rate differences were not significant (p = .164), suggesting that extinction of alternative responding in Context A during baseline increased renewal during extinction plus DRA.
In addition to examining difference in the magnitude of renewal across experiments, we also analyzed renewal effects in terms of the persistence of behavior. Figure 7 displays the mean number of consecutive sessions with target responding during each renewal test across participants and experiments. When participants exhibited renewal during extinction plus DRA, renewal tended to occur for only a single session. Alternatively, renewal of target responding during extinction alone often persisted for multiple sessions. Renewal during extinction plus DRA tended to be a more transient phenomenon.
Figure 7.
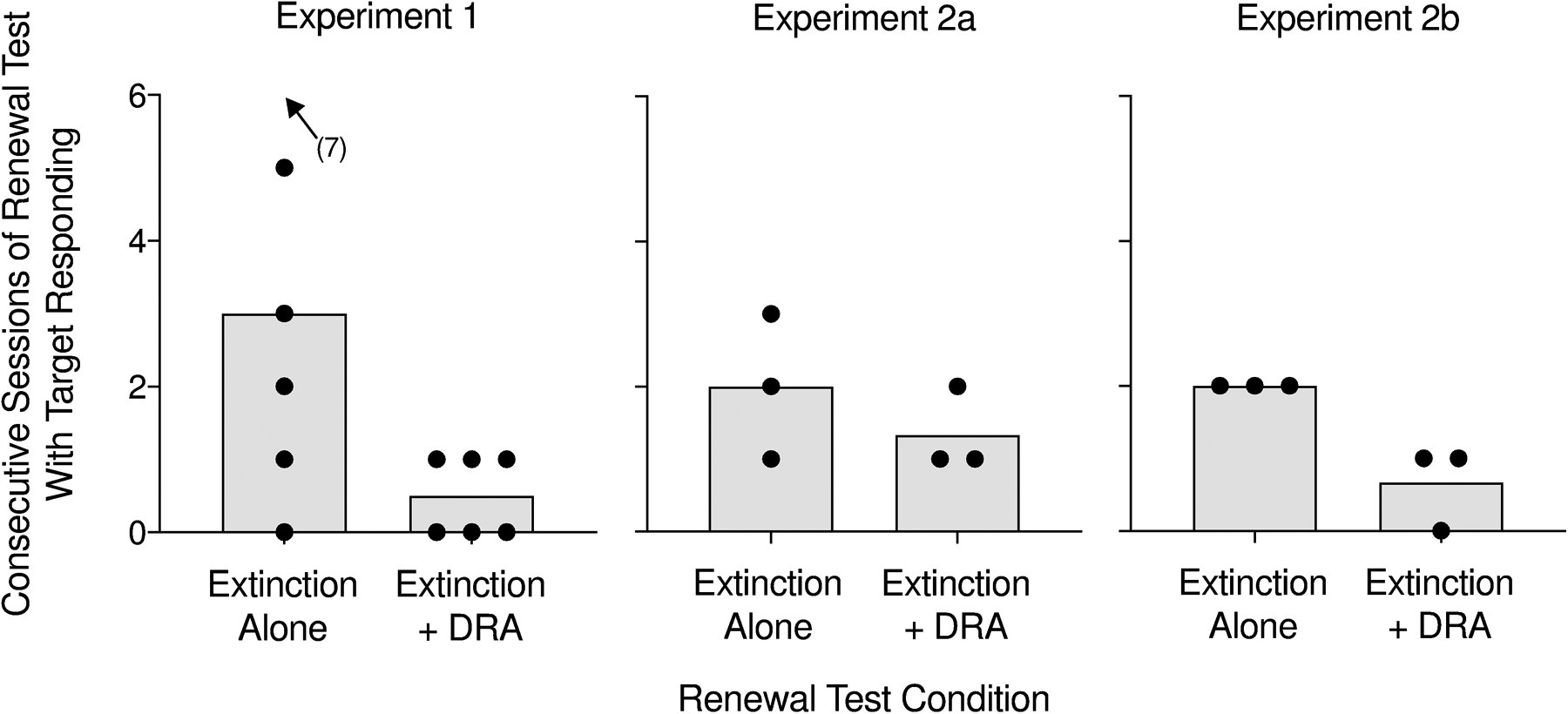
Bars depict mean consecutive sessions with target responding across participants during the renewal tests in Experiments 1, 2a, and 2b. The data points represent mean consecutive sessions with target responding for each participant.
Finally, Figure 8 displays mean alternative responses per min during each renewal test across experiments. For Figure 8, we analyzed the data in the same manner as for Figure 6 but did so for alternative responding instead of target responding. We observed lower rates of alternative responding across Experiments 2a and 2b during renewal tests with extinction alone relative to renewal tests with extinction plus DRA, suggesting that alternative reinforcement was necessary to increase alternative response rates during the renewal test.
Figure 8.
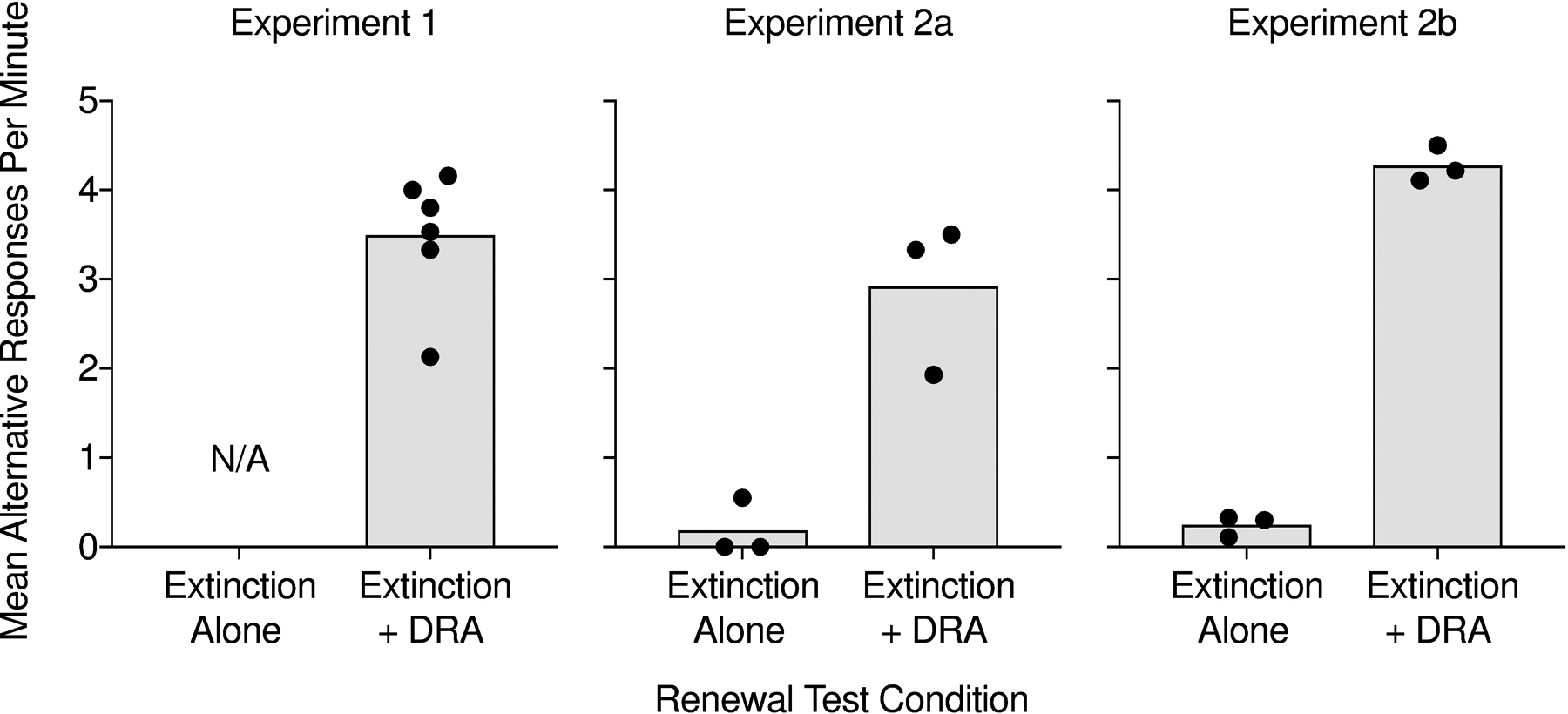
Bars depict mean rates of alternative responding across participants during the renewal tests in Experiments 1, 2a, and 2b. The data points represent mean response rates for each participant.
General Discussion
We compared renewal of target responding when differential reinforcement was present or absent during an ABA renewal test. The results of the present research yielded several general findings. First, extinction plus differential reinforcement disrupted target behavior more consistently in Context B relative to extinction alone. Second, renewal tended to be greater and more persistent during extinction alone relative to extinction plus differential reinforcement. Third, the renewal effect appeared to depend on whether the alternative response had a history of extinction in Context A. Repeated exposure to extinction and renewal tests did not seem to impact the magnitude or persistence of the renewal effect.
We observed more robust renewal during extinction plus DRA when the alternative response had a history of extinction in baseline than when the alternative response was absent in baseline. This across-experiment observation is hampered somewhat by the small number of individuals who participated in Experiments 2a and 2b. Nevertheless, future research on how histories of extinction for alternative behavior affect relapse seems warranted. Results from such investigations are likely to be of interest to applied behavior analysts who are often tasked with selecting an alternative response to reinforce during DRA-based interventions for problem behavior.
We hypothesized that differences in renewal in Experiment 1 during extinction alone and extinction plus DRA might have been due to the different number of response alternatives programmed across the two renewal tests. That is, during the renewal test with extinction plus DRA, the mere presence of a second response option may have occasioned distributed responding between the two responses (i.e., response competition). However, the results from Experiments 2a and 2b suggest that this was likely not the case. All six participants across Experiments 2a and 2b demonstrated renewal during the renewal test with extinction alone (Figures 3 and 4), replicating our findings from Experiment 1 (Figures 1 and 2). Additionally, target responding exceeded alternative responding during renewal tests with extinction alone for all participants across Experiments 2a and 2b, suggesting that the mere presence of an alternative response did little to supress the renewal effect.
A clear alternative explanation for the lack of robust renewal during extinction plus DRA in Experiments 1 and 2b is that the dense VR 2 schedule of reinforcement for alternative responding mitigated renewal. Still, with small sample sizes, it is somewhat difficult to compare results across the experiments. Nevertheless, we hypothesize that access to the reinforcer via the contingency for the alternative response competed with the stimulus control exerted by Context A. With a dense VR 2 schedule of reinforcement for the alterative response, participants spent most of the session in the reinforcement interval and very little time without access to reinforcement during renewal tests with extinction plus DRA. In contrast, during the extinction alone renewal tests, participants did not access reinforcement. Parametric analyses of alternative reinforcement rates appear to be clear next steps for testing this hypothesis. Renewal effects may be greater during extinction plus DRA as the schedule of alternative reinforcement becomes leaner. It should also be noted that applied behavior analysts often thin DRA schedules when treating problem behavior (Greer, Fisher, Saini, Owen, & Jones, 2016; Hagopian, Boelter, & Jarmolowicz, 2011). Thus, renewal tests under lean DRA schedules may have greater external validity than renewal tests under dense DRA schedules.
It should be noted that our procedures were incapable of determining whether the DRA contingency was necessary to mitigate renewal or if the reinforcer in the absence of the contingency (i.e., NCR) would have had similar suppresive effects. Results from Trask and Bouton (2016) provided some evidence that NCR alone impacts ABA renewal. In Experiment 1, rats’ lever presses produced one type of edible item (e.g., grain pellets) on a VI 30-s schedule in Context A. Next, experimenters placed lever pressing on extinction and concurrently introduced response-independent delivery of a second edible item (e.g., sucrose pellets) on a random-time (RT) 30-s schedule in Context B. The experimenters tested for renewal of lever pressing in Context A under two conditions. In one condition, extinction continued for lever pressing, as did the RT 30-s schedule with the second edible item. In the second condition, extinction continued for lever pressing, and the RT 30-s schedule was discontinued. Trask and Bouton concluded that response-independent reinforcement attenuated renewal in the return to Context A. One question worthy of future study is whether simply introducing reinforcers with or without a response-dependent contingency during a renewal test would have similar effects or whether renewal mitigation in the present study and that of Trask and Bouton was achieved by programming common stimuli across the otherwise distinct contexts.
The continued availability of an alternative response, its associated contingency, or the reinforcers produced as a result of satisfying that contingency may contribute in part or in combination to the renewal effect. In this regard, some of our experimental procedures arranged renewal tests more analogous to ABB renewal than ABA renewal. What role each of these potential variables played in mitigating the renewal effect is unclear. Furthermore, whether these potential variables neutralized the programmed differences across contexts of the renewal test moreso than they disrupted target responding by way of their direct, operant effects on choice behavior also remains unanswered.
One limitation of the present study is that our laboratory model of ABA renewal may not reflect the dynamic stimulus and reinforcement conditions that are likely to occur in the natural environment. That is, we programmed only a limited number of context changes in our set of experiments, but during clinical practice, patients in intensive-outpatient and day-treatment programs return to Context A each day after appointments in the clinic (Sullivan et al., 2018).
A second limitation might be our ability to compare renewal effects following two different independent variable manipulations. We found that extinction plus DRA was more effective at decreasing and suppressing target responding in Context B than extinction alone. Thus, some may find it difficult to compare renewal effects following extinction plus DRA and extinction alone. To circumvent this issue, researchers investigating resistance to extinction and relapse-related phenomena often transform absolute response rates to proportions of baseline to compare behavior equitably during test conditions. Proportion of baseline, a relative measure, allows for the identification of levels of responding during extinction that meet or exceed the levels of responding during baseline (Cançado, Abreu-Rodrigues, & Alo, 2016). However, in the case of our experiments, proportion of baseline was an unnecessary transformation because baseline responding did not differ substantially within any participant. Whenever possible, we suggest that future researchers use strict criteria for changing phases when comparing renewal following two different procedures.
A third limitation of the study is the topographical similarity between the target and alternative responses. In practice, response topographies nearly always differ for the target and alternative response. Their similarity in the present study differs in this regard. In future translational research, experimenters may consider evaluating renewal during extinction plus DRA using two topographically distinct responses (e.g., an object-permanence box and a card exchange).
We provided access to a single ball during sessions, which some participants played with during phases of extinction, but this behavior occurred infrequently. However, our decision to provide a single ball eliminated the opportunity for participants to engage simultaneously in the target and the alternative response (see also, Liggett et al., 2018). In clinical settings, it is often possible for an individual to engage in both responses simultaneously. This procedural distinction could impact the degree to which our findings are generalizable to clinical practice.
A clinical implication from our research stems from the disparate findings of Experiments 2a and 2b. We found that levels of renewal during extinction plus DRA seemed to depend on whether a history of extinction for the alternative response occurred in Context A during baseline. A common recommendation is for practitioners to select alternative responses to destructive behavior based partly on those that already appear in the individual’s repertoire (Tiger et al., 2008). Our data suggest that practitioners should be cognizant of the histories of extinction they arrange (and the contexts in which they arrange them) for such responses, as this may have direct bearing on susceptibility to treatment relapse.
The present study fills a gap in the literature in the form of comparing renewal during extinction alone and renewal during extinction plus DRA. Practitioners rarely recommend extinction alone as a treatment for problem behavior. Studying renewal during extinction plus DRA has the advantage of more closely simulating the stimulus and reinforcement conditions that occur in practice. The present study, as well as recent studies by Kelley et al. (2018) and Saini et al. (2018), represent first steps toward a better understanding of renewal in the natural environment.
Acknowledgments
This research was conducted in partial fulfillment of the first author’s doctoral degree from the University of Nebraska Medical Center and was funded in part by grants 5R01HD079113, 5R01HD083214, and 1R01HD093734 from The National Institute of Child Health and Human Development.
Footnotes
All participant names are pseudonyms.
Data available upon request.
References
- Bandarian Balooch S, & Neumann DL (2011). Effects of multiple contexts and context similarity on the renewal of extinguished conditioned behaviour in an ABA design with humans. Learning and Motivation, 42, 53–63. doi: 10.1016/j.lmot.2010.08.008 [DOI] [Google Scholar]
- Baron A, & Perone M (1998). Experimental design and analysis in the laboratory study of human operant behavior In Lattal KA, & Perone M (Eds.), Handbook of research methods in human operant behavior (pp. 45–91). New York, NY, US: Plenum Press. doi: 10.1007/978-1-4899-1947-2_3 [DOI] [Google Scholar]
- Bernal-Gamboa R, Nieto J, & Uengoer M (2017). Effects of extinction in multiple contexts on renewal of instrumental responses. Behavioural Processes, 14, 264–269. doi: 10.1016/j.beproc.2017.06.003 [DOI] [PubMed] [Google Scholar]
- Berry MS, Sweeney MM, & Odum AL (2014). Effects of baseline reinforcement rate on operant ABA and ABC renewal. Behavioural Processes, 108, 87–93. doi: 10.1016/j.beproc.2014.09.009 [DOI] [PubMed] [Google Scholar]
- Bouton ME (1993). Context, time, and memory retrieval in the interference paradigms of Pavlovian learning. Psychological Bulletin, 114, 80–99. doi: 10.1037/0033-2909.114.1.80 [DOI] [PubMed] [Google Scholar]
- Bouton ME (2002). Context, ambiguity, and unlearning: Sources of relapse after behavioral extinction. Biological Psychiatry, 52, 976–986. doi: 10.1016/S0006-3223(02)01546-9 [DOI] [PubMed] [Google Scholar]
- Bouton ME (2014). Why behavior change is difficult to sustain. Preventive Medicine: An International Journal Devoted to Practice and Theory, 68, 29–36. doi: 10.1016/j.ypmed.2014.06.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouton ME, Todd TP, Vurbic D, & Winterbauer NE (2011). Renewal after the extinction of free operant behavior. Learning & Behavior, 39, 57–67. doi: 10.3758/s13420-011-0018-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bullock CE, Fisher WW, & Hagopian LP (2017). Description and validation of a computerized behavioral data program: “BDataPro.” The Behavior Analyst, 40, 275–285. doi: 10.1007/s40614-016-0079-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cançado CR, Abreu-Rodrigues J, & Aló RM (2016). A note on measuring recurrence. Revista Mexicana de Análisis de la Conducta, 42, 75–86. [Google Scholar]
- Carr EG, & Durand VM (1985). Reducing behavior problems through functional communication training. Journal of Applied Behavior Analysis, 18, 111–126. doi: 10.1901/jaba.1985.18-111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charlop MH, Schreibman L, & Thibodeau MG (1985). Increasing spontaneous verbal responding in autistic children using a time delay procedure. Journal of Applied Behavior Analysis, 18, 155–166. doi: 10.1901/jaba.1985.18-155 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohenour JM, Volkert VM, & Allen KD (2018). An experimental demonstration of AAB renewal in children with autism spectrum disorder. Journal of the Experimental Analysis of Behavior, 110, 63–73. doi: 10.1002/jeab.443 [DOI] [PubMed] [Google Scholar]
- DeLeon IG, & Iwata BA (1996). Evaluation of a multiple-stimulus presentation format for assessing reinforcer preferences. Journal of Applied Behavior Analysis, 29, 519–533. doi: 10.1901/jaba.1996.29-519 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greer BD, Fisher WW, Saini V, Owen TM, & Jones JK, (2016). Improving functional communication training during reinforcement schedule thinning: An analysis of 25 applications. Journal of Applied Behavior Analysis, 49, 105–121. doi: 10.1002/jaba.265 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagopian LP, Boelter EW, & Jarmolowicz DP (2011). Reinforcement schedule thinning following functional communication training: Review and recommendations. Behavior Analysis in Practice, 4, 4–16. doi: 10.1007/BF03391770 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ibañez VF, Piazza CC, & Peterson KM (2019). A translational evaluation of renewal of inappropriate mealtime behavior. Journal of Applied Behavior Analysis, 52, 1005–1020. doi: 10.1002/jaba.647 [DOI] [PubMed] [Google Scholar]
- Kelley ME, Jimenez-Gomez C, Podlesnik CA, & Morgan A (2018). Evaluation of renewal mitigation of negatively reinforced socially significant operant behavior. Learning and Motivation, 63, 133–141. doi: 10.1016/j.lmot.2018.05.003 [DOI] [Google Scholar]
- Kelley ME, Liddon CL, Ribeiro A, Greif AE, & Podlesnik CA (2015). Basic and translational evaluation of renewal of operant responding. Journal of Applied Behavior Analysis, 48, 390–401. doi: 10.1002/jaba.209 [DOI] [PubMed] [Google Scholar]
- Kimball RT, Kelley ME, Podlesnik CA, Forton A, & Hinkle B (2018). Resurgence with and without an alternative response. Journal of Applied Behavior Analysis, 51, 854–865. doi: 10.1002/jaba.466 [DOI] [PubMed] [Google Scholar]
- Kurtz PF, Fodstad JC, Huete JM, & Hagopian LP (2013). Caregiver- and staff‐conducted functional analysis outcomes: A summary of 52 cases. Journal of Applied Behavior Analysis, 46, 738–749. doi: 10.1002/jaba.87 [DOI] [PubMed] [Google Scholar]
- Liggett AP, Nastri R, & Podlesnik CA (2018). Assessing the combined effects of resurgence and reinstatement in children diagnosed with autism spectrum disorder. Journal of the Experimental Analysis of Behavior, 109, 408–421. doi: 10.1002/jeab.315 [DOI] [PubMed] [Google Scholar]
- Lowry MA, & Lachter GD (1977). Response elimination: A comparison of four procedures. Learning and Motivation, 8, 69–76. doi: 10.1016/0023-9690(77)90068-6 [DOI] [Google Scholar]
- Liddon CJ, Kelley ME, Rey CN, Liggett AP, & Ribeiro A (2018). A translational analysis of ABA and ABC renewal of operant behavior. Journal of Applied Behavior Analysis, 51, 819–830. doi: 10.1002/jaba.496 [DOI] [PubMed] [Google Scholar]
- Mace FC, & Lalli JS (1991). Linking descriptive and experimental analyses in the treatment of bizarre speech. Journal of Applied Behavior Analysis, 24, 553–562. doi: 10.1901/jaba.1991.24-553 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McConnell BL, & Miller RR (2014). Associative accounts of recovery-from-extinction effects. Learning and Motivation, 4, 1–15. doi: 10.1016/j.lmot.2014.01.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakajima S, Tanaka S, Urushihara K, & Imada H (2000). Renewal of extinguished lever-press responses upon return to the training context. Learning and Motivation, 31, 416–431. doi: 10.1006/lmot.2000.1064 [DOI] [Google Scholar]
- Nelson JB, Sanjuan MC, Vadillo-Ruiz S, Pérez J, & León SP (2011). Experimental renewal in human participants. Journal of Experimental Psychology: Animal Behavior Processes, 37, 58–70. doi: 10.1037/a0020519 [DOI] [PubMed] [Google Scholar]
- Petscher ES, Rey C, & Bailey JS (2009). A review of empirical support for differential reinforcement of alternative behavior. Research in Developmental Disabilities, 30, 409–425. doi: 10.1016/j.ridd.2008.08.008 [DOI] [PubMed] [Google Scholar]
- Phillips CL, Iannaccone JA, Rooker GW, & Hagopian LP (2017). Noncontingent reinforcement for the treatment of severe problem behavior: An analysis of 27 consecutive applications. Journal of Applied Behavior Analysis, 50, 357–376. doi: 10.1002/jaba.376 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Podlesnik CA, & Kelley ME (2014). Resurgence: Response competition, stimulus control, and reinforcer control. Journal of the Experimental Analysis of Behavior, 102, 231–240. doi: 10.1002/jeab.102 [DOI] [PubMed] [Google Scholar]
- Podlesnik CA, Kelley ME, Jimenez-Gomez C, & Bouton ME (2017). Renewed behavior produced by context change and its implications for treatment maintenance: A review. Journal of Applied Behavior Analysis, 50, 675–697. doi: 10.1002/jaba.400 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Podlesnik CA, & Miranda-Dukoski L (2015). Stimulus generalization and operant context renewal. Behavioural Processes, 11, 93–98. doi: 10.1016/j.beproc.2015.07.015 [DOI] [PubMed] [Google Scholar]
- Podlesnik CA, & Shahan TA (2009). Behavioral momentum and relapse of extinguished operant responding. Learning & Behavior, 37, 357–364. doi: 10.3758/LB.37.4.357 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pritchard D, Hoerger M, & Mace FC (2014). Treatment relapse and behavioral momentum theory. Journal of Applied Behavior Analysis, 47, 814–833. doi: 10.1002/jaba.163 [DOI] [PubMed] [Google Scholar]
- Querim AC, Iwata BA, Roscoe EM, Schlichenmeyer KJ, Ortega JV, & Hurl KE (2013). Functional analysis screening for problem behavior maintained by automatic reinforcement. Journal of Applied Behavior Analysis, 46, 47–60. doi: 10.1002/jaba.26 [DOI] [PubMed] [Google Scholar]
- Rescorla RA, & Skucy JC (1969). Effect of response-independent reinforcers during extinction. Journal of Comparative and Physiological Psychology, 67, 381–389. doi: 10.1037/h0026793 [DOI] [Google Scholar]
- Saini V, Sullivan WE, Baxter EL, DeRosa NM, & Roane HS (2018). Renewal during functional communication training. Journal of Applied Behavior Analysis, 51, 603–619. doi: 10.1002/jaba.471 [DOI] [PubMed] [Google Scholar]
- Shiban Y, Pauli P, & Mühlberger A (2013). Effect of multiple context exposure on renewal in spider phobia. Behaviour Research and Therapy, 51, 68–74. doi: 10.1016/j.brat.2012.10.007 [DOI] [PubMed] [Google Scholar]
- Sullivan WE, Saini V, & Roane HS (2018). A nonsequential approach to the study of operant renewal: A reverse translation. Journal of the Experimental Analysis of Behavior, 110, 74–86. doi: 10.1002/jeab.456 [DOI] [PubMed] [Google Scholar]
- Thompson RH, & Iwata BA (2001). A descriptive analysis of social consequences following problem behavior. Journal of Applied Behavior Analysis, 34, 169–178. doi: 10.1901/jaba.2001.34-169 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tiger JH, Hanley GP, & Bruzek J (2008). Functional communication training: A review and practical guide. Behavior Analysis in Practice, 1, 16–23. doi: 10.1007/bf03391716 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trask S, & Bouton ME (2016). Discriminative properties of the reinforcer can be used to attenuate the renewal of extinguished operant behavior. Learning & Behavior, 44, 151–161. doi: 10.3758/s13420-015-0195-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todd TP, Winterbauer NE, & Bouton ME (2012). Effects of the amount of acquisition and contextual generalization on the renewal of instrumental behavior after extinction. Learning & Behavior, 40, 145–157. doi: 10.3758/s13420-011-0051-5 [DOI] [PubMed] [Google Scholar]
- Vollmer TR, Iwata BA, Zarcone JR, Smith RG, & Mazaleski JL (1993). The role of attention in the treatment of attention-maintained self-injurious behavior: Noncontingent reinforcement and differential reinforcement of other behavior. Journal of Applied Behavior Analysis, 26, 9–21. doi: 10.1901/jaba.1993.26-9 [DOI] [PMC free article] [PubMed] [Google Scholar]