

1. Introduction
Differential reinforcement of alternative behavior is a simple and effective procedure for reducing a wide range of problematic behaviors (see Petscher, Rey, & Bailey, 2009). Generally, alternative-reinforcement based therapies involve withholding reinforcement (i.e., extinction) for a problematic target behavior while reinforcement is provided for a more acceptable alternative behavior (i.e., alternative reinforcement). Such procedures tend to be highly effective at suppressing problem behavior while these contingencies for target and alternative behaviors remain in place. However, when alternative reinforcers become unavailable during lapses in treatment or upon treatment cessation, relapse of problem behavior can occur (e.g., Volkert, Lerman, Call, & Trosclair-Lasserre, 2009; Wacker et al., 2013). Relapse following omission or reduction of alternative reinforcement has been termed resurgence (Epstein, 1985), and represents a serious challenge for maintenance of treatment effects (e.g., Briggs et al., 2018; St. Peter, 2015).
Resurgence has been studied extensively in basic research with animals (e.g., Craig, Browning, Nall, Marshall, & Shahan, 2017; Lieving & Lattal, 2003; Podlesnik & Kelley, 2014; Winterbauer & Bouton, 2010). The typical animal model of resurgence consists of three phases. In Phase 1, animals earn reinforcers by performing a target behavior. In Phase 2, target behavior no longer produces reinforcement (i.e., extinction) and an alternative behavior is reinforced. Finally, in Phase 3, target behavior typically increases when the alternative behavior is also extinguished.
Animal studies of resurgence may have translational utility for clinical treatment of problem behaviors. Phase 1 can serve as an analogue of a history of reinforcement for problem behavior, Phase 2 as an analogue of treatment with extinction and alternative reinforcement, and Phase 3 as an analogue of omission errors following treatment cessation or treatment integrity violations (e.g. Fuhrman, Fisher, & Greer, 2016; Nevin et al., 2016). As such, animal studies have been used to evaluate resurgence mitigation techniques such as thinning alternative reinforcer rates and magnitudes (Craig et al., 2017; Schepers & Bouton, 2015; Sweeney & Shahan, 2013), lengthening durations of Phase 2 treatment (e.g., Nall, Craig, Browning, & Shahan, 2018; Winterbauer & Bouton, 2010), providing stimuli associated with alternative reinforcement during extinction of alternative responding (Craig, Browning, & Shahan, 2017), and others (e.g., Bouton & Trask, 2016; Jarmolowicz & Lattal, 2014; Lieving & Lattal, 2003).
Although the typical resurgence procedure examines relapse of a previously-extinguished target behavior, in some cases extinction of target behavior in the clinic may be relatively ineffective (e.g., DeRosa, Roane, Bishop, & Silkowski, 2016; Hagopian, Fisher, Sullivan, Acquisto, & LeBlanc, 1998; Hanley, Piazza, Fisher, & Maglieri, 2005), difficult to achieve (e.g., Baker, Rapp, & Carroll, 2010; DeRosa et al., 2016; Kurtz et al., 2003; Lang et al., 2011; Matson & LoVullo, 2008; Sanders-Dewey & Larson, 2006; Singh, Manning, & Angell, 1982), or contraindicated due to risk of harm (e.g., Goh & Iwata, 1994). In such cases, punishment rather than extinction may be implemented.
Punishment procedures reduce problem behavior by presenting response-dependent aversive stimuli (e.g., verbal reprimand) or removing access to positive reinforcers (e.g., time-out). In addition to explicit punishment-based interventions, common treatment procedures employed in the clinic may have punishment-like properties. For example, guided compliance, overcorrection, and response blocking procedures have all been suggested to have punishment-like properties (Lerman & Iwata, 1996; Lerman & Vorndran, 2002; Mazaleski, Iwata, Rodgers, Vollmer, & Zarcone, 1994; Rolider & Van Houten, 1990). Finally, punishment contingencies in the natural environment may serve an important role in controlling behavior as well (see Vollmer, 2002 for discussion). Thus, while its use is controversial, there are many instances in which punishment or punishment-like procedures are effective at reducing problem behavior.
There is considerable evidence that the response-suppressive effects of punishment are enhanced by the addition of reinforcement for an alternative response, both from basic (e.g., Herman & Azrin, 1964; Pelloux, Murray, & Everitt, 2015; Rawson & Leitenberg, 1973) and clinical studies (e.g., DeRosa et al., 2016; Fisher et al., 1993; Hagopian et al., 1998; Hanley et al., 2005; Rooker, Jessel, Kurtz, & Hagopian, 2013). Indeed, reinforcement of alternative behavior is recommended whenever punishment is implemented in clinical settings (Behavior Analyst Certification Board, 2016; Cooper, Heron, & Heward, 2007). However, as noted above, the removal of alternative reinforcement often induces resurgence of previously-extinguished problem behavior. Thus, it is possible that the removal of alternative reinforcement could leave problem behaviors previously suppressed by punishment susceptible to resurgence as well. Because the clinical use of punishment procedures is typically reserved for severe or dangerous problem behaviors (Doughty, Poe, & Anderson, 2005; Goh & Iwata, 1994; Griffin, Locke, & Landers, 1975; Thompson, Iwata, Conners, & Roscoe, 1999), research on resurgence of behavior previously suppressed by punishment may be particularly important.
There is some mixed evidence from basic research that the removal of alternative reinforcement can induce relapse of behavior previously suppressed by a combination of punishment, extinction, and alternative reinforcement. In human studies by Okouchi (2015) and Kestner and colleagues (2018), undergraduate students performed a target response on a computer to earn points in baseline. In the next phase, target responses no longer produced points, but would remove points (i.e., punishment of target responding). At the same time, an alternative response produced points. In the final phase, target responding remained on extinction, the punishment contingency was removed (i.e., target responding produced no consequences), and alternative responses were extinguished such that they no longer produced points. Both studies found resurgence of target responding under these conditions (Kestner et al., 2018; Okouchi, 2015).
Similarly, Rawson and Leitenberg (1973) found resurgence of lever pressing in rats when it was previously suppressed by a combination of extinction, punishment, and alternative reinforcement. Rats earned food by performing a target response in the first phase. In the second phase, target responding no longer produced food, but performing the target response resulted in a mild foot shock. At the same time, performing an alternative response produced food. In the final phase, target responding remained on extinction, the punishment contingency was removed, and alternative responding was extinguished. Rawson and Leitenberg (1973) found resurgence of target responding under these conditions. However, more recently, Kestner, Redner, Watkins, & Poling (2015) failed to find resurgence of rats’ lever pressing suppressed by extinction, punishment, and alternative reinforcement using a procedure similar to that of Rawson and Leitenberg (1973). Although the reasons for the difference in results between Rawson and Leitenberg (1973) and Kestner et al. (2015) remain unclear, it is important to note that none of the above studies have examined resurgence of a target behavior previously eliminated by punishment without also using extinction. Given that combinations of punishment and alternative reinforcement are often used in clinical situations where extinction is contraindicated, it is important to know if behavior suppressed by a combination of punishment and alternative reinforcement is susceptible to resurgence when alternative reinforcement is omitted.
Thus, the goal of the present experiment was to determine if omission of alternative reinforcement produces resurgence of a target behavior previously suppressed by a combination of punishment and alternative reinforcement. Accordingly, rats were trained to press a lever to earn food pellets in Phase 1. During Phase 2, food pellets could be earned for lever pressing as in Phase 1, but lever pressing also intermittently produced brief mild foot shocks. In addition, for an Alternative + Punishment group food pellets could also be earned for performing a nose poke response which did not produce shock. Finally, in Phase 3, resurgence was examined by removing all consequences for both responses as in the previous punishment-based resurgence procedures discussed above (Kestner et al., 2018; Kestner, Redner, Watkins, & Poling, 2015; Okouchi, 2015; Rawson & Leitenberg, 1973). Because both reinforcement for the alternative behavior and punishment for the target behavior are removed in such a resurgence test, it is possible that any increase in responding in Phase 3 could be due to the removal of punishment alone. Thus, the present experiment also included a Punishment Control group for which target behavior was punished in Phase 2, as with the Alternative + Punishment group, but no alternative reinforcement was available. In Phase 3, the foot-shock punishment alone was removed. Thus, any additional increase in target responding in the Alternative + Punishment group relative to the Punishment Control group should be due to the history of exposure to and then removal of alternative reinforcement (i.e. resurgence).
2. Materials and Methods
2.1. Subjects
Nineteen male Long-Evans rats (Charles River, Portage, MI), approximately 90 days old at the beginning of the experiment, served as subjects. Rats were housed individually in a climate-controlled colony under a 12:12 light/dark cycle (lights on at 7:00am). Rats were maintained at 80% of their free feeding weights by post-session feeding and had free access to water in their home cages. Animal housing, care, and all procedures reported below were approved by Utah State University’s Institutional Animal Care and Use Committee.
2.2. Apparatus
Ten identical Med Associates (St. Albans, VT) operant chambers measuring 30 cm × 24 cm × 21 cm and housed in light and sound attenuating cubicles were used. The chambers consisted of two side panels and a ceiling made of Plexiglas. On the front wall was an aluminum response panel equipped with two retractable levers below stimulus lights. Centered on the front wall and between the two levers was an aperture that could be illuminated during the delivery of 45-mg food pellets (which served as target and alternative reinforcers; Bio-Serv, Flemington, NJ). All food deliveries throughout the experiment were accompanied by a 3-s blackout of the chamber and illumination of the food aperture light. A house light centered at the top of the front wall provided general chamber illumination. The rear wall of the chamber consisted of an aluminum array of five nose poke apertures that could be illuminated yellow and were equipped to detect head entries. Each chamber was also equipped to deliver scrambled foot shock (detailed in section 2.3.6 below) through the metal grid floor. All sessions throughout were 25-min exclusive of reinforcer deliveries.
2.3. Procedure
2.3.1. Magazine Training.
Rats were initially trained to consume food pellets from the food aperture. In three daily sessions, rats were placed in the chamber with levers retracted and no lights illuminated. After an average of 60s, the food aperture was illuminated for 3 s and a single food pellet was delivered response independently (Variable Time, VT 60s schedule).
2.3.2. Alternative Response Training.
Immediately following magazine training, rats were exposed to a single session during which only the left-most nose poke (i.e., the alternative response) was illuminated and each response to it was reinforced (Fixed Ratio, FR 1 schedule). This session served to ensure that rats could readily perform the alternative response when it was introduced later.
2.3.4. Target Response Training.
Following Alternative Response Training, all sessions began with the illumination of the house light and insertion of both levers. The stimulus light above one lever was illuminated (i.e., the target lever; left-right counterbalanced across rats), and responses to that lever were initially reinforced according to an FR 2 schedule of reinforcement (i.e., every other lever press produced food). Once responding was stable under FR 2 conditions, responding was shaped by increasing the FR requirement by 2, within sessions, following consistent responding at each interim FR value. All rats had reached an FR 20 by the end of four sessions. During this phase and for the remainder of the experiment, responses to the other lever (i.e., inactive lever) were recorded but produced no consequence. This inactive lever was included to allow for differentiation between general increases in responding induced by extinction and responding directed to the target lever (i.e. resurgence) during later resurgence testing.
2.3.5. Phase 1: Baseline.
Next, during Phase 1, target responding was reinforced for all rats according to an FR 20 schedule for 15 sessions.
2.3.6. Phase 2: Punishment.
At the end of Phase 1, rats were divided into two groups matched on mean target response rates across the last three sessions of Phase 1. For both groups during Phase 2, target responding continued to produce reinforcement according to an FR 20 schedule. However, with probability =.5 target lever presses also produced a brief, mild foot shock (0.6mA, 50ms) for both groups. For the Alternative + Punishment group (N=9), the left-most nose poke in the rear of the chamber (i.e., the alternative response) was illuminated and responses to it produced food according to a VI 10 s schedule (i.e., the first nose poke after an average of 10 s was reinforced). Conditions for the Punishment Control group (N=10) where identical, except that no alternative reinforcement was available for nose poking. Phase 2 lasted for 5 sessions.
2.3.7. Phase 3: Resurgence Test.
Following Phase 2, all consequences were omitted for all rats. That is, food delivery was suspended for both target and alternative options, and shock was no longer delivered for target responding. Phase 3 lasted for 5 sessions.
2.3.8. Statistical Analyses.
The effects of contingencies on target responding were analyzed across phases using linear mixed-effects modeling in R (R Core Team, 2013) via the lme4 package (Bates, Mächler, Bolker, & Walker, 2015). In this type of analysis, the effects of the independent variables are simultaneously modeled for the group (fixed effects; population-level estimates) and individual subjects (random effects; or the effects of the independent variables accounting for individual differences in degree of responding and/or sensitivity to the independent variables). One of the benefits to the mixed-effects modeling approach is that while these individual subject differences are taken into account in the estimation of fixed-effects, outlying or inconsistent data are given less weight in the fixed effects estimates—in other analytic approaches such as ANOVA, such problematic values must often be excluded (or otherwise modified) because their presence violates model assumptions and their influence cannot be moderated internally (see Young, 2017, and Young, 2018 for similar discussions pertaining to multilevel techniques for delay discounting data). Thus, the chosen modeling technique is particularly well-suited to simultaneously preserve the richness of individual data and allow for more accurate estimation of overall group differences and ultimately, the effects of independent variables.
The primary mixed-effects analysis was conducted across several steps. First, an initial model was developed to evaluate and control for changes in target responding due to acclimation to the contingencies within-phase and the different levels/trajectories of target responding arranged by the phase-specific contingencies. Thus, this initial model included fixed effects of Session, Phase, a Session by Phase interaction, and a random intercept of Subject (i.e., the overall degree of responding was allowed to vary across rats). Next, the effect of theoretical interest was examined by testing the addition of a Group by Phase interaction to the initial model: a significant interaction between these predictors would capture differences in the degree (or presence) of resurgence across groups. Last, the necessity of a random slope effect of Session was explored (i.e., allowing for individual differences in the rate at which responding changed across sessions). The significance of these latter additions (Group by Phase interaction and random slope effect) was evaluated using likelihood ratio tests. Specific comparisons (e.g., last session of Phase 2 vs. first session of Phase 3) were then conducted using the lsmeans package (Lenth, 2016).
Alternative nose pokes and Inactive lever response rates were evaluated using a similar approach as described above. For these analyses, models were confined to examining response rates in specific phases (e.g., Phase 2) and/or groups (e.g., Alternative + Punishment group only) where relevant. In instances where these models included fixed effects of Session, they also included a random slope of Session due to its necessity in the primary analyses above. All models included random intercepts.
3. Results
The initial mixed-effects model included a significant Session by Phase interaction (χ2[2] = 33.82, p < .001), and the final model a significant Group by Phase interaction (χ2 [3] = 17.34, p = .001). The latter warranted inclusion of a random slope effect of session (χ2[2] = 30.32, p < .001), indicating there was variability in how quickly responding adapted for each rat in each phase. These significant interactions are discussed below as they pertain to the rate of change in responding upon the introduction of new contingencies across phases (Session by Phase interaction) and effects of prior conjoint punishment and alternative reinforcement on resurgence in Phase 3 (Group by Phase interaction).
3.1. Phase 1.
Target responding initially increased across sessions of baseline for both groups, indicating successful acquisition of the target response (see Figure 1). At the end of baseline, Alternative + Punishment and Punishment Control rats responded at similar rates on the target lever (see Figure 1 & Table 1). The significant Group by Phase interaction supported this finding, in that unlike in other phases (e.g., Phase 2 below) degree of responding was not significantly different across groups in the final session of Baseline, t(39.92) = 0.31, p = .76.
Figure 1.
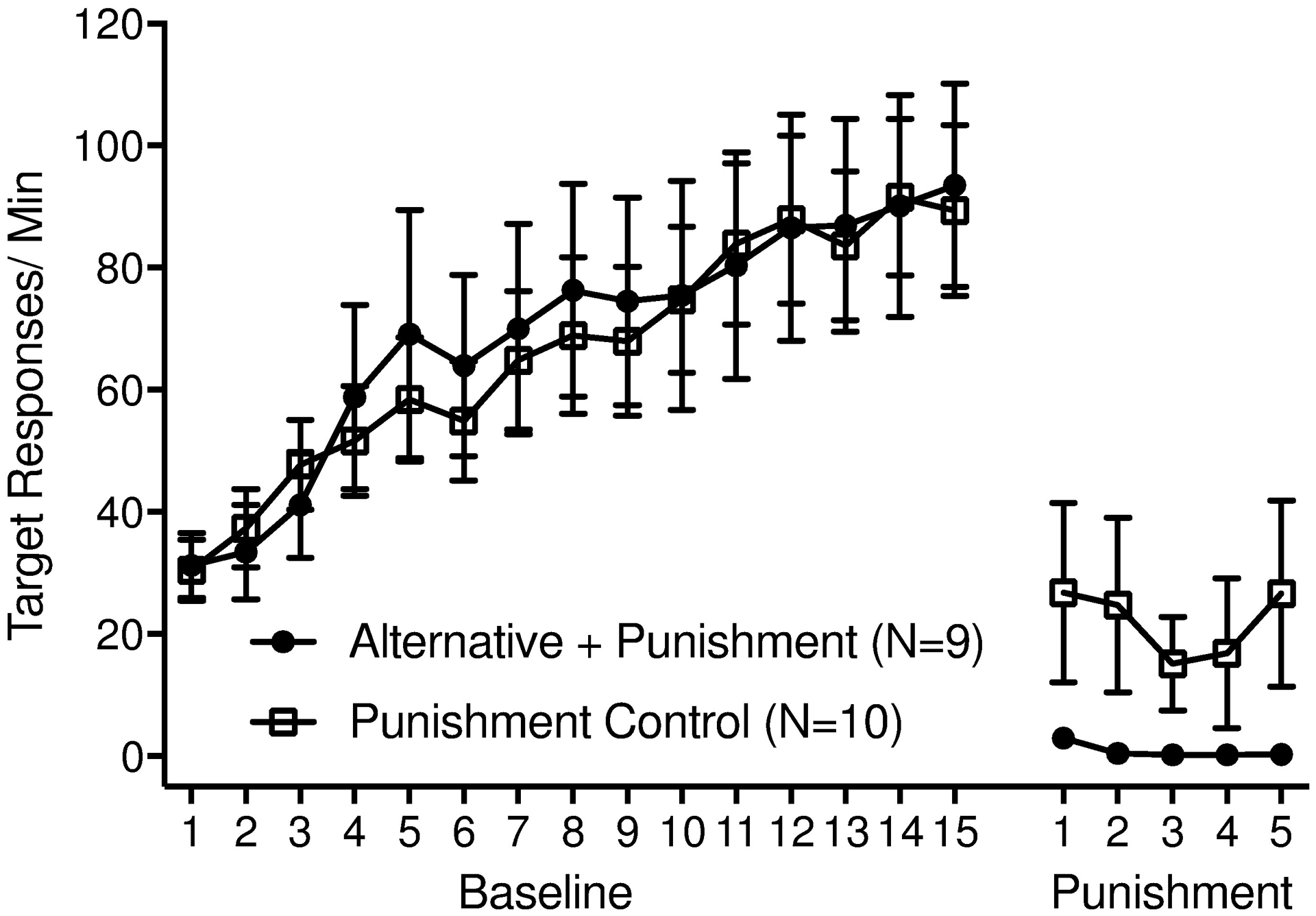
Target response rates from all sessions of Phase 1 (Baseline) and Phase 2 (Punishment) for both groups. Error bars represent standard errors of the mean.
Table 1.
Summary of Mean Response, Reinforcer, and Punishment rates from each phase of the present experiment.
| Groups | ||||||
|---|---|---|---|---|---|---|
| Alternative + Punishment | Punishment Control | |||||
| Phase 1a | Phase 2b | Phase 3c | Phase 1a | Phase 2b | Phase 3c | |
| Target/Min | 90.15 | 0.29 | 20.42 | 88.14 | 26.64 | 29.92 |
| (SEM) | (17.34) | (0.14) | (6.08) | (12.80) | (14.70) | (8.78) |
| Alt./Min | - | 80.14 | 14.30 | - | 0.32 | 0.18 |
| (SEM) | - | (7.14) | (1.61) | - | (0.09) | (0.04) |
| Inactive/Min | 0.48 | 0.14 | 0.56 | 0.20 | 0.42 | 0.42 |
| (SEM) | (0.23) | (0.06) | (0.21) | (0.05) | (0.11) | (0.11) |
| Pellets/Min | 4.63 | 8.21* | - | 4.50 | 1.33 | - |
| (SEM) | (0.72) | (1.21) | - | (0.62) | (0.77) | - |
| Shocks/Min | - | 0.15 | - | - | 13.23 | - |
| (SEM) | - | (0.07) | - | - | (7.61) | - |
Data averaged across the last three sessions of Phase 1 are shown.
Data from the last session of Phase 2 are shown.
Data from the first session of Phase 3 are shown.
Includes target and alternative reinforcers
3.2. Phase 2.
Target responding decreased for both groups during Phase 2 (see Figure 1). This was confirmed via the Session by Phase interaction, indicating that responding in the final session of Baseline was significantly higher than that in the final session of Phase 2, t(431.52) = 17.90, p < .001. While target responding did decrease for both groups during Phase 2, the significant Group by Phase interaction indicated that rats in the Alternative + Punishment group showed a greater reduction in responding: by the end of Phase 2, target response rates in the Alternative + Punishment group were significantly lower than those in the Punishment Control group, t(28.79) = 2.16, p = .04 (see Table 1).
Alternative responding increased for the Alternative + Punishment group but not the Punishment Control group during Phase 2 (see Figure 2 & Table 1). This finding was supported by a significant Group by Session interaction (χ2[1] = 30.02, p < .001) in a separate model predicting alternative response rates in Phase 2.
Figure 2.
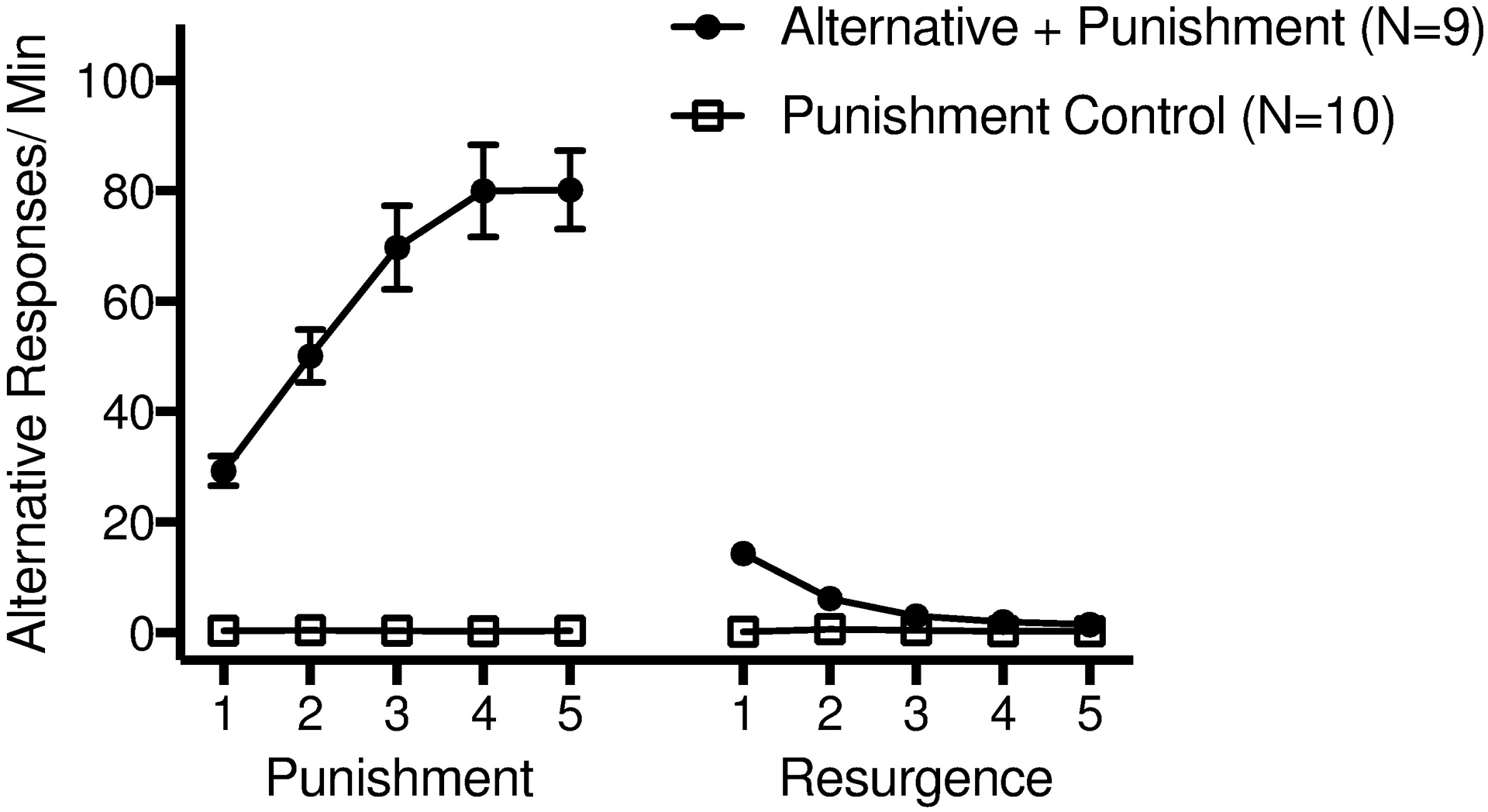
Alternative response rates from all sessions of Phases 2 (Punishment) and 3 (Resurgence) for both groups. Error bars represent standard errors of the mean.
3.3. Phase 3.
Upon removal of the contingencies in place during Phase 2, only rats in the Alternative + Punishment group showed evidence of relapse (see Figure 3). This finding was supported by the significant Group by Phase interaction, such that responding in the first session of Phase 3 increased relative to that in the final session of Phase 2 for rats in the Alternative + Punishment group (t[440.72] = 2.98, p = .003) but not for those in the Punishment Control group (t[439.58] = 0.28, p = .80) (see Table 1). Although target responding increased in the transition from Phase 2 to Phase 3 for the Alternative + Punishment group, overall response rates during Phase 3 did not differ across groups, t(19.59) = .26, p = .80.
Figure 3.
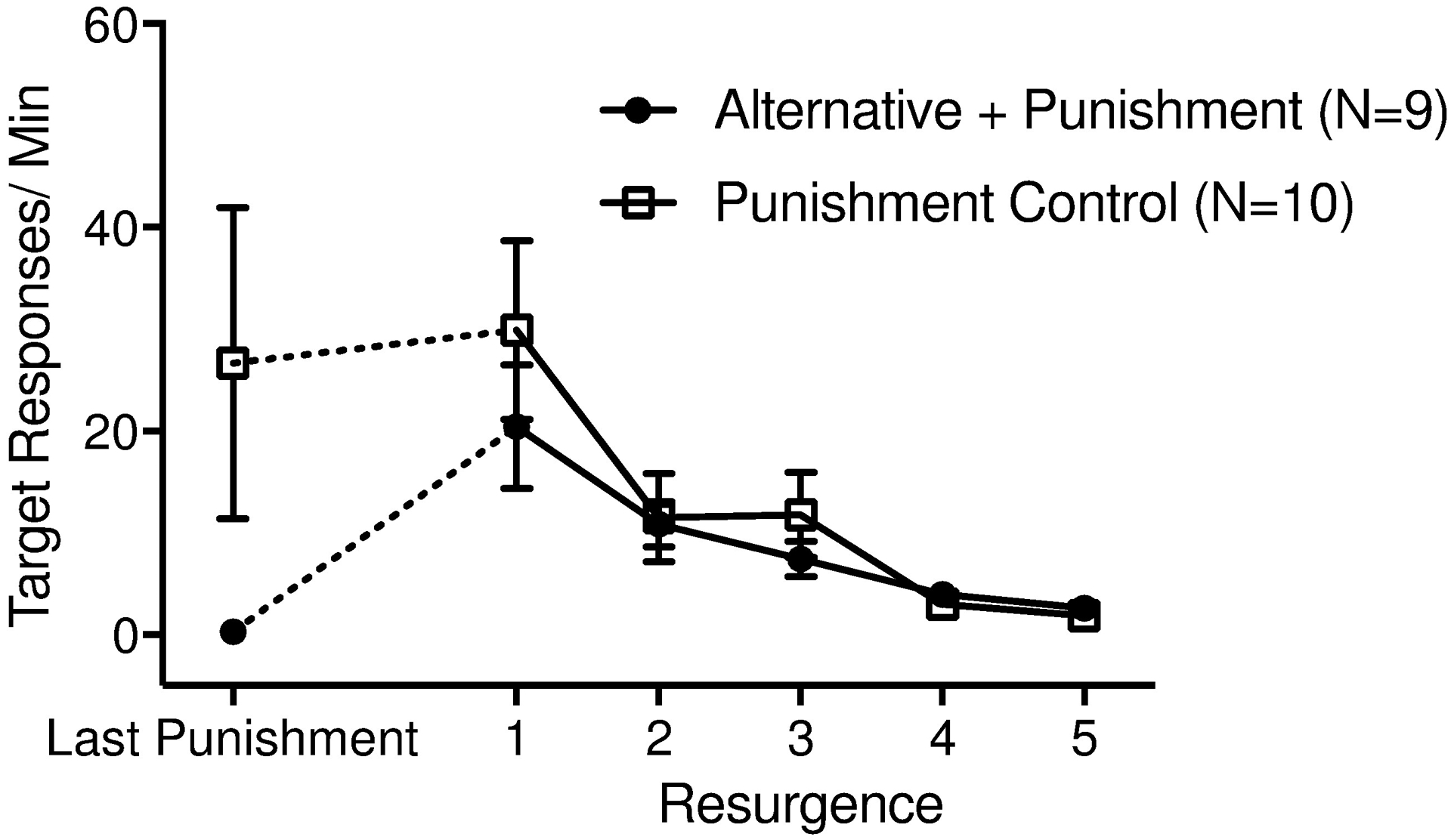
Target response rates during the last session of Phase 2 (Last Pun) and all sessions of Phase 3 (Resurgence) for both groups. Error bars represent standard errors of the mean.
Alternative responding decreased for the Alternative + Punishment group and remained low for the Punishment Control group during Phase 3 (see Figure 2). The mixed-model conducted on alternative response rates in Phase 3 revealed a significant interaction between Session and Group (χ2[1] = 28.71, p < .001), verifying that alterative responding decreased significantly only for the Alternative + Punishment group.
Inactive responding did increase slightly between the last session of Phase 2 and the first session of Phase 3 for the Alternative + Punishment group (see Table 1), but this increase was less than that for target responding. This finding was supported by a significant interaction between Lever (Target/Inactive) and Phase (Phase 2/Phase 3; χ2[1] = 10.16, p = .001) in a separate mixed-model predicting inactive and target lever response rates during the last session of Phase 2 and the first session of Phase 3 for rats in the Alternative + Punishment group. These results indicate that while responding increased regardless of lever when transitioning from Phase 2 to 3, that the increase was larger for target responding. These results provide further evidence for resurgence of target responding as opposed to a general increase in activity.
4. Discussion
The goal of the present experiment was to examine resurgence of behavior previously suppressed by a combination of punishment and alternative reinforcement. Rats initially pressed levers for food. Next, lever pressing continued to produce food, but also produced intermittent mild foot shocks. At the same time, food was made available for a nose poke response for the Alternative + Punishment group, but not for the Punishment Control group. Finally, all consequences were removed from both responses for both groups and resurgence of lever pressing was assessed.
When punishment and alternative reinforcement were discontinued during Phase 3, resurgence of target responding occurred in the Alternative + Punishment group. Importantly, the removal of punishment alone was not sufficient to induce relapse in the Punishment Control group, suggesting that resurgence in the Alternative + Punishment group was the product of exposure to and then removal of alternative reinforcement and not the product of removing the punishment contingency alone (see Figure 3). However, in addition to differences in alternative reinforcement, there were also differences in obtained rates of target reinforcement and punishment between groups during Phase 2 (see Table 1). Because the goal of the present study was to evaluate resurgence following treatment with a clinically-relevant package of target punishment and alternative reinforcement, and because the differences in reinforcement and punishment rates were inherent to this treatment package, it was important to allow differences in reinforcement and punishment rates to occur. As such, the relative contributions of each component to the differences in resurgence between groups were not assessed. Future studies should seek to evaluate how much each component or combination of components contributes to resurgence in the paradigm developed herein.
Regardless of which components may be driving resurgence, the present results indicate that the removal of alternative reinforcement can induce relapse of punished behavior as it does with extinguished behavior, and are generally consistent with previous studies examining resurgence of behavior previously suppressed by a combination of extinction, punishment, and alternative reinforcement (Kestner et al., 2018; Okouchi, 2015; Rawson & Leitenberg, 1973, but see Kesnter et al., 2015). These results are also consistent with findings from animal models demonstrating other forms of relapse following suppression by punishment (e.g., contextual renewal, Bouton & Schepers, 2015; cue-induced reinstatement, Campbell et al., 2017; incubation of craving, Krasnova et al., 2014). Thus, like extinction-based procedures, alternative reinforcement procedures using punishment may be susceptible to relapse of target behavior when alternative reinforcement becomes unavailable.
The present findings are generally consistent with Context Theory which suggests that resurgence is a special case of ABC renewal wherein the contexts are differentiated by the reinforcement contingencies in place for each response (Trask, Schepers, & Bouton, 2015). In traditional extinction-based resurgence procedures, context A is represented by target reinforcement, context B is represented by extinction of target behavior and reinforcement of alternative behavior, and context C is represented by extinction of alternative behavior. Bouton and colleagues (e.g., Bouton, Winterbauer, & Todd, 2012; Trask et al., 2015) have suggested that the inhibition of responding learned during extinction is highly context specific. Thus, inhibition of responding does not generalize to the novel context C and relapse of responding occurs. Bouton and Schepers (2015) demonstrated that the context specificity of inhibitory learning may also apply when responding is reduced by punishment. Rats were trained to press a lever for food in context A. Next, lever pressing produced mild foot shock in context B. Finally, rats were exposed to a novel context C and previously-punished responding increased. Bouton and Schepers (2015) suggested that the renewal of previously punished responding in context C may too have been the product of context-specific inhibitory learning failing to generalize.
A Context Theory interpretation may also be extended to the results of the current experiment. The response inhibition learned in Phase 2 may have failed to generalize to Phase 3, thus generating resurgence. In general, the findings of the current experiment are consistent with this approach. However, it is unclear how to quantify the magnitude of a contextual change, making it difficult to interpret the lack of resurgence in the Punishment Control group. On one hand, rats in the Punishment Control group did experience changes in response contingencies that could serve as contextual changes (i.e., target reinforcement in Phase 1, target punishment in Phase 2, and target extinction and removal of target punishment in Phase 3). On the other hand, a proponent of Context Theory could argue that those contextual changes might have been smaller in magnitude than those experienced by the Alternative + Punishment group, which included additional changes in alternative reinforcement contingencies, resulting in resurgence in the Alternative + Punishment group but not the Punishment Control group. Thus, the lack of a method for quantifying the magnitude of a contextual change makes generating and testing predictions about subsequent relapse effects difficult. Context Theory remains flexible enough to account for the current results by assuming an arbitrarily large impact of the contextual change in the Alternative + Punishment group and an arbitrarily small impact of the contextual change in the Punishment Control group. While the current findings are generally consistent with Context Theory, the flexibility and lack of specificity makes it difficult to generate data that are inconsistent with the theory and has led some authors to question its general utility and falsifiability (Craig & Shahan, 2016; McConnell & Miller, 2014; Nall et al., 2018; Podlesnik & Kelley, 2014; Shahan & Craig, 2017).
Another theory of resurgence, Resurgence as Choice (RaC; Shahan & Craig, 2017) suggests that resurgence can be understood within the general framework of the concatenated matching law (Baum & Rachlin, 1969). In its most general form, RaC suggests that the conditional probability of the target response is determined by:
| (1) |
where pT is the conditional probability of a target response and VT and VAlt are the values of the target and alternative response options, respectively (see Shahan & Craig, 2017 for a detailed discussion of how pT is converted to absolute response rates). Using a version of the temporal weighting rule (e.g., Devenport, Hill, Wilson, & Ogden, 1997), RaC provides a means to calculate how the value of the target and alternative options change across time in extinction, and thus provides a quantitative description of why extinction of an alternative behavior generates resurgence of a previously extinguished target behavior. As a result of the temporal weighting rule, RaC suggests that when alternative reinforcement is discontinued, the value of the alternative option decreases precipitously, producing an increase in the conditional probability of target responding despite continued extinction of the target response via Equation 1 (i.e., resurgence; see Shahan & Craig, 2017 for quantitative details).
RaC has been recently suggested to provide a means to understand resurgence induced by the punishment of an alternative behavior. Fontes, Todorov, & Shahan (2018) demonstrated resurgence of target behavior following the punishment of alternative behavior. First, rats performed a target response to earn food. Next, target behavior was extinguished and alternative behavior was reinforced. Finally, target behavior remained on extinction and a punishment contingency was superimposed onto the reinforcement schedule for alternative behavior. When alternative behavior was punished, target behavior increased (i.e., resurgence occurred). Fontes et al. (2018) suggested that punishment produced a decrease in the value of the alternative behavior, thus producing an increase in the conditional probability of target behavior according to Equation 1 above, despite continued extinction of the target behavior.
The data from the present experiment may also be interpreted within the general choice-based framework suggested by RaC. During Phase 2, punishment may have decreased the value (and thus response rates) of the target option for both groups. The inclusion of alternative reinforcement for the Alternative + Punishment group would be expected to further reduce the conditional probability of target responding, explaining the difference in target response rates between groups during Phase 2. Extinction of alternative responding during Phase 3 for the Alternative + Punishment group would be expected to produce a decrease in the value of the alternative option, and thus, an increase in the conditional probability of target responding and predicted target response rates despite extinction of the target response. While the data from the present experiment are consistent with RaC in a general sense, to date, an effective way to account for punishment quantitatively within a matching-law framework remains elusive (see Klapes, Riley, & McDowell, 2018 for discussion). Thus, a formal quantitative extension of RaC to resurgence of punishment-suppressed behavior must await an effective theory for quantifying how punishment reduces the value of an option over time. Until then, interpretation of the present data in terms of RaC remains speculative and lacking in quantitative specificity, and as such, it remains subject to the same criticisms as Context Theory detailed above. Thus, further formal development of both theories will be necessary before they can be meaningfully compared with respect to the present findings.
Regardless of whether the contextual signaling properties of different contingencies, the relative values of different response options, or both are responsible for driving resurgence, there are several effects from the current experiment that may be directly relevant for clinical treatments using punishment. First, target responding during treatment (i.e. punishment alone or punishment and alternative reinforcement) was reliably greater in the Punishment Control group than in the Alternative + Punishment group (i.e., Phase 2, see Figure 1). This finding is consistent with ample existing evidence that alternative reinforcement can increase the efficacy of punishment effects (Boe, 1964; Carr, Dozier, Patel, Adams, & Martin, 2002; Herman & Azrin, 1964; Holz, Azrin, & Ayllon, 1963; Pelloux et al., 2015; Rawson & Leitenberg, 1973; Thompson et al., 1999). Second, because punishment was response contingent, the Alternative + Punishment group received fewer punishers over the course of Phase 2 than did the Punishment Control group (see Table 1). Thus, the presence of alternative reinforcement reduced the frequency of punished responses, which may be advantageous for clinical application. Finally, while alternative reinforcement did expedite suppression of target responding, resurgence occurred only for those individuals that received alternative reinforcement. Because punishment is typically reserved for particularly resilient, severe, or dangerous behaviors (e.g., Doughty, Poe, & Anderson, 2005; Goh & Iwata, 1994; Griffin, Locke, & Landers, 1975; Thompson, Iwata, Conners, & Roscoe, 1999), resurgence under those conditions may be especially problematic. Thus, it may be important to weigh the consequences of using alternative reinforcement: quickly reducing severe problem behavior with alternative reinforcement may leave the problem behavior susceptible to resurgence when alternative reinforcement is omitted.
Finally, the procedures herein combining alternative reinforcement and punishment may also be useful for evaluating resurgence of drug use, as it has been suggested that punishment models better reflect the conditions facing humans with substance use disorders than do traditional extinction-based models of relapse (e.g., Burman, 1997; Laudet, Savage, & Mahmood, 2002; Marchant, Li, & Shaham, 2013; Panlilio, Thorndike, & Schindler, 2003). Because recent evidence suggests the involvement of different neurobiological mechanisms in relapse of drug seeking previously suppressed by punishment versus extinction (Pelloux, Minier-Toribio, Hoots, Bossert, & Shaham, 2018), the present procedure could be useful for investigating the neurobiological processes underlying resurgence of behavior suppressed by punishment.
5. Conclusions.
The major finding of the present experiment is that resurgence of target behavior can occur when alternative reinforcement is discontinued following suppression of that behavior by punishment and alternative reinforcement. This finding is consistent with previous work showing that resurgence of target behavior can occur when alternative reinforcement is omitted following suppression of the target behavior by extinction alone and combinations of punishment and extinction. The results of the current experiment are generally consistent with the Context Theory and Resurgence as Choice approaches for understanding resurgence. Finally, the animal model developed herein could serve as a base for studying clinically relevant resurgence mitigation strategies, resurgence of drug seeking, and the neurobiological underpinnings of resurgence following punishment.
Acknowledgments
This research was supported by NIH grant R21DA038950 (TAS). Jillian M. Rung is now at the division of Behavioral Medicine in the Jacobs School of Medicine and Biomedical Sciences at the University at Buffalo, State University of New York. The authors have no competing interests to declare.
References
- Bates D, Mächler M, Bolker BM, & Walker SC (2015). Fitting linear mixed-effects models using lme4. Journal of Statistical Software, 67(1), 1–48. [Google Scholar]
- Baum WM, & Rachlin HC (1969). Choice as time allocation. Journal of the Experimental Analysis of Behavior, 12(6), 861–874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behavior Analyst Certification Board. (2016). Professional and Ethical Compliance Code for Behavior Analysts. Professional and Ethical Compliance Code for Behavior Analysts
- Boe EE (1964). Extinction As a Function of Intensity of Punishment, Amount of. Canadian Journal of Psychology, 18(1962), 328–342. [DOI] [PubMed] [Google Scholar]
- Bouton ME, & Schepers ST (2015). Renewal after the punishment of free operant behavior. Journal of Experimental Psychology: Animal Learning and Cognition, 41(1), 81–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouton ME, & Trask S (2016). Role of the discriminative properties of the reinforcer in resurgence. Learning & Behavior, 44(2011), 137–150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouton M, Winterbauer N, & Todd T (2012). Relapse processes after the extinction of instrumental learning: renewal, resurgence, and reacquisition. Behavioural Processes. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burman S (1997). The challenge of sobriety: Natural recovery without treatment and self- help groups. Journal of Substance Abuse, 9(1), 41–61. [DOI] [PubMed] [Google Scholar]
- Campbell EJ, Barker DJ, Nasser HM, Kaganovsky K, Dayas CV, & Marchant NJ (2017). Cue-induced food seeking after punishment is associated with increased fos expression in the lateral hypothalamus and basolateral and medial amygdala. Behavioral Neuroscience, 131(2), 155–167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carr JE, Dozier CL, Patel MR, Adams AN, & Martin N (2002). Treatment of automatically reinforced object mouthing with noncontingent reinforcement and response blocking: Experimental analysis and social validation. Research in Developmental Disabilities, 23(1), 37–44. [DOI] [PubMed] [Google Scholar]
- Cooper J, Heron T, & Heward W (2007). Applied behavior analysis. [DOI] [PMC free article] [PubMed]
- Craig AR, Browning KO, Nall RW, Marshall CM, & Shahan TA (2017). Resurgence and alternative-reinforcer magnitude. Journal of the Experimental Analysis of Behavior, 107(2), 218–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Craig AR, Browning KO, & Shahan TA (2017). Stimuli previously associated with reinforcement mitigate resurgence. Journal of the Experimental Analysis of Behavior, 108(2), 139–150. [DOI] [PubMed] [Google Scholar]
- Craig AR, & Shahan TA (2016). Behavioral momentum theory fails to account for the effects of reinforcement rate on resurgence. Journal of the Experimental Analysis of Behavior, 105(3), 375–392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeRosa NM, Roane HS, Bishop JR, & Silkowski EL (2016). The combined effects of noncontingent reinforcement and punishment on the reduction of rumination. Journal of Applied Behavior Analysis, 49(3), 680–685. [DOI] [PubMed] [Google Scholar]
- Devenport L, Hill T, Wilson M, & Ogden E (1997). Tracking and averaging in variable environments: A transition rule. Journal of Experimental Psychology: Animal Behavior Processes, 23(4), 450–460. [Google Scholar]
- Doughty SS, Poe SG, & Anderson CM (2005). Effects of punishment and response-independent attention on severe problem behavior and appropriate toy play. Journal of Early and Intensive Behavior Intervention, 2(2), 91–98. [Google Scholar]
- Epstein R (1985). Extinction-induced resurgence: Preliminary investigations and possible applications. The Psychological Record. [Google Scholar]
- Fisher W, Piazza C, Cataldo M, Harrell R, Jefferson G, & Conner R (1993). Functional communication training with and without extinction and punishment. Journal of Applied Behavior Analysis, 26(1), 23–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuhrman AM, Fisher WW, & Greer BD (2016). A preliminary investigation on improving functional communication training by mitigating resurgence of destructive behavior. Journal of Applied Behavior Analysis, 49(4), 884–899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goh HL, & Iwata B. a. (1994). Behavioral persistence and variability during extinction of self-injury maintained by escape. Journal of Applied Behavior Analysis, 27(1), 173–174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griffin JC, Locke BJ, & Landers WF (1975). Manipulation of potential punishment parameters in the treatment of self-injury. Journal of Applied Behavior Analysis, 8(4), 458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagopian LP, Fisher WW, Sullivan MT, Acquisto J, & LeBlanc LA (1998). Effectiveness of functional communication training with and without extinction and punishment: A Summary of 21 inpatient cases. Journal of Applied Behavior Analysis, 31(2), 211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanley GP, Piazza CC, Fisher WW, & Maglieri KA (2005). On the effectiveness of and preference for punishment and extinction components of function-based interventions. Journal of Applied Behavior Analysis, 38(1), 51–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herman RL, & Azrin NH (1964). Punishment By Noise in an Alternative Response Situation. Journal of the Experimental Analysis of Behavior, 7(2), 185–188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holz WC, Azrin NH, & Ayllon T (1963). Elimination of behavior of mental patients by response-produced extinction. Journal of the Experimental Analysis of Behavior, 6(3), 407–412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jarmolowicz DP, & Lattal KA (2014). Resurgence Under Delayed Reinforcement. Psychological Record, 64(2), 189–193. [Google Scholar]
- Kestner KM, Romano LM, St. Peter CC, & Mesches GA (2018). Resurgence Following Response Cost in a Human-Operant Procedure. The Psychological Record, 68(1), 81–87. [Google Scholar]
- Kestner K, Redner R, Watkins EE, & Poling A (2015). The Effects of Punishment on Resurgence in Laboratory Rats. Psychological Record, 65(2), 315–321. [Google Scholar]
- Klapes B, Riley S, & McDowell JJ (2018). Toward a contemporary quantitative model of punishment. Journal of the Experimental Analysis of Behavior, 109(2), 336–348. [DOI] [PubMed] [Google Scholar]
- Krasnova IN, Marchant NJ, Ladenheim B, McCoy MT, Panlilio LV, Bossert JM, … Cadet JL (2014). Incubation of methamphetamine and palatable food craving after punishment-induced abstinence. Neuropsychopharmacology, 39(8), 2008–2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laudet AB, Savage R, & Mahmood D (2002). Pathways to long-term recovery: a preliminary investigation. Journal of Psychoactive Drugs, 34(3), 305–311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lenth RV (2016). Least-Squares Means: The R Package lsmeans. Journal of Statistical Software, 69(1), 1–33. [Google Scholar]
- Lerman DC, & Iwata BA (1996). A methodology for distinguishing between extinction and punishment effects associated with response blocking. Journal of Applied Behavior Analysis, 29(2), 231–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lerman DC, & Vorndran CM (2002). On the status of knowledge for using punishment implications for treating behavior disorders. Journal of Applied Behavior Analysis, 35(4), 431–464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lieving G. a, & Lattal K. a. (2003). Recency, repeatability, and reinforcer retrenchment: An experimental analysis of resurgence. Journal of the Experimental Analysis of Behavior, 80(2), 217–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mace FC, McComas JJ, Mauro BC, Progar PR, Taylor B, Ervin R, & Zangrillo AN (2010). Differential reinforcement of alternative behavior increases resistance to extinction: clinical demonstration, animal modeling, and clinical test of one solution. Journal of the Experimental Analysis of Behavior, 93(3), 349–367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marchant NJ, Li X, & Shaham Y (2013). Recent developments in animal models of drug relapse. Current Opinion in Neurobiology, 23(4), 675–683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McConnell BL, & Miller RR (2014). Associative accounts of recovery-from-extinction effects. Learning and Motivation, 46, 1–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nall RW, Craig AR, Browning KO, & Shahan TA (2018). Longer treatment with alternative non-drug reinforcement fails to reduce resurgence of cocaine or alcohol seeking in rats. Behavioural Brain Research, 341, 54–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nevin JA, Mace FC, DeLeon IG, Shahan TA, Shamlian KD, Lit K, … Craig AR (2016). Effects of signaled and unsignaled alternative reinforcement on persistence and relapse in children and pigeons. Journal of the Experimental Analysis of Behavior, 106(1), 34–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okouchi H (2015). Resurgence of Two-Response Sequences Punished By Point-Loss Response Cost in Humans. Mexican Journal of Behavior Analysis, 41(2), 137–154. [Google Scholar]
- Panlilio LV, Thorndike EB, & Schindler CW (2003). Reinstatement of punishment-suppressed opioid self-administration in rats: An alternative model of relapse to drug abuse. Psychopharmacology, 168(1–2), 229–235. [DOI] [PubMed] [Google Scholar]
- Pelloux Y, Minier-Toribio A, Hoots JK, Bossert JM, & Shaham Y (2018). Opposite Effects of Basolateral Amygdala Inactivation on Context-Induced Relapse to Cocaine Seeking after Extinction versus Punishment. Neuroscience, 38(1), 51–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pelloux Y, Murray JE, & Everitt BJ (2015). Differential vulnerability to the punishment of cocaine related behaviours: Effects of locus of punishment, cocaine taking history and alternative reinforcer availability. Psychopharmacology, 232(1), 125–134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petscher ES, Rey C, & Bailey JS (2009). A review of empirical support for differential reinforcement of alternative behavior. Research in Developmental Disabilities, 30(3), 409–425. [DOI] [PubMed] [Google Scholar]
- Podlesnik CA, & Kelley ME (2014). Resurgence: Response competition, stimulus control, and reinforcer control. Journal of the Experimental Analysis of Behavior, 102(2), 231–240. [DOI] [PubMed] [Google Scholar]
- R Core Team. (2013). R Software. R: A Language and Environment for Statistical Computing. [Google Scholar]
- Rawson RA, & Leitenberg H (1973). Reinforced alternative behavior during punishment and extinction with rats. Journal of Comparative and Physiological Psychology, 85(3), 593–600. [Google Scholar]
- Rolider A, & Van Houten R (1990). The role of reinforcement in reducing inapropriate behavior: Some myths and misconceptions. In Perspectives on the use of nonaversive and aversive interventions for persons with developmental disabilities (pp. 119–127). [Google Scholar]
- Rooker GW, Jessel J, Kurtz PF, & Hagopian LP (2013). Functional communication training with and without alternative reinforcement and punishment: An analysis of 58 applications. Journal of Applied Behavior Analysis, 46(4), 708–722. [DOI] [PubMed] [Google Scholar]
- Schepers ST, & Bouton ME (2015). Effects of reinforcer distribution during response elimination on resurgence of an instrumental behavior. Journal of Experimental Psychology: Animal Learning and Cognition, 41(2), 179–192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shahan TA, & Craig AR (2017). Resurgence as Choice. Behavioural Processes, 141(Part 1), 100–127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith MR, & Lerman DC (1999). A Preliminary Comparison of Guided Compliance and High-Probability Instructional Sequences as Treatment for Noncompliance in Children with Developmental Disabilities. Pergamon Research in Developmental Disabilities (Vol. 20). [DOI] [PubMed] [Google Scholar]
- St. Peter CC (2015). Six reasons why applied behavior analysts should know about resurgence. Mexican Journal of Behavior Analysis, 41(2), 252–268. [Google Scholar]
- Sweeney M, & Shahan T (2013). Effects of high, low, and thinning rates of alternative reinforcement on response elimination and resurgence. Journal of the Experimental. [DOI] [PubMed] [Google Scholar]
- Thompson RH, Iwata BA, Conners J, & Roscoe EM (1999). Effects of reinforcement for alternative behavior during punishment of self-injury. Journal of Applied Behavior Analysis, 32(3), 317–328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trask S, Schepers ST, & Bouton ME (2015). Context Change Explains Resurgence After the Extinction of Operant Behavior. Mexican Journal of Behavior Analysis, 41(2), 187–210. [PMC free article] [PubMed] [Google Scholar]
- Volkert VM, Lerman DC, Call NA, & Trosclair-Lasserre N (2009). An evaluation of resurgence during treatment with functional communication training. Journal of Applied Behavior Analysis, 42(1), 145–160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vollmer TR (2002). “Punishment happens: Some comments on Lerman and Vorndran’s review.” Journal of Apllied Behavior Analysis 35.4 (2002): 469–473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wacker D, Harding JW, Morgan TA, Berg WK, Schieltz KM, Lee JF, & Padilla YC (2013). An Evaluation of Resurgence During Functional Communication Training. The Psychological Record, 63(1), 3–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winterbauer NE, & Bouton ME (2010). Mechanisms of resurgence of an extinguished instrumental behavior. Journal of Experimental Psychology: Animal Behavior Processes, 36(3), 343–353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Young ME (2017). Discounting: A practical guide to multilevel analysis of indifference data. Journal of the Experimental Analysis of Behavior, 108(1), 97–112. [DOI] [PubMed] [Google Scholar]
- Young ME (2018). Discounting: A practical guide to multilevel analysis of choice data. Journal of the Experimental Analysis of Behavior, 109(2), 293–312. [DOI] [PubMed] [Google Scholar]