

Abstract
Purpose
Growth factors and inflammatory and angiogenetic proteins are involved in the development of retinopathy of prematurity (ROP). However, no early biochemical markers are in clinical use to predict ROP. By performing cluster analysis of multiple biomarkers, we aimed to determine patient groups with high and low risk for developing ROP.
Methods
In total, 202 protein markers in plasma were quantified by proximity extension assay from 35 extremely preterm infants on day 2 of life. Infants were sorted in groups by automated two-dimensional hierarchical clustering of all biomarkers. ROP was classified as stages I to III with or without surgical treatment. Predictive biomarkers were evaluated by analysis of variance and detected differences by two-sided paired t-test with Bonferroni corrections for multiple comparisons.
Results
Differences in 39 biochemical markers divided infants without ROP into two control groups (control 1, n = 7; control 2, n = 5; P < 0.05). Sixty-six biochemical markers defined differences between the control groups (n = 13) and all ROP infants (n = 23; P < 0.05). PARK7, VIM, MPO, CD69, and NEMO were markedly increased in control 1 compared to all ROP infants (P < 0.001). Lower TNFRSF4 and higher HER2 and GAL appeared in infants with ROP as compared to control 1 and/or 2 (P < 0.05, respectively).
Conclusions
Our data suggest that early elevated levels of PARK7, VIM, MPO, CD69, and NEMO may be associated with lower risk of developing ROP. Lower levels of TNFRSF4 with higher levels of HER2 and GAL may predict ROP development.
Translational Relevance
Cluster analysis of early postnatal biomarkers may help to identify infants with low or high risk of developing ROP.
Keywords: ROP, biomarkers, prematurity, cluster analysis
Introduction
Retinopathy of prematurity (ROP) is a vasoproliferative retinal disorder affecting premature infants and a leading cause of childhood blindness worldwide.1 The number of infants at risk for ROP has been increasing as the survival rates of premature births improve due to advances in neonatal care.2 The retinal developmental mechanisms behind ROP have been extensively studied in animal models during the past decades.3,4 The dysregulation of retinal vascular development is a major factor in ROP pathogenesis,5 where vascular endothelial growth factor (VEGF) and insulin-like growth factor 1 (IGF-1) are important factors. However, other vascular and inflammatory proteins as well as growth factors, angiogenetic proteins, and neurotrophins have also been suggested to be associated with ROP.6–11 This multitude of ROP-associated factors demonstrates that numerous signaling pathways may be involved in the development of ROP and that maybe not a single biomarker but a combination of biomarkers should be investigated.
There are five stages used to describe ROP progression.12 Stages 1 and 2 are mild and likely to regress spontaneously. From stage 3, ROP is classified as severe since extraretinal neovascularization can become progressive enough to cause total retinal detachment.5 The incidence and severity of ROP are inversely related to birth weight (BW) and gestational age (GA)13,14 and directly related to intrauterine growth restriction.15 Male gender contributes to severe ROP.15 Models for oxygen-induced retinopathy exist,16 although the duration of mechanical ventilation seems to be a greater predictive factor for ROP development than the total duration of oxygen supplementation.17
Previous studies indicate that plasma proteins can be useful in identifying preterm newborn infants at high risk for developing diseases such as bronchopulmonary dysplasia, respiratory distress syndrome, and persistent ductus arteriosus.18–23 However, only a few investigators have aimed to find a relationship between protein levels at birth and ROP development.10,24,25 None of these early biochemical markers is in clinical use for the prediction of ROP development. To define predictive biomarkers one by one for ROP may be difficult because of the numerous interacting regulatory mechanisms and the heterogeneity of study populations. Therefore, investigating groups together with expression profiles of proteins, instead of separate protein testing, can be an alternative for detecting predictive protein-level patterns in ROP development.26
Our aim was to find predictive biomarkers for determining patient groups at high and low risk of developing ROP by investigating plasma proteins during the early postnatal period with comprehensive protein screening. The hypothesis was that proteins involved in ROP may be detected and categorized in clusters.
Methods
Study Population
Thirty-five infants were included from our previously described DAPPR (Ductus arteriosus and pulmonary circulation in premature infants) cohort.23,27 Infants born at Uppsala University Children's Hospital between November 2012 and May 2015 with a GA of less than 28 weeks and without heart defects or major congenital anomalies were eligible for inclusion. Infants were enrolled after informed and written consent was obtained from the parents. The study was approved by Regional Ethical Review Board.
ROP Screening
All infants were screened for ROP in the neonatal period, with weekly examinations from a postmenstrual age of around 31 weeks and up to at least full-term age. Treatment was performed when indicated (ETROP (Early Treatment for Retinopathy Of Prematurity) 2003).28 The data on various aspects of screening and treatment for ROP were extracted from SWEDROP (Swedish national register for retinopathy of prematurity), a national web-based ophthalmologic ROP register.29
Biochemical Markers
Blood samples were collected from umbilical arterial catheters during the second day of life. A minimum volume of 20 µL was required for proximity extension assay (PEA). After centrifugation at 2400 × g for 7 minutes, the supernatant serum was obtained and stored at −80°C until further analysis. In total, 202 biochemical markers were analyzed with the PEA technique using the Proseek Multiplex 96 × 96 CVD I, Oncology I, and Inflammation I biomarker panels (Olink Bioscience, Uppsala, Sweden) as previously described.30 The list of 202 biochemical markers is presented in Table 1. All data from the PEA analyses are presented as arbitrary units in linear values. Measurements were performed without knowledge of clinical data, and treating physicians were blinded to the results of the biochemical marker analyses.
Table 1.
List of All Tested Biochemical Markers (n = 202).
| 4E-BP1 | CCL28 | CXCL10 | ESM-1 | hGDNF | IL-1ra | LEP | MMP-7 | PTPN22 | TM | VE-Statin |
|---|---|---|---|---|---|---|---|---|---|---|
| ADA | CCL4 | CXCL11 | EZR | HGF | IL-2 | LIF | MPO | PTX3 | TNF | VIM |
| AGRP | CD244 | CXCL13 | FABP4 | hK11 | IL-20 | LIF-R | MYD88 | RAGE | TNFB | |
| AM | CD40 | CXCL16 | FADD | HSP 27 | IL-20RA | LITAF | NEMO | REG-4 | TNF-R1 | |
| AR | CD40-L | CXCL5 | FAS | ICOSLG | IL-22 RA1 | LOX-1 | NRTN | REN | TNF-R2 | |
| ARTN | CD5 | CXCL6 | FasL | IFN-γ | IL-24 | LYN | NT-3 | RETN | TNFRSF4 | |
| AXIN1 | CD6 | CXCL9 | FGF-19 | IL-1α | IL27-A | mAmP | NT-pro-BNP | SCF | TNFRSF9 | |
| BAFF | CD69 | Dkk-1 | FGF-21 | IL-10 | IL-2RB | MB | NTRK3 | SELE | TNFSF14 | |
| BDNF | CDCP1 | DNER | FGF-23 | IL-10RA | IL-33 | MCP-1 | OPG | SIRT2 | t-PA | |
| Beta-NGF | CDH3 | ECP | FGF-5 | IL-10RB | IL-4 | MCP-2 | OSM | SLAMF1 | TRAIL | |
| BNP | CDKN1A | EGF | Flt3L | IL-12 | IL-5 | MCP-3 | PAPPA | SPON1 | TRAIL-R2 | |
| CA-125 | CEA | EGFR | FR-α | IL-12B | IL-6 | MCP-4 | PAR-1 | SRC | TRANCE | |
| CAIX | CHI3L1 | eIF-4B | FS | IL-13 | IL-6RA | MIA | PARK7 | ST1A1 | TR-AP | |
| CASP-3 | CSF-1 | EMMPRIN | FUR | IL-15RA | IL-7 | MIC-A | PDGF subunit B | ST2 | TSLP | |
| CASP-8 | CST5 | EN-RAGE | GAL | IL-16 | IL-8 | MIP-1 α | PD-L1 | STAMPB | TWEAK | |
| CCL11 | CSTB | Ep-CAM | Gal-3 | IL-17A | ILT-3 | MK | PECAM-1 | TF | uPA | |
| CCL19 | CTSD | EPO | GDF-15 | IL-17C | ITGA1 | MMP-1 | PlGF | TGF-α | U-PAR | |
| CCL20 | CTSL1 | ErbB2/HER2 | GH | IL-17RB | ITGB1BP2 | MMP-10 | PRL | THPO | VEGF-A | |
| CCL23 | CX3CL1 | ErbB3/HER3 | HB-EGF | IL-18 | KLK6 | MMP-12 | PRSS8 | TIE2 | VEGF-D | |
| CCL25 | CXCL1 | ErbB4/HER4 | HE4 | IL-18R1 | LAP TGF-β-1 | MMP-3 | PSGL-1 | TIM | VEGFR-2 |
Cluster Analysis
The biochemical marker levels were analyzed together by two-dimensional hierarchical clustering (Cluster 3.0 freeware)31 as previously described.32 The clustering automatically sorted infants into groups depending on the relationship of their clinical and biochemical parameters. All biochemical marker levels were used and weighted equally in the clustering algorithm. The results were visualized by Java Treeview33 as a map of color pixels. A higher red intensity corresponded to a higher value of a certain parameter.
Statistical Analysis
One-way analysis of variance tests were performed to evaluate differences within a variable and between the hierarchically clustered groups, comparing both clinical data (GA, BW, BW percentile, and BW z score) and biochemical marker levels. Detected differences were analyzed with a two-sided paired Student's t-test for direct comparisons and Bonferroni corrections for multiple comparisons. The level of significance was set at P < 0.05 for all tests.
Results
Infants’ Clinical Characteristics
The infants’ clinical characteristics are presented in Figure 1. ROP was classified as stages I to III according to the International Classification of ROP.12 Infants with ROP stage III were divided into nontreated and treated groups. Prenatal steroids were used in all cases. No significant differences were found in BW percentile (BW%) or z score in BW between any of the groups. Infants with ROP stage I or ROP stage II were similar to infants in the group without ROP in GA and BW. Nontreated or treated infants with ROP stage III had lower GA and BW than infants without ROP (Fig. 1). The gender ratio of infants with ROP (male, n = 13; female, n = 10) was similar to infants without ROP (male, n = 6; female, n = 6).
Figure 1.

Infants’ clinical characteristics. The patients without ROP showed significant differences compared to patients with ROP III nontreated or ROP III treated in GA and BW (marked with gray). No significant differences were found in BW percentile (BW%) or in z score BW between any of the studied groups.
Clustering of Infants without ROP (Controls 1 and 2)
The two-dimensional hierarchical clustering sorted infants without ROP into two control groups (controls 1 and 2). Both control groups manifested a unique and individual visual pattern of red pixels, mainly related to significant differences in 39 of 202 biochemical markers (P < 0.05). The levels of all 39 biochemical markers were higher in control 1 than in control 2 (Fig. 2). Of these 39 markers, 5 were associated with angiogenesis, 8 with apoptosis, and 16 with inflammation34 (Fig. 2).
Figure 2.
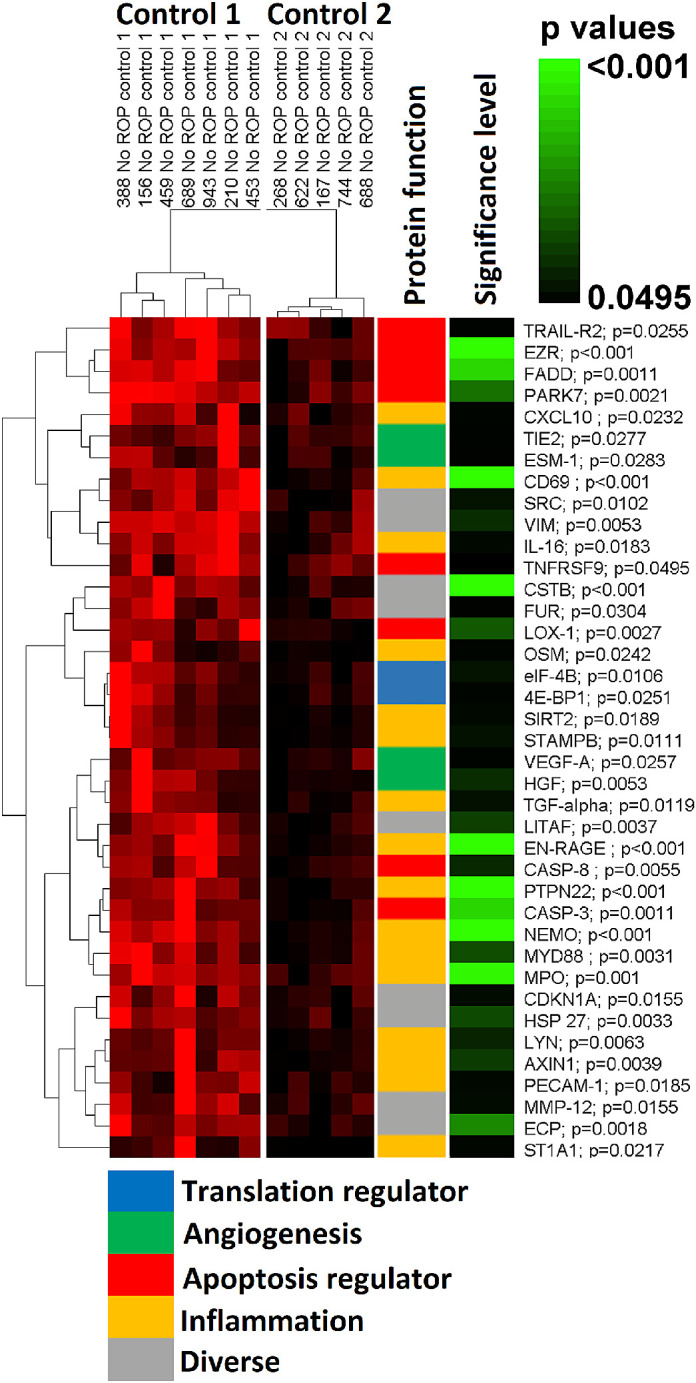
Clustering of infants without ROP (controls 1 and 2) according to biomarker expression (n = 39). Two control groups could be defined by clustering. Thirty-nine of 202 biochemical markers showed significant differences between the two control groups (controls 1 and 2). Higher intensity in red corresponds to higher value of a certain biochemical marker expression level. The function of the proteins and the significance level of differences between the two controls are labeled with colors. Higher intensity in green corresponds to higher level of significance; P values are shown next to each biochemical marker.
Clinical Characteristics of Infants without ROP (Controls 1 and 2)
There was no significant difference in the gender ratio between the two control groups, and they did not differ in GA (P = 0.316). However, infants in control 2 had higher BW (P = 0.036), higher z score (P = 0.03), and higher percentile values in BW (P = 0.027). None of the controls were growth restricted (z minimum: −1.46), but these findings could indicate that infants in control 2 had a higher intrauterine growth (Fig. 3).
Figure 3.

Differences in GA and BW between the two control groups (controls 1 and 2). The control groups did not differ in GA. However, infants in control 2 had higher BW, higher z score, and higher percentile values in BW.
Biochemical Markers Associated with ROP
The levels of 66 biochemical markers were significantly different between control 1, control 2, or both controls and all ROP infants, and the combination of respectively significant differences (P < 0.05) resulted in six groups (Table 2):
Group 1. Control 1 versus all ROP
Group 2. Control 1 and all controls versus all ROP
Group 5. Control 2 versus all ROP
Group 4. Control 2 and all controls versus all ROP
Group 3. All controls versus all ROP
Group 6. Control 1, control 2, and all controls versus all ROP
Table 2.
Biochemical Markers (n = 69) with Significant Differences between Control 1, Control 2, or Both Controls versus All Patients with ROP
| Group | Control 1 vs All ROP | Control 1–2 vs All ROP | Control 2 vs All ROP | |
|---|---|---|---|---|
| Group 1 | EPO | 0.0036 | NS | NS |
| OSM | 0.0348 | NS | NS | |
| 4E-BP1 | 0.0469 | NS | NS | |
| SIRT2 | 0.0304 | NS | NS | |
| STAMPB | 0.0232 | NS | NS | |
| CSTB | 0.0013 | NS | NS | |
| PARK7 | 0.0001 | NS | NS | |
| VEGF-A | 0.0194 | NS | NS | |
| ESM-1 | 0.0255 | NS | NS | |
| SPON1 | 0.0212 | NS | NS | |
| SRC | 0.0037 | NS | NS | |
| MMP-12 | 0.0267 | NS | NS | |
| TNFRSF9 | 0.0285 | NS | NS | |
| IL-16 | 0.0012 | NS | NS | |
| Group 2 | Flt3L | 0.0163 | 0.0064 | NS |
| VIM | <0,0001 | 0.0222 | NS | |
| FADD | 0.0013 | 0.0390 | NS | |
| MYD88 | 0.0029 | 0.0160 | NS | |
| CASP-3 | 0.0021 | 0.0351 | NS | |
| NEMO | 0.0002 | 0.0142 | NS | |
| HSP 27 | 0.0014 | 0.0078 | NS | |
| eIF-4B | 0.0111 | 0.0358 | NS | |
| PTPN22 | 0.0011 | 0.0354 | NS | |
| LITAF | 0.0036 | 0.0203 | NS | |
| MPO | 0.0001 | 0.0102 | NS | |
| EN-RAGE | 0.0020 | 0.0237 | NS | |
| CASP-8 | 0.0050 | 0.0155 | NS | |
| CD69 | 0.0002 | 0.0157 | NS | |
| LYN | 0.0067 | 0.0179 | NS | |
| AXIN1 | 0.0044 | 0.0132 | NS | |
| ECP | 0.0018 | 0.0177 | NS | |
| RETN | 0.0034 | 0.0027 | NS | |
| BAFF | 0.0051 | 0.0107 | NS | |
| RAGE | 0.0105 | 0.0088 | NS | |
| MMP-10 | 0.0170 | 0.0148 | NS | |
| MMP-3 | 0.0291 | 0.0163 | NS | |
| IL-10 | 0.0268 | 0.0450 | NS | |
| Group 3 | FGF-21 | NS | 0.0303 | NS |
| ILT-3 | NS | 0.0454 | NS | |
| mAmP | NS | 0.0385 | NS | |
| Group 4 | KLK6 | NS | 0.0174 | 0.0276 |
| VE-statin | NS | 0.0251 | 0.0315 | |
| FGF-19 | NS | 0.0197 | 0.0267 | |
| ErbB4/HER4 | NS | 0.0122 | 0.0041 | |
| TIM | NS | 0.0303 | 0.0063 | |
| Group 5 | TRAIL-R2 | NS | NS | 0.0074 |
| TR-AP | NS | NS | 0.0410 | |
| TIE2 | NS | NS | 0.0113 | |
| NT-3 | NS | NS | 0.0014 | |
| TNF-R2 | NS | NS | 0.0338 | |
| IL-10RB | NS | NS | 0.0296 | |
| GH | NS | NS | 0.0297 | |
| MIC-A | NS | NS | 0.0145 | |
| IL-8 | NS | NS | 0.0076 | |
| CEA | NS | NS | 0.0462 | |
| HGF | NS | NS | 0.0001 | |
| TGF-α | NS | NS | 0.0191 | |
| EZR | NS | NS | 0.0007 | |
| PTX3 | NS | NS | 0.0378 | |
| LOX-1 | NS | NS | 0.0005 | |
| IL-17C | NS | NS | 0.0186 | |
| ST1A1 | NS | NS | 0.0397 | |
| ITGB1BP2 | NS | NS | 0.0357 | |
| Group 6 | TNFRSF4 | 0.0333 | 0.0082 | 0.0303 |
| ErbB2/HER2 | 0.0059 | 0.0009 | 0.0099 | |
| GAL | 0.0032 | 0.0008 | 0.0306 |
NS, not significant.
Clustering of Infants without ROP
The biochemical markers were clustered separately in each group and are presented together in Figure 4A. Figure 4B presents the level of significant differences between controls and ROP infants. The direction (±) and amplitude of differences in biochemical marker levels of the two controls and ROP infants were compared to the joint mean value of all control infants (Fig. 4C). Thus, the existence of groups 1 to 6 could be confirmed both with the individual pattern of marker levels and with the direction of differences from the clustering (Figs. 4A–C). Figure 4 shows that most markers from groups 1 and 2 had higher levels in control 1 compared to control 2 or infants with ROP independent of grade. There were no significant differences in these markers between control 2 versus all the ROP infants (Fig. 4B, Table 2). Furthermore, the pattern for these markers seems to be homogeneous within control 2 and ROP infants.
Figure 4.

Clustered biochemical markers with significant differences (n = 66) between controls and all ROP patients. Patients labeled with ROP III are nontreated ROP cases. Sixty-six biochemical markers were significantly different between control 1, control 2, or both controls versus all ROP patients. Six groups (groups 1–6) could be defined depending on their combination of significances: Group 1. Control 1 versus all ROP. Group 2. Control 1 and all controls versus all ROP. Group 3. All controls versus all ROP. Group 4. Control 2 and all controls versus all ROP. Group 5. Control 2 versus all ROP. Group 6. Control 1, control 2, and all controls versus all ROP. (A) Biomarkers in each group clustered separately and presented together. (B) Level of significant differences between the two controls and all ROP patients. Increased lighter purple corresponds to higher level of significance; gray, not significant (numeric values can be followed in Table 2). (C) The direction (±) and amplitude of differences in biochemical marker levels of the two control groups. All ROP patients and combinations of patients with grade I to III ROP were compared to all control patients. Higher intensity in red corresponds to higher positive value; higher intensity in green corresponds to lower negative value.
Parkinson disease protein 7 (PARK7), vimentin (VIM), myeloperoxidase (MPO), CD69, and NF-κB essential modulator (NEMO) show the most significant differences between control 1 and all ROP infants (P < 0.001; Table 2). No correlation could be found between the levels of these five markers and the GA or the BW (data not shown). The levels of these five biochemical markers are represented in Figure 56.
Figure 5.

Biochemical markers (PARK7, VIM, MPO, CD69, NEMO) with the most significant differences (P < 0.001) between control 1 and all ROP groups. The significant differences are marked with dark gray in the table.
Figure 6.
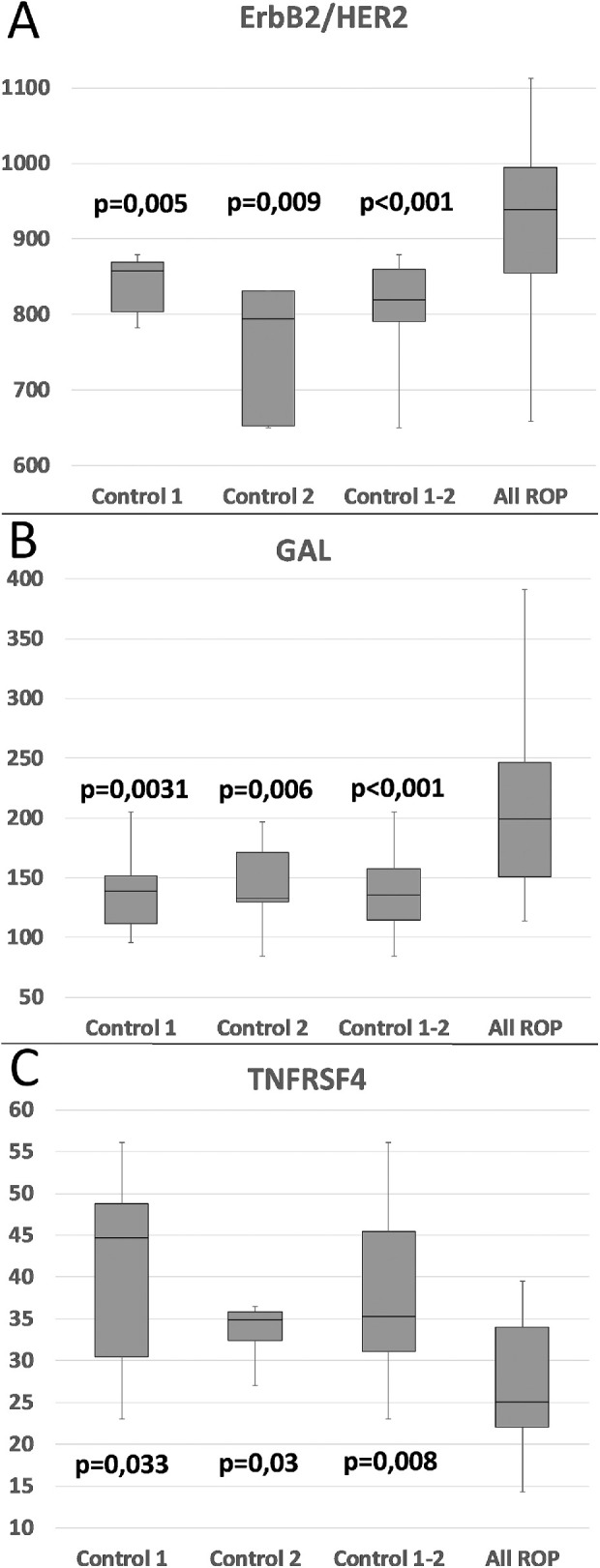
Biochemical markers (ErbB2/HER2, GAL, TNFRSF4) in group 6. All three biochemical markers from group 6 show significant differences between all combinations of controls (control 1, control 2, and all controls) and all patients with ROP. P values correspond to significant differences between controls and all patients with ROP. Note that ErbB2/HER2 and GAL are higher in controls, whereas TNFRSF4 is lower than in ROP.
The levels of hepatocyte growth factor (HGF), ezrin (EZR), and lectin-like oxidized low-density lipoprotein receptor 1 (LOX-1) in group 5 showed strong significant differences (P < 0.001; Table 2) between control 2 and all ROP infants, but the expression levels were highly heterogonous in the separate ROP groups (Fig. 4). Thirty-four of the 39 biochemical markers that distinguish the two controls from each other were represented in groups 1, 2, and 5 but not in group 3, 4, or 6.
Group 6 included three biochemical markers with significant differences between any combination of the controls (control 1, control 2, or both controls) and all ROP infants; tumor necrosis factor receptor superfamily member 4 (TNFRSF4) had lower expression in all ROP infants, whereas human epidermal growth factor receptor 2 (HER2) and galanin (GAL) had higher expression in all ROP infants Figure 6. There was no significant difference between the expression levels of these three markers and the different ROP stages, that is, no correlation to future severity of ROP could be observed (data not shown). Although GA and BW correlate negatively to the severity of ROP (Fig. 1), no correlation was seen between these three markers and the GA or BW (data not shown).
Discussion
Although numerous proinflammatory cytokines in the systemic circulation are associated with ROP,35 several studies have failed to find early predictive markers for ROP development with clinical relevance. This is likely due to the process being multifactorial and that the group of infants with potential risk of developing ROP is highly heterogeneous. Clustering of infants and plasma protein levels enabled us to identify biochemical markers with possible predictive value on ROP development in the early postnatal period in extremely premature infants. Our study has two main findings: first, simultaneously elevated levels of PARK7, VIM, MPO, CD69, and NEMO were associated with a protective effect on ROP development, and second, decreased levels of TNFRSF4 and elevated levels of HER2 and GAL were associated with ROP development independently of the final stage of ROP. All these proteins are involved in inflammatory or vascularizing processes; however, no study has previously associated any of them with ROP development. We propose the following possible explanations for the role these eight biomarkers might play in the development of ROP.
PARK7 is as a multifunctional protein with transcriptional regulation, protein chaperone, protease, and antioxidative functions.36 Xu et al.37 showed that PARK7 has the ability to protect neurons against oxidative stress and apoptosis. PARK7 may also influence angiogenesis indirectly by inhibiting production of reactive oxygen species and by enhancing the antioxidant capacity to reduce apoptosis in retinal pericytes.38 As pericytes regulate vessel permeability and endothelial cell proliferation through direct contact with endothelial cells,39 a lack of pericytes might lead to abnormal vascular morphogenesis.39 Choi et al.40 reported that the proper amount of pericytes is crucial for the normal retinal angiogenesis by controlling endothelial cell proliferation. These findings are in accordance with our results that PARK7 may have a protective effect on ROP development.
VIM, a major type III intermediate filament protein, is expressed in cells with mesenchymal origin such as astrocytes41 and endothelial cells.42 In addition to the effects that retinal astrocytes have on blood vessels, a reverse interaction also occurs, where the developing vessels influence retinal astrocyte development by promoting their differentiation.43 Extracellular VIM can be secreted by unstimulated astrocytes41 and activate insulin-like growth factor 1 receptor (IGF1R) as an alternative ligand in neurons and thus promote axonal growth.44 Interestingly, Hellström et al.45 showed that sufficient postnatal level of IGF-1 (the main ligand in IGF1R activation) is associated with normal vessel development and reduced risk of developing ROP. VIM-deficient mice show decreased flow-induced dilation during arterial remodeling, suggesting that VIM plays an important role in regulating structural responses of arteries in chronic blood flow46 and hypoxia-induced retinal neovascularization.47
Our finding that higher levels of MPO may be associated with a reduced risk of developing ROP is in contrast with observations from previous studies. Inflammation is in general found to be associated with ROP,35 and sepsis is closely related to the development of any stage of ROP.48 Bulka et al.49 found that greater CpG methylation (repressed gene transcription) of MPO in the placenta serum at birth was associated with a lower risk of prethreshold ROP, and increased MPO activity and lipid peroxide levels have been demonstrated in patients with diabetic retinopathy.50 However, the time point for inflammatory response may be of importance, in which late neonatal bacteremia appears to be a more prominent risk factor for severe ROP than early neonatal bacteremia.51 Holm et al.6 found that the risk for ROP increased when vascular endothelial growth factor receptor 1 (VEGFR-1) was elevated on day 1 but not when MPO was elevated at them same time. Although MPO is known as a local mediator of tissue damage by catalyzing the formation of reactive oxygen intermediates,52 it has multifaceted functions.53 Studies in murine models of inflammation show that MPO has an anti-inflammatory role,53,54 which appears even in complex inflammatory responses without present infectious agents.54 Odobasic et al.55 showed that MPO inhibits the generation of adaptive immunity by suppressing dendritic cell activation, antigen uptake/processing, and migration to lymph nodes to limit pathologic tissue inflammation. Furthermore, infants with total or subtotal lack of MPO have an increased incidence of chronic inflammatory conditions.56 These findings do not exclude the possibility that elevated early postnatal levels of MPO together with the other four biomarkers might have an anti-inflammatory and modulatory effect on angiogenesis in ROP.
CD69 is an early inflammation marker, which is expressed on leukocytes, and the expression increases following cell stimulation.57 There are indirect associations between lower CD69 expression and neoangiogenesis. CD69 may act as a negative regulator of endothelial cell activation,58 and Falk et al.59 found a lower percentage of CD4+CD69+CXCR3+ T cells in patients with neovascular age-related macular degeneration compared to age-matched controls. The coexpression of CD69 and chemokine receptor CXCR3 can have further consequences, as CXCR3 plays an important role in C-X-C motif chemokine ligand 10 (CXCL10)–mediated inhibition of VEGF-induced angiogenesis,60 a vascular factor implicated in the development of ROP.
NEMO is a key regulator in NF-κB–mediated signaling.61 The mutation in the gene of NEMO is a cause of incontinentia pigmenti (IP) as NF-κB activation is defective in IP cells.62 The eye involvement in infants with IP can manifest as areas of ischemia and reactive neovascularization, similar to the development of ROP.63–65
We could not find any clear explanation for the possible connection between TNFRSF4 (synonyms: OX40, CD134) and ROP in the literature. TNFRSF4 is expressed by T cells that are largely dependent on antigen recognition and participate in activation, clonal expansion, and differentiation of both CD4+ and CD8+ T cells.66 T cells can facilitate postischemic angiogenesis by recruiting macrophages to ischemic tissues, promoting secretion of cytokines in murine hindlimb ischemic models.67,68 OX40 ligand (OX40L) expression has been observed on antigen-presenting dendritic cells,69 B cells,70 microglia cells,71 and endothelial cells.72 Cunningham et al.73 showed that OX40L abolishes the retinal pigment epithelium–mediated immunosuppression. TNFRSF4 is involved in the pathogenesis of autoimmune vasculitis diseases (e.g., systemic lupus erythematosus,74 Behcet disease,75 and Henoch-Schönlein purpura).76 Inherited TNFRSF4 deficiency in a patient was associated with childhood-onset classic Kaposi sarcoma (an endothelial tumor with inflammatory origin), in which TNFRSF4 was not able to bind to its ligand (OX40L) expressed by endothelial cells.77 Gong et al.78 found upregulated TNFRSF4 gene expression in active fibrovascular membranes of proliferative diabetic retinopathy. Nakano et al.79 observed that VEGF-induced angiogenesis was suppressed by the genetic deletion of the OX40/OX40L signaling in a mouse model. Angiogenic cytokines can recruit immune cells that cause continuous secretion of cytokines and further recruitment of immune cells.80 OX40/OX40L interaction induces the phospholipase C signal transduction pathway,81 which induces diacylglycerol-protein kinase C and the inositol trisphosphate (IP(3))-intracellular free calcium ([Ca(2+)](i)) pathway.82 These pathways are also known as downstream signal pathways for VEGF-induced angiogenesis,83 which could be in conflict with our results in which ROP development was associated with lower TNFRSF4 expression. However, markers such as VEGF can be suppressed immediately after birth and increased later during development of ROP.11
HER2 (synonym: ErbB2, CD340) is known as an oncogene and a member of the human epidermal growth factor receptor family. HER2 activation is followed by several intracellular signaling pathways such as the phospholipase C pathway, which ultimately affects proliferation, survival, motility, and adhesion of different cells.84 As discussed above, the phospholipase C pathway has a connection to VEGF-induced angiogenesis,83 and studies have shown consequently that HER2 signaling is involved in angiogenesis. 85,86 HER2 overexpression correlates with increased expression of VEGF in human breast carcinoma87,88 and ovarian carcinoma,89 whereas neutralizing antibodies against HER2 result in the reduction of VEGF production in a dose-dependent manner in cancer cells.90 HER2 signaling induces protein synthesis of hypoxia-inducible factor 1α, which is playing a predominant role in the direct transcriptional upregulation of VEGF.91 Furthermore, hypoxia-inducible factor 1 was presented earlier as a potential target in the treatment of ocular neovascular diseases.92 These findings correspond well with our results that higher HER2 levels may be associated with ROP development.
GAL is a vasoactive neuropeptide acting via three known receptors: GALR 1, 2, and 3.93 GAL receptors have been detected in various ocular tissues and cells in normal human eye structures, but the role of GAL in the eye is not fully understood.94 GAL is involved in the regulation of inflammatory processes, and the expression of GAL receptors is upregulated in inflammatory conditions.95 There are potential associations to ROP development since it is involved in both vascular homeostasis96 and neuronal differentiation,97 and several authors suggest that GAL is an important angiogenetic factor in cancer.98,99 No previous reports are in conflict with our results that ROP development may be associated with elevated GAL levels.
Our study has the following limitations: we performed semiquantitative measurements of plasma proteins, which make the values not comparable with other studies. Furthermore, there are no reference intervals available for extremely premature infants, which makes it difficult to interpret and compare the measured protein levels to values in physiologic circumstances. A larger span in maturational ages and sequential samplings might have also given correlations between the tested biomarkers and GA and/or PMA (postmenstrual age), which we did not see in our study. The small number of infants in each group did not allow us to perform multivariate analysis.
The strength of our study is that we analyzed and could interpret the levels of over 200 plasma proteins simultaneously. We also present clustering as a powerful tool to describe multidimensional associations between the plasma levels of our biomarkers and to find possible relevant early markers associated with ROP development.
Conclusions
Our data suggest that simultaneously elevated plasma levels of PARK7, VIM, MPO, CD69, and NEMO after birth may help to identify extremely preterm infants at low risk of developing ROP. Lower levels of TNFRSF4 and simultaneously higher levels of HER2 and GAL may predict ROP development. The signaling networks of these proteins may help to understand the mechanisms of ROP development. Further studies are needed on larger populations to confirm the predictive relevance of the presented biochemical markers, using cluster analysis, in which comprehensive protein screening on small blood samples enables sequential testing.
Acknowledgments
Supported by HKH Kronprinsessan Lovisas förening för barnsjukvård/Stiftelsen Axel Tielmans minnesfond (Stockholm, Sweden) and Gillbergska Stiftelsen (Uppsala, Sweden).
Disclosure: L. Markasz, None; K.-W. Olsson, None; G. Holmström, None; R. Sindelar, None
References
- 1. Gilbert C, Fielder A, Gordillo L, et al.. Characteristics of infants with severe retinopathy of prematurity in countries with low, moderate, and high levels of development: implications for screening programs. Pediatrics. 2005; 115(5): e518–e525. [DOI] [PubMed] [Google Scholar]
- 2. Blencowe H, Moxon S, Gilbert C. Update on blindness due to retinopathy of prematurity globally and in India. Indian Pediatr. 2016; 53(suppl 2): S89–S92. [PubMed] [Google Scholar]
- 3. Stahl A, Connor KM, Sapieha P, et al.. The mouse retina as an angiogenesis model. Invest Ophthalmol Vis Sci. 2010; 51(6): 2813–2826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Fruttiger M. Development of the retinal vasculature. Angiogenesis. 2007; 10(2): 77–88. [DOI] [PubMed] [Google Scholar]
- 5. Hellström A, Smith LEH, Dammann O. Retinopathy of prematurity. Lancet. 2013; 382(9902): 1445–1457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Holm M, Morken TS, Fichorova RN, et al.. Systemic inflammation-associated proteins and retinopathy of prematurity in infants born before the 28th week of gestation. Invest Ophthalmol Vis Sci. 2017; 58(14): 6419–6428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Hartnett ME, Penn JS.. Mechanisms and management of retinopathy of prematurity. N Engl J Med. 2012; 367(26): 2515–2526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Hellström A, Engström E, Hård A-L, et al.. Postnatal serum insulin-like growth factor I deficiency is associated with retinopathy of prematurity and other complications of premature birth. Pediatrics. 2003; 112(5): 1016–1020. [DOI] [PubMed] [Google Scholar]
- 9. Ishida S, Yamashiro K, Usui T, et al.. Leukocytes mediate retinal vascular remodeling during development and vaso-obliteration in disease. Nat Med. 2003; 9(6): 781–788. [DOI] [PubMed] [Google Scholar]
- 10. Park YJ, Woo SJ, Kim YM, Hong S, Lee YE, Park KH. Immune and inflammatory proteins in cord blood as predictive biomarkers of retinopathy of prematurity in preterm infants. Invest Ophthalmol Vis Sci. 2019; 60(12): 3813–3820. [DOI] [PubMed] [Google Scholar]
- 11. Smith LEH. Pathogenesis of retinopathy of prematurity. Growth Horm IGF Res. 2004; 14(suppl A): S140–S144. [DOI] [PubMed] [Google Scholar]
- 12. International Committee for the Classification of Retinopathy of Prematurity. The International Classification of Retinopathy of Prematurity revisited. Arch Ophthalmol. 2005; 123(7): 991–999. [DOI] [PubMed] [Google Scholar]
- 13. Akkawi MT, Shehadeh MM, Shams ANA, et al.. Incidence and risk factors of retinopathy of prematurity in three neonatal intensive care units in Palestine. BMC Ophthalmol. 2019; 19(1): 189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Lermann VL, Fortes Filho JB, Procianoy RS. The prevalence of retinopathy of prematurity in very low birth weight newborn infants. J Pediatr (Rio J). 2006; 82(1): 27–32. [DOI] [PubMed] [Google Scholar]
- 15. Darlow BA, Hutchinson JL, Henderson-Smart DJ, et al.. Prenatal risk factors for severe retinopathy of prematurity among very preterm infants of the Australian and New Zealand Neonatal Network. Pediatrics. 2005; 115(4): 990–996. [DOI] [PubMed] [Google Scholar]
- 16. Gammons MV, Bates DO.. Models of oxygen induced retinopathy in rodents. Methods Mol Biol. 2016; 1430: 317–332. [DOI] [PubMed] [Google Scholar]
- 17. Chang JW. Risk factor analysis for the development and progression of retinopathy of prematurity. PLoS ONE. 2019; 14(7): e0219934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Bhandari A, Bhandari V.. Biomarkers in bronchopulmonary dysplasia. Paediatr Respir Rev. 2013; 14(3): 173–179. [DOI] [PubMed] [Google Scholar]
- 19. Förster K, Sass S, Ehrhardt H, et al.. Early identification of bronchopulmonary dysplasia using novel biomarkers by proteomic screening. Am J Respir Crit Care Med. 2018; 197(8): 1076–1080. [DOI] [PubMed] [Google Scholar]
- 20. Rocha G, Proença E, Guedes A, et al.. Cord blood levels of IL-6, IL-8 and IL-10 may be early predictors of bronchopulmonary dysplasia in preterm newborns small for gestational age. Dis Markers. 2012; 33(1): 51–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Shim YJ, Choi BY, Park KH, Lee H, Jung YM, Kim YM. Inflammatory and immune proteins in umbilical cord blood: association with hearing screening test failure in preterm neonates. Mediators Inflamm. 2018; 2018: 4209359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Takahashi N, Uehara R, Kobayashi M, et al.. Cytokine profiles of seventeen cytokines, growth factors and chemokines in cord blood and its relation to perinatal clinical findings. Cytokine. 2010; 49(3): 331–337. [DOI] [PubMed] [Google Scholar]
- 23. Olsson KW, Larsson A, Jonzon A, Sindelar R. Exploration of potential biochemical markers for persistence of patent ductus arteriosus in preterm infants at 22–27 weeks’ gestation. Pediatr Res. 2019; 86(3): 333–338. [DOI] [PubMed] [Google Scholar]
- 24. Woo SJ, Park KH, Lee SY, et al.. The relationship between cord blood cytokine levels and perinatal factors and retinopathy of prematurity: a gestational age-matched case-control study. Invest Ophthalmol Vis Sci. 2013; 54(5): 3434–3439. [DOI] [PubMed] [Google Scholar]
- 25. Madan A, El-Ferzli G, Carlson SM, et al.. A potential biomarker in the cord blood of preterm infants who develop retinopathy of prematurity. Pediatr Res. 2007; 61(2): 215–221. [DOI] [PubMed] [Google Scholar]
- 26. Wang Y, Wu S, Yang Y, et al.. Differentially expressed miRNAs in oxygen-induced retinopathy newborn mouse models. Mol Med Rep. 2017; 15(1): 146–152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Olsson KW, Sindelar R.. Early biochemical markers associated with development of necrotizing enterocolitis—selected abstracts of the 2nd Congress of joint European Neonatal Societies (jENS 2017); Venice (Italy); October 31–November 4, 2017; Session “Neonatal Gastrointestinal Physiology and NEC.” J Pediatr Neonat Individualized Med. 2017; 6(2): e060244. [Google Scholar]
- 28. Early Treatment for Retinopathy of Prematurity Cooperative Group. Revised indications for the treatment of retinopathy of prematurity: results of the early treatment for retinopathy of prematurity randomized trial. Arch Ophthalmol. 2003; 121(12): 1684–1694. [DOI] [PubMed] [Google Scholar]
- 29. Holmström GE, Hellström A, Jakobsson PG, Lundgren P, Tornqvist K, Wallin A. Swedish National Register for Retinopathy of Prematurity (SWEDROP) and the evaluation of screening in Sweden. Arch Ophthalmol. 2012; 130(11): 1418–1424. [DOI] [PubMed] [Google Scholar]
- 30. Assarsson E, Lundberg M, Holmquist G, et al.. Homogenous 96-plex PEA immunoassay exhibiting high sensitivity, specificity, and excellent scalability. PLoS ONE. 2014; 9(4): e95192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. de Hoon MJL, Imoto S, Nolan J, Miyano S. Open source clustering software. Bioinformatics. 2004; 20(9): 1453–1454. [DOI] [PubMed] [Google Scholar]
- 32. Markasz L, Savani RC, Sedin G, Sindelar R. The receptor for hyaluronan-mediated motility (RHAMM) expression in neonatal bronchiolar epithelium correlates negatively with lung air content. Early Hum Dev. 2018; 127: 58–68. [DOI] [PubMed] [Google Scholar]
- 33. Saldanha AJ. Java Treeview—extensible visualization of microarray data. Bioinformatics. 2004; 20(17): 3246–3248. [DOI] [PubMed] [Google Scholar]
- 34. UniProt. Available at: https://www.uniprot.org/. Accessed November 7, 2019.
- 35. Sood BG, Madan A, Saha S, et al.. Perinatal systemic inflammatory response syndrome and retinopathy of prematurity. Pediatr Res. 2010; 67(4): 394–400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Hijioka M, Inden M, Yanagisawa D, Kitamura Y. DJ-1/PARK7: a new therapeutic target for neurodegenerative disorders. Biol Pharm Bull. 2017; 40(5): 548–552. [DOI] [PubMed] [Google Scholar]
- 37. Xu J, Zhong N, Wang H, et al.. The Parkinson's disease–associated DJ-1 protein is a transcriptional co-activator that protects against neuronal apoptosis. Hum Mol Genet. 2005; 14(9): 1231–1241. [DOI] [PubMed] [Google Scholar]
- 38. Zeng J, Zhao H, Chen B. DJ-1/PARK7 inhibits high glucose-induced oxidative stress to prevent retinal pericyte apoptosis via the PI3K/AKT/mTOR signaling pathway. Exp Eye Res. 2019; 189: 107830. [DOI] [PubMed] [Google Scholar]
- 39. Hellström M, Gerhardt H, Kalén M, et al.. Lack of pericytes leads to endothelial hyperplasia and abnormal vascular morphogenesis. J Cell Biol. 2001; 153(3): 543–553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Choi SH, Chung M, Park SW, Jeon NL, Kim JH, Yu YS. Relationship between pericytes and endothelial cells in retinal neovascularization: a histological and immunofluorescent study of retinal angiogenesis. Korean J Ophthalmol. 2018; 32(1): 70–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Greco TM, Seeholzer SH, Mak A, Spruce L, Ischiropoulos H. Quantitative mass spectrometry–based proteomics reveals the dynamic range of primary mouse astrocyte protein secretion. J Proteome Res. 2010; 9(5): 2764–2774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Dave JM, Bayless KJ.. Vimentin as an integral regulator of cell adhesion and endothelial sprouting. Microcirculation. 2014; 21(4): 333–344. [DOI] [PubMed] [Google Scholar]
- 43. Selvam S, Kumar T, Fruttiger M. Retinal vasculature development in health and disease. Prog Retin Eye Res. 2018; 63: 1–19. [DOI] [PubMed] [Google Scholar]
- 44. Shigyo M, Kuboyama T, Sawai Y, Tada-Umezaki M, Tohda C. Extracellular vimentin interacts with insulin-like growth factor 1 receptor to promote axonal growth. Sci Rep. 2015; 5: 12055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Hellström A, Perruzzi C, Ju M, et al.. Low IGF-I suppresses VEGF-survival signaling in retinal endothelial cells: direct correlation with clinical retinopathy of prematurity. Proc Natl Acad Sci U S A. 2001; 98(10): 5804–5808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Schiffers PM, Henrion D, Boulanger CM, et al.. Altered flow-induced arterial remodeling in vimentin-deficient mice. Arterioscler Thromb Vasc Biol. 2000; 20(3): 611–616. [DOI] [PubMed] [Google Scholar]
- 47. Lundkvist A, Reichenbach A, Betsholtz C, Carmeliet P, Wolburg H, Pekny M. Under stress, the absence of intermediate filaments from Müller cells in the retina has structural and functional consequences. J Cell Sci. 2004; 117(pt 16): 3481–3488. [DOI] [PubMed] [Google Scholar]
- 48. Wang X, Tang K, Chen L, Cheng S, Xu H. Association between sepsis and retinopathy of prematurity: a systematic review and meta-analysis. BMJ Open. 2019; 9(5): e025440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Bulka CM, Dammann O, Santos HP, et al.. Placental CpG methylation of inflammation, angiogenic, and neurotrophic genes and retinopathy of prematurity. Invest Ophthalmol Vis Sci. 2019; 60(8): 2888–2894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Augustin AJ, Breipohl W, Böker T, Lutz J, Spitznas M. Increased lipid peroxide levels and myeloperoxidase activity in the vitreous of patients suffering from proliferative diabetic retinopathy. Graefes Arch Clin Exp Ophthalmol. 1993; 231(11): 647–650. [DOI] [PubMed] [Google Scholar]
- 51. Tolsma KW, Allred EN, Chen ML, Duker J, Leviton A, Dammann O. Neonatal bacteremia and retinopathy of prematurity: the ELGAN study. Arch Ophthalmol. 2011; 129(12): 1555–1563. [DOI] [PubMed] [Google Scholar]
- 52. Winterbourn CC, Vissers MC, Kettle AJ. Myeloperoxidase. Curr Opin Hematol. 2000; 7(1): 53–58. [DOI] [PubMed] [Google Scholar]
- 53. Aratani Y. Myeloperoxidase: its role for host defense, inflammation, and neutrophil function. Arch Biochem Biophys. 2018; 640: 47–52. [DOI] [PubMed] [Google Scholar]
- 54. Odobasic D, Kitching AR, Holdsworth SR. Neutrophil-mediated regulation of innate and adaptive immunity: the role of myeloperoxidase. J Immunol Res. 2016; 2016: 2349817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Odobasic D, Kitching AR, Yang Y, et al.. Neutrophil myeloperoxidase regulates T-cell-driven tissue inflammation in mice by inhibiting dendritic cell function. Blood. 2013; 121(20): 4195–4204. [DOI] [PubMed] [Google Scholar]
- 56. Kutter D, Devaquet P, Vanderstocken G, Paulus JM, Marchal V, Gothot A. Consequences of total and subtotal myeloperoxidase deficiency: risk or benefit ? Acta Haematol. 2000; 104(1): 10–15. [DOI] [PubMed] [Google Scholar]
- 57. Laguna T, Notario L, Pippa R, et al.. New insights on the transcriptional regulation of CD69 gene through a potent enhancer located in the conserved non-coding sequence 2. Mol Immunol. 2015; 66(2): 171–179. [DOI] [PubMed] [Google Scholar]
- 58. Brait VH, Miró-Mur F, Pérez-de-Puig I, et al.. CD69 plays a beneficial role in ischemic stroke by dampening endothelial activation. Circ Res. 2019; 124(2): 279–291. [DOI] [PubMed] [Google Scholar]
- 59. Falk MK, Singh A, Faber C, Nissen MH, Hviid T, Sørensen TL. Dysregulation of CXCR3 expression on peripheral blood leukocytes in patients with neovascular age-related macular degeneration. Invest Ophthalmol Vis Sci. 2014; 55(7): 4050–4056. [DOI] [PubMed] [Google Scholar]
- 60. Yates-Binder CC, Rodgers M, Jaynes J, Wells A, Bodnar RJ, Turner T. An IP-10 (CXCL10)–derived peptide inhibits angiogenesis. PLoS ONE. 2012; 7(7): e40812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Maubach G, Schmädicke A-C, Naumann M. NEMO links nuclear factor-κB to human diseases. Trends Mol Med. 2017; 23(12): 1138–1155. [DOI] [PubMed] [Google Scholar]
- 62. Smahi A, Courtois G, Vabres P, et al.. Genomic rearrangement in NEMO impairs NF-kappaB activation and is a cause of incontinentia pigmenti. The International Incontinentia Pigmenti (IP) Consortium. Nature. 2000; 405(6785): 466–472. [DOI] [PubMed] [Google Scholar]
- 63. O'Doherty M, Mc Creery K, Green AJ, Tuwir I, Brosnahan D. Incontinentia pigmenti—ophthalmological observation of a series of cases and review of the literature. Br J Ophthalmol. 2011; 95(1): 11–16. [DOI] [PubMed] [Google Scholar]
- 64. Araki Y, Abe Y, Takeda Y, et al.. Incontinentia pigmenti with retinal vascular anomaly and deletion of exons 4-10 in NEMO. J Dermatol. 2017; 44(8): 976–977. [DOI] [PubMed] [Google Scholar]
- 65. Holmström G, Thorén K.. Ocular manifestations of incontinentia pigmenti. Acta Ophthalmol Scand. 2000; 78(3): 348–353. [DOI] [PubMed] [Google Scholar]
- 66. So T, Ishii N.. The TNF-TNFR family of co-signal molecules. Adv Exp Med Biol. 2019; 1189: 53–84. [DOI] [PubMed] [Google Scholar]
- 67. Stabile E, Burnett MS, Watkins C, et al.. Impaired arteriogenic response to acute hindlimb ischemia in CD4-knockout mice. Circulation. 2003; 108(2): 205–210. [DOI] [PubMed] [Google Scholar]
- 68. Stabile E, Kinnaird T, la Sala A, et al.. CD8+ T lymphocytes regulate the arteriogenic response to ischemia by infiltrating the site of collateral vessel development and recruiting CD4+ mononuclear cells through the expression of interleukin-16. Circulation. 2006; 113(1): 118–124. [DOI] [PubMed] [Google Scholar]
- 69. Ohshima Y, Tanaka Y, Tozawa H, Takahashi Y, Maliszewski C, Delespesse G. Expression and function of OX40 ligand on human dendritic cells. J Immunol. 1997; 159(8): 3838–3848. [PubMed] [Google Scholar]
- 70. Stüber E, Neurath M, Calderhead D, Fell HP, Strober W. Cross-linking of OX40 ligand, a member of the TNF/NGF cytokine family, induces proliferation and differentiation in murine splenic B cells. Immunity. 1995; 2(5): 507–521. [DOI] [PubMed] [Google Scholar]
- 71. Weinberg AD, Wegmann KW, Funatake C, Whitham RH. Blocking OX-40/OX-40 ligand interaction in vitro and in vivo leads to decreased T cell function and amelioration of experimental allergic encephalomyelitis. J Immunol. 1999; 162(3): 1818–1826. [PubMed] [Google Scholar]
- 72. Imura A, Hori T, Imada K, et al.. The human OX40/gp34 system directly mediates adhesion of activated T cells to vascular endothelial cells. J Exp Med. 1996; 183(5): 2185–2195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Cunningham MA, Li Z, Liu B, Yeh S, Nussenblatt RB. OX40 ligand expression abrogates the immunosuppressive function of retinal pigment epithelium. J Ophthalmic Inflamm Infect. 2013; 3(1): 12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Jacquemin C, Schmitt N, Contin-Bordes C, et al.. OX40 ligand contributes to human lupus pathogenesis by promoting T follicular helper response. Immunity. 2015; 42(6): 1159–1170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Lu S, Song S, Hou S, Li H, Yang P. Association of TNFSF4 polymorphisms with Vogt-Koyanagi-Harada and Behcet's disease in Han Chinese. Sci Rep. 2016; 6: 37257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Qin W, Hongya W, Yongjing C, et al.. Increased OX40 and soluble OX40 ligands in children with Henoch-Schonlein purpura: association with renal involvement. Pediatr Allergy Immunol. 2011; 22(1, pt 1): 54–59. [DOI] [PubMed] [Google Scholar]
- 77. Byun M, Ma CS, Akçay A, et al.. Inherited human OX40 deficiency underlying classic Kaposi sarcoma of childhood. J Exp Med. 2013; 210(9): 1743–1759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Gong M-T, Li W-X, Zhang Q, et al.. Comprehensive analysis of gene expression profiles associated with proliferative diabetic retinopathy. Exp Ther Med. 2018; 16(4): 3539–3545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Nakano M, Fukumoto Y, Satoh K, et al.. OX40 ligand plays an important role in the development of atherosclerosis through vasa vasorum neovascularization. Cardiovasc Res. 2010; 88(3): 539–546. [DOI] [PubMed] [Google Scholar]
- 80. Leibovich SJ, Polverini PJ, Shepard HM, Wiseman DM, Shively V, Nuseir N. Macrophage-induced angiogenesis is mediated by tumour necrosis factor-alpha. Nature. 1987; 329(6140): 630–632. [DOI] [PubMed] [Google Scholar]
- 81. Yan J, Wang C, Du R, Liu P, Chen G. OX40-OX40 ligand interaction may activate phospholipase C signal transduction pathway in human umbilical vein endothelial cells. Chem Biol Interact. 2009; 180(3): 460–464. [DOI] [PubMed] [Google Scholar]
- 82. Kim MJ, Kim E, Ryu SH, Suh PG. The mechanism of phospholipase C-gamma1 regulation. Exp Mol Med. 2000; 32(3): 101–109. [DOI] [PubMed] [Google Scholar]
- 83. Takahashi T, Yamaguchi S, Chida K, Shibuya M. A single autophosphorylation site on KDR/Flk-1 is essential for VEGF-A-dependent activation of PLC-gamma and DNA synthesis in vascular endothelial cells. EMBO J. 2001; 20(11): 2768–2778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Moasser MM. The oncogene HER2: its signaling and transforming functions and its role in human cancer pathogenesis. Oncogene. 2007; 26(45): 6469–6487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85. Koukourakis MI, Manolas C, Minopoulos G, Giatromanolaki A, Sivridis E. Angiogenesis relates to estrogen receptor negativity, c-erbB-2 overexpression and early relapse in node-negative ductal carcinoma of the breast. Int J Surg Pathol. 2003; 11(1): 29–34. [DOI] [PubMed] [Google Scholar]
- 86. Russell KS, Stern DF, Polverini PJ, Bender JR. Neuregulin activation of ErbB receptors in vascular endothelium leads to angiogenesis. Am J Physiol. 1999; 277(6): H2205–2211. [DOI] [PubMed] [Google Scholar]
- 87. Yang W, Klos K, Yang Y, Smith TL, Shi D, Yu D. ErbB2 overexpression correlates with increased expression of vascular endothelial growth factors A, C, and D in human breast carcinoma. Cancer. 2002; 94(11): 2855–2861. [DOI] [PubMed] [Google Scholar]
- 88. Schoppmann SF, Tamandl D, Roberts L, et al.. HER2/neu expression correlates with vascular endothelial growth factor-C and lymphangiogenesis in lymph node-positive breast cancer. Ann Oncol. 2010; 21(5): 955–960. [DOI] [PubMed] [Google Scholar]
- 89. Zhang A, Shen G, Zhao T, et al.. Augmented inhibition of angiogenesis by combination of HER2 antibody chA21 and trastuzumab in human ovarian carcinoma xenograft. J Ovarian Res. 2010; 3: 20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90. Petit AM, Rak J, Hung MC, et al.. Neutralizing antibodies against epidermal growth factor and ErbB-2/neu receptor tyrosine kinases down-regulate vascular endothelial growth factor production by tumor cells in vitro and in vivo: angiogenic implications for signal transduction therapy of solid tumors. Am J Pathol. 1997; 151(6): 1523–1530. [PMC free article] [PubMed] [Google Scholar]
- 91. Laughner E, Taghavi P, Chiles K, Mahon PC, Semenza GL. HER2 (neu) signaling increases the rate of hypoxia-inducible factor 1α (HIF-1α) synthesis: novel mechanism for HIF-1-mediated vascular endothelial growth factor expression. Mol Cell Biol. 2001; 21(12): 3995–4004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Vadlapatla RK, Vadlapudi AD, Mitra AK. Hypoxia-inducible factor-1 (HIF-1): a potential target for intervention in ocular neovascular diseases. Curr Drug Targets. 2013; 14(8): 919–935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93. Webling KEB, Runesson J, Bartfai T, Langel U. Galanin receptors and ligands. Front Endocrinol (Lausanne). 2012; 3: 146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94. Schrödl F, Kaser-Eichberger A, Trost A, et al.. Distribution of galanin receptors in the human eye. Exp Eye Res. 2015; 138: 42–51. [DOI] [PubMed] [Google Scholar]
- 95. Lang R, Kofler B.. The galanin peptide family in inflammation. Neuropeptides. 2011; 45(1): 1–8. [DOI] [PubMed] [Google Scholar]
- 96. Brooke-Smith ME, Carati CJ, Bhandari M, Toouli J, Saccone GTP. Galanin in the regulation of pancreatic vascular perfusion. Pancreas. 2008; 36(3): 267–273. [DOI] [PubMed] [Google Scholar]
- 97. Cordero-Llana O, Rinaldi F, Brennan PA, Wynick D, Caldwell MA. Galanin promotes neuronal differentiation from neural progenitor cells in vitro and contributes to the generation of new olfactory neurons in the adult mouse brain. Exp Neurol. 2014; 256: 93–104. [DOI] [PubMed] [Google Scholar]
- 98. Yamamoto H, Okada R, Tanaka R, Unno K, Iguchi K. Expression of a urokinase-type plasminogen activator during tumor growth leads to angiogenesis via galanin activation in tumor-bearing mice. FEBS Open Bio. 2017; 7(11): 1784–1792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99. Banerjee R, Van Tubergen EA, Scanlon CS, et al.. The G protein-coupled receptor GALR2 promotes angiogenesis in head and neck cancer. Mol Cancer Ther. 2014; 13(5): 1323–1333. [DOI] [PMC free article] [PubMed] [Google Scholar]