

Abstract
Lymphatic filariasis (LF) is a disease caused by parasitic filarial nematodes that is endemic in 49 countries of the world and affects or threatens over 890 million people. Strategies to control LF rely heavily on mass administration of anthelmintic drugs including ivermectin (IVM), a macrocyclic lactone drug considered an Essential Medicine by the WHO. However, despite its widespread use the therapeutic mode of action of IVM against filarial nematodes is not clear. We have previously reported that filarial nematodes secrete extracellular vesicles (EVs) and that their cargo has immunomodulatory properties. Here we investigate the effects of IVM and other anti‐filarial drugs on parasitic nematode EV secretion, motility, and protein secretion. We show that inhibition of EV secretion was a specific property of IVM, which had consistent and significant inhibitory effects across nematode life stages and species, with the exception of male parasites. IVM inhibited EV secretion, but not parasite motility, at therapeutically relevant concentrations. Protein secretion was inhibited by IVM in the microfilariae stage, but not in any other stage tested. Our data provides evidence that inhibiting the secretion of immunomodulatory EVs by parasitic nematodes could explain, at least in part, IVM mode of action and provides a phenotype for novel drug discovery.
Keywords: Brugia malayi, extracellular vesicle, ivermectin, lymphatic filariasis, macrocyclic lactone, parasite
1. INTRODUCTION
Lymphatic filariasis (LF) is a neglected tropical disease caused by thread‐like parasitic filarial nematodes, including Wuchereria bancrofti (responsible for approximately 90% of LF cases) and Brugia malayi, that establish in the lymphatic vasculature. LF is often subclinical but symptoms manifest in approximately 40% of cases with lymphedema, hydrocoele, dermatitis and long‐term disability characterizing clinical disease. It is estimated that LF is endemic in 49 countries and that over 890 million people are infected or at risk of infection (World Health Organization, 2019). In 2000 the Global Programme to Eliminate Lymphatic Filariasis was created with the goal of eliminating this disease by 2020 and although there has been reduction in the prevalence of LF in some areas, this disease is far from being eliminated. Strategies to control LF and other filarial parasitic nematode infections rely heavily on mass administration of the anthelmintic drugs ivermectin (IVM), albendazole (ABZ) and diethylcarbamazine (DEC) in endemic areas. The well‐established potential for adverse reactions to DEC alone or in combination with ABZ emphasizes the importance of IVM to LF chemotherapeutic control strategies. IVM is classified as an essential medication by the World Health Organization (World Health Organization, 2019) and since 2000, over 7 billion treatments have been delivered to at risk individuals (World Health Organization, 2019); however, this disease still remains an issue. One challenge to eliminating LF centres on the inadequate drugs that are currently available; neither IVM, ABZ or DEC effectively kill adult parasites, thus established infections are incurable (Ottesen, 2006). Compounding this and despite their widespread use, the therapeutic modes of action of IVM, and to a lesser extent DEC, are not entirely clear.
A current working hypothesis for the therapeutic activity of IVM is that it inhibits the release of excretory‐secretory (ES) products from parasites. This inhibition is postulated to prevent active host immunomodulation by the parasite via these ES products allowing for host recognition and parasite clearance (Moreno et al., 2010). In support is the acceptance that the host immune system is involved in filarial parasite elimination, especially in the clearance of microfilaria (mf) stage worms (Carithers, 2017; Wolstenholme, Maclean, Coates, McCoy, & Reaves, 2016) and data from experiments such as Berrafato et al. showing that IVM enhanced leukocyte binding to Dirofilaria immitis mf and Semnani et al who showed that IVM could reverse the modified Th2 phenotypes caused in filaria infected patients (Berrafato, Coates, Reaves, Kulke, & Wolstenholme, 2019; Semnani et al., 2006). There is widespread support that ES products from filarial nematodes do modulate host immune responses. Early filarial nematode infection elicits a canonical Th2 immune response characterized by increased production of the cytokines interleukin (IL)‐4, IL‐5, IL‐9, IL‐10 and IL‐13 and the antibody isotypes IgG1, IgG4 (in humans) and IgE and increased production of Th2 cells, eosinophils, alternatively activated macrophages, and innate lymphoid cells 2 (ILC2) (Allen & Maizels, 2011; Geary et al., 2010). With development of chronic filarial infection the Th2 response becomes 'modified' to a more tolerant and regulatory environment with increased IL‐4, IL‐10, Treg and alternatively activated macrophage proliferation and reduction in IL‐5, IL‐13 and T cell proliferation coupled with T cell anergy and decreased antigen presenting capabilities (Babu & Nutman, 2014). There is considerable evidence that filarial nematode parasites contribute to this 'modified' phenotype but the exact parasite factors driving this manipulation remain uncertain.
Extracellular vesicles (EVs) are membrane‐bound vesicles secreted into the extracellular environment by eukaryotic and prokaryotic cells. Although once thought to be carriers of waste products, it has been shown that EVs function in many physiological processes and are important mediators of cell‐to‐cell signalling (Bobrie, Colombo, Raposo, & Théry, 2011; Lee, EL Andaloussi, & Wood, 2012; Raposo et al., 1996; Valadi et al., 2007). EVs are considered a heterogenous group of sub‐cellular structures that can be subdivided based on size and biogenesis. Primary focus has been on two subsets of EVs, microvesicles that range from 150 to 1500 nm and a smaller grouping (30–150 nm) originally termed exosomes (Johnstone, Adam, Hammond, Orr, & Turbide, 1987). Exosomes are products of the endosomal pathway and are derived from multivesicular bodies (MVBs) that fuse with the cell membrane to secrete the vesicles into the extracellular space (Catalano & O'Driscoll, 2020; Riaz & Cheng, 2017; Vlassov, Magdaleno, Setterquist, & Conrad, 2012). Consistent with a role in cell‐to‐cell communication, EVs contain diverse functional cargo that varies depending on the cellular origin of the EVs, but in general include bioactive proteins, RNA and lipids (Thery, Zitvogel, & Amigorena, 2002; Valadi et al., 2007). EV secretion from diverse parasitic nematodes has been described (Buck et al., 2014; Coakley et al., 2017; Eichenberger et al., 2018; Eichenberger et al., 2018; Gu et al., 2017; Hansen, Kringel, Williams, & Nejsum, 2015, 2019; Harischandra, Yuan, Loghry, Zamanian, & Kimber, 2018; Shears, Bancroft, Hughes, Grencis, & Thornton, 2018; Tritten et al., 2017; Tzelos et al., 2016; Zamanian et al., 2015) and the cargo of these EVs have immunomodulatory functions (Buck et al., 2014; Quintana, Babayan, & Buck, 2017; Tritten, Clarke, Timmins, McTier, & Geary, 2016). We have previously reported that B. malayi secretes EVs and that their cargo has putative immunomodulatory properties (Harischandra et al., 2018; Zamanian et al., 2015). Driven by these emerging data, EVs have been proposed as a potential mechanism by which parasites modulate host immune responses (Buck et al., 2014; Coakley et al., 2017; Eichenberger, Sotillo, & Loukas, 2018; Harischandra et al., 2018).
We propose that nematode EVs are essential for filarial nematode parasitism and hypothesize that effective anti‐filarial drugs inhibit their secretion. To investigate this hypothesis, a panel of anti‐filarial drugs was screened for their ability to reduce EV secretion from parasitic nematodes. We found that IVM had the most consistent inhibitory effects on EV secretion by various parasite species and life stages of Brugia malayi. Importantly, IVM had insignificant effects on motility and limited effects on protein secretion at therapeutically relevant concentrations and timepoints. These observations provide insight into the mechanism of action of IVM and may support prioritizing inhibition of EV secretion as a screenable phenotype for novel anti‐filarial drug development.
2. MATERIALS AND METHODS
2.1. 2.1 Parasite culture and maintenance
Brugia malayi and B. pahangi parasites were obtained from the NIH/NIAID Filariasis Research Reagent Resource Center (FR3) at the University of Georgia, USA. Persistent B. malayi infections at FR3 are maintained in domestic short‐haired cats. To obtain adult stage B. malayi, jirds were infected intraperitoneally with approximately 400 L3 stage parasites. A total of 120 days post‐infection jirds were necropsied to collect adult stage parasites. L3 stage B. malayi were obtained from dissection of anesthetized Aedes aegypti 14 days post‐infection. Microfilaria stage B. malayi were obtained from a lavage of the peritoneal cavity of a euthanized gerbil. B. pahangi stages were obtained in the same manner as B. malayi with the exception that infective L3 stage parasites were collected 11 days and 16 days post‐infection, respectively. B. malayi parasites were also obtained from TRS Labs LLC (Athens, Georgia, USA). These parasites were tested and responded to treatments in the same manner as parasites from FR3. Upon receipt at ISU, all B. malayi and B. pahangi parasites were washed several times in warmed worm culture media (RPMI with 1% HEPES, 1% L‐glutamine, 0.2% Penicillin/Streptomycin, and 1% w/v glucose [all Thermo Fisher Scientific, Waltham, MA, USA]) and then counted and cultured at 37°C with 5% CO2. Adult female Ascaris suum were collected from an abattoir in Marshalltown, Iowa, USA. These parasites were washed multiple times in warmed Ascaris Ringer's Solution (13.14 mM NaCl, 9.67 mM CaCl2, 7.83 mM MgCl2, 12.09 mM C4H11NO3, 99.96 mM C2H3NaO2, 19.64 mM KCl with Gentamycin (100 μg/ml), Ciprofloxacin Hydrochloride (20 μg/ml), penicillin (10,000 units/ml), streptomycin (10,000 μg/ml), and Amphotericin B (25 μg/ml) at pH 7.87 [all Sigma‐Aldrich, St Louis, MO, USA]) and then incubated overnight at 34°C. After 24 h in culture the parasites were checked for visible signs of bacterial or fungal contamination; if present the parasites were discarded.
2.2. Drug treatments of parasites
Parasites were cultured in the presence or absence of drug to examine effects on extracellular vesicle (EV) secretion. For B. malayi and B. pahangi, 10 adult female and 10 adult males were cultured as previously described for 24 h in 10 and 3 ml culture media, respectively, in 15 ml polypropylene centrifuge tubes (Thermo Fisher Scientific). 100 L3 or 1 × 106 microfilariae were cultured as previously described for 24 h in 1 ml culture media in 1.5 ml microcentrifuge tubes (Thermo Fisher Scientific). Single adult female A. suum were cultured in 100 ml culture media in 250 ml sterile Erlenmeyer flask for 24 h as previously described. Four drugs, ivermectin, albendazole, diethylcarbamazine, and levamisole (all Sigma‐Aldrich) were investigated for their effects on each life stage of the parasite species. The various drugs or DMSO (vehicle control) were added to the culture media at a final concentration of 1 μM and 0.01% respectively. This dose was chosen for initial screening purposes because it is at the high end of the therapeutically relevant concentration range for each drug under examination and therefore should be sufficient to manifest a phenotype. If this dose did elicit a phenotype, then a full concentration response was established. Spent media was collected after a 24 h incubation. Additionally, drug and control treated A. suum and B. pahangi media was collected at 2, 4, 6 and 12‐h intervals to investigate the time course of the effects of the drugs. A dose curve for the effects of ivermectin on B. malayi were conducted in the same manner as described above with concentrations ranging from 0.1 Nm to 10 μM. Unless otherwise stated, all drug treatments were performed at 37°C with 5% CO2.
2.3. EV isolation and quantification
EVs were collected as previously described using differential ultracentrifugation (Harischandra et al., 2018; Zamanian et al., 2015). Media was filtered through 0.2 μm PVDF filtered syringes (GE Healthcare, Chicago, IL) or PVDF vacuum filters (Sigma‐Aldrich) and centrifuged at 120,000 x g for 90 min at 4°C. The supernatant was decanted leaving approximately 1.5 ml media to ensure that the EV pellet was not disrupted. The retained media and pellet were filtered through a PVDF 0.2 μm syringe filter and centrifuged at 186,000 x g for a further 2 h at 4°C. Pelleted EV samples were resuspended to 500 μl in dPBS (Thermo Fisher Scientific). EV quantification and size determination were performed using nanoparticle tracking analysis (NTA; Nano‐Sight LM10, Malvern Instruments, Malvern, UK).
2.4. Motility analysis
The Worminator system developed and described by Marcellino et al. (Marcellino et al., 2012) was used to quantitatively measure motility of adult filarial nematodes in microtiter plates. Microscopic parasite life stages were quantitatively analysed by the same software, but with methods previously described by Storey et al. (2014). Briefly, single adult male and female worms were cultured individually in one well of a standard 24‐well cell culture plate (Sigma‐Aldrich). For infective L3 stage worms, 10 worms were cultured per well of a 96‐well plate (Corning Inc, Corning, NY, USA). Drug or DMSO (vehicle control) was added to each well to a final concentration of 1 μM or 0.01% respectively. Worms were incubated at 37°C and 5% CO2 and measurements were briefly taken, at room temperature, prior to treatment, immediately after treatment (0 h) and at 2, 4, 6 and 24‐h post treatment. Measurements of the effects of doses of ivermectin ranging from 0.1 Nm to 10 μM on adult female B. malayi were conducted at 24 h post treatment.
2.5. Protein Quantification Assay
A single B. malayi adult or 100,000 microfilariae were cultured per well of a 24‐well plate with either drug or DMSO at a final concentration of 1.0 μM or 0.01%, respectively, for 24 h. A concentration‐response curve for ivermectin was conducted on adult female worms with concentrations ranging from 0.1 nM to 10 μM. Spent media was collected and filtered through a 0.2 μM PVDF membrane filter (GE Healthcare) 500 μl of media were concentrated using a 0.5 ml, 3000 Da Amicon Ultra centrifugal filter unit (Sigma‐Aldrich) according to manufacturer's instructions. Media samples were concentrated by centrifuging at 14,000 x g for 30 min. Samples were then washed with 500 μl dPBS for 30 min at 14,000 x g. The washing step was repeated four times. The volume of each sample was determined, and all samples were normalized to 150 μl with dPBS. Adult female samples were then further diluted four‐fold with dPBS while adult male, L3 stage and microfilariae were diluted 2‐fold with dPBS to ensure that readings would fall within the standard curve. Total protein was quantified with Pierce micro BCA kit (Thermo Fisher Scientific) according to manufacturers’ instructions. Protein assay plates were quantified using a SpectraMax M2e plate reader (Molecular Devices, San Jose, CA, USA).
2.6. Statistical analysis
Due to some variation among individual parasites and between batches of parasites, experiments were conducted across multiple batches of parasite shipments, with each N representing parasites from independent shipments. Individual control and treated worms within each batch were paired together to help account for batch variation. EV NTA data were analysed via a ratio‐paired T‐test with P‐values < 0.05 being considered significant. ANOVA/mixed effects model analysis was used to compare the means of EV size profiles following drug treatment with statistical significance determined using post‐hoc Šídák test (P‐values < 0.05 being considered significant). Non‐linear regressions with least squares fit were used to analyse the dose response curves for ivermectin on B. malayi adult female EV secretion, motility, and protein secretion. Motility data was analysed via a RM 2‐way ANOVA with Geisser‐Greenhouse correction followed by a Dunnet's multiple comparison test. Paired T‐test between each treatment for each life stage were used to analyse data from the protein assay as the data contained values unsuited for a ratio‐paired T‐test. Due to the variability among batches and individual parasites the ROUT outlier identification method was used to identify outliers in the data (Q = 0.5%). All statistical analyses were performed using Prism 8.4.1. (GraphPad Software, San Diego, CA, USA).
3. RESULTS
3.1. Ivermectin inhibits EV secretion from Brugia malayi in a sex‐ and stage‐ specific manner
In this study we investigated the effects of ivermectin (IVM) on B. malayi EV secretion in vitro. Parasites were cultured at 37°C in the presence or absence of IVM and the number of EVs secreted by the worms was quantified by nanoparticle tracking analysis (NTA) (Figure 1) and visually confirmed as EVs using electron microscopy (Figure S1). An initial screening concentration of 1.0 μM IVM significantly reduced EV secretion after 24 h incubation from B. malayi adult females by 59% (P = 0.0204, N = 13) and from L3 stage parasites by 31% (P = 0.0067, N = 19) compared to vehicle controls. There was no significant effect on EV secretion from adult male (P = 0.4028, N = 10) or microfilariae (mf) (P = 0.2081, N = 13) life stages (Figure 1A‐D). The IVM concentration response in adult female B. malayi was further profiled and the IC50 determined to be 7.7 nM (Figure 1E). Studies conducted on the pharmacokinetics of a single dose of IVM in human subjects have determined serum levels to be between 20 and 70 nM (González Canga et al., 2008). IVM therefore inhibits EV secretion in adult female parasites at therapeutically relevant concentrations suggesting that this phenomenon may contribute to IVM therapeutic mode of action. It is important to recognize that some portion of the EVs secreted by adult female B. malayi may originate from mf worms in uteri. However, the observation that 1.0 μM IVM significantly inhibited EV release from adult females at 37°C, but did not significantly inhibit EV release from mfs at the same temperature, suggests the inhibitory effect of IVM against adult females is due to bioactivity at that life stage and not mfs in uteri.
Figure 1.
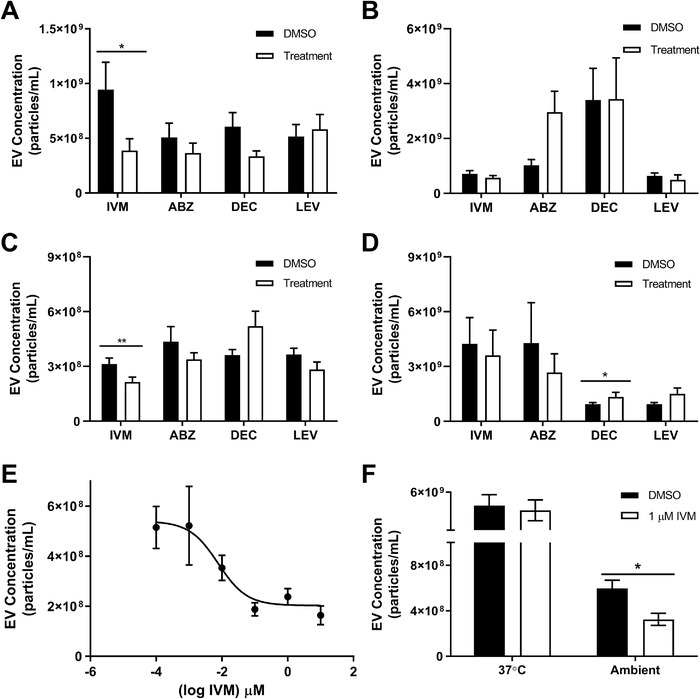
Ivermectin inhibits Brugia malayi EV secretion in a stage‐ and sex‐ specific manner. B. malayi life stages were cultured at 37°C in RPMI with either drug (1.0 μM) or DMSO (vehicle control). Media was collected after 24 h and EVs were isolated and quantified. 1.0 μM IVM significantly reduced EV secretion from adult female worms (A) and L3 stage parasites (C) but not from adult males (B) or microfilaria (D) Albendazole (ABZ), diethylcarbamazine (DEC) or levamisole (LEV) had no effect on EV secretion from any life stage except in microfilaria, where DEC increased EV secretion. (E) The IC50 for IVM on adult female worms was determined to be 7.7 nM. (F) 1 μM IVM has temperature dependent effects on EV secretion from B. malayi microfilariae with inhibition occurring at 22°C, but not at 37°C. N = 7 (minimum), Mean ± SEM, *P < 0.05, **P < 0.01. ■ = DMSO control, □ = Treatment
These observations on the inhibitory effect of IVM on EV secretion from adult female and L3 stage B. malayi generally align with preliminary data previously reported (Harischandra et al., 2018) with the exception of the lack of inhibition in adult males and a reduced inhibition in mf stages. The previously reported inhibitory effect on EV secretion from adult males was marginal, but the lack of effect on mf is more surprising considering its prior robustness. Previously, mf were incubated with IVM at ambient temperature whereas here they were incubated at 37°C. To test the impact of temperature on the IVM phenotype in mf, we repeated the mf IVM incubation at ambient temperature. Unlike at 37°C, 1.0 μM IVM significantly inhibited EV secretion by 46% (P = 0.0177, N = 8) at ambient temperature (22°C) (Figure 1F). Control and treated parasites were still viable at the end of the experiment indicating that it was not loss of viability or death of the parasites that had caused inhibition of EV secretion. There are clear temperature‐dependent effects on EV secretion from mf stage worms, not only did incubation of mf at 37°C abrogate the inhibitory effect of IVM on EV secretion observed at ambient temperature, but it also increased EV secretion in untreated worms by approximately a factor of 7. Whilst logical to assume temperature changes impact worm physiology, there is a lack of data on the effects of temperature on specific processes and functions in mf stage nematodes. We do know that host temperature has no effect on the nocturnal periodicity of mf (Hawking, 1967) or on the ability of mf to bind to vascular endothelial cells (Schroeder et al., 2017). Environmental temperature may affect other physiological processes in mf leading to this increased production of EVs, but further investigation into this phenomenon is necessary.
To test the hypothesis that IVM treatment altered the size profile of secreted EVs, we compared the size‐distribution of EV populations secreted by IVM‐treated and DMSO‐treated parasites (Figure 2). We initially focused specifically on those B. malayi stages for which 1.0 μM significantly inhibited EV release – adult females and L3 stage worms. Secreted EVs were binned by size into 50 nm increments; the number of EVs in each bin was then totalled and the effect of 24 h drug treatment determined by comparing the means of EVs in each bin between control and IVM‐treated worms. In B. malayi adult females, 24 h 1.0 μM IVM treatment reduced secretion of 51–100 nm EVs by 59% (P = < 0.0001, N = 11) and 101–150 nm by 69% (P = 0.0093, N = 11) (Figure 2A). For B. malayi L3, 24 h 1.0 μM IVM treatment reduced secretion of 51–100 nm EVs by 41% (P = < 0.0001, N = 15) and 101–150 nm by 58% (P = < 0.0001, N = 15) (Figure 2B). The bioactivity of ivermectin to inhibit secretion of specific EV cohorts based on their size may reflect the effect of this drug on specific anatomical locations where EVs are generated or from where they are released. As summarized by Drurey, Coakley and Maizels (Drurey, Coakley, & Maizels, 2020), EV release from nematodes may occur from different structures throughout their lifecycle including the ES pore and gastrointestinal tract (both oral and anal routes). It is possible that inhibition of multiple discrete EV size ranges reflects IVM activity at one or more of these sites. The lack of a through gut in mf stage worms would imply only the ES pore is responsible in that stage and in B. pahangi mf, we observed the narrowest size distribution of inhibition (151–200 nm) (see Section 3.3).
Figure 2.
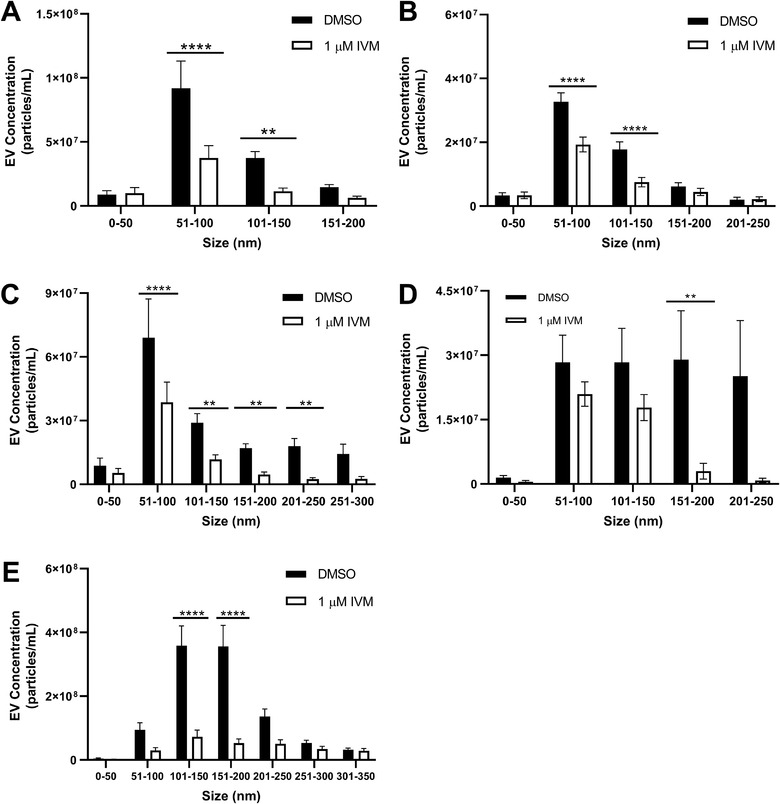
Secretion of distinct EV size subsets are inhibited by ivermectin. B. malayi and B. pahangi life stages were cultured at 37°C in RPMI, and Ascaris suum adult females were cultured at 34°C in Ascaris Ringers Solution with either 1.0 μM IVM or DMSO (vehicle control). Media was collected after 24 h and EVs were isolated and quantified. The size distributions of EVs secreted from parasites were analysed by nanoparticle tracking analysis. 1.0 μM IVM significantly inhibited secretion of 51–100 nm and 101–150 nm EVs and from both B. malayi adult females (A) and L3 (B) life stages. In B. pahangi, secretion of a broader range of EV sizes ranging from 51 to 250 nm was inhibited from adult females (C) while only a distinct subset of larger sized EVs (151‐200 nm) was inhibited from microfilariae (D). In adult female Ascaris suum 101–150 nm and 151–200 nm EV subsets were significantly inhibited (E). N = 6 (minimum).**P < 0.01, ****P < 0.0001.. ■ = DMSO, □ = 1 μM IVM
3.2. Other drugs with anti‐filarial activity do not inhibit EV secretion from B. malayi
To examine if inhibition of EV secretion is a general feature of drugs with anti‐filarial activity, we tested a panel of drugs with known anti‐filarial activity including albendazole (ABZ), diethylcarbamazine (DEC) and levamisole (LEV). LEV is a nicotinic agonist and although more typically used to treat gastrointestinal nematode infection, was included in the panel because it is an anthelmintic drug with known neuromuscular effects on filarial nematodes (Martin, 1997; Robertson, Buxton, & Martin, 2013) and also because of reported microfilaricidal effects on canine heartworm, D. immitis (Carlisle, Atwell, & Robinson, 1984; Mills & Amis, 1975). Parasites were cultured with or without an initial screening concentration of 1.0 μM drug and EVs were quantified using NTA. 1.0 μM DEC significantly increased EV secretion from B. malayi mf by 43% (P = 0.0177, N = 7) after 24 h (Figure 1D). DEC also seemed to increase EV secretion in L3 (Figure 1C) though not significantly (P = 0.2236, N = 10). Additionally, DEC had a moderate, but not significant, inhibition on EV secretion in adult females (P = 0.1704, N = 10) and had no effect on adult males (P = 0.2323, N = 9) (Figure 1A,B). There was a minor, but not significant, inhibition of EV secretion due to ABZ on B. malayi adult females (P = 0.4042, N = 10), L3 (P = 0.1564, N = 11), and mf (P = 0.7815, N = 7) (Figure 1A,C,D). In contrast, ABZ treatment seemed to increase EV secretion from adult males (Figure 1B) though not significantly (P = 0.0821, N = 9). LEV did not have an effect on EV secretion from either male (P = 0.1091, N = 11) or female adults (P = 0.8659, N = 9) (Figure 1A,B). L3 stage parasites showed a minor although not significant inhibition in EV secretion due to LEV treatment (P = 0.0719, N = 10) while mf had a moderate, but not significant increase in EV secretion due to LEV treatment (P = 0.0770, N = 7) (Figure 1C,D). In summary, none of the drugs in the panel significantly inhibited EV secretion from any of the B. malayi life stages when tested at 1.0 μM, except for DEC which significantly increased EV secretion from B. malayi microfilariae. This is significant because it suggests that inhibition of EV secretion by filarial nematodes may be a phenotype specific to IVM treatment and is not observed upon treatment with other anthelmintic drugs that are known to have anti‐filarial activity, including a drug (LEV) that has clear neuromuscular effects on filarial worms. This helps support the hypothesis that the mechanism of action of IVM, and perhaps other macrocyclic lactones, includes inhibition of EV secretion.
3.3. Ivermectin has broad inhibitory effects on EV secretion across other filarial and gastrointestinal nematode parasites
To test whether the inhibitory IVM phenotype in B. malayi was broadly consistent in nematodes, we repeated the same screening experiment with our same drug panel using first a related species of filarial nematode, B. pahangi. Our analysis was limited to B. pahangi adult females, adult males and mf based on worm availability. All drugs in the panel significantly inhibited EV secretion from B. pahangi adult female parasites (Figure 3A). IVM treatment had the greatest reduction in EV secretion with an inhibition of 63% (P = 0.0083, N = 10), while ABZ had an inhibition of 61% (P = 0.0066, N = 12), DEC by 59% (P = 0.0322, N = 12) and LEV by 44% (P = 0.0416, N = 11). There was no significant difference in magnitude of inhibition across the four drug treatments. The IVM, ABZ and DEC results generally paralleled the trends seen in B. malayi adult females, but with LEV now also active. Neither IVM (P = 0.9897, N = 11), ABZ (P = 0.9369, N = 13), DEC (P = 0.0925, N = 12), nor LEV (P = 0.7558, N = 11) had any effect on adult male B. pahangi (Figure 3B). Again, this is consistent with the results seen in B. malayi adult male parasites. In mf stage B. pahangi, IVM significantly reduced EV secretion by 40% at 37°C (P = 0.0358, N = 4) (Figure 3C), which contrasts sharply with what was seen in B. malayi mf and perhaps better aligns with the expected bioactivity of IVM at this life stage (Moreno et al., 2010). While DEC significantly increased EV secretion in B. malayi mf it did not have any effect on B. pahangi mf (P = 0.6428, N = 3) (Figure 3C). Lastly, ABZ (P = 0.6605, N = 3) and LEV (P = 0.4125, N = 3) had no effect on EV secretion from B. pahangi mf (Figure 3C). To investigate whether the inhibitory effects of IVM on EV secretion were seen in more divergent nematode species we again repeated our screen on single adult female Ascaris suum, a soil‐transmitted gastrointestinal nematode. IVM (P = 0.0013, N = 14) and LEV (P = 0.0021, N = 16) both significantly inhibited EV secretion from individual adult female A. suum after 24 h by 99.4% and 99.1% respectively (Figure 3D). However, neither ABZ (P = 0.3769, N = 12) nor DEC (P = 0.9680, N = 16) had any effect on EV secretion.
Figure 3.

Ivermectin has broad inhibitory effects across filarial and gastrointestinal parasites. B. pahangi life stages in RPMI and adult female A. suum in Ascaris Ringers solution were cultured at 37°C with either drug (1.0 uM) or DMSO (vehicle control). Media was collected after 24 h and EVs were isolated and quantified. 1.0 μM IVM, ABZ, DEC, and LEV all significantly inhibited EV secretion from B. pahangi adult female parasites (A) while only IVM significantly reduced EV secretion in the microfilariae life stage (C) No treatment had any effect on EV secretion from B. pahangi adult male parasites (B) For A. suum adult females, both IVM and LEV significantly inhibited EV secretion (E) IVM rapidly inhibits EV secretion from adult female B pahangi and A. suum 24 h post‐treatment (E‐F). N = 3 (minimum). Mean ± SEM, *P < 0.05, **P < 0.01. ■ = DMSO, □ = Treatment
B. pahangi and A. suum adult female worms secrete EVs more robustly than B. malayi. In the case of A. suum, they secrete approximately 250 times more EVs than B. malayi in 24 h. This positioned us to use these two species to better understand how rapidly IVM inhibits EV secretion from susceptible parasitic nematodes. Adult female B. pahangi and A. suum parasites were treated with 1.0 μM IVM as before and spent media collected at 2, 4, 6, 12, and 24 h post IVM treatment. IVM significantly inhibited both B. pahangi and A. suum EV secretion by 63% (P = 0.0076, N = 3) and 99.4% (P = 0.0054, N = 3), respectively, at the 24 h post‐treatment timepoint (Figure 3E,F). In addition, EV secretion was inhibited as early as at 12 h post‐treatment by 72% (P = 0.1929, N = 3) for A. suum and by 57% (P = 0.0762, N = 3) for B. pahangi, though not statistically significant (Figure 3E,F). In vivo studies have shown IVM reduces microfilaremia in mice experimentally infected with B. malayi 24 h post‐treatment (Halliday et al., 2014). Thus, the rapidity of onset for this EV secretion phenotype is consistent with the therapeutic action of IVM. For context, other IVM phenotypes have been identified at 24 h post‐treatment timepoint, in vitro IVM reduces the release of B. malayi mf from adult female worms (Tompkins, Stitt, & Ardelli, 2010) and increases the binding of polymorphonuclear leukocytes to B. malayi mf (Berrafato et al., 2019) after 24 h. In canine heartworms, IVM inhibits the motility of Missouri strain D. immitis 24 h post‐treatment in vitro (Maclean et al., 2017).
As with B. malayi, we examined the effects of IVM treatment on the size distribution of secreted EVs in these other species. Compared to B. malayi, a broader distribution of B. pahangi adult female EVs was inhibited with more pronounced effects on larger EV cohorts. 24 h 1.0 μM IVM treatment reduced secretion of 51–100 nm EVs by 44% (P = < 0.0001, N = 12), 101–150 nm by 60% (P = 0.0022, N = 12), 151–200 nm by 73% (P = 0.0019, N = 12), and 201–250 nm by 86% (P = < 0.0001, N = 12) (Figure 2C). By contrast, in B. pahangi mfs, only EVs in the range 151–200 nm were inhibited (90%, P = 0.0056, N = 6) (Figure 2D). In Ascaris, IVM treatment inhibited secretion of 101–150 nm EVs by 80% (P = < 0.0001, N = 18) and 151–200 nm by 85% (P = < 0.0001, N = 11) (Figure 2E).
3.4. Ivermectin inhibition of EV secretion is not driven by loss of gross motor function
Glutamate‐gated chloride channels (GluCl) and nicotinic acetylcholine receptors (nAChRs), are known targets for IVM and LEV, respectively (Arena, Liu, Paress, & Cully, 1991, 1992; Harrow & Gration, 1985). GluCl have been identified in motor neurons and interneurons in various parasitic nematode species (Wolstenholme & Rogers, 2005) and nAChRs have been identified at the neuromuscular junction in filarial nematodes (Martin, 1997). Their locations lead to the discovery that IVM can induce paralysis of pharyngeal pumping in Haemonchus contortus (Geary et al., 1993) and both IVM and LEV can cause paralysis of B. malayi parasites (Mostafa et al., 2015; Tompkins et al., 2010). Due to these documented effects it is plausible that the EV phenotype is driven by gross motor function defects. To test this we examined the effect of our screening panel on gross motor function by analysing motility quantified using the Worminator software system (Marcellino et al., 2012). Our analysis was limited to B. malayi adults and L3 stage parasites due to parasite availability and difficulties in consistently recording the smaller mf life stage. A single B. malayi adult or 10 L3 stage parasites were cultured in a 24‐well or 96‐well plate respectively with or without 1.0 μM drug. Video recordings were taken prior to treatment, immediately after treatment (0 h), and at 2, 4, 6 and 24 h post‐treatment. IVM significantly reduced adult female motility by 57% beginning at 4 h post‐treatment (P < 0.0001, N = 5) (Figure 4A). However, when the kinetics of IVM treatment on adult female parasites was investigated it was observed that more therapeutically relevant concentrations (≤0.1 μM) did not affect motility (Figure 4D). This is corroborated by other data that shows that motility of B. malayi parasites was not inhibited by concentrations of IVM < 2 μM (Storey et al., 2014; Tompkins et al., 2010). This provides additional evidence that therapeutically relevant concentrations of IVM do not affect filarial nematode motility. The IC50 for IVM was determined to be 0.203 μM. The IC50 for EV secretion (7.7 nM) was below that of motility indicating that IVM is not reducing EV secretion by paralyzing the parasites. DEC (P < 0.001, N = 5) significantly inhibited adult female motility immediately upon treatment, but parasites began to recover at 1 h (P < 0.01, N = 5) and completely recovered by 4 h post‐treatment. LEV (P < 0.001, N = 5) significantly inhibited adult female motility by 88% immediately upon treatment, but parasites began to recover during the remaining 24 h. At 1 h post treatment LEV significantly inhibited adult female motility by 71%, at 4 h by 40%, at 6 h by 35% and at 24 h by 29% (1‐6 h post‐treatment: P < 0.0001, N = 5; 24 h: P < 0.01, N = 5). As was discussed earlier, no drug in our panel had any effect on EV secretion in adult male B. malayi, but it was discovered that LEV was a potent inhibitor of motility in adult male B. malayi (Figure 4B). Motility in adult males was significantly inhibited by 70% upon treatment with LEV. Adult male parasites treated with LEV did not recover with significant inhibition ranging from 70% to 76% over the 24 h tested (1–24 h post treatment: P < 0.0001, N = 5). The only drug that had any effect on B. malayi L3 parasites was also LEV with inhibition of motility by 90% immediately upon treatment (P < 0.001, N = 5) (Figure 4C). However, a very quick recovery of motility was seen in just one hour. In summary, 1.0 μM IVM had inhibitory effects on B. malayi adult female motility, but this concentration does not compare to therapeutically relevant concentrations or to the concentrations that inhibited EV secretion. LEV also had inhibitory effects on motility in all life stages tested, but adult female and L3 life stages recovered over 24 h while adult males did not recover. Due to the differences in IVM effects on motility compared to EV secretion we can conclude that inhibition of EV secretion is not a factor of parasite gross motor function being compromised.
Figure 4.
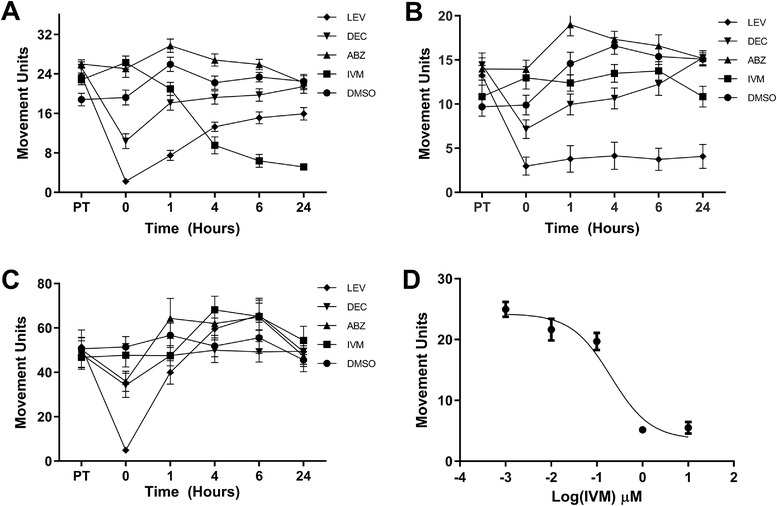
Ivermectin inhibition of EV secretion is not driven by loss of gross motor function. Single adult or 10 L3 stage Brugia malayi parasites were cultured in a 24‐well plate with either 1.0 μM drug or DMSO (vehicle control). Video recordings of worms were taken at timepoints ranging from pre‐treatment to 24 h using the Worminator system. (A) IVM significantly reduced adult female B. malayi motility as compared to control from 4 to 24 h post treatment (P < 0.0001). However, further investigation revealed that more therapeutically relevant concentrations of IVM did not affect adult female motility. The IC50 for IVM on adult female parasites was determined to be 203 nM. (D) DEC significantly reduced adult female motility immediately upon treatment and 1 h post treatment (P < 0.001). LEV significantly reduced adult female motility from 0 to 24 h (P < 0.0001‐P < 0.01) though the parasites began to recover after initial treatment. (B) DEC significantly reduced adult male motility from 1 to 4 h post treatment (P < 0.05, P < 0.01) and IVM began to reduce motility at 24 h post treatment (P < 0.05). LEV significantly reduced motility with no recovery from 0 to 24 h post treatment (P < 0.0001). (C) LEV significantly reduced L3 stage motility immediately upon treatment (P < 0.0001), but the parasites quickly recovered within 1 h. N = 11 (minimum), Mean ± SEM. ● = DMSO, ■ = IVM, ▲ = ABZ, ▼ = DEC, ⧫ = LEV
3.5. Ivermectin does not have parallel effects on EV and protein secretion
Data indicate that IVM and ABZ inhibit protein secretion from B. malayi mf (Moreno et al., 2010). We have already shown that IVM can inhibit EV secretion from B. malayi so we hypothesized that excretory‐secretory (ES) proteins and EVs would similarly be affected by the screening drug panel. Parasites were cultured with or without drug and ES protein secreted into the culture media was quantified 24 h after treatment using BCA. We chose to examine the 24 h time point as it is the most consistent with IVM therapeutic mechanism of action. Unlike EV secretion, neither 1.0 μM IVM (P = 0.8152, N = 5), ABZ (P = 0.9962, N = 5), DEC (P = 0.8863, N = 5) or LEV (P = 0.1571, N = 5) had any effect on protein secretion from B. malayi adult females (Figure 5A,D). Similarly, no effect of any drug on protein secretion was observed in adult males (IVM P = 0.9400, N = 5, ABZ P = 0.4906, N = 5, DEC P = 0.9902, N = 5, LEV P = 0.8043, N = 5) (Figure 5B). Inhibition of ES protein secretion from B. malayi mf was noted after treatment with 1.0 μM IVM but this was not statistically significant (23%, P = 0.0535, N = 5) (Figure 5C); however, ABZ had no effect on protein secretion (P = 0.5827, N = 5). DEC (0.5789, N = 5) and LEV (P = 0.1648, N = 5) also had no inhibitory effect on ES protein secretion from B. malayi mf. The data do not exactly correlate to previously published work describing clear inhibitory effects of IVM and ABZ protein release from B. malayi mf (Moreno et al., 2010). The assay we used to quantify ES protein secretion from B. malayi was slightly modified from that study but was fundamentally the same. Despite tight technical replication, there was challenging biological variability between worm batches and the low quantities of ES protein secreted necessitated a concentration step, potentially exacerbating variability. Despite this, we have high confidence in comparing this data with those of Moreno et al. (2010). Both studies observed a rapid inhibition of ES protein secretion from B. malayi mf following 24 h treatment with 1.0 μM IVM, and although our observed inhibition was moderately higher at 23% inhibition than Moreno et al. with 14%, it was not statistically significant. Further, Moreno et al described an inhibitory effect of ABZ (more potent than that of IVM) that was not observed here. In an effort to better describe the qualitative impact of IVM on secretion of ES products, we separated ES proteins on a 12% mini‐protean TGX gel and visualized using Coomassie Blue staining (Supplemental Figure 2). 7.5 μg total ES protein from IVM‐treated and control B. malayi adult females, and 1.8 μg total ES protein from IVM‐treated and control B. malayi mfs, cultured at both 37°C and 22°C, were so visualized. No conclusive qualitative changes could be determined using this approach but alternative approaches such as comparative proteomic analyses may be more revealing.
Figure 5.
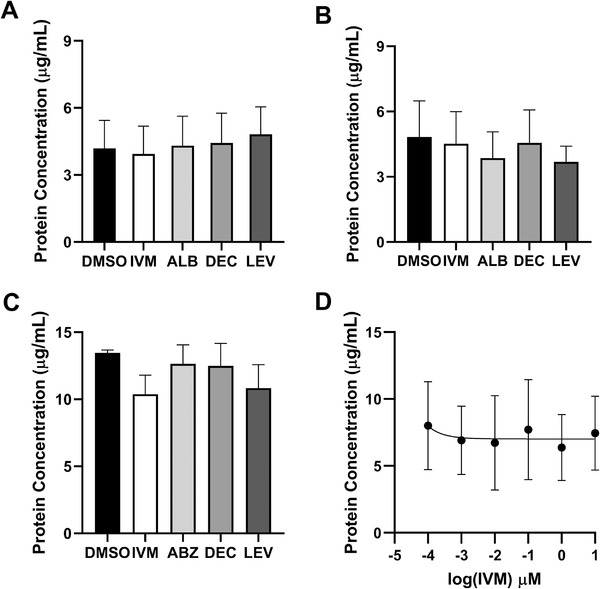
EV and protein secretion are differentially affected by ivermectin. Single adult or 100,000 microfilariae Brugia malayi parasites were cultured per well of a 24‐well plate with either drug (1.0 μM) or DMSO (0.01%) for 24 h. Spent media was collected and proteins were concentrated and washed. Protein concentration was determined by absorbance at 562 nm. No drug had any effect on protein secretion from B. malayi adult females (A) B. malayi adult males (B) had a decrease in protein secretion due to LEV though not statistically significant. Microfilariae protein secretion (C) was inhibited by ivermectin (P = 0.0535). (D) A dose response curve for IVM on adult female parasites showed no effect from any concentration of IVM (10 μM–0.1 nM). N = 5 (minimum), mean ± SEM
4. DISCUSSION
IVM is a broad spectrum, anti‐parasitic drug that is commonly used to treat and prevent multiple diseases caused by parasitic nematodes. Even with its extensive usage, the therapeutic mechanism of action of this drug is not completely understood (Wolstenholme et al., 2016). The current accepted hypothesis is that it functions, at least in part, by inhibiting secretion of ES proteins from parasites; this impairs the ability of the parasite to modulate host immune responses thereby facilitating host clearance (Moreno et al., 2010). Recent work has led to the characterization of the B. malayi secretome which, combined with RNA sequencing approaches, has defined a complex milieu of proteins and miRNAs secreted from these and other filarial parasites (Bennuru et al., 2009; Hewitson et al., 2008; Hoy et al., 2014; Kaushal, Hussain, Nash, & Ottesen, 1982; Tritten et al., 2016). Within this heterogenous mix of ES products are documented host immunomodulatory effectors, including leucyl aminopeptidase (ES‐62) and macrophage inhibiting factor 1 (MIF‐1), among others (Harnett, Deehan, Williams, & Harnett, 1998; Lal, Kumaraswami, Steel, & Nutman, 1990; Pastrana et al., 1998). Therefore, the observation that IVM can inhibit the secretion of immunomodulatory ES products is consistent with the rapid mf clearance observed after treatment in infected individuals. Although there is evidence tying the inhibition of ES product secretion to the mode of action of IVM, the critical ES products being inhibited are not immediately clear. In addition to freely secreted proteins, we have identified that prodigious numbers of EVs are also found in the ES products of filarial nematodes (Zamanian et al., 2015). In this study we show that IVM significantly and consistently inhibits EV secretion from B. malayi adult female, L3 and mf life stages, from B. pahangi adult females and mfs, and from female gastrointestinal A. suum nematodes. This inhibition occurs at therapeutically relevant concentrations (IC50 = 7.7 nM in adult female B. malayi) and time frame (within 24 h and perhaps even by 12 h). Given these properties, it is reasonable to hypothesize that the therapeutic mechanism of action of IVM against filarial nematodes may, in part, involve inhibition of EV secretion. Although premature, there is growing evidence to support this hypothesis. First, filarial nematode EVs are discrete structures that are enriched in immunomodulatory molecules. The cargo of B. malayi EVs includes proteins and miRNAs that have immunomodulatory functions and include modulatory proteins such as galectins and MIF‐1 as well as miRNAs with identity to immunomodulatory host miRNAs (Harischandra et al., 2018; Zamanian et al., 2015). EVs from other nematode species have similar composition; protein and small RNA profiling of EV cargo from a range of gastrointestinal and filarial nematodes reveals a multitude of putative effector molecules with emerging functionality at the host‐parasite interface (Buck et al., 2014; Eichenberger et al., 2018; Eichenberger et al., 2018; Gu et al., 2017; Hansen et al., 2015, 2019; Shears et al., 2018; Tritten et al., 2017). It is reasonable to posit that specifically inhibiting EV secretion would obstruct the immunomodulatory capabilities of these parasites. Second, the pharmacological disruption of EV secretion does not perfectly correlate with the secretion of other ES products, hinting that the regulation of EV secretion may be distinct to that of other ES products and therefore differentially 'druggable.' IVM (1.0 μM) inhibited EV secretion from mf stage B. malayi and B. pahangi after 24 h by 46% and 40%, respectively. In comparison, we found the same treatment inhibited protein secretion by B. malayi mf more modestly at 23% (not statistically significant, P = 0.0535). Further, whilst IVM (1.0 μM) inhibited EV secretion in adult female B. malayi by 59% after 24 h, the same treatment had no significant effect on protein secretion from those worms. Clearly, more work is needed to understand how parasite secretions are regulated but moving forward it may be advisable to disentangle the broad panoply of ES products and investigate them individually to help better understand host‐parasite interactions and particularly how drugs affect the secretion of parasite effector molecules.
Parasite motility has long been used as an assay to identify and measure the anthelmintic activity of compounds and as marker of parasite health and viability. Impaired motility alone, however, does not adequately account for the therapeutic effects of IVM in filarial disease. The in vitro IVM concentrations that are required to produce detrimental effects on gross filarial nematode motility are significantly higher than the bioavailable concentrations found in vivo after therapeutic administration (González Canga et al., 2008; Marcellino et al., 2012; Storey et al., 2014; Tompkins et al., 2010). Our data support that the IC50 of IVM in the B. malayi adult female EV secretion assay was 7.7 nM but was over 200 nM for the motility assay. IVM inhibits EV secretion but not motility in key stages at therapeutically relevant concentrations, supporting inhibition of EV secretion as a component of IVM mode of action. IVM also exerted inhibitory effects on B. malayi adult females but not adult males, and larval stages. This stage‐ and sex‐specific activity does correlate to the expression patterns of genes encoding subunits of glutamate‐gated chloride channels (GluCls), a proposed target for IVM (Arena et al., 1991, 1992). Li et al. (2014) found that avr‐14, a gene encoding a putative GluCl subunit in B. malayi, was expressed in both female and male reproductive tissues but consistently more strongly in female tissues (ovaries and surrounding body wall muscle) than male. This differential expression profile may help explain why EV secretion was inhibited in female worms but not male worms and may also point to reproductive structures as a source of these EVs in adult female worms. Proteomic analyses of EV cargo has proved valuable in identifying markers of tissue origin in other nematodes (Buck et al., 2014) but our previous nano‐scale proteome profiling of B. malayi female and male EVs did not identify any clear markers supporting a reproductive tissue origin for these vesicles (Harischandra et al., 2018). A more focused investigation of the fluid found in these structures may prove more illuminating, as would demonstration that putative IVM targets are similarly expressed in reproductive tissues of adult female B. pahangi and A. suum to account for the potent IVM activity we noted in those species. Li et al., (2014) also observed tissue‐specific avr‐14 expression in embryonic stages within gravid females. This corroborates the findings of Moreno et al. (2010) who noted strong localization of avr‐14 around the ES pore of B. malayi mfs, earmarking this structure as another, perhaps more predictable, site of EV secretion in this stage that lacks reproductive tissues or a through gut.
Whether these EVs have their biogenesis in reproductive tissues, the excretory system or some other secretory route (Drurey et al., 2020), the pathways by which IVM inhibits their secretion is obscure and will require a more thorough description of the microscopic anatomy of key tissues and a better understanding of IVM targets expressed therein. For example, despite the recognition that parasitic nematode ES systems secrete a complex suite of molecules believed to be essential for successful parasitism (Allen & Maizels, 2011; Hewitson, Grainger, & Maizels, 2009; Hoerauf, Satoguina, Saeftel, & Specht, 2005; van Riet, Hartgers, & Yazdanbakhsh, 2007), the ultrastructure and transcriptional topography of the ES pore region has largely been uninvestigated. The intersection between EVs as an important mechanism for host manipulation during infection, the inhibition of their secretion by IVM at therapeutically relevant concentrations and time frames, and the localization of putative IVM targets in critical stage‐specific tissues, provides strong rationale for addressing this knowledge gap.
A significant outcome from the work presented here is the demonstration that EV secretion from adult female and mf stage filarial nematodes (the stages that one could argue are most relevant to LF control programs) can be quantified and the effect of extraneously applied compounds on this secretion measured. Using this assay, we detected an IVM‐sensitive EV secretion phenotype that perhaps correlates better with therapeutically relevant IVM concentrations than does assaying parasite motility, and in our experience is a more convenient and reproducible assay than that used to measure protein secretion from these worms. It may also be a better predictor of IVM mode of action. If the therapeutic mechanism of action of IVM is to inhibit immunomodulatory protein secretion from mf parasites then albendazole, which has been reported to inhibit protein secretion from B. malayi mf faster and more comprehensively than IVM (Moreno et al., 2010) (although we did not observe this), should also be an effective microfilaricide. Albendazole, however, is ineffective against mf stage filarial nematodes (Critchley et al., 2005). This suggests inhibition of EV secretion may be a preferred characteristic of anti‐filarial drugs and therefore assaying this phenotype would be of significance to future drug discovery efforts aimed at developing new anti‐filarial compounds, certainly those that function like IVM. In its favour, EV quantification would provide a consistent screening assay that would be comparable across different species of parasites, however, EV quantification is not high‐throughput and does require additional EV isolation steps and specialized equipment for EV visualization. Recent technological advances may provide platforms that could be leveraged to streamline EV quantification assays and overcome these drawbacks. For example, we have contributed to an on‐chip microfluidic device that utilizes a label‐free photonic crystal biosensor to detect and discriminate host EVs from those secreted by parasitic nematodes based on differential expression of EV surface markers (Wang, Yuan, Kimber, Lu, & Dong, 2018). This type of platform combines minimal sample processing with high throughput potential and does not require EV labelling, overcoming the disadvantages of traditional EV quantification and could be leveraged in drug discovery efforts centred on EV secretion as an assay endpoint. Another potential application for such a sensitive platform for EV quantification is the detection of parasite EVs in host biofluids as an early diagnostic marker for parasite infection. The use of EVs as a nematode diagnostic has been seeded by the focus on EVs as diagnostic markers for cancer detection. Current advanced technologies involve using surface enhanced Raman scattering (SERS) or localized surface plasmon resonance to detect tumour‐derived EVs in body fluids (Mehmet, R, Aysun, & Sebastian, 2017; Thakur et al., 2017; Zong et al., 2016). In addition, the miRNA cargo of EVs has been of interest as biomarkers for various cancers (Kosaka et al., 2019). Similarly, a microfluidic on‐chip device has potential to identify parasite EVs from host biofluids. Current efforts towards this goal are aimed at identifying secreted parasite markers that that could be incorporated in such a design, or used in more simple assay formats such as PCR (Quintana et al., 2017; Tritten et al., 2014). Finally, the assay we describe here may be an example of a relatively simple in vitro assay to test or validate the emergence of anthelmintic resistance. The Fecal Egg Count Reduction Test is the gold standard for detecting resistance to anthelmintics like IVM. Alternative in vitro assays complement FECRT and include hatching and development assays, molecular tests and, of course, motility assays (Kotze & Prichard, 2016). The EV secretion assay could be added to this list if it could reliably, and with sensitivity, detect resistance to drugs such as IVM in a standardized fashion. There is some evidence for this potential; we previously detected differences in IVM susceptibility based on EV secretion for two strains of canine heartworm, D. immitis (Harischandra et al., 2018).
Collectively, our data show that the secretion of EVs from different parasitic nematode species can be assayed and the effects of anthelmintic drugs or lead compounds on this physiological process can be measured. IVM consistently inhibited EV secretion against all species and most life stages investigated, with the exception of male worms; other anti‐filarial drugs did not. These findings provide new insight into the stage‐, sex‐ and species‐specific pathways and pharmacological regulation of EV secretion in parasitic nematodes. The data are significant because, given the emerging immunomodulatory role of EVs at the host‐parasite interface, it provides new evidence that the therapeutic mechanism of IVM, in part, involves inhibition of parasite EV secretion.
CONFLICTS OF INTEREST
The authors report no conflict of interest.
Supporting information
Supporting information.
ACKNOWLEDGEMENT
This work was supported by the National Institutes of Health under Grant AI117204 (MK) and AI151171 (MZ).
Loghry HJ, Yuan W, Zamanian M, Wheeler NJ, Day TA, Kimber MJ. Ivermectin inhibits extracellular vesicle secretion from parasitic nematodes. J Extracell Vesicles. 2020;10:e12036 10.1002/jev2.12036
Present Address: Michael J. Kimber, Department of Pharmacology, University of Texas Southwestern Medical Center, Dallas, Texas, USA
REFERENCES
- Allen, J. E. , & Maizels, R. M. (2011). Diversity and dialogue in immunity to helminths. Nature Reviews Immunology, 11(6), 375–388. [DOI] [PubMed] [Google Scholar]
- Arena, J. P. , Liu, K. K. , Paress, P. S. , & Cully, D. F. (1991). Avermectin‐sensitive chloride currents induced by Caenorhabditis elegans RNA in Xenopus oocytes. Molecular Pharmacology, 40(3), 368–374. [PubMed] [Google Scholar]
- Arena, J. P. , Liu, K. K. , Paress, P. S. , Schaeffer, J. M. , & Cully, D. F. (1992). Expression of a glutamate‐activated chloride current in Xenopus oocytes injected with Caenorhabditis elegans RNA: evidence for modulation by avermectin. Molecular Brain Research, 15(3), 339–348. [DOI] [PubMed] [Google Scholar]
- Babu, S. , & Nutman, T. B. (2014). Immunology of lymphatic filariasis. Parasite Immunology, 36(8), 338–346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bennuru, S. , Semnani, R. , Meng, Z. , Ribeiro, J. M. C. , Veenstra, T. D. , & Nutman, T. B. (2009). Brugia malayi Excreted/secreted proteins at the host/parasite interface: stage‐ and gender‐specific proteomic profiling. PLOS Neglected Tropical Diseases, 3(4), e410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berrafato, T. , Coates, R. , Reaves, B. J. , Kulke, D. , & Wolstenholme, A. J. (2019). Macrocyclic lactone anthelmintic‐induced leukocyte binding to Dirofilaria immitis microfilariae: Influence of the drug resistance status of the parasite. International Journal for Parasitology: Drugs and Drug Resistance, 10, 45–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bobrie, A. , Colombo, M. , Raposo, G. , & Théry, C. (2011). Exosome secretion: Molecular mechanisms and roles in immune responses. Traffic, 12(12), 1659–1668. [DOI] [PubMed] [Google Scholar]
- Buck, A. H. , Coakley, G. , Simbari, F. , McSorley, H. J. , Quintana, J. F. , Le Bihan, T. , … Maizels, R. M. (2014). Exosomes secreted by nematode parasites transfer small RNAs to mammalian cells and modulate innate immunity. Nature Communications, 5(1), 5488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carithers, D. S. (2017). Examining the role of macrolides and host immunity in combatting filarial parasites. Parasites & Vectors, 10(1), 182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carlisle, C. H. , Atwell, R. B. , & Robinson, S. (1984). The effectiveness of levamisole hydrochloride against the microfilaria of Dirofilaria immitis. Australian Veterinary Journal, 61(9), 282–284. [DOI] [PubMed] [Google Scholar]
- Catalano, M. , & O'Driscoll, L. (2020). Inhibiting extracellular vesicles formation and release: A review of EV inhibitors. Journal of Extracellular Vesicles, 9(1), 1703244 10.1080/20013078.2019.1703244 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coakley, G. , McCaskill, J. L. , Borger, J. G. , Simbari, F. , Robertson, E. , Millar, M. , … Buck, A. H. (2017). Extracellular vesicles from a helminth parasite suppress macrophage activation and constitute an effective vaccine for protective immunity. Cell Reports, 19(8), 1545–1557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Critchley, J. , Addiss, D. , Ejere, H. , Gamble, C. , Garner, P. , & Gelband, H. (2005). Albendazole for the control and elimination of lymphatic filariasis: Systematic review. Tropical Medicine & International Health, 10(9), 818–825. [DOI] [PubMed] [Google Scholar]
- Drurey, C. , Coakley, G. , & Maizels, R. M. (2020). Extracellular vesicles: New targets for vaccines against helminth parasites. International Journal for Parasitology, 50(9), 623–633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eichenberger, R. M. , Ryan, S. , Jones, L. , Buitrago, G. , Polster, R. , Montes de Oca, M. , … Loukas, A. (2018). Hookworm secreted extracellular vesicles interact with host cells and prevent inducible colitis in mice. Frontiers in Immunology, 9, 850 10.3389/fimmu.2018.00850 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eichenberger, R. M. , Sotillo, J. , & Loukas, A. (2018). Immunobiology of parasitic worm extracellular vesicles. Immunology & Cell Biology, 96(7), 704–713. 10.1111/imcb.12171 [DOI] [PubMed] [Google Scholar]
- Eichenberger, R. M. , Talukder, M. H. , Field, M. A. , Wangchuk, P. , Giacomin, P. , Loukas, A. , & Sotillo, J. (2018). Characterization of Trichuris muris secreted proteins and extracellular vesicles provides new insights into host–parasite communication. Journal of Extracellular Vesicles, 7(1), 1428004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geary, T. G. , Sims, S. M. , Thomas, E. M. , Vanover, L. , Davis, J. P. , Winterrowd, C. A. , … Thompson, D. P. (1993). Haemonchus contortus: Ivermectin‐induced paralysis of the pharynx. Experimental Parasitology, 77(1), 88–96. [DOI] [PubMed] [Google Scholar]
- Geary, T. G. , Woo, K. , McCarthy, J. S. , Mackenzie, C. D. , Horton, J. , Prichard, R. K. , … Bundy, D. A. (2010). Unresolved issues in anthelmintic pharmacology for helminthiases of humans. International Journal for Parasitology, 40(1), 1–13. [DOI] [PubMed] [Google Scholar]
- González Canga, A. , Sahagún Prieto, A. M. , Diez Liébana, M. J. , Fernández Martínez, N. , Sierra Vega, M. , & García Vieitez, J. J. (2008). The pharmacokinetics and interactions of ivermectin in humans—A mini‐review. The AAPS Journal, 10(1), 42–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gu, H. Y. , Marks, N. D. , Winter, A. D. , Weir, W. , Tzelos, T. , McNeilly, T. N. , … Devaney, E. (2017). Conservation of a microRNA cluster in parasitic nematodes and profiling of miRNAs in excretory‐secretory products and microvesicles of Haemonchus contortus. PLOS Neglected Tropical Diseases, 11(11), e0006056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halliday, A. , Guimaraes, A. F. , Tyrer, H. E. , Metuge, H. M. , Patrick, C. N. W. , Arnaud, K.‐O. J. , … Turner, J. D. (2014). A murine macrofilaricide pre‐clinical screening model for onchocerciasis and lymphatic filariasis. Parasites & Vectors, 7(1), 472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hansen, E. P. , Fromm, B. , Andersen, S. D. , Marcilla, A. , Andersen, K. L. , Borup, A. , … Nejsum, P. (2019). Exploration of extracellular vesicles from Ascaris suum provides evidence of parasite–host cross talk. Journal of Extracellular Vesicles, 8(1), 1578116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hansen, E. P. , Kringel, H. , Williams, A. R. , & Nejsum, P. (2015). Secretion of rna‐containing extracellular vesicles by the porcine whipworm, Trichuris suis. The Journal of Parasitology, 101(3), 336–340. JSTOR. [DOI] [PubMed] [Google Scholar]
- Harischandra, H. , Yuan, W. , Loghry, H. J. , Zamanian, M. , & Kimber, M. J. (2018). Profiling extracellular vesicle release by the filarial nematode Brugia malayi reveals sex‐specific differences in cargo and a sensitivity to ivermectin. PLOS Neglected Tropical Diseases, 12(4), e0006438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harnett, M. M. , Deehan, M. R. , Williams, D. M. , & Harnett, W. (1998). Induction of signalling anergy via the T‐cell receptor in cultured Jurkat T cells by pre‐exposure to a filarial nematode secreted product. Parasite Immunology, 20(11), 551–563. [DOI] [PubMed] [Google Scholar]
- Harrow, I. D. , & Gration, K. A. F. (1985). Mode of action of the anthelmintics morantel, pyrantel and levamisole on muscle cell membrane of the nematode Ascaris suum. Pesticide Science, 16(6), 662–672. [Google Scholar]
- Hewitson, J. P. , Grainger, J. R. , & Maizels, R. M. (2009). Helminth immunoregulation: The role of parasite secreted proteins in modulating host immunity. Molecular and Biochemical Parasitology, 167(1), 1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hewitson, J. P. , Harcus, Y. M. , Curwen, R. S. , Dowle, A. A. , Atmadja, A. K. , Ashton, P. D. , … Maizels, R. M. (2008). The secretome of the filarial parasite, Brugia malayi: Proteomic profile of adult excretory–secretory products. Molecular and Biochemical Parasitology, 160(1), 8–21. [DOI] [PubMed] [Google Scholar]
- Hoerauf, A. , Satoguina, J. , Saeftel, M. , & Specht, S. (2005). Immunomodulation by filarial nematodes. Parasite Immunology, 27(10–11), 417–429. [DOI] [PubMed] [Google Scholar]
- Hoy, A. M. , Lundie, R. J. , Ivens, A. , Quintana, J. F. , Nausch, N. , Forster, T. , … Buck, A. H. (2014). Parasite‐derived microRnas in host serum as novel biomarkers of helminth infection. PLoS Neglected Tropical Diseases, 8(2), e2701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnstone, R. M. , Adam, M. , Hammond, J. R. , Orr, L. , & Turbide, C. (1987). Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). Journal of Biological Chemistry, 262(19), 9412–9420. [PubMed] [Google Scholar]
- Kaushal, N. A. , Hussain, R. , Nash, T. E. , & Ottesen, E. A. (1982). Identification and characterization of excretory‐secretory products of Brugia malayi, adult filarial parasites. The Journal of Immunology, 129(1), 338–343. [PubMed] [Google Scholar]
- Kosaka, N. , Kogure, A. , Yamamoto, T. , Urabe, F. , Usuba, W. , Prieto‐Vila, M. , & Ochiya, T. (2019). Exploiting the message from cancer: the diagnostic value of extracellular vesicles for clinical applications. Experimental & Molecular Medicine, 51(3), 1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kotze, A. C. , & Prichard, R. K. (2016). Anthelmintic resistance in Haemonchus contortus: History, mechanisms and diagnosis In Gasser R. B. & Samson‐Himmelstjerna G. V. (Eds.), Advances in parasitology. Cambridge, MA: Academic Press, 93, 397–428. [DOI] [PubMed] [Google Scholar]
- Lal, R. B. , Kumaraswami, V. , Steel, C. , & Nutman, T. B. (1990). Phosphocholine‐containing antigens of brugia malayi nonspecifically suppress lymphocyte function. The American Journal of Tropical Medicine and Hygiene, 42(1), 56–64. [DOI] [PubMed] [Google Scholar]
- Lee, Y. , EL Andaloussi, S. , & Wood, M. J. A. (2012). Exosomes and microvesicles: Extracellular vesicles for genetic information transfer and gene therapy. Human Molecular Genetics, 21(R1), R125–R134. [DOI] [PubMed] [Google Scholar]
- Li, B. W. , Rush, A. C. , & Weil, G. J. (2014). High level expression of a glutamate‐gated chloride channel gene in reproductive tissues of Brugia malayi may explain the sterilizing effect of ivermectin on filarial worms. International Journal for Parasitology: Drugs and Drug Resistance, 4(2), 71–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maclean, M. J. , Savadelis, M. D. , Coates, R. , Dzimianski, M. T. , Jones, C. , Benbow, C. , … Wolstenholme, A. J. (2017). Does evaluation of in vitro microfilarial motility reflect the resistance status of Dirofilaria immitis isolates to macrocyclic lactones? Parasites & Vectors, 10(2), 480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marcellino, C. , Gut, J. , Lim, K. C. , Singh, R. , McKerrow, J. , & Sakanari, J. (2012). WormAssay: A novel computer application for whole‐plate motion‐based screening of macroscopic parasites. PLOS Neglected Tropical Diseases, 6(1), e1494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin, R. J. (1997). Modes of action of anthelmintic drugs. The Veterinary Journal, 154(1), 11–34. [DOI] [PubMed] [Google Scholar]
- Mehmet, K. , R, M. E. , Aysun, K. , & Sebastian, W.‐H. (2017). Fundamentals and applications of SERS‐based bioanalytical sensing. Nanophotonics, 6(5), 831–852. [Google Scholar]
- Mills, J. N. , & Amis, T. C. (1975). Levamisole as a microfilaricidal agent in the control of canine dirofilariasis. Australian Veterinary Journal, 51(6), 310–314. [DOI] [PubMed] [Google Scholar]
- Moreno, Y. , Nabhan, J. F. , Solomon, J. , Mackenzie, C. D. , Geary, T. G. , & Campbell, W. C. (2010). Ivermectin disrupts the function of the excretory‐secretory apparatus in microfilariae of Brugia malayi. Proceedings of the National Academy of Sciences of the United States of America, 107(46), 20120–20125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mostafa, E. , Storey, B. , Farghaly, A. M. , Afify, H. A. E.‐H. , Taha, A. A. E.‐R. , & Wolstenholme, A. J. (2015). Transient effects of levamisole on Brugia malayi microfilariae. Invertebrate Neuroscience, 15(3), 5. [DOI] [PubMed] [Google Scholar]
- Ottesen, E. A. (2006). Lymphatic filariasis: Treatment, control and elimination In Molyneux D. H. (Ed.), Advances in parasitology. Cambridge, MA: Academic Press, 61, 395–441. [DOI] [PubMed] [Google Scholar]
- Pastrana, D. V. , Raghavan, N. , FitzGerald, P. , Eisinger, S. W. , Metz, C. , Bucala, R. , … Scott, A. L. (1998). Filarial nematode parasites secrete a homologue of the human cytokine macrophage migration inhibitory factor. Infection and Immunity, 66(12), 5955–5963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quintana, J. F. , Babayan, S. A. , & Buck, A. H. (2017). Small RNAs and extracellular vesicles in filarial nematodes: From nematode development to diagnostics. Parasite Immunology, 39(2), e12395. [DOI] [PubMed] [Google Scholar]
- Raposo, G. , Nijman, H. W. , Stoorvogel, W. , Liejendekker, R. , Harding, C. V. , Melief, C. J. , & Geuze, H. J. (1996). B lymphocytes secrete antigen‐presenting vesicles. Journal of Experimental Medicine, 183(3), 1161–1172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riaz, F. , & Cheng, G. (2017). Exosome‐like vesicles of helminths: Implication of pathogenesis and vaccine development. Annals of Translational Medicine, 5(7), 175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robertson, A. P. , Buxton, S. K. , & Martin, R. J. (2013). Whole‐cell patch‐clamp recording of nicotinic acetylcholine receptors in adult Brugia malayi muscle. Parasitology International, 62(6), 616–618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schroeder, J.‐H. , McCarthy, D. , Szestak, T. , Cook, D. A. , Taylor, M. J. , Craig, A. G. , … Lawrence, R. A. (2017). Brugia malayi microfilariae adhere to human vascular endothelial cells in a C3‐dependent manner. PLoS Negl Trop Dis, 11(5), e0005592 10.1371/journal.pntd.0005592 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Semnani, R. T. , Keiser, P. B. , Coulibaly, Y. I. , Keita, F. , Diallo, A. A. , Traore, D. , … Nutman, T. B. (2006). Filaria‐induced monocyte dysfunction and its reversal following treatment. Infection and Immunity, 74(8), 4409–4417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shears, R. K. , Bancroft, A. J. , Hughes, G. W. , Grencis, R. K. , & Thornton, D. J. (2018). Extracellular vesicles induce protective immunity against Trichuris muris. Parasite Immunology, 40(7), e12536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Storey, B. , Marcellino, C. , Miller, M. , Maclean, M. , Mostafa, E. , Howell, S. , … Kaplan, R. (2014). Utilization of computer processed high definition video imaging for measuring motility of microscopic nematode stages on a quantitative scale: “The Worminator.” International Journal for Parasitology: Drugs and Drug Resistance, 4(3), 233–243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thakur, A. , Qiu, G. , Ng, S.‐P. , Guan, J. , Yue, J. , Lee, Y. , & Wu, C.‐M. L. (2017). Direct detection of two different tumor‐derived extracellular vesicles by SAM‐AuNIs LSPR biosensor. Biosensors and Bioelectronics, 94, 400–407. [DOI] [PubMed] [Google Scholar]
- Thery, C. , Zitvogel, L. , & Amigorena, S. (2002). Exosomes: composition, biogenesis and function. Nature Reviews Immunology, 2(8), 569–579. [DOI] [PubMed] [Google Scholar]
- Tompkins, J. B. , Stitt, L. E. , & Ardelli, B. F. (2010). Brugia malayi: In vitro effects of ivermectin and moxidectin on adults and microfilariae. Experimental Parasitology, 124(4), 394–402. [DOI] [PubMed] [Google Scholar]
- Tritten, L. , Burkman, E. , Moorhead, A. , Satti, M. , Geary, J. , Mackenzie, C. , & Geary, T. (2014). Detection of circulating parasite‐derived microRNAs in filarial infections. PLoS Neglected Tropical Diseases, 8(7), e2971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tritten, L. , Clarke, D. , Timmins, S. , McTier, T. , & Geary, T. G. (2016). Dirofilaria immitis exhibits sex‐ and stage‐specific differences in excretory/secretory miRNA and protein profiles. Veterinary Parasitology, 232, 1–7. [DOI] [PubMed] [Google Scholar]
- Tritten, L. , Tam, M. , Vargas, M. , Jardim, A. , Stevenson, M. M. , Keiser, J. , & Geary, T. G. (2017). Excretory/secretory products from the gastrointestinal nematode Trichuris muris. Experimental Parasitology, 178, 30–36. [DOI] [PubMed] [Google Scholar]
- Tzelos, T. , Matthews, J. B. , Buck, A. H. , Simbari, F. , Frew, D. , Inglis, N. F. , … McNeilly, T. N. (2016). A preliminary proteomic characterisation of extracellular vesicles released by the ovine parasitic nematode, Teladorsagia circumcincta. Veterinary Parasitology, 221, 84–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valadi, H. , Ekstrom, K. , Bossios, A. , Sjostrand, M. , Lee, J. J. , & Lotvall, J. O. (2007). Exosome‐mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nature Cell Biology, 9(6), 654–659. [DOI] [PubMed] [Google Scholar]
- van Riet, E. , Hartgers, F. C. , & Yazdanbakhsh, M. (2007). Chronic helminth infections induce immunomodulation: Consequences and mechanisms. Immunobiology, 212(6), 475–490. [DOI] [PubMed] [Google Scholar]
- Vlassov, A. V. , Magdaleno, S. , Setterquist, R. , & Conrad, R. (2012). Exosomes: Current knowledge of their composition, biological functions, and diagnostic and therapeutic potentials. Biochimica et Biophysica Acta (BBA) ‐ General Subjects, 1820(7), 940–948. [DOI] [PubMed] [Google Scholar]
- Wang, Y. , Yuan, W. , Kimber, M. , Lu, M. , & Dong, L. (2018). Rapid differentiation of host and parasitic exosome vesicles using microfluidic photonic crystal biosensor. ACS Sensors, 3(9), 1616–1621. [DOI] [PubMed] [Google Scholar]
- Wolstenholme, A. J. , Maclean, M. J. , Coates, R. , McCoy, C. J. , & Reaves, B. J. (2016). How do the macrocyclic lactones kill filarial nematode larvae? Invertebrate Neuroscience, 16(3), 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolstenholme, A. J. , & Rogers, A. T. (2005). Glutamate‐gated chloride channels and the mode of action of the avermectin/milbemycin anthelmintics. Parasitology, 131(S1), S85–S95. [DOI] [PubMed] [Google Scholar]
- World Health Organization . (2019). World Health Organization model list of essential medicines: 21st list 2019 (WHO/MVP/EMP/IAU/2019.06). Article WHO/MVP/EMP/IAU/2019.06. Retrieved from https://apps.who.int/iris/handle/10665/325771
- World Health Organization . (2019). Global programme to eliminate lymphatic filariasis: progress report, 2018. Weekly Epidemiological Record, 94(41), 457–472. [Google Scholar]
- Zamanian, M. , Fraser, L. M. , Agbedanu, P. N. , Harischandra, H. , Moorhead, A. R. , Day, T. A. , … Kimber, M. J. (2015). Release of small RNA‐containing exosome‐like vesicles from the human filarial parasite Brugia malayi. PLoS Neglected Tropical Diseases, 9(9), e0004069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zong, S. , Wang, L. , Chen, C. , Lu, J. , Zhu, D. , Zhang, Y. , … Cui, Y. (2016). Facile detection of tumor‐derived exosomes using magnetic nanobeads and SERS nanoprobes. Analytical Methods, 8(25), 5001–5008. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supporting information.