

Abstract
Coronavirus disease 2019 (COVID-19) is a virus-induced respiratory disease that may progress to acute respiratory distress syndrome (ARDS) and is triggered by immunopathological mechanisms that cause excessive inflammation and leukocyte dysfunction. Neutrophils play a critical function in the clearance of bacteria with specific mechanisms to combat viruses. The aim of this review is to highlight the current advances in the pathways of neutrophilic inflammation against viral infection over the past ten years, focusing on the production of neutrophil extracellular traps (NETs) and its impact on severe lung diseases, such as COVID-19. We focused on studies regarding hyperinflammation, cytokine storms, neutrophil function, and viral infections. We discuss how the neutrophil's role could influence COVID-19 symptoms in the interaction between hyperinflammation (overproduction of NETs and cytokines) and the clearance function of neutrophils to eliminate the viral infection. We also propose a more in-depth investigation into the neutrophil response mechanism targeting NETosis in the different phases of COVID-19.
1. Introduction
The World Health Organization (WHO) established the coronavirus disease 2019 (COVID-19) as a pandemic on March 11, 2020. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is a member of the coronavirus family, a class of enveloped viruses with a positive-sense single-stranded RNA genome. This virus can cross species barriers and induce illnesses ranging from the usual cold to severe interstitial pneumonia, respiratory failure, and septic shock [1]. While there is a global effort in the development of vaccines and improvement of diagnostic methods [2, 3] and therapies that relieve the symptoms and prognosis of COVID-19 patients under severe infection [4], there remain gaps in our understanding of the pathophysiology of COVID-19 related to innate immunity.
In a scenario where patients with severe COVID-19 could develop dysfunction of the immune response that aggravates the hyperinflammation [5, 6], it is hypothesized that neutrophils can amplify pathological damage or control other cell subsets depending on the infection features. Therefore, to use the potential of NETs with minimal damage to the hosts, there must be a right balance of NET formation and reduction of the amount of NETs that accumulate in tissues [7].
Notwithstanding the rapid progress in the field, there are many critical unknown features of neutrophils in fighting viral infections. We highlighted the current progress in the pathways of neutrophilic inflammation in viral infection, with a focus on the release of NETs and its influence on lung disease. The knowledge summarized in this study should benefit researchers in integrating neutrophil biology to design new and more efficient virus-targeted interventions concerning COVID-19.
2. Hyperinflammation
Although a well-regulated innate immune process is the first protection action against viral infections [8], in severe COVID-19 condition occurs hyperinflammation (“cytokine storm”) that might lead to the acute respiratory distress syndrome (ARDS) [6, 9].
Cytokines play a relevant function in immunopathology during virus infections. The host-viral interactions are established via host identification of pathogen-associated molecular patterns (PAMPs) of the virus [10]. This identification occurs through host pattern recognition receptors (PRRs) manifested on innate immune cells (e.g., neutrophils, dendritic cells, epithelial cells, and macrophages) [11], and the recognition of PAMPs and viral danger-associated molecular patterns (DAMPs) by conserved PRRs marks the first line of defense against pathogens, involving toll-like receptors (TLRs) [11].
TLR stimulation activates the nuclear factor-κB (NF-κB) signaling cascade, causing the production of inflammatory markers from monocytes (interleukin- (IL-) 1, tumor necrosis factor-alpha (TNF-α), and IL-6) to control virus infections [8] by direct antiviral pathways and the recruitment of other leukocytes [10]. Moreover, the exacerbated oxidative stress induced by elevated concentrations of cytokines, along with reduced concentrations of interferon α and interferon β (IFN-α, IFN-β), influences the severity of COVID-19 [12].
Several mediators control the release of chemoattractants and neutrophil activity [10], and studies have demonstrated that higher values of proinflammatory markers are related to extensive lung damage and pulmonary inflammation in MERS-CoV [13] and ARDS infection [14]. COVID-19 in the severe state exhibits a cytokine storm with elevated plasma levels of chemokine ligand 2 (CCL2), IFNγ, IFNγ-inducible protein 10, G-CSF, chemokine C-C motif ligand 3 (CCL3), IL-1β, IL-2, IL-6, IL-7, IL-8, IL-10, IL-17, and TNF-α [12, 15]. Nucleotide-binding oligomerization domain- (NOD-) like receptor and increased plasma levels of chemokines and cytokines in COVID-19 patients relate to the severity of the disease rather than did those nonsevere patients [5]. In this sense, Huang et al. [15] found that patients in the intensive care unit (ICU) with laboratory-confirmed COVID-19 infection had higher plasma levels of IL-2, IL-7, IL-10, interferon-inducible protein 10, granulocyte colony-stimulating factor, CCL2, CCL3, and TNF-α when compared with non-ICU patients [15].
3. Neutrophils: The First Cell Recruitment
Neutrophils are innate immune cells with a brief lifespan after leaving the bone marrow and exist in a quiescent, primed, or active state. These leukocytes are the leading players in innate immunity since they are among the first innate leukocytes recruited during infections [16]. The primary function of neutrophil is clearance of pathogens and debris through phagocytosis [17]. They also have a distinct array of other immune roles, such as the liberation of NETs for viral infection inactivation [18] and cytokine production to restrict virus replication [16].
The release of neutrophil-chemoattractive elements and the resulting recruitment of neutrophils are a global host response to viral infection [19]. In this scenario, the neutrophil cell membrane also expresses a complex array of receptors and adhesion molecules for various ligands, including immunoglobulins, membrane molecules on other cells, and cytokines [20].
In addition to the trafficking to infection places to phagocytize viruses, the neutrophils can initiate, enlarge, and/or repress adaptive immune effector processes by promoting bidirectional cross-talk with T cells [21, 22]. Following the acute inflammation arising from immunological processes, such as viral infections, neutrophils with decreased expression of CD62L weaken T cell migration via the CXCL11 chemokine gradient by releasing H2O2 into an immunological synapse [23]. Thus, neutrophils that uncovered viral antigens can home to draining lymph nodes, acting as antigen-presenting cells (APC) [24]. Hufford et al. [25] evidenced that neutrophils expressing viral antigen as an outcome of direct infection by influenza A virus (IAV) display the most potent APC activity and that viral antigen-presenting neutrophils infiltrating the IAV-infected lungs act as APC for effector CD8(+) T lymphocytes in the infected lungs [25]. Neutrophils recruit the T cell molecular mechanism during the influenza virus infection and associate to CXCL12 reservoirs left behind. CD8+ T cells follow the chemoattractant trail left behind by neutrophil uropods to the influenza virus infection site [26].
Decreased cell number or impaired leukocyte function can play a part in advance of mild to severe clinical disease conditions [16]. Regarding the new coronavirus, the neutrophil-to-lymphocyte ratio (NLR), a well-known marker of infection and systemic inflammation, has evidenced an enhanced inflammatory response in COVID-19 patients [5]. Since the ARDS is the primary cause of mortality in patients with COVID-19, the elevated NLR values suggest a poor prognosis in COVID-19 disease [27], especially severe COVID-19 compared to mild patients. Sun et al. [28] studied 116 patients with COVID-19 and showed a higher NLR [28]. The authors compared severe COVID-19 patients admitted to the ICU with others or severe patients not admitted to the ICU. They reported that COVID-19 patients have the lowest count of lymphocytes and the highest neutrophil count and NLR [28]. Wang et al. [29] also showed that several COVID-19 patients have a rising neutrophil count and a falling lymphocyte count during the severe phase [29]. Similarly, Barnes et al. [30] found extensive neutrophil infiltration in pulmonary capillaries from a COVID-19 patient [30]. Nevertheless, even though severe cases of COVID-19 appear to be related to increased NLR levels [5], whether NLR could be an independent predictor of mortality in COVID-19 patients still requires investigation.
4. Neutrophil Extracellular Traps (NETs) and Viral Infection
Neutrophils can develop a sophisticated network of DNA called NETs through NETosis, a liberation of web-like structures of nucleic acids wrapped with histones that detain viral particles [31]. Upon discovery, the researchers believed that the production of NETs defended only against fungi and bacteria [32]. However, the NETosis process plays an important function in the response to viral diseases [33], thereby protecting the host during the virus response by trapping and eliminating distinct pathogens [31].
The formation of NETs is a controlled process, even though the related signals remain unknown. NETosis is conditional on the production of reactive oxygen species (ROS) by nicotinamide adenine dinucleotide phosphate oxidase (NADPH oxidase) [34]. There is evidence of NETosis produced in a ROS-independent mechanism [35]. In general, the NETosis process includes the release of nuclear chromatin lined with effector proteins and peptidyl arginine deiminase type IV (PAD4) activation [36]. After stimulation, the neutrophil nuclear envelope disintegrates to enable the mixing of chromatin with granular proteins [37]. Myeloperoxidase (MPO) and neutrophil elastase (NE) stimulate chromatin condensation and deteriorate histones [38]. In the presence of histone hypercitrullination, PAD4 mediates chromatin decondensation, and the DNA-protein complexes are released extracellularly as NETs [37]. Therefore, differently from apoptosis or necrosis, both the granular membrane and nuclear membrane deteriorate during NETosis, whereas plasma membrane integrity remains [36].
The overproduction of NETs induces lung tissue damage by NETosis-related enzymes such as NE and MPO [39]. Uncontrolled NET production correlates with disease gravity and lung injury extension. For instance, NETosis markers are related to bacterial burden and local inflammation in the lung [40] and patients with pneumonia-associated ARDS have neutrophils in a “primed” condition to generate NETs [41].
During chronic obstructive pulmonary disease aggravation, the production of NETs increases in people with acute respiratory failure [39] and in ARDS patients [40, 42]. The elevated NET production, as noted in patients with severe IAV infection [43], increased injury to the pulmonary endothelial and epithelial cells [44], directing to severe pneumonia. Zhu et al. [43] also noted that the production of NETs positively correlates with multiple organ dysfunction syndromes [43].
The inflammatory process is a trigger for thrombotic complications usually noted in COVID-19 patients, and the immunothrombotic dysregulation seems to be an important key marker for the disease severity [45]. Skendros et al. [46] found that complement activation potentiates the platelet/NET/tissue factor/thrombin axis during SARS-CoV-2 infection [46]. In contrast, Nicolai et al. [47] noted that, in COVID-19, inflammatory microvascular thrombi are found in the kidney, lung, and heart, containing NETs related to the fibrin and platelets. In blood, Nicolai et al. also show that COVID-19 patients have neutrophil-platelet aggregates and a different platelet and neutrophil activation pattern, which alters with the disease severity [47]. Middleton et al. [48] also found that plasma MPO-DNA complexes increased in COVID-19 and that the elevated NET formation correlates with COVID-19-related ARDS. Together, these findings suggest the timely application of therapeutic strategies that can disrupt the vicious cycle of COVID-19 immunothrombosis/thromboinflammation by targeting neutrophil activation and NET formation.
In addition to the physical containment promoted by NETosis [33], NETs contain DNA, modified extracellular histones, proteases, and cytotoxic enzymes that allow neutrophils to centralize lethal proteins at infection sites [7]. The mechanisms of NETs' release in the viral response seem to involve neutrophil NE production attributed to the change of macrophage role by the cleavage of TLRs [49]. A range of stimuli, including toxic factors, viruses, and proinflammatory cytokines, such as TNF-α and IL-8, can lead neutrophils to release NETs [7, 33]. Mechanisms that determine strain specificity to induce NETosis formation during viral infection are still unknown.
Lung inflammation is the leading cause of the life-threatening respiratory complication at the severe levels of COVID-19 [50]. Veras et al. [51] investigated the potentially detrimental function of NETs in the pathophysiology of 32 hospitalized severe COVID-19 patients and found that the levels of NETs increase in tracheal aspirate and plasma from patients with COVID-19 and their neutrophils naturally produced more significant concentrations of NETs [51]. The authors also reported NETs in the lung tissue specimens from autopsies of COVID-19 patients. In vitro, they noted that viable SARS-CoV-2 cause NET production by healthy neutrophils through a PAD-4-dependent manner and that NETs produced by SARS-CoV-2-activated neutrophils instigated lung epithelial cell death [51]. Zuo et al. [52] also investigated sera from COVID-19 patients and found higher cell-free DNA, myeloperoxidase-DNA (MPO-DNA), and citrullinated histone H3 (Cit-H3) [52]. In vitro, they also noted that sera from COVID-19 patients trigger NET release from control neutrophils [52].
Although the literature does not report direct evidence linking NETs and SARS-CoV2 clearance, virus entrapping by NETs was already found in syncytial respiratory virus infection [53] or influenza [54]. Furthermore, in virus infection, NETs are efficient to block viruses at the infection site, entrapping them in a DNA web [22]. Therefore, the NETosis process induced by the virus could operate as a double-edged sword: on the one hand, there are essential and efficient mechanisms for trapping the virus [55], and on the other, there are highly intense immunological and inflammatory processes triggered by NET release causing damage to the organism [7]. These interactions could influence the COVID-19 symptoms in the relationship between hyperinflammation (overproduction of NETs and cytokine storm) and the function of neutrophils to destroy the viral infection (Figure 1).
Figure 1.
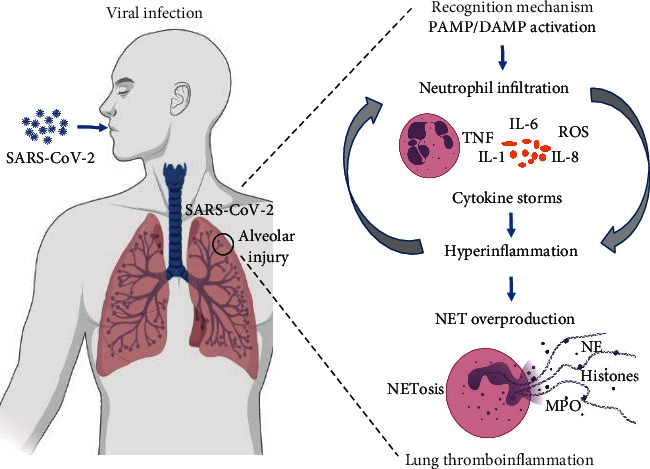
The interaction hypothesis between neutrophil and hyperinflammation in COVID-19. After the host-viral interaction, the virus signaling leads to a cascade of interactions between the virus recognition mechanism, neutrophil activation, and inflammatory stimuli. The NETosis process can protect the host during the virus response or exacerbate lung hyperinflammation in COVID-19 patients. The figure is made with BioRender (https://app.biorender.com/). Abbreviations: SARS-CoV-2: severe acute respiratory syndrome coronavirus 2; PAMP: pathogen-associated molecular pattern; DAMP: danger-associated molecular pattern; TNF: tumor necrosis factor; IL-6: interleukin-6; IL-1: interleukin-1; IL-8: interleukin-8; ROS: reactive oxygen species; NE: neutrophil elastase; MPO: myeloperoxidase.
5. Concluding Remarks and Future Directions
The exacerbated NET formation can drive to a cascade of inflammatory reactions that destroys surrounding tissues, favors microthrombosis, contributes to the progress of cancer cell metastasis, and results in permanent damage to the pulmonary, cardiovascular, and renal systems [56]. Whether by coincidence or a cause-and-effect relationship, these organs are affected in the severe state of the COVID-19 disease [57, 58]. The uncontrolled and poorly acknowledged host response regarding the cytokine storm is one of the major causes of severe COVID-19 conditions [12]. In this pandemic scenario, there is a compelling need to investigate the mechanisms associated with hyperinflammation process and NET production in response to COVID-19.
The NLR is an independent risk factor for severe COVID-19 [27], and neutrophilia forecasts poor outcomes in COVID-19 patients [29]. In this sense, new frontiers in NET assessment regarding COVID-19 may be expressed by analyzing NETosis directly after sputum induction or after bronchoscopy using the bronchial alveolar fluid of COVID-19 patients [42]. Since patient samples usually become accessible at the hospital, it could investigate whether the existence of NETs is associated with the severity of COVID-19.
Treatments using NET-targeting approaches, although would not directly target the new coronavirus, could reduce the damage caused by hyperinflammation [59], thereby decreasing the disease's severity and avoiding invasive mechanical ventilation, consequently diminishing mortality. Drugs that target NETs include inhibitors of the molecules necessary for NET formation, such as gasdermin D [60], PAD4 [61], and NE [62]. Studies on treatment of inflammatory state in COVID-19 patients with NET inhibitors are still in development (please see Table 1).
Table 1.
Interventional studies registered at the ClinicalTrials.gov database relating the treatment of COVID-19 with NET inhibitors.
| NCT identifier | Status | Location | Study type | Condition or disease | Intervention and phase | Primary outcome | Estimated completion date |
|---|---|---|---|---|---|---|---|
| NCT04409925 | Not yet recruiting | Canada | Nonrandomized pilot study | COVID-19 | Dornase Alfa Phase: 1 |
(1) Rate of all adverse events | January 2021 |
| NCT04359654 | Not yet recruiting | United Kingdom | Randomized clinical trial | COVID-19 Hypoxia |
Drug: Dornase Alfa Phase: 2 |
(1) Change in inflammation (C-reactive protein) | November 2020 |
| NCT04445285 | Recruiting | United States | Randomized clinical trial | COVID-19 | Dornase Alfa Phase: 2 |
(1) All-cause mortality (2) Systemic therapeutic response |
February 2021 |
| NCT04432987 | Recruiting | Turkey | Randomized clinical trial | COVID-19 | Dornase Alfa Phase: 2 |
(1) Clinical improvement and inflammatory markers in blood (2) Intubation or extubation |
September 2020 |
| NCT04402944 | Not yet recruiting | United States | Randomized clinical trial | COVID-19 | Dornase Alfa Phase: 2 |
(1) Ventilator-free days | December 2021 |
| NCT04322565 | Recruiting | Italy | Randomized clinical trial | COVID-19 Pneumonia |
Colchicine Phase: 2 |
(1) Clinical improvement (2) Hospital discharge |
December 2020 |
| NCT04326790 | Recruiting | Greece | Randomized clinical trial | COVID-19 | Colchicine Phase: 2 |
(1) Time to clinical deterioration (2) Concentration of cardiac troponin |
September 2020 |
| NCT04402970 | Recruiting | United States | Nonrandomized clinical trial | COVID-19 ARDS |
Dornase Alfa Phase: 3 |
(1) Improvement in partial pressure of O2 to fraction of inspired O2 ratio | May 2022 |
| NCT04355364 | Recruiting | France | Randomized clinical trial | COVID-19 ARDS |
Dornase Alfa Phase: 3 |
(1) Occurrence of at least one grade improvement (ARDS scale severity) | August 2020 |
| NCT04322682 | Recruiting | United States | Randomized clinical trial | COVID-19 | Colchicine Phase: 3 |
(1) Number of participants who die or require hospitalization | December 2020 |
| NCT04328480 | Recruiting | Argentina | Randomized clinical trial | COVID-19 | Colchicine Phase: 3 |
(1) Number of participants who die (all-cause mortality) | August 2020 |
Retrieved October 30, 2020, from https://www.https://www.clinicaltrials.gov/ct2/home. Abbreviations: NCT: National Clinical Trial; O2: oxygen; ARDS: acute respiratory distress syndrome.
Caution is needed to define which people would advantage from suppressing the neutrophil response and which would help more from a strengthened neutrophil action during viral infections. Despite prior studies linking pulmonary diseases to aberrant NET formation [3, 4], our understanding of NETosis mechanisms in viral infection is still limited.
The hyperinflammation is related to the severity of COVID-19 by influencing the pulmonary inflammation [12]. Neutrophils exhibit an intense response to virus infection, promoting bidirectional cross-talk with T cells [21]. Neutrophils also express a complex array of receptors and adhesion molecules for various ligands, including immunoglobulins and inflammatory markers [20]. In this sense, severe cases of COVID-19 appear to be related to increased NLR levels [5], and treatments using NET-targeting approaches have the potential to decrease the damage caused by hyperinflammation [40, 41]. The researchers should consider hyperinflammation in the different phases of COVID-19, neutrophil response mechanisms, and NETosis.
Acknowledgments
This work was supported by the CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) (grant number 88882.314890/2013-01); FAPESP (Fundação de Amparo a Pesquisa do Estado de São Paulo); and CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico).
Data Availability
The data supporting this narrative review are from previously reported studies and datasets, which have been cited.
Conflicts of Interest
The authors declare that there is no conflict of interest regarding the publication of this paper.
References
- 1.Kutti-Sridharan G., Vegunta R., Vegunta R., Mohan B. P., Rokkam V. R. P. SARS-CoV2 in different body fluids, risks of transmission, and preventing COVID-19: a comprehensive evidence-based review. International Journal of Preventive Medicine. 2020;11:p. 97. doi: 10.4103/ijpvm.IJPVM_255_20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Fernandes L. L., Pacheco V. B., Borges L., et al. Saliva in the diagnosis of COVID-19: a review and new research directions. Journal of Dental Research. 2020;99(13):1435–1443. doi: 10.1177/0022034520960070. [DOI] [PubMed] [Google Scholar]
- 3.Fernandes L. L., Borges L., Pacheco V. B., et al. SARS-CoV-2 : a promising path in salivary diagnosis. The Open Dentistry Journal. 2020;14(1):343–344. doi: 10.2174/1874210602014010343. [DOI] [Google Scholar]
- 4.Fernandes A. C. L., Vale A. J. M., Guzen F. P., Pinheiro F. I., Cobucci R. N., de Azevedo E. P. Therapeutic options against the new coronavirus: updated clinical and laboratory evidences. Frontiers in Medicine. 2020;7:p. 546. doi: 10.3389/fmed.2020.00546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Qin C., Zhou L., Hu Z., et al. Dysregulation of immune response in patients with coronavirus 2019 (COVID-19) in Wuhan, China. Clinical Infectious Diseases. 2020;71(15):762–768. doi: 10.1093/cid/ciaa248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ruan Q., Yang K., Wang W., Jiang L., Song J. Clinical predictors of mortality due to COVID-19 based on an analysis of data of 150 patients from Wuhan, China. Intensive Care Medicine. 2020;46(5):846–848. doi: 10.1007/s00134-020-05991-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cheng O. Z., Palaniyar N. NET balancing: a problem in inflammatory lung diseases. Frontiers in Immunology. 2013;4:p. 1. doi: 10.3389/fimmu.2013.00001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zhu G., Xu Y., Cen X., Nandakumar K. S., Liu S., Cheng K. Targeting pattern-recognition receptors to discover new small molecule immune modulators. European Journal of Medicinal Chemistry. 2018;144:82–92. doi: 10.1016/j.ejmech.2017.12.026. [DOI] [PubMed] [Google Scholar]
- 9.Min C.-K., Cheon S., Ha N.-Y., et al. Comparative and kinetic analysis of viral shedding and immunological responses in MERS patients representing a broad spectrum of disease severity. Scientific Reports. 2016;6(1, article 25359) doi: 10.1038/srep25359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Channappanavar R., Perlman S. Pathogenic human coronavirus infections: causes and consequences of cytokine storm and immunopathology. Seminars in Immunopathology. 2017;39(5):529–539. doi: 10.1007/s00281-017-0629-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kolli D., Velayutham T. S., Casola A. Host-viral interactions: role of pattern recognition receptors (PRRs) in human pneumovirus infections. Pathogens. 2013;2(2):232–263. doi: 10.3390/pathogens2020232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Mehta P., McAuley D. F., Brown M., Sanchez E., Tattersall R. S., Manson J. J. COVID-19: consider cytokine storm syndromes and immunosuppression. The Lancet. 2020;395(10229):1033–1034. doi: 10.1016/S0140-6736(20)30628-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mahallawi W. H., Khabour O. F., Zhang Q., Makhdoum H. M., Suliman B. A. MERS-CoV infection in humans is associated with a pro-inflammatory Th1 and Th17 cytokine profile. Cytokine. 2018;104:8–13. doi: 10.1016/j.cyto.2018.01.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Prete M., Favoino E., Catacchio G., Racanelli V., Perosa F. SARS-CoV-2 inflammatory syndrome. Clinical features and rationale for immunological treatment. International Journal of Molecular Sciences. 2020;21(9, article 3377) doi: 10.3390/ijms21093377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Huang C., Wang Y., Li X., et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. The Lancet. 2020;395(10223):497–506. doi: 10.1016/s0140-6736(20)30183-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Lamichhane P. P., Samarasinghe A. E. The role of innate leukocytes during influenza virus infection. Journal of Immunology Research. 2019;2019:17. doi: 10.1155/2019/8028725.8028725 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Rosales C. Neutrophils at the crossroads of innate and adaptive immunity. Journal of Leukocyte Biology. 2020;108(1):377–396. doi: 10.1002/JLB.4MIR0220-574RR. [DOI] [PubMed] [Google Scholar]
- 18.Barr F. D., Ochsenbauer C., Wira C. R., Rodriguez-Garcia M. Neutrophil extracellular traps prevent HIV infection in the female genital tract. Mucosal Immunology. 2018;11(5):1420–1428. doi: 10.1038/s41385-018-0045-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bordon J., Aliberti S., Fernandez-Botran R., et al. Understanding the roles of cytokines and neutrophil activity and neutrophil apoptosis in the protective versus deleterious inflammatory response in pneumonia. International Journal of Infectious Diseases. 2013;17(2):e76–e83. doi: 10.1016/j.ijid.2012.06.006. [DOI] [PubMed] [Google Scholar]
- 20.Russell C. D., Unger S. A., Walton M., Schwarze J. The human immune response to respiratory syncytial virus infection. Clinical Microbiology Reviews. 2017;30(2):481–502. doi: 10.1128/CMR.00090-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Costa S., Bevilacqua D., Cassatella M. A., Scapini P. Recent advances on the crosstalk between neutrophils and B or T lymphocytes. Immunology. 2019;156(1):23–32. doi: 10.1111/imm.13005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Giacalone V. D., Margaroli C., Mall M. A., Tirouvanziam R. Neutrophil adaptations upon recruitment to the lung: new concepts and implications for homeostasis and disease. International Journal of Molecular Sciences. 2020;21(3):p. 851. doi: 10.3390/ijms21030851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Pillay J., Kamp V. M., van Hoffen E., et al. A subset of neutrophils in human systemic inflammation inhibits T cell responses through Mac-1. The Journal of Clinical Investigation. 2012;122(1):327–336. doi: 10.1172/JCI57990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lukens M. V., van de Pol A. C., Coenjaerts F. E. J., et al. A systemic neutrophil response precedes robust CD8+ T-cell activation during natural respiratory syncytial virus infection in infants. Journal of Virology. 2010;84(5):2374–2383. doi: 10.1128/JVI.01807-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hufford M. M., Richardson G., Zhou H., et al. Influenza-infected neutrophils within the infected lungs act as antigen presenting cells for anti-viral CD8(+) T cells. PLoS One. 2012;7(10, article e46581) doi: 10.1371/journal.pone.0046581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lim K., Hyun Y.-M., Lambert-Emo K., et al. Neutrophil trails guide influenza-specific CD8+ T cells in the airways. Science. 2015;349(6252, article aaa 4352) doi: 10.1126/science.aaa4352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lagunas-Rangel F. A. Neutrophil-to-lymphocyte ratio and lymphocyte-to-C-reactive protein ratio in patients with severe coronavirus disease 2019 (COVID-19): a meta-analysis. Journal of Medical Virology. 2020;92(10):1733–1734. doi: 10.1002/jmv.25819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sun S., Cai X., Wang H., et al. Abnormalities of peripheral blood system in patients with COVID-19 in Wenzhou, China. Clinica Chimica Acta. 2020;507:174–180. doi: 10.1016/j.cca.2020.04.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wang D., Hu B., Hu C., et al. Clinical characteristics of 138 hospitalized patients with 2019 novel coronavirus-infected pneumonia in Wuhan, China. JAMA. 2020;323(11):1061–1069. doi: 10.1001/jama.2020.1585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Barnes B. J., Adrover J. M., Baxter-Stoltzfus A., et al. Targeting potential drivers of COVID-19: neutrophil extracellular traps. The Journal of Experimental Medicine. 2020;217(6, article e20200652) doi: 10.1084/jem.20200652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Jenne C. N., Wong C. H. Y., Zemp F. J., et al. Neutrophils recruited to sites of infection protect from virus challenge by releasing neutrophil extracellular traps. Cell Host & Microbe. 2013;13(2):169–180. doi: 10.1016/j.chom.2013.01.005. [DOI] [PubMed] [Google Scholar]
- 32.Brinkmann V., Reichard U., Goosmann C., et al. Neutrophil extracellular traps kill bacteria. Science. 2004;303(5663):1532–1535. doi: 10.1126/science.1092385. [DOI] [PubMed] [Google Scholar]
- 33.Saitoh T., Komano J., Saitoh Y., et al. Neutrophil extracellular traps mediate a host defense response to human immunodeficiency virus-1. Cell Host & Microbe. 2012;12(1):109–116. doi: 10.1016/j.chom.2012.05.015. [DOI] [PubMed] [Google Scholar]
- 34.Delgado-Rizo V., Martínez-Guzmán M. A., Iñiguez-Gutierrez L., García-Orozco A., Alvarado-Navarro A., Fafutis-Morris M. Neutrophil extracellular traps and its implications in inflammation: an overview. Frontiers in Immunology. 2017;8:p. 81. doi: 10.3389/fimmu.2017.00081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Tatsiy O., McDonald P. P. Physiological stimuli induce PAD4-dependent, ROS-independent NETosis, with early and late events controlled by discrete signaling pathways. Frontiers in Immunology. 2018;9, article 2036 doi: 10.3389/fimmu.2018.02036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Yousefi S., Stojkov D., Germic N., et al. Untangling “NETosis” from NETs. European Journal of Immunology. 2019;49(2):221–227. doi: 10.1002/eji.201747053. [DOI] [PubMed] [Google Scholar]
- 37.Fuchs T. A., Abed U., Goosmann C., et al. Novel cell death program leads to neutrophil extracellular traps. The Journal of Cell Biology. 2007;176(2):231–241. doi: 10.1083/jcb.200606027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Papayannopoulos V., Metzler K. D., Hakkim A., Zychlinsky A. Neutrophil elastase and myeloperoxidase regulate the formation of neutrophil extracellular traps. The Journal of Cell Biology. 2010;191(3):677–691. doi: 10.1083/jcb.201006052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Grabcanovic-Musija F., Obermayer A., Stoiber W., et al. Neutrophil extracellular trap (NET) formation characterises stable and exacerbated COPD and correlates with airflow limitation. Respiratory Research. 2015;16(1):p. 59. doi: 10.1186/s12931-015-0221-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Mikacenic C., Moore R., Dmyterko V., et al. Neutrophil extracellular traps (NETs) are increased in the alveolar spaces of patients with ventilator-associated pneumonia. Critical Care. 2018;22(1):p. 358. doi: 10.1186/s13054-018-2290-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bendib I., de Chaisemartin L., Granger V., et al. Neutrophil extracellular traps are elevated in patients with pneumonia-related acute respiratory distress syndrome. Anesthesiology. 2019;130(4):581–591. doi: 10.1097/ALN.0000000000002619. [DOI] [PubMed] [Google Scholar]
- 42.Wong J. J. M., Leong J. Y., Lee J. H., Albani S., Yeo J. G. Insights into the immuno-pathogenesis of acute respiratory distress syndrome. Annals of Translational Medicine. 2019;7(19):p. 504. doi: 10.21037/atm.2019.09.28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Zhu L., Liu L., Zhang Y., et al. High level of neutrophil extracellular traps correlates with poor prognosis of severe influenza A Infection. The Journal of Infectious Diseases. 2018;217(3):428–437. doi: 10.1093/infdis/jix475. [DOI] [PubMed] [Google Scholar]
- 44.Narasaraju T., Yang E., Samy R. P., et al. Excessive neutrophils and neutrophil extracellular traps contribute to acute lung injury of influenza pneumonitis. The American Journal of Pathology. 2011;179(1):199–210. doi: 10.1016/j.ajpath.2011.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Zuo Y., Zuo M., Yalavarthi S., et al. Neutrophil extracellular traps and thrombosis in COVID-19. Journal of Thrombosis and Thrombolysis. 2020;1-8 doi: 10.1101/2020.04.30.20086736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Skendros P., Mitsios A., Chrysanthopoulou A., et al. Complement and tissue factor-enriched neutrophil extracellular traps are key drivers in COVID-19 immunothrombosis. The Journal of Clinical Investigation. 2020;130(11):6151–6157. doi: 10.1172/JCI141374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Nicolai L., Leunig A., Brambs S., et al. Immunothrombotic dysregulation in COVID-19 pneumonia is associated with respiratory failure and coagulopathy. Circulation. 2020;142(12):1176–1189. doi: 10.1161/CIRCULATIONAHA.120.048488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Middleton E. A., He X.-Y., Denorme F., et al. Neutrophil extracellular traps contribute to immunothrombosis in COVID-19 acute respiratory distress syndrome. Blood. 2020;136(10):1169–1179. doi: 10.1182/blood.2020007008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Domon H., Nagai K., Maekawa T., et al. Neutrophil elastase subverts the immune response by cleaving toll-like receptors and cytokines in pneumococcal pneumonia. Frontiers in Immunology. 2018;9:p. 732. doi: 10.3389/fimmu.2018.00732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Guan W.-J., Ni Z.-Y., Hu Y., et al. Clinical characteristics of coronavirus disease 2019 in China. The New England Journal of Medicine. 2020;382(18):1708–1720. doi: 10.1056/NEJMoa2002032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Veras F. P., Pontelli M., Silva C., et al. SARS-CoV-2–triggered neutrophil extracellular traps mediate COVID-19 pathology. The Journal of Experimental Medicine. 2020;217(12, article e20201129) doi: 10.1084/jem.20201129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Zuo Y., Yalavarthi S., Shi H., et al. Neutrophil extracellular traps in COVID-19. JCI Insight. 2020;5(11, article e138999) doi: 10.1172/jci.insight.138999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Funchal G. A., Jaeger N., Czepielewski R. S., et al. Respiratory syncytial virus fusion protein promotes TLR-4-dependent neutrophil extracellular trap formation by human neutrophils. PLoS One. 2015;10(4, article e0124082) doi: 10.1371/journal.pone.0124082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Perrone L. A., Plowden J. K., García-Sastre A., Katz J. M., Tumpey T. M. H5N1 and 1918 pandemic influenza virus infection results in early and excessive infiltration of macrophages and neutrophils in the lungs of mice. PLoS Pathogens. 2008;4(8, article e1000115) doi: 10.1371/journal.ppat.1000115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Muraro S. P., De Souza G. F., Gallo S. W., et al. Respiratory syncytial virus induces the classical ROS-dependent NETosis through PAD-4 and necroptosis pathways activation. Scientific Reports. 2018;8(1, article 14166) doi: 10.1038/s41598-018-32576-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Jorch S. K., Kubes P. An emerging role for neutrophil extracellular traps in noninfectious disease. Nature Medicine. 2017;23(3):279–287. doi: 10.1038/nm.4294. [DOI] [PubMed] [Google Scholar]
- 57.Bonow R. O., Fonarow G. C., O’Gara P. T., Yancy C. W. Association of coronavirus disease 2019 (COVID-19) with myocardial injury and mortality. JAMA Cardiology. 2020;5(7):751–753. doi: 10.1001/jamacardio.2020.1105. [DOI] [PubMed] [Google Scholar]
- 58.Chen N., Zhou M., Dong X., et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: a descriptive study. The Lancet. 2020;395(10223):507–513. doi: 10.1016/s0140-6736(20)30211-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Cicco S., Cicco G., Racanelli V., Vacca A. Neutrophil extracellular traps (NETs) and damage-associated molecular patterns (DAMPs): two potential targets for COVID-19 treatment. Mediators of Inflammation. 2020;2020:25. doi: 10.1155/2020/7527953.7527953 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Hu J. J., Liu X., Xia S., et al. FDA-approved disulfiram inhibits pyroptosis by blocking gasdermin D pore formation. Nature Immunology. 2020;21(7):736–745. doi: 10.1038/s41590-020-0669-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Yost C. C., Schwertz H., Cody M. J., et al. Neonatal NET-inhibitory factor and related peptides inhibit neutrophil extracellular trap formation. The Journal of Clinical Investigation. 2016;126(10):3783–3798. doi: 10.1172/JCI83873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Tagami T., Tosa R., Omura M., et al. Effect of a selective neutrophil elastase inhibitor on mortality and ventilator-free days in patients with increased extravascular lung water: a post hoc analysis of the PiCCO Pulmonary Edema Study. Journal of Intensive Care. 2014;2(1):p. 67. doi: 10.1186/s40560-014-0067-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data supporting this narrative review are from previously reported studies and datasets, which have been cited.