

Abstract
Background
Glutathione Peroxidase 8 (GPX8) as a member of the glutathione peroxidase (GPx) family plays an important role in anti-oxidation. Besides, dysregulation of GPX8 has been found in gastric cancer, but its detailed molecular mechanism in gastric cancer has not been reported.
Methods
Our study detected the expression of GPX8 in gastric cancer tissues and cell lines using immunohistochemistry (IHC), western blot and qRT-PCR, and determined the effect of GPX8 on gastric cancer cells using CCK-8, colony formation, transwell migration and invasion assays. Besides, the effect of GPX8 on the Wnt signaling pathway was determined by western blot. Furthermore, the transcription factor of GPX8 was identified by bioinformatics methods, dual luciferase reporter and chromatin immunoprecipitation (CHIP) assays. In addition, the effect of GPX8 on tumor formation was measured by IHC and western blot.
Results
The over-expression of GPX8 was observed in gastric cancer tissues and cells, which facilitated the proliferation, migration and invasion of gastric cancer cells as well as the tumor growth. GPX8 knockdown effectively inhibited the growth of gastric cancer cells and tumors. Moreover, GPX8 could activate the Wnt signaling pathway to promote the cellular proliferation, migration and invasion through. Furthermore, FOXC1 was identified as a transcription factor of GPX8 and mediated GPX8 expression to affect cell development processes.
Conclusions
These findings contribute to understanding the molecular mechanism of GPX8 in gastric cancer. Additionally, GPX8 can be a potential biomarker for gastric cancer therapy.
Keywords: GPX8, FOXC1, Gastric cancer, Wnt signaling pathway
Background
Gastric cancer is one of the leading causes of cancer-related death in the world [1]. The risk factors for gastric cancer mainly include Helicobacter pylori infection, unhealthy lifestyle (i.e. smoking, high consumption of salts, low intake of vegetables and fruits), and genetic alterations [2]. Up to now, surgical resection is an only curative therapy for locally gastric cancer at the early stage, and surgery combined with neoadjuvant or adjuvant therapy has been applied to treat locally advanced and metastatic disease [3]. Despite advances in the therapeutic methods, the patients have a poor prognosis with approximately 25% of the 5-year survival and about 1 year of median overall survival [4]. To improve the patient’s outcome, the targeted therapy has got more and more attention [5], and finding novel biomarkers facilitates diagnosing gastric cancer early as well as monitoring recurrence or progression.
Glutathione peroxidases (GPxs) are a kind of enzyme family that can reduce hydroperoxides or hydrogen peroxide to corresponding alcohols or water [6]. The GPX family is divided into two groups, selenocysteine GPxs (GPX1, 2, 3, 4, and 6) and cysteine GPXs (GPX5, 7, and 8), based on their catalytic center’s amino acid tetrad [7]. Despite their antioxidation, GPXs are proved to be critical for the development of cancers [8]. For instance, GPX1 is regarded as a potential biomarker for kidney cancer treatment and diagnosis and its high expression promotes the disease’s progression [9]. GPX3 hypermethylation has been found in gastric cancer, which is associated with tumor recurrence in gastric cancer patients over 60 years old [10]. GPX8, as a unique GPX, is located in the membranes of ER (endoplasmic reticulum) and prevent the spill of hydrogen peroxide to control redox status [11]. Dysregulation of GPX8 has been found in several tumors such as gastric cancer [12–14], but its molecular mechanism in gastric cancer has not been reported.
To address this issue, we firstly used bioinformatics methods to identify the potential transcription factors of GPX8 and chose Forkhead box C1 (FOXC1) as an upstream candidate molecule for further analysis. Next, we selected the Wnt signaling pathway as a downstream pathway of GPX8 according to the previous report [13]. Then, we determined the effects and molecular mechanism of GPX8 in gastric cancer cells using a series of biological experiments.
Methods
Bioinformatics methods
We used the Gene Global of Biotechnology Information (GCBI) website (https://www.gcbi.com.cn/gclib/html/index) to find the underlying transcription factors of GPX8. The GCBI website analyzed the datasets from Transfac, COSMIC and dbSNP databases. In the GCBI website, we firstly registered a paid member, clicked the column of Gene radar, and then entered “GPX8” to search its transcription factors. Finally, we obtained 37 transcription factors of GPX8. The information of 37 transcription factors was listed in Additional file 1: Table S1. Compared with other transcription factors, FOXC1 was chosen for the following experiments because it had the higher prediction score and more DNA-binding locations and plays an important role in the metastasis and development of tumors [15].
According to KEGG (Kyoto Gene and Genome Encyclopedia) pathway, Zhang et al. used GSEA (Gene set enrichment analysis) to indicate a positive correlation between GPX8 expression and basal cell carcinoma signaling pathway [13]. Based on the above literature, we clicked the column of “KEGG PATHWAY” in the KEGG website (https://www.kegg.jp/) and entered “basal cell carcinoma signaling pathway” to obtain the pathway information (map05217, Additional file 2: Figure S1).
Clinical specimens
We collected 30 paired tumor and adjacent non-tumor tissues from gastric cancer patients who diagnosed and underwent surgery at the First Affiliated Hospital of Soochow University between January 2018 and October 2018. All patients received no treatment before surgery. They provided informed consent regarding this experiment. The Human Research Ethics Committee of the First Affiliated Hospital of Soochow University approved this study (Ethical Approval No. 2017028).
Cell culture and transfection
Normal human gastric epithelial cells (GES-1) were obtained from the Beijing Institute of Cancer Research (Beijing, China). Gastric cancer cell lines (NCI-N87, SNU-16, SNU-5 and HGC-27) were purchased from ATCC (American Type Culture Collection). All cell lines were cultured in RPMI-1640 (GIBCO, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS, GIBCO), streptomycin (100 mg/ml, GIBCO) and penicillin (100 U/ml, GIBCO) at 37℃ under 5% CO2 in a humidified incubator.
GPX8 siRNA (si-GPX8), FOXC1 siRNA (si-FOXC1) and corresponding negative control siRNAs (si-NC) were purchased from Thermo Fisher Scientific, and they were transfected into HGC-27 cells using Lipofectamine® RNAiMAX Reagent (Invitrogen, Carlsbad, USA). GPX8 overexpression plasmids (pcDNA-GPX8), FOXC1 overexpression plasmids (pcDNA-FOXC1) and corresponding plasmids (pcDNA-control) were obtained from HANBIO (Shanghai, China). These plasmids were respectively transfected into HGC-27 cells via Lipofectamine™ 3000 Reagent (Invitrogen).
RNA isolation and qRT-PCR
Total RNA from gastric cancer tissues and cells were isolated by TRIzol reagent (Invitrogen), followed by the quantification of RNA concentration using NanoDrop2000c (Thermo Fisher Scientific). The reverse transcription of RNA to cDNA was carried out using a PrimeScript RT reagent kit (Takara, Dalian, China). Next, in an ABI 7500 system, a SYBR® Premix Ex Taq™ RT-PCR Kit (Takara) was used to perform the quantitative PCR according to the manufacturer’s protocol. The relative expression of target molecules was calculated by the 2−ΔΔCq method, with normalization of the internal control, β-actin. The premier sequences were listed in Table 1.
Table 1.
The sequences of the primers
| Gene name | Sequence (5′-3′) |
|---|---|
| GPX8 forward | GTTTCACTAGTTGTAAACGTGGC |
| GPX8 reverse | CGATTCTCCAAACTGATTGCAGG |
| Foxc1 forward | ACTCGGTGCGGGAGATGTT |
| Foxc1 reverse | CCTTGATGGGTTCCTTTAGC |
| β-actin forward | TCCTCCTGAGCGCAAGTACTCC |
| β-actin reverse | CATACTCCTGCTTGCTGATCCAC |
Western blot
Gastric cancer tissues and cells were lysed by RIPA lysis buffer containing protease inhibitor cocktail (Beyotime, Jiangsu, China) to obtain total protein, and its concentration was detected and quantified using a BCA assay kit (Beyotime). Next, 20 μg of equal protein was separated on 10% SDS-PAGE, followed by transferring to PVDF membranes. After blocking with 5% non-fat milk at room temperature for 1 h, the membranes were incubated at 4℃ overnight with primary antibodies against GPX8 (1/1000 dilution, ab183664), FOXC1 (1/1000 dilution, #8758, ab227977), Wnt1 (1/1000 dilution, ab15251), Wnt3a (1/1000 dilution, ab219412), p-GSK3β (1/1000 dilution, ab131097), GSK3β (1/5000 dilution, ab32391), β-catenin (1/5000 dilution, ab32572), and β-actin (1/1000 dilution, ab8227), washed with 0.1 M PBS with 0.05% Tween20, and then cultured with HRP (horseradish peroxidase)-conjugated secondary antibodies (Goat Anti-Rabbit IgG H&L, 1/2000 dilution, ab205718) at room temperature for 1 h. Finally, the membranes were determined using a chemiluminescence assay kit (Bio-Rad Laboratories). The primary and secondary antibodies were purchased from Abcam. The quantification of protein bands was assessed by the ImageJ software.
Immunohistochemistry (IHC)
Gastric cancer tissues or adjacent non-tumor tissues were fixed with 2% paraformaldehyde at room temperature for 24 h, rinsed with running tap water for 5 min, dehydrated with 70%, 80%, 95% and 100% alcohol for 5 min each, cleared with xylene twice for 5 min each, then embedded with paraffin, and sectioned at 5–8 μm thickness. Next, the tissue sections were deparaffinized in xylene twice for 5 min each, rehydrated respectively using 100%, 95%, 70% and 50% alcohols for 3 min each. The sections were incubated in methanol with 3% H2O2 for 10 min at room temperature to block endogenous peroxidase activity. After washing with PBS twice, antigen retrieval was performed using a citrate buffer method. The sections were washed with PBS twice, blocked with 10% FBS in PBS for 1 h at room temperature, and incubated at 37℃ for 2 h with primary antibodies against GPX8 (1/1000 dilution, ab183664, Abcam) and Ki-67 (1/200 dilution, ab16667, Abcam). After rinsing with PBS twice, the sections were incubated with secondary antibodies (Goat Anti-Rabbit IgG H&L (HRP), 1/2000 dilution, ab205718) for 30 min at room temperature. Next, the sections were washed with PBS twice, incubated with Sav-HRP conjugates solution (Thermo Fisher Scientific) for 30 min at room temperature, washed with PBS, stained with DAB (3,3′-diaminobenzidine, Abcam) substrate solution, counterstained with hematoxylin, dehydrated with alcohol, cleared with xylene and mounted with resin. The antibody staining was recorded under a light microscope (Olympus Corporation, ×200 magnification).
Cell counting kit-8 (CCK-8) assay
After transfection for 48 h, HGC-27 cells were harvested, inoculated in 96 well-plates (1 × 104 cells/well), and respectively incubated in complete medium for 1, 2, 3 and 4 days. Next, the cells were treated with CCK-8 solution (10 μl per well) for 4 h of culture. The sample’s absorbance was examined by a microplate reader at 450 nm.
Colony formation assay
48 h post-transfection, HGC-27 cells were collected and inoculated in culture dishes, followed by incubating in complete culture medium for 2 weeks at 37 °C in a humidified incubator under 5% CO2. During cultivation, the medium was changed every 3 or 4 days. After incubation, the cells were cleared with PBS, fixed with 4% paraformaldehyde for 15 min and stained by GIMSA for 20 min. The image of the cell colony was recorded by an Olympus BX51 light microscope (Olympus Corporation).
Transwell migration and invasion assays
HGC-27 cells, 48 h post-transfection, were harvested, suspended in serum-free medium, and seeded on the upper chamber of transwell (Corning, 8 μm pore) coated with or without Matrigel (BD Biosciences). RPMI-1640 medium with 10% FBS was added in the transwell’s lower chamber and was used as a chemical inducer. After 24 h of culture, cotton swabs were utilized to remove the cells that stayed on the upper chamber. The migrated or invaded cells were fixed with 10% formaldehyde for 20 min, followed by staining using hematoxylin for 10 min as well as recording their images with an Olympus BX51 microscope (Olympus Corporation).
Dual luciferase reporter assay
The fragments of GPX8 that contained the DNA-binding sites between GPX8 and FOXC1 were cloned into pGL3-vectors (Promega), obtaining GPX8 promoter reporter vectors (GPX8-WT). The corresponding control reporter vectors (GPX8-MUT) were obtained from the fragments of GPX8 with the mutation DNA-binding sites cloned into pGL3-vectors. These reporter vectors with FOXC1 overexpression plasmids (pcDNA-FOXC1) or corresponding plasmids (pcDNA-control) were co-transfected into HGC-27 cells using Lipofectamine™ 3000 Reagent (Invitrogen). The relative luciferase activity = firefly luciferase activity/Renill luciferase activity.
Chromatin immunoprecipitation (CHIP) assay
HGC-27 cells were cross-linked by a 1% final concentration of formaldehyde for 10 min, quenched using a final concentration of 125 mM glycine for 5 min and cleared with PBS. The cells were added with lysis buffer containing protease inhibitor for 10 min and then sonicated to shear chromatin to fragments of 200–500 bp. The lysates were centrifuged at 12,000g for 10 min. The supernatant was collected in a clean tube, and added FOXC1 antibodies and normal rabbit lgG, followed by incubating 15 min at room temperature. Then, secondary antibodies (Goat Anti-Rabbit IgG H&L (Biotin)) were added to the samples for 15 min incubation at room temperature. Streptavidin beads were added to the samples with gentle rotation for 30 min at 4 °C A High salt solution (5 M NaCl) was used to perform reverse cross-linking to obtain DNA fragments. The DNA fragments containing the DNA-binding domain of GPX8 with FOXC1 were further validated by PCR.
Tumor formation in mice
BALB/c nude male mice, 6-week-old, 15–18 g, were purchased from Laboratory Animal Resources, Chinese Academy of Sciences (Beijing, China). The mice were housed in a SPF (specific pathogen-free) room at 25–27 °C, 40–50% humidity under a 12 h light/dark cycle. This experiment was approved by the Animal Care and Use Committee of the First Affiliated Hospital of Soochow University and carried out based on the rules of the Guide for Care. HGC-27 cells were respectively transfected with blank control vector, pcDNA-control, pcDNA-GPX8, sh-NC and sh-GPX8, and then harvested using trypsinization at the logarithmic growth phase. The cell suspension at the concentration of 5 × 106 cells per mouse was subcutaneously injected into mice. After inoculation, the tumor sizes were recorded every seven days. The formula of the tumor volume = 1/2 L × W2; L presents the long diameter, and W represents the short diameter. After 28 days, the mice were euthanized and sacrificed. The expression of GPX8 and Ki67 in mice tumors was measured by IHC assay.
Statistical analysis
The data from three independently repeated experiments were presented as mean ± SD (standard deviation) and analyzed by GraphPad Prism 7 software. The comparison between two groups was calculated by Student’s t test, and data among multiple groups was compared by one-way ANOVA with a Tukey multiple comparison test. A p-value of less than 0.05 indicates a significant difference.
Results
The up-regulation of GPX8 in gastric cancer
To detect the expression of GPX8 in gastric cancer, we firstly performed IHC, western blot and qRT-PCR assays to determine the level of GPX8 in tumor and adjacent non-tumor tissues from gastric cancer patients. The images of IHC assay showed that GPX8 expression was positive in gastric cancer tumors compared with adjacent non-tumor (normal control) tissues (Fig. 1a, b). Western blot revealed that the protein expression of GPX8 was up-regulated in gastric cancer tumors as compared to the normal control group (Fig. 1c,d). The mRNA expression of GPX8 detected by qRT-PCR was also up-regulated in gastric cancer tumors (Fig. 1e).
Fig. 1.

The expression of GPX8 in gastric cancer tissues and cell lines. a, b The expression of GPX8 in the tumor and adjacent non-tumor tissues from gastric cancer patients was detected by immunohistochemistry (IHC). The c, d protein and e mRNA expression of GPX8 in the patient’s tumors was respectively assessed by western blot and qRT-PCR. The f, g protein and h mRNA expression levels of GPX8 in normal human gastric epithelial cells (GES-1) and Gastric cancer cell lines (NCI-N87, SNU-16, SNU-5 and HGC-27) were respectively measured by western blot and qRT-PCR. **p < 0.01, ***p < 0.001, ****p < 0.0001 versus Normal control or GES-1
Furthermore, the expression of GPX8 in normal human gastric epithelial cells (GES-1) and gastric cancer cell lines (NCI-N87, SNU-16, SNU-5 and HGC-27) was validated by western blot and qRT-PCR. Gastric cancer cell lines showed a relative high level of GPX8 compared with GSE-1 cells (Fig. 1f–h). Collectively, GPX8 was over-expressed in gastric cancer tissues and cell lines.
The effect of GPX8 on gastric cancer cells
To study the effect of GPX8 on gastric cancer cells, we measured the cellular proliferation, migration and invasion. HGC-27 cells were transfected with blank control vector, pcDNA-control, pcDNA-GPX8, si-NC and si-GPX8, and the changes of GPX8 expression was detected by western blot and qRT-PCR. The results indicated that pcDNA-GPX8 significantly increased the protein and mRNA expression of GPX8 compared with pcDNA-control, and si-GPX8 obviously decreased the expression of GPX8 as compared to siRNA (Fig. 2a–c). The overexpression of GPX8 enhanced the cell viability and colony formation ability, and the knockdown of GPX8 suppressed the cell viability and colony formation ability compared with the control groups (Fig. 2d–f). Thus, the high expression of GPX8 could promote the proliferation of gastric cancer cells. In addition, transwell assays suggested that GPX8 overexpression promoted the migration and invasion of gastric cancer cells and yet GPX8 knockdown inhibited the cell migration and invasion (Fig. 2e, g, h).
Fig. 2.

The effect of GPX8 on gastric cancer cells. After HGC-27 cells were transfected with blank control vector, pcDNA-control, pcDNA-GPX8, si-NC and si-GPX8, the a, b protein and c mRNA expression of GPX8 were respectively measured by western blot and qRT-PCR; d the proliferation of HGC-27 cells were examined by CCK-8 assay; e The colony formation, migration and migration abilities were respectively detected by colony formation assay, transwell migration and invasion assays. The images of data quantification of f colony formation, g transwell migration and h migration assays. **p < 0.01, ***p < 0.001, ****p < 0.0001 versus pcDNA-control; ###p < 0.001, ####p < 0.0001 Versus si-NC
The effect of GPX8 on the Wnt signaling pathway
Zhang et al. have analyzed the gastric cancer data in TCGA (the Cancer Genome Atlas) database, found a potential prognostic biomarker- GPX8,and proved the high expression of GPX8 was closely correlated with basal cell carcinoma signaling pathway [13]. Based on KEGG pathway, Basal cell carcinoma signaling includes the Wnt signaling pathway (Additional file 2: Figure S1) that is critical for tumorigenesis and development [16]. Accordingly, we selected the Wnt signaling pathway as a potential downstream pathway of GPX8.
We determined the effect of GPX8 on the Wnt signaling pathway using a series of experiments. Firstly, compared with GES-1 cells, the increase of wnt1, wnt3a and β-catenin and the decrease of p-GSK3β were found in gastric cancer cell lines (NCI-N87, SNU-16, SNU-5 and HGC-27), and consequently the Wnt signaling pathway was activated in gastric cancer cells (Fig. 3a, b). After HGC-27 cells were transfected with blank control vector, pcDNA-control, pcDNA-GPX8, si-NC and si-GPX8, the overexpression of GPX8 greatly up-regulated the expression of wnt1, wnt3a and β-catenin and down-regulated the expression of p-GSK3β as compared to pcDNA-control (Fig. 3c, d). In addition, the knockdown of GPX8 decreased the levels of wnt1, wnt3a and β-catenin and increased the levels of p-GSK3β compared with the si-NC group (Fig. 3c, d). Thus, the high expression of GPX8 could activate the Wnt signaling pathway.
Fig. 3.

The effects of GPX8 on the Wnt signaling pathway. a, b The expression levels of key proteins (wnt1, wnt3a, p-GSK3β, GSK3β and β-catenin) of the Wnt signaling pathway were examined using western blot in normal human gastric epithelial cells (GES-1) and Gastric cancer cell lines (NCI-N87, SNU-16, SNU-5 and HGC-27). c, d After HGC-27 cells were transfected with blank control vector, pcDNA-control, pcDNA-GPX8, si-NC and si-GPX8, the expression levels of wnt1, wnt3a, p-GSK3β, GSK3β and β-catenin were detected using western blot. After pcDNA-GPX8 combined with or without Wnt inhibitor was co-transfected into HGC-27 cells, c the cellular proliferation was assessed by CCK-8 assay; f Colony formation assay, transwell migration and invasion assays were utilized to respectively detect the cellular colony formation, migration and invasion abilities. The images of data quantification of g colony formation, h transwell migration and i migration assays. ****p < 0.0001 Versus GES-1 or pcDNA-control; ###p < 0.001, ####p < 0.0001 versus si-NC or pcDNA-GPX8
Furthermore, CCK-8, colony formation and transwell assays indicated that the promoting effect of the proliferation, migration, invasion of gastric cancer cells induced by GPX8 overexpression was weakened by the introduction of JW 55 as the Wnt signaling pathway inhibitor (Fig. 3e–i). Therefore, the high expression of GPX8 promoted the proliferation, migration and invasion of gastric cancer cells through activating the Wnt signaling pathway.
The prediction of the underlying transcription factors of GPX8
To further explore the molecular mechanism of GPX8 in gastric cancer cells, we used the GCBI website to find the potential transcription factors of GPX8. As shown in Fig. 4, a total of 37 transcription factors of GPX8 were obtained, among which FOXC1 was chosen as a candidate for further analysis. The reason for our choice was FOXC1 with the higher prediction score, more binding locations and its key roles in the progression of tumors [15]. The DNA-binding domains of GPX8 with FOXC1 are listed in Table 2.
Fig. 4.
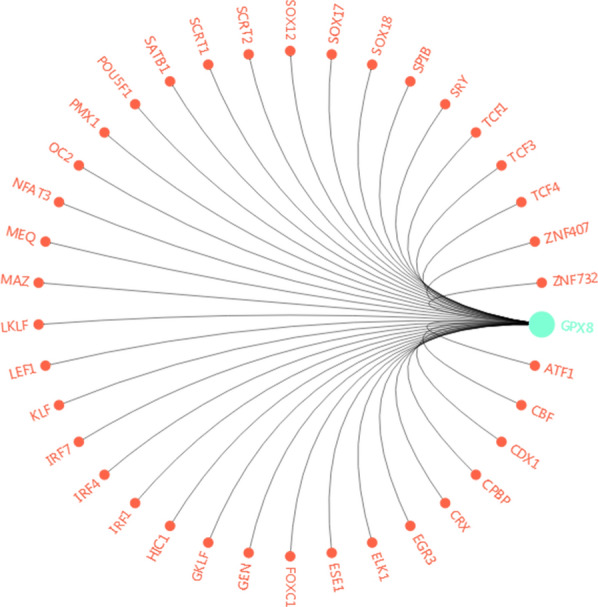
The image of the potential transcription factors of GPX8
Table 2.
The prediction binding domain between GPX8 and FOXC1
| Gene name | Transcript name | Transcription factor | Chromosome location | Starting position | Ending position | Sequence | Prediction score |
|---|---|---|---|---|---|---|---|
| GPX8 | ENST00000296734.6 | FOXC1 | 5 | 55158430 | 55158439 | TTGCTtttt | 1 |
| GPX8 | ENST00000296734.6 | FOXC1 | 5 | 55159007 | 55159016 | TTGCTtttt | 1 |
| GPX8 | ENST00000503787.5 | FOXC1 | 5 | 55158430 | 55158439 | TTGCTtttt | 1 |
| GPX8 | ENST00000503787.5 | FOXC1 | 5 | 55159007 | 55159016 | TTGCTtttt | 1 |
| GPX8 | ENST00000515370.1 | FOXC1 | 5 | 55158430 | 55158439 | TTGCTtttt | 1 |
| GPX8 | ENST00000515370.1 | FOXC1 | 5 | 55159007 | 55159016 | TTGCTtttt | 1 |
The roles of FOXC1 in gastric cancer cells and the relationship between FOXC1 and GPX8
To explore the relationship between GPX8 and FOXC1, we separately detected the roles of FOXC1 in gastric cancer cells and assessed the targeting relationship of GPX8 with FOXC1. Firstly, gastric cancer cell lines (NCI-N87, SNU-16, SNU-5 and HGC-27) showed higher protein and mRNA expression of FOXC1 than GES-1 cells (Fig. 5a–c). After HGC-27 cells were transfected with blank control vector, pcDNA-control, pcDNA-FOXC1, si-NC or si-FOXC1, pcDNA-FOXC1 obviously up-regulated the protein and mRNA expression of FOXC1 in comparison to pcDNA-control and yet si-FOXC1 greatly down-regulated FOXC1 expression compared with si-NC (Fig. 5d–f). Furthermore, FOXC1 overexpression enhanced the cell viability and colony formation ability but FOXC1 knockdown suppressed the cell viability and colony formation ability (Fig. 5g–i). Additionally, the overexpression of FOXC1 promoted the migration and invasion of gastric cancer cells, and yet knockdown of FOXC1 inhibited the cell migration and invasion (Fig. 5h–k). Hence, the high expression of FOXC1 promoted the proliferation, migration and invasion of gastric cancer cells. Furthermore, Dual luciferase reporter and CHIP assays were used to detect the binding relationship of GPX8 with FOXC1. Dual luciferase reporter assay indicated that the luciferase activity of the wild GPX8 promoter reporter vector was significantly up-regulated by the introduction of pcDNA-FOXC1 compared with pcDNA-control, and yet the luciferase activity in mutant GPX8 promoter reporter vector had no significant changes after co-transfection with pcDNA-control or pcDNA-FOXC1 (Fig. 6a). Thus, FOXC1 could target the DNA-binding domain of GPX8 to enhance the luciferase activity. CHIP assay showed that GPX8 DNA fragment could be immunoprecipitated by anti-FOXC1 antibodies compared with the control (lgG), and consequently further validated that FOXC1 protein directly combined with GPX8 DNA (Fig. 6b, c). After pcDNA-FOXC1, si-FOXC1 and corresponding controls were respectively transfected into HGC-27 cells, pcDNA-FOXC1 up-regulated the protein and mRNA expression of GPX8 compared with pcDNA-control but si-FOXC1 down-regulated the expression of GPX8 as compared to si-NC (Fig. 6d–f), indicating that FOXC1 induced GPX8 expression. Moreover, GPX8 overexpression enhanced cell viability and colony formation ability and promoted cell migration and invasion, which was reversed by the introduction of si-FOXC1 (Fig. 6g–k). Hence, FOXC1 could mediate GPX8 to regulate the proliferation, migration and invasion of gastric cancer cells.
Fig. 5.

The roles of FOXC1 in gastric cancer cells. The a, b protein and c mRNA expression levels of FOXC1 in normal human gastric epithelial cells (GES-1) and gastric cancer cell lines (NCI-N87, SNU-16, SNU-5 and HGC-27) were determined using western blot and qRT-PCR. HGC-27 cells were transfected with blank control vector, pcDNA-control, pcDNA-FOXC1, si-NC and si-FOXC1, respectively, and then the d, e protein and f mRNA expression levels of FOXC1 were assessed by western blot and qRT-PCR. g The cellular proliferation was examined by CCK-8 assay; h The colony formation, migration and migration abilities were respectively measured using colony formation assay, transwell migration and invasion assays. The images of data quantification of i colony formation, j transwell migration and (K) migration assays. ***p < 0.001, ****p < 0.0001 Versus GES-1 or pcDNA-control; ##p < 0.01, ###p < 0.001, ####p < 0.0001 versus si-NC
Fig. 6.

The relationship between FOXC1 and GPX8. The binding relationship of GPX8 with FOXC1 was determined by a dual luciferase reporter and b, c chromatin immunoprecipitation (CHIP) assays. After HGC-27 cells were transfected with pcDNA-control, pcDNA-FOXC1, si-NC or si-FOXC1, the d, e protein and f mRNA expression levels of GPX8 were detected by western blot and qRT-PCR. After HGC-27 cells were respectively transfected with pcDNA-control + si-NC, pcDNA-GPX8 + si-NC and pcDNA-GPX8 + si-FOXC1, g CCK-8 was used to determine the cell viability; h colony formation assay, transwell migration and invasion assays were applied to detect the cell proliferation, migration and invasion. The images of data quantification of i colony formation, (J) migration and (K) migration assays. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 Versus pcDNA-control, lgG or pcDNA-control + si-NC; #p < 0.05, ##p < 0.01, ###p < 0.001, ####p < 0.0001 Versus si-NC or pcDNA-GPX8 + si-NC
Taken together, FOXC1 was a transcription factor of GPX8, and the interaction of GPX8 with FOXC1 had promoting roles in the growth of gastric cancer cells.
The effect of GPX8 on tumor growth
To investigate the effect of GPX8 on gastric cancer in vivo, we constructed the gastric cancer tumor model in nude mice. The tumors were treated with blank control vector, pcDNA-control, pcDNA-GPX8, sh-NC and sh-GPX8, respectively. After 28 days of feeding, the mice were sacrificed and the tumors were removed. IHC assays showed that pcDNA-GPX8 up-regulated the expression of GPX8 compared with pcDNA-control and sh-GPX8 down-regulated the levels of GPX8 as compared to sh-NC (Fig. 7a, b). In addition, the proliferation biomarker, ki-67, was up-regulated by GPX8 overexpression and down-regulated by GPX8 knockdown (Fig. 7a, c).
Fig. 7.

The effects of GPX8 on tumor growth. HGC-27 cells were respectively transfected with blank control vector, pcDNA-control, pcDNA-GPX8, sh-NC and sh-GPX8, collected and subcutaneously injected into mice. After inoculation, the tumor sizes were recorded every seven days, and the mice were euthanized and sacrificed on day 28. a The expression of GPX8 and Ki-67 in the mice tumors were determined by immunohistochemistry (IHC) assay. b, c The images of positive expression quantification of GPX8 and Ki-67 in IHC assay. d The images of the mice tumors on day 28. e The changes of tumor volumes during 28 days of feeding. *p < 0.05, **p < 0.01, ****p < 0.0001 Versus pcDNA-control; ##p < 0.01, ####p < 0.0001 versus si-NC
Furthermore, as compared to pcDNA-control, the greater size of the tumor was observed in the GPX8 overexpression group, and yet GPX8 knockdown decreased the tumor size compared with the sh-NC group (Fig. 8d). In addition, the volume of the tumors showed an obvious growth trend in the GPX8 overexpression group, but GPX8 knockdown suppressed the growth trend of the tumors (Fig. 8e). Therefore, the high expression of GPX8 promoted the growth of gastric cancer tumors, and GPX8 knockdown inhibited the tumor growth.
Fig. 8.

Proposed molecular mechanisms of GPX8 in gastric cancer cells. GPX8 is regulated by its transcription factor FOXC1 and activates the Wnt signaling pathway to promote the proliferation, migration and invasion of gastric cancer cells
Discussion
Our work indicated that the aberrant high expression of GPX8 was detected in gastric cancer tissues and cell lines, which promoted the proliferation, migration and invasion of gastric cancer cells. Furthermore, GPX8 overexpression promoted the cell growth through activating the Wnt signaling pathway. FOXC1 was proved to be a transcription factor of GPX8 and mediated GPX8 expression to regulate cell development processes. Additionally, GPX8 overexpression in nude mice promoted the growth of gastric cancer tumors and yet GPX8 knockdown significantly suppressed the tumor growth.
The targeted therapy is a research hotspot for improving the prognosis of gastric cancer patients [5]. Finding novel and potent biomarkers is a key step for investigating targeted therapy. GPX8 as a member of the GPX family plays an important role in protecting cells against oxidative stress [11]. Moreover, several reports indicated GPX8 dysregulation in gastric cancers through bioinformatics analysis [12, 13]. However, the detailed molecular mechanism of GPX8 in gastric cancer has not been reported. To study this issue, we detected the expression of GPX8 in gastric cancer and detected the overexpression of GPX8 in gastric cancer tissues and cell lines. This result was consistent with the previous report that revealed the aberrant high level of GPX8 in gastric cancer tissues compared with normal controls based on TCGA database [13]. Additionally, our study showed that GPX8 overexpression promoted the proliferation, migration and invasion of gastric cancer cells. Besides, GPX8 knockdown obviously inhibited the cell growth. Moreover, the overexpression of GPX8 promoted the growth of gastric cancer tumors in nude mice and yet GPX8 knockdown suppressed the tumor growth. Therefore, GPX8 had the potential to be a biomarker for gastric cancer therapy.
Based on the previous report [13], we predicted the Wnt signaling pathway as the down-stream molecular pathway of GPX8. The Wnt signaling pathway is the crucial regulatory pathway of tumorigenesis and progression [17]. It is closely associated with the development of many cellular processes such as proliferation, apoptosis and metastasis [16]. Wnt1 and wnt3a are involved in oncogenesis as well as in biological processes such as cell survival [18, 19]. The phosphorylation of GSK3β can effectively suppress the activity of the Wnt pathway [20]. β-catenin can translocate cytoplasm to the nucleus to activate the expression of Wnt target genes [21]. Our study found that GPX8 overexpression increased the expression of wnt1, wnt3a and β-catenin and decreased the expression of p-GSK3β, thereby activating the Wnt signaling pathway. Besides, the knockdown of GPX8 suppressed the activation of the Wnt signaling pathway. Moreover, the promoting effect of the proliferation, migration, invasion of gastric cancer cells induced by GPX8 overexpression was weakened by the introduction of the Wnt signaling pathway inhibitor, JW 55. Therefore, the overexpression of GPX8 promoted the cellular proliferation, migration and invasion through activating the Wnt signaling pathway.
To explore the detail molecular mechanism of GPX8 in gastric cancer, we used the GCBI website to identify FOXC1 as a potential transcription factor of GPX8. Furthermore, dual luciferase reporter and CHIP assays confirmed that FOXC1 was a transcription factor of GPX8. FOXC1 transcription factor is critical for the occurrence and progression of a wide range of cancers [15]. Up-regulated FOXC1 expression is reported to be positively correlated with poor prognosis in various cancers such as gastric cancer [22]. FOXC1 can enhance the proliferation, differentiation, survival and metastasis of different cancer cells, including gastric cancer cells [23, 24]. To agreement with the above previous reports [22–24], our study showed that FOXC1 was over-expressed in gastric cancer cells. FOXC1 overexpression promoted the proliferation, migration and invasion of gastric cancer cells, and FOXC1 knockdown had the opposite effect. Furthermore, FOXC1 overexpression increased GPX8 expression and FOXC1 knockdown decreased GPX8 expression. Moreover, FOXC1 could target GPX8 to regulate the growth of gastric cancer cells.
Conclusions
In conclusion, the overexpression of GPX8 was shown in gastric cancer tissues and cell lines. In addition, GPX8 was regulated by its transcription factor FOXC1, and promoted the proliferation, migration and invasion of gastric cancer cells by activating the Wnt signaling pathway (Fig. 8). Therefore, GPX8 can be a potential biomarker for the targeted therapy of gastric cancer and these results contribute to understanding the molecular mechanism of GPX8 in gastric cancer.
Supplementary Information
Additional file 1: Table S1. The information of predicted transcription factors of GPX8.
Additional file 2: Figure S1. The images of basal cell carcinoma signaling pathway from the KEGG database.
Acknowledgements
Not applicable.
Abbreviations
- GPX8
Glutathione peroxidase 8
- GPx
The glutathione peroxidase
- IHC
Immunohistochemistry
- CHIP
Chromatin immunoprecipitation
Authors’ contributions
HC, LX, HH: Conceptualization. HC, LX: Writing- Original draft preparation. ZLS, SC, HH: Formal analysis. All authors read and approved the final manuscript.
Funding
Not applicable.
Availability of data and materials
All data generated or analysed during this study are included in this published article and its additional information files.
Ethics approval and consent to participate
The Human Research Ethics Committee of the First Affiliated Hospital of Soochow University approved this study.
This experiment was approved by the Animal Care and Use Committee of the First Affiliated Hospital of Soochow University and carried out based on the rules of the Guide for Care.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
This article has been retracted. Please see the retraction notice for more detail:https://doi.org/10.1186/s12935-024-03269-6
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Hong Chen and Lu Xu contributed equally to this work.
Change history
2/22/2024
This article has been retracted. Please see the Retraction Notice for more detail: 10.1186/s12935-024-03269-6
Supplementary Information
The online version contains supplementary material available at 10.1186/s12935-020-01692-z.
References
- 1.Kinoshita H, Hayakawa Y, Koike K. Metaplasia in the Stomach-Precursor of Gastric Cancer? Int J Mol Sci. 2017;18(10):2063. doi: 10.3390/ijms18102063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Padmanabhan N, Ushijima T, Tan P. How to stomach an epigenetic insult: the gastric cancer epigenome. Nat Rev Gastroenterol Hepatol. 2017;14(8):467–478. doi: 10.1038/nrgastro.2017.53. [DOI] [PubMed] [Google Scholar]
- 3.Van Cutsem E, Sagaert X, Topal B, Haustermans K, Prenen H. Gastric cancer. Lancet (London, England) 2016;388(10060):2654–2664. doi: 10.1016/S0140-6736(16)30354-3. [DOI] [PubMed] [Google Scholar]
- 4.Figueiredo C, Camargo MC, Leite M, Fuentes-Pananá EM, Rabkin CS, Machado JC. Pathogenesis of gastric cancer: genetics and molecular classification. Curr Top Microbiol Immunol. 2017;400:277–304. doi: 10.1007/978-3-319-50520-6_12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.McLean MH, El-Omar EM. Genetics of gastric cancer. Nat Rev Gastroenterol Hepatol. 2014;11(11):664–674. doi: 10.1038/nrgastro.2014.143. [DOI] [PubMed] [Google Scholar]
- 6.Toppo S, Vanin S, Bosello V, Tosatto SC. Evolutionary and structural insights into the multifaceted glutathione peroxidase (Gpx) superfamily. Antioxid Redox Signal. 2008;10(9):1501–1514. doi: 10.1089/ars.2008.2057. [DOI] [PubMed] [Google Scholar]
- 7.Ramming T, Appenzeller-Herzog C. Destroy and exploit: catalyzed removal of hydroperoxides from the endoplasmic reticulum. Int J Cell Biol. 2013;2013:180906. doi: 10.1155/2013/180906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zhang ML, Wu HT, Chen WJ, Xu Y, Ye QQ, Shen JX, et al. Involvement of glutathione peroxidases in the occurrence and development of breast cancers. J Transl Med. 2020;18(1):247. doi: 10.1186/s12967-020-02420-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cheng Y, Xu T, Li S, Ruan H. GPX1, a biomarker for the diagnosis and prognosis of kidney cancer, promotes the progression of kidney cancer. Aging. 2019;11(24):12165–12176. doi: 10.18632/aging.102555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zhou C, Pan R, Li B, Huang T, Zhao J, Ying J, et al. GPX3 hypermethylation in gastric cancer and its prognostic value in patients aged over 60. Future Oncol (London, England) 2019;15(11):1279–1289. doi: 10.2217/fon-2018-0674. [DOI] [PubMed] [Google Scholar]
- 11.Bosello Travain V, Miotto G, Vučković AM, Cozza G, Roveri A, Toppo S, et al. Lack of glutathione peroxidase-8 in the ER impacts on lipid composition of HeLa cells microsomal membranes. Free Radical Biol Med. 2020;147:80–89. doi: 10.1016/j.freeradbiomed.2019.12.010. [DOI] [PubMed] [Google Scholar]
- 12.Wang L, Li C, Tian J, Liu J, Zhao Y, Yi Y, et al. Genome-wide transcriptional analysis of Aristolochia manshuriensis induced gastric carcinoma. Pharm Biol. 2020;58(1):98–106. doi: 10.1080/13880209.2019.1710219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zhang X, Zhan D, Li Y, Wang H, Cheng C, Yao Y, et al. Glutathione Peroxidase 8 as a prognostic biomarker of gastric cancer: an analysis of The Cancer Genome Atlas (TCGA) Data. Med Sci Monit. 2020;26:e921775. doi: 10.12659/MSM.921775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Mazzio EA, Soliman KFA. Whole-transcriptomic profile of SK-MEL-3 melanoma cells treated with the histone deacetylase inhibitor: trichostatin A. Cancer Genomics Proteom. 2018;15(5):349–364. doi: 10.21873/cgp.20094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Han B, Bhowmick N, Qu Y, Chung S, Giuliano AE, Cui X. FOXC1: an emerging marker and therapeutic target for cancer. Oncogene. 2017;36(28):3957–3963. doi: 10.1038/onc.2017.48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Duchartre Y, Kim YM, Kahn M. The Wnt signaling pathway in cancer. Crit Rev Oncol Hematol. 2016;99:141–149. doi: 10.1016/j.critrevonc.2015.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nusse R, Clevers H. Wnt/β-catenin signaling, disease, and emerging therapeutic modalities. Cell. 2017;169(6):985–999. doi: 10.1016/j.cell.2017.05.016. [DOI] [PubMed] [Google Scholar]
- 18.Dunn KJ, Brady M, Ochsenbauer-Jambor C, Snyder S, Incao A, Pavan WJ. WNT1 and WNT3a promote expansion of melanocytes through distinct modes of action. Pigment Cell Res. 2005;18(3):167–180. doi: 10.1111/j.1600-0749.2005.00226.x. [DOI] [PubMed] [Google Scholar]
- 19.Kaur N, Chettiar S, Rathod S, Rath P, Muzumdar D, Shaikh ML, et al. Wnt3a mediated activation of Wnt/β-catenin signaling promotes tumor progression in glioblastoma. Mol Cell Neurosci. 2013;54:44–57. doi: 10.1016/j.mcn.2013.01.001. [DOI] [PubMed] [Google Scholar]
- 20.Badimon L, Casaní L, Camino-Lopez S, Juan-Babot O, Borrell-Pages M. GSK3β inhibition and canonical Wnt signaling in mice hearts after myocardial ischemic damage. PLoS ONE. 2019;14(6):e0218098. doi: 10.1371/journal.pone.0218098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.MacDonald BT, Tamai K, He X. Wnt/beta-catenin signaling: components, mechanisms, and diseases. Dev Cell. 2009;17(1):9–26. doi: 10.1016/j.devcel.2009.06.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Xu Y, Shao QS, Yao HB, Jin Y, Ma YY, Jia LH. Overexpression of FOXC1 correlates with poor prognosis in gastric cancer patients. Histopathology. 2014;64(7):963–970. doi: 10.1111/his.12347. [DOI] [PubMed] [Google Scholar]
- 23.Subramani R, Camacho FA, Levin CI, Flores K, Clift A, Galvez A, et al. FOXC1 plays a crucial role in the growth of pancreatic cancer. Oncogenesis. 2018;7(7):52. doi: 10.1038/s41389-018-0061-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Shang YK, Li C, Liu ZK, Kong LM, Wei D, Xu J, et al. System analysis of the regulation of the immune response by CD147 and FOXC1 in cancer cell lines. Oncotarget. 2018;9(16):12918–12931. doi: 10.18632/oncotarget.24161. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1: Table S1. The information of predicted transcription factors of GPX8.
Additional file 2: Figure S1. The images of basal cell carcinoma signaling pathway from the KEGG database.
Data Availability Statement
All data generated or analysed during this study are included in this published article and its additional information files.