

Abstract
Purpose
A multitude of animal studies substantiates the beneficial effects of Ang-(1–7), a peptide hormone in the protective axis of the renin angiotensin system, in diabetes and its associated complications including diabetic retinopathy (DR). However, the clinical application of Ang-(1–7) is limited due to unfavorable pharmacological properties. As emerging evidence implicates gut dysbiosis in pathogenesis of diabetes and supports beneficial effects of probiotics, we sought to develop probiotics-based expression and delivery system to enhance Ang-(1–7) and evaluate the efficacy of engineered probiotics expressing Ang-(1–7) in attenuation of DR in animal models.
Methods
Ang-(1–7) was expressed in the Lactobacillus species as a secreted fusion protein with a trans-epithelial carrier to allow uptake into circulation. To evaluate the effects of Ang-(1–7) expressed from Lactobacillus paracasei (LP), adult diabetic eNOS−/− and Akita mice were orally gavaged with either 1 × 109 CFU of LP secreting Ang-(1–7) (LP-A), LP alone or vehicle, 3 times/week, for 8 and 12 weeks, respectively.
Results
Ang-(1–7) is efficiently expressed from different Lactobacillus species and secreted into circulation in mice fed with LP-A. Oral administration of LP-A significantly reduced diabetes-induced loss of retinal vascular capillaries. LP-A treatment also prevented loss of retinal ganglion cells, and significantly decreased retinal inflammatory cytokine expression in both diabetic eNOS−/− and Akita mice.
Conclusions
These results provide proof-of-concept for feasibility and efficacy of using engineered probiotic species as live vector for delivery of Ang-(1–7) with enhanced bioavailability.
Translational Relevance
Probiotics-based delivery of Ang-(1–7) may hold important therapeutic potential for the treatment of DR and other diabetic complications.
Keywords: renin angiotensin system, angiotensin-(1–7), diabetes, diabetic complications, diabetic retinopathy, probiotics, Lactobacillus; Lactobacillus paracasei
Introduction
Diabetic retinopathy (DR) is a major microvascular complication of diabetes mellitus and the leading cause of blindness in the working-age population.1,2 A large body of experimental and clinical evidence has demonstrated that dysregulation of the renin angiotensin system (RAS), resulting in elevated concentrations of Angiotensin II (Ang II) systemically and locally at tissue level, contributes to increased oxidative stress, inflammation, development of metabolic syndrome, obesity, diabetes, and its associated complications.3−7 Angiotensin-(1–7) (Ang-(1–7)) is a peptide hormone in the protective axis of the RAS, generated through cleavage of Ang II by the angiotensin converting enzyme 2 (ACE2).8,9 Ang-(1–7) binds to the G protein coupled receptor Mas and activate signaling pathways that counteract the deleterious effects of Ang II.10−12 Ang-(1–7) has been shown beneficial in improving metabolic dysfunction and diabetic complications by inhibiting apoptosis, oxidative stress, and inflammation,12−17 and is also protective in diabetic retina in rodents.18 However, clinical application of Ang-(1–7) is limited due to its extreme short half-life and rapid clearance in circulation and tissues.19,20
As emerging evidence indicates that impaired gut microbiota contributes to the pathogenesis of both type 1 and type 2 diabetes21−27 and probiotic supplements are beneficial in managing diabetes and other metabolic diseases,28−34 we sought to develop an expression and delivery system based on the use of recombinant probiotic species of Lactobacillus bacteria to serve as a live vector for the oral delivery of Ang-(1–7) and evaluate the efficacy of recombinant probiotics expressing Ang-(1–7) in improving diabetes-induced retinal damage in animal models of diabetes. Lactobacillus bacteria are components of the normal gut microbiota35 and are also commonly used in production of fermented food and beverages in the food industry. Many species of Lactobacillus bacteria are also used as probiotic supplements with beneficial effects in humans.36−38 As ingested bacteria can survive both gastric acid and bile to reach the small intestine and colon, where they exert their effects, making them ideal vehicle for delivery of protein and peptide drugs. We generated an expression system based on the use of recombinant Lactobacillus paracasei (LP) to serve as a live vector for the oral delivery of the Ang-(1–7) using the strategy that we have reported previously.39 The Ang-(1–7) peptide is expressed as a secreted fusion protein with the chorea toxin binding protein subunit B (CTB), which facilitates the transmucosal transport into circulation and tissue uptake by GM1 receptor mediated endocytosis. We show that oral administration of LP expressing Ang-(1–7) significantly attenuated diabetes-induced loss of retinal capillaries and retinal ganglion cells (RGCs), and reduced the inflammatory cytokine expression in diabetic animals. These results provide proof-of concept for the feasibility of using recombinant probiotic species as live vector for delivery of Ang-(1–7) and this approach may have important therapeutic potential for treating metabolic diseases and diabetic complications.
Results
Vector Construction to Express Secreted Ang-(1–7) in Lactobacillus Species and In Vivo Characterization in Mice
As reported previously,39 the backbone Lactobacillus shuttle plasmid containing a GFP reporter gene driven by the lactate dehydrogenase (LDH) promoter from Lactobacillus acidophilus was from Addgene (Plasmid #27167).40 The original GFP reporter gene was replaced by a synthetic gene construct in which the Ang-(1–7) peptide is expressed as a secreted fusion protein with the CTB (Fig. 1A), which facilitates transepithelial transport into circulation and tissue uptake.39 The CTB is separated by a furin cleavage site to release Ang-(1–7) once it is secreted.
Figure 1.
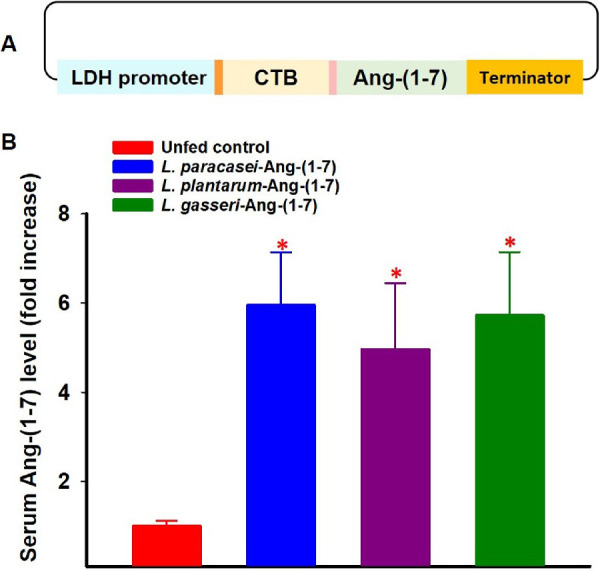
Schematics of Lactobacillus vector expressing secreted Ang-(1–7) (A) and serum levels of Ang-(1–7) in mice fed with different species of Lactobacillus expressing Ang-(1–7) (B). The Ang-(1–7) expression is under the control of the LDH promoter and expressed as a secreted fusion protein to the non-toxic subunit of cholera toxin B (CTB), separated by a furin cleavage site. Ang-(1–7) is efficiently expressed and secreted into circulation in mice fed with these bacteria. N = 4. *P < 0.001 (versus unfed control). Error bars represent standard deviation.
The expression of fusion protein (CTB-Ang-1–7) in Lactobacillus strains was confirmed by Western blotting (data not shown). The ability of the Lactobacillus-expressed proteins to enter the circulation and uptake by different tissues following oral administration in mice was evaluated by ELISA. Six week old C57Bl/6J mice were orally fed with Lactobacillus expressing Ang-(1–7) at 1 × 1010 cfu/mouse daily for 3 days. Mice were then euthanized, and serum and tissue samples were collected 6 hours after the last oral gavage. Ang-(1–7) levels were determined by a commercial enzyme immunoassay (EIA) kit (Peninsula Laboratories International, Inc., San Carlos, CA). As shown in Figure 1B, serum levels of Ang-(1–7) in mice fed with these bacteria were approximately five-fold higher than unred control animals, suggesting that Ang-(1–7) is efficiently expressed from three probiotic species and secreted into circulation.
Oral Administration of Recombinant L. paracasei Expressing Ang-(1–7) did not Affect Body Weight
Oral feeding of L. paracasei expressing Ang-(1–7) (LP-A) did not have any effects on blood glucose levels (data not shown). Non-diabetic animals gained approximately 20% body weight during the period of the study. The body weight was significantly reduced in all diabetic animals. Treatment with LP or LP-A did not affect the body weight (Fig. 2).
Figure 2.

Body weight of non-diabetes mellitus (NDM) and diabetes mellitus (DM) eNOS−/− (A) and Akita (B) mice treated with vehicle (PBS), wild-type Lactobacillus paracasei (LP) and recombinant LP expressing Ang-(1–7) (LP-A) at week 0 (wk 0) and the end of the experiment. Error bars represent standard deviation. *P < 0.05 (DM versus NDM, N = 12 for eNOS−/− mice and Akita versus wild-type littermate control, N = 8).
Oral Administration of Recombinant L. paracasei Expressing Ang-(1–7) (LP-A) Prevents Diabetes-Induced Retinal Capillary Loss in Mice
The protective effect of Ang-(1–7) in DR has been demonstrated previously using AAV vector mediated gene delivery.18 To evaluate the efficacy of Ang-(1–7) expressed from LP-A in diabetic retinopathy, we used two mouse models of diabetes: the streptozotocin (STZ)-induced diabetes in eNOS−/− mice, which develop an accelerated time course and increased severity of retinopathy;18,39,41 and the Akita mice, which carry a mutation in the insulin 2 gene resulting in mice exhibiting reduced ß cell mass and reduced insulin secretion.42 The Akita mice develop progressive retinal abnormalities, including increased vascular permeability, apoptosis, and inner retinal thinning as early as 12 weeks after the onset of hyperglycemia.43 Mice were gavaged 3 times/week with either 1 × 109 CFU of LP-A, wild-type bacteria (LP), or vehicle (PBS) for 8 weeks after STZ-induced diabetes in eNOS−/− mice. The Akita mice were treated with the same dose for 12 weeks beginning at 6 weeks of age.
Ang-(1–7) level in serum and retinal samples collected at the end points was measured by EIA using a commercial kit. Both serum and retinal Ang-(1–7) levels were significantly reduced in diabetic eNOS−/− and Akita mice compared with non-diabetic controls and were restored to normal levels by LP-A treatment (Fig. 3).
Figure 3.

Ang-(1–7) levels in serum (A) and retina (B) samples from eNOS−/− and Akita mice fed with vehicle (PBS), wild-type L. paracasei (LP), and L. paracasei expressing Ang-(1–7) (LP-A) measured by EIA (n = 5 per group). Error bars represent standard deviation. NDM: non-diabetes mellitus. *: P < 0.01 (versus nondiabetic controls). # P < 0.01 (versus vehicle treated diabetic animals). There is not significant difference in Ang-(1–7) levels between LP and vehicle treated groups (ns: not significant; P > 0.1) in both serum and retina of eNOS−/− and Akita mice.
Diabetes resulted in increased capillary loss in eNOS−/− (Fig. 4A) as reported previously.18,39,41 Akita mice also showed increased retinal acellular capillaries compared with age-matched littermate controls (Fig. 4C). LP-A treatment significantly reduced the number of acellular capillaries in both diabetic eNOS−/− (approximately 29% reduction) and Akita (approximately 40% reduction) mice compared with untreated diabetic animals (Fig. 4). Wild type LP alone also showed small but insignificant reduction of capillary loss in both diabetic eNOS−/− and Akita mice.
Figure 4.

Evaluation of retinal acellular capillary in diabetic eNOS−/− (A, B) and Akita (C, D) mice. A and C Representative images of trypsin-digested retinal vascular preparations from eNOS−/− (A) and Akita (C) mice treated with vehicle (PBS), WT-LP, or LP-A; and quantitative measurements of acellular capillaries of eNOS−/− (B) and Akita (D) mice. Arrows indicate the acellular capillaries. Error bars represent standard deviation. NDM, non-diabetes mellitus; DM, diabetes mellitus. *: P < 0.05; **P < 0.01. ns: not significant (P > 0.1). N = 6/group. Treatments with LP-A significantly reduced acellular capillaries in both eNOS−/− and Akita mice.
Oral Administration of L. paracasei Ang-(1–7) Reduce Diabetes-Induced Retinal Ganglion Cell Loss
In additional to microvascular change, diabetic retina showed considerable loss of retinal ganglion cells (RGCs), as detected by Brn3a immunostaining, a specific marker for RGCs,44 in both diabetic eNOS−/− and Akita mice (Fig. 5), as reported previously.39 RGCs loss also occurs in human patients with diabetes as well as other diabetic rodents.45−50 LP-A treatment, but not wild-type LP, prevented RGCs loss in both diabetic eNOS−/− and Akita mice (Fig. 5).
Figure 5.

Quantification of retinal ganglion cell (RGC) density detected by Brn3a immunostaining in diabetic eNOS−/− (A, B) and Akita (C, D) mice. Representative immunofluorescence images of Brn3a staining from eNOS−/− (A) and Akita (C) mice treated with vehicle (PBS), WT-LP, or LP-A and quantification of Brn3a positive cells of eNOS−/− (B) and Akita (D) mice. Error bars represent standard deviation. NDM, nondiabetes mellitus; DM, diabetic mellitus. **: P < 0.01. ns: not significant (P > 0.1). N = 6/group.
Oral Administration of L. paracasei- Ang-(1–7) Reduces Diabetes-Induced Expression of Inflammatory Cytokines in Diabetic Retina in Mice
Diabetes is associated with increased inflammatory responses in the retina. The expression level of pro-inflammatory cytokines and chemokines was evaluated by real-time RT-polymerase chain reaction (PCR) in the retina from each experimental group. LP-A treated animals show significantly decreased retinal expression of all these cytokines and chemokines in both diabetic eNOS−/− mice (Fig. 6A) and Akita mice (Fig. 6B). LP alone did not show any effect on the expression of these genes in diabetic eNOS−/− mice (Fig. 6A), however, the expression levels of MCP-1 and VEGF were slightly but significantly reduced in Akita mice (Fig. 6B). There was also a slight decrease in ICAM-1 expression in LP treated Akita mice, but not statistically significant. The expression of Iba-1, a marker of microglia, was also elevated in diabetic retina of both eNOS−/− and Akita mice, and was significantly reduced by LP-A treatment but not affected by LP alone (Fig. 6). Elevated expression of Iba-1 in diabetic retina was also detected by immunofluorescence. Diabetic eNOS−/− mouse retina showed increased number of Iba-1 positive microglial cells, which was significantly reduced in LP-A treated mice (Fig. 7).
Figure 6.

Retinal inflammatory cytokine expression measured by real-time RT-PCR in diabetic eNOS−/− (A) and Akita mice (B) treated with vehicle (PBS), WT-LP, or LP-A. Values on y-axis represent relative expression level compared to PBS treated group for each gene. Error bars represent standard deviation. NDM, non-diabetes mellitus. *: P < 0.05; **: P < 0.01 (versus PBS treated diabetic groups). #: P < 0.01 (versus PBS treated diabetic groups). There is no significant difference between vehicle and WT-LP treated groups. N = 4/group.
Figure 7.

Quantification of microglial cells by Iba-1 immunofluorescence. (A) Representative immunofluorescence images of Iba-1 staining from eNOS−/− mice treated with vehicle (PBS), LP, or LP-A. (B) quantification of Iba-1 positive cells. Error bars represent standard deviation. NDM: non-diabetes mellitus; DM: diabetes mellitus. *: P < 0.05; **: P <0.01. ns: not significant (P > 0.1). N = 4/group.
Materials and Methods
Bacterial Strains and Growth Conditions
The Lactobacillus strains used in this study were from American Type Culture Collection (ATCC, Manassas, VA): L. paracasei (LP) (ATCC 27092), L. gasseri (ATCC 4963), and L. plantarum (ATCC 8014) and were cultured in de Man, Rogosa, and Sharpe (MRS) broth (Thermo Fisher Scientific, #DF0881-17-5) at 37°C for 18 hours without shaking. The plasmid pTRKH3-ldhGFP (Addgene, plasmid #27170) was used as a backbone for cloning of secreted Ang-(1–7) in fusion with the CTB, which allows for uptake of the protein into the enterocytes via its binding to the GM1 receptor. A mutant form of CTB, which retains the binding to GM1-ganglioside for cellular uptake but lacks immunomodulatory and toxic activity51,52 was used as reported previously39 to construct the fusion construct. The resulting plasmid was electroporated into different Lactobacillus species by electroporation as described by Welker et al.53 Recombinant Lactobacillus bacteria expressing Ang-(1–7) were grown in the MRS media supplemented with 5 μg/mL erythromycin (Sigma-Aldrich, St. Louis, MO). For oral gavage of mice, bacteria were harvested by centrifugation at 5,000 × g for 20 minutes and resuspended in sterile PBS.
Animals and Experimental Procedures
Wild-type C57Bl/6J (Stock Number: 000664), eNOS−/− (Stock Number: 002684) and Akita mice (Stock Number: 003548) were purchased from Jackson Laboratories (Bar Harbor, ME) and maintained in the Animal Care Service at the University of Florida. All procedures adhered to the ARVO statement for the Use of Animals in Ophthalmic and Vision Research, and the protocol was approved by the Animal Care and Use Committee of the University of Florida. The animals were fed standard laboratory chow and allowed free access to water in an air-conditioned room with a 12 and 12-hour light dark cycle. Diabetes in adult eNOS−/− mice (8–10 weeks old) was induced by STZ injection, as reported previously.41 Gavage of diabetic eNOS−/− mice was performed with either 1 × 1010 CFU of wild-type L. paracasei (WT-LP), LP expressing Ang-(1–7) (LP-A) or vehicle (PBS), 3 times/week for 8 weeks. Akita mice were gavaged with the same dose for 12 weeks. Mice were euthanized 2 days after oral gavage for final tissue collection and analysis.
ELISA Analysis
Serum and retinal level of Ang-(1–7) was measured using a commercial EIA kit (Bachem, San Carlos, CA), following the manufacturer's instructions. All measurements were performed in duplicate and the data represent the mean of two separate assay results.
Retinal Vascular Preparation by Trypsin-Digestion
Retinal vasculature was prepared using trypsin digest, as described previously.41 Briefly, eyes were fixed in 4% paraformaldehyde freshly made in PBS overnight. Retinas were dissected out from the eyecups and digested in 3% trypsin (GIBCO-BRL) for 2–3 hours at 37°C. Retinal vessels were separated from other retinal neuronal cells by gentle shaking and manipulation under a dissection microscope. The vessels were then mounted on a clean slide, allowed to dry, and stained with Periodic Acid Solution hematoxylin and eosin, Gill No. 3 (PAS-H&E; , Sigma, St. Louis, MO) according to the instruction manual. After staining and washing in water, the tissue was dehydrated and mounted using Permount mounting media (Sigma).
Immunofluorescence and Immunocytochemistry
For immunofluorescence studies, eyes were fixed in 4% paraformaldehyde overnight at 4°C and subsequently processed for either quick freezing in optical cutting temperature (OCT) compound or paraffin embedding. For OCT embedding, the eyes were cryoprotected in 30% sucrose/PBS for several hours or overnight before quick freezing in OCT compound, then 12-μm-thick sections were cut at −20 to –22 degrees. For paraffin embedded eyes, 4 μm-thick paraffin sections were cut and mounted on Superfrost Plus slides. The paraffin sections were first deparaffinized followed by antigen retrieval in low pH citric acid buffer for 20 minutes. The sections were then incubated in blocking solution (5% BSA + 0.3% Triton X-100 in PBS) for 1 hour. This was followed by incubation overnight at 4°C with primary antibody (mouse anti-Brn3a, 1:200, MAB1585; Millipore, Billerica, MA). Iba-1 (1:500, 019-19741; Wako, Richmond, VA) immunostaining was performed using OCT-embedded frozen sections without antigen retrieval. After washing, secondary antibody conjugated to Alexa 488 (Molecular Probes/Invitrogen, Carlsbad, CA) was incubated for 1 hour at room temperature (RT). Sections were washed in PBS containing the nuclear counterstain DAPI (4′,6 diamidino-2-phenylindole), and mounted in Dako mounting media. The images were captured with a Leica Fluorescence Microscope LAS X System (Leica Microsystems Inc., Buffalo Grove, IL).
Real Time RT-PCR analysis
Total RNA was isolated from freshly enucleated eyes using Trizol Reagent (Invitrogen, Carlsbad, CA) according to manufacturer's instructions. Reverse transcription was performed using Enhanced Avian HS RT-PCR kit (Sigma-Aldrich, Inc., St. Louis, MO) following manufacturer's instructions. Real time PCR was carried out on real time thermal cycler (iCycler; Bio-Rad Life Sciences, Hercules, CA) using iQTM Sybr Green Supermix (Bio-Rad Life Sciences). The threshold cycle number (Ct) for real-time PCR was set by the cycler software. Optimal primer concentration for PCR was determined separately for each primer pair. Each reaction was run in duplicate or in triplicate, and reaction tubes with target primers and those with Actin primers were always included in the same PCR run. Primer sequences used in this study are shown in the Table. All the reactions were repeated at least twice.
Table.
Primers Used for Real-Time RT-PCR Analysis
| Gene Name | Accession Number | Sequences |
|---|---|---|
| IL-1β | NM_008361.3 | Forward: 5'-AAAGCCTCGTGCTGTCGGACC -3' |
| Reverse: 5'-CAGCTGCAGGGTGGGTGTGC -3' | ||
| TNF-α | NM_013693.2 | Forward: 5'-AGGCGCCACATCTCCCTCCA-3' |
| Reverse: 5'-CGGTGTGGGTGAGGAGCACG-3' | ||
| ICAM-1 | NM_010493 | Forward: 5'-AGATGACCTGCAGACGGAAG-3' |
| Reverse: 5'-GGCTGAGGGTAAATGCTGTC-3' | ||
| MCP-1 | NM_011333 | Forward: 5'-CCCCACTCACCTGCTGCTACT-3' |
| Reverse: 5'-GGCATCACAGTCCGAGTCACA-3' | ||
| β-Actin | X03672 | Forward: 5'-AGCAGATGTGGATCAGCAAG-3' |
| Reverse: 5'-ACAGAAGCAATGCTGTCACC-3' | ||
| Iba-1 | XM_006523503.4 | Forward: 5'-TCCCCCAGCCAAGAAAGCTA- 3' |
| Reverse: 5' -TGACCCACTAGGAGCGTCAT- 3' | ||
| VEGF | NM_001025250.3 | Forward: 5'-TTCAGCTCGCTCCTCCACTT- 3' |
| Reverse: 5'TTTTCTCTGCCTCCGTGAGG- 3' |
Statistical Analysis
Data are expressed as the mean + SD of at least two independent experiments. Differences between mean values of multiple groups were analyzed by 1-way ANOVA with Dunnett's test for post hoc comparisons. A P value < 0.05 was considered statistically significant.
Discussion
Dysregulation of RAS, resulting in elevated Ang II, contributes diabetes and diabetic complications, including DR. The protective axis of RAS, involving ACE2/Ang-(1–7)/Mas, opposes these effects by degradation of Ang II to generate Ang-(1–7), which binds to a G-protein coupled receptor, Mas, and activates signaling pathways that counteract the effects of Ang II.10−12 We have previously demonstrated that increased expression of ACE2 or Ang-(1–7) diminishes diabetes-induced retinal pathophysiology18,54 and ocular inflammation55−57; providing the “proof-of-principle” that enhancing the ACE2/Ang-(1–7) axis is a promising approach for treating DR. Moreover, a large number of studies have shown that enhanced expression of Ang-(1–7) and ACE2 reduces inflammation58−61 and oxidative damage,62−65 increases glucose uptake,63 improves lipid and glucose metabolism,66,67 ameliorates insulin resistance and dyslipidemia,14,67−69 improves pancreatic β-cell function,70−72 improves the reparative function of dysfunctional endothelial cells and progenitors,73−75 and confers protection against a variety of pathological conditions including diabetes-induced nephropathy76−83 and cardiovascular dysfunction.83−88 Despite the mounting evidence for beneficial effects of Ang-(1–7), its clinical studies and applications are limited, largely due to extreme short half-life and rapid clearance in circulation and tissues,19,20 making it challenging to develop a pharmaceutical composition of Ang-(1–7) that delivers the peptide to target tissues with sufficient bioavailability.
In this study, we designed an expression and delivery system based on the use of Lactobacillus bacteria to serve as a live vector for the oral delivery of Ang-(1–7) peptide. Using the similar strategy as reported previously,39 the expression of Ang-(1–7) is driven by the LDH promoter from Lactobacillus acidophilus, a strong promoter that is active in different bacterial hosts.40,89 The peptide is expressed as a secreted fusion protein with the CTB, which facilitates the transmucosal transport into circulation and tissue uptake by monosialotetrahexosylganglioside (GM1) mediated endocytosis. The Usp45 secretion signal peptide, CTB, and furin cleavage site enable Ang-(1–7) to be secreted into the gut lumen and its transepithelial transport into circulation as described previously.39 We show that the Ang-(1–7) peptide is efficiently expressed from three different Lactobacillus species and secreted into circulation in mice fed with these bacteria. Treatment with L. paracasei expressing Ang-(1–7) reduced diabetes-induced increase in retinal acellular capillaries, prevented RGC loss and decreased inflammatory cytokine expression in the retina in both diabetic eNOS−/− and Akita mice.
We sought to develop probiotics-based expression and delivery system to enhance Ang-(1–7) based on the following rationale. First, diabetes inflicts multiple tissues including the retina and Ang-(1–7) has been shown to be beneficial in many tissues. Second, increasing evidence implicates the gut and its microbiota in pathogenesis of diabetes and its associated complications,21−24,26,90−104 including DR,105−107 and probiotics are beneficial in management of diabetes.108−112 The probiotic-based delivery of Ang-(1–7) offers many advantages. First, probiotics have been consumed by humans for thousands of years, are generally recognized as safe (GRAS) to consume and offer many beneficial effects on their own.113 Probiotics are known to promote host health by modulating immune system, interfere with potential pathogens, improving intestinal barrier function, positively modulating the composition and function of the commensal microbiota, as well as many other protective actions.114−121 Second, ingested probiotics can survive both gastric acid and bile to reach the small intestine and colon, where they exert their effects, making them an ideal vehicle for delivery of protein drugs. Third, unlike conventional approaches, the probiotics-based oral delivery system, using a carrier fused with the therapeutic protein, facilitates efficient transmucosal transport into the circulation, increases half-life and target tissue uptake, thus enhancing bioavailability.39,122,123 Moreover, probiotic bacteria are inexpensive to produce and oral delivery of therapeutics is patient-friendly, thus probiotics-based approach is more cost-effective. Thus, a probiotic enriched with Ang-(1–7) represent potential therapy to improve metabolism, and intestinal and immune functions, thereby preventing DR and other diabetic complications. Our results provide proof-of concept for feasibility using probiotics-based oral delivery of Ang-(1–7) with enhanced bioavailability and efficacy in two mouse models of DR.
In current study, we have used LP expressing Ang-(1–7) to test the efficacy in animal models of DR. The rationale for choosing LP is that, first, it is part of the healthy gut microbiota124 and is widely used as a probiotic125−127 and, second, LP has been shown to be beneficial in colitis-induced gut inflammation and barrier dysfunction,128−130 as well as improving diabetes in experimental models.131−134 LP-A treatment protected development of DR in both models. However, wild-type LP alone did not show protective effects, as reported previously,39 this is likely due to the strain used in this study. A number of experimental and clinical studies has demonstrated that probiotics supplements of several Lactobacillus species (including LP) are beneficial in management of both type 1 and type 2 diabetes and other metabolic diseases; however, these various beneficial effects are strain and species-specific.109−112 For example, Dang et al. evaluated eight strains of LP, only one of those showed antidiabetic effects.132 It would be interesting to determine whether Ang-(1–7) expressed from this strain would provide better protection against DR. Moreover, our results showed that Ang-(1–7) can be expressed from several species of Lactobacillus, each of which may offer different beneficial effects, future studies will be required to determine whether specific probiotics species or strains can be selected for oral delivery to optimally target specific patient populations to achieve personalized precision medicine paradigm. It is interesting to note that although LP alone did not provide significant protection against diabetes-induced loss of retinal capillaries and RGCs, but it did slightly reduce the expression of some inflammatory cytokines (e.g. MCP-1 and VEGF), but not other cytokines measured in this study. The reduction of these cytokines was only detected in Akita mice but not in diabetic eNOS−/−. This likely due to the fact that diabetic eNOS−/− mice not only develop more severe retinopathy, but also other tissue damage and impaired immune functions compared with Akita mice.
Ang-(1–7) has been shown to be beneficial under various pathologic conditions in various tissues, including diabetes and diabetic complications, by improving metabolism and insulin sensitivity, inhibiting apoptosis, oxidative stress, and inflammation.12−17 Here, we showed that Ang-(1–7) is increased in both circulation and retina in mice fed with LP-A, suggesting that the protective effects of LP-A in diabetic retina may be mediated by both local effects of Ang-(1–7) in the retina as well as general beneficial effects in other tissues of diabetic animals. It is also possible that LP-A may provide protective effects by modulating gut microbiome. A previous study showed that oral administration of LP-A in rats increased abundance of gut Akkermansia muciniphila (A. muciniphila).123 As decreased abundance of A. muciniphila has been associated with increased prevalence of metabolic disorders, such as obesity and type 2 diabetes,135−137 such increase in A. muciniphila by LP-A treatment, which remains to be confirmed in diabetic mice by ongoing study, may provide additional beneficial effects in diabetic animals. More comprehensive studies will be required to further understand the effects and underlying mechanisms of LP-A treatment in various tissues.
In conclusion, Ang-(1–7) is efficiently expressed from different probiotic species and secreted into circulation in mice fed with these bacteria. Oral administration of L. paracasei secreting Ang-(1–7) provides protection against diabetes-induced DR. Thus, LP-based delivery of Ang-(1–7) may hold important therapeutic potential for the treatment of DR and other complications.
Acknowledgments
Supported in part by NIH grants EY021752 and EY024564, American Diabetes Association, and BrightFocus Foundation (Q.L.); and HL102033 (M.K.R.). Core facilities were supported by NEI grant P30 EY02172 and Research to Prevent Blindness to University of Florida.
Disclosure: A. Verma, None; P. Zhu, None; K. Xu, None; T. Du, None; S. Liao, None; Z. Liang, None; M.K. Raizada, None; Q. Li, None
References
- 1. Yau JW, Rogers SL, Kawasaki R, et al.. Global prevalence and major risk factors of diabetic retinopathy. Diabetes Care. 2012; 35: 556–564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Pascolini D, Mariotti SP. Global estimates of visual impairment: 2010. Br J Ophthalmol. 2012; 96: 614–618. [DOI] [PubMed] [Google Scholar]
- 3. Goossens GH. The renin-angiotensin system in the pathophysiology of type 2 diabetes. Obes Facts. 2012; 5: 611–624. [DOI] [PubMed] [Google Scholar]
- 4. Dikalov SI, Nazarewicz RR. Angiotensin II-induced production of mitochondrial reactive oxygen species: potential mechanisms and relevance for cardiovascular disease. Antioxid Redox Signal. 2013; 19: 1085–1094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Das UN. Renin-angiotensin-aldosterone system in insulin resistance and metabolic syndrome. J Transl Int Med. 2016; 4: 66–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Cabandugama PK, Gardner MJ, Sowers JR. The renin angiotensin aldosterone system in obesity and hypertension: roles in the cardiorenal metabolic syndrome. Med Clin North Am. 2017; 101: 129–137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Rein J, Bader M. Renin-angiotensin system in diabetes. Protein Pept Lett. 2017; 24: 833–840. [DOI] [PubMed] [Google Scholar]
- 8. Crackower MA, Sarao R, Oudit GY, et al.. Angiotensin-converting enzyme 2 is an essential regulator of heart function. Nature. 2002; 417: 822–828. [DOI] [PubMed] [Google Scholar]
- 9. Donoghue M, Hsieh F, Baronas E, et al.. A novel angiotensin-converting enzyme-related carboxypeptidase (ACE2) converts angiotensin I to angiotensin 1-9. Circ Res. 2000; 87: E1–E9. [DOI] [PubMed] [Google Scholar]
- 10. Santos RA, Simoes e Silva AC, Maric C, et al.. Angiotensin-(1–7) is an endogenous ligand for the G protein-coupled receptor Mas. Proc Natl Acad Sci USA. 2003; 100: 8258–8263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Karnik SS, Singh KD, Tirupula K, Unal H. Significance of angiotensin 1–7 coupling with MAS1 receptor and other GPCRs to the renin-angiotensin system: IUPHAR Review 22. Br J Pharmacol. 2017; 174: 737–753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Santos RAS, Sampaio WO, Alzamora AC, et al.. The ACE2/angiotensin-(1–7)/MAS axis of the renin-angiotensin system: focus on angiotensin-(1–7). Physiol Rev. 2018; 98: 505–553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Simoes e Silva AC, Silveira KD, Ferreira AJ, Teixeira MM. ACE2, angiotensin-(1–7) and Mas receptor axis in inflammation and fibrosis. Br J Pharmacol. 2013; 169: 477–492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Passos-Silva DG, Verano-Braga T, Santos RA. Angiotensin-(1–7): beyond the cardio-renal actions. Clin Sci (Lond). 2013; 124: 443–456. [DOI] [PubMed] [Google Scholar]
- 15. Santos SH, Andrade JM. Angiotensin 1–7: a peptide for preventing and treating metabolic syndrome. Peptides. 2014; 59: 34–41. [DOI] [PubMed] [Google Scholar]
- 16. Patel VB, Basu R, Oudit GY. ACE2/Ang 1–7 axis: a critical regulator of epicardial adipose tissue inflammation and cardiac dysfunction in obesity. Adipocyte. 2016; 5: 306–311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Lelis DF, Freitas DF, Machado AS, Crespo TS, Santos SHS. Angiotensin-(1–7), adipokines and inflammation: inflammation and adipokines. Metabolism. 2019; 95: 36–45. [DOI] [PubMed] [Google Scholar]
- 18. Verma A, Shan Z, Lei B, et al.. ACE2 and Ang-(1–7) confer protection against development of diabetic retinopathy. Mol Ther. 2012; 20: 28–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Mordwinkin NM, Russell JR, Burke AS, Dizerega GS, Louie SG, Rodgers KE. Toxicological and toxicokinetic analysis of angiotensin (1–7) in two species. J Pharm Sci. 2012; 101: 373–380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Yamada K, Iyer SN, Chappell MC, Ganten D, Ferrario CM. Converting enzyme determines plasma clearance of angiotensin-(1–7). Hypertension. 1998; 32: 496–502. [DOI] [PubMed] [Google Scholar]
- 21. Blandino G, Inturri R, Lazzara F, Di Rosa M, Malaguarnera L. Impact of gut microbiota on diabetes mellitus. Diabetes Metab. 2016; 42: 303–315. [DOI] [PubMed] [Google Scholar]
- 22. Hartstra AV, Bouter KE, Backhed F, Nieuwdorp M. Insights into the role of the microbiome in obesity and type 2 diabetes. Diabetes Care. 2015; 38: 159–165. [DOI] [PubMed] [Google Scholar]
- 23. Needell JC, Zipris D. The role of the intestinal microbiome in type 1 diabetes pathogenesis. Curr Diab Rep. 2016; 16(10): 89. [DOI] [PubMed] [Google Scholar]
- 24. Abdellatif AM, Sarvetnick NE. Current understanding of the role of gut dysbiosis in type 1 diabetes. J Diabetes. 2019; 11(8): 9632–644. [DOI] [PubMed] [Google Scholar]
- 25. Chen X, Devaraj S. Gut microbiome in obesity, metabolic syndrome, and diabetes. Curr Diab Rep. 2018; 18: 129. [DOI] [PubMed] [Google Scholar]
- 26. Zheng P, Li Z, Zhou Z. Gut microbiome in type 1 diabetes: a comprehensive review. Diabetes Metab Res Rev. 2018; 34: e3043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Sharma S, Tripathi P. Gut microbiome and type 2 diabetes: where we are and where to go? J Nutr Biochem. 2019; 63: 101–108. [DOI] [PubMed] [Google Scholar]
- 28. Akbari V, Hendijani F. Effects of probiotic supplementation in patients with type 2 diabetes: systematic review and meta-analysis. Nutr Rev. 2016; 74: 774–784. [DOI] [PubMed] [Google Scholar]
- 29. Samah S, Ramasamy K, Lim SM, Neoh CF. Probiotics for the management of type 2 diabetes mellitus: a systematic review and meta-analysis. Diabetes Res Clin Pract. 2016; 118: 172–182. [DOI] [PubMed] [Google Scholar]
- 30. Saez-Lara MJ, Robles-Sanchez C, Ruiz-Ojeda FJ, Plaza-Diaz J, Gil A. Effects of probiotics and synbiotics on obesity, insulin resistance syndrome, type 2 diabetes and non-alcoholic fatty liver disease: a review of human clinical trials. Int J Mol Sci. 2016; 17(6): 928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Li C, Li X, Han H, et al.. Effect of probiotics on metabolic profiles in type 2 diabetes mellitus: a meta-analysis of randomized, controlled trials. Medicine (Baltimore). 2016; 95: e4088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Sun J, Buys NJ. Glucose- and glycaemic factor-lowering effects of probiotics on diabetes: a meta-analysis of randomised placebo-controlled trials. Br J Nutr. 2016; 115: 1167–1177. [DOI] [PubMed] [Google Scholar]
- 33. Zhang Q, Wu Y, Fei X. Effect of probiotics on glucose metabolism in patients with type 2 diabetes mellitus: a meta-analysis of randomized controlled trials. Medicina (Kaunas). 2016; 52: 28–34. [DOI] [PubMed] [Google Scholar]
- 34. Razmpoosh E, Javadi M, Ejtahed HS, Mirmiran P. Probiotics as beneficial agents in the management of diabetes mellitus: a systematic review. Diabetes Metab Res Rev. 2016; 32: 143–168. [DOI] [PubMed] [Google Scholar]
- 35. Heeney DD, Gareau MG, Marco ML. Intestinal Lactobacillus in health and disease, a driver or just along for the ride? Curr Opin Biotechnol. 2018; 49: 140–147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Lebeer S, Vanderleyden J, De Keersmaecker SC. Genes and molecules of lactobacilli supporting probiotic action. Microbiol Mol Biol Rev. 2008; 72: 728–764, Table of Contents. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Di Cerbo A, Palmieri B, Aponte M, Morales-Medina JC, Iannitti T. Mechanisms and therapeutic effectiveness of lactobacilli. J Clin Pathol. 2016; 69: 187–203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Salvetti E, O'Toole PW. The genomic basis of lactobacilli as health-promoting organisms. Microbiol Spectr, 10.1128/microbiolspec.BAD-0011-2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Verma A, Xu K, Du T, et al.. Expression of human ACE2 in Lactobacillus and beneficial effects in diabetic retinopathy in mice. Mol Ther Methods Clin Dev. 2019; 14: 161–170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Lizier M, Sarra PG, Cauda R, Lucchini F. Comparison of expression vectors in Lactobacillus reuteri strains. FEMS Microbiol Lett. 2010; 308: 8–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Li Q, Verma A, Han PY, et al.. Diabetic eNOS-knockout mice develop accelerated retinopathy. Invest Ophthalmol Vis Sci. 2010; 51: 5240–5246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Wang J, Takeuchi T, Tanaka S, et al.. A mutation in the insulin 2 gene induces diabetes with severe pancreatic beta-cell dysfunction in the Mody mouse. J Clin Invest. 1999; 103: 27–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Barber AJ, Antonetti DA, Kern TS, et al.. The Ins2Akita mouse as a model of early retinal complications in diabetes. Invest Ophthalmol Vis Sci. 2005; 46: 2210–2218. [DOI] [PubMed] [Google Scholar]
- 44. Quina LA, Pak W, Lanier J, et al.. Brn3a-expressing retinal ganglion cells project specifically to thalamocortical and collicular visual pathways. J Neurosci. 2005; 25: 11595–11604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Sohn EH, van Dijk HW, Jiao C, et al.. Retinal neurodegeneration may precede microvascular changes characteristic of diabetic retinopathy in diabetes mellitus. Proc Natl Acad Sci USA. 2016; 113: E2655–E2664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Verma A, Raman R, Vaitheeswaran K, et al.. Does neuronal damage precede vascular damage in subjects with type 2 diabetes mellitus and having no clinical diabetic retinopathy? Ophthalmic Res. 2012; 47: 202–207. [DOI] [PubMed] [Google Scholar]
- 47. van Dijk HW, Verbraak FD, Kok PH, et al.. Decreased retinal ganglion cell layer thickness in patients with type 1 diabetes. Invest Ophthalmol Vis Sci. 2010; 51: 3660–3665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Zeng Y, Cao D, Yu H, et al.. Early retinal neurovascular impairment in patients with diabetes without clinically detectable retinopathy. Br J Ophthalmol. 2019; 103(12): 1747–1752. [DOI] [PubMed] [Google Scholar]
- 49. Kim K, Kim ES, Yu SY. Longitudinal relationship between retinal diabetic neurodegeneration and progression of diabetic retinopathy in patients with type 2 diabetes. Am J Ophthalmol. 2018; 196: 165–172. [DOI] [PubMed] [Google Scholar]
- 50. Prasad T, Roksnoer LC, Zhu P, et al.. Beneficial effects of combined AT1 receptor/neprilysin inhibition (ARNI) versus AT1 receptor blockade alone in the diabetic eye. Invest Ophthalmol Vis Sci. 2016; 57: 6722–6730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Rodighiero C, Fujinaga Y, Hirst TR, Lencer WI. A cholera toxin B-subunit variant that binds ganglioside G(M1) but fails to induce toxicity. J Biol Chem. 2001; 276: 36939–36945. [DOI] [PubMed] [Google Scholar]
- 52. Aman AT, Fraser S, Merritt EA, et al.. A mutant cholera toxin B subunit that binds GM1- ganglioside but lacks immunomodulatory or toxic activity. Proc Natl Acad Sci USA. 2001; 98: 8536–8541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Welker DL, Hughes JE, Steele JL, Broadbent JR. High efficiency electrotransformation of Lactobacillus casei. FEMS Microbiol Lett. 2015; 362: 1–6. [DOI] [PubMed] [Google Scholar]
- 54. Dominguez JM 2nd, Hu P, Caballero S, et al.. Adeno-associated virus overexpression of angiotensin-converting enzyme-2 reverses diabetic retinopathy in type 1 diabetes in mice. Am J Pathol. 2016; 186: 1688–1700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Shil PK, Kwon KC, Zhu P, Verma A, Daniell H, Li Q. Oral delivery of ACE2/Ang-(1–7) bioencapsulated in plant cells protects against experimental uveitis and autoimmune uveoretinitis. Mol Ther. 2014; 22: 2069–2082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Qiu Y, Shil PK, Zhu P, et al.. Angiotensin-converting enzyme 2 (ACE2) activator diminazene aceturate ameliorates endotoxin-induced uveitis in mice. Invest Ophthalmol Vis Sci. 2014; 55: 3809–3818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Qiu Y, Tao L, Zheng S, et al.. AAV8-mediated angiotensin-converting enzyme 2 gene delivery prevents experimental autoimmune uveitis by regulating MAPK, NF-kappaB and STAT3 pathways. Sci Rep. 2016; 6: 31912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. da Silveira KD, Coelho FM, Vieira AT, et al.. Anti-inflammatory effects of the activation of the angiotensin-(1–7) receptor, MAS, in experimental models of arthritis. J Immunol. 2010; 185: 5569–5576. [DOI] [PubMed] [Google Scholar]
- 59. Jiang T, Gao L, Guo J, Lu J, Wang Y, Zhang Y. Suppressing inflammation by inhibiting the NF-kappaB pathway contributes to the neuroprotective effect of angiotensin-(1–7) in rats with permanent cerebral ischaemia. Br J Pharmacol. 2012; 167: 1520–1532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Santos SH, Andrade JM, Fernandes LR, et al.. Oral angiotensin-(1–7) prevented obesity and hepatic inflammation by inhibition of resistin/TLR4/MAPK/NF-kappaB in rats fed with high-fat diet. Peptides. 2013; 46: 47–52. [DOI] [PubMed] [Google Scholar]
- 61. Souza LL, Costa-Neto CM. Angiotensin-(1–7) decreases LPS-induced inflammatory response in macrophages. J Cell Physiol. 2012; 227: 2117–2122. [DOI] [PubMed] [Google Scholar]
- 62. Mordwinkin NM, Meeks CJ, Jadhav SS, et al.. Angiotensin-(1–7) administration reduces oxidative stress in diabetic bone marrow. Endocrinology. 2012; 153: 2189–2197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Liu C, Lv XH, Li HX, et al.. Angiotensin-(1–7) suppresses oxidative stress and improves glucose uptake via Mas receptor in adipocytes. Acta Diabetol. 2012; 49: 291–299. [DOI] [PubMed] [Google Scholar]
- 64. Jiang T, Gao L, Shi J, Lu J, Wang Y, Zhang Y. Angiotensin-(1–7) modulates renin-angiotensin system associated with reducing oxidative stress and attenuating neuronal apoptosis in the brain of hypertensive rats. Pharmacol Res. 2013; 67: 84–93. [DOI] [PubMed] [Google Scholar]
- 65. Raffai G, Durand MJ, Lombard JH. Acute and chronic angiotensin-(1–7) restores vasodilation and reduces oxidative stress in mesenteric arteries of salt-fed rats. Am J Physiol Heart Circ Physiol. 2011; 301: H1341–H1352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Bindom SM, Hans CP, Xia H, Boulares AH, Lazartigues E. Angiotensin I-converting enzyme type 2 (ACE2) gene therapy improves glycemic control in diabetic mice. Diabetes. 2010; 59: 2540–2548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Santos SH, Braga JF, Mario EG, et al.. Improved lipid and glucose metabolism in transgenic rats with increased circulating angiotensin-(1–7). Arterioscler Thromb Vasc Biol. 2010; 30: 953–961. [DOI] [PubMed] [Google Scholar]
- 68. Marcus Y, Shefer G, Sasson K, et al.. Angiotensin 1–7 as means to prevent the metabolic syndrome: lessons from the fructose-fed rat model. Diabetes. 2013; 62: 1121–1130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Giani JF, Mayer MA, Munoz MC, et al.. Chronic infusion of angiotensin-(1–7) improves insulin resistance and hypertension induced by a high-fructose diet in rats. Am J Physiol Endocrinol Metab. 2009; 296: E262–E271. [DOI] [PubMed] [Google Scholar]
- 70. Sahr A, Wolke C, Maczewsky J, Krippeit-, et al.. The angiotensin-(1–7)/Mas axis improves pancreatic beta-cell function in vitro and in vivo. Endocrinology. 2016; 157: 4677–4690. [DOI] [PubMed] [Google Scholar]
- 71. Yu X, Cui L, Hou F, et al.. Angiotensin-converting enzyme 2-angiotensin (1–7)-Mas axis prevents pancreatic acinar cell inflammatory response via inhibition of the p38 mitogen-activated protein kinase/nuclear factor-kappaB pathway. Int J Mol Med. 2018; 41: 409–420. [DOI] [PubMed] [Google Scholar]
- 72. Lu CL, Wang Y, Yuan L, Li Y, Li XY. The angiotensin-converting enzyme 2/angiotensin (1–7)/Mas axis protects the function of pancreatic beta cells by improving the function of islet microvascular endothelial cells. Int J Mol Med. 2014; 34: 1293–1300. [DOI] [PubMed] [Google Scholar]
- 73. Jarajapu YP, Bhatwadekar AD, Caballero S, et al.. Activation of the ACE2/angiotensin-(1–7)/Mas receptor axis enhances the reparative function of dysfunctional diabetic endothelial progenitors. Diabetes. 2013; 62: 1258–1269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Chen J, Xiao X, Chen S, et al.. Angiotensin-converting enzyme 2 priming enhances the function of endothelial progenitor cells and their therapeutic efficacy. Hypertension. 2013; 61: 681–689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Vasam G, Joshi S, Thatcher SE, Bartelmez SH, Cassis LA, Jarajapu YP. Reversal of bone marrow mobilopathy and enhanced vascular repair by angiotensin-(1–7) in diabetes. Diabetes. 2017; 66: 505–518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Batlle D, Wysocki J, Soler MJ, Ranganath K. Angiotensin-converting enzyme 2: enhancing the degradation of angiotensin II as a potential therapy for diabetic nephropathy. Kidney Int. 2012; 81: 520–528. [DOI] [PubMed] [Google Scholar]
- 77. Giani JF, Burghi V, Veiras LC, et al.. Angiotensin-(1–7) attenuates diabetic nephropathy in Zucker diabetic fatty rats. Am J Physiol Renal Physiol. 2012; 302: F1606–F1615. [DOI] [PubMed] [Google Scholar]
- 78. Moon JY, Tanimoto M, Gohda T, et al.. Attenuating effect of angiotensin-(1–7) on angiotensin II-mediated NAD(P)H oxidase activation in type 2 diabetic nephropathy of KK-A(y)/Ta mice. Am J Physiol Renal Physiol. 2011; 300: F1271–F1282. [DOI] [PubMed] [Google Scholar]
- 79. Oudit GY, Liu GC, Zhong J, et al.. Human recombinant ACE2 reduces the progression of diabetic nephropathy. Diabetes. 2010; 59: 529–538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Nadarajah R, Milagres R, Dilauro M, et al.. Podocyte-specific overexpression of human angiotensin-converting enzyme 2 attenuates diabetic nephropathy in mice. Kidney Int. 2012; 82: 292–303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Singh T, Singh K, Sharma PL. Ameliorative potential of angiotensin1–7/Mas receptor axis in streptozotocin-induced diabetic nephropathy in rats. Methods Find Exp Clin Pharmacol. 2010; 32: 19–25. [DOI] [PubMed] [Google Scholar]
- 82. Shi Y, Lo CS, Padda R, et al.. Angiotensin-(1–7) prevents systemic hypertension, attenuates oxidative stress and tubulointerstitial fibrosis, and normalizes renal angiotensin-converting enzyme 2 and Mas receptor expression in diabetic mice. Clin Sci (Lond). 2015; 128: 649–663. [DOI] [PubMed] [Google Scholar]
- 83. Zhang K, Meng X, Li D, et al.. Angiotensin(1–7) attenuates the progression of streptozotocin-induced diabetic renal injury better than angiotensin receptor blockade. Kidney Int. 2015; 87: 359–369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Dong B, Yu QT, Dai HY, et al.. Angiotensin-converting enzyme-2 overexpression improves left ventricular remodeling and function in a rat model of diabetic cardiomyopathy. J Am Coll Cardiol. 2012; 59: 739–747. [DOI] [PubMed] [Google Scholar]
- 85. Murca TM, Almeida TC, Raizada MK, Ferreira AJ. Chronic activation of endogenous angiotensin-converting enzyme 2 protects diabetic rats from cardiovascular autonomic dysfunction. Exp Physiol. 2012; 97: 699–709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Murca TM, Moraes PL, Capuruco CA, et al.. Oral administration of an angiotensin-converting enzyme 2 activator ameliorates diabetes-induced cardiac dysfunction. Regul Pept. 2012; 177: 107–115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Singh K, Singh T, Sharma PL. Beneficial effects of angiotensin (1–7) in diabetic rats with cardiomyopathy. Ther Adv Cardiovasc Dis. 2011; 5: 159–167. [DOI] [PubMed] [Google Scholar]
- 88. Patel VB, Parajuli N, Oudit GY. Role of angiotensin-converting enzyme 2 (ACE2) in diabetic cardiovascular complications. Clin Sci (Lond). 2014; 126: 471–482. [DOI] [PubMed] [Google Scholar]
- 89. Kim SF, Baek SJ, Pack MY. Cloning and nucleotide sequence of the Lactobacillus casei lactate dehydrogenase gene. Appl Environ Microbiol. 1991; 57: 2413–2417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90. Paun A, Danska JS. Modulation of type 1 and type 2 diabetes risk by the intestinal microbiome. Pediatr Diabetes. 2016; 17: 469–477. [DOI] [PubMed] [Google Scholar]
- 91. Han JL, Lin HL. Intestinal microbiota and type 2 diabetes: from mechanism insights to therapeutic perspective. World J Gastroenterol. 2014; 20: 17737–17745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Vaarala O. Human intestinal microbiota and type 1 diabetes. Curr Diab Rep. 2013; 13: 601–607. [DOI] [PubMed] [Google Scholar]
- 93. Zipris D. The interplay between the gut microbiota and the immune system in the mechanism of type 1 diabetes. Curr Opin Endocrinol Diabetes Obes. 2013; 20: 265–270. [DOI] [PubMed] [Google Scholar]
- 94. Slyepchenko A, Maes M, Machado-Vieira R, et al.. Intestinal dysbiosis, gut hyperpermeability and bacterial translocation: missing links between depression, obesity and type 2 diabetes. Curr Pharm Des. 2016; 22: 6087–6106. [DOI] [PubMed] [Google Scholar]
- 95. Bibbo S, Dore MP, Pes GM, Delitala G, Delitala AP. Is there a role for gut microbiota in type 1 diabetes pathogenesis? Ann Med. 2017; 49: 11–22. [DOI] [PubMed] [Google Scholar]
- 96. Scott FW, Pound LD, Patrick C, Eberhard CE, Crookshank JA. Where genes meet environment-integrating the role of gut luminal contents, immunity and pancreas in type 1 diabetes. Transl Res. 2017; 179: 183–198. [DOI] [PubMed] [Google Scholar]
- 97. Paun A, Yau C, Danska JS. Immune recognition and response to the intestinal microbiome in type 1 diabetes. J Autoimmun. 2016; 71: 10–18. [DOI] [PubMed] [Google Scholar]
- 98. Paun A, Yau C, Danska JS. The influence of the microbiome on type 1 diabetes. J Immunol. 2017; 198: 590–595. [DOI] [PubMed] [Google Scholar]
- 99. Brunkwall L, Orho-Melander M. The gut microbiome as a target for prevention and treatment of hyperglycaemia in type 2 diabetes: from current human evidence to future possibilities. Diabetologia. 2017; 60: 943–951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100. Siljander H, Honkanen J, Knip M. Microbiome and type 1 diabetes. EBioMedicine. 2019; 46: 512–521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101. Han H, Li Y, Fang J, et al.. Gut microbiota and type 1 diabetes. Int J Mol Sci. 2018; 19(4): 995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102. Knip M, Honkanen J. Modulation of type 1 diabetes risk by the intestinal microbiome. Curr Diab Rep. 2017; 17: 105. [DOI] [PubMed] [Google Scholar]
- 103. Harsch IA, Konturek PC. The role of gut microbiota in obesity and type 2 and type 1 diabetes mellitus: new insights into "old" diseases. Med Sci (Basel). 2018; 6(2): 32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104. Gurung M, Li Z, You H, et al.. Role of gut microbiota in type 2 diabetes pathophysiology. EBioMedicine. 2020; 51: 102590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105. Beli E, Yan Y, Moldovan L, et al.. Restructuring of the gut microbiome by intermittent fasting prevents retinopathy and prolongs survival in db/db mice. Diabetes. 2018; 67: 1867–1879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106. Duan Y, Prasad R, Feng D, et al.. Bone marrow-derived cells restore functional integrity of the gut epithelial and vascular barriers in a model of diabetes and ACE2 deficiency. Circ Res. 2019; 125: 969–988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. Rowan S, Taylor A. The role of microbiota in retinal disease. Adv Exp Med Biol. 2018; 1074: 429–435. [DOI] [PubMed] [Google Scholar]
- 108. Panwar H, Rashmi HM, Batish VK, Grover S. Probiotics as potential biotherapeutics in the management of type 2 diabetes - prospects and perspectives. Diabetes Metab Res Rev. 2013; 29: 103–112. [DOI] [PubMed] [Google Scholar]
- 109. Hampe CS, Roth CL. Probiotic strains and mechanistic insights for the treatment of type 2 diabetes. Endocrine. 2017; 58: 207–227. [DOI] [PubMed] [Google Scholar]
- 110. Mishra SP, Wang S, Nagpal R, et al.. Probiotics and prebiotics for the amelioration of type 1 diabetes: present and future perspectives. Microorganisms. 2019; 7(3): 67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111. Tiderencel KA, Hutcheon DA, Ziegler J. Probiotics for the treatment of type 2 diabetes: a review of randomized controlled trials. Diabetes Metab Res Rev. 2020; 36: e3213. [DOI] [PubMed] [Google Scholar]
- 112. Sun Z, Sun X, Li J, et al.. Using probiotics for type 2 diabetes mellitus intervention: advances, questions, and potential. Crit Rev Food Sci Nutr. 2020; 60: 670–683. [DOI] [PubMed] [Google Scholar]
- 113. Hill C, Guarner F, Reid G, et al.. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat Rev Gastroenterol Hepatol. 2014; 11: 506–514. [DOI] [PubMed] [Google Scholar]
- 114. Sanchez B, Delgado S, Blanco-Miguez A, Lourenco A, Gueimonde M, Margolles A. Probiotics, gut microbiota, and their influence on host health and disease. Mol Nutr Food Res, 10.1002/mnfr.201600240. [DOI] [PubMed] [Google Scholar]
- 115. Suchodolski JS, Jergens AE. Recent advances and understanding of using probiotic-based interventions to restore homeostasis of the microbiome for the prevention/therapy of bacterial diseases. Microbiol Spectr, 10.1128/microbiolspec.VMBF-0025-2015. [DOI] [PubMed] [Google Scholar]
- 116. Grimm V, Riedel CU. Manipulation of the microbiota using probiotics. Adv Exp Med Biol. 2016; 902: 109–117. [DOI] [PubMed] [Google Scholar]
- 117. Liu X, Cao S, Zhang X. Modulation of gut microbiota-brain axis by probiotics, prebiotics, and diet. J Agric Food Chem. 2015; 63: 7885–7895. [DOI] [PubMed] [Google Scholar]
- 118. Vitetta L, Briskey D, Alford H, Hall S, Coulson S. Probiotics, prebiotics and the gastrointestinal tract in health and disease. Inflammopharmacology. 2014; 22: 135–154. [DOI] [PubMed] [Google Scholar]
- 119. Butel MJ. Probiotics, gut microbiota and health. Med Mal Infect. 2014; 44: 1–8. [DOI] [PubMed] [Google Scholar]
- 120. Vandenplas Y, Huys G, Daube G. Probiotics: an update. J Pediatr (Rio J). 2015; 91: 6–21. [DOI] [PubMed] [Google Scholar]
- 121. Reid G. Probiotics: definition, scope and mechanisms of action. Best Pract Res Clin Gastroenterol. 2016; 30: 17–25. [DOI] [PubMed] [Google Scholar]
- 122. Carter CS, Morgan D, Verma A, et al.. Therapeutic delivery of Ang(1–7) via genetically modified probiotic: a dosing study. J Gerontol A Biol Sci Med Sci. 2020; 75: 1299–1303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123. Buford TW, Sun Y, Roberts LM, et al.. Angiotensin (1–7) delivered orally via probiotic, but not subcutaneously, benefits the gut-brain axis in older rats. Geroscience. 2020; 42: 1307–1321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Smokvina T, Wels M, Polka J, et al.. Lactobacillus paracasei comparative genomics: towards species pan-genome definition and exploitation of diversity. PLoS One. 2013; 8: e68731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125. Maldonado Galdeano C, Novotny Nunez I, Carmuega E, de Moreno de LeBlanc A, Perdigon G. Role of probiotics and functional foods in health: gut immune stimulation by two probiotic strains and a potential probiotic yoghurt. Endocr Metab Immune Disord Drug Targets. 2015; 15: 37–45. [DOI] [PubMed] [Google Scholar]
- 126. Mizock BA. Probiotics. Dis Mon. 2015; 61: 259–290. [DOI] [PubMed] [Google Scholar]
- 127. Sanders ME. Probiotics in 2015: their scope and use. J Clin Gastroenterol. 2015; 49(suppl 1): S2–S6. [DOI] [PubMed] [Google Scholar]
- 128. Simeoli R, Mattace Raso G, Lama A, et al.. Preventive and therapeutic effects of Lactobacillus paracasei B21060-based synbiotic treatment on gut inflammation and barrier integrity in colitic mice. J Nutr. 2015; 145: 1202–1210. [DOI] [PubMed] [Google Scholar]
- 129. Pan T, Guo HY, Zhang H, Liu AP, Wang XX, Ren FZ. Oral administration of Lactobacillus paracasei alleviates clinical symptoms of colitis induced by dextran sulphate sodium salt in BALB/c mice. Benef Microbes. 2014; 5: 315–322. [DOI] [PubMed] [Google Scholar]
- 130. Oliveira M, Bosco N, Perruisseau G, et al.. Lactobacillus paracasei reduces intestinal inflammation in adoptive transfer mouse model of experimental colitis. Clin Dev Immunol. 2011; 2011: 807483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131. Tian P, Li B, He C, et al.. Antidiabetic (type 2) effects of Lactobacillus G15 and Q14 in rats through regulation of intestinal permeability and microbiota. Food Funct. 2016; 7: 3789–3797. [DOI] [PubMed] [Google Scholar]
- 132. Dang F, Jiang Y, Pan R, et al.. Administration of Lactobacillus paracasei ameliorates type 2 diabetes in mice. Food Funct. 2018; 9: 3630–3639. [DOI] [PubMed] [Google Scholar]
- 133. Tunapong W, Apaijai N, Yasom S, et al.. Chronic treatment with prebiotics, probiotics and synbiotics attenuated cardiac dysfunction by improving cardiac mitochondrial dysfunction in male obese insulin-resistant rats. Eur J Nutr. 2018; 57: 2091–2104. [DOI] [PubMed] [Google Scholar]
- 134. Zeng Z, Yuan Q, Yu R, Zhang J, Ma H, Chen S. Ameliorative effects of probiotic Lactobacillus paracasei NL41 on insulin sensitivity, oxidative stress, and beta-cell function in a type 2 diabetes mellitus rat model. Mol Nutr Food Res. 2019; 63: e1900457. [DOI] [PubMed] [Google Scholar]
- 135. Derrien M, Belzer C, de Vos WM. Akkermansia muciniphila and its role in regulating host functions. Microbial Pathogenesis. 2017; 106: 171–181. [DOI] [PubMed] [Google Scholar]
- 136. Macchione IG, Lopetuso LR, Ianiro G, et al.. Akkermansia muciniphila: key player in metabolic and gastrointestinal disorders. Eur Rev Med Pharmacol Sci. 2019; 23: 8075–8083. [DOI] [PubMed] [Google Scholar]
- 137. Xu Y, Wang N, Tan HY, Li S, Zhang C, Feng YB. Function of Akkermansia muciniphila in obesity: interactions with lipid metabolism, immune response and gut systems. Front Microbiol. 2020; 11: 219. [DOI] [PMC free article] [PubMed] [Google Scholar]