

Abstract
Recent assessment reports by the Intergovernmental Panel on Climate Change (IPCC) and the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES) have highlighted the risks to humanity arising from the unsustainable use of natural resources. Thus far, land, freshwater, and ocean exploitation have been the chief causes of biodiversity loss. Climate change is projected to be a rapidly increasing additional driver for biodiversity loss. Since climate change and biodiversity loss impact human societies everywhere, bold solutions are required that integrate environmental and societal objectives. As yet, most existing international biodiversity targets have overlooked climate change impacts. At the same time, climate change mitigation measures themselves may harm biodiversity directly. The Convention on Biological Diversity’s post-2020 framework offers the important opportunity to address the interactions between climate change and biodiversity and revise biodiversity targets accordingly by better aligning these with the United Nations Framework Convention on Climate Change Paris Agreement and the Sustainable Development Goals. We identify the considerable number of existing and proposed post-2020 biodiversity targets that risk being severely compromised due to climate change, even if other barriers to their achievement were removed. Our analysis suggests that the next set of biodiversity targets explicitly addresses climate change-related risks since many aspirational goals will not be feasible under even lower-end projections of future warming. Adopting more flexible and dynamic approaches to conservation, rather than static goals, would allow us to respond flexibly to changes in habitats, genetic resources, species composition, and ecosystem functioning and leverage biodiversity’s capacity to contribute to climate change mitigation and adaptation.
Keywords: biodiversity, ecosystem services, sustainability, policy
Changes in biodiversity and ecosystem function must be considered within the context of multiple pressures. Over the last decades to centuries, the intensive use of land, fresh water, and oceans with the extraction of marine and freshwater organisms, wood, and agricultural commodities has dominated the loss of biodiversity and the deterioration of ecosystems globally (1, 2). Approximately 70–75% of the ice-free land area is affected by human use, nearly 50% intensively so. Since 1961, cropland production increased by about 3.5 times and production of animal products by 2.5 times, supported by a massive enhancement of fertilizer input (+800%) and freshwater withdrawal (+100%) (3, 4). Demand for fish has increased by >3% per year (5), and more than half of edible fish consumption stems from capture fisheries (6). In absence of strong conservation policies and changes in per capita consumption, agricultural expansion is projected to further hasten species extinctions (4, 7–9), while the world fish production (capture and aquaculture) is projected to increase by 18% between 2016 and 2030 (5, 9). In addition to the pressure from direct exploitation, the detrimental impacts of multiple pollution sources all are also harmful to marine, freshwater, and terrestrial biodiversity (9) (Fig. 1). Continued human population growth and the concomitant increase in per capita consumption raise serious concerns about the acceleration of overexploitation and pollution of ecosystems (3, 9). Recent assessment reports by the Intergovernmental Panel on Climate Change (IPCC) and the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES) have highlighted the risks to humanity arising from the unprecedented and unsustainable use of natural resources (1–3), with serious negative consequences for biodiversity and the supply of multiple ecosystem services and hence, for human well-being and the achievement of many conservation targets and Sustainable Development Goals (SDGs).
Fig. 1.

Examples of future projected impacts of climate change (CC) and CO2 on biodiversity and ecosystem processes, which can interact with other major drivers of change (such as land use change, resource extraction, and pollution, among others). Examples are given for terrestrial and marine biomes. Impacts of CC and atmospheric CO2 concentration are in bold. BAU, business as usual and/or high-emission scenarios; SLR, sea-level rise; TL, trophic level. Modified with permission from ref. 9.
Climate Change, the “New Kid on the Biodiversity Block”
The existing pressures will continue to aggravate the effects of human-driven climate change on biodiversity. Although climate change has so far not been shown to be a primary driver of the current loss of biodiversity and ecosystems (1), a rapidly increasing range of observations is indicative of widespread and pervasive impacts of climate change across the structural, compositional, and functional dimensions of biodiversity, from genes to ecosystems (9, 10) (Figs. 1 and 2 and SI Appendix). For instance, recent climate change has likely worsened habitat loss and fragmentation in >18% of terrestrial ecoregions, containing >50% of terrestrial vertebrate species (7). Species have been documented to be moving poleward, with an estimated decadal average shift of 17 km for terrestrial and 72 km for marine taxa (11). In mountain regions, significant upslope movements have been documented (11).
Fig. 2.

Interrelations and feedbacks between hierarchical levels that are important for the future of biodiversity and ecosystem functioning. Direct drivers of global change affect all levels of biodiversity, either directly (white arrows) or indirectly through interactions between levels (gray arrows). Self-reinforcing feedbacks can potentially significantly increase expected negative effects of global change drivers. Modified with permission from ref. 9. E, effects of changes in structure and functioning of ecosystems; ED, effects of changes in diversity of ecosystems, and heterogeneity of landscapes and seascapes; GD, effects of changes in genetic and phenotypic diversity; S, effects of changes in functioning, population size, and range of individual species; SD, effects of changes in local species diversity, species composition, and interspecific relations (SI Appendix, section 2).
Projections of climate change impacts on species diversity are fraught with large uncertainties (9), but model projections showing that climate change will become a significant driver of change for all systems are coherent with the existing observations. In some regions, climate change is expected to outpace other important drivers of biodiversity loss in the coming decades (9). A substantial fraction of marine, aquatic, and terrestrial species may be at risk of extinction during the 21st century due to climate change (9, 10, 12, 13). These extinction risks are based on projected decreases in populations’ range or size, with large variability depending on regions, taxonomic groups, whether or not time inertia referred to as “extinction debt” is considered, and whether or not adaptation of species to climate change (due to both intraspecific diversity and rapid evolution) could reduce species vulnerability (9) (SI Appendix, section 2).
Other than species extinctions, a large variety of other consequences for biodiversity, ecosystem functioning, and ecosystem services is also expected (Figs. 1 and 2). Many of these impacts are likely even for mean global temperature increases below 2 °C and have been reviewed elsewhere (e.g., refs. 9 and 14). We thus only briefly revisit these here and in doing so, identify specifically the common features across marine, terrestrial, and freshwater systems.
Direct temperature impacts are expected to be strongest for ectotherms. Marine corals have received particular attention since even for a warming of only 1.5 °C globally, 70 to 90% of tropical coral reefs may be severely degraded or go extinct by 2050 (14, 15), projections that are in line with recent observations (16). Warming, combined with deoxygenation or food limitation, may cause reductions in the mean body size and abundance of fish and other marine ectotherms by the end of century and thus, lead to negative interactions with fishing, which also reduces fish size and abundance significantly (Fig. 1) (17, 18). On land and in freshwater ecosystems, tropical ectotherms will be negatively affected in response to warmer temperatures since they already live at temperatures close to their optimum. In case of amphibians and strictly aquatic species, changes in precipitation patterns will play an additional crucial role (19). It remains unresolved whether shrinking body size will be a uniform response across terrestrial, freshwater, and marine ecto- as well as endotherms (20–22) since endotherms (and homeotherms) are physiologically buffered from some direct effects of rising temperatures. For marine, freshwater, and land mammals or birds, climate change impacts thus are expected to be realized most strongly through restriction in their range (23, 24).
Where there are no barriers to movement, warmer temperatures likely result in continued poleward and altitudinal shifts of species and entire biomes, with particularly rapid changes in high latitudes due to the greater warming in these regions compared with the rest of the earth (25–27). Within ecoregions, climate change will alter active-season lengths and seasonal patterns. Species are expected to respond to these changes by shifting their phenology: for instance, leaf out and flowering, seasonal migration, or breeding (10). The changing interplay between species and their abiotic environment can also result in shifts in consumer–resource interactions, as already demonstrated through empirical evidence, including losses of top predators or top herbivores (10, 22, 28, 29).
Rising levels of CO2 in the atmosphere, in addition to advancing climate change, have contributed to the observed shifts from grasslands to increasingly woody-dominated vegetation in semiarid regions (30). They also lead to ocean and freshwater acidification, which is expected to reduce growth rates in calcifying phytoplankton and organisms like gastropods, crustaceans, shellfish, or corals, whereas some primary producers might benefit from increased CO2 (31–33). Direct effects of CO2 thus can have knock-on effects across all systems on food web structures and for the integrity of the habitats (32, 33). In many regions, climate change together with rising atmospheric CO2 is expected to enhance the already observed biotic homogenization caused by direct human impacts, such as fishing and agriculture by favoring generalists (34, 35) (Fig. 2 and SI Appendix, section 2).
The impacts of the frequency and severity of weather extremes have been much less studied than impacts of mean climate change effects but may be important determinants of the resilience of communities: for instance, through the capacity of species to follow shifting habitats (11). More pronounced or frequent periods of drought or heat waves could impact disproportionally diversity in regions already hot and/or dry today (36), as species already survive close to physiological limits. As an additional factor, the combinations of abiotic and biotic characteristics that have not been observed in the past might lead to the emergence of novel communities, in which species will co-occur in historically unknown associations (22). Climate change (and on land, atmospheric CO2) can also be favorable to some species in cases when it provides more resources for growth, reproduction, and distribution (10). As such, climate change may enhance the accelerating pressure from biological invasions that follow from direct human introductions (37).
Taken together, the expected climate-driven changes in species ranges, community composition, and physiology will not only greatly enhance the existing pressures on biodiversity but also lead to “no-analog” challenges for conservation. Without the use of climate change impact scenarios for their implementation, static biodiversity conservation goals, as stated in the current Aichi Targets 11 and 15 of the Convention of Biological Diversity (CBD; on protected areas and restoration of degraded ecosystems, respectively) (Table 1 and SI Appendix, Table S1) are therefore inadequate for achieving conservation goals and their contributions to multiple other sustainable development objectives.
Table 1.
Conservation targets directly impacted by climate change
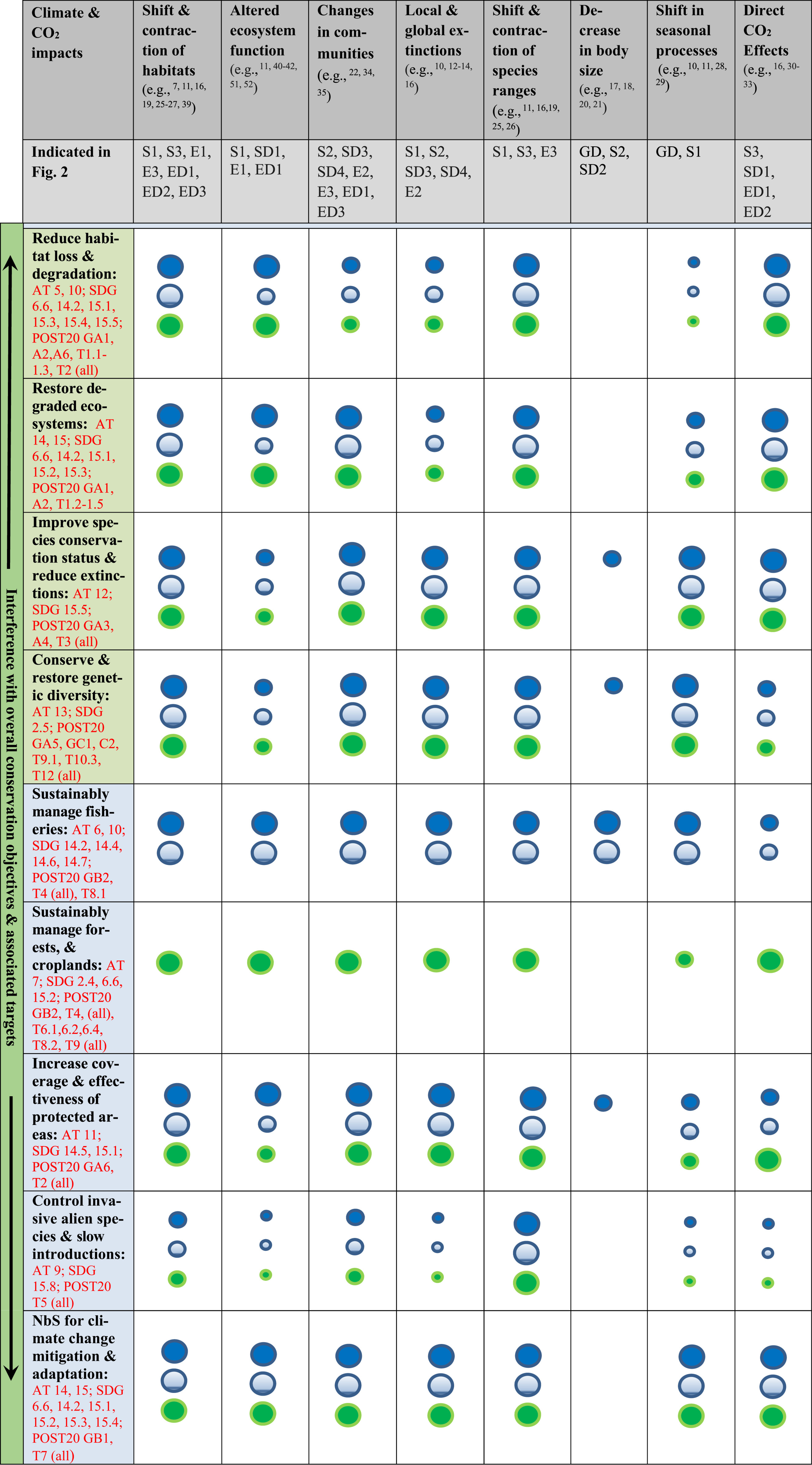 |
Matrix-table of projected effects of climate change on biodiversity (top row) that can interfere with conservation objectives and hence the achievement of Aichi Targets (AT), the draft action targets in the post-2020 Global Biodiversity Framework (POST20, with Components of the Goals = G and the Target Actions = T; see https://www.cbd.int/sbstta/sbstta-24/post2020-monitoring-en.pdf), or Sustainable Development Goals (SDG & target number)(left column). Objectives related to the improvements of biodiversity status: cells highlighted in pale green; those related to addressing direct drivers: cells highlighted in pale blue. For the climate change and CO2 impacts on species, communities and ecosystems see literature cited in the table, the text, Fig 2, and the SI Appendix. An overview of the listed AT, POST20 and SDGs is provided in the SI Appendix, Table S1. The associations with AT, POST20 and SDG are expert-based, made by the authors of the manuscript; the interference of climate change and rising CO2 with conservation objectives and associated targets is indicated for:  marine;
marine;  freshwater;
freshwater;  terrestrial ecosystems. Some of the targets relate to several conservation objectives. Large symbols indicate strong, well-documented pervasive interference; regular size indicates moderate, well-documented interference; small size indicates weak interference or lack of strong evidence (but reasonable supposition); empty cells: little evidence for interference. NbS, Nature-based solutions.
terrestrial ecosystems. Some of the targets relate to several conservation objectives. Large symbols indicate strong, well-documented pervasive interference; regular size indicates moderate, well-documented interference; small size indicates weak interference or lack of strong evidence (but reasonable supposition); empty cells: little evidence for interference. NbS, Nature-based solutions.
Climate Change Impacts on Ecosystem Functioning and Services through Changes in Biodiversity
Observed impacts of climate change and increasing atmospheric CO2 concentrations on biodiversity, habitat quality, biotic interactions, and organism’s physiology raise concerns about accelerating the overall loss of functional diversity and ecosystem services (35, 38, 39). How altered species composition and ecosystem functioning will impact multiple ecosystem services in different regions remains largely unquantified: appropriate process-based models that capture the complex interactions between biodiversity and ecosystem functioning globally do not yet exist (9). Nevertheless, some model simulations address how climate and CO2-induced changes in species and functional groups composition will alter regulating and provisioning services.
In a high CO2 world, plants with the C3 photosynthetic pathway generally increase the efficiency with which they use water (2), which underpins projections of increased catchment runoff, impacting freshwater ecosystems both directly and through the enhanced inflow of dissolved organic carbon. The CO2 effects compensate to some degree the expected reduced net primary productivity and ecosystem carbon uptake arising from hotter and drier climate, but accelerated carbon losses from the large stores in northern peatlands and permafrost soils or from tropical ecosystem vegetation in response to warming and drying are very likely (40, 41). Likewise, future uptake of carbon from the atmosphere in marine systems will depend on net primary productivity from phytoplankton in addition to physical processes (dissolution and mixing), with a projected decline in net primary productivity by the end of the century (42).
In dry and hot regions, projected negative impacts of reduced herbaceous productivity on land have been shown to translate into reduction in livestock production, with detrimental economic impacts (43), and to reduce crop yields (44). Moreover, crop production serves as an illustrative example of the links between provisioning ecosystem services, regulating services, biodiversity, and climate change. Approximately 5 to 8% of production (by volume) would be lost annually without pollinators (45). Climate change thus may affect food supply not only through suitability of crops and its direct impact on yields but also, through shifts in the range of plant–pollinator networks. This loss would have overproportional effects on human health and the economy since pollination-dependent crops tend to be nutritious (fruits, nuts) and/or cash crops (coffee) (45). Other climate change impacts on human societies through changes in biodiversity range from the distribution of vector-borne diseases such as malaria, which are reaching new areas, to changes in cultural (nonmaterial) ecosystem services through impacts on recreation and cultural identity driven by species migrations and extinctions (11, 46).
Despite the inherent limitations of a purely economic valuation, the large estimated total economic value of marine, freshwater, and terrestrial ecosystems (nearly 125 trillion US dollars [2007 values]) (46) is indicative of the importance of ecosystem services to human society and the potential risks arising from climate change to both economic wealth and human well-being. While global-scale simulation studies are scarce, the existing literature points unequivocally toward disproportionate climate change impacts on poor societies both for material and nonmaterial benefits (e.g., refs. 43 and 47–50).
Climate Regulation and Feedbacks Related to Biodiversity
Feedbacks could either enhance the solution space regarding climate change mitigation and/or biodiversity conservation measures or increase the risk of breaching thresholds and “tipping points.” In the global climate system, for instance, changes in marine and terrestrial carbon uptake and losses underpin prominent feedbacks, which amplify the original CO2 and climate forcing in many scenarios (51). These feedbacks are directly caused by changes in species’ physiologies and shifts in functional diversity, which have received only limited attention (51). It is the altered metabolism of marine phyto- and zooplankton in response to climate change and rising CO2 that feeds back to atmospheric CO2 and climate via effects on the biological pump (52). On land, shifts toward coniferous forests in what is currently tundra or toward drought-tolerant vegetation in the tropics could dampen the expected negative impacts of climate change on productivity and carbon uptake (51). Additionally, climate–ecosystem–carbon feedbacks exist beyond shifts in autotrophic organisms. Even though animals have long been considered to play a negligible role in carbon cycling in natural ecosystems, herbivory demonstrably reduces autotrophic biomass, enhances light transfer into plant canopies or deeper water, and increases nutrient input to the soil and sediments through its impact on litter amount and quality (38, 52). Cascading trophic effects triggered by top predators or the largest herbivores propagate through food webs and reverberate through to the functioning of whole ecosystems. On land, experimental evidence shows the ecosystem consequences of the presence or absence of herbivores and carnivores to be quantitatively as large as the effects of other environmental change drivers such as warming, enhanced CO2, or variable nitrogen deposition (53). If “zoogeochemistry” (38) is indeed as important as abiotic climate change factors, as suggested by experiments and field observations, neglecting the broader role of carnivores, grazers, and browsers in modulating ecosystems’ responses to climate change is an important oversight.
Biodiversity is also integral to ecosystem–climate feedback loops beyond carbon cycle processes. Changes in vegetation cover alter the local surface–atmosphere exchange of energy and feedback to regional climate, which can either enhance or reduce warming, depending on the regions where this occurs (54). Global warming exacerbates changes in plankton species composition, blooms of cyanobacteria, and eutrophication, which will in turn contribute to ocean/fresh water–atmosphere feedbacks: for instance, through shifts in the emissions of dimethyl sulfide, a component contributing notably to aerosol formation and cloud processes (55). Quantifying the role of functional diversity across all trophic levels in the climate system therefore can provide highly relevant information in view of the expected continued strong declines in species diversity arising from climate change but also when designing conservation measures such as the expansion of protected areas, sustainable fishing, or rewilding.
The complex interactions between ecosystem processes, atmospheric composition, and climate highlight the need to account for climate change in conservation goals. Similar to species-based goals to protect biodiversity, climate change will also render static ecosystem protection goals inadequate. Ignoring reinforcements between populations’ decline and loss of genetic diversity, alteration of species composition, and ecosystem functioning can lead to underestimation of possible negative effects of climate change or other environmental pressures (Fig. 2)—while the dynamic role of biodiversity in ecosystem–climate feedbacks in turn will have to be factored in when designing measures and assessing their potential success in reaching the Paris Agreement’s goals.
Indirect Climate Change Effects: Cobenefits and Pitfalls of Land-Based Climate Change Mitigation
Despite the wide range of benefits arising from limiting warming to below 2 °C for all ecosystems and their biodiversity (ref. 14 and references therein), the way climate change mitigation will be implemented is as critical for the future for land and freshwater ecosystems as limiting warming itself. Several prominent measures to achieve mitigation have been identified as being in conflict with biodiversity conservation, the supply of many ecosystem services and human well-being.
The current large annual CO2 uptake in terrestrial ecosystems [∼30% of anthropogenic emissions (51)] underpins climate change mitigation scenarios of large-scale growth of bioenergy crops or expansion of forest area, both aiming to achieve negative emissions (56). As today’s land area is already heavily used and food demand is projected to increase substantially, conversion of areas (on average) equivalent to about one-third of today’s food crop area or 10 to 15% of today’s forest area for mitigation purposes (56) would enhance competition for land, place enormous pressure on existing conservation areas, and impede land–area-related biodiversity conservation measures (8, 57, 58). It would also further aggravate hunger and loss of ecosystem services related to SDGs such as availability of clean water or clean air (1, 3, 58). These results are particularly pertinent in the light of studies that have raised doubts on whether the projected cumulative carbon uptake on land at the massive scales proposed could, in fact, be achieved (59).
In contrast, avoiding further conversion of natural ecosystems into managed lands should be a foremost priority in order to maintain carbon sinks and avoid large carbon emissions, as well as provide large benefits for biodiversity. Given that land use contributed on average 23% to total annual emissions (CO2 eq in 2007 to 2016) of CO2, N2O, and CH4, multiple mitigation options exist on managed land (3), especially when complemented by altered consumption and reduced per capita consumption in high-income countries. When ecosystem processes are properly taken into consideration, ecosystem-based mitigation measures can have significant and cost-effective adaptation cobenefits that target conservation purposes, supply of multiple ecosystem services, and human well-being (3, 60, 61).
Climate Change and Global Biodiversity Targets
The Aichi Targets of the CBD and some of the SDGs have been adopted to motivate actions to sustain biodiversity, and its contributions to human well-being and societal development. Global targets for biodiversity have been missed in the past, and the world is still far from achieving most of the Aichi Targets by this year’s deadline (1, 62). Even if these targets had been set for much later in the future (2050 and beyond), evidence from scenarios and modeling analyses show that many of them are unlikely to be achieved for the vast majority of socioeconomic projections (9).
Biodiversity directly supports the achievement of at least 13 SDGs, rising to all 17 SDGs after indirect interactions are considered (63). At the same time, climate change has the potential to undermine 16 SDGs (64). Climate change will exacerbate existing conservation challenges if species lose suitable climate conditions in currently protected areas and no replacement for these protected areas can be created or if economically valuable species come under double pressure from human demand and climate change. Considering climate change more directly in the formulation and implementation of future biodiversity targets therefore has clear cobenefits for addressing a wide range of environmental and societal challenges. It also reduces the risk that measures put in place for achieving a target will be ineffective.
Table 1 summarizes some of the chief impacts of climate change on marine, freshwater, and/or terrestrial biodiversity and assesses how these impacts hinder the achievement of two overarching conservation objectives and related Aichi Targets and SDGs: improving the status of biodiversity and addressing direct drivers that underlie the loss of biodiversity. Several Aichi Targets were not realistic or not well formulated under even low-end scenarios of climate change. None explicitly address the urgency of climate mitigation and adaptation as a critical component of biodiversity conservation. For example, Aichi Target 10 calls for action to be taken to minimize multiple anthropogenic pressures on vulnerable ecosystems impacted by climate change, especially coral reefs, but makes no mention of the urgency to hold climate change to the lowest levels possible while at the same time implementing conservation measures. Aichi Target 12, which aims to prevent the extinction of all known threatened species and improve their conservation status would seem unrealistic given the evidence that climate change has already resulted in significant range shifts and that even very low levels of future climate change (i.e., 1.5 °C to 2 °C global warming) are projected to reduce the range size of a substantial fraction of species and put many species at high risk of extinction (10, 12–14). The projected climate change impacts on terrestrial and marine ecosystems also seriously compromise meeting the targets for reducing the degradation of terrestrial habitats and managing and harvesting sustainably all fish and invertebrate stocks (Aichi Targets 5 and 6).
The next set of targets in the post-2020 global biodiversity framework are currently being designed (65). At this point in time, four goals corresponding to the 2050 Vision on Biodiversity are proposed, together with 20 action-oriented targets with a time horizon of 2030, many of which can be mapped onto the existing targets (SI Appendix, Table S1). In a similar spirit to the Aichi Targets, Goal A of the post-2020 biodiversity framework calls for substantial (as yet unquantified) reduction of the number of species threatened with extinction, without acknowledging climate change as a factor that could potentially impede the success of any of the implemented conservation measures. Minimizing additional stressors on ecosystems is vital for maintaining resilience in the face of climate change, but ecosystems in many locations, including tropical coral reefs, rainforest and savannas, or (sub-)arctic tundra (Fig. 1), will be unable to adapt to moderate to high levels of climate change even when other stressors are addressed; climate change will enhance the risk of extinction for a considerable number of species (7, 10, 14). Climate change interactions with land use and fisheries management (9) are not considered in the relevant draft post-2020 action targets (POST20 Targets 8 and 9). The current version of the post-2020 targets only explicitly addresses climate change and climate impacts in 1 of 20 proposed targets (POST20 Target 7).
On the whole, suitable conservation measures will be hard to implement based on static targets. For example, the size, placement, and connectivity of protected areas need to account for shifts and contractions of habitats and species distributions (66) (Table 1). Like the current formulation of the protected areas target (Aichi Target 11), the corresponding draft post-2020 targets might lead to static approaches to setting up and maintaining protected areas, whereas flexible and dynamic measures will be needed that allow us to account for spatial shifts in species and communities (66). At the same time, it will be necessary to acknowledge that in the face of climate change, conservation will be much more about managing change; a return to a historical state of biodiversity will be hard to achieve. References to adaptation in the proposed 2020 targets rightly emphasize the role of nature-based solutions in mitigating climate related disaster risk and attenuating climate impacts on human livelihoods (e.g., POST20 Targets 7.1, 10.2, and 10.3). However, the framework, as drafted, inadequately omits the need to reduce climate impacts and risks to biodiversity itself and to enhance biodiversity with ecosystem-based adaptation. This would warrant a separate target.
Because protection and restoration of habitats can contribute substantially to climate change mitigation and adaptation, targets related to reducing the degradation and unsustainable use of land, freshwater, and marine resources thus offer clear cobenefits for jointly achieving biodiversity and climate change objectives. The post-2020 action targets explicitly call for ecosystem-based management to factor in climate change mitigation (POST20 Target 7), strengthening considerably the earlier formulations in the related Aichi Target 15. Enhancing the contribution of biodiversity to carbon stocks through conservation and restoration is important since measures taken could also support a broader range of objectives, such as those related to water (POST20 Target 10) and human health (POST20 Target 11). Successful implementation of these measures is particularly likely if accompanied by actions targeting changes in human consumption (3), which are being asked for in draft Target 15, which promises to be a considerable step up from Aichi Target 4.
Given the well-established strong interactions between climate change and biodiversity, it is critical that these interactions be more explicitly addressed in the post-2020 global biodiversity framework by
• highlighting the need to rapidly reduce anthropogenic greenhouse gas emissions for strong climate change mitigation as a fundamental contribution to protecting ecosystems and biodiversity,
• critically assessing which mitigation and adaptation measures to adopt in order to increase biodiversity or reduce risks to it (including a related target),
• setting realistic targets given the unavoidable impacts of future climate change on biodiversity,
• clearly identifying the contributions that biodiversity can make to climate change mitigation and adaptation, and
• devising a process that will result in successful, continuous monitoring of progress toward targets’ achievement in view of impacts of climate change.
We suggest that measures taken to implement the post-2020 biodiversity targets need to consider all of the five points raised above and should explicitly address climate change-related issues within individual targets on drivers, impacts, and responses. This should also contribute to position the POST20 targets as one further piece to synergistically tackle climate change, together with the SDGs and the Paris Agreement.
Climate, biodiversity, and sustainable development policies are often disconnected and sometimes in conflict. However, climate change awareness across policy sectors comes with potentially large gains as, for example, natural resources management and conservation measures could also be reinforced by synergistic approaches with engineering climate change mitigation solutions. Putting in practice the cross-sectoral policy strategies required to flexibly adjust conservation measures is challenging and has so far not been achieved. It is critical therefore to identify different visions, uncover hidden synergies, and identify barriers toward implementation of goal-achieving policies. In dialogue with policy decision makers and other key stakeholders, linkages between societal values and conservation in a climate change context can be identified and used to explore the effects of low-regrets options as well as climate-targeted options for marine, freshwater, and terrestrial conservations to respond flexibly to changing circumstances (67–70). The concept of technological–ecological synergies is based on the premise that mutually beneficial relationships can be developed between technological/interventional and nature-based management options and has large potential for climate–biodiversity cobenefits (67). Still, many of the options are likely to also lead to trade-offs between different conservation goals (such as optimizing ecological function or genetic resources via interventionistic approaches vs. maintaining pristine ecosystems), which calls for adaptation actions adjusted to location-specific conditions that both acknowledge these trade-offs and seek to minimize these across broader spatial scales (69, 70). Rather than relying on fixed decisions, conservation management strategies and their underpinning goals need to be adjusted and reassessed over time to account for climate or socioeconomic developments, new knowledge or technologies, and changing societal values (67–70).
Concluding Remarks
Taking climate change into account as an up-front challenge for biodiversity affects the formulation of new biodiversity targets (71), the balance between biodiversity-focused adaptation and ecosystem-based adaptation (72, 73), and their interactions with other sustainable development objectives, as well as the measures that will be nationally and locally implemented to achieve these. “Bending the curve” of biodiversity loss (73) and the ambition to retain, restore, and protect natural ecosystems (72) will be difficult, if not impossible, to achieve, unless climate change is considered explicitly as a main threat to biodiversity and ecosystems. Since the strongest impacts of climate change are expected to emerge in many regions from around midcentury onward, even if biodiversity goals were seemingly achieved by 2030, such an apparent achievement could well lead to a false sense of success as it could be rapidly reversed in the decades to follow. Formulating “climate-informed targets” and the associated dynamically responsive policies is an immense challenge. Enhancing the dialogue between important conventions, such as the United Nations Framework Convention on Climate Change (UNFCCC) and on Biological Diversity (CBD), and the associated science-policy platforms of the IPCC and IPBES could help align policy and create added value to address this challenge. The POST20 targets could become one further element in the global strategy against climate change. A better alignment of conventions and scientific assessments could also stimulate the urgent decarbonization of the economy and ensure that climate change is minimized through actions that benefit from rather than compromise biodiversity protection.
Supplementary Material
Acknowledgments
We acknowledge the support by Rainer Krug, Hien Ngo, and Max Gueze in parts of the literature review. Pierre Lopez drew Fig. 1, using spatial data layers provided by Aidin Niamir (https://doi.org/10.5281/zenodo.3975694).
Footnotes
The authors declare no competing interest.
This article is a PNAS Direct Submission.
This article contains supporting information online at https://www.pnas.org/lookup/suppl/doi:10.1073/pnas.2009584117/-/DCSupplemental.
Data Availability.
All study data are included in the article and SI Appendix.
References
- 1.IPBES , Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science—Policy Platform on Biodiversity and Ecosystem Services, Brondizio E. S., et al., Eds. (IPBES Secretariat, Bonn, Germany, 2019). [Google Scholar]
- 2.Settele J., et al. , “Terrestrial and water systems” in Climate Change 2014: Impacts, Adaptation, Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the IPCC, Field C. B., et al., Eds. (Cambridge University Press, Cambridge, United Kingdom, 2014), pp. 271–359. [Google Scholar]
- 3.IPCC , “Summary for policy makers” in Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems, Shukla P. R., Ed. et al. (Intergovernmental Panel on Climate Change, Geneva, Switzerland, 2019), pp. 3–36. [Google Scholar]
- 4.Arneth A., et al. , “Chapter 1: Framing and context” in IPCC Special Report on Climate Change and Land, Shukla P. R. et al., Eds. (Intergovernmental Panel on Climate Change Geneva, Switzerland: ). [Google Scholar]
- 5.FAO , The State of World Fisheries and Aquaculture 2018—Meeting the Sustainable Development Goals (Food and Agricultural Organisation, Rome, Italy, 2018). [Google Scholar]
- 6.Edwards P., et al. , Misunderstandings, myths and mantras in aquaculture: Its contribution to world food supplies has been systematically over reported. Mar. Policy 106, 103547 (2019). [Google Scholar]
- 7.Segan D. B., Murray K. A., Watson J. E. M., A global assessment of current and future biodiversity vulnerability to habitat loss-climate change interactions. Glob. Ecol. Conserv. 5, 12–21 (2016). [Google Scholar]
- 8.Hof C., et al. , Bioenergy cropland expansion may offset positive effects of climate change mitigation for global vertebrate diversity. Proc. Natl. Acad. Sci. U.S.A. 115, 13294–13299 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Shin Y.-J. et al.; IPBES Global Assessment on Biodiversity and Ecosystem Services , “Chapter 4: Plausible futures of nature, its contributions to people and their good quality of life” in Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science—Policy Platform on Biodiversity and Ecosystem Services, Brondizio E. S., Ed. et al. (IPBES Secretariat, Bonn, Germany, 2019), pp. 1–264. [Google Scholar]
- 10.Bellard C., Bertelsmeier C., Leadley P., Thuiller W., Courchamp F., Impacts of climate change on the future of biodiversity. Ecol. Lett. 15, 365–377 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Pecl G. T., et al. , Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 355, 355 (2017). [DOI] [PubMed] [Google Scholar]
- 12.Visconti P., et al. , Projecting global biodiversity indicators under future development scenarios. Conserv. Lett. 9, 5–13 (2016). [Google Scholar]
- 13.Urban M. C., Climate change. Accelerating extinction risk from climate change. Science 348, 571–573 (2015). [DOI] [PubMed] [Google Scholar]
- 14.Hoegh-Guldberg O., et al. , The human imperative of stabilizing global climate change at 1.5°C. Science 365, eaaw6974 (2019). [DOI] [PubMed] [Google Scholar]
- 15.Eakin C. M., Sweatman H. P. A., Brainard R. E., The 2014-2017 global-scale coral bleaching event: Insights and impacts. Coral Reefs 38, 539–545 (2019). [Google Scholar]
- 16.Gattuso J.-P., et al. , Contrasting futures for ocean and society from different anthropogenic CO2 emissions scenarios. Science 349, aac4722 (2015). [DOI] [PubMed] [Google Scholar]
- 17.Cheung W. W. L., et al. , Shrinking of fishes exacerbates impacts of global ocean changes on marine ecosystems. Nat. Clim. Chang. 3, 254–258 (2013). [Google Scholar]
- 18.Jones D. O. B., et al. , Global reductions in seafloor biomass in response to climate change. Glob. Change Biol. 20, 1861–1872 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Tedesco P. A., et al. , A scenario for impacts of water availability loss due to climate change on riverine fish extinction rates. J. Appl. Ecol. 50, 1105–1115 (2013). [Google Scholar]
- 20.Naya D. E., Naya H., Cook J., Climate change and body size trends in aquatic and terrestrial endotherms: Does habitat matter? PLoS One 12, e0183051 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ohlberger J., Climate warming and ectotherm body size—from individual physiology to community ecology. Funct. Ecol. 27, 991–1001 (2013). [Google Scholar]
- 22.Lurgi M., López B. C., Montoya J. M., Novel communities from climate change. Philos. Trans. R. Soc. Lond. B Biol. Sci. 367, 2913–2922 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kubisch A., et al. , Predicting range shifts under global change: The balance between local adaptation and dispersal. Ecography 36, 873–882 (2013). [Google Scholar]
- 24.Li Y. M., et al. , Climate and topography explain range sizes of terrestrial vertebrates. Nat. Clim. Chang. 6, 498–502 (2016). [Google Scholar]
- 25.Cheung W. W. L. L., et al. , Projecting global marine biodiversity impacts under climate change scenarios. Fish. 10, 235–251 (2009). [Google Scholar]
- 26.Comte L., et al. , Climate-induced changes in the distribution of freshwater fish: Observed and predicted trends. Freshw. Biol. 58, 625–639 (2013). [Google Scholar]
- 27.Ostberg S., et al. , Critical impacts of global warming on land ecosystems. Earth Syst. Dyn. 4, 347–357 (2013). [Google Scholar]
- 28.Damien M., Tougeron K., Prey-predator phenological mismatch under climate change. Curr. Opin. Insect Sci. 35, 60–68 (2019). [DOI] [PubMed] [Google Scholar]
- 29.Prop J., et al. , Climate change and the increasing impact of polar bears on bird populations. Front. Ecol. Evol. 3, 33 (2015). [Google Scholar]
- 30.Zhu Z. C., et al. , Greening of the earth and its drivers. Nat. Clim. Chang. 6, 791–795 (2016). [Google Scholar]
- 31.Hasler C. T., et al. , Biological consequences of weak acidification caused by elevated carbon dioxide in freshwater ecosystems. Hydrobiologia 806, 1–12 (2018). [Google Scholar]
- 32.Heinze C., et al. , The ocean carbon sink—impacts, vulnerabilities and challenges. Earth Syst. Dyn. 6, 327–358 (2015). [Google Scholar]
- 33.Rogers A., Blanchard J. L., Mumby P. J., Vulnerability of coral reef fisheries to a loss of structural complexity. Curr. Biol. 24, 1000–1005 (2014). [DOI] [PubMed] [Google Scholar]
- 34.Magurran A. E., Dornelas M., Moyes F., Gotelli N. J., McGill B., Rapid biotic homogenization of marine fish assemblages. Nat. Commun. 6, 8405 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Clavel J., Julliard R., Devictor V., Worldwide decline of specialist species: Toward a global functional homogenization? Front. Ecol. Environ. 9, 222–228 (2011). [Google Scholar]
- 36.Li J., et al. , Global priority conservation areas in the face of 21st century climate change. PLoS One 8, e54839 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Bellard C., Jeschke J. M., Leroy B., Mace G. M., Insights from modeling studies on how climate change affects invasive alien species geography. Ecol. Evol. 8, 5688–5700 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Schmitz O. J., et al. , Animals and the zoogeochemistry of the carbon cycle. Science 362, eaar3213 (2018). [DOI] [PubMed] [Google Scholar]
- 39.Eigenbrod F., Gonzalez P., Dash J., Steyl I., Vulnerability of ecosystems to climate change moderated by habitat intactness. Glob. Change Biol. 21, 275–286 (2015). [DOI] [PubMed] [Google Scholar]
- 40.Nichols J. E., Peteet D. M., Rapid expansion of northern peatlands and doubled estimate of carbon storage. Nat. Geosci. 12, 917–921 (2019). [Google Scholar]
- 41.Huntingford C., et al. , Simulated resilience of tropical rainforests to CO2-induced climate change. Nat. Geosci. 6, 268–273 (2013). [Google Scholar]
- 42.Bopp L., et al. , Multiple stressors of ocean ecosystems in the 21st century: Projections with CMIP5 models. Biogeosciences 10, 6225–6245 (2013). [Google Scholar]
- 43.Boone R. B., Conant R. T., Sircely J., Thornton P. K., Herrero M., Climate change impacts on selected global rangeland ecosystem services. Glob. Change Biol. 24, 1382–1393 (2018). [DOI] [PubMed] [Google Scholar]
- 44.Rosenzweig C., et al. , Assessing agricultural risks of climate change in the 21st century in a global gridded crop model intercomparison. Proc. Natl. Acad. Sci. U.S.A. 111, 3268–3273 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Potts S. G., et al. , Safeguarding pollinators and their values to human well-being. Nature 540, 220–229 (2016). [DOI] [PubMed] [Google Scholar]
- 46.Costanza R., et al. , Changes in the global value of ecosystem services. Glob. Environ. Change 26, 152–158 (2014). [Google Scholar]
- 47.Markkula I., Turunen M., Rasmus S., A review of climate change impacts on the ecosystem services in the Saami Homeland in Finland. Sci. Total Environ. 692, 1070–1085 (2019). [DOI] [PubMed] [Google Scholar]
- 48.Lindegren M., Brander K., Adapting fisheries and their management to climate change: A review of concepts, tools, frameworks, and current progress toward implementation. Rev. Fish. Sci. Aquacult. 26, 400–415 (2018). [Google Scholar]
- 49.Immerzeel W. W., et al. , Importance and vulnerability of the world’s water towers. Nature 577, 364–369 (2020). [DOI] [PubMed] [Google Scholar]
- 50.Allison E. A., The spiritual significance of glaciers in an age of climate change. Wiley Interdiscip. Rev. Clim. Change 6, 493–508 (2015). [Google Scholar]
- 51.Ciais P., et al. , “Carbon and other biogeochemical cycles” in Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, Stocker T. F., Ed. et al. (Cambridge University Press, Cambridge, United Kingdom, 2013), pp. 465–570. [Google Scholar]
- 52.Boyd P. W., Claustre H., Levy M., Siegel D. A., Weber T., Multi-faceted particle pumps drive carbon sequestration in the ocean. Nature 568, 327–335 (2019). [DOI] [PubMed] [Google Scholar]
- 53.Hooper D. U., et al. , A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 486, 105–108 (2012). [DOI] [PubMed] [Google Scholar]
- 54.Perugini L., et al. , Biophysical effects on temperature and precipitation due to land cover change. Environ. Res. Lett. 12, 053002 (2017). [Google Scholar]
- 55.Hopkins F. E., et al. , A meta-analysis of microcosm experiments shows that dimethyl sulfide (DMS) production in polar waters is insensitive to ocean acidification. Biogeosciences 17, 163–186 (2020). [Google Scholar]
- 56.Rogelj J., et al. , Scenarios towards limiting global mean temperature increase below 1.5 degrees C. Nat. Clim. Chang. 8, 325–332 (2018). [Google Scholar]
- 57.Veldman J. W., et al. , Where tree planting and forest expansion are bad for biodiversity and ecosystem services. Bioscience 65, 1011–1018 (2015). [Google Scholar]
- 58.Fuss S., et al. , Negative emissions. Part 2: Costs, potentials and side effects. Environ. Res. Lett. 13, 063002 (2018). [Google Scholar]
- 59.Harper A. B., et al. , Land-use emissions play a critical role in land-based mitigation for Paris climate targets. Nat. Commun. 9, 2938 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Seddon N., et al. , Grounding nature-based climate solutions in sound biodiversity science. Nat. Clim. Chang. 9, 84–87 (2019). [Google Scholar]
- 61.Donatti C. I., et al. , Indicators to measure the climate change adaptation outcomes of ecosystem-based adaptation. Clim. Change 158, 413–433 (2019). [Google Scholar]
- 62.Tittensor D. P., et al. , A mid-term analysis of progress toward international biodiversity targets. Science 346, 241–244 (2014). [DOI] [PubMed] [Google Scholar]
- 63.Blicharska M., et al. , Biodiversity’s contributions to sustainable development. Nat. Sustain. 2, 1083–1093 (2019). [Google Scholar]
- 64.Fuso Nerini F., et al. , Connecting climate action with other sustainable development goals. Nat. Sustain. 2, 674–680 (2019). [Google Scholar]
- 65.CBD-SBSTTA , Draft monitoring framework for the post-2020 global biodiversity framework for review. Convention on Biological Diversity, Subsidiary Body on Scientific, Technical and Technological Advice. https://www.cbd.int/sbstta/sbstta-24/post2020-monitoring-en.pdf. Accessed 12 November 2020.
- 66.Hanson J. O., et al. , Global conservation of species’ niches. Nature 580, 232–234 (2020). [DOI] [PubMed] [Google Scholar]
- 67.Hernandez R. R., et al. , Techno-ecological synergies of solar energy for global sustainability. Nat. Sustain. 2, 560–568 (2019). [Google Scholar]
- 68.Kwakkel J. H., Haasnoot M., Walker W. E., Comparing robust decision-making and dynamic adaptive policy pathways for model-based decision support under deep uncertainty. Environ. Model. Softw. 86, 168–183 (2016). [Google Scholar]
- 69.Prober S. M., et al. , Shifting the conservation paradigm: A synthesis of options for renovating nature under climate change. Ecol. Monogr. 89, e01333 (2019). [Google Scholar]
- 70.Tittensor D. P., et al. , Integrating climate adaptation and biodiversity conservation in the global ocean. Sci. Adv. 5, eaay9969 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Butchart S. H. M., Di Marco M., Watson J. E. M., Formulating smart commitments on biodiversity: Lessons from the Aichi targets. Conserv. Lett. 9, 457–468 (2016). [Google Scholar]
- 72.Watson J. E. M., et al. , Set a global target for ecosystems. Nature 578, 360–362 (2020). [DOI] [PubMed] [Google Scholar]
- 73.Mace G. M., et al. , Aiming higher to bend the curve of biodiversity loss. Nat. Sustain. 1, 448–451 (2018). [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All study data are included in the article and SI Appendix.