

Petit-Pedrol and Groc review the regulatory role of NMDAR nanoscale organization and surface dynamics in glutamate synapse transmission and plasticity.
Abstract
Understanding neurotransmitter system crosstalk in the brain is a major challenge in neurobiology. Several intracellular and genomic cascades have been identified in this crosstalk. However, the discovery that neurotransmitter receptors are highly diffusive in the plasma membrane of neurons, where they form heterocomplexes with other proteins, has profoundly changed our view of neurotransmitter signaling. Here, we review new insights into neurotransmitter crosstalk at the plasma membrane. We focus on the membrane organization and interactome of the ionotropic glutamate N-methyl-D-aspartate receptor (NMDAR) that plays a central role in excitatory synaptic and network physiology and is involved in the etiology of several major neuropsychiatric disorders. The nanoscale organization and dynamics of NMDAR is a key regulatory process for glutamate synapse transmission, plasticity, and crosstalk with other neurotransmitter systems, such as the monoaminergic ones. The plasma membrane appears to be a prime regulatory compartment for spatial and temporal crosstalk between neurotransmitter systems in the healthy and diseased brain. Understanding the molecular mechanisms regulating membrane neurotransmitter receptor crosstalk will likely open research avenues for innovative therapeutical strategies.
Introduction
The N-methyl-D-aspartate receptors (NMDARs) are ionotropic glutamate receptors whose activation and calcium permeability play central roles in glutamate synapse physiology. Consequently, dysfunctions of NMDAR signaling are associated with neurological and psychiatric diseases (Paoletti et al., 2013; Wyllie et al., 2013). Several decades of research have provided valuable insight into how the NMDARs are trafficked to glutamatergic synapses, how they are regulated by neuronal activity or modulatory systems, and how they become dysfunctional in neuropsychiatric disorders. Schematically, NMDAR activation is essential for numerous brain cell communication processes, including synaptogenesis, long-term synaptic plasticity (e.g., NMDAR-dependent long-term synaptic potentiation [LTP]), dendritic integration, learning, and memory (Collingridge et al., 2004; Lau et al., 2009; Hansen et al., 2018). The NMDAR activation requires (a) a depolarization of the membrane to remove the magnesium block and (b) the presence of both agonist (glutamate) and coagonist (glycine or D-serine), making this receptor a coincident detector at the basis of Hebbian-type synaptic plasticity processes (Collingridge et al., 2004). Besides the effect of NMDAR-induced depolarization, the NMDAR permeability to calcium triggers calcium-dependent signaling cascades that regulate functional and structural plasticity of synapses. These calcium-dependent processes play key roles in the maturation of synapses, structural plasticity of synapses and dendritic spines, and many other key plasticity processes (Collingridge et al., 2004; Lau et al., 2009; Paoletti et al., 2013; Hansen et al., 2018). During NMDAR-dependent LTP, it has for instance been shown that calcium entry from NMDAR activates Ca2+/calmodulin-dependent protein kinase II (CaMKII), which phosphorylates and favors the trafficking of α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors (AMPARs) to the synapses (Herring and Nicoll, 2016).
NMDAR subunit composition determines its location and function. NMDARs are heterotetramers composed of two obligatory GluN1 subunits that associate with two GluN2 (A–D) or GluN3 (A and B) subunits (Paoletti et al., 2013; Wyllie et al., 2013). The GluN2 subunits determine most NMDAR biophysical properties, including agonist affinity, open probability, and deactivation kinetics (Paoletti et al., 2013; Wyllie et al., 2013). Whereas the GluN1 subunit is widely distributed over the nervous system, the different GluN2 subunits show regional distributions that change during development and aging (Paoletti et al., 2013). The NMDARs are expressed not only by neurons but also by glial and endothelium cells in the brain, as well as by cells from the immune system, kidney, heart, and pancreas (Leboyer et al., 2016), suggesting that the understanding of NMDAR function in brain cell networks is a complex challenge. Exocytosis, endocytosis, and local dendritic synthesis control most of the membrane pool of NMDARs, including their delivery to synaptic areas (Trepanier et al., 2012; Sanz-Clemente et al., 2013b; Horak et al., 2014; Vieira et al., 2020). Once inserted in the plasma membrane through exocytosis, NMDARs explore large segments of the dendrite through lateral diffusion (Tovar and Westbrook, 2002; Groc et al., 2004, 2006; McQuate and Barria, 2020). Diffusing NMDARs eventually enter synapses, in which they interact with a myriad of scaffold proteins from the postsynaptic density (PSD) or trans-synaptic complexes (Frank and Grant, 2017).
Besides the canonical role of NMDARs in mediating long-term synaptic plasticity through their calcium permeability, non-ionotropic functions of the receptor have also been well described (Gray et al., 2016; Dore et al., 2016, 2017). Among these, the specific role of the NMDAR surface dynamics in tuning synaptic plasticity and associative memory highlight the essential role of receptor movement, organization, and direct interactions with other proteins (Bard and Groc, 2011; Ladépêche et al., 2014; Groc and Choquet, 2020). Yet our mechanistic understanding of the crosstalk between neurotransmitter systems is far from complete. Besides well-identified intracellular signaling cascades that are often shared between systems, direct interactions between neuronal receptors occur within the plasma membrane and play major roles in network physiology, behavioral adaptations, and even neuropsychiatric disorders. Here, we review our current understanding of two fundamental processes that shape the organization and functions of neuronal receptors: receptor lateral diffusion and receptor–receptor interaction at the plasma membrane, focusing on the NMDAR.
Nanoscale organization and surface dynamics of NMDAR
More than a third of surface NMDARs are located extrasynaptically, the rest being concentrated within the postsynaptic compartment (Harris and Pettit, 2007; Petralia et al., 2010; McQuate and Barria, 2020; Tovar and Westbrook, 1999). Although the activation of synaptic NMDARs leads to survival-promoting mechanisms and synaptic adaptations, the activation of extrasynaptic NMDARs preferentially leads to mitochondrial dysfunction, loss of integrity of neuronal structures, neurotoxicity, and cell death (Hardingham and Bading, 2010). The mechanism behind these distinct fates is likely the activation of different intracellular pathways triggered by calcium influx (Bading, 2013; Hardingham, 2019). The surface distribution of NMDARs results from the equilibrium between synaptic and extrasynaptic receptor exchange. The surface dynamics through Brownian diffusion of a given receptor is solely powered by thermal agitation. Yet NMDAR surface organization within these compartments is not homogeneous, indicating the presence of local regulatory mechanisms (Groc and Choquet, 2020). Advances in superresolution microscopy show that in the postsynaptic compartment, the nanoscale organization of neurotransmitter receptors, scaffolds, and signaling molecules is well structured (Frost et al., 2010; MacGillavry et al., 2013; Nair et al., 2013; Broadhead et al., 2016). Nanoscale surface mapping of NMDAR subtypes revealed a small number (5–10) of nanodomains (∼55 nm wide) per synaptic area (MacGillavry et al., 2013; Kellermayer et al., 2018). The presence of synaptic nanodomains for most glutamate receptors and scaffold proteins in hippocampal synapses strengthens the view that the postsynapse is a highly compartmentalized entity. The nanoscale compartmentalization of NMDARs plays a role in postsynaptic transmission and synaptic plasticity (Zeng et al., 2016; Kellermayer et al., 2018). It has been proposed that the postsynaptic organization in nanodomains better aligns the receptors to presynaptic release sites (Tang et al., 2016). Within the NMDAR family, GluN2A and GluN2B subunits have differential and largely nonoverlapping nanoscale organization in synapses from hippocampal neurons, indicating the presence of specific sorting mechanisms for different NMDAR subtypes. Although much less explored, extrasynaptic NMDARs are also structured in nanodomains, forming a star-like mesh of small nanodomains that outnumber the synaptic ones (Fig. 1 A; Kellermayer et al., 2018). Yet the mechanism governing the nanodomain organization of extrasynaptic NMDARs remains mostly unknown. While in synapses, intracellular scaffold proteins from the PSD mainly contribute to the nanodomain organization, other putative mechanisms involving transmembrane interaction and/or extracellular matrix are likely to be organizers for extrasynaptic NMDARs.
Figure 1.

Surface distribution and dynamics of NMDAR. (A) Live immunostaining of surface GluN2B-NMDAR in days in vitro 15 hippocampal neurons followed by dSTORM imaging. SR-Tesseler segmented clusters are represented. NMDARs can be observed to form nanodomains within glutamatergic synapses and in the extrasynaptic (Extrasyn.) compartment (black arrow heads). Scale bars = 30 nm. (B) Schematic representation of NMDAR lateral diffusion along neuronal dendrites. A given NMDAR will be anchored within synaptic and extrasynaptic NMDAR nanodomains. (C) NMDAR surface dynamics is regulated (Δ) by intracellular modulators (PDZ scaffold proteins at the postsynaptic compartment) and by extracellular modulators (NMDAR coagonists, D-serine and glycine, matrix metalloproteases, tissue plasminogen, and steroid hormones). Glut., Glutamate.
The surface dynamics of a single NMDAR at the neuronal plasma membrane classically exhibits distinct behaviors: (a) strong confined diffusion and anchoring of receptors within PSD nanodomains, (b) high diffusion with low confinement in the extrasynaptic membrane, and (c) confined diffusion within small extrasynaptic nanodomains (Fig. 1 B; Groc et al., 2009; Groc and Choquet, 2020). In parallel to the observation that GluN2A- and GluN2B-NMDARs form different nanodomains, their surface dynamics are also different (Groc et al., 2006; Bard et al., 2010). During sustained synaptic activity in immature neurons, GluN2B-NMDARS, but not GluN2A-NMDARs, exhibit a high degree of exchange between synaptic and extrasynaptic compartments, redistributing CaMKII and favoring LTP (Groc et al., 2004; Bellone and Nicoll, 2007; Dupuis et al., 2014). Within synapses, PDZ domain–containing membrane-associated guanylate kinase proteins, such as PSD95, serve as scaffolding proteins that anchor NMDARs as well as other transmembrane proteins (Sheng and Sala, 2001; Nair et al., 2013; Chen et al., 2015; Zhu et al., 2016). Interactions of NMDARs with other intracellular partners are also important to regulate their organization, such as direct interaction with CaMKII. Indeed, activated CaMKII interacts with the C-terminus of the GluN2B subunit and regulates receptor function (Halt et al., 2012; Incontro et al., 2018; Barcomb et al., 2016) and surface trafficking (Sanz-Clemente et al., 2013a; Dupuis et al., 2014). Note that CaMKIIα is recruited to the PSD in an activity-dependent manner during LTP, whereas CaMKIIβ stabilizes the actin cytoskeleton at the PSD, acting as scaffolding protein for NMDAR (Incontro et al., 2018).
The extracellular environment is also a major regulator of NMDAR surface dynamics, particularly for extrasynaptic receptors. Extracellular NMDAR surface dynamics is regulated by components of the extracellular matrix such as matrix metalloproteinase 9, tissue plasminogen activator, stress hormone (corticosterone), and sex hormone (estrogen; Groc et al., 2008; Michaluk et al., 2009; Lesept et al., 2016; Potier et al., 2016; Mikasova et al., 2017). In addition, the NMDAR coagonists, glycine and D-serine, differentially regulate the surface dynamics of both synaptic and extrasynaptic NMDARs (Fig. 1 C; Papouin and Oliet, 2014; Ferreira et al., 2017). For instance, D-serine alters the surface dynamics and synaptic content of GluNB-NMDAR, but not GluN2A-NMDAR, through interaction between its C-terminus and PDZ-binding scaffold proteins (Ferreira et al., 2017). Other, yet unknown, molecules of the extracellular environment could also be potent regulators of NMDAR dynamics, e.g., polyamines, protons, zinc, or neurosteroids, as they all contribute to NMDAR-mediated synaptic regulation (Paoletti et al., 2013; Hansen et al., 2018). Thus, the nanoscale distribution and surface dynamics of synaptic and extrasynaptic NMDARs are regulated by multiple well-defined and yet still unknown mechanisms. The latter is particularly true for extrasynaptic NMDARs that form small nanodomains whose molecular composition and regulation remain enigmatic.
Protein–protein interactions at the plasma membrane: Detection methods
The term interactome describes the molecular interaction networks in a living organism (Sanchez et al., 1999). Interactions can occur between transmembrane proteins from opposed cells (trans-interactions) or between transmembrane proteins embedded within the same membrane (cis-interactions). Transient interactions associate and dissociate rapidly and can be divided into strong or weak based on their properties (Iacobucci et al., 2021). Weak transient interactions associate and dissociate continuously (e.g., high dissociation rate), like most receptor–receptor interactions (Perkins et al., 2010; Acuner Ozbabacan et al., 2011). The characterization of these interactions in their native environment has been challenging compared with strong and high-affinity interactions that can be captured by most available biochemical approaches (Xing et al., 2016; Titeca et al., 2019; Portillo et al., 2020). Different methods are available to investigate the interaction of hydrophobic proteins, such as membrane proteins (Snider et al., 2015). Although these techniques have brought to light a series of new interactions, their respective advantages and disadvantages support the claim that different approaches should be used to validate membrane receptor interaction. Several methods based on different strategies exist and provide a powerful toolbox to cell biologists. In addition to these methods, the criteria defined for the well-studied G protein–coupled receptor (GPCR) heterometers could be used to define membrane complexes: (a) heteromer components should colocalize in the same compartment and physically interact, (b) heteromers should exhibit properties distinct from those of the protomers, and (c) heteromer disruption should lead to a loss of heteromer-specific properties (Gomes et al., 2016). Depending on whether the interactors are known/unknown or form putative strong/weak interaction, the choice of complementary methods will be essential. Yet still today, there is a relative lack of methods to study protein–protein interaction within the plasma membrane in a live and native environment with both high spatial (interaction of spots that are eventually at low density) and temporal (short time of interaction) resolution. Future technological developments are needed to define a receptor membrane interactome, which can be composed of dozens of different proteins, with high spatial and temporal resolutions, in a given biological tissue.
Current methods to decipher protein–protein interaction in cells
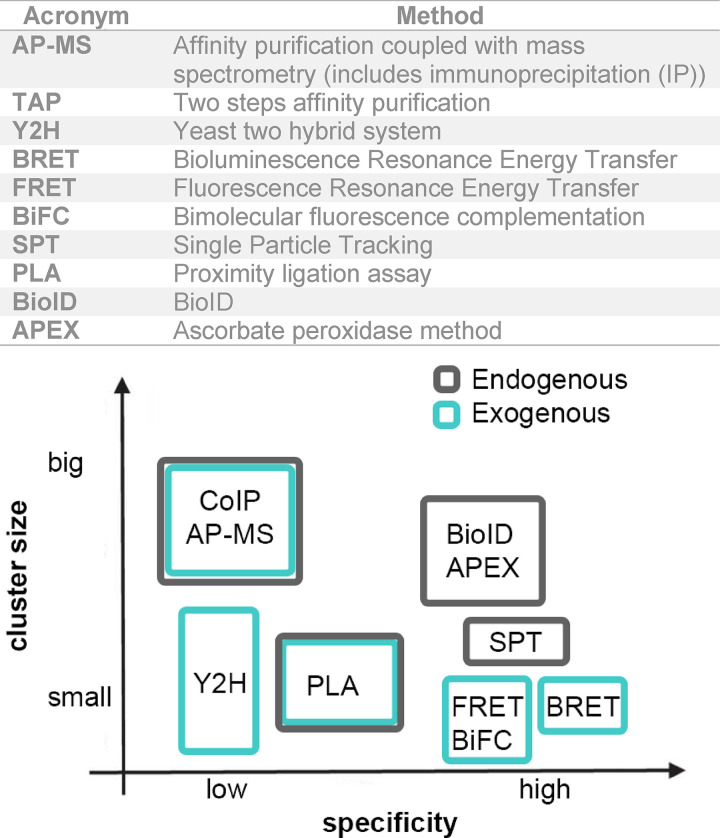
The relative specificity of the methods relates here to their capacities to minimize the production of false-positive or -negative outcomes. The size of the putative cluster formed by interacting proteins can range from small (tens of nanometers) to larger (hundreds of nanometers to submicrons) entities.
The NMDAR surface cis-interactome: Current view
To define the NMDAR membrane interactome, we have classified the amount of the evidence of interaction for NMDAR interactors based on the number of different techniques used to unveil specific heterodimer (Fig. 2 and Table 1). Note that part of the synaptic NMDAR interactome with intracellular proteins from the PSD has been successfully characterized using immunoprecipitation and mass spectrometry (Frank and Grant, 2017). The NMDAR involvement in major neurological and psychiatric disorders has made the mapping of its protein interactions a longstanding goal in neurophysiology and drug therapy. The cycling of NMDARs between intracellular pool and the plasma membrane relies on protein interactions and phosphorylation processes that have been well characterized (Trepanier et al., 2012; Sanz-Clemente et al., 2013b; Horak et al., 2014; Vieira et al., 2020). Yet our understanding of the NMDAR membrane cis-interactome and its spatiotemporal regulation in live neurons is far less complete. Here, we review the experimentally based evidence of cis-interactions between NMDAR and neurotransmitter receptors or ion channels at the neuronal plasma membrane and discuss their main physiological functions (Fig. 2).
Figure 2.

Known neurotransmitter receptor and ion channels cis-interactors of the NMDAR at the neuronal membrane. Described cis-interactions that NMDARs can establish at the neuronal cell surface and the multiple interactions within the NMDAR cell-surface interactome. Because methods to demonstrate protein–protein interactions can give false positives, the interaction will be more relevant if it has been demonstrated by several means. The different grade of evidence for its given NMDAR cis-interaction has been represented by thick dark red lines, indicating that the interaction has been proven by three or more techniques, middle-thickness red lines, indicating two techniques, and thin light pink lines, indicating one technique. Note that GluN1 (nine interactions), mGluR5 (six), D1R (five), µOR (five), Sig1R (five), mGluR1 (four), and EphB2 (four) are the most connected receptors in this macrocomplex.
Table 1. Evidence of NMDAR interactors.
| Interaction | Method | Sources |
|---|---|---|
| D1R-GluN1 | CoIP of rat brain tissue and hippocampal cultures | Lee et al., 2002; Fiorentini et al., 2003 |
| BRET in COS cells expressing the subunits | Fiorentini et al., 2003 | |
| CoIP using rat hippocampal extract | Pei et al., 2004 | |
| CoIP after disruption of interaction using peptide in mouse striatum neurons and slices | Cahill et al., 2014 | |
| SPT in rat hippocampal neurons | Ladepeche et al., 2013a | |
| D1R-GluN2A | CoIP of rat brain tissue, hippocampal cultures, and transfected heterologous cells | Lee et al., 2002; Fiorentini et al., 2003 |
| D2R-GluN2B | CoIP of GST-fused subunits using striatal extracts | Liu et al., 2006 |
| mGluR1-GluN1 | BRET in transfected HEK293 cells | Perroy et al., 2008 |
| mGluR1-GluN2A | CoIP with rat brain hippocampal lysates | Lai et al., 2019 |
| mGluR5-GluN1 | BRET in transfected HEK293 cells | Perroy et al., 2008 |
| SPT in rat hippocampal neurons | Aloisi et al., 2017 | |
| EphB2-GluN1 | CoIP of transfected HEK293 cells | Dalva et al., 2000 |
| CoIP of brain lysates; cultured neurons | Nolt et al., 2011; Hanamura et al., 2017 | |
| SPT in rat hippocampal neurons | Mikasova et al., 2012 | |
| ICC in synaptosomes from mouse brain (colocalization) and PLA | Washburn et al., 2020 | |
| µOR-GluN1 | CoIP assays with mice brain synaptosomes; BiFC approach in live transfected CHO cells | Rodríguez-Muñoz et al., 2012 |
| ApoE-GluN1 | CoIP of transfected COS7 cells and CoIP of mouse brain lysates | Hoe et al., 2006 |
| BKα R–GluN1 | CoIP of rodent brain | Zhang et al., 2018 |
| α7 nAchR-GluN2A | IP of rat hippocampal homogenate | Li et al., 2012 |
| Sig1R-GluN1 | CoIP, AFM, PLA | Balasuriya et al., 2013 |
| NL1–GluN1 | PLA in COS7 cells | Budreck et al., 2013 |
| H3R-GluN1, H3R-GluN2A | CoIP, PLA, BRET, BiFC in heterologous cells, and/or rodent brain tissue | Rodríguez-Ruiz et al., 2017 |
| IL1R-GluN1, IL1R-GluN2B | CoIP of rat hippocampi | Gardoni et al., 2011 |
| P2X-GluN1 | Electrophysiology experiments on exogenously expressed receptors in oocytes | Rodriguez et al., 2020 |
| TRPM4-GluN2A, TRPM4-GluN2B | CoIP of rodent brain tissue | Yan et al., 2020 |
CoIP, coimmunoprecipitation; BRET, bioluminescence resonance energy transfer; COS cells, CV-1 (simian) in Origin, and carrying the SV40 cells; SPT, single particle tracking; HEK293 cells, human embryonic kidney 293 cells; ICC, immunocytochemestry; PLA, proximity ligase assay; BiFC, bimolecular fluorescence complementation; AFM, atomic force microscopy.
Interaction with dopamine receptors
Dopamine receptors are GPCRs involved in numerous brain functions such as motor activity, emotion, cognition, and reward. Of the five types of dopamine receptors, only D1R and D2R directly interact with NMDARs at the neuronal membrane (Wang et al., 2012). Coimmunoprecipitation of fusion proteins expressed in heterologous cells and bioluminescence resonance energy transfer (BRET) experiments demonstrated that D1R interacts with GluN1 and GluN2A subunits but not with the GluN2B subunit (Lee et al., 2002; Fiorentini et al., 2003). Imaging approaches further confirmed that GluN1-NMDAR and D1R interact at the plasma membrane of live hippocampal neurons (Scott et al., 2006; Ladepeche et al., 2013b; Gréa et al., 2019). NMDAR and D1R interact via their intracellular C-terminal tails (Lee et al., 2002; Pei et al., 2004; Fiorentini et al., 2003; Cahill et al., 2014). The D1R-NMDAR interaction favors the retention of NMDARs at extrasynaptic location, increases D1R insertion into the cell membrane, and inhibits NMDAR currents and NMDAR-mediated excitotoxicity through a phosphatidylinositol 3-kinase–dependent pathway (Lee et al., 2002; Pei et al., 2004; Ladepeche et al., 2013b). Activation of D1R promotes the phosphorylation of GluN1 subunit through the recruitment of cAMP/PKA signaling, Fyn-related pathways, or alternatively via the activation of the PKC/Src cascades (Trepanier et al., 2012). As a result, D1R-NMDAR interaction is disrupted, which increases NMDAR surface dynamics and synaptic content and promotes NMDA-dependent LTP and working memory (Nai et al., 2010; Ladepeche et al., 2013b). Note that different, sometimes opposite, regulatory processes have been defined for NMDAR-D1R interactions in striatal neurons (Cahill et al., 2014). Heterocomplexes also exist between D2R and GluN2B-NMDAR (Liu et al., 2006). This interaction, enhanced by dopamine stimulation (e.g., cocaine), prevents the binding of CaMKII to GluN2B-NMDAR, leading to GluN2B subunit dephosphorylation and a decrease in NMDAR signaling (Liu et al., 2006). Therefore, NMDARs and dopamine receptors can interact at different loci whose characteristics and fine molecular regulation remain still to be fully uncovered.
Interaction with metabotropic glutamate receptors (mGluRs)
The mGluRs are GPCRs that participate in the modulation of glutamatergic synaptic transmission and neuronal excitability throughout the central nervous system (CNS). Of the eight mGluRs, only mGluR1 and mGluR5 directly interact with NMDARs. The direct interaction between mGluR5 and GluN1 subunits was first demonstrated using BRET in HEK293 cells expressing both receptors (Perroy et al., 2008). The interaction regulates their respective trafficking and inhibits receptor activity (Perroy et al., 2008). Single-particle tracking (SPT) experiments further demonstrated that GluN1-NMDAR and mGluR5 colocalize at the surface of neurons, especially in synapses and their vicinities (Aloisi et al., 2017). Coimmunoprecipitation studies using rat brain hippocampal lysates also showed that mGluR1 and GluN2A subunits interact (Lai et al., 2019). NMDAR and mGluR1/5 also have a fast and indirect interplay via Homer proteins (O’Neill et al., 2018; Sylantyev et al., 2013). As NMDARs and mGluR1/5s are both glutamate receptors, their interaction is likely a key regulator of glutamate synapse physiology and pathology (Bockaert et al., 2010).
Interaction with opioid receptors
µ-Opioid receptors (µORs) are inhibitory GPCRs responsible for analgesia and mediate rewarding and positive reinforcement, a critical initial step leading to opioid addiction, after activation (Cuitavi et al., 2020). Coimmunoprecipitation experiments with mice brain synaptosomes identified that µORs form a heterocomplex with the GluN1 subunit through their C-terminus but do not interact with GluN2A or GluN2B subunit (Rodríguez-Muñoz et al., 2012). This interaction was observed in midbrain periaqueductal gray cells, cerebral cortex, striatum, and dorsal spinal cord. Application of the opioid morphine above analgesic doses disrupts the NMDAR–µOR complex and favors GluN1 C-terminal phosphorylation through PKC. Inhibition of PKC strengthens the NMDAR–µOR complex, restoring analgesic effects of morphine (Rodríguez-Muñoz et al., 2012; Garzón et al., 2012).
Interaction with Ephrin B2 receptor
Ephrin receptors are divided in two families: EphA (EphA1–A9) and EphB (EphB1–B6R) receptors, and generally mediate cell–cell communication by interacting with surface-associated ligands (ephrins) on neighboring cells. EphB2Rs are tyrosine kinases crucial for migration, repulsion, and adhesion during neuronal, vascular, and epithelial development. Direct cis-interaction between the extracellular domains of EphB2R and GluN1 subunit was demonstrated by coimmunoprecipitation (Dalva et al., 2000; Washburn et al., 2020), and later confirmed with SPT (Mikasova et al., 2012) and proximity ligase assay (PLA) experiments (Washburn et al., 2020). NMDAR–EphB2R heterocomplex formation increases NMDAR-dependent Ca2+ influx and synaptic retention and modulates NMDAR channel function and phosphorylation of GluN2B subunits (Dalva et al., 2000; Nolt et al., 2011; Mikasova et al., 2012). Coimmunoprecipitation experiments revealed that the GluN1 subunit interacts with other Eph receptors, such as EphB1R, EphB3R, and EphB4R (Dalva et al., 2000; Calò et al., 2005). During development, the EphBR-NMDAR association plays an important role in synaptogenesis and regulates synaptic plasticity at later stages (Grunwald et al., 2001; Hruska and Dalva, 2012). EphB2R thus appears, among other signaling functions, to anchor and stabilize NMDARs within synapses through cis-interactions.
Interaction with ApoE receptor
Apolipoprotein E receptor 2 (ApoER2) is a member of the low-density lipoprotein receptor family, involved in neural signaling, neurogenesis, neuroinflammation, cholesterol metabolism, and trafficking of cholesterol and other lipids in the plasma and the CNS (Holtzman et al., 2012; Lee and D’Arcangelo, 2016; Zhao et al., 2018; Jossin, 2020). Activation of ApoER2 with reelin modulates glutamatergic transmission by enhancing NMDAR-mediated synaptic transmission, stabilizing GluN2B-NMDAR at synapses, and favoring NMDAR-dependent LTP in hippocampal neurons (Groc et al., 2007; Qiu et al., 2006). ApoER2 and GluN1-NMDAR have been found to interact through their extracellular domains, as demonstrated by coimmunoprecipitation experiments (Hoe et al., 2006). ApoER2 also interacts with PSD95, potentially forming a bridge with other GluN2 subunits (Hoe et al., 2006).
Interaction with nicotinic receptors
Nicotinic acetylcholine receptors (nAChRs) are major signaling molecules in both the central and peripheral nervous system that bind to the neurotransmitter acetylcholine and drugs such as the agonist nicotine (Koukouli and Maskos, 2015). There are 17 subtypes of nAchR, divided into muscle-type and neuronal-type subunits. The neuronal α7 nAchRs, which are highly expressed in the hippocampus, physically interact with GluN1 and GluN2A subunits, forming a multimeric complex that modulates NMDAR function (Li et al., 2012; Zhang et al., 2016). Nicotine promotes α7 nAchR-NMDAR interaction, up-regulates NMDAR currents, and favors synaptic NMDAR-dependent LTP (Bürli et al., 2010; Li et al., 2013). NMDAR-α7 nAchR complex formation leads to changes in extracellular signal–regulated kinase activity, suggesting that it is part of the downstream pathway associated with the formation of this heterocomplex (Li et al., 2012). In mice, disruption of the interaction causes memory and learning deficits (Li et al., 2013).
Interaction with neuroligin
Neuroligin 1 (NL1) is a postsynaptic cell adhesion molecule that binds to presynaptic β-neurexins to mediate the formation, maintenance, and plasticity of glutamate synapses (Shipman and Nicoll, 2012; Chamma et al., 2020). Neuroligin mutations have been associated with developmental disorders and autism (Etherton et al., 2011; Bemben et al., 2015). Neuroligins, present at inhibitory and excitatory synapses, have a key role in the formation of synapses by recruiting PSD95 and NMDAR at the postsynaptic site (Bang and Owczarek, 2013) and are required for the induction of NMDAR-dependent LTP (Shipman and Nicoll, 2012; Wu et al., 2019). NL1 interacts with the GluN1 subunit and regulates the abundance of NMDARs at postsynaptic sites (Budreck et al., 2013). Using chimeric NL1/NL2 proteins, it was demonstrated that the extracellular domain of NL1 is required to recruit GluN1-NMDAR at the postsynaptic terminals (Budreck et al., 2013). The overexpression of NL1 promotes the clustering of synaptic NMDARs in cultured neurons (Chih, 2005). Thus, NL1 anchors and stabilizes NMDARs within synapses through cis-interaction.
Interaction with voltage-activated potassium (BK) channels
The large conductance, calcium-activated, and BK channels are channels with a voltage sensor formed by four pore-forming subunits, which repolarize the membrane potential in response to depolarization by allowing an outward flow of potassium. BK channels are distributed in the whole brain, and they regulate membrane excitability, neurotransmitter release, and calcium signaling (Contet et al., 2016). Coimmunoprecipitation studies unveiled that BK channels and NMDARs form complexes via direct interactions with their obligatory subunits, BKα and GluN1 subunit (Zhang et al., 2018). NMDARs activate BK channels by Ca2+ influx, which mediates rapid membrane repolarization through potassium efflux (Ghatta et al., 2006). Disruption of the BK-NMDAR interaction using a specific competing peptide mimicking BKα intracellular region prevents NMDAR-induced BK channel activation and increased glutamate synapse transmission (Zhang et al., 2018).
Interaction with Sigma-1 receptor (Sig1R)
Sig1R is a transmembrane protein widely distributed in the CNS that regulates neurotransmitter release, learning, and memory (Maurice and Su, 2009; Bolshakova et al., 2016). Sig1R regulates the inositol-3-phosphate receptor and modulates calcium signaling in ER and mitochondria. Although Sig1R is mainly an ER protein, it has been proposed to translocate to the plasma membrane (Ryskamp et al., 2019), where it interacts with NMDARs. The interaction between Sig1Rs and NMDARs was demonstrated using different methods: protein coimmunoprecipitation, atomic force microscopy (AFM) imaging, and PLA (Balasuriya et al., 2013). Sig1R directly interacts with GluN1, but not GluN2A, subunits through their extracellular domains. The role of this interaction at the plasma membrane, however, remains to be defined in neurons.
Interaction with histamine 3 receptor (H3R)
H3R is a member of the GPCR superfamily widely expressed in the CNS, particularly in the frontal cortex and hippocampus (Pillot et al., 2002). H3Rs are located at the presynaptic and postsynaptic membrane, where they regulate the synthesis and release of histamine (Panula et al., 2015). Not only do they function as a presynaptic autoreceptor by inhibiting histamine release in the brain, but they also regulate the release of other neurotransmitters such as acetylcholine, noradrenaline, serotonin (5-HT), and dopamine (Esbenshade et al., 2008). It has been demonstrated that H3Rs act as allosteric regulators of NMDARs and D1Rs (Moreno et al., 2011; Rodríguez-Ruiz et al., 2017). Several approaches to study protein–protein interactions such as coimmunoprecipitation, PLA, BRET, and bimolecular fluorescence complementation (BiFC) in heterologous cells or rodent brain tissue have demonstrated that H3Rs directly interact with GluN1 and GluN2A subunits, and also with D1Rs, forming an heteromeric complex (Rodríguez-Ruiz et al., 2017). Antagonists of H3Rs reduce the excitotoxicity of NMDAR. Acting on this heterocomplex has thus been suggested as a target for neurodegenerative diseases (Grove et al., 2014; Moreno-Delgado et al., 2020). H3Rs form heterocomplexes with D1Rs and D2Rs that modulate dopaminergic neurotransmission and play a putative role in drug addiction (Ellenbroek, 2013).
Interaction with IL-1 receptor (IL-1R)
IL-1R is a cytokine receptor activated by the cytokine IL1 that promotes proinflammatory processes (Boraschi et al., 2018). Coimmunoprecipitation using homogenates from rat hippocampus demonstrated that IL1R is expressed at the dendrites, is enriched at the postsynaptic compartment, and interacts with GluN1 and GluN2B subunits. NMDAR activation increases IL-1R interaction with NMDARs, as well as the surface expression and localization of IL-1R at synaptic membranes, by favoring the membrane insertion of new IL-1R (Gardoni et al., 2011). IL-1R activation activates tyrosine kinases that trigger GluN2B subunit phosphorylation, promoting the formation of IL-1R–NMDAR complexes and synaptic localization (Viviani et al., 2003; Johansson et al., 2020).
Interaction with purinergic P2X receptors
Purinergic P2X receptors are ATP-gated, calcium-permeable, nonselective cation channels (seven subunits: P2X1–7) implicated in the modulation of synaptic transmission, neuroinflammation, pain, neurological dysfunctions, vascular tone, cardiac rhythm and contractility, and immune response (Khakh and North, 2012; Burnstock, 2018). They are usually located at the PSDs of excitatory neurons. Binding of ATP results in channel opening, cation influx, and membrane depolarization (Coddou et al., 2011). Among the P2X receptor members, P2X4 has been found to play a role in the induction of NMDAR-dependent LTP via the modulation of NMDAR calcium influx (Sim et al., 2006; Pankratov et al., 2009). In the hippocampus, the activation of postsynaptic P2X2 or P2X4 triggers surface AMPAR internalization, leading to P2X-mediated long-term synaptic depression (Pougnet et al., 2014, 2016). P2X receptors further modulate the activity of GABAa receptors (Boué-Grabot et al., 2004; Toulmé et al., 2007; Jo et al., 2011), nACh receptors (Khakh et al., 2000), and 5-hydroxytryptamine receptor 3A (Boué-Grabot et al., 2003; Emerit et al., 2016; Soto et al., 2020). Coexpression of P2X receptors and NMDARs in oocytes showed that they interact via the C-terminal domain of NMDAR, and their activation inhibits NMDAR-dependent currents (Rodriguez et al., 2020).
Interaction with transient receptor potential cation channel subfamily M member 4 (TRPM4)
TRPM4, also known as melastatin-4, is a cation channel that mediates cell depolarization. TRPM4 provides a mechanism for cells to depolarize in a Ca2+-dependent manner and shape action potential duration and spiking frequency (Launay et al., 2002; Nilius et al., 2003). TRPM4 has been identified as a key component of NMDAR-mediated cell death signaling. Indeed, TRPM4 is not expressed in synapses but in extrasynaptic membranes, where NMDAR activation has been related to cell death. Coimmunoprecipitation experiments using lysates from cultured mouse hippocampal neurons and brain lysates from mouse hippocampus and cortex have demonstrated that GluN2A and GluN2B subunits, but not GluN1 subunit, interact with TRPM4. While interaction between TRPM4 and extrasynaptic NMDAR triggers mitochondrial dysfunction and cell death, and inhibits cAMP response element–binding protein activation and early gene expression, disruption of this interaction has a neuroprotective effect against NMDAR-mediated toxicity (Yan et al., 2020).
NMDAR surface interactome: Regulation and dynamics
The above multiple known cis-interactions between NMDARs and neurotransmitter receptors, ion channels, and adhesion proteins suggest that NMDAR is centrally positioned to integrate various key signals in brain circuits. Indeed, receptors from monoaminergic, opioid, and cell-adhesion molecule signaling directly modulate NMDAR signaling through protein–protein interactions. In the plasma membrane of neurons, neurotransmitter receptors and ion channels laterally diffuse and explore large somatic and dendritic membrane domains. This is not specific for neurons, as neurotransmitter receptors and transporters also exhibit high diffusion at the surface of astrocytes (Ciappelloni et al., 2017; Renner and Triller, 2020). NMDARs and their interacting receptors and ion channels are often expressed in the same neuronal types and dendritic compartments. The lateral diffusion of all these interacting membrane proteins is likely to be modulated by their random interactions (Fig. 3). An approximate comparison of the lateral diffusion characteristics (e.g., diffusion coefficients) of NMDAR and its receptor interactome demonstrates that NMDAR diffusion is relatively slower (Fig. 3), eventually reflecting a role as central integrator. At the synapse, NMDARs are structurally organized in nanodomains, and direct interactions between NMDARs and EphB2Rs or NL1Rs are important determinants of NMDAR synaptic anchoring. In the perisynaptic and extrasynaptic compartments, NMDARs likely associate with other interacting receptors allowing modulatory processes. For instance, interaction of NMDARs with dopamine receptors forms an extracellular pool of “reserve” NMDARs (Scott et al., 2002; Ladepeche et al., 2013b). Upon dopamine release, the NMDAR–dopamine receptor interaction is disrupted in hippocampal neurons, inducing a lateral redistribution of NMDARs that favors their recruitment at synapse and NMDAR-dependent LTP (Liu et al., 2000; Ladepeche et al., 2013a). Although the exact molecular mechanism underpinning the regulation of such protein–protein interaction remain unclear, a change in the conformational structure of the receptors that impacts their interaction is a likely scenario. In support, the activation of NMDARs alters the intracellular C-terminus conformation that impairs direct interaction with PDZ scaffold proteins (Ferreira et al., 2017). Whether such a molecular scenario applies to the other members of the NMDAR cis-interactome remains an open question.
Figure 3.

NMDAR surface dynamics are highly regulated by the crosstalk between NMDAR surface interactors. NMDAR surface interactors show different diffusion rates at the cell surface, undergoing Brownian diffusion until they interact to form nanodomains (upper panel). Neurotransmitters, such as glutamate (Glut.), opioid, acetylcholine, and dopamine (Dop.), released by projecting neurons, and neuronal activity modulate the diffusion of the NMDAR and its interactors, creating nanoclusters at different neuronal compartments (lower panel). syn., synthesis.
Distribution of these putative interactions is likely organized and strategically positioned by agonists such as dopamine, acetylcholine, and opioid (Fig. 3). In volume transmission, i.e., long-distance communication between brain cells, the extracellular space may regulate the NMDAR cis-interactome not only locally but more globally and tonically at the scale of brain region areas (Borroto-Escuela et al., 2015). Investigations are needed to define the spatial (short versus long distance) and temporal (phasic versus tonic release) regulation of the NMDAR cis-interactome. Different mechanisms likely coexist to tune short- and long-term neuronal integration. In hippocampal neurons, the extrasynaptic compartment contains a myriad of small NMDAR nanodomains, i.e., NMDAR interactome “hubs,” that can be mobilized on demand by the extracellular interacting receptor agonists (e.g., glutamate, dopamine, acetylcholine, and opioids) through fast lateral diffusion. The advantages for such interplay are that receptor lateral diffusion (a) is purely based on thermodynamics and does not directly require cell energy, (b) is rapidly translated (millisecond range), (c) does integrate multiple biophysical characteristics besides interacting receptor agonists (e.g., membrane lipid composition, viscosity, and temperature), (d) is sufficiently sensitive to weak interactions, and (e) easily translocates receptor complexes to different nanoscale subcompartments. Developing new imaging approaches with high spatiotemporal resolution in live neurons will likely shed new unsuspected light on the regulation of the NMDAR interactome and its physiological role in brain circuits. Furthermore, one could envision that drugs designed to alter NMDAR signaling will have to take into account the interactome to better control excitatory-modulatory balance in brain disorders.
Putative role of the NMDAR membrane interactome in brain diseases
Human genetic and postmortem brain studies, as well as animal models, have unveiled that dysfunctions of NMDAR signaling are associated with the etiology of several major neurological and psychiatric disorders, such as Alzheimer’s, Huntington’s, and Parkinson’s diseases (Wenk, 2006; Bonuccelli and Del Dotto, 2006; Lipton, 2006; Fan and Raymond, 2007); schizophrenia (Kantrowitz and Javitt, 2012; Weinberger, 2019); depression (Maeng and Zarate, 2007); epilepsy; and ischemic stroke (Choi et al., 1988; Villmann and Becker, 2007). For instance, the pharmacological blockade of NMDAR with phencyclidine or ketamine recapitulates a full range of cognitive, positive, and negative symptoms of schizophrenia. In addition, psychomimetic drugs that block NMDAR transmission or genetic mutations of the receptor produce major deficits in synaptic plasticity, neuronal networks, and cognitive functions. In developmental models of schizophrenia, alterations of NMDAR functions early in development perturb, among others, cell proliferation and migration and synaptogenesis and generate psychotic-like behavior at adult stage (Millan et al., 2016). Thus, dysfunction of NMDAR signaling constitutes today one of the main biological hypotheses at the basis of psychotic disorders. Yet antipsychotic treatments, developed in the 1950s, primarily target the dopaminergic receptors, and new pharmacological drugs targeting the NMDAR have failed to meet therapeutical end points (Miyamoto et al., 2005; Girgis et al., 2019). In addition to these two receptor types, several other neurotransmitter receptors and ion channels have been associated with the emergence of psychotic disorders, including voltage-gated calcium channels, α7 nAchR, mGluR, opioid receptors, EphB2R, Sig1R, and neurexin-neuroligin (Heyes et al., 2015; Young and Geyer, 2013; Nicoletti et al., 2019; Ashok et al., 2019; Zhang et al., 2010; Ishiguro et al., 1998; Südhof, 2008). Strikingly, a large fraction of these neurotransmitter receptors belong to the NMDAR interactome (Fig. 4 A). One may thus hypothesize that the implication of all these neurotransmitter receptors is somehow related to their physical interaction and constant crosstalk with the NMDAR. Theoretically, a sustained alteration of one of these receptors (e.g., NMDAR) will reverberate on its membrane partner signaling. The current antipsychotic drugs, which mainly target dopamine and monoamine receptors, may thus restore balance in the whole NMDAR interactome.
Figure 4.

Altered NMDAR interactome in pathological conditions. (A) A high interconnection within the NMDAR interactome is established. The majority of the components of the macrocomplex have been linked to neuropsychiatric disorders (e.g., schizophrenia; indicated in gray rectangles), suggesting that disorders altering any of these interactions can potentially alter the whole interactome. (B) NMDAR interaction with dopamine receptor and EphrinB2 receptor is impaired by patients’ NMDAR autoantibodies (left). For most of the NMDAR-interacting proteins, whether autoantibodies alter their interaction, location, or function is unknown, although a domino effect is likely (right).
Is this proposal purely speculative? In other words, does any experimental evidence support the claim that specific disruption of the NMDAR membrane organization, related to psychosis in humans, alters NMDAR partners in the interactome hub? The discovery of anti-NMDAR encephalitis, an autoimmune disorder of the brain mediated by the presence of autoantibodies in the serum and cerebrospinal fluid that target the extracellular domain of the NMDAR (NMDAR-Abs), has been of prime importance in the field. The initial clinical presentation usually includes prodromal headache or fever, followed by psychiatric manifestations that may include anxiety, insomnia, delusional thinking, hallucinations, paranoid thoughts, pressured speech, mood disorder, or aggressive behavior, with episodes of agitation and catatonia. At this stage, the symptoms suggest drug abuse or an acute psychotic break, and a psychiatric consultation is often considered. However, the disease usually progresses to a more severe stage including seizures, reduced verbal output, decreased level of consciousness, highly characteristic orofacial and limb dyskinesias, dystonic postures, rigidity, and autonomic dysfunction (with tachycardia, high blood pressure, hyperthermia, profuse salivation, and hypoventilation), suggesting a disorder other than primary psychiatric disease (Dalmau et al., 2011, 2017; Titulaer et al., 2013). All these symptoms, including the psychotic ones, are only due to the presence of the NMDAR-Abs, providing a direct link between the presence of a given molecule and the various symptoms. At the molecular and cellular levels, the NMDAR-Abs target the GluN1 subunit and cause a massive surface reorganization of the receptor. Indeed, synaptic NMDARs are laterally displaced to the extrasynaptic compartment, since NMDAR-Abs prevent interaction between NMDAR and EphB2R (Mikasova et al., 2012). Then, NMDAR-Abs decrease the content of extrasynaptic NMDARs over time through a cross-linking effect that likely favors receptor internalization; the effects are reversed 4 d after the removal of NMDAR-Abs (Hughes et al., 2010; Mikasova et al., 2012). Of note, NMDAR-Abs do not alter dendritic branching, dendritic spine density, or number of glutamatergic synapses (e.g., density of PSD95 or Bassoon), indicating that the autoantibodies have a specific effect on NMDAR trafficking and organization.
In mice, the chronic intraventricular infusion of NMDAR-Abs recapitulates the in vitro effects, i.e., decreases total surface and synaptic NMDAR clusters and induces behavioral alterations (Planagumà et al., 2015). The key question is then whether such a change in NMDAR surface dynamics impacts other interactome receptors. Neither the surface dynamics of the glutamatergic AMPA and GABAa receptors nor that of Kv1.3 potassium channels were affected by NMDAR-Abs (Mikasova et al., 2012). However, sustained exposure of hippocampal neurons to NMDAR-Abs significantly altered the surface dynamics of D1R (Gréa et al., 2019). Altered D1R trafficking was observed mainly in the extrasynaptic compartment, where D1Rs interact with NMDARs. Truncating the D1R C-terminal domain, which is required for direct interaction with NMDARs, prevented the effect of NMDAR-Abs, indicating that the disorganization of surface NMDARs by NMDAR-Abs reverberates to the interacting dopamine receptors. Consistently, the intraventricular injection of NMDAR-Abs, which cause memory deficits and psychotic-like behaviors, altered the content of surface dopamine receptors (D1R and D2R; Carceles-Cordon et al., 2020). Besides the dopamine receptors, NMDAR-Abs also disrupt the direct interaction between synaptic NMDAR and EphB2R, leading to the rapid dispersal of both receptors outside synapses. Thus, an antibody targeting one identified receptor causes at least two collateral damage effects, disorganization of the dopaminergic and ephrin signaling (Mikasova et al., 2012; Planagumà et al., 2016). We propose thus a “domino effect” hypothesis whereby altered membrane trafficking of NMDARs will mechanically translate to nearby interactors, altering not only NMDAR signaling but that of all the ones mediated by receptors in the NMDAR interactome. It would thus be of great interest to test the domino effect theory on other members of the NMDAR interactome, such as neuroligin and mGluR (Fig. 4 B). The development of new approaches to identify interactions and their signaling effects in live brain tissue are certainly necessary to uncover the rules underpinning dysfunction of the NMDAR interactome in neuropsychiatric conditions. It could be predicted that the presence of NMDAR autoantibodies, for instance, will corrupt the NMDAR interactome, and an unbalance in glutamatergic-dopaminergic-cholinergic-opioid-ephrin-neuroligin signaling.
Conclusion and perspectives
NMDARs form protein–protein interactions with several transmembrane neurotransmitter receptors and ion channels at the surface of neurons, in which they all actively diffuse and explore large dendritic areas. Besides the fundamental physiological processes that have been related to these cis-interactions, the formation of these membrane hubs are the first regulatory level of this interplay. The size, distribution, and duration of the NMDAR interactome is currently unknown, calling for in-depth investigations. These interactions are likely rather transient, allowing fast remodeling and ensuring a great sensitivity to changes in extracellular agonist levels. Measuring with high time resolution (i.e., milliseconds) the interaction between membrane receptors using innovative imaging approaches will surely shed light on this question. Furthermore, elucidating the rules governing the NMDAR interactome may also shed unsuspected light on the etiology of major neurological and psychiatric disorders. Autoimmune brain disorders in which a given autoantibody targets a membrane neurotransmitter receptor constitute great investigation tools to dissect the primary (i.e., direct target) and secondary (i.e., multiple associated receptor signaling) effects and their translation to clinical symptoms. The use of heterobivalent drugs, designed to bind to two different membrane receptors simultaneously or not, has been suggested for the treatment of drug abuse with molecules targeting adenosine A2A receptor–D2R heterodimers (Borroto-Escuela et al., 2016). Innovative therapeutical approaches to counterbalance the receptor interactome (e.g., favor stabilization) rather than acting with agonists/antagonists, as traditionally envisioned, could thus revolutionize our treatment of brain diseases.
Acknowledgments
We thank J. Ferreira for providing the NMDAR STORM image.
This research was supported by the Centre National de la Recherche Scientifique, Agence Nationale de la Recherche (L. Groc, ANR DopamineHub, 19-CE16-0003-04), Fondation pour la Recherche Médicale (M. Petit-Pedrol, SPF201909009269), Human Frontier Science Program (L. Groc, RGP0019/2016), Conseil Régional d'Aquitaine, and Labex Bordeaux BRAIN.
The authors declare no competing financial interests.
References
- Acuner Ozbabacan S.E., Engin H.B., Gursoy A., and Keskin O.. 2011. Transient protein-protein interactions. Protein Eng. Des. Sel. 24:635–648. 10.1093/protein/gzr025 [DOI] [PubMed] [Google Scholar]
- Aloisi E., Le Corf K., Dupuis J., Zhang P., Ginger M., Labrousse V., Spatuzza M., Georg Haberl M., Costa L., Shigemoto R., et al. . 2017. Altered surface mGluR5 dynamics provoke synaptic NMDAR dysfunction and cognitive defects in Fmr1 knockout mice. Nat. Commun. 8:1103 10.1038/s41467-017-01191-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashok A.H., Myers J., Reis Marques T., Rabiner E.A., and Howes O.D.. 2019. Reduced mu opioid receptor availability in schizophrenia revealed with [11C]-carfentanil positron emission tomographic Imaging. Nat. Commun. 10:4493 10.1038/s41467-019-12366-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bading H. 2013. Nuclear calcium signalling in the regulation of brain function. Nat. Rev. Neurosci. 14:593–608. 10.1038/nrn3531 [DOI] [PubMed] [Google Scholar]
- Balasuriya D., Stewart A.P., and Edwardson J.M.. 2013. The σ-1 receptor interacts directly with GluN1 but not GluN2A in the GluN1/GluN2A NMDA receptor. J. Neurosci. 33:18219–18224. 10.1523/JNEUROSCI.3360-13.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bang M.L., and Owczarek S.. 2013. A matter of balance: role of neurexin and neuroligin at the synapse. Neurochem. Res. 38:1174–1189. 10.1007/s11064-013-1029-9 [DOI] [PubMed] [Google Scholar]
- Barcomb K., Hell J.W., Benke T.A., and Bayer K.U.. 2016. The CaMKII/GluN2B protein interaction maintains synaptic strength. J. Biol. Chem. 291:16082–16089. 10.1074/jbc.M116.734822 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bard L., and Groc L.. 2011. Glutamate receptor dynamics and protein interaction: lessons from the NMDA receptor. Mol. Cell. Neurosci. 48:298–307. 10.1016/j.mcn.2011.05.009 [DOI] [PubMed] [Google Scholar]
- Bard L., Sainlos M., Bouchet D., Cousins S., Mikasova L., Breillat C., Stephenson F.A., Imperiali B., Choquet D., and Groc L.. 2010. Dynamic and specific interaction between synaptic NR2-NMDA receptor and PDZ proteins. Proc. Natl. Acad. Sci. USA. 107:19561–19566. 10.1073/pnas.1002690107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bellone C., and Nicoll R.A.. 2007. Rapid bidirectional switching of synaptic NMDA receptors. Neuron. 55:779–785. 10.1016/j.neuron.2007.07.035 [DOI] [PubMed] [Google Scholar]
- Bemben M.A., Nguyen Q.A., Wang T., Li Y., Nicoll R.A., and Roche K.W.. 2015. Autism-associated mutation inhibits protein kinase C-mediated neuroligin-4X enhancement of excitatory synapses. Proc. Natl. Acad. Sci. USA. 112:2551–2556. 10.1073/pnas.1500501112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bockaert J., Perroy J., Bécamel C., Marin P., and Fagni L.. 2010. GPCR interacting proteins (GIPs) in the nervous system: Roles in physiology and pathologies. Annu. Rev. Pharmacol. Toxicol. 50:89–109. 10.1146/annurev.pharmtox.010909.105705 [DOI] [PubMed] [Google Scholar]
- Bolshakova A.V., Kukanova E.O., Gainullina A.N., Zhemkov V.A., Korban S.A., and Bezprozvanny I.B.. 2016. Sigma-1 receptor as a potential pharmacological target for the treatment of neuropathology. St. Petersbg. Polytech. Univ. J. Phys. Math. 2:31–40. 10.1016/j.spjpm.2016.03.003 [DOI] [Google Scholar]
- Bonuccelli U., and Del Dotto P.. 2006. New pharmacologic horizons in the treatment of Parkinson disease. Neurology. 67(7, Suppl 2):S30–S38. 10.1212/WNL.67.7_suppl_2.S30 [DOI] [PubMed] [Google Scholar]
- Boraschi D., Italiani P., Weil S., and Martin M.U.. 2018. The family of the interleukin-1 receptors. Immunol. Rev. 281:197–232. 10.1111/imr.12606 [DOI] [PubMed] [Google Scholar]
- Borroto-Escuela D.O., Agnati L.F., Bechter K., Jansson A., Tarakanov A.O., and Fuxe K.. 2015. The role of transmitter diffusion and flow versus extracellular vesicles in volume transmission in the brain neural-glial networks. Philos. Trans. R. Soc. Lond. B Biol. Sci. 370:1–14. 10.1098/rstb.2014.0183 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borroto-Escuela D.O., Pintsuk J., Schäfer T., Friedland K., Ferraro L., Tanganelli S., Liu F., and Fuxe K.. 2016. Multiple D2 heteroreceptor complexes: new targets for treatment of schizophrenia. Ther. Adv. Psychopharmacol. 6:77–94. 10.1177/2045125316637570 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boué-Grabot E., Barajas-López C., Chakfe Y., Blais D., Bélanger D., Émerit M.B., and Séguéla P.. 2003. Intracellular cross talk and physical interaction between two classes of neurotransmitter-gated channels. J. Neurosci. 23:1246–1253. 10.1523/JNEUROSCI.23-04-01246.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boué-Grabot E., Toulmé E., Émerit M.B., and Garret M.. 2004. Subunit-specific coupling between γ-aminobutyric acid type A and P2X2 receptor channels. J. Biol. Chem. 279:52517–52525. 10.1074/jbc.M410223200 [DOI] [PubMed] [Google Scholar]
- Broadhead M.J., Horrocks M.H., Zhu F., Muresan L., Benavides-Piccione R., DeFelipe J., Fricker D., Kopanitsa M.V., Duncan R.R., Klenerman D., et al. . 2016. PSD95 nanoclusters are postsynaptic building blocks in hippocampus circuits. Sci. Rep. 6:24626 10.1038/srep24626 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Budreck E.C., Kwon O.B., Jung J.H., Baudouin S., Thommen A., Kim H.S., Fukazawa Y., Harada H., Tabuchi K., Shigemoto R., et al. . 2013. Neuroligin-1 controls synaptic abundance of NMDA-type glutamate receptors through extracellular coupling. Proc. Natl. Acad. Sci. USA. 110:725–730. 10.1073/pnas.1214718110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bürli T., Baer K., Ewers H., Sidler C., Fuhrer C., and Fritschy J.-M.. 2010. Single particle tracking of α7 nicotinic AChR in hippocampal neurons reveals regulated confinement at glutamatergic and GABAergic perisynaptic sites. PLoS One. 5:e11507 10.1371/journal.pone.0011507 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnstock G. 2018. Purine and purinergic receptors. Brain Neurosci. Adv. 2:2398212818817494 10.1177/2398212818817494 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cahill E., Pascoli V., Trifilieff P., Savoldi D., Kappès V., Lüscher C., Caboche J., and Vanhoutte P.. 2014. D1R/GluN1 complexes in the striatum integrate dopamine and glutamate signalling to control synaptic plasticity and cocaine-induced responses. Mol. Psychiatry. 19:1295–1304. 10.1038/mp.2014.73 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calò L., Bruno V., Spinsanti P., Molinari G., Korkhov V., Esposito Z., Patanè M., Melchiorri D., Freissmuth M., and Nicoletti F.. 2005. Interactions between ephrin-B and metabotropic glutamate 1 receptors in brain tissue and cultured neurons. J. Neurosci. 25:2245–2254. 10.1523/JNEUROSCI.4956-04.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carceles-Cordon M., Mannara F., Aguilar E., Castellanos A., Planagumà J., and Dalmau J.. 2020. NMDAR antibodies alter dopamine receptors and cause psychotic behavior in mice. Ann. Neurol. 88:603–613. 10.1002/ana.25829 [DOI] [PubMed] [Google Scholar]
- Chamma I., Sainlos M., and Thoumine O.. 2020. Biophysical mechanisms underlying the membrane trafficking of synaptic adhesion molecules. Neuropharmacology. 169:107555 10.1016/j.neuropharm.2019.02.037 [DOI] [PubMed] [Google Scholar]
- Chen X., Levy J.M., Hou A., Winters C., Azzam R., Sousa A.A., Leapman R.D., Nicoll R.A., and Reese T.S.. 2015. PSD-95 family MAGUKs are essential for anchoring AMPA and NMDA receptor complexes at the postsynaptic density. Proc. Natl. Acad. Sci. USA. 112:E6983–E6992. 10.1073/pnas.1517045112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chih B. 2005. Control of Excitatory and Inhibitory Synapse Formation by Neuroligins. Science. 307:1324–1328. 10.1126/science.1107470 [DOI] [PubMed] [Google Scholar]
- Choi D.W., Koh J.Y., and Peters S.. 1988. Pharmacology of glutamate neurotoxicity in cortical cell culture: attenuation by NMDA antagonists. J. Neurosci. 8:185–196. 10.1523/JNEUROSCI.08-01-00185.1988 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ciappelloni S., Murphy-Royal C., Dupuis J.P., Oliet S.H.R., and Groc L.. 2017. Dynamics of surface neurotransmitter receptors and transporters in glial cells: Single molecule insights. Cell Calcium. 67:46–52. 10.1016/j.ceca.2017.08.009 [DOI] [PubMed] [Google Scholar]
- Coddou C., Yan Z., Obsil T., Huidobro-Toro J.P., and Stojilkovic S.S.. 2011. Activation and regulation of purinergic P2X receptor channels. Pharmacol. Rev. 63:641–683. 10.1124/pr.110.003129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collingridge G.L., Isaac J.T.R., and Wang Y.T.. 2004. Receptor trafficking and synaptic plasticity. Nat. Rev. Neurosci. 5:952–962. 10.1038/nrn1556 [DOI] [PubMed] [Google Scholar]
- Contet C., Goulding S.P., Kuljis D.A., and Barth A.L.. 2016. BK Channels in the Central Nervous System. Int. Rev. Neurobiol. 128:281–342. 10.1016/bs.irn.2016.04.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cuitavi J., Hipólito L., and Canals M.. 2020. The Life Cycle of the Mu-Opioid Receptor. Trends Biochem. Sci. 10.1016/j.tibs.2020.10.002 [DOI] [PubMed] [Google Scholar]
- Dalmau J., Geis C., and Graus F.. 2017. Autoantibodies to synaptic receptors and neuronal cell surface proteins in autoimmune diseases of the central nervous system. Physiol. Rev. 97:839–887. 10.1152/physrev.00010.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dalmau J., Lancaster E., Martinez-Hernandez E., Rosenfeld M.R., and Balice-Gordon R.. 2011. Clinical experience and laboratory investigations in patients with anti-NMDAR encephalitis. Lancet Neurol. 10:63–74. 10.1016/S1474-4422(10)70253-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dalva M.B., Takasu M.A., Lin M.Z., Shamah S.M., Hu L., Gale N.W., and Greenberg M.E.. 2000. EphB receptors interact with NMDA receptors and regulate excitatory synapse formation. Cell. 103:945–956. 10.1016/S0092-8674(00)00197-5 [DOI] [PubMed] [Google Scholar]
- Dore K., Aow J., and Malinow R.. 2016. The emergence of NMDA receptor metabotropic function: Insights from imaging. Front. Synaptic Neurosci. 8:20 10.3389/fnsyn.2016.00020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dore K., Stein I.S., Brock J.A., Castillo P.E., Zito K., and Sjöström P.J.. 2017. Unconventional NMDA receptor signaling. J. Neurosci. 37:10800–10807. 10.1523/JNEUROSCI.1825-17.2017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dupuis J.P., Ladépêche L., Seth H., Bard L., Varela J., Mikasova L., Bouchet D., Rogemond V., Honnorat J., Hanse E., and Groc L.. 2014. Surface dynamics of GluN2B-NMDA receptors controls plasticity of maturing glutamate synapses. EMBO J. 33:842–861. 10.1002/embj.201386356 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellenbroek B.A. 2013. Histamine H3 receptors, the complex interaction with dopamine and its implications for addiction. Br. J. Pharmacol. 170:46–57. 10.1111/bph.12221 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Emerit M.B., Baranowski C., Diaz J., Martinez A., Areias J., Alterio J., Masson J., Boué-Grabot E., and Darmon M.. 2016. A new mechanism of receptor targeting by interaction between two classes of ligand-gated ion channels. J. Neurosci. 36:1456–1470. 10.1523/JNEUROSCI.2390-15.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Esbenshade T.A., Browman K.E., Bitner R.S., Strakhova M., Cowart M.D., and Brioni J.D.. 2008. The histamine H3 receptor: an attractive target for the treatment of cognitive disorders. Br. J. Pharmacol. 154:1166–1181. 10.1038/bjp.2008.147 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Etherton M., Földy C., Sharma M., Tabuchi K., Liu X., Shamloo M., Malenka R.C., and Südhof T.C.. 2011. Autism-linked neuroligin-3 R451C mutation differentially alters hippocampal and cortical synaptic function. Proc. Natl. Acad. Sci. USA. 108:13764–13769. 10.1073/pnas.1111093108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fan M.M.Y., and Raymond L.A.. 2007. N-methyl-D-aspartate (NMDA) receptor function and excitotoxicity in Huntington’s disease. Prog. Neurobiol. 81:272–293. 10.1016/j.pneurobio.2006.11.003 [DOI] [PubMed] [Google Scholar]
- Ferreira J.S., Papouin T., Ladépêche L., Yao A., Langlais V.C., Bouchet D., Dulong J., Mothet J.-P., Sacchi S., Pollegioni L., et al. . 2017. Co-agonists differentially tune GluN2B-NMDA receptor trafficking at hippocampal synapses. eLife. 6:e25492 10.7554/eLife.25492 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fiorentini C., Gardoni F., Spano P., Di Luca M., and Missale C.. 2003. Regulation of dopamine D1 receptor trafficking and desensitization by oligomerization with glutamate N-methyl-D-aspartate receptors. J. Biol. Chem. 278:20196–20202. 10.1074/jbc.M213140200 [DOI] [PubMed] [Google Scholar]
- Frank R.A., and Grant S.G.. 2017. Supramolecular organization of NMDA receptors and the postsynaptic density. Curr. Opin. Neurobiol. 45:139–147. 10.1016/j.conb.2017.05.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frost N.A., Shroff H., Kong H., Betzig E., and Blanpied T.A.. 2010. Single-molecule discrimination of discrete perisynaptic and distributed sites of actin filament assembly within dendritic spines. Neuron. 67:86–99. 10.1016/j.neuron.2010.05.026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardoni F., Boraso M., Zianni E., Corsini E., Galli C.L., Cattabeni F., Marinovich M., Di Luca M., and Viviani B.. 2011. Distribution of interleukin-1 receptor complex at the synaptic membrane driven by interleukin-1β and NMDA stimulation. J. Neuroinflammation. 8:14 10.1186/1742-2094-8-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garzón J., Rodríguez-Muñoz M., and Sánchez-Blázquez P.. 2012. Direct association of Mu-opioid and NMDA glutamate receptors supports their cross-regulation: molecular implications for opioid tolerance. Curr. Drug Abuse Rev. 5:199–226. 10.2174/1874473711205030199 [DOI] [PubMed] [Google Scholar]
- Ghatta S., Nimmagadda D., Xu X., and O’Rourke S.T.. 2006. Large-conductance, calcium-activated potassium channels: structural and functional implications. Pharmacol. Ther. 110:103–116. 10.1016/j.pharmthera.2005.10.007 [DOI] [PubMed] [Google Scholar]
- Girgis R.R., Zoghbi A.W., Javitt D.C., and Lieberman J.A.. 2019. The past and future of novel, non-dopamine-2 receptor therapeutics for schizophrenia: A critical and comprehensive review. J. Psychiatr. Res. 108:57–83. 10.1016/j.jpsychires.2018.07.006 [DOI] [PubMed] [Google Scholar]
- Gomes I., Ayoub M.A., Fujita W., Jaeger W.C., Pfleger K.D.G., and Devi L.A.. 2016. G Protein-Coupled Receptor Heteromers. Annu. Rev. Pharmacol. Toxicol. 56:403–425. 10.1146/annurev-pharmtox-011613-135952 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gray J.A., Zito K., and Hell J.W.. 2016. Non-ionotropic signaling by the NMDA receptor: controversy and opportunity. F1000 Res. 5:1010 10.12688/f1000research.8366.1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gréa H., Bouchet D., Rogemond V., Hamdani N., Le Guen E., Tamouza R., Darrau E., Passerieux C., Honnorat J., Leboyer M., and Groc L.. 2019. Human Autoantibodies Against N-Methyl-D-Aspartate Receptor Modestly Alter Dopamine D1 Receptor Surface Dynamics. Front. Psychiatry. 10:670 10.3389/fpsyt.2019.00670 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Groc L., and Choquet D.. 2020. Linking glutamate receptor movements and synapse function. Science. 368:eaay4631. https://doi.org/. 10.1126/science.aay4631 [DOI] [PubMed] [Google Scholar]
- Groc L., Bard L., and Choquet D.. 2009. Surface trafficking of N-methyl-D-aspartate receptors: physiological and pathological perspectives. Neuroscience. 158:4–18. 10.1016/j.neuroscience.2008.05.029 [DOI] [PubMed] [Google Scholar]
- Groc L., Choquet D., Stephenson F.A., Verrier D., Manzoni O.J., and Chavis P.. 2007. NMDA receptor surface trafficking and synaptic subunit composition are developmentally regulated by the extracellular matrix protein Reelin. J. Neurosci. 27:10165–10175. 10.1523/JNEUROSCI.1772-07.2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Groc L., Choquet D., and Chaouloff F.. 2008. The stress hormone corticosterone conditions AMPAR surface trafficking and synaptic potentiation. Nat. Neurosci. 11:868–870. 10.1038/nn.2150 [DOI] [PubMed] [Google Scholar]
- Groc L., Heine M., Cognet L., Brickley K., Stephenson F.A., Lounis B., and Choquet D.. 2004. Differential activity-dependent regulation of the lateral mobilities of AMPA and NMDA receptors. Nat. Neurosci. 7:695–696. 10.1038/nn1270 [DOI] [PubMed] [Google Scholar]
- Groc L., Heine M., Cousins S.L., Stephenson F.A., Lounis B., Cognet L., and Choquet D.. 2006. NMDA receptor surface mobility depends on NR2A-2B subunits. Proc. Natl. Acad. Sci. USA. 103:18769–18774. 10.1073/pnas.0605238103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grove R.A., Harrington C.M., Mahler A., Beresford I., Maruff P., Lowy M.T., Nicholls A.P., Boardley R.L., Berges A.C., Nathan P.J., and Horrigan J.P.. 2014. A randomized, double-blind, placebo-controlled, 16-week study of the H3 receptor antagonist, GSK239512 as a monotherapy in subjects with mild-to-moderate Alzheimer’s disease. Curr. Alzheimer Res. 11:47–58. 10.2174/1567205010666131212110148 [DOI] [PubMed] [Google Scholar]
- Grunwald I.C., Korte M., Wolfer D., Wilkinson G.A., Unsicker K., Lipp H.P., Bonhoeffer T., and Klein R.. 2001. Kinase-independent requirement of EphB2 receptors in hippocampal synaptic plasticity. Neuron. 32:1027–1040. 10.1016/S0896-6273(01)00550-5 [DOI] [PubMed] [Google Scholar]
- Halt A.R., Dallapiazza R.F., Zhou Y., Stein I.S., Qian H., Juntti S., Wojcik S., Brose N., Silva A.J., and Hell J.W.. 2012. CaMKII binding to GluN2B is critical during memory consolidation. EMBO J. 31:1203–1216. 10.1038/emboj.2011.482 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanamura K., Washburn H.R., Sheffler-Collins S.I., Xia N.L., Henderson N., Tillu D.V., Hassler S., Spellman D.S., Zhang G., Neubert T.A., et al. . 2017. Extracellular phosphorylation of a receptor tyrosine kinase controls synaptic localization of NMDA receptors and regulates pathological pain. PLoS Biol. 15:e2002457 10.1371/journal.pbio.2002457 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hansen K.B., Yi F., Perszyk R.E., Furukawa H., Wollmuth L.P., Gibb A.J., and Traynelis S.F.. 2018. Structure, function, and allosteric modulation of NMDA receptors. J. Gen. Physiol. 150:1081–1105. 10.1085/jgp.201812032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hardingham G. 2019. NMDA receptor C-terminal signaling in development, plasticity, and disease. F1000 Res. 8:1547 10.12688/f1000research.19925.1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hardingham G.E., and Bading H.. 2010. Synaptic versus extrasynaptic NMDA receptor signalling: implications for neurodegenerative disorders. Nat. Rev. Neurosci. 11:682–696. 10.1038/nrn2911 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harris A.Z., and Pettit D.L.. 2007. Extrasynaptic and synaptic NMDA receptors form stable and uniform pools in rat hippocampal slices. J. Physiol. 584:509–519. 10.1113/jphysiol.2007.137679 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herring B.E., and Nicoll R.A.. 2016. Long-Term Potentiation: From CaMKII to AMPA Receptor Trafficking. Annu. Rev. Physiol. 78:351–365. 10.1146/annurev-physiol-021014-071753 [DOI] [PubMed] [Google Scholar]
- Heyes S., Pratt W.S., Rees E., Dahimene S., Ferron L., Owen M.J., and Dolphin A.C.. 2015. Genetic disruption of voltage-gated calcium channels in psychiatric and neurological disorders. Prog. Neurobiol. 134:36–54. 10.1016/j.pneurobio.2015.09.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoe H.S., Pocivavsek A., Chakraborty G., Fu Z., Vicini S., Ehlers M.D., and Rebeck G.W.. 2006. Apolipoprotein E receptor 2 interactions with the N-methyl-D-aspartate receptor. J. Biol. Chem. 281:3425–3431. 10.1074/jbc.M509380200 [DOI] [PubMed] [Google Scholar]
- Holtzman D.M., Herz J., and Bu G.. 2012. Apolipoprotein E and apolipoprotein E receptors: normal biology and roles in Alzheimer disease. Cold Spring Harb. Perspect. Med. 2:a006312 10.1101/cshperspect.a006312 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horak M., Petralia R.S., Kaniakova M., and Sans N.. 2014. ER to synapse trafficking of NMDA receptors. Front. Cell. Neurosci. 8:394 10.3389/fncel.2014.00394 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hruska M., and Dalva M.B.. 2012. Ephrin regulation of synapse formation, function and plasticity. Mol. Cell. Neurosci. 50:35–44. 10.1016/j.mcn.2012.03.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hughes E.G., Peng X., Gleichman A.J., Lai M., Zhou L., Tsou R., Parsons T.D., Lynch D.R., Dalmau J., and Balice-Gordon R.J.. 2010. Cellular and synaptic mechanisms of anti-NMDA receptor encephalitis. J. Neurosci. 30:5866–5875. 10.1523/JNEUROSCI.0167-10.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iacobucci I., Monaco V., Cozzolino F., and Monti M.. 2021. From classical to new generation approaches: An excursus of -omics methods for investigation of protein-protein interaction networks. J. Proteomics. 230:103990 10.1016/j.jprot.2020.103990 [DOI] [PubMed] [Google Scholar]
- Incontro S., Díaz-Alonso J., Iafrati J., Vieira M., Asensio C.S., Sohal V.S., Roche K.W., Bender K.J., and Nicoll R.A.. 2018. The CaMKII/NMDA receptor complex controls hippocampal synaptic transmission by kinase-dependent and independent mechanisms. Nat. Commun. 9:2069 10.1038/s41467-018-04439-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ishiguro H., Ohtsuki T., Toru M., Itokawa M., Aoki J., Shibuya H., Kurumaji A., Okubo Y., Iwawaki A., Ota K., et al. . 1998. Association between polymorphisms in the type 1 sigma receptor gene and schizophrenia. Neurosci. Lett. 257:45–48. 10.1016/S0304-3940(98)00797-6 [DOI] [PubMed] [Google Scholar]
- Jo Y.H., Donier E., Martinez A., Garret M., Toulmé E., and Boué-Grabot E.. 2011. Cross-talk between P2X4 and γ-aminobutyric acid, type A receptors determines synaptic efficacy at a central synapse. J. Biol. Chem. 286:19993–20004. 10.1074/jbc.M111.231324 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johansson E.M., Bouchet D., Tamouza R., Ellul P., Morr A.S., Avignone E., Germi R., Leboyer M., Perron H., and Groc L.. 2020. Human endogenous retroviral protein triggers deficit in glutamate synapse maturation and behaviors associated with psychosis. Sci. Adv. 6:eabc0708 10.1126/sciadv.abc0708 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jossin Y. 2020. Reelin Functions, Mechanisms of Action and Signaling Pathways During Brain Development and Maturation. Biomolecules. 10:964 10.3390/biom10060964 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kantrowitz J., and Javitt D.C.. 2012. Glutamatergic transmission in schizophrenia: from basic research to clinical practice. Curr. Opin. Psychiatry. 25:96–102. 10.1097/YCO.0b013e32835035b2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kellermayer B., Ferreira J.S., Dupuis J., Levet F., Grillo-Bosch D., Bard L., Linarès-Loyez J., Bouchet D., Choquet D., Rusakov D.A., et al. . 2018. Differential Nanoscale Topography and Functional Role of GluN2-NMDA Receptor Subtypes at Glutamatergic Synapses. Neuron. 100:106–119.e7. 10.1016/j.neuron.2018.09.012 [DOI] [PubMed] [Google Scholar]
- Khakh B.S., and North R.A.. 2012. Neuromodulation by extracellular ATP and P2X receptors in the CNS. Neuron. 76:51–69. 10.1016/j.neuron.2012.09.024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khakh B.S., Zhou X., Sydes J., Galligan J.J., and Lester H.A.. 2000. State-dependent cross-inhibition between transmitter-gated cation channels. Nature. 406:405–410. 10.1038/35019066 [DOI] [PubMed] [Google Scholar]
- Koukouli F., and Maskos U.. 2015. The multiple roles of the α7 nicotinic acetylcholine receptor in modulating glutamatergic systems in the normal and diseased nervous system. Biochem. Pharmacol. 97:378–387. 10.1016/j.bcp.2015.07.018 [DOI] [PubMed] [Google Scholar]
- Ladepeche L., Dupuis J.P., Bouchet D., Doudnikoff E., Yang L., Campagne Y., Bézard E., Hosy E., and Groc L.. 2013a Single-molecule imaging of the functional crosstalk between surface NMDA and dopamine D1 receptors. Proc. Natl. Acad. Sci. USA. 110:18005–18010. 10.1073/pnas.1310145110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ladépêche L., Dupuis J.P., and Groc L.. 2014. Surface trafficking of NMDA receptors: gathering from a partner to another. Semin. Cell Dev. Biol. 27:3–13. 10.1016/j.semcdb.2013.10.005 [DOI] [PubMed] [Google Scholar]
- Ladepeche L., Yang L., Bouchet D., and Groc L.. 2013b Regulation of dopamine D1 receptor dynamics within the postsynaptic density of hippocampal glutamate synapses. PLoS One. 8:e74512 10.1371/journal.pone.0074512 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lai T.K.Y., Zhai D., Su P., Jiang A., Boychuk J., and Liu F.. 2019. The receptor-receptor interaction between mGluR1 receptor and NMDA receptor: a potential therapeutic target for protection against ischemic stroke. FASEB J. 33:14423–14439. 10.1096/fj.201900417R [DOI] [PubMed] [Google Scholar]
- Lau C.G., Takeuchi K., Rodenas-Ruano A., Takayasu Y., Murphy J., Bennett M.V.I., and Zukin R.S.. 2009. Regulation of NMDA receptor Ca2+ signalling and synaptic plasticity. Biochem. Soc. Trans. 37:1369–1374. 10.1042/BST0371369 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Launay P., Fleig A., Perraud A.-L., Scharenberg A.M., Penner R., and Kinet J.-P.. 2002. TRPM4 is a Ca2+-activated nonselective cation channel mediating cell membrane depolarization. Cell. 109:397–407. 10.1016/S0092-8674(02)00719-5 [DOI] [PubMed] [Google Scholar]
- Leboyer M., Berk M., Yolken R.H., Tamouza R., Kupfer D., and Groc L.. 2016. Immuno-psychiatry: an agenda for clinical practice and innovative research. BMC Med. 14:173 10.1186/s12916-016-0712-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee F.J.S., Xue S., Pei L., Vukusic B., Chéry N., Wang Y., Wang Y.T., Niznik H.B., Yu X.M., and Liu F.. 2002. Dual regulation of NMDA receptor functions by direct protein-protein interactions with the dopamine D1 receptor. Cell. 111:219–230. 10.1016/S0092-8674(02)00962-5 [DOI] [PubMed] [Google Scholar]
- Lee G.H., and D’Arcangelo G.. 2016. New insights into reelin-mediated signaling pathways. Front. Cell. Neurosci. 10:122 10.3389/fncel.2016.00122 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lesept F., Chevilley A., Jezequel J., Ladépêche L., Macrez R., Aimable M., Lenoir S., Bertrand T., Rubrecht L., Galea P., et al. . 2016. Tissue-type plasminogen activator controls neuronal death by raising surface dynamics of extrasynaptic NMDA receptors. Cell Death Dis. 7:e2466 10.1038/cddis.2016.279 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S., Li Z., Pei L., Le A.D., and Liu F.. 2012. The α7nACh-NMDA receptor complex is involved in cue-induced reinstatement of nicotine seeking. J. Exp. Med. 209:2141–2147. 10.1084/jem.20121270 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S., Nai Q., Lipina T.V., Roder J.C., and Liu F.. 2013. α7nAchR/NMDAR coupling affects NMDAR function and object recognition. Mol. Brain. 6:58 10.1186/1756-6606-6-58 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lipton S.A. 2006. Paradigm shift in neuroprotection by NMDA receptor blockade: memantine and beyond. Nat. Rev. Drug Discov. 5:160–170. 10.1038/nrd1958 [DOI] [PubMed] [Google Scholar]
- Liu F., Wan Q., Pristupa Z.B., Yu X.M., Wang Y.T., and Niznik H.B.. 2000. Direct protein-protein coupling enables cross-talk between dopamine D5 and γ-aminobutyric acid A receptors. Nature. 403:274–280. 10.1038/35002014 [DOI] [PubMed] [Google Scholar]
- Liu X.Y., Chu X.P., Mao L.M., Wang M., Lan H.X., Li M.H., Zhang G.C., Parelkar N.K.K., Fibuch E.E.E., Haines M., et al. . 2006. Modulation of D2R-NR2B interactions in response to cocaine. Neuron. 52:897–909. 10.1016/j.neuron.2006.10.011 [DOI] [PubMed] [Google Scholar]
- MacGillavry H.D., Song Y., Raghavachari S., and Blanpied T.A.. 2013. Nanoscale scaffolding domains within the postsynaptic density concentrate synaptic AMPA receptors. Neuron. 78:615–622. 10.1016/j.neuron.2013.03.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maeng S., and Zarate C.A. Jr. 2007. The role of glutamate in mood disorders: results from the ketamine in major depression study and the presumed cellular mechanism underlying its antidepressant effects. Curr. Psychiatry Rep. 9:467–474. 10.1007/s11920-007-0063-1 [DOI] [PubMed] [Google Scholar]
- Maurice T., and Su T.P.. 2009. The pharmacology of sigma-1 receptors. Pharmacol. Ther. 124:195–206. 10.1016/j.pharmthera.2009.07.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McQuate A., and Barria A.. 2020. Rapid exchange of synaptic and extrasynaptic NMDA receptors in hippocampal CA1 neurons. J. Neurophysiol. 123:1004–1014. 10.1152/jn.00458.2019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michaluk P., Mikasova L., Groc L., Frischknecht R., Choquet D., and Kaczmarek L.. 2009. Matrix metalloproteinase-9 controls NMDA receptor surface diffusion through integrin β1 signaling. J. Neurosci. 29:6007–6012. 10.1523/JNEUROSCI.5346-08.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mikasova L., De Rossi P., Bouchet D., Georges F., Rogemond V., Didelot A., Meissirel C., Honnorat J., and Groc L.. 2012. Disrupted surface cross-talk between NMDA and Ephrin-B2 receptors in anti-NMDA encephalitis. Brain. 135:1606–1621. 10.1093/brain/aws092 [DOI] [PubMed] [Google Scholar]
- Mikasova L., Xiong H., Kerkhofs A., Bouchet D., Krugers H.J., and Groc L.. 2017. Stress hormone rapidly tunes synaptic NMDA receptor through membrane dynamics and mineralocorticoid signalling. Sci. Rep. 7:8053 10.1038/s41598-017-08695-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Millan M.J., Andrieux A., Bartzokis G., Cadenhead K., Dazzan P., Fusar-Poli P., Gallinat J., Giedd J., Grayson D.R., Heinrichs M., et al. . 2016. Altering the course of schizophrenia: progress and perspectives. Nat. Rev. Drug Discov. 15:485–515. 10.1038/nrd.2016.28 [DOI] [PubMed] [Google Scholar]
- Miyamoto S., Duncan G.E., Marx C.E., and Lieberman J.A.. 2005. Treatments for schizophrenia: a critical review of pharmacology and mechanisms of action of antipsychotic drugs. Mol. Psychiatry. 10:79–104. 10.1038/sj.mp.4001556 [DOI] [PubMed] [Google Scholar]
- Moreno E., Hoffmann H., Gonzalez-Sepúlveda M., Navarro G., Casadó V., Cortés A., Mallol J., Vignes M., McCormick P.J., Canela E.I., et al. . 2011. Dopamine D1-histamine H3 receptor heteromers provide a selective link to MAPK signaling in GABAergic neurons of the direct striatal pathway. J. Biol. Chem. 286:5846–5854. 10.1074/jbc.M110.161489 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moreno-Delgado D., Puigdellívol M., Moreno E., Rodríguez-Ruiz M., Botta J., Gasperini P., Chiarlone A., Howell L.A., Scarselli M., Casadó V., et al. . 2020. Modulation of dopamine D1 receptors via histamine H3 receptors is a novel therapeutic target for Huntington’s disease. eLife. 9:e51093 10.7554/eLife.51093 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nai Q., Li S., Wang S.H., Liu J., Lee F.J.S., Frankland P.W., and Liu F.. 2010. Uncoupling the D1-N-methyl-D-aspartate (NMDA) receptor complex promotes NMDA-dependent long-term potentiation and working memory. Biol. Psychiatry. 67:246–254. 10.1016/j.biopsych.2009.08.011 [DOI] [PubMed] [Google Scholar]
- Nair D., Hosy E., Petersen J.D., Constals A., Giannone G., Choquet D., and Sibarita J.B.. 2013. Super-resolution imaging reveals that AMPA receptors inside synapses are dynamically organized in nanodomains regulated by PSD95. J. Neurosci. 33:13204–13224. 10.1523/JNEUROSCI.2381-12.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicoletti F., Orlando R., Di Menna L., Cannella M., Notartomaso S., Mascio G., Iacovelli L., Matrisciano F., Fazio F., Caraci F., et al. . 2019. Targeting mGlu receptors for optimization of antipsychotic activity and disease-modifying effect in schizophrenia. Front. Psychiatry. 10:49 10.3389/fpsyt.2019.00049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nilius B., Prenen J., Droogmans G., Voets T., Vennekens R., Freichel M., Wissenbach U., and Flockerzi V.. 2003. Voltage dependence of the Ca2+-activated cation channel TRPM4. J. Biol. Chem. 278:30813–30820. 10.1074/jbc.M305127200 [DOI] [PubMed] [Google Scholar]
- Nolt M.J., Lin Y., Hruska M., Murphy J., Sheffler-Colins S.I., Kayser M.S., Passer J., Bennett M.V.L., Zukin R.S., and Dalva M.B.. 2011. EphB controls NMDA receptor function and synaptic targeting in a subunit-specific manner. J. Neurosci. 31:5353–5364. 10.1523/JNEUROSCI.0282-11.2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Neill N., McLaughlin C., Komiyama N., and Sylantyev S.. 2018. Biphasic modulation of NMDA receptor function by metabotropic glutamate receptors. J. Neurosci. 38:9840–9855. 10.1523/JNEUROSCI.1000-18.2018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pankratov Y., Lalo U., Krishtal O.A., and Verkhratsky A.. 2009. P2X receptors and synaptic plasticity. Neuroscience. 158:137–148. 10.1016/j.neuroscience.2008.03.076 [DOI] [PubMed] [Google Scholar]
- Panula P., Chazot P.L., Cowart M., Gutzmer R., Leurs R., Liu W.L.S., Stark H., Thurmond R.L., and Haas H.L.. 2015. International union of basic and clinical pharmacology. XCVIII. histamine receptors. Pharmacol. Rev. 67:601–655. 10.1124/pr.114.010249 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paoletti P., Bellone C., and Zhou Q.. 2013. NMDA receptor subunit diversity: impact on receptor properties, synaptic plasticity and disease. Nat. Rev. Neurosci. 14:383–400. 10.1038/nrn3504 [DOI] [PubMed] [Google Scholar]
- Papouin T., and Oliet S.H.R.. 2014. Organization, control and function of extrasynaptic NMDA receptors. Philos. Trans. R. Soc. Lond. B Biol. Sci. 369:20130601 10.1098/rstb.2013.0601 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pei L., Lee F.J.S.S., Moszczynska A., Vukusic B., and Liu F.. 2004. Regulation of dopamine D1 receptor function by physical interaction with the NMDA receptors. J. Neurosci. 24:1149–1158. 10.1523/JNEUROSCI.3922-03.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perkins J.R., Diboun I., Dessailly B.H., Lees J.G., and Orengo C.. 2010. Transient protein-protein interactions: structural, functional, and network properties. Structure. 18:1233–1243. 10.1016/j.str.2010.08.007 [DOI] [PubMed] [Google Scholar]
- Perroy J., Raynaud F., Homburger V., Rousset M.C., Telley L., Bockaert J., and Fagni L.. 2008. Direct interaction enables cross-talk between ionotropic and group I metabotropic glutamate receptors. J. Biol. Chem. 283:6799–6805. 10.1074/jbc.M705661200 [DOI] [PubMed] [Google Scholar]
- Petralia R.S., Wang Y.X., Hua F., Yi Z., Zhou A., Ge L., Stephenson F.A., and Wenthold R.J.. 2010. Organization of NMDA receptors at extrasynaptic locations. Neuroscience. 167:68–87. 10.1016/j.neuroscience.2010.01.022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pillot C., Heron A., Cochois V., Tardivel-Lacombe J., Ligneau X., Schwartz J.C., and Arrang J.M.. 2002. A detailed mapping of the histamine H(3) receptor and its gene transcripts in rat brain. Neuroscience. 114:173–193. 10.1016/S0306-4522(02)00135-5 [DOI] [PubMed] [Google Scholar]
- Planagumà J., Haselmann H., Mannara F., Petit-Pedrol M., Grünewald B., Aguilar E., Röpke L., Martín-García E., Titulaer M.J., Jercog P., et al. . 2016. Ephrin-B2 prevents N-methyl-D-aspartate receptor antibody effects on memory and neuroplasticity. Ann. Neurol. 80:388–400. 10.1002/ana.24721 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Planagumà J., Leypoldt F., Mannara F., Gutiérrez-Cuesta J., Martín-García E., Aguilar E., Titulaer M.J., Petit-Pedrol M., Jain A., Balice-Gordon R., et al. . 2015. Human N-methyl D-aspartate receptor antibodies alter memory and behaviour in mice. Brain. 138:94–109. 10.1093/brain/awu310 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Portillo F., Vázquez J., and Pajares M.A.. 2020. Protein-protein interactions involving enzymes of the mammalian methionine and homocysteine metabolism. Biochimie. 173:33–47. 10.1016/j.biochi.2020.02.015 [DOI] [PubMed] [Google Scholar]
- Potier M., Georges F., Brayda-Bruno L., Ladépêche L., Lamothe V., Al Abed A.S., Groc L., and Marighetto A.. 2016. Temporal Memory and Its Enhancement by Estradiol Requires Surface Dynamics of Hippocampal CA1 N-Methyl-D-Aspartate Receptors. Biol. Psychiatry. 79:735–745. 10.1016/j.biopsych.2015.07.017 [DOI] [PubMed] [Google Scholar]
- Pougnet J.T., Compans B., Martinez A., Choquet D., Hosy E., and Boué-Grabot E.. 2016. P2X-mediated AMPA receptor internalization and synaptic depression is controlled by two CaMKII phosphorylation sites on GluA1 in hippocampal neurons. Sci. Rep. 6:31836 10.1038/srep31836 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pougnet J.T., Toulme E., Martinez A., Choquet D., Hosy E., and Boué-Grabot E.. 2014. ATP P2X receptors downregulate AMPA receptor trafficking and postsynaptic efficacy in hippocampal neurons. Neuron. 83:417–430. 10.1016/j.neuron.2014.06.005 [DOI] [PubMed] [Google Scholar]
- Qiu S., Zhao L.F., Korwek K.M., and Weeber E.J.. 2006. Differential reelin-induced enhancement of NMDA and AMPA receptor activity in the adult hippocampus. J. Neurosci. 26:12943–12955. 10.1523/JNEUROSCI.2561-06.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Renner M., and Triller A.. 2020. Approaching Protein-Protein Interactions in Membranes Using Single-Particle Tracking and Packing Coefficient Analysis. In Neuromethods. Vol. 154. Single Molecule Microscopy in Neurobiology. Yamamoto N. and Okada Y., editors. Humana, New York: 115–129. 10.1007/978-1-0716-0532-5_6 [DOI] [Google Scholar]
- Rodriguez L., Yi C., Chu C., Duriez Q., Watanabe S., Ryu M., Reyes B., Asatryan L., Boué-Grabot E., and Davies D.. 2020. Cross-Talk between P2X and NMDA Receptors. Int. J. Mol. Sci. 21:1–20. 10.3390/ijms21197187 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodríguez-Muñoz M., Sánchez-Blázquez P., Vicente-Sánchez A., Berrocoso E., and Garzón J.. 2012. The mu-opioid receptor and the NMDA receptor associate in PAG neurons: implications in pain control. Neuropsychopharmacology. 37:338–349. 10.1038/npp.2011.155 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodríguez-Ruiz M., Moreno E., Moreno-Delgado D., Navarro G., Mallol J., Cortés A., Lluís C., Canela E.I., Casadó V., McCormick P.J., and Franco R.. 2017. Heteroreceptor Complexes Formed by Dopamine D1, Histamine H3, and N-Methyl-D-Aspartate Glutamate Receptors as Targets to Prevent Neuronal Death in Alzheimer’s Disease. Mol. Neurobiol. 54:4537–4550. 10.1007/s12035-016-9995-y [DOI] [PubMed] [Google Scholar]
- Ryskamp D.A., Korban S., Zhemkov V., Kraskovskaya N., and Bezprozvanny I.. 2019. Neuronal sigma-1 receptors: Signaling functions and protective roles in neurodegenerative diseases. Front. Neurosci. 13:862 10.3389/fnins.2019.00862 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanchez C., Lachaize C., Janody F., Bellon B., Röder L., Euzenat J., Rechenmann F., and Jacq B.. 1999. Grasping at molecular interactions and genetic networks in Drosophila melanogaster using FlyNets, an Internet database. Nucleic Acids Res. 27:89–94. 10.1093/nar/27.1.89 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanz-Clemente A., Gray J.A., Ogilvie K.A., Nicoll R.A., and Roche K.W.. 2013a Activated CaMKII couples GluN2B and casein kinase 2 to control synaptic NMDA receptors. Cell Rep. 3:607–614. 10.1016/j.celrep.2013.02.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanz-Clemente A., Nicoll R.A., and Roche K.W.. 2013b Diversity in NMDA receptor composition: many regulators, many consequences. Neuroscientist. 19:62–75. 10.1177/1073858411435129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scott L., Kruse M.S., Forssberg H., Brismar H., Greengard P., and Aperia A.. 2002. Selective up-regulation of dopamine D1 receptors in dendritic spines by NMDA receptor activation. Proc. Natl. Acad. Sci. USA. 99:1661–1664. 10.1073/pnas.032654599 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scott L., Zelenin S., Malmersjö S., Kowalewski J.M., Markus E.Z., Nairn A.C., Greengard P., Brismar H., and Aperia A.. 2006. Allosteric changes of the NMDA receptor trap diffusible dopamine 1 receptors in spines. Proc. Natl. Acad. Sci. USA. 103:762–767. 10.1073/pnas.0505557103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheng M., and Sala C.. 2001. PDZ domains and the organization of supramolecular complexes. Annu. Rev. Neurosci. 24:1–29. 10.1146/annurev.neuro.24.1.1 [DOI] [PubMed] [Google Scholar]
- Shipman S.L., and Nicoll R.A.. 2012. Dimerization of postsynaptic neuroligin drives synaptic assembly via transsynaptic clustering of neurexin. Proc. Natl. Acad. Sci. USA. 109:19432–19437. 10.1073/pnas.1217633109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sim J.A., Chaumont S., Jo J., Ulmann L., Young M.T., Cho K., Buell G., North R.A., and Rassendren F.. 2006. Altered hippocampal synaptic potentiation in P2X4 knock-out mice. J. Neurosci. 26:9006–9009. 10.1523/JNEUROSCI.2370-06.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snider J., Kotlyar M., Saraon P., Yao Z., Jurisica I., and Stagljar I.. 2015. Fundamentals of protein interaction network mapping. Mol. Syst. Biol. 11:848 10.15252/msb.20156351 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soto P., Gaete P.S., Fuentes C., Lozano B., Naulin P.A., Figueroa X.F., and Barrera N.P.. 2020. Function of P2X4 Receptors Is Directly Modulated by a 1:1 Stoichiometric Interaction With 5-HT3A Receptors. Front. Cell. Neurosci. 14:106 10.3389/fncel.2020.00106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Südhof T.C. 2008. Neuroligins and neurexins link synaptic function to cognitive disease. Nature. 455:903–911. 10.1038/nature07456 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sylantyev S., Savtchenko L.P., Ermolyuk Y., Michaluk P., and Rusakov D.A.. 2013. Spike-driven glutamate electrodiffusion triggers synaptic potentiation via a homer-dependent mGluR-NMDAR link. Neuron. 77:528–541. 10.1016/j.neuron.2012.11.026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang A.H., Chen H., Li T.P., Metzbower S.R., MacGillavry H.D., and Blanpied T.A.. 2016. A trans-synaptic nanocolumn aligns neurotransmitter release to receptors. Nature. 536:210–214. 10.1038/nature19058 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Titeca K., Lemmens I., Tavernier J., and Eyckerman S.. 2019. Discovering cellular protein-protein interactions: Technological strategies and opportunities. Mass Spectrom. Rev. 38:79–111. 10.1002/mas.21574 [DOI] [PubMed] [Google Scholar]
- Titulaer M.J., Mccracken L., Gabilondo I., Armangué T., Glaser C., Iizuka T., Honig L.S., Benseler S.M., Kawachi I., Martinez-Hernandez E., et al. . 2013. Treatment and prognostic factors for long-term outcome in patients with anti-NMDA receptor encephalitis: a cohort study. Lancet Neurol. 12:157–165. 10.1016/S1474-4422(12)70310-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toulmé E., Blais D., Léger C., Landry M., Garret M., Séguéla P., and Boué-Grabot E.. 2007. An intracellular motif of P2X(3) receptors is required for functional cross-talk with GABA(A) receptors in nociceptive DRG neurons. J. Neurochem. 102:1357–1368. 10.1111/j.1471-4159.2007.04640.x [DOI] [PubMed] [Google Scholar]
- Tovar K.R., and Westbrook G.L.. 1999. The incorporation of NMDA receptors with a distinct subunit composition at nascent hippocampal synapses in vitro. J. Neurosci. 19:4180–4188. 10.1523/JNEUROSCI.19-10-04180.1999 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tovar K.R., and Westbrook G.L.. 2002. Mobile NMDA receptors at hippocampal synapses. Neuron. 34:255–264. 10.1016/S0896-6273(02)00658-X [DOI] [PubMed] [Google Scholar]
- Trepanier C.H., Jackson M.F., and MacDonald J.F.. 2012. Regulation of NMDA receptors by the tyrosine kinase Fyn. FEBS J. 279:12–19. 10.1111/j.1742-4658.2011.08391.x [DOI] [PubMed] [Google Scholar]
- Vieira M., Yong X.L.H., Roche K.W., and Anggono V.. 2020. Regulation of NMDA glutamate receptor functions by the GluN2 subunits. J. Neurochem. 154:121–143. 10.1111/jnc.14970 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Villmann C., and Becker C.M.. 2007. On the hypes and falls in neuroprotection: targeting the NMDA receptor. Neuroscientist. 13:594–615. 10.1177/1073858406296259 [DOI] [PubMed] [Google Scholar]
- Viviani B., Bartesaghi S., Gardoni F., Vezzani A., Behrens M.M., Bartfai T., Binaglia M., Corsini E., Di Luca M., Galli C.L., and Marinovich M.. 2003. Interleukin-1β enhances NMDA receptor-mediated intracellular calcium increase through activation of the Src family of kinases. J. Neurosci. 23:8692–8700. 10.1523/JNEUROSCI.23-25-08692.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang M., Wong A.H., and Liu F.. 2012. Interactions between NMDA and dopamine receptors: a potential therapeutic target. Brain Res. 1476:154–163. 10.1016/j.brainres.2012.03.029 [DOI] [PubMed] [Google Scholar]
- Washburn H.R., Xia N.L., Zhou W., Mao Y.-T.T., and Dalva M.B.. 2020. Positive surface charge of GluN1 N-terminus mediates the direct interaction with EphB2 and NMDAR mobility. Nat. Commun. 11:570 10.1038/s41467-020-14345-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weinberger D.R. 2019. Thinking about schizophrenia in an era of genomic medicine. Am. J. Psychiatry. 176:12–20. 10.1176/appi.ajp.2018.18111275 [DOI] [PubMed] [Google Scholar]
- Wenk G.L. 2006. Neuropathologic changes in Alzheimer’s disease: potential targets for treatment. J. Clin. Psychiatry. 67(Suppl 3):3–7, quiz:23. [PubMed] [Google Scholar]
- Wu X., Morishita W.K., Riley A.M., Hale W.D., Südhof T.C., and Malenka R.C.. 2019. Neuroligin-1 Signaling Controls LTP and NMDA Receptors by Distinct Molecular Pathways. Neuron. 102:621–635.e3. 10.1016/j.neuron.2019.02.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wyllie D.J.A., Livesey M.R., and Hardingham G.E.. 2013. Influence of GluN2 subunit identity on NMDA receptor function. Neuropharmacology. 74:4–17. 10.1016/j.neuropharm.2013.01.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xing S., Wallmeroth N., Berendzen K.W., and Grefen C.. 2016. Techniques for the analysis of protein-protein interactions in vivo. Plant Physiol. 171:727–758. 10.1104/pp.16.00470 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan J., Bengtson C.P., Buchthal B., Hagenston A.M., and Bading H.. 2020. Coupling of NMDA receptors and TRPM4 guides discovery of unconventional neuroprotectants. Science. 370:eaay3302 10.1126/science.aay3302 [DOI] [PubMed] [Google Scholar]
- Young J.W., and Geyer M.A.. 2013. Evaluating the role of the alpha-7 nicotinic acetylcholine receptor in the pathophysiology and treatment of schizophrenia. Biochem. Pharmacol. 86:1122–1132. 10.1016/j.bcp.2013.06.031 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeng M., Shang Y., Araki Y., Guo T., Huganir R.L., and Zhang M.. 2016. Phase Transition in Postsynaptic Densities Underlies Formation of Synaptic Complexes and Synaptic Plasticity. Cell. 166:1163–1175.e12. 10.1016/j.cell.2016.07.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang H., Li T., Li S., and Liu F.. 2016. Cross-talk between α7 nAchR and NMDAR revealed by protein profiling. J. Proteomics. 131:113–121. 10.1016/j.jprot.2015.10.018 [DOI] [PubMed] [Google Scholar]
- Zhang J., Guan X., Li Q., Meredith A.L., Pan H.L., and Yan J.. 2018. Glutamate-activated BK channel complexes formed with NMDA receptors. Proc. Natl. Acad. Sci. USA. 115:E9006–E9014. 10.1073/pnas.1802567115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang R., Zhong N.N., Liu X.G., Yan H., Qiu C., Han Y., Wang W., Hou W.K., Liu Y., Gao C.G., et al. . 2010. Is the EFNB2 locus associated with schizophrenia? Single nucleotide polymorphisms and haplotypes analysis. Psychiatry Res. 180:5–9. 10.1016/j.psychres.2010.04.037 [DOI] [PubMed] [Google Scholar]
- Zhao N., Liu C.C., Qiao W., and Bu G.. 2018. Apolipoprotein E, Receptors, and Modulation of Alzheimer’s Disease. Biol. Psychiatry. 83:347–357. 10.1016/j.biopsych.2017.03.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu J., Shang Y., and Zhang M.. 2016. Mechanistic basis of MAGUK-organized complexes in synaptic development and signalling. Nat. Rev. Neurosci. 17:209–223. 10.1038/nrn.2016.18 [DOI] [PubMed] [Google Scholar]