

Abstract
Glycomic and glycoproteomic analyses involve the characterization of oligosaccharides (glycans) conjugated to proteins. Glycans are produced through a complicated nontemplate driven process involving the competition of enzymes that extend the nascent chain. The large diversity of structures, the variations in polarity of the individual saccharide residues, and the poor ionization efficiencies of glycans all conspire to make the analysis arguably much more difficult than any other biopolymer. Furthermore, the large number of glycoforms associated with a specific protein site makes it more difficult to characterize than any post-translational modification. Nonetheless, there have been significant progress, and advanced separation and mass spectrometry methods have been at its center and the main reason for the progress. While glycomic and glycoproteomic analyses are still typically available only through highly specialized laboratories, new software and workflow is making it more accessible. This review focuses on the role of mass spectrometry and separation methods in advancing glycomic and glycoproteomic analyses. It describes the current state of the field and progress toward making it more available to the larger scientific community.
1. INTRODUCTION
Glycosylation plays a key role in maintaining health and in diseases. While the importance of glycosylation has long been known, the ability to determine and quantitate structures have hindered the ability to obtain deeper and more refined details regarding the role of glycosylation. Indeed, the modification of proteins by glycosylation has been a long-standing problem hampered by the dearth of methods to characterize and quantitate glycans. Unlike the genome and the proteome, there is no template for the glycome. Glycans are produced by a set of competing enzymes with the addition of each monosaccharide dictated by the one before. Furthermore, there is no “completed” structure. The protein may exit anywhere along the glycosylation pathway producing a suite of structures that vary homogeneously by linkages, length, number of antennae, and composition. For these and other reasons, glycomic and glycoproteomic methods have not advanced as rapidly as genomic and proteomic methods. Nonetheless, there have been considerable recent improvements. Glycan profiling tools whether by mass to yield compositions or separation to yield structures measure the scope of the glycome, while proteomic and lipidomic methods are advancing to yield intact glycoconjugates. Mass spectrometry (MS) has been at the center of this effort. Mass spectrometry methods that yield accurate mass, structurally informative fragments, coupled to advanced separation methods including capillary electrophoresis (CE), high performance liquid chromatography (HPLC), and ultrahigh pressure liquid chromatography (UPLC) have contributed significantly to the effort.
Glycosylation is a post-translational modification of proteins, but glycans can also be found on lipids and as free compounds in, for example, human milk. Combined in various forms, it represents one of the most common types of modification of proteins and the one that is also most structurally complicated. On proteins, they add an additional level of information but play outsized roles on protein function.
Glycans are short carbohydrate chains consisting of a single monosaccharide to large polysaccharides consisting of thousands of saccharide units. The monosaccharide includes the most notably O-GlcNAc (N-acetylglucosamine),1 however, other monosaccharides modifications have also been observed.2 Large polysaccharides include polysialic acids and glycosaminoglycans (GAG).3 This review will focus on those that are on serine or threonine, the O-glycans, and those on asparagine, the N-glycans. Those interested in other types of glycans such as O-GlcNAc and GAG are referred to other current reviews.4,5
Figure 1 illustrates the many levels of information associated with protein glycosylation. The glycans may be released and measured by mass spectrometry to yield compositional information (the apex of the pyramid). The majority of early MS analyses of glycans have focused on composition because glycans can be released and profiled according to mass. The analysis yields the number of hexose (Glc, Gal, and Man), N-acetylhexosamine (GlcNAc and GalNAc), deoxyhexose (Fuc), and sialic acid (Sia or NeuAc and NeuGc). Although this information is limited, it is sufficient for observing, for example, the rise of fucose or sialic acid in N-glycans during biomarker discovery.
Figure 1.
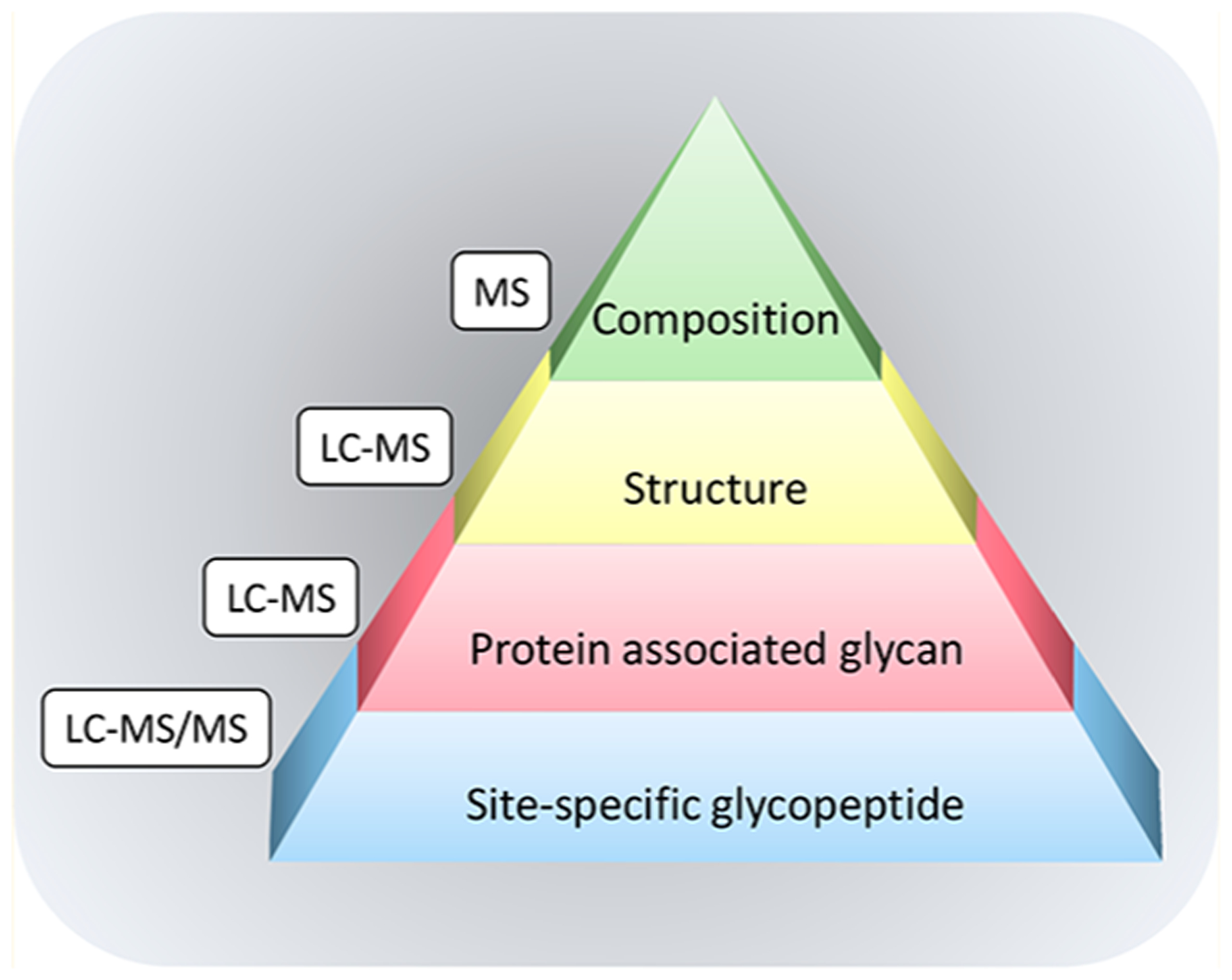
An illustration of the several levels of complexity in glycoprotein analysis.
Glycans may be separated chromatographically and analyzed by mass spectrometry or spectrophotometric detection to yield the glycan profile, structurally separated and typically elucidated array of structures (Figure 1). Glycan profiles have increased our understanding of the large breadth in glycan structures from biological samples. A common early belief was that there are too many structures resulting in heterogeneity too great to perform comprehensive analysis. It was believed, for example, that the large number of possible structures were nearly infinite. Six oligosaccharides can indeed chemically produce 1012 possible combinations,6 however, there is a limited number of glycosyltransferases, which severely limits the number of structures. The notion was perhaps first noted when HPLC using porous graphitized carbon stationary phase became available for oligosaccharides profiling. The analysis of free oligosaccharides in human milk (human milk oligosaccharides or HMOs) with LC-MS showed that rather than the potentially millions of structures thought to be present, it was observed that five different mothers produced less than 500 structures spanning over 5 orders of magnitude in abundances. A single mother produced approximately between 23 and 130 structures.7
Using a neural network of potential oligosaccharides, Kronewitter et al. deduced the different compositions that can be produced and hence the size of the serum N-glycome.8 They estimated that there are less than 500 compositions employing standard monosaccharides found in humans. On the basis of this analysis, we assumed that the glycome is limited and can be characterized entirely using annotated glycan libraries. There have since been efforts to create comprehensive libraries of complete structures for serum N-glycans9 and for human milk oligosaccharides.10,11
The glycans are associated with specific proteins providing large diversities among protein glycoforms (Figure 1). An example of the complexity of glycoproteins is illustrated with IgA, an abundant immune protein in blood and human milk (Figure 2).12 The secretory IgA found in human milk is composed of the secretory component (SC), J chain, and IgA1 or IgA2, while IgA in blood is mainly monomeric. The permutations between the different glycoforms in the polypeptides units can be almost endless.
Figure 2.
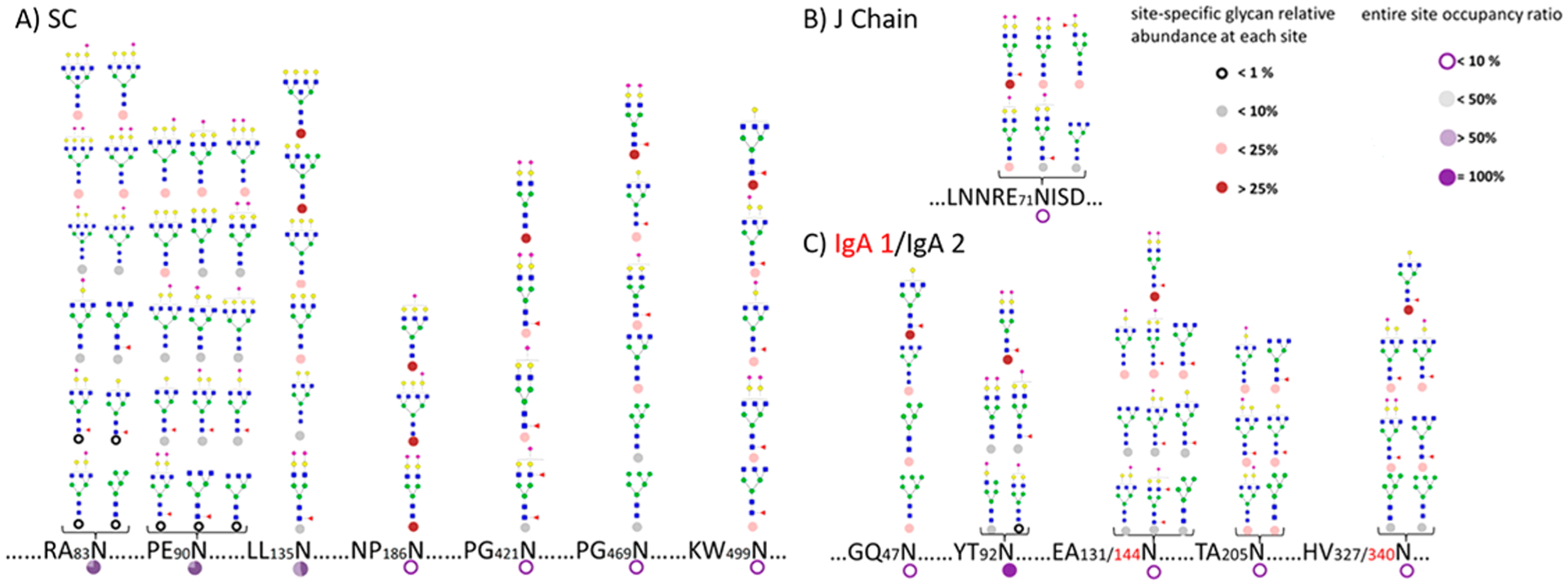
Site-specific glycosylation with occupancy information on secretory IgA from human milk. An example of the complexity of glycosylation even on a single protein. Reprinted with permission from ref 12. Copyright 2015 American Chemical Society.
Protein enrichment followed by glycan release provide information regarding protein-specific glycosylation (lower tier of the pyramid in Figure 1). This method has been used extensively, for example, with immunoglobulin G (IgG), where the glycoprotein is enriched by protein G and the glycans are released from the enriched fraction.13 This method has also been used to discover markers for a whole host of diseases including various cancers.14
Despite the difficulty in glycoproteomic analysis, the protein and the site-specific glycans can now be simultaneously characterized. An initial effort toward the extensive characterization of glycosylation on proteins using an automated software was first described by An et al. on a small set of proteins.15 Recent glycoproteomic software are now commercially available that yield extensive characterization of intact glycopeptides.
1.1. Understanding the Biological Pathways Is Necessary for Elucidating Structures
The key to developing methods for glycoproteomic analysis lies in understanding the glycosylation process and the products manufactured by cells. The structural properties of glycans are complicated and as such highly challenging from the perspective of structural elucidation. First, glycans may be subdivided based on the way they are attached to the peptide (N-or O-glycans as well as O-monosaccharide modifications). N-Glycans and O-glycans are produced in a series of competing enzymatic steps through nontemplate driven processes. The processes differ for the two glycan types. A recent review by Moremen et al. described well the diversity, structure, and function in vertebrate glycosylation.16
N-Glycans are produced at first in the endoplasmic reticulum on a lipid (dolichol) to yield ultimately a high mannose structure consisting of nine mannoses with a triglucose terminus (Figure 3). This structure is then transferred to a nascent polypeptide chain where it guides the protein folding process. When folding is complete, the glycan structure first losses the triglucose structure and a stepwise process disassembles the high mannose structure and rebuilds a variety of structures in the Golgi. Typically, N-glycans have a common core consisting of two GlcNAc residues attached to three mannoses. This core structure may be extended using multiple substitutions to form different branching patterns as well as a large number of linkages.17 N-Glycans may be classified into three groups: the complex type N-glycans, the hybrid type N-glycans, and the high-mannose type N-glycans (Figure 4). On the basis of the biosynthesis of N-glycans, the structures follow a strict regime involving a single core but differ in the composition and lengths of the antenna. Glycoproteomic analysis of N-glycans is further simplified by a consensus sequence for a glycosylation site of N-X-S/T, where N is asparagine, X is any amino acid but proline, and S/T is serine or threonine.
Figure 3.
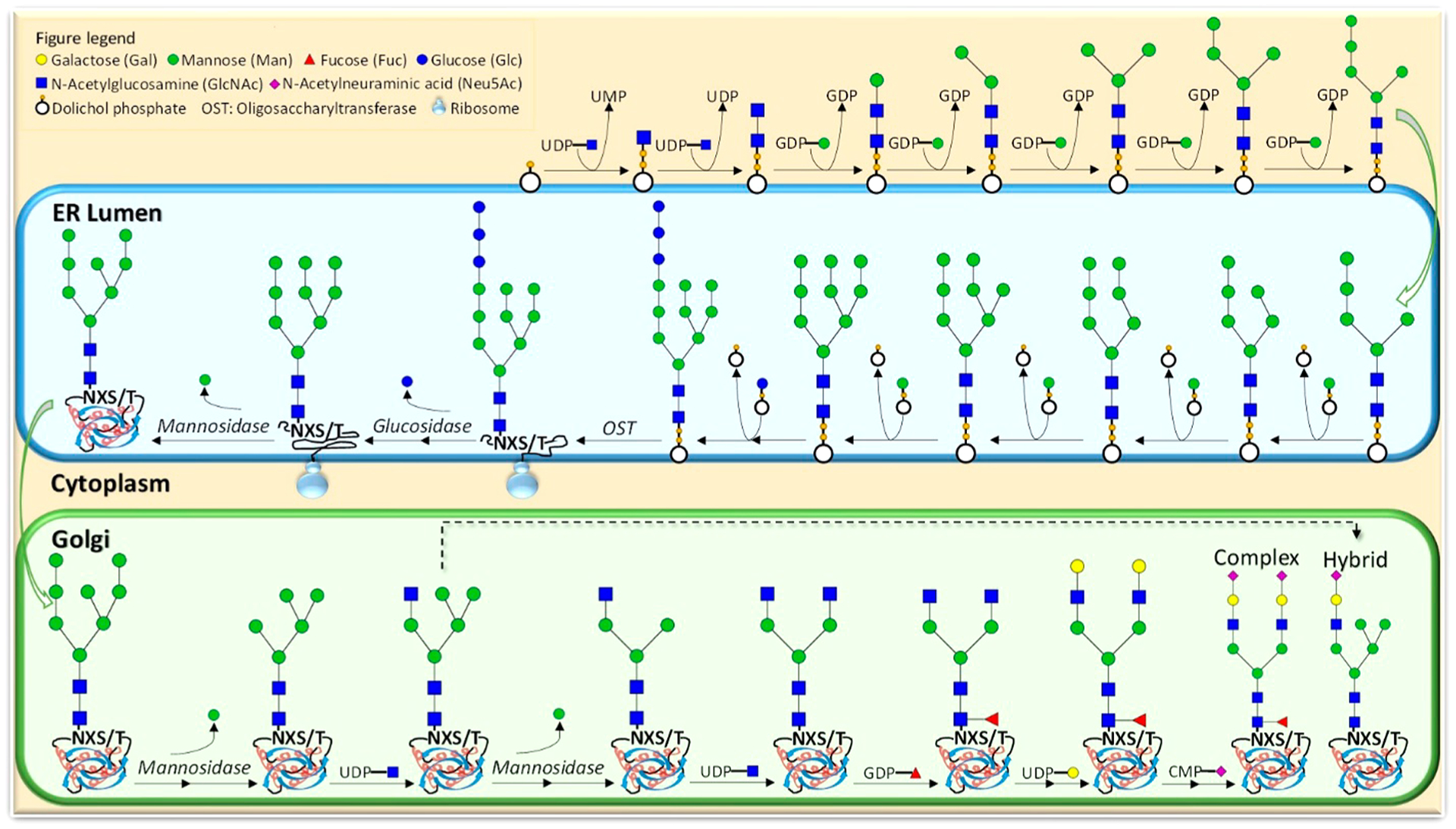
Mechanistic pathway for the formation of N-glycans.
Figure 4.
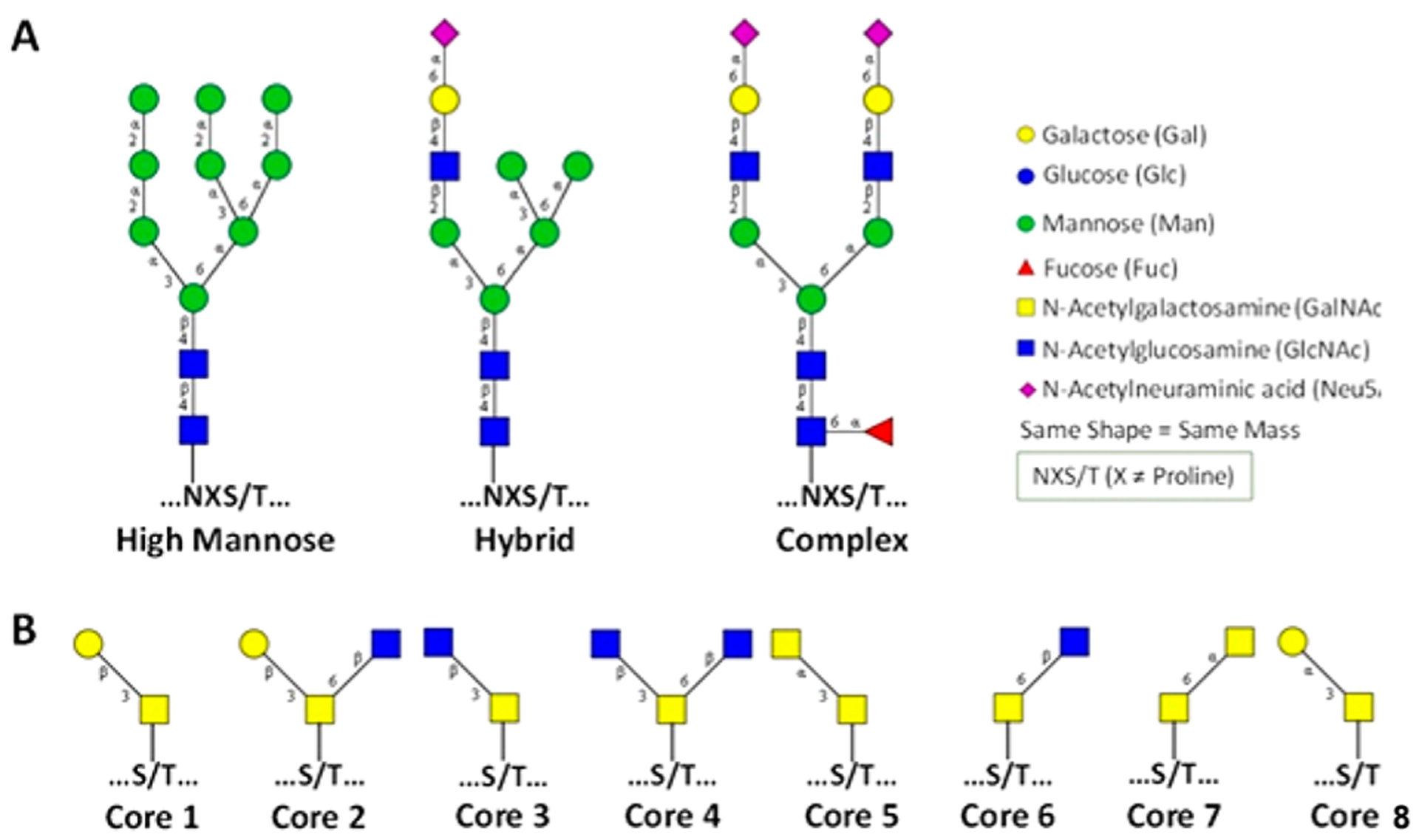
Variation in the N- and O-glycosylation in proteins. (A) Three types of N-glycans. (B) Eight core structures of O-glycans.
In contrast to N-glycans, O-glycans represent a greater challenge for structural analysis. There is no unique consensus sequence for O-glycosylation. There are several types of O-glycans including α-linked O-GalNAc, β-linked O-GlcNAc, α-linked O-fucose, α-linked O-mannose, β-linked O-xylose, α- or β-glucose glycans, and α- or β-linked O-galactose.18 However, there have been many more studies performed on O-GalNAc and O-GlcNAc types of glycosylation due to their known biological functions. There are currently eight known core structures with the mucin-type O-glycans varying in length and in branching antennae (Figure 4).19 The difficulty of O-glycan analysis also lies in the lack of universal enzymes to release O-glycans. There are enzymes that can release mono- and disaccharides from proteins but not larger more complicated structures. The current methods for release rely on chemical approaches that are discussed in the glycomics section below.
Despite the complexity of glycan synthesis, the resulting glycosylation appears to be biologically (phenotypically) conserved. The serum glycomic profile of an individual is highly reproducible in the structures and their abundances.20 Furthermore, the glycoprotein receives considerable structural variability due to glycosylation. A protein such as IgG has been shown to have over 70 different glycoforms at one site.21 A protein with a single polypeptide backbone and three glycosylation sites with 10 possible glycan structures at each site may have over a thousand unique glycoforms. It is arguably due to this complexity that glycans have been largely neglected in the large proteomic effort. Even at this point, 20 years after “proteomic” was coined,22 glycoproteomic analysis remains far from routine compared to proteomics.
1.2. Previous Reviews on MS Methods for Glycomics and Glycoproteomics
While the majority of the analytical chemistry advancements over the last decades have focused on analyzing nucleic acids in the form of DNA and RNA and amino acids in the form of proteins and peptides, it has become evident that glycans are also important biopolymers and play important functions in areas as broad as embryonic development and evolution but also in essentially every area of cellular processing. As with all other “omics”, it is clear that technological advances in analytics and informatics are major drivers for the in-depth analysis of glycans and glycoproteins. Mass spectrometry, in particular with its multitude ionization, fragmentation, and detection techniques, plays a central role in glycomic and glycoproteomic analyses.
There have been recent notable reviews in this area. A series of comprehensive reviews have been written by Harvey on glycans analyzed by matrix-assisted laser desorption/ionization mass spectrometry (MALDI-MS).23–27 Mulagapati et al. have described the analysis of O-glycosylation by current mass spectrometric methods.28 Banazadeh et al. examined the recent advances in glycoprotein analysis by mass spectrometry with a focus on N-glycosylation.29 Oligosaccharide analysis by MS was reviewed by Kailemia et al.30 Although not in the scope of this review, but important to glycosylation nonetheless, Ma et al. reviewed the state of O-GlcNAc analysis in proteins and the proteome.4 The concept of chemical glycoproteomics, which employs chemical approaches to glycoprotein analysis, was reviewed by Palaniappan et al.31
2. INSTRUMENTATION FOR GLYCOMIC AND GLYCOPROTEOMIC ANALYSES
2.1. Key Characteristics of MS Needs/Requirements
Mass spectrometry has several important features that make it ideal for glycomic and glycoproteomic analyses. It has the capability of providing structural information on small amounts of material. Limits of detection have reached femtomolar to attomolar levels for glycans, while the dynamic range is typically 4 or 5 orders of magnitude.32,33 For an abundant signal at 100% relative intensity, signals with corresponding relative abundances of 0.01% are also observed. Structural analysis of glycans can be performed on abundances typically in the femtomolar range. There is no other analytical method that can provide such low limit of detection with the structural information. Further mass analyzers can be combined in tandem (e.g., QTOF and QIT-FTMS), and these hybrid mass analyzers can further improve the dynamic range.
The instrumental requirements for glycomics, glycoproteomics, and intact glycoprotein analysis are summarized in Figure 5. For comparison, bottom-up proteomics is also included. Specifically for glycomics experiments, the sensitivity of the method should be slightly higher compared to bottom-up proteomics strategies due to the poorer ionization efficiency of glycans. The need for sensitivity is even higher for glycoproteomics experiments, where the glycan heterogeneity is also subdivided over the peptide backbones. In this regard, extensive separation is less of a requirement in glycomics experiments where glycan compositional information may be obtained by direct MALDI-TOF or MALDI-FTICR analysis of released glycans. However, it is needed for glycoproteomics and even more so for intact glycoprotein analysis, where isolation of a single glycoprotein is highly beneficial. Indeed, compositional glycomic analysis is typically performed using MALDI-MS, which can be configured for high-throughput applications. The requirement of LC or CE separation prior to MS analysis for glycopeptides and intact glycoproteins typically results in a loss of throughput. While CID fragmentation is traditionally the method of choice for bottom-up strategies, this fragmentation strategy may provide information in a glycomics experiment but is generally not sufficient in glycoproteomics and intact glycoprotein analysis. For better identification of intact glycopeptides and glycoproteins, alternative dissociation techniques such as ETD, EThcD, UVPD, or AI-ETD are needed to provide more comprehensive fragmentation of both the peptide backbone and the glycan. As a result, more extensive software tools are required in these experiments to assign the MS spectra for structural elucidation.
Figure 5.
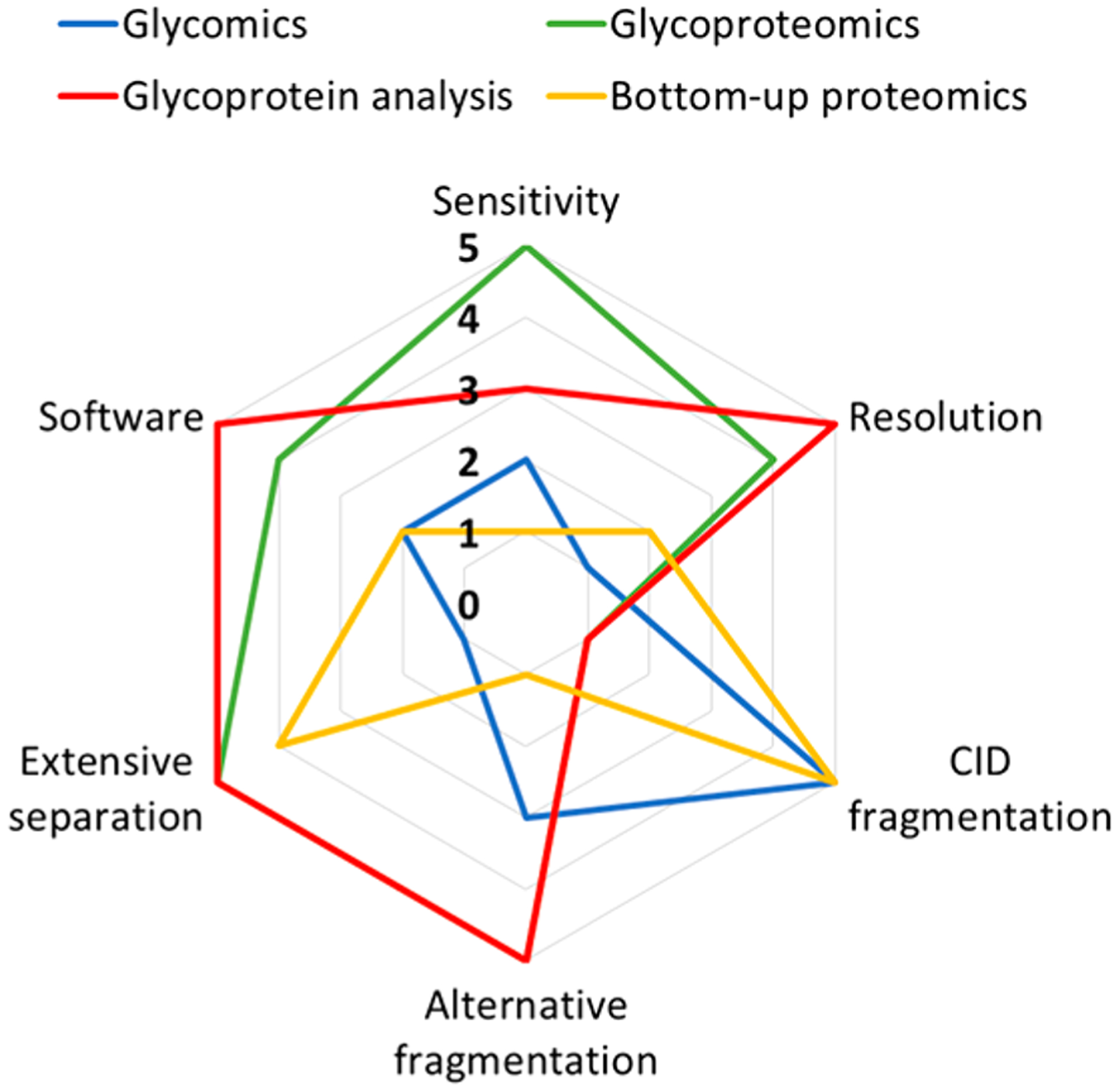
Spider graph showing the instrument requirements for glycomics, proteomics, glycoproteomics, and intact glycoprotein analysis.
Mass analyzer performance also varies depending on the resolution, mass accuracy, and scan rate. Resolution is defined in mass spectrometry as m/Δm, where Δm is the full width at half-maximum. Because of the relatively large mass differences between monosaccharides, it was generally believed that high resolution was unnecessary for mass spectrometry. However, there is a need for accurate mass, particularly in global profiling of released glycans. High mass accuracy provides rapid differentiation of glycan peaks from nonglycan peaks. Peptides and even lipid contaminants can have masses that nominally corresponds to glycan compositions but are rapidly differentiated when the accurate masses are known. Fourier transform ion cyclotron resonance mass spectrometry (FTICR MS) was the earliest technique used for high resolution analysis of glycans.34 N-Glycans and O-glycans released from serum proteins and analyzed by MALDI-FTICR illustrated the utility of global glycan profiling for disease and specifically cancer biomarker discovery.35 Time-of-flight analyzers employed in QTOF and MALDI-TOF instruments have improved considerably in terms of resolution and are used for global glycan profiling with great success. More recently, orbitrap FT MS with high resolution and mass accuracy has also been used, however, more for glycoproteomic analysis.36
Glycoproteomic methods generally require high resolution to be most effective and are best performed with orbitrap MS. Both top-down and bottom-up methods require high resolution for more specific identification and elucidation of glycoforms. FTICR generally lacks the capability of fast LC-MS analysis and are not used for glycoproteomics, although there are numerous examples of glycoproteins analyzed by FTICR.37 Quadrupole and TOF mass analyzers have high scan rates and are highly essential in high-throughput analyses to get reliable quantitative data. In glycoproteomics, these instruments are used to quantitate glycans and site-specific glycosylation.38,39
2.2. Ionization Considerations
Glycans contain labile residues such as fucose and sialic acid that might easily be fragmented either in-source or post-source during ionization. Soft ionization techniques that impart little excess energy and thus generate intact molecular ions are normally used for glycan and glycopeptide analyses. Milder source conditions are often needed for the analysis of glycans, especially native glycans, than those for peptides and small molecules.40 Among the various ionization methods available, MALDI and ESI are the most widely used.
MALDI-MS has been extensively used for profiling of released glycans derived from biological mixtures. The energetic conditions of the process did not initially favor the formation of intact species. MALDI ionization produced in-source fragmentation that made it unsuitable for early high performance instruments such as FTICR MS.41 Efforts were placed on finding matrices that yielded intact species. Permethylation was one of the methods used to produce intact species in both time-of-flight and FTICR MS. Partial derivatization including esterification of sialic acids achieved similar results in a more specific manner. For native compounds, post-collisional cooling was used in FTICR MS to remove the excess energy. Metastable dissociation of MALDI produced ions were studied by Ngoka et al. showed that ions generated by MALDI dissociated in the millisecond time scale.42 Because oligosaccharides have weak gas-phase basicities, they were typically doped with alkali metals to yield quasimolecular ions. They further showed that alkali metals yielded variable rates of dissociation with the fastest corresponding to the smallest metal, thus lithium yielded the fastest rate. Similarly, Fenselau and co-workers showed that peptides produced by MALDI also decayed, with in-source collisional cooling found to minimize dissociation.43 Later FTICR MS included collisional cooling to decrease fragmentation of oligosaccharide ions, thereby allowing native analysis of multiply sialylated and fucosylated species.44
MALDI ionization was used extensively for biomarker discovery. The first glycomic biomarker study of cancer was performed by MALDI.45 MALDI-MS has the advantage of being more amenable to biological species containing salt residues. Indeed, sodium chloride was often used as dopant to yield sodium coordinated oligosaccharide ions.41 MALDI-MS is used less for profiling because of the difficulty in mating with chromatography. The method is used more today for MS imaging of oligosaccharides as discussed further below.
Electrospray ionization (ESI) is more commonly used for glycan analysis due to its greater sensitivity and its ability to couple to liquid chromatography. ESI imparts less internal energy on the ion compared to MALDI. Unlike MALDI, where the neutral compounds are better ionized in the positive mode while sialylated compounds are better ionized in the negative mode, ESI can be used for both neutral and anionic native glycans in the positive mode. ESI is also most amenable for glycopeptides, however, glycosylation tends to diminish the glycopeptide signal compared to the peptide signal considerably.46
2.3. Fragmentation Methods
The ability to obtain fragments gives MS the structural elucidation capability with high sensitivity. Nearly every fragmentation method has been attempted with glycans and glycopeptides. They have been sufficiently characterized to determine the best methods for glycan and glycopeptide analyses. The methods for glycoproteins are discussed in greater detail below for top-down analysis.
2.3.1. Low Energy Dissociation Methods.
Collision-induced dissociation (CID) involves the use of collisions with neutral gas molecules or surfaces to yield fragmentation of isolated ions. For glycans (and glycopeptides), collision induced dissociation with gases is more common. Surface induced dissociation is less common but yields the same results.47 Infrared multiphoton dissociation (IRMPD) is a photon-based dissociation method with energetics similar to CID, yielding the same fragments and hence included in this group. The fragmentation of oligosaccharides under CID and IRMPD conditions have been reviewed extensively previously.48
Low energy tandem MS methods are typically employed in analyzers such as quadrupole and FTICR. The fragmentation behavior of N- and O-glycans follow regular rules of fragmentation (Figure 6). This includes primarily cleavage along glycosidic bonds for native glycans. Low energy dissociation methods follow the tendency order of the bond to break. Fucosylated glycans tend to lose fucose more readily, resulting in abundant fragment ions with fucose loss from the quasimolecular ion in immediate succession. Sialic acids are also readily lost when present and are even more labile than fucose. In the positive mode, the loss of sialic acids are the major fragment ions. In the negative mode, sialic acid is the charge containing unit. The most abundant fragment tends to be the lone negatively charged sialic acid.
Figure 6.
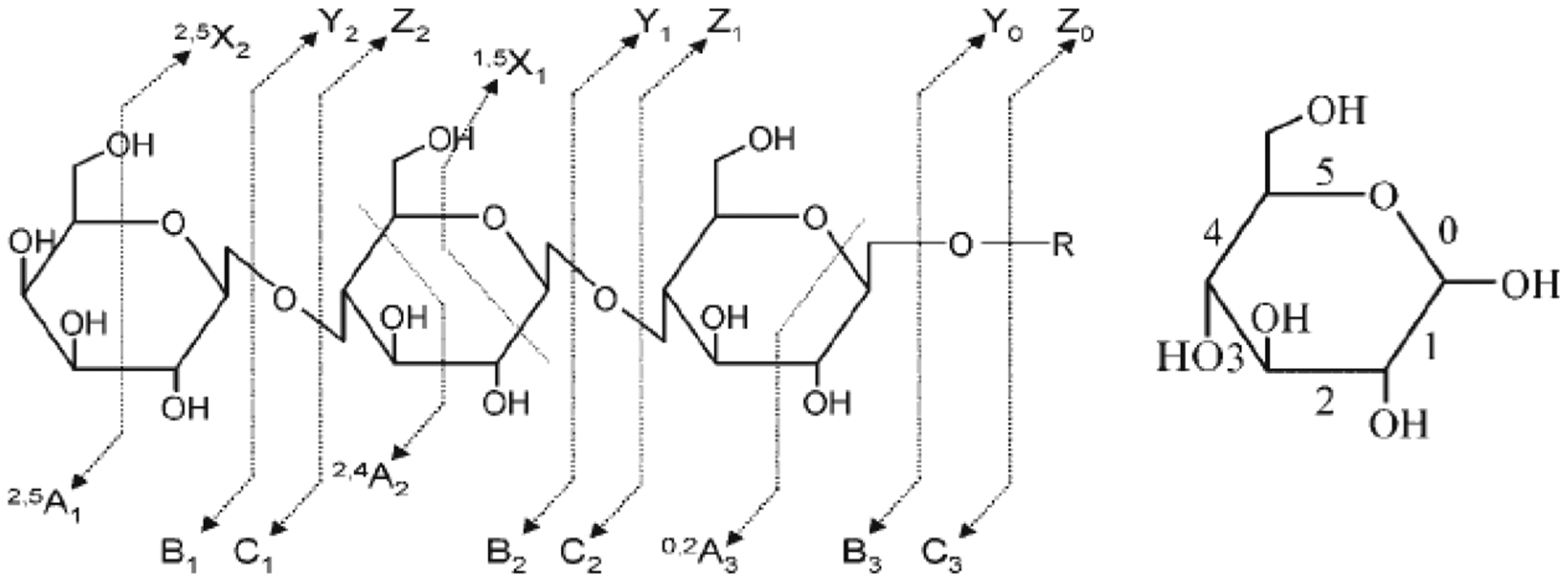
Fragmentation behavior of native oligosaccharides under collision-induced dissociation conditions. Reprinted with permission from ref 48. Copyright 2011 Wiley Online Library.
An important issue with native glycans is the occurrence of migrating species during CID. Fucose tends to transfer with high probability from the termini to the reducing end in native oligosaccharides performed in the positive mode, resulting in the loss of internal residues. This is diminished significantly when the reducing end is reduced to the alditol or derivatized. The topic has been covered extensively in a previous review.49
Permethylated compounds yield fragments that diverge from native species. The permethylation reaction of oligosaccharides originates from the early work on carbohydrates, where permethylation of oligosaccharides allows thermally labile species to be more volatile.50 For oligosaccharide analysis, proponents claim that it yields higher sensitivity. However, for native glycans, limits of detection as low as attomole and even zeptomole have been reported,51 while no similar reports of permethylated species have been found. Fragments tend to concentrate along glycosidic bonds in permethylated species, however, cross-ring cleavages are also present in the tandem mass spectra. The use of MSn where n > 2 has been employed as a method for obtaining linkage information.52 Cross-ring cleavages generally identify the linkage position, however, they do not readily determine the stereochemistry nor the anomeric character of the linkage.
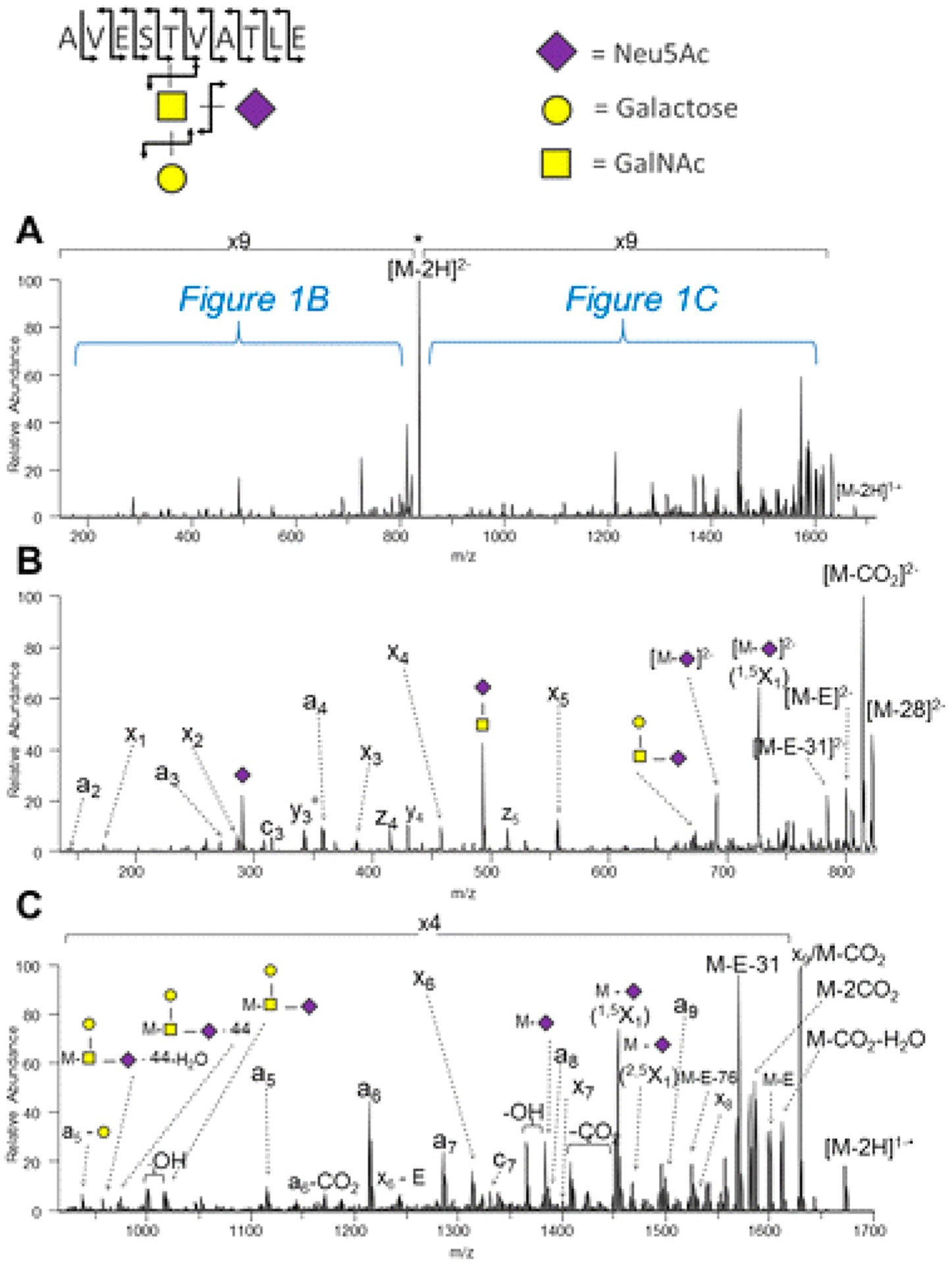
Fragmentation of glycopeptides is complicated by the presence of two chemically dissimilar groups. Peptides create their own difficulties under CID conditions. Glycan bonds are more labile than peptide bonds. Glycans also have significantly lower intrinsic basicity than peptides. For these reasons, both the ionization and the CID conditions of glycans differ significantly from that of glycopeptides. Early studies using both CID and IRMPD yielded predominantly glycan fragments but peptide fragments as well. Glycan fragmentations are low energy processes, and primarily B and Y ions produced by cleavages of glycosidic bonds are the most abundant fragments.53 Charge states and coordinating cations varied the product ion in proportion depending on how the charge was retained. Sodiated species produced information primarily on the glycan sequence, while protonated species yielded both glycan and peptide information (Figure 7).53 The same fragmentation patterns were reported for both N-glycopeptides and O-glycopeptides.53,54
Figure 7.
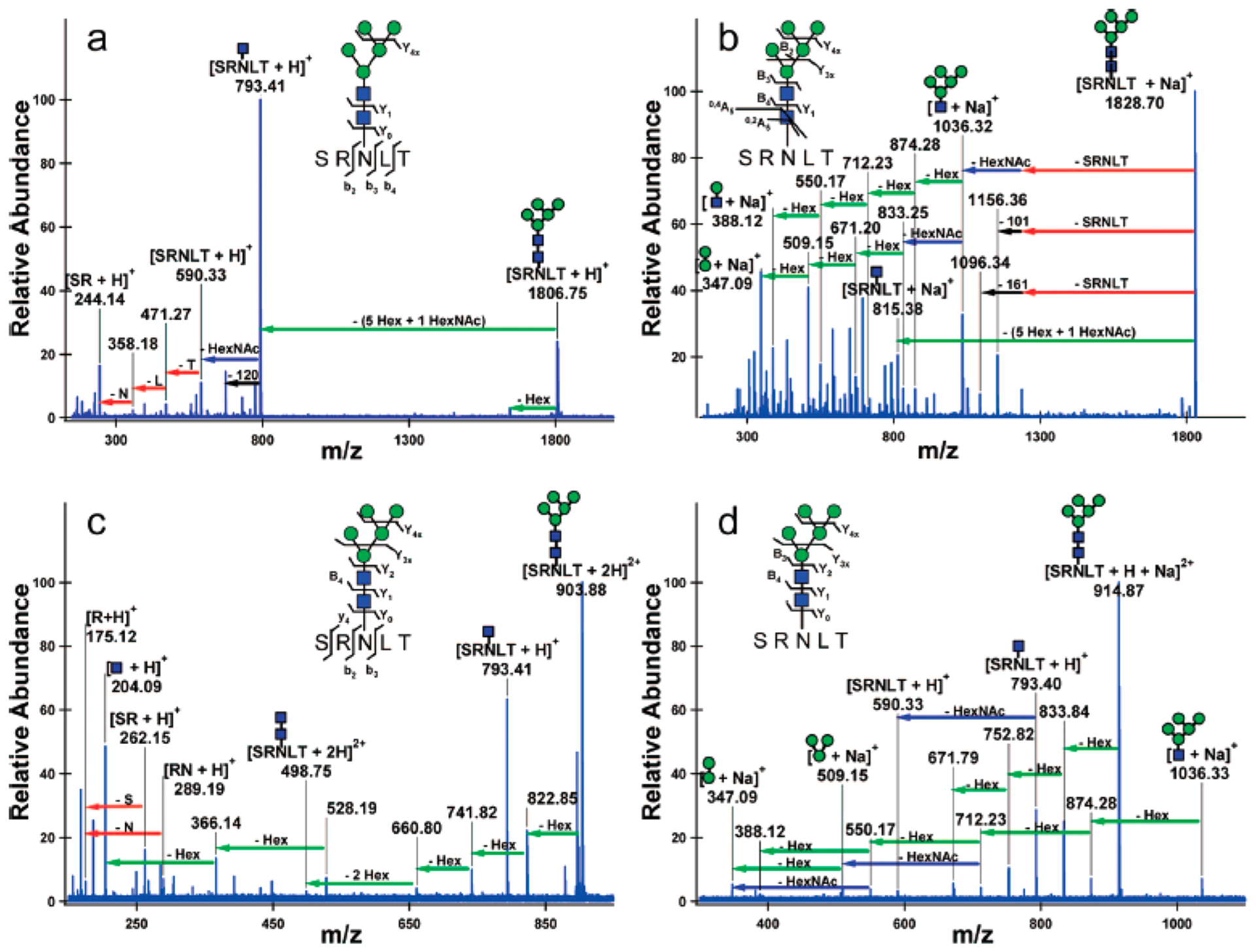
IRMPD of glycopeptides yields low energy fragmentation corresponding to glycan and peptide fragments. Protonated species yield information regarding the peptide, while sodiated species yield fragmentation regarding the glycan. Conditions can be varied to yield both glycan and peptide fragmentation for protonated species under CID and IRMPD. Reprinted with permission from ref 53. Copyright 2008 American Chemical Society.
2.3.2. High Energy Dissociation Methods.
High energy dissociation methods include high energy CID, as well as electron activation methods and UV photodissociation. High energy CID, those involving keV energies, yield extensive dissociation and are available in two types of instrumentation, namely TOF/TOF and sector instruments. Sector instruments are no longer commonly used in bioanalysis and will not be discussed further. TOF/TOF can be used but are not directly coupled to chromatographic methods, which are necessary for separating glycan isomers. Furthermore, the precursor mass selecting capabilities of TOF/TOF instruments remain crude compared to the quadrupoles used in LC-MS methods.55 These methods have not received wide utility in glycomic and glycoproteomic analysis.
Electron Aided Dissociation.
Besides high energy CID, another widely used method for fragmentation of glycopeptides and glycans is electron aided dissociation, where fragments are due to interactions between electrons and multiply charged ions. The most common is electron transfer dissociation (ETD), where an electron in a molecular carrier is transferred to a precursor ion. For peptide cations, the fragment ions produced by ETD are mainly c and z ions along peptide backbone compared to the b and y ions produced by CID.56 The method also yields fragmentation of the peptide backbone for glycopeptides, where CID of the same precursor ions yield more extensive glycan cleavages. ETD has the advantage of keeping the glycan moieties intact and retained on the amino acid while cleaving the peptide backbone(Figure 8).57–59
Figure 8.
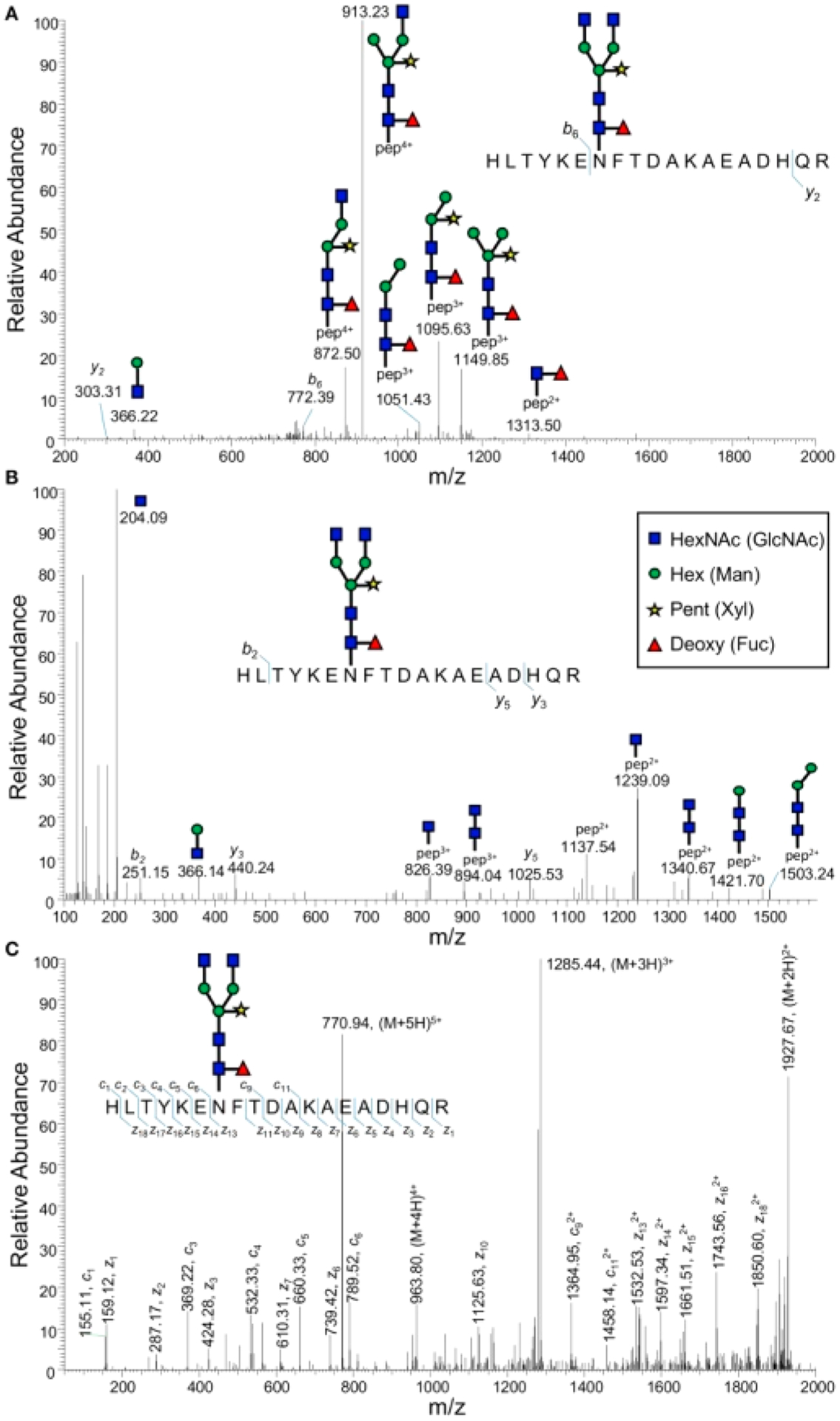
Comparison of (A) CID, (B) HCD, and (C) ETD fragmentation of an enriched plant glycopeptide. Reprinted with permission from ref 59. Under Creative Commons license (https://creativecommons.org/licenses/by/4.0/).
For both N- and O-glycopeptides, ETD has been shown to provide structurally characteristic fragment ions. Windwarder et al.60 characterized the O-linked glycosylation of the recombinant membrane proximal (c/MAM) domain of neuropilin-1 using ETD MS/MS, revealing that the c/MAM domain is O-glycosylated and contains mainly disialylated core structure. N-Glycopeptides of recombinant polymeric human IgM protein were probed with ETD MS/MS, showing the heterogeneity of glycosylation on different glycosylation sites.61
ETD has also been combined with other dissociation methods to generate complementary information in different spectra of one precursor or more comprehensive fragmentation patterns in one spectrum. One commonly used hybrid dissociation method is the combination of electron-transfer and higher-energy collision dissociation (EThcD) whereby HCD fragmentation is further applied to all the ions generated from the ETD step.62 It should be noted that HCD in the context of this review is only high energy CID and is used by the manufacturer to distinguish from a lower energy CID, which they term as CID. EThcD was first developed to obtain higher peptide sequence coverage, but was later applied to studying post-translational modifications (PTM) of proteins.
Often, the combination of several fragamentation methods provides better coverage than a single method. Parker et al. studied the glycosylation changes on adipocyte cell membrane under the state of insulin resistance using LC-MS/MS with triggered EThcD.63 The MS/MS spectra of enriched N-glycopeptides were first collected with HCD. Then, the precursor ions with HexNAc fragments in the MS/MS spectra were reisolated and subjected to EThcD. To confirm the glycan structures of N-glycopeptides, CID was also conducted on the reisolated precursor ions. Due in part to the inefficiencies of triggering EThcD, the number of identified glycopeptides was lower than for HCD alone. The manual interpretation of EThcD spectra provided precise localization of glycosites. However, they identified more unique glycopeptides by combining HCD and EThcD data into one matching algorithm. By modifying the EThcD format to collect HCD and ETD fragments simultaneously, Yu et al. were able to perform the large-scale N-glycopeptide characterization of both human plasma and rat carotid artery samples.64 With the improved fragmentation and shortened ETD reaction times, more N-glycopeptides were identified and mapped on their respective glycosylation sites.
Other electron aided dissociation methods that have been applied in LC-MS/MS for glycan or glycopeptide analysis include electron capture dissociation (ECD) and electron excitation dissociation (EED). ECD has been used together with IRMPD for characterizing tryptic N-glycopeptides in Erythrina corallodendron lectin, providing complementary structural information including peptide backbone fragmentation generated from ECD and monosaccharide composition generated from IRMPD.65 Studies on improving the efficiency of ECD for glycopeptide dissociation have been performed by optimizing the electron energy ranging from 0.2 to 9 eV. Higher sequence coverages of complex N-glycans with sialylation were obtained from “hot” ECD, which they defined as ECD at the higher energy range.66
Electron energy larger than 9 eV is considered as EED, which was found to generate even more fragments than hot ECD. To obtain efficient EED, decreasing the electron flux to minimize electron repulsion in the ion cyclotron resonance (ICR) cell was required to increase the electron interaction. This method was applied toward analyzing glycans with sialylation and fucosylation in tumor antigens.67 An improvement of EED, compared to ECD or ETD, is that there is no charge reduction in EED so that glycans with lower charges can also be fragmented efficiently.67
Ultraviolet Photodissociation (UVPD).
UVPD is an emerging new method that has great potential in both bottom-up and top-down glycoproteomics. Pulsed UV photons generated from lasers are used to excite ions to higher electronic states, raising the internal energy of the ion to the range of 3.5–7.9 eV.68 This method differs from IRMPD, where the fragmentation mechanism follows lower energy dissociation pathways due to the small energy step size from lower photon energy. UVPD provides higher energy UV photons that excite ions to higher energy levels that generate new fragmentation pathways. Because the dissociation of ions takes place either right after the excitation or after energy redistribution, different dissociation pathways are accessed, resulting in more diverse fragmentation products. Two commonly used wavelengths in UVPD for analyzing biological macromolecules such as peptides, glycopeptides, and glycans are 157 and 193 nm.68 For oligosaccharides, both glycosidic and cross-ring cleavages take place in UVPD, thereby generating more A and X-type ions, which is useful for determining the branching patterns of glycans.
Both N- and O-linked glycopeptides have been studied with UVPD mass spectrometry. Zhang and Reilly have employed 157 nm UVPD on N-linked glycopeptides, thereby generating more varied fragmentation of both peptide and N-glycan.69 UVPD has also been employed on O-linked glycopeptides (Figure 9)70 to yield a greater amount of fragmentation of both the peptide backbone and the glycan in contrast to CID.71 Additionally, UVPD at 193 nm has been applied to oligosaccharides with labile groups such as sialic acids, thereby facilitating their locations in highly branched structures.72 Middle-down UVPD at 193 nm was recently used as a promising strategy to characterize therapeutics by providing both the peptide sequence and the glycosylation site of immunoglobulin G.73
Figure 9.
UVPD of a doubly deprotonated O-glycopeptide from κ casein yields fragmentation of both the glycan and peptide backbone. Reprinted with permission from ref 70. Copyright 2013 American Chemical Society.
3. CURRENT METHODS FOR COMPREHENSIVE GLYCOMICS
The rapid development of MS methods for glycomic analysis has made it possible to obtain comprehensive glycan profiles of biological samples. Glycomics is less complicated than glycoproteomics because the carrier proteins and glycosylation sites of the glycans are not considered. However, glycomics data alone can often identify aberrant glycosylation changes in diseases and facilitate the characterization of glycoproteins. This section will cover the essential techniques for glycomics experiments including strategies for sample preparation, glycan separation, and structural elucidation. Other emerging techniques such as ion mobility and imaging will also be discussed.
3.1. Strategies for Releasing Glycans
3.1.1. Enzymatic Release.
A critical step in glycomic analysis is the efficient release of N- or O-glycans from glycoproteins or glycopeptides. Enzymatic digestion using endoglycosidase or glycoamidase is preferable because it can provide specific and complete sugar removal under mild conditions. The most common method for N-glycan release is enzymatic digestion using peptide-N-glycosidase F (PNGase F). Complete release of N-glycans from glycoproteins or glycopeptides in complex mixtures is normally achieved by overnight incubation with PNGase F at 37 °C. Alternatively, faster release can be accomplished within 10 min using microwave74,75 or within 2 min using immobilized PNGase F under ultrasonication.76 PNGase F works for all kinds of high-mannose, complex, and hybrid N-glycans except the ones with α(1,3)-linked core fucose.77 Another enzyme, PNGase A, can cleave N-glycans with or without α(1,3)-linked core fucose residues. However, the efficiency of PNGase A in N-glycan release from glycoproteins is lower than that from glycopeptides. A novel bacterial enzyme, PNGase H+, has recently been reported and proven to have combined advantages of PNGase F and A at an optimum pH of 2.6.78 At such low pH, however, loss of sialic acid will occur. A variety of endoglycosidases that cleave between the two GlcNAc residues in the core region and leave one GlcNAc bound to the protein are also available. Unlike PNGase F and A, endoglycosidases have specificity for different types of glycans. For example, endoglycosidase H (Endo H) and Endo F1 cleave high-mannose and some hybrid-type oligosaccharides but not complex-type oligosaccharides. While Endo F2 cleaves high-mannose and biantennary complex-type oligosaccharides, and Endo F3 cleaves biantennary and triantennary complex-type oligosaccharides, especially the core-fucosylated structures.79 Enzymatic release of O-glycans is more challenging due to both the diversity of core structures and the lack of enzymes with broad substrate specificity. Commercially available O-glycosidases are only able to catalyze the removal of core 1 or core 3 O-linked disaccharides from serine/threonine residues of glycoproteins.80 O-glycans with extended chains need to be trimmed to core structures by exoglycosidases before releasing using O-glycosidases.
3.1.2. Chemical Release.
For complete and unbiased release of glycans, especially O-glycans, numerous chemical release methods have been developed. Reductive β-elimination under alkaline conditions is the most commonly used liberation approach for the analysis of O-glycans. The released O-glycans in alditol forms can be characterized using MS-based methods. Other approaches such as hydrazinolysis and nonreductive β-elimination can be used to obtain N- and O-glycans in their nonreduced forms for subsequent or one-pot labeling of the reducing end using fluorescent or ultraviolet (UV) tags.81,82 More recently, an oxidative strategy named oxidative release of natural glycans (ORNG) using NaClO from household bleach have been reported to liberate all types of glycans from glycoproteins and glycosphingolipids.83 However, most of these chemical release methods are problematic because they can cause peeling of the released glycans, resulting in altered glycans compositions and poor reproducibility. Goso et al. recently evaluated the release of mucin-type O-glycans using hydrazine, ammonia, or sodium hydroxide treatment under nonreductive conditions.84 Although peeling was minimized when the porcine gastric mucin or bovine fetuin samples were treated with hydrazine gas in the presence of malonic acid, “peeled” products were detected under all of the conditions. In addition, chemical release often introduces degradation of the polypeptide chain, making it impossible to analyze the deglycosylated proteins for glycoproteomic study.
3.2. Glycan Derivatization
Native glycans do not have a chromophore and cannot be directly monitored using fluorescence or UV detection. Label-free qualitative analysis can be achieved by obtaining the accurate masses of intact glycans using MS-based methods. As glycans are composed of monosaccharides with unique masses, the composition of a glycan represented by the numbers of monosaccharide units can be easily deduced. Quantitative analysis can be performed using the absolute ion counts of the compounds. However, due to the variation of glycan structures and existence of labile groups such as sialic acid, derivatization of glycans is often needed to improve the stability, ionization efficiency, and quantitation accuracy for glycomic analysis. A variety of glycan labeling strategies and their use in glycan separation and detection by mass spectrometry have been reviewed by Ruhaak et al.85
3.2.1. Derivatization of Reducing End.
The most commonly used glycan labeling strategy involves derivatization of the reducing end through reductive amination. In this protocol, primary amine group from the label reacts with the aldehyde group of the glycan to form a secondary amine under reducing conditions. A variety of labels have been developed such as 2-aminobenzamide (2-AB), 2-aminobenzoic acid (2-AA), 2-aminopyridine (PA), 2-aminonaphthalene trisulfonic acid (ANTS), and 1-aminopyrene-3,6,8-trisulfonic acid (APTS).86,87 These labels can add a chromophore or fluorophore to the glycan, making it possible to perform quantitation using UV or fluorescence detectors. When applied to MS, the introduced secondary amine group can greatly improve the ionization efficiency of glycans in positive ion mode. Besides reductive amination, labeling of the reducing terminus can also be conducted using 1-phenyl-3-methyl-5-pyrazolone (PMP) or its analogues through Michael addition reaction,88,89 or alternatively using hydrazide labeling reagents such as (carboxymethyl)trimethylammonium chloride hydrazide (Girard’s reagent T).90,91
Recent efforts in glycan reducing end labeling include the development of novel labels, improvement of sample preparation efficiency, and introduction of isotope tags for quantification. Hydrazinonicotinic acid (HYNIC) is a novel derivatization reagent developed by Jiao et al. for glycans analyzed by MALDI-MS.92 Because HYNIC can also act as a matrix, removal of excess reagent and addition of matrix are not needed. Comparison of excess HYNIC with a traditional matrix, DHB, showed that peptide signal was greatly suppressed by HYNIC in this method, enabling direct analysis of the released glycans by MALDI. However, sample cleanup is still needed for LC-MS. Zhao et al. synthesized 10 hydrazino-s-triazine based labeling reagents and compared their efficiencies using maltoheptaose as model.93 The most hydrophobic reagent, n-Pr2N, was found to yield the most enhanced signal and applied to N-glycans released from human serum. Other new labels that can facilitate the glycan enrichment after derivatization were also investigated such as 4-amino-phenylphosphate tag combined with Ti4+-SPE94 and heptadecafluoroundecylamine tag combined with fluorous SPE.95 Jiang et al. developed a global solid-phase approach for the reductive amination of glycans by streamlined glycan extraction, derivatization, and purification on nonporous graphitized carbon sorbents.96 They compared this method with traditional in-solution derivatization using multiple common labels including 2-AA, 2-AB, and 2-amino-N-(2-aminoethyl)-benzamide (AEAB) and found 20–30% increase in glycan discovery. Another strategy for simplifying sample preparation is the one-pot method for simultaneous release and labeling of glycans. This method is generally applied to O-glycans released by nonreductive β-elimination or hydrazinolysis that contain reducing terminus. Labels such as 2-AB and PMP either with or without deuterium label have been reported.81,97,98 More recently, Yang et al. developed a glycoprotein immobilization for glycan extraction (GIG) method that combined a series of enzymatic and chemical reactions including the immobilization of glycoproteins by reductive amination, derivatization of sialic acids by carbodiimide coupling, release of N-glycans by PNGase F digestion, and release of O-glycans by β-elimination and one-pot PMP labeling (Figure 10).99,100 The approach allowed for the simultaneous extraction and analysis of N- and O-glycans from biological samples using a solid support.
Figure 10.
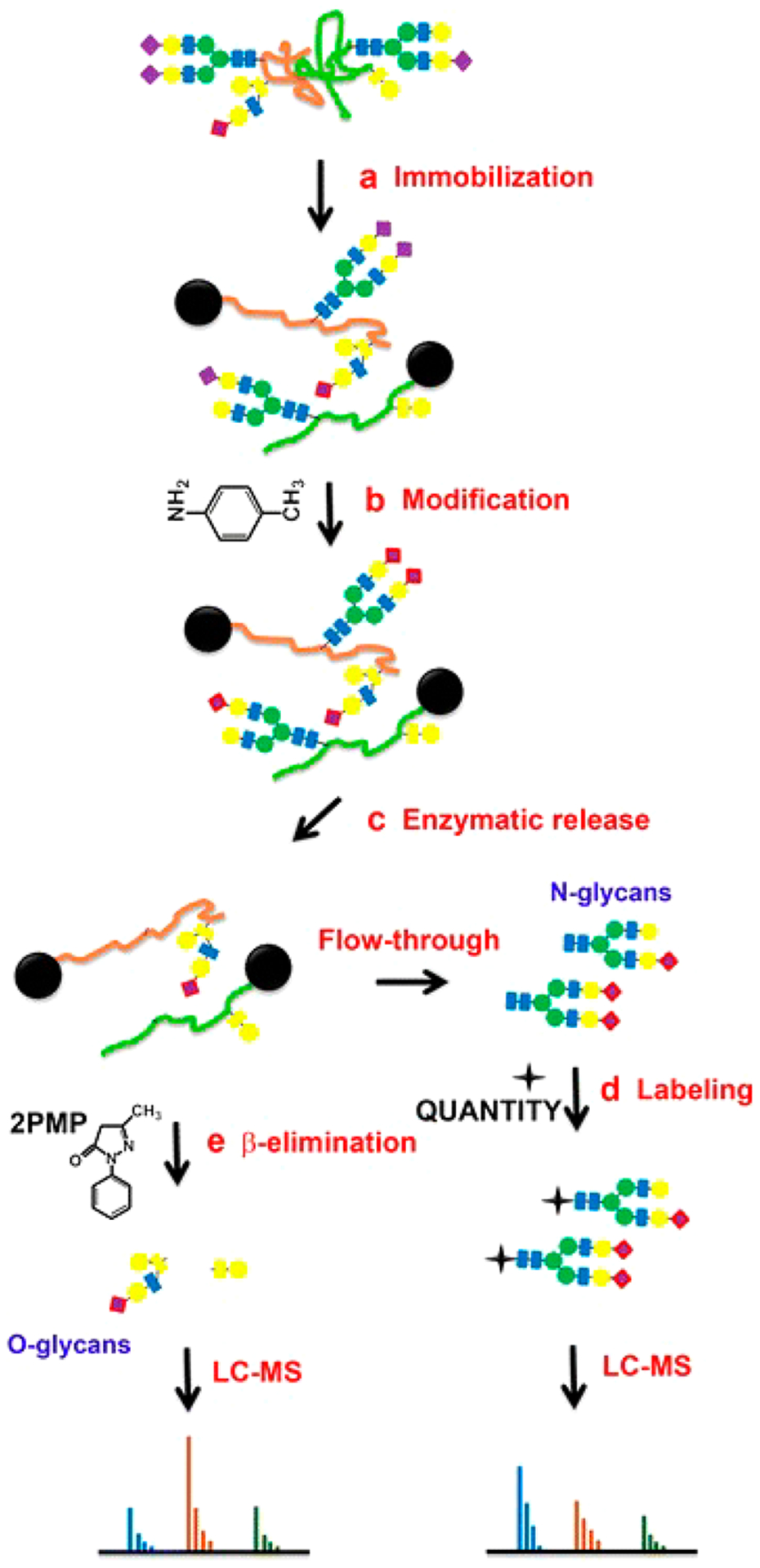
A chemoenzymatic method for sequential releases and analyses of N-linked and O-linked glycans. Reprinted with permission from ref 99. Under Creative Commons license (CC BY 4.0) (http://creativecommons.org/licenses/by/4.0/).
3.2.2. Perderivatization of Oligosaccharides.
Permethylation is another glycan derivatization approach that can be used either alone or together with reducing end labeling approaches. In permethylation, hydrogens on the highly polar hydroxyl groups, amine groups, and carboxyl groups are converted to nonpolar methyl groups.85 Compared to their native forms, permethylated glycans have improved stability and ionization efficiency and are more readily separated by reversed phase liquid chromatography.29 The most widely used permethylation procedure was introduced by Ciucanu and Kerek using dimethyl sulfoxide and methyl iodide with solid hydroxide.101 Modifications and improvements have been made to the protocol such as adding a trace of water to eliminate oxidative degradation102 and performing online reaction using capillary reactors packed with powdered NaOH.103 However, permethylation efficiency is affected by various factors such as differential reactivities of monosaccharides, oxidative degradation, and peeling of the glycans and repeatability of liquid-liquid extractions.103
Recent researches have been focusing on the automation of sample processing and improvement of efficiency. Shubhakar et al. developed an automated high-throughput protocol in 96-well microplate format using a liquid handling robot that can perform N-glycan release, enrichment, permethylation, and extraction.104 Released IgG N-glycans analyzed by HILIC UHPLC after 2-AB labeling or by MALDI-TOF-MS after permethylation showed good correlation for abundant glycans, while slight variation was observed for sialylated glycans (Figure 11). This method was applied to derivatize and extract glycans from 96 biopharmaceutical samples in less than 5 h. Its applicability to complicated biological samples remains to be tested. Hu et al. discussed the low permethylation yield of HexNAc residues using traditional protocol and reported a spin column-free approach that provided comparable or better yields than some widely used spin column-based procedures.105 Permethylation efficiencies of >98% for hexose and >99% for HexNAc residues were obtained for disaccharide standards before this method was applied to plasma samples collected from 20 breast cancer patients and age-matched controls. Nevertheless, the reaction efficiency for anionic residues such as sialic acid was not discussed in this research. The complicated matrix of biological samples, which may significantly affect the reaction efficiency, was also not considered. Incomplete conversion limits the dynamic range because the baseline is chemically noisy with partially methylated species. Furthermore, the issue of isomer separation of permethylated glycans has yet to be adequately addressed.
Figure 11.
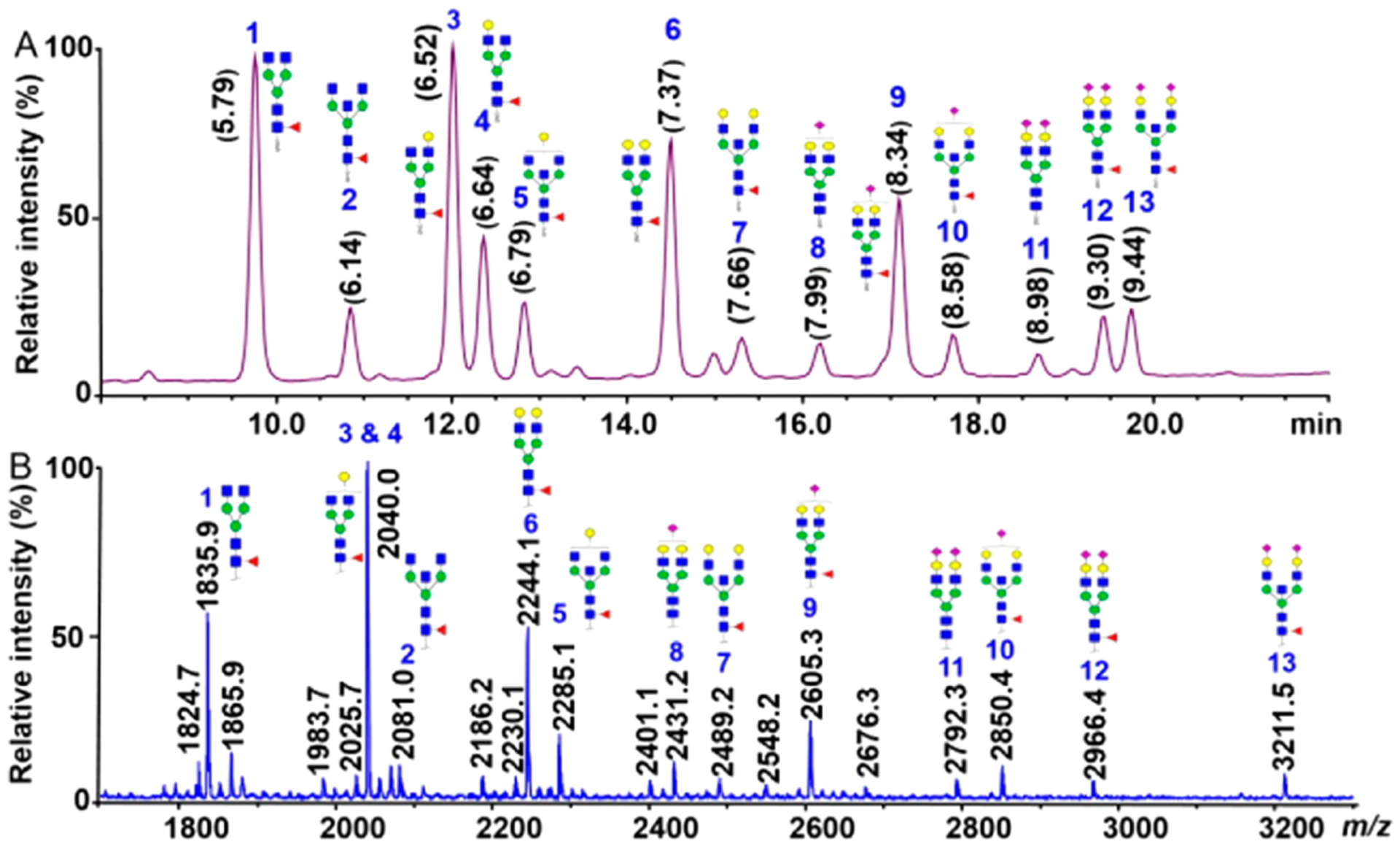
Comparison of IgG N-glycans prepared by an automated workflow using a liquid handling robot and analyzed by (A) HILIC UHPLC after 2-AB labeling and (B) MALDI-TOF-MS after permethylation. Reprinted with permission from ref 104. Copyright 2016 American Chemical Society.
3.2.3. Linkage-Specific Derivatization of Sialic Acids.
A recent advancement in glycan derivatization is the development of linkage-specific sialic acid derivatization strategies to stabilize the labile sialic acid residues and differentiate the α(2,3) and α(2,6) linkages directly by mass. The original approach was reported by Wheeler et al. in 2009, where the glycans were treated with methanol and 4-(4,6-dimethoxy-1,3,5-triazin-2-yl)-4-methylmorpholinium chloride (DMT-MM) to produce methyl esters (+14 Da) for α(2,6)-linked sialic acids and lactones (−18 Da) for α(2,3)-linked sialic acids.106 This method was applied to breast and other cancer serum samples using MALDI-TOF MS or HILIC-MS/MS analyses.107,108 To reduce derivatization time and side reactions, Reiding et al. modified the method by using 1-ethyl-3-(3-(dimethylamino)propyl)-carbodiimide (EDC) and 1-hydroxybenzotriazole (HOBt) as activators to ethyl esterify α(2,6)-linked sialic acids in ethanolic solution under mild conditions.109 Derivatized N-glycans from human plasma were analyzed by MALDI-TOF MS, and 77 glycan compositions corresponding to 108 glycan species with different sialic acid linkages were identified (Figure 12). It was also shown that the efficiency for lactonization of α(2,3)-linked sialic acids was higher in ethanol than in methanol. This method is more rapid and higher throughput compared to the original protocol, making it more readily used in large glycan studies.110–112 However, because the lactones formed by intramolecular dehydration are not stable in aqueous solution, further derivatization such as permethylation and methylamidation is often performed.113–115
Figure 12.
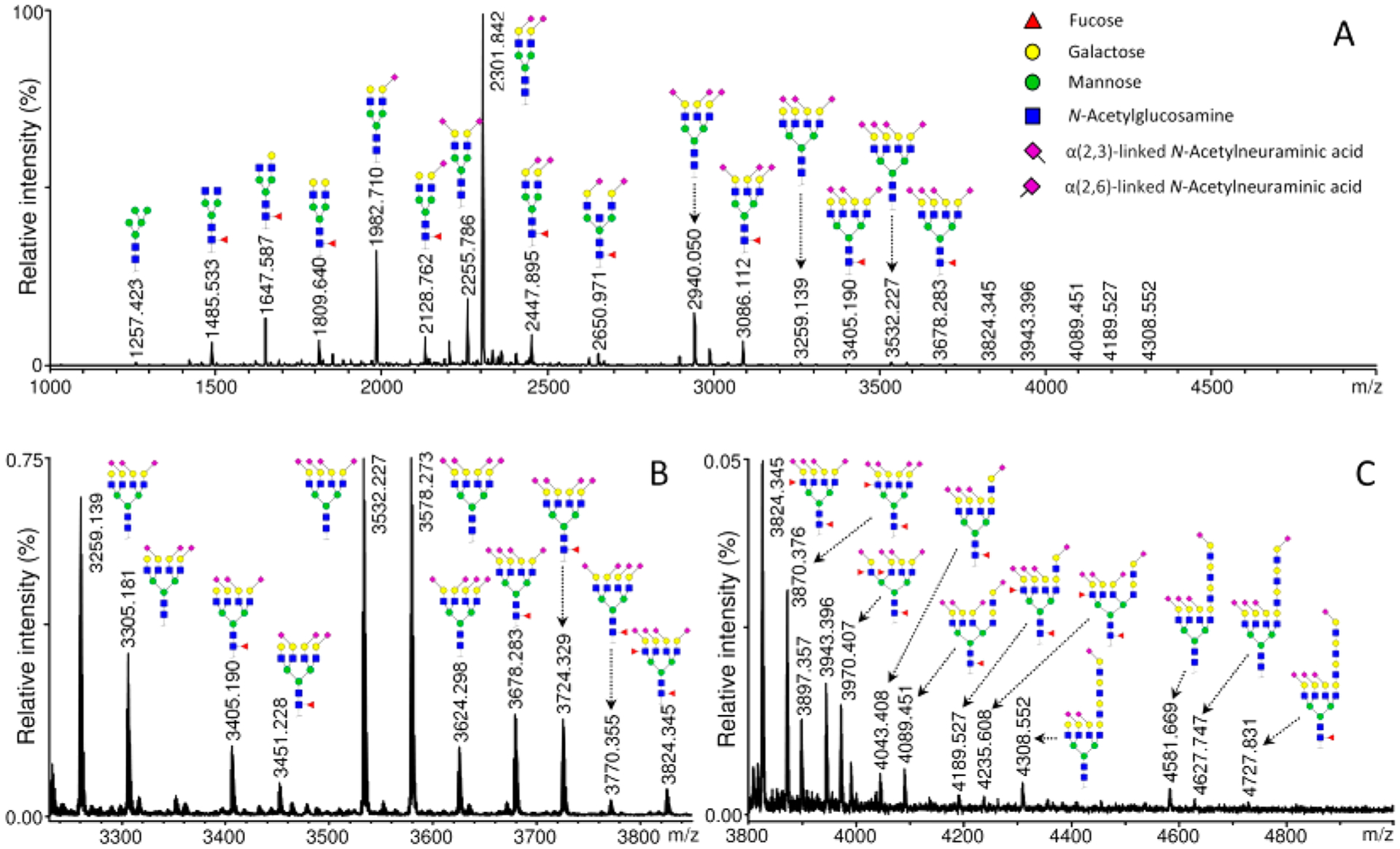
MALDI-TOF MS spectrum of human plasma N-glycans after linkage-specific sialic acid ethyl esterification. Reprinted with permission from ref 109. Copyright 2014 American Chemical Society.
3.3. Separation Methods for Oligosaccharides
Glycans are composed of monosaccharide units that have unique masses. Monosaccharide composition of a glycan can be calculated using its accurate mass obtained by MS detection. However, structural isomers due to variations in the sequence and linkages between the monosaccharides cannot be fully differentiated by mass spectrometry. To characterize the structural heterogeneity of glycans and reduce the signal suppression from concurrent ions, an efficient separation technique is often needed for glycomic analysis, especially for complicated biological samples that may contain hundreds if not thousands of glycan structures.
3.3.1. Liquid Chromatography of Oligosaccharides and Glycans.
Liquid chromatography is currently the most prevalent glycan separation technique for several reasons: (1) Various methods are available for either native or derivatized glycans based on the interaction between glycans and stationary phases, such as RPLC, HILIC, and graphitized carbon LC. (2) Efficient isomeric separation can normally be achieved without the use of nonvolatile salts, making it compatible with MS detection. (3) When adapted to UHPLC and nanoLC systems, the separation efficiency and sensitivity can be further improved. Different chromatographic separation techniques have been applied to glycan analysis, including reversed-phase, normal-phase (hydrophilic interaction), and porous graphitized carbon (PGC) chromatography.
Reversed-Phase Liquid Chromatography.
Reversed-phase liquid chromatography (RPLC) uses hydrophobic materials such as C18 or C8-bonded silica as stationary phase to retain nonpolar compounds. Native glycans are hydrophilic and not well retained in RPLC. Therefore, RPLC separation is only applied to glycans that are permethylated or derivatized with a hydrophobic tag on the reducing end (such as those described above). The strength of interaction between a glycan and the hydrophobic stationary phase is mainly determined by the tagging agent. However, monosaccharide composition and linkage also influence the retention, enabling separation of different glycan types and structural isomers. Pabst et al. compared the separation of IgG N-glycans derivatized by PA, 2-AB, and p-aminobenzoic acid ethyl ester (ABEE), respectively, using a C18 column.86 PA and ABEE labeled glycans were better separated than 2-AB labeled ones, while the effect of galactosylation on elution order was opposite for PA and ABEE labeled glycans. Walker et al. derivatized plasma N-glycans with 4-phenethylbenzohydrazide (P2PGN) via hydrazone formation and compared the separation efficiencies of nanoflow RPLC and HILIC.116 A total of 42 glycan compositions were detected by both methods, while 18 additional compositions were detected only by RPLC-MS with narrower peak widths. RPLC has also been applied to the separation of permethylated glycans.117–119 Improved chromatographic resolution with increasing temperature117 and linear correlation between the retention times and glucose units of N-glycans118,119 were observed. A recent review by Vreeker and Wuhrer have discussed the application of reversed phase separation methods to released glycans.120
Hydrophilic Interaction Liquid Chromatography.
Glycans contain multiple hydroxyl groups that can interact with HILIC stationary phases like amine, amide, or zwitterion-bonded silica through hydrogen bonding, dipole-dipole, or ion-dipole interaction. HILIC separation has been applied to the analysis of underivatized glycans derived from fetal bovine serum106 and human plasma121,122 in conjunction with MS analysis. Lam et al. built an online-coupled RP-HILIC system to separate peptides, glycopeptides, and glycans from a single injection.123 Glycomics data obtained from the same sample could provide valuable complementary information for studying the glycoproteome. To improve the sensitivity and isomeric separation efficiency, however, glycans are often derivatized on the reducing end before HILIC analysis. With a UHPLC platform developed by Ahn et al. using a column packed with <2 μm amide sorbent, excellent separation on the basis of both sequence and linkage was achieved for 2-AB labeled glycans with fluorescence detection.124 A total of 143 human serum N-glycans were separated and assigned with a 30 min HILIC gradient using fluorescence detection.125 This platform has been automated, coupled to mass spectrometry, and widely applied to glycomic analysis of monoclonal antibodies.126–128
Porous Graphitized Carbon Chromatography.
PGC is currently the most widely used stationary phase for the purification and separation of underivatized glycans. Comparison of PGC separation of reduced glycans and ZIC-HILIC separation of 2-AB derivatized glycans from monoclonal antibodies has demonstrated that PGC is a more efficient sorbent for the separation of isobaric glycans with various degrees of mannosylation and galactosylation.129 PGC has been used extensively to separate sialylated N-glycans from bovine fetuin.130 Up to six isomers were observed for a disialylated glycan composition. Isomers of sialylated N-glycans from serum131,132 and cell lines133,134 were also efficiently separated by PGC employing LC-MS platforms. The retention of a glycan on PGC is greatly dependent on its size, monosaccharide composition, and linkage. For example, sialylated and/or fucosylated complex type glycans generally elute later than high-mannose type glycans.74,135 (Figure 13) Increased retention times are often associated with higher degrees of sialylation and fucosylation or lower degrees of mannosylation.131 The α(2,3)-linked isomers of sialylated N-glycans were found to elute later compared with the α(2,6)-linked isomers.133 Although peak broadening was occasionally observed on PGC for highly sialylated species,121 Kronewitter et al. were able to characterize polysialylated glycans containing up to 18 sialic acid residues with a 60 cm graphite fused silica column.136 Using improved sample preparation steps and optimized LC conditions, they identified 290 glycan compositions and observed 994 potential isomer peaks from human serum.
Figure 13.
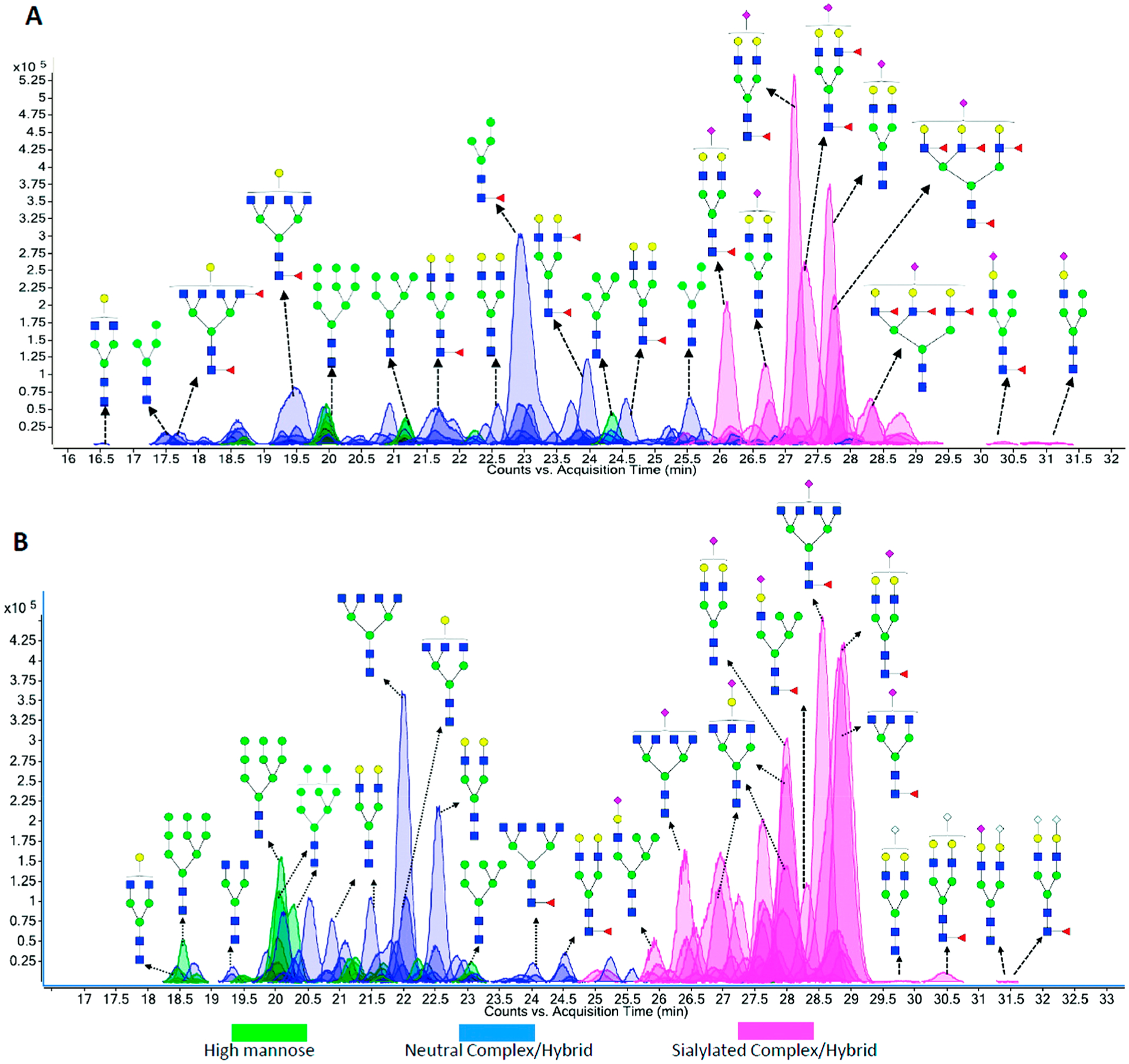
PGC separation of N-linked glycans released from (A) human milk and (B) bovine milk. Reprinted with permission from ref 135. Copyright 2012 American Chemical Society.
3.3.2. Capillary Electrophoresis of Oligosaccharides and Glycans.
Capillary electrophoresis has long been recognized as a highly efficient separation technique that has been applied to various types of compounds. The analysis of native or labeled glycans has been accomplished by employing several electromigrative separation techniques, including capillary zone electrophoresis (CZE), capillary gel electrophoresis (CGE), micellar electrokinetic chromatography (MEKC), and capillary electrochromatography (CEC).137 The migration rate of a glycan is determined by a number of factors including its own structure as well as the separation mechanism, strength of electric field, buffer pH, and column temperature. Guttman et al. studied the effect of column temperature on the structure specific separation of glycans using capillary gel electrophoresis138 and greatly enhanced the separation of APTS labeled glycans of biotherapeutic interest using an optimized temperature gradient.139 Feng et al. discovered the high orthogonalities between the CZE and MEKC separation of glycans and developed a multiplexing technique by combining these two mechanisms to achieve higher peak capacity for the analysis of complicated glycan mixtures.140 The coupling of CE to MS, however, has been challenging due to the low flow rates employed in CE.141 This issue has been partially addressed by the implement of sheath-flow142,143 or sheathless-flow144,145 interfaces with compromised sensitivity or reproducibility. Development of novel low-dilution sheath-flow or liquid junction interface designs that can improve sensitivity while creating stable spray is of great interest for CE-MS. Chen and co-workers reported a flow-through microvial interface146 that was applied to underivatized N-glycans released from IgG and recombinant human erythropoietin (rHuEPO).147 The presence of O-acetyl groups and polyLacNAc chains were observed for rHuEPO-derived glycans. Another type of interfaces are microfluidic devices that fully integrate CE separation system with a nanoESI source on a single chip.148 These integrated devices do not require precise alignment by the user and are therefore more readily commercialized. Khatri et al. recently applied a commercial ZipChip CE-ESI device for the analysis of released glycans and glycopeptides from standard glycoproteins.149 However, the application of microfluidic CE devices to glycans from complicated biological samples has not yet been widely pursued.
3.3.3. Ion Mobility of Oligosaccharides and Glycans.
Ion mobility-mass spectrometry (IMS-MS) has been of great interest in the past decade for its potential applications in glycomic, proteomic, and glycoproteomic analyses. The ability of IMS to separate ions in the gas phase based on their differences in shape and charge makes it possible to resolve some structural isomers. A review by Gray et al. has discussed different types of ion mobility instruments and their applications in the analyses of free saccharides, released glycans, and glycoconjugates.150 A more recent mini-review by Hofmann and Pagel explores the history and recent advancements of glycan analysis by IMS-MS.151 Earlier efforts for the isomeric separation of carbohydrates were focused on mono-, di-, and trisaccharides from simple mixtures.152–155 The differences of the compounds in shape can be represented by their collision cross sections (CCS). Fenn and McLean published a CCS database of 303 carbohydrates including three sets of isobaric isomers.156 Certain structural patterns such as branching and 1–3 linkage versus 1–4 linkage could result in more compact structures and therefore shorter drift times.
Studying N-glycan isomers using IMS-MS is more challenging due to the high structural similarities accompanied by large variations in glycan structures. The existence of conformers, while fundamentally interesting, further interferes with structure assignment. The earliest studies on IMS-MS of N-glycans were published by Clemmer and co-workers in 2008.157,158 Glycans from human serum were directly ionized by an ESI source and analyzed by IMS-MS with a 1 m long drift tube. Separated features were assigned based on their m/z and drift times and compared between control and liver disease groups.157 Another study by the same group focused on the separation of Man7 glycan isomers.159 The broad drift time distribution of this composition was assigned to four potential isomers, which were distinguished by their fragment ion drift time distributions. To facilitate the identification and assignment of glycan structures and isomers, Pagel and Harvey reported a calibration protocol for calculating CCS values of sodiated N-glycans using traveling wave (TW) IMS instruments.160 They published a CCS library for N-glycans released from standard glycoproteins. Certain structural isomers such as those for Hex4HexNAc3 and Hex5HexNAc4 could be distinguished based on their CCS values. The list was expanded to over 350 CCS values corresponding to 70 precursor glycan structures and added to a freely accessible database, GlycoMob.161
To increase the peak capacity and identify more structures, IMS-MS is coupled to LC as a complement to chromatographic separation.162,163 However, IMS-MS currently has limitations as a separation technique for glycomic and glycoproteomic analyses. First, overlapping peak is often observed for isomers, especially for glycans, with only minor differences in the position or linkage of one monosaccharide residue.164,165 Several strategies have been developed to increase the resolution of IMS separation. One effective method is to use metal cations as charge carriers because the formation of metal cation adducts with isomeric glycans can induce greater differences in the conformations and thereby improve separation by IMS.166,167 Huang and Dodds systematically studied the ion-neutral collisional cross sections of carbohydrate isomers as group I168 and group II169 metal ion adducts. Different metal ion adducts were found to be useful in separating isomeric groups, while the most commonly used metal ion adduct, sodium, was not necessarily as effective for improving IMS resolution. Morrison et al. evaluated the effects of seven metal ion adducts on the separation of five isomeric tetrasaccharide-alditols.170 Transition metal cations such as Co, Cu, and Mn notably improved the separation. Similar results were obtained by Zheng et al. for oligosaccharide standards and synthetic O-glycans.171 Despite these improvements, baseline separation of glycan structural isomers by ion mobility remains difficult. Additionally, IMS-MS data is often further complicated by the existence of conformers that overlap with structural isomers.172 Molecular modeling techniques, used to calculate theoretical cross sections, may provide insightful information regarding experimental collisional cross sections of the glycans.158
3.4. Imaging MS of Glycans
The glycan and glycoprotein distributions in living organisms are rarely homogeneous. The spatial distributions of glycans in tissues and in organisms can provide fundamental understanding of the glycobiology as well as locate specifically diseased areas. Imaging mass spectrometry (IMS) has emerged as a tool for this purpose owing to its high sensitivity and specificity for detecting glycan analytes.173 Mass spectrometry techniques for imaging include secondary ion mass spectrometry (SIMS), MALDI-MS, desorption electrospray ionization mass spectrometry (DESI-MS), and direct analysis in real-time mass spectrometry (DART-MS). Among them, MALDI-MS is the most widely used method and has already been applied to a variety of biomolecules.174 Imaging of glycans in tissues using MALDI-IMS is relatively new, but important progress has been made by Drake and co-workers in the past few years. The original workflow involved on-tissue release of N-glycans by spraying PNGase F onto prepared tissue slices, followed by matrix application and MALDI-IMS analysis.175,176 Images of individual glycans could then be extracted to illustrate their distributions within the tissue (Figure 14). This method has been applied to tissues in different formats such as frozen tissues, formalin-fixed paraffin embedded (FFPE) tissue blocks, and tissue microarrays for biomarker analysis.175,177,178 To stabilize labile sialic acid residues and characterize their linkages, in situ linkage specific derivatization was performed on FFPE tissues.179 Additional multiply sialylated species with m/z > 2500 were detected after derivatization. A new application is a multimodal approach to sequentially analyze N-glycans and proteins in the same tissue by digestions with both PNGase F and trypsin.180 For more detailed structural identification, however, other techniques such as permethylation, exoglycosidase digestion, and LC-MS/MS are often necessary to provide complementary information for MALDI-IMS.175,181
Figure 14.
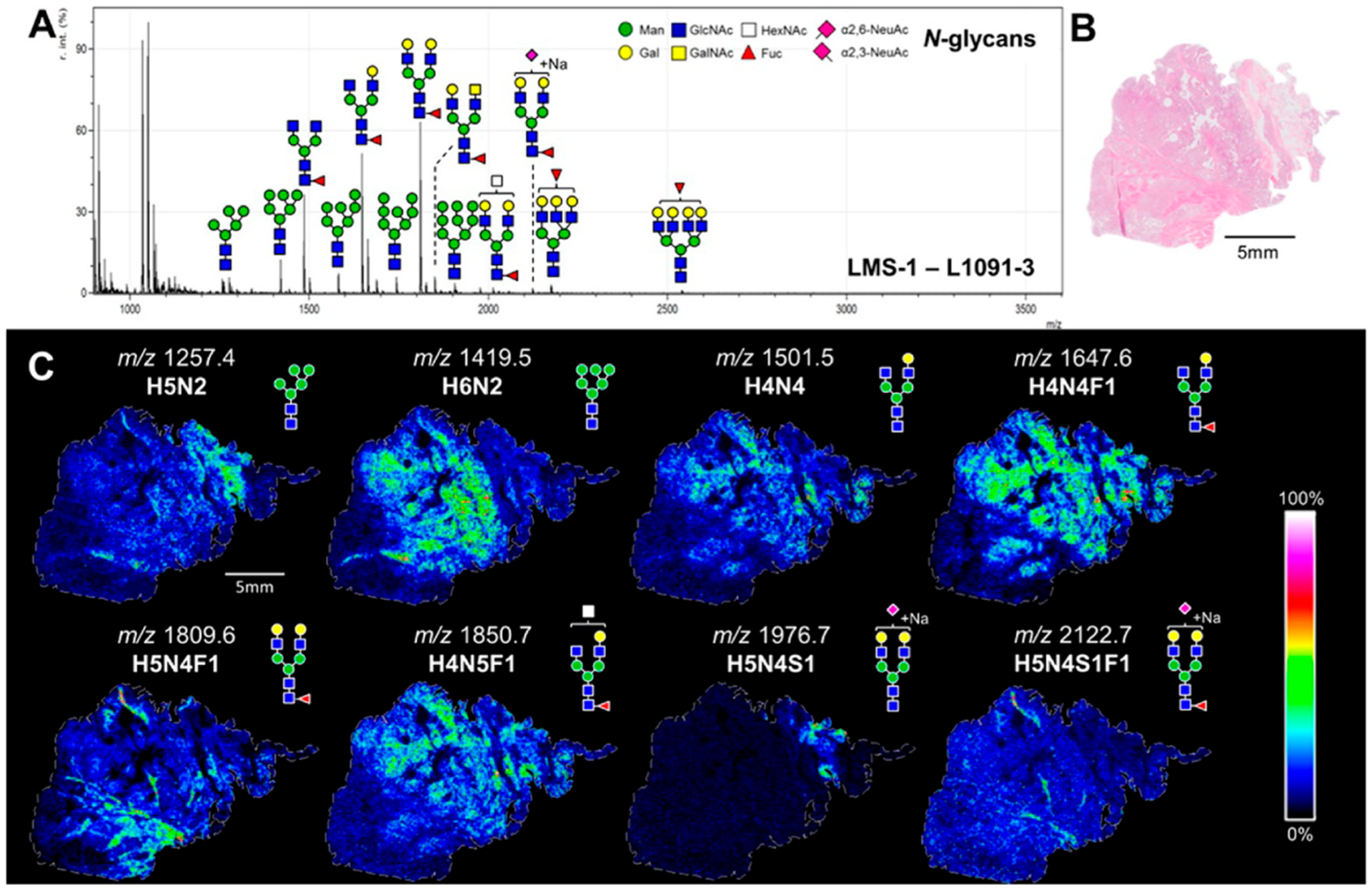
MALDI-IMS for visualizations of the distribution of various N-glycans throughout a leiomyosarcoma tissue. Reprinted with permission from ref 180. Copyright 2016 American Chemical Society.
3.5. Structural Elucidation of Glycans
Although separation and linkage-specific derivatization techniques can yield some structural information such as the linkage of sialic acid109 and position of fucose,182 the structural elucidation of glycans is still challenging due to the complexity and large variability in the structures. Complete structural elucidation requires extensive determinations of monosaccharide composition, sequence, branching, and the unique linkages of each monosaccharide residue. There are two commonly used approaches for structural analysis of N- and O-glycans. The first is based on tandem MS techniques including the low and high energy dissociation methods. The second one involves enzymatic digestion using exoglycosidases and detection by UV, fluorescence, or MS.
3.5.1. Structural Elucidation by Tandem MS.
Several aspects of the data obtained from MS and tandem MS can be used to elucidate partial glycan structures. For example, monosaccharide composition of a glycan can be easily deduced from the accurate mass of the quasimolecular ion. The connectivities between monosaccharides can be determined by the sequential cleavage of glycosidic bonds under low-energy conditions such as CID and IRMPD,183,184 and the glycosidic linkage positions can be obtained through cross-ring cleavages under high-energy fragmentation conditions such as electron dissociation methods (ExD) and UVPD.72,185,186 Isomers having variable linkage of a single monosaccharide residue are identified by MS/MS based on the variations in distinct bond energies.187–189 However, interpretation of glycan tandem MS spectra can be complicated by the rearrangement of monosaccharide residues, especially fucose, and the presence of spurious MS peaks.190 One strategy to address this issue is to develop searchable MSn spectra libraries by using specifically developed software tools. A universal software for annotating glycan structures based on tandem MS data is still lacking, perhaps due to the diversity of glycan structures and the large variations in fragmentation patterns under different conditions.
Tandem MS alone is not sufficient for complete structural elucidation of glycans because it cannot provide some essential information such as the anomeric nature of the glycosidic bonds and the stereochemistry of the monosaccharides. It can be used in combination with other techniques to enhance the efficiency and accuracy for glycan identification. Fellenberg et al. applied an integrated LC-MS/MS and 1D 1H NMR workflow to the analysis of 10 desialylated underivatized glycans from bovine fibrinogen.191 By comparing and combining data from both methods, the branching and linkage information partially assigned by MS/MS was further confirmed by an NMR database search. LC-MS profiles were additionally used to determine the sample purity for NMR interpretation. An improved method by the same group was able to extract the NMR spectra of N-glycans from heavily overlapping chromatographic separations by mathematically dissecting the NMR spectra obtained from chromatographic fractions.192 Due to the limitations of NMR sensitivity, however, only abundant glycans could be unambiguously characterized.
Glycan microarray is another technique that can be combined with tandem MS for detailed structural analysis. An approach named metadata-assisted glycan sequencing (MAGS) was developed by Cummings and co-workers,193 where undefined glycan arrays were interrogated by glycan-binding proteins to filter glycans with specific motifs or significant biological relevance. Additional structural information on specific glycans could be obtained further by tandem MS.
3.5.2. Structural Elucidation by Exoglycosidase Digestion.
Exoglycosidases cleave glycosidic bonds of monosaccharide residues at the nonreducing end of oligosaccharides or polysaccharides. There is a variety of exoglycosidases; some have general reactivity, for example, they may cleave all terminal galactoses, while others are highly specific to one type of glycosidic linkage with defined anomeric character. Therefore, the linkage of a terminal monosaccharide residue in a glycan can be unambiguously determined by the detection of expected product after exoglycosidase digestion. Complete structure of a glycan can then be elucidated by sequential or enzyme matrix digestions using a series of exoglycosidases. The combination of exoglycosidase digestion and mass spectrometry was extensively used by Wu et al. to create a library of structures for human milk oligosaccharides.10,11 This approach also has been widely used in conjunction with various separation and detection methods such as CE-LIF,194,195 HPLC-LFD,196 and LC-MS.197 Exoglycosidase digestion has been combined in a rapid-throughput analytical HPLC platform called GlycoBase and autoGU by Rudd and co-workers.198,199 GlycoBase is a database containing more than 350 2-AB-labeled N-linked glycan structures (117 from human serum) and their corresponding HILIC elution times expressed as glucose unit values. AutoGU automatically assigns glycan structures using combined data from a set of sequential exoglycosidase digestions. An experimental strategy combining these two tools helped less experienced glycobiologists to identify glycan structures from HPLC data without the need for tandem MS knowledge. The method has some drawbacks because limited isomer separation with HILIC columns can complicate the identification of coeluting isomeric glycans. Aberrant compounds from the matrices might also interfere with glycan analysis for complicated biological samples. Aldredge et al. addressed these issues by combining exoglycosidase digestion with nanoLC-MS/MS by employing PGC as stationary phase.200 A N-glycan pool from human serum glycoprotein standards was fractionated by off-line HPLC and subjected to sequential exoglycosidase digestions. Compounds before and after digestion were analyzed by nanoLC-MS/MS to confirm the compositions by both accurate masses and tandem MS spectra (Figure 15). The effective separation with PGC and sensitive detection with nanoLC-MS/MS facilitated the construction of a serum N-glycome library that allowed rapid identification of exact glycan structures simply by matching their nanoLC retention times and accurate masses. The initial library contained over 300 entries with 50 completely elucidated structures derived from serum glycoprotein standards. A more refined library was later created with a larger number of elucidated structures using glycans released directly from human serum.9 The relative abundances and variations in the N-glycans from several individuals were determined using this library. More recently, Abrahams et al. used a similar approach to assess the elution behavior of previously characterized N-glycans derived from standard glycoproteins on capillary PGC-LC-MS/MS.201 The glycan retention times along with their associated MS/MS spectra were added to an elution centric database GlycoStore. Enzymatic digestion methods for glycan structural elucidation will continue to be developed and facilitate the extension of existing glycan libraries. These libraries will greatly improve the efficiency and accuracy of glycomics experiments in biomarker and biotherapeutic studies.
Figure 15.
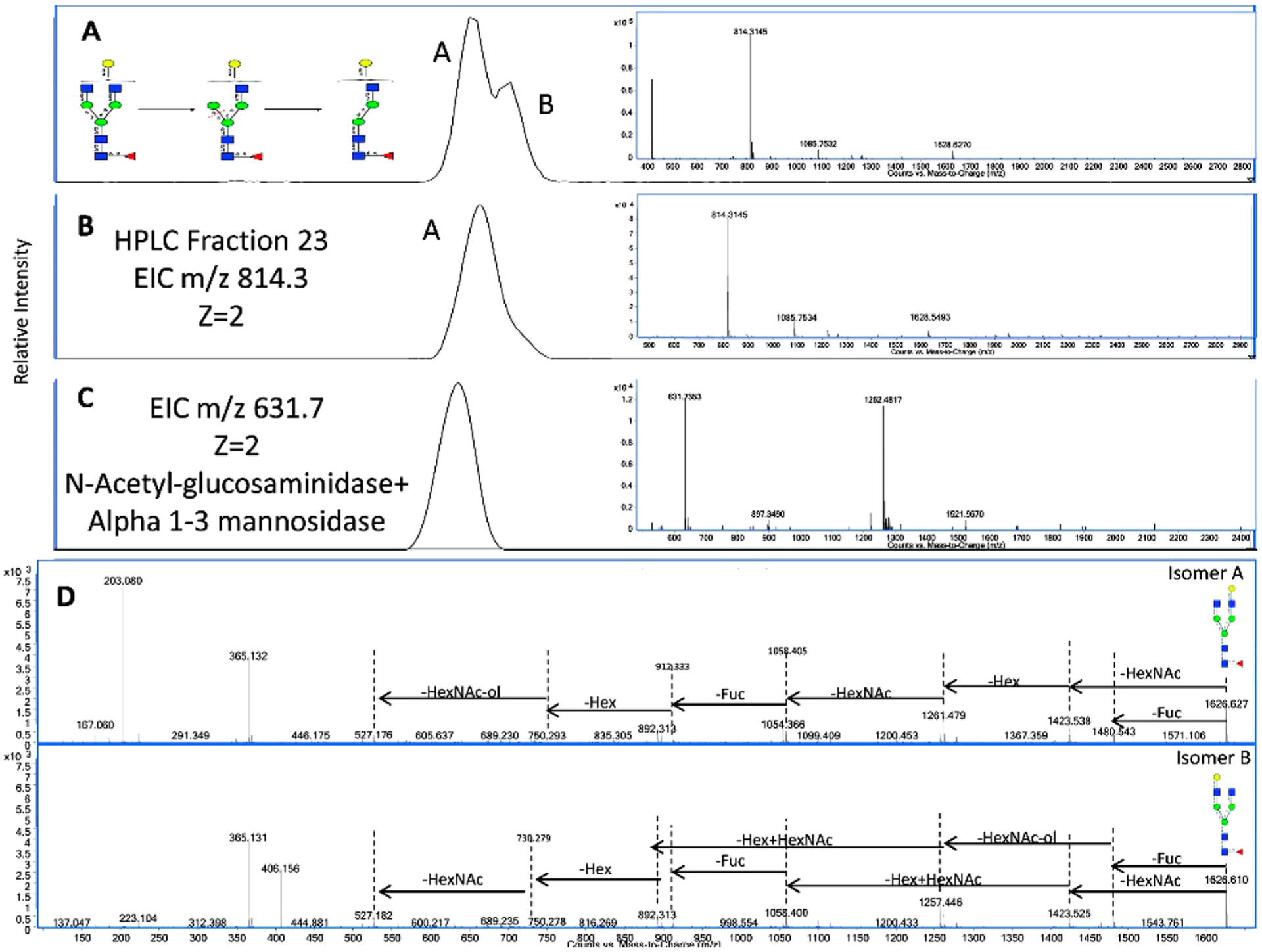
N-Glycan structural elucidation by exoglycosidase digestion and tandem MS. (A) EIC of m/z 814.29 shows two isomers for Hex4HexNAc4Fuc1. (B) HPLC fractionation to isolate one isomer. (C) EIC of exoglycosidase digestion product with m/z 631.7. (D) Differential tandem MS spectra for the identified isomers. Reprinted with permission from ref 200. Copyright 2012 American Chemical Society.
3.6. Minimum Information Required for a Glycomics Experiment (MIRAGE)
The rapid development of sample preparation methods, as well as separation, ionization, and detection techniques for glycomics studies has brought valuable tools for understanding the key roles of glycans in biological processes. However, the diversity of methods has also led to inconsistencies between laboratories and scientists in generating and reporting glycomics data. To establish standards for the glycobiology community, a group of scientists published the minimum information required for a glycomics experiment (MIRAGE) guidelines for reporting mass spectrometry-based glycoanalytic data.202 The guidelines were developed according to the processes published in 2014.203 Five major sections were described as essential information that authors should provide in their glycomics experiment reports; they include ion source conditions and software tools for data interpretation, etc. Recently, more MIRAGE guidelines for sample preparation204 and for reporting glycan microarray-based data204 were also added. Although these guidelines have not yet been widely applied to glycomics studies, they provide an important starting point and standardized criteria for authors and reviewers for handling glycoanalytic data.205
4. MS METHODS FOR GLYCOPROTEOMICS
The characterization of glycoproteins can provide both glycan and protein information, thereby linking glycomic and proteomic analyses. Glycoproteomics combines the tools of proteomic analysis with site-specific glycomic information. However, it also takes the difficulties in protein analysis and compounds them with the difficulties associated with glycan analysis.
The analyses of peptides and glycopeptides, as in bottom up, provide the simple route for the development of glycoproteomic platforms. Site-specific analysis of glycoproteins employing intact glycopeptides is challenging due to the presence of multiple glycosylation sites and the glycan heterogeneity associated with each site. Glycopeptide analysis by mass spectrometry is also complicated by several factors. Glycans tend to diminish severely the ionization efficiencies of the respective peptides. The many glycoforms distribute further the peptide signals over several species. Tandem MS methods were not sufficient to obtain both peptide and glycan information simultaneously.
Early attempts at glycoproteomics released glycans from the carrier proteins or peptides so that the glycans and deglycosylated peptides can be analyzed separately, or the glycans not at all.206 This proteomic-centered approach simplified proteomics data but typically lost glycan information. However, with the emergence of newer fragmentation methods and bioinformatics tools focused on glycosylation, determining glycosite heterogeneity and occupancy has become more feasible. The improvements in enrichment and separation methods have also contributed greatly to the increased sensitivity and coverage.
4.1. Enrichment Methods for Sample Preparation
Enrichment methods have been used primarily to determine glycosylation in specific glycoproteins of interest. In this targeted approach, glycans and glycopeptides are released to obtain protein-specific glycosylation. In untargeted glycoproteomic approaches, glycopeptides are enriched to minimize competition for charge with the more abundant and more ionizable nonglycosylated peptides. Ion suppression from highly abundant peptides and glycan heterogeneity at the glycosylation site makes the detection and identification of glycoproteins challenging. To improve the sensitivity and coverage in glycoproteomic analysis, a diverse array of enrichment methods has been developed such as immunoaffinity and single-lectin affinity for targeted enrichment of one or a small group of glycoproteins, and multilectin affinity, HILIC, and covalent interaction-based methods for nonspecific enrichment. Several reviews have covered different aspects of these methods.207–210
4.1.1. Enrichment Based on Immunoaffinity.
Antibodies or antibody-like agents, immobilized on beads or columns, are used to target glycoproteins through immunoprecipitation or immunoaffinity chromatography. The purified glycoproteins or glycopeptides are typically quantified by traditional immunoassays such as enzyme-linked immunosorbent assay (ELISA) but are increasingly probed by mass spectrometry techniques. The development and application of technologies combining immunoaffinity assays and mass spectrometry for quantitative proteomics in biological fluids have recently been reviewed by Borchers and co-workers.211
Immunoaffinity chromatography or immunoprecipitation as an enrichment step for subsequent MS analysis can greatly improve the sensitivity of MS detection for target glycoproteins. Immunoprecipitation using antibody-bound agarose beads has been used to obtain comprehensive glycan maps of selected glycoproteins. For example, Hong et al. used antihuman-IgA and antihuman-IgM bound to agarose to enrich IgA and IgM from human serum. Glycopeptide mapping was then performed on the enriched glycoproteins to identify glycopeptides that could be quantified by MRM.20 A more common application of immunoaffinity enrichment is to purify specific glycoprotein of interest for biomarker discovery and validation. Recent examples include the characterization of clusterin in the plasma of clear cell renal cell carcinoma patients before and after curative nephrectomy,212 glycoprofiling of intact transferrin for diagnosis and subtype identification of glycosylation in the congenital disorders, and213 site-specific and linkage analyses of fucosylated N-glycans on haptoglobin in the sera of patients with various types of cancer.214 In general, immunoaffinity enrichment can greatly improve the sensitivity for the detection of targeted protein or glycoprotein with acceptable reproducibility. Due to the high specificity of antibody capture and limited number of available antibodies, however, immuno-enrichment assays are not suited for untargeted glycoproteomics.
4.1.2. Enrichment Based on Lectin Affinity.
Lectins are carbohydrate binding proteins that are widely used for selectively capturing and identifying glycoproteins. Unlike antibodies that may target individual proteins, each type of lectin can enrich a group of glycopeptides or glycoproteins with a unique glycan motif. Thus, lectin-based enrichment methods can provide a wider coverage of glycoproteome compared to immunoaffinity enrichment. Various lectin-based capturing techniques have been used in combination with mass spectrometry for glycoproteomic analysis, including lectin affinity chromatography, lectin microarrays, and lectin magnetic bead arrays.
Lectin affinity chromatography (LAC) can be performed using a single type of lectin to extract selectively one group of glycoforms or a combination of multiple types of lectins for the enrichment of a broader glycoproteome. Single-lectin affinity chromatography is generally applied to complicated biological samples such as serum, plasma, and tissue lysate to monitor specific glycosylation changes in deseases.215,216 A variety of immobilized lectins with high specificities are commercially available for this purpose such as Sambucus nigra agglutinin (SNA) lectin for α(2,6) sialylated glycans and Lens culinaris agglutinin (LCA) lectin for core-fucosylated glycans.216,217 To simplify data processing, the enriched glycoproteins were often deglycosylated before analysis, with the assumption that all of the identified proteins containing DXS/T concensus sequence were originally modified by the specific glycan epitope.218 One problem with this approach is that the lectins may have broader specificity than expected, especially for complicated biological samples. The presence of naturally occurring aspartic acid and nonspecifically bound glycoproteins will often lead to misinterpretation of glycoproteomic data.
For more global analysis of glycoproteome, multilectin affinity chromatography (M-LAC) that can simultaneously capture glycoproteins with various glycan motifs was developed by mixing multiple lectins of interest. M-LAC can be combined with other depletion methods when applied to complicated biological samples such as human plasma.219 For example, Gbormittah et al. incorporated 12 high-abundance proteins (12P) depletion with multilectin affinity enrichment by employing equal amounts of Aleuria aurantia lectin (AAL), SNA lectin, and Phaseolus vulgaris leucoagglutinin (PHA-L). Tryptic digests of M-LAC bound and unbound proteins were separately analyzed using nanoLC-MS/MS to compare the glycoproteome and proteome of clear cell renal cell carcinoma plasma before and after curative nephrectomy.220 Alternatively, M-LAC bound glycoproteins could be eluted sequentially to fractionate glycoproteins specifically bound to each individual lectin. Totten et al. incorporated this approach with isotopic labeling during alkylation for relative quantitation of glycopeptides from two plasma samples.221 The use of multiple lectins, however, still cannot fully resolve the glycopeptide loss caused by the biases of individual lectins during untargeted enrichment. Other complementary enrichment methods are often needed to completely cover different types of glycoforms.222
Lectin microarrays have proven to be a simple and rapid technique that has been applied to the high-throughput detection and differentiation of glycoforms for disease diagnostics.223 When combined with in situ proteolysis and MALDI-TOF MS analysis, lectin array-based detection and identification of the specifically bound proteins can be achieved.224–226 More recently, lectin magnetic bead array (LeMBA) platforms have been developed to facilitate the isolation of lectin-bound glycoproteins. Isolated glycoproteins can be readily eluted and digested before LC-MS/MS analysis.227–229 Shah et al. reported a semiautomated LeMBA-based pipeline for serum glycoprotein biomarker discovery and candidate qualification.230 Serum samples were incubated with magnetic beads coated with 20 different lectins to pull down glycoproteins, followed by on-bead trypsin digestion, proteomic analysis by nanoLC-MS/MS, and candidate glycoprotein quantitation by LC-MRM-MS. However, glycoproteome mapping and quantitation only performed at the protein level in this and other similar studies.228
4.1.3. Enrichment Based on HILIC.
Glycopeptides contain hydrophilic glycan moieties and are thereby better retained on polar materials than nonglycosylated peptides. This feature allows glycopeptides to be isolated from the highly abundant and hydrophobic peptides using HILIC. Various HILIC stationary phases have been applied to glycopeptide enrichment such as silica-based functional groups including amine,231 amide,232 cyclodextrin,233 maltose,234,235 and zwitterion,236–239 and polysaccharide-based materials including cellulose,240 cotton wool,241 and sepharose.221,242,243 Compared to LAC, HILIC generally has broader glycan specificity that enables the enrichment of a wider range of glycopeptides. However, the strength of interaction between a glycopeptide and the HILIC material is largely dependent on the hydrophobicity of the underlying peptide backbone as well as the size and composition of the glycan moiety. Some O-glycopeptides and large tryptic glycopeptides are very weakly retained on HILIC and may be lost during enrichment.244
To increase the effectiveness of the enrichment and thereby increase the coverage of glycoproteome, efforts have been made to refine HILIC techniques by combining it with other enrichment methods. Electrostatic repulsion hydrophilic interaction chromatography (ERLIC), originally introduced by Alpert245 for isolation of phosphopeptides, has become a popular technique for glycopeptide enrichment.246–248 Its combined action of hydrophilic and electrostatic interactions allows for the simultaneous enrichment of neutral and sialylated glycopeptides.127 Totten et al. recently compared the efficiencies of strong anion exchange ERLIC (SAX-ERLIC), M-LAC, and Sepharose-HILIC for the enrichment of N-glycopeptides from human plasma.221 SAX-ERLIC was found to be the most robust method for glycopeptide enrichment, yielding over 800 unique glycopeptides from 95 plasma glycoproteins (Figure 16). This method could provide a potentially effective tool for system-wide site-specific mapping of the glycoproteome. Another refinement is the use of magnetic beads and nanoparticles functionalized with various hydrophilic ligands. This includes cross-linked cyclodextrin-metal-organic frameworks,74 chitosan-coated magnetic colloidal nanocrystal clusters,249 and graphene oxide-polyethylenimine-Au-L-Cys ZIC-HILIC nanocomposites.250 The high biocompatibility and large surface area of these nanomaterials make them readily applicable to biological samples. However, no significant improvement in glycoproteome coverage has been reported for these materials compared to conventional HILIC approaches.
Figure 16.
Comparison of different glycopeptide enrichment and detection methods showed ERLIC enrichment with HCD/ETD fragmention yielded the largest percentage and number of glycopeptides from palsma. Reprinted with permission from ref 36. Copyright 2017 American Chemical Society.
A more direct approach toward improving glycopeptide coverage is to use a combination of different enrichment regime. A study by Calvano et al. compared M-LAC and ZIC-HILIC enrichment protocols and discovered that different groups of glycopeptides were enriched when a single method was used.238 A combined approach involving lectin enrichment of intact proteins followed by ZIC-HILIC enrichment of tryptic glycopeptides provided more comprehensive coverage of the serum glycoproteome. Other strategies such as TiO2 chromatography and even chemical methods have also been used sequentially or in parallel with HILIC for glycopeptide enrichment.251,252 The combination of different approaches can normally increase the number of captured glycopeptides dramatically. However, glycopeptide loss could also occur when enrichment methods with similar specificities are used sequentially such as the HILIC-ERLIC enrichment strategy tested by Zacharias et al.127
4.1.4. Enrichment Based on Covalent Interactions.
Enrichment methods based on covalent interactions between glycopeptides and binding ligands are thought to be more universal and unbiased because chemical reactions are less affected by variations in glycan structures. The two most common types of glycopeptide enrichment involve boronic acid and hydrazide chemistry. Both methods produce covalent bonds between the cis-diol groups present in monosaccharides and the boronic acid or hydrazide groups, respectively. The major difference between these two methods lies in the reversibility of the reaction.209 The five- or six-membered cyclic esters formed between boronic acids and glycopeptides can easily be dissociated under acidic conditions to release the captured glycopeptides intactly. The hydrazone formation between hydrazide groups and the aldehyde groups formed by oxidization of glycan cis-diols, however, is irreversible. Glycans must be cleaved by PNGase F or chemically to release the enriched peptides. In this method, only deglycosylated peptides are observed, while the glycan and perhaps even the site-specific information are lost. The same problems occur with other chemical enrichment methods such as reductive amination253 and oxime click chemistry.74 For this reason, only boronic acid chemistry is further discussed as a universal enrichment method for site-specific glyoproteomic analysis.
Various types of solid supports have been functionalized with boronic acids for the enrichment of glycoproteins or glycopeptides. Some functionalized materials including mesoporous silica,254,255 magnetic nanoparticles,147,256,257 and carbon nanotubes258 have been applied to the glyoproteomic analysis of complicated biological samples.259,260 For example, magnetic beads conjugated with boronic acid were combined with stable isotope labeling of amino acids in cell culture (SILAC) to capture and quantitatively characterize glycoproteins in the secretome and whole cell lysates of yeast.261,262 Although boronic acid chemistry is suitable for the enrichment of a wide range of glycoforms, its nonspecific binding of free saccharides and other compounds in the matrices can compete with glycopeptide capture and interfere with subsequent LC-MS/MS analysis. Novel materials such as dendrimeric boronic acid-functionalized magnetic nanoparticles may enhance the binding capacity and address this problem to some extent.263 However, further improvement in specificity is still necessary.
Another emerging approach involves the use of bioorthogonal functional groups, such as azides, alkynes, or ketones for targeted labeling and enrichment of glycoproteins from cultured cells.264–266 More recently, Spiciarich et al. applied this approach to human prostate cancer tissue.267 Peracetylated N-azidoacetylmannosamine (Ac4ManNAz) was used in sliced normal and cancerous prostate tissue cultures to add the azide groups to the sialic acid residues on cell surface and secreted glycoproteins. The labeled sialoglycoproteins were then reacted with a biotin-alkyne probe, captured with avidin resin, digested with trypsin, and analyzed by LC-MS/MS. (Figure 17) These bioorthorgonal approaches have high specificity for enriching sialylated glycoproteins and other types of glycoproteins when proper metabolic labels are utilized. For global labeling of N-glycoproteome, for example, comparison of three sugar analogues, N-azidoacetylgalactosamine (GalNAz), N-azidoacetylglucosamine (GlcNAz), and N-azidoacetylmannosamine (ManNAz), revealed that GalNAz could yield the greatest number of glycoproteins and glycosylation sites for HepG2 liver cells.268
Figure 17.
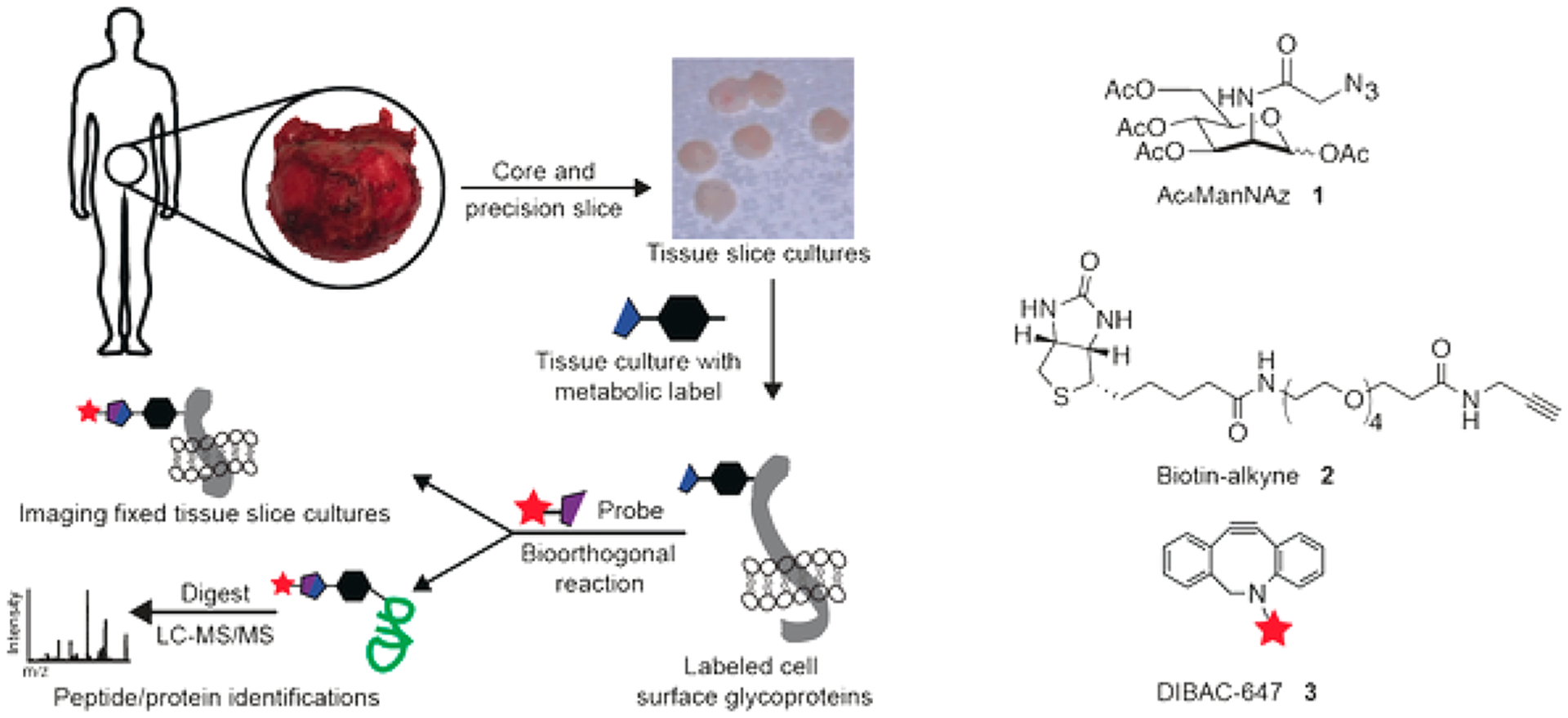
Bioorthogonal labeling of human prostate tissue slice cultures with azido sialic acids for imaging, enrichment, and LC-MS/MS analysis of sialoglycoproteins. Reprinted with permission from Spiciarich, ref 267. Copyright 2017 Wiley Online Library.
4.2. Separation Methods for Glycopeptides
4.2.1. Liquid Chromatography of Glycopeptides.
Reversed-phase liquid chromatography (RPLC) employing a C18 column is the most widely used method for untargeted site-specific glycoproteome analysis. Separation of glycopeptides with RPLC is mainly based on the binding interaction with the peptide backbone and can be predicted based on the amino acid sequence.269 The glycan has only minor influence in the interaction between glycopeptide and RPLC. The effect of glycan composition and structure on the retention behavior of glycopeptides was recently studied by Kozlik et al.270 Reduction of the retention time was observed with increasing number of neutral monosaccharide units of glycans attached to the same peptide backbone (Figure 18). Relative retention times of different glycoforms could thereby be calculated and used as a qualitative parameter to reduce false positive rate of glycopeptide identification. However, care should be taken when using this prediction approach because the actual retention times of glycopeptides could be complicated by variations in chromatographic conditions and existence of sialic acids residues.271
Figure 18.
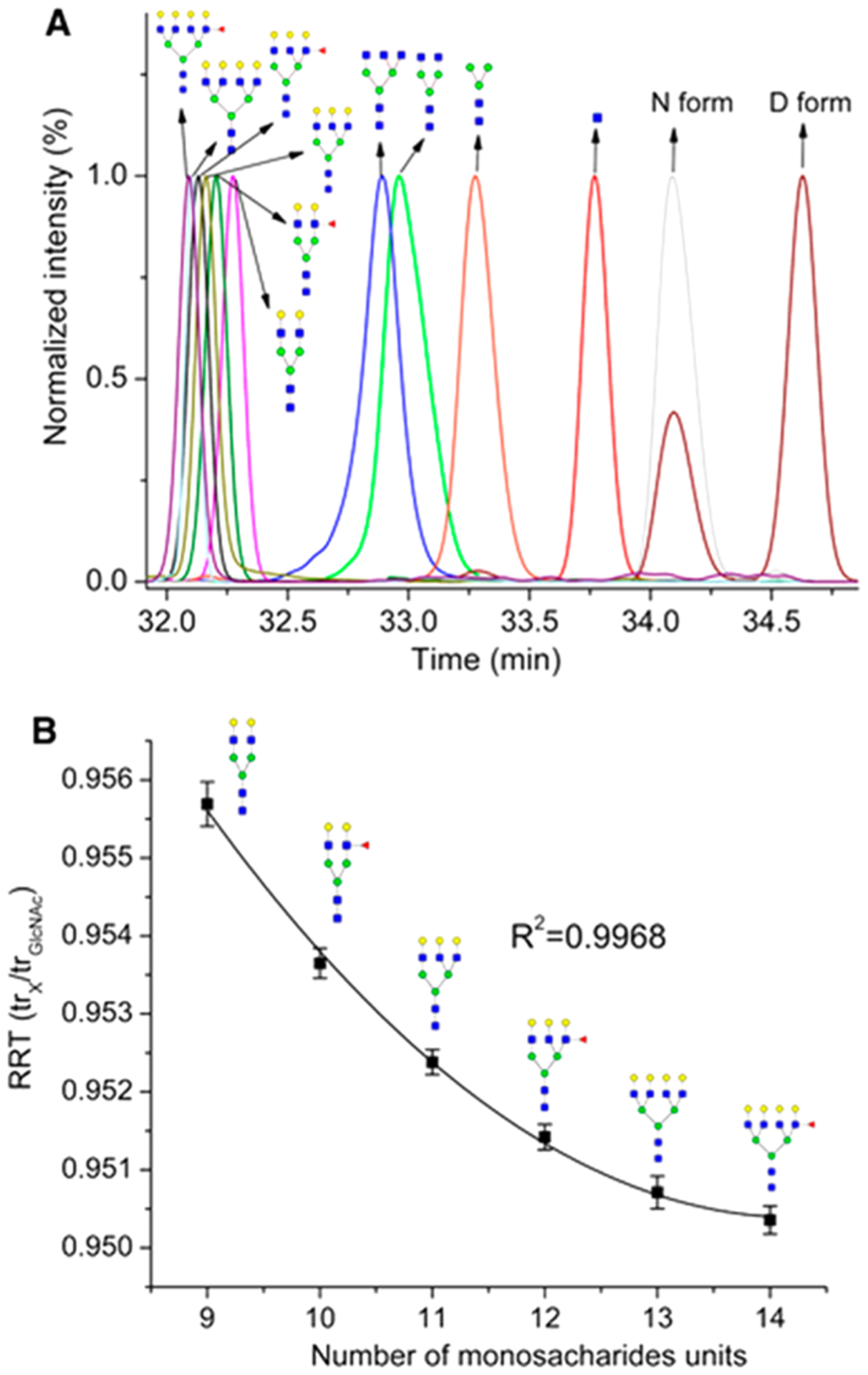
Analysis of the VVLHPNYSQVDIGLIK peptide of haptoglobin by nanoLC. Decreased retention of glycopeptides was observed with increasing number of neutral monosaccharide units. Reprinted with permission from ref 270. Copyright 2017 Wiley Online Library.
The major drawbacks of RPLC for glycopeptide separation include the poor retention of highly hydrophilic glycopeptides and the limited separation of different glycoforms. To address these problems, orthogonal separation methods combining RPLC and PGC or HILIC have been developed and applied to site-specific analysis of N- and O-glycopeptides derived from trypsin272,273 and pronase274 digestion. Complementary separation by PGC or HILIC can efficiently recapture the hydrophilic glycopeptides that might be lost in a conventional RPLC-only approach (Figure 19). PGC or HILIC alone was also used to enrich and separate intact glycopeptides derived from specific and nonspecific protease digestion of glycoproteins.275 Isomeric separation was achieved for both N-glycopeptides and O-glycopeptides, especially for those obtained from pronase digestion with short peptide backbones.54,244,276–279 These methods are ideal for glycosylation site and structural heterogeneity mapping of purified glycoproteins. Although HILIC separation has also been used to separate IgG N-glycopeptides from human serum,280 neither PGC nor HILIC alone is sufficient for comprehensive glycoproteome mapping because important information from the unretained hydrophobic peptides and glycopeptides are lost.
Figure 19.
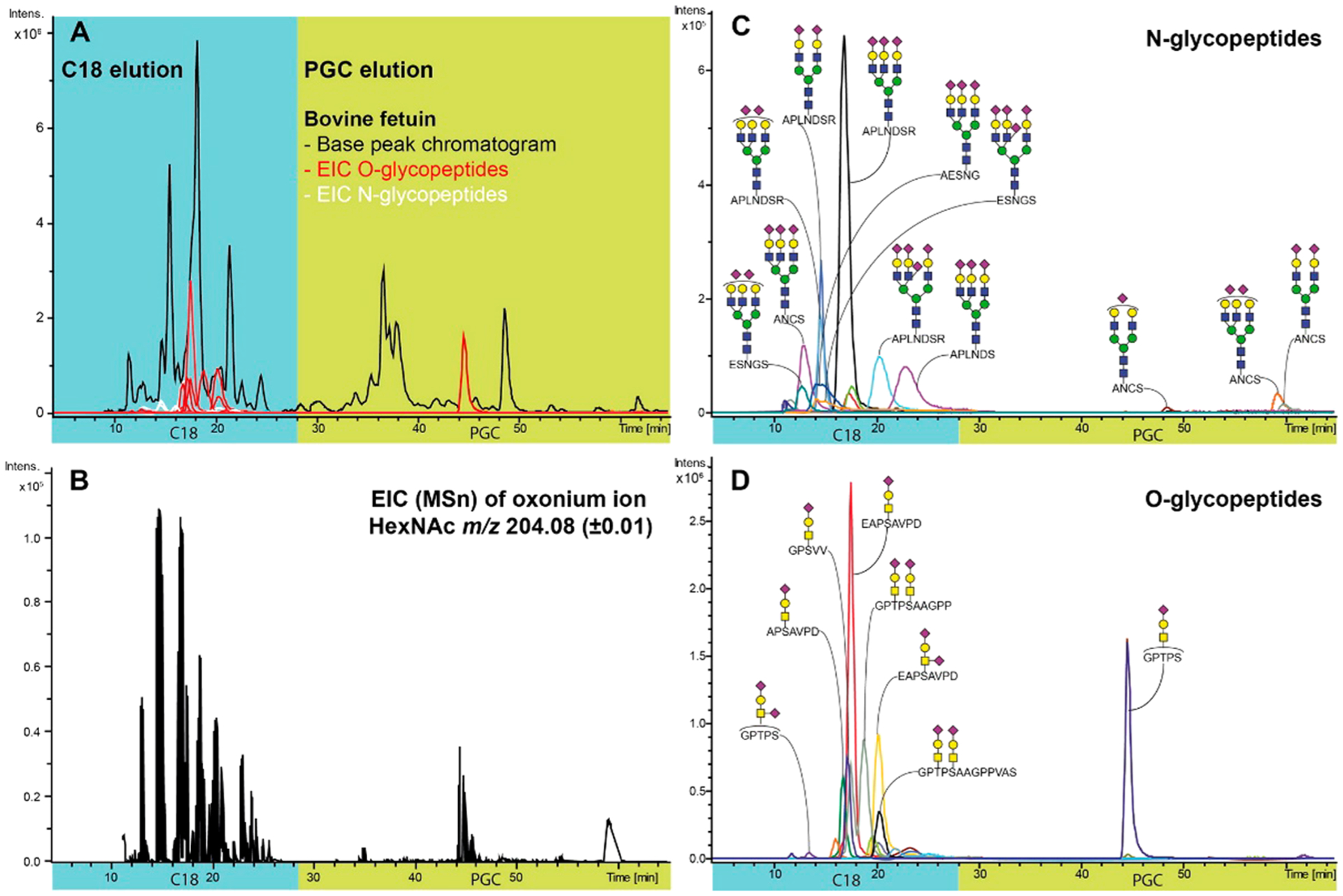
C18-PGC-LC-ESI-QTOF-MS/MS analysis of N- and O-glycopeptides from pronase digest of bovine fetuin. Reprinted with permission from ref 274. Copyright 2015 American Chemical Society.
4.2.2. Capillary Electrophoresis of Glycopeptides.
Early researches on capillary electrophoresis of glycopeptides have applied sheath-flow CE-MS to hydrophilic glycopeptides such as tryptic N-glycopeptides from ribonuclease B281 and antithrombin282 glycoprotein standards. Sanz-Nebot and co-workers have applied this approach to a series of studies on the characterization of N- and O-glycopeptides from recombinant human erythropoietin283 and other glycoproteins.284,285 They have also systematically studied the electrophoretic migration behavior of peptides and glycopeptides that can be used to simulate their migration and separation on CE.286 However, the limitations of CE-MS discussed above for glycomics are further amplified for glycoproteomics because higher sensitivity and more robust separation are required to characterize the structural heterogeneity of glycopeptides. For example, comparison of CE-MS and μLC-MS showed that CE had poorer separation for glycopeptides when a surfactant was used to enhance enzymatic digestion.284 To improve the sensitivity of CE-ESI-MS in glycopeptide analysis, Kammeijer et al. employed dopant enriched nitrogen gas with a coaxial gas flow to build a sheathless nanoESI interface.287 The addition of dopant gas greatly facilitated the desolvation and ionization, thus achieving higher sensitivity for glycopeptide detection. More recently, Qu et al. reported a rapid CZE-ESI-MS method with an electrokinetically pumped sheath-flow nanospray interface for the analysis of glycopeptides from IgG and haptoglobin standards. The development of these novel interfaces has increased the applicability of CE to glycoproteomics as a complementary separation approach to RPLC.
4.2.3. Ion Mobility of Glycopeptides.
Ion mobility separation of glycopeptide isomers is even more challenging than that of glycans due to the high structural similarities, numerous possibilities for isomerization, and lack of commercial standards. Isomeric separation of intact glycopeptides achieved so far include the separation of peptides modified with different monosaccharide residues (GlcNAc or GalNAc) at the same site,288 GalNAc at different locations,289 or a sialylated N-glycan at different sites,290 and the separation of isomeric glycans attached to a single amino acid.291 For N-glycopeptide isomers that differ only in the linkage of one monosaccharide residue, partial separation was achieved for their highly charged species.292 Baseline separation is nearly impossible. A novel strategy to address this issue involves IM separation of the diagnostic isomeric product ions after fragmenting the glycopeptides using low-energy CID. This strategy has been used to distinguish sialic acid linkage isomers of synthesized N-glycopeptide standards (Figure 20)290 and N- and O-glycopeptides derived from glycoprotein standards.293 However, the application of this approach to complicated mixtures remains challenging because other coeluting sialylated glycopeptides will generate the same fragments.
Figure 20.
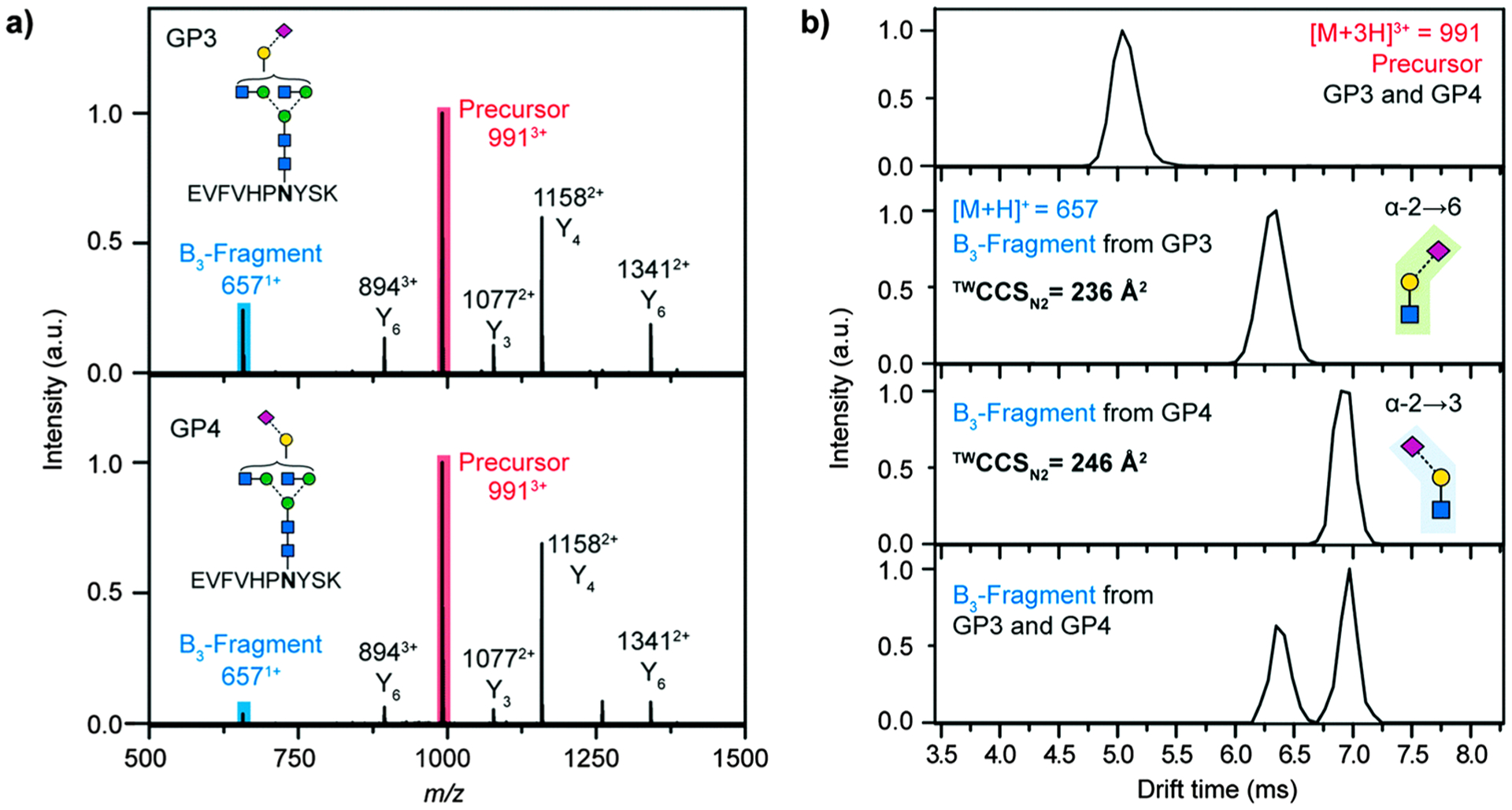
Distinguishing sialic acid linkage isomers on glycopeptides using CID fragmentation and subsequent IM-MS analysis. Reprinted with permission from ref 290. Under Creative Commons Attribution 3.0 license (https://creativecommons.org/licenses/by/3.0/).
4.3. Determination of Glycosylation Sites
The most common approach for identifying site-specific glycosylation involves a series of steps including enzymatic proteolysis of glycoproteins as well as chromatographic separation and mass spectrometry analysis of glycopeptides.294 For enzymatic digestion, trypsin, which exclusively cleaves a protein at the C-terminus of arginine and lysine except when they are bound to a proline, is widely used. Because of its high specificity, trypsin digestion generates predictable peptide backbones. Furthermore, the ionization efficiency of tryptic digested glycopeptides are improved during MS analysis because a basic residue is contained in each peptide backbone. A huge drawback of trypsin digestion is the possibility of generating missed cleavages due to steric hindrance at the cleavage site near the glycosylation site.295 The resulting glycopeptides can be too large for the current proteomic workflow. Additionally, some glycoproteins may not be compatible with trypsin digestion conditions.296 These issues can make identification of glycosylation sites more challenging.
To minimize the limitations of trypsin digestion, nonspecific digestion using proteases like pronase and proteinase K can be applied.20,297 These nonspecific proteases cleave glycoproteins into glycopeptides with shorter peptide backbones or nonglycosylated dipeptides and amino acids. The resulting glycopeptides can be further enriched by solid phase extraction using PGC prior to MS analysis. Compared to specific protease digestion using trypsin, nonspecific digestion could provide much better glycosite coverage but fewer glycoforms on some sites (Figure 21).20 This method has been extensively used to identify the site heterogeneity (N- and O-glycosylation site) of many glycoproteins.298 However, it is limited mainly to the analysis of either one purified glycoprotein or small pools of glycoproteins because the resulting glycopeptides contain fewer amino acids. Multienzyme strategies employing combinations of specific proteases could also be used to improve glycosite identification. The combinations of trypsin and Glu-C as well as trypsin and chymotrypsin have been used to identify O-glycosylation sites in human serum proteins.299 All of the methods mentioned above can be further improved by enriching glycoproteins prior to the digestion or by enriching glycopeptides after the digestion.
Figure 21.
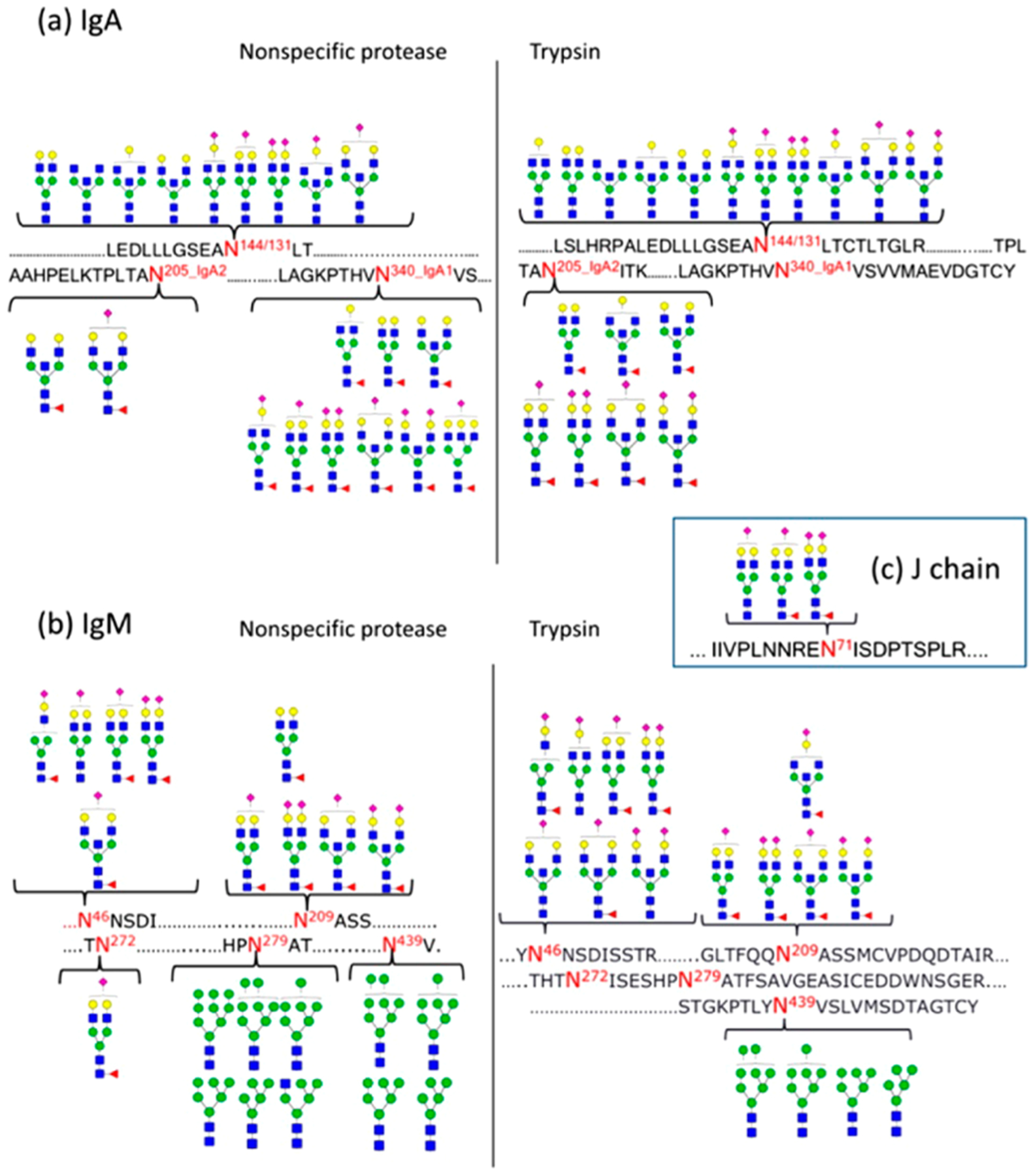
Comparison of nonspecific protease and trypsin for site-specific glycan mapping. Reprinted with permission from ref 20. Copyright 2015 American Chemical Society.
A simple approach for identifying glycosylation sites is to remove the glycans completely or partially using enzymatic means. The entire N-glycans can be cleaved off from the attached asparagine by peptide-N-glycosidase F (PNGase F), which converts asparagine into aspartic acid. Detection of aspartic acid would be considered as a direct indication of an occupied N-glycosylation site. However, deamidation of asparagine, a common post-translational modification, can result in the false positive identifications. This problem can be addressed by performing the deglycosylation in isotopically labeled water.300 Another approach is to perform partial deglycosylation using endoglycosidases301–303 or chemical methods,304 thus leaving a single monosaccharide (GlcNAc) attached to the peptide while removing the rest of the glycan moiety. The resulting glycopeptide with a single GlcNAc is a valid indication of an occupied site.
4.4. Applications of Glycoproteomics
The development of LC-MS/MS techniques in glycoproteomics makes it more widely used for studying the pathogenesis of several different diseases including cancer305–307 and neurological diseases.308 In addition, glycoproteomics research can now be conducted on various types of biological samples such as blood,309–311 urine,312 cell lines,313–315 and other tissues.127,316 Many of these studies focus on the biomarker discovery for a specific disease by quantifying glycopeptides and determining glycosite occupancies.317–319 Other studies expand the glycoproteome with regard to unique organisms.320 While the application of glycoproteomics has expanded considerably, this section highlights more recent examples related to advances in dealing with various sample types with specific focus on diseases.
Goyallon et al. studied the site-specific glycosylation of proteins in cerebrospinal fluid using an approach that combines N-glycoproteomic and N-glycomic analyses.321 The N-glycans from human serum and cerebrospinal fluid were released, permethylated, and analyzed with MALDI-TOF MS. Tryptic glycopeptides from the same samples were analyzed after enrichment as intact glycopeptides or deglycosylated peptides with a nanoLC-MS/MS system (Figure 22). In total, over 100 glycopeptides from 36 glycoproteins were identified in a pooled cerebrospinal fluid sample. Unique “brain-type” glycoforms were identified in this sample compared to those found in serum.
Figure 22.
Overall strategy used for the site-specific glycopeptide analysis of cerebrospinal fluid samples. Reprinted with permission from ref 321. Copyright 2015 Wiley Online Library.
Glycoproteomic analysis of mouse and human embryonic stem cells (mESC and hESC) were performed by Stadlmann et al. to identify new proteins involved in the ricin toxicity.322 Glycopeptides were enriched from digested cell lysis using HILIC cartridges and analyzed by nanoLC-MS/MS (Figure 23). By applying the method to mESC, the total number of experimentally identified N-glycosylation sites was almost twice the number previously determined in the mouse glycoproteome. The method was also applied to hESC, and more novel glycoproteins were identified. The specificity of the method was validated by studying the glycoproteomes of Fut9 and Slc35c1 knockout cell lines, which are ricin-resistant. Several fucosylated glycoproteins that are essential in ricin toxicity resistance were revealed.
Figure 23.
Workflow of glycoproteomics by combining proteomcis platform and novel algorism for intact glycopeptide identification. Reprinted with permission from ref 322. Copyright 2017 Springer Nature.
Mouse brain glycoproteome was also studied by Fang et al.320 using multienzyme approaches by employing both the enzymes with high specificity such as trypsin and Glu-C and those with low specificity such as chymotrypsin and pepsin. Compared to the single-enzyme approach using only trypsin, the number of identified N-glycosylated sites was greatly improved with the multienzyme approach. The efficiencies of four glycopeptide enrichment methods, including hydrazide chemistry, TiO2 chromatography, HILIC (Sepharose CL-4B), and ZIC-HILIC, were also evaluated in the study. While complementary N-glycosylation sites were obtained from different methods, HILIC and ZIC-HILIC yielded the largest numbers of identified glycopeptides. Furthermore, two different fractionation techniques, SCX prefractionation before ZIC-HILIC and bRP prefractionation after ZIC-HILIC, were also compared. Altogether, over 10000 N-glycopeptides from nearly 4000 proteins were identified in this study, revealing the large diversity of N-glycoproteome in mouse brain. Sample preparation methods optimized in these types of studies therein will be helpful for the further study of neurological diseases.
The N-glycoproteome of dried blood spot (DBS) samples has been studied by Choi et al. with a relatively simple sample preparation method.323 The DBS samples were prepared and stored for different periods of time before analysis to evaluate the stability of N-glycoproteins. N-Glycopeptides were obtained after denaturation and tryptic digestion without depletion, desalting, or enrichment. With their optimal sample preparation method, more than 30 N-glycopeptides of 14 glycoproteins were quantified consistently over different storing periods. N-Glycan profiles of DBS samples have also been reported by Ruhaak et al., showing the applicability of DBS for long-term and large cohort clinical studies.324
Glycoproteomic analyses have been performed extensively on cancer. One widely studied target is ovarian cancer, a disease that desperately needs good diagnostic markers. Ji et al. performed both proteomic and N-glycoproteomc analyses of ovarian cancer cell line OVCAR8 and its doxorubicin-resistant variant, NIC-ADR/RES, to unravel the underlying mechanism for multidrug resistance (MDR).325 Quantitation of peptides and glycopeptides were conducted using stable isotope labeling of amino acids in cell culture (SILAC). After the enrichment of N-glycopeptides with HILIC, PNGase F was used to remove the N-glycans. The resulting deglycosylated peptides containing glycosites were analyzed with LC-MS/MS. Quantitative proteomic information was obtained simultaneously. Several peptides with N-glycosylation sites were found to be differentially expressed with and without changes in site occupancy, which may be relatible to MDR properties of ovarian cancer cells.
To understand the biology of epithelial ovarian carcinomas, Li et al. studied the overall variation of glycoproteome and site occupancy of glycoproteins in high-grade ovarian serous carcinoma (HGSC) and benign epithelial ovarian tumor.326 Both tumor tissues and benign tissues were collected from female patients and homogenized for protein extraction. Glycopeptides were enriched using hydrazide chemistry and labeled with an isobaric tag for relative and absolute quantitation (iTRAQ) reagent. Samples were prefractionated on basic RPLC prior to analysis with LC-MS/MS to provide better coverage. Approximately 40 deglycosylated peptides showed significant changes between ovarian tumor and benign tumor, while the site occupancies of 10 glycopeptides varied, revealing that specific changes in glycosylation site occupancy may be potential biomarkers for ovarian cancer.
To obtain potential biomarkers of hepatocellular carcinoma (HCC), Liu et al. analyzed serum samples collected from patients with early HCC related to chronic hepatitis B virus (HBV) infection and controls.327 Glycoproteins with α(2,3)-linked sialic acid were enriched with Maackia amurensis lectin (MAL) and quantified using the iTRAQ labeling method on nanoLC-MS/MS. The quantitation results showed significant fold changes of eight proteins. The differential expression of these proteins were further validated by Western blotting, and galactin 3 binding protein (Gal-3BP) was selected for ELISA analysis as a candidate biomarker to identify HBV-related HCC.
5. SOFTWARE TOOLS FOR GLYCOMICS AND GLYCOPROTEOMICS
Mass spectrometry of glycans and glycoconjugates in biological samples generates large amounts of data. To fully interpret the data, bioinformatics tools are necessary. Unlike proteins and DNA, glycans and glycoproteins are highly heterogeneous due to their nontemplate biosynthesis. This makes it difficult to automatically annotate glycan and glycopeptide structures. The ability to assign peptide sequence, glycan composition, and glycosylation site simultaneously is challenging and considerably more difficult than elucidating point modifications on proteins such as phosphorylation. For the analyis, tandem MS data are usually deconvoluted and deisotoped. Effective searches need to be performed against both protein and glycan databases. The theoretical protein and glycan search space should also be defined properly to reduce false discovery rates (FDRs)328 in the database searches. A recent review by Hu et al. has discussed the issues associated with interpretation of glycopeptide tandem MS data.329
Software for glycomic analysis should facilitate the annotation of structures of N-glycan, O-glycans, glycosaminoglycans, and other oligosaccharides from mass spectrometry data. Xu et al. recently developed GlycoMaid with a graphic user interface (GUI) for automatic interpretation of N-glycan data from MALDI-TOF MS based on the masses and the isotopic distribution of the signals.83 The software was improved from a previously developed version using a matching glycan isotope abundance (mGIA) algorithm, enhancing the ability for deconvolution of overlapping isotopic distributions of glycans.
Other approaches produce glycan structures based on tandem mass spectrometry data. A machine learning tool developed by Kumozaki et al. conducts de novo sequencing of glycans based on fragmented glycan peaks in MS/MS spectra without database searches.330 The fragmentation model of glycans is first established, and a solver using Lagrangian relaxation together with a dynamic programming technique was utilized. The algorithm scores were then optimized by making use of structured support vector machines (SVMs), which is a machine learning technique by studying the parameters for glycan structures using training data sets such as MS/MS spectra of known glycans. To annotate glycan structures more accurately, known glycan core structures including O-linked and N-linked glycans were more constrained. The algorithm enables the de novo sequencing of complicated glycan structures.
For glycoproteomic analysis, commercial and in-lab software have been developed. Byonic is arguably one of the most commonly used comercial software packages for interpreting the tandem MS data of peptides and glycopeptides.331 With the target protein database and N-linked or O-linked glycan library, the software can perform automatic glycan search based on MS/MS spectra of glycopeptides without knowing glycosylation sites or glycan compositions. Besides glycosylation, several other modifications such as carbamidomethylation of cysteine (C [+57.021]) and oxidation of methionine (M [+15.995]) can be set as fixed or variable modifications on peptides. In addition, enzymes with variable specificities can be specified for in silico digestion when searching spectra. The results generated from Byonic include a summary of assigned peptides and a list of proteins. In the list of peptides, the possible variable and fixed modifications with their masses on each peptide and glycosylation modifications including glycan compositions and masses are illustrated. The charges of observed glycopeptide precursor ions are also listed. Several scores are used to evaluate the quality of the site-specific assignments of glycopeptides, including Delta Mod and Log Prob. The Delta Mod evaluates the quality of the assigned modification on the peptide and Log Prob shows the possibility that the assigned peptide sequence is random. Both of these two parameters are used in further data analysis to remove those poorly matched glycopeptides. The original spectra together with matched peptide fragments can be accessed from the software to check the coverage of protein sequence.
A software that was developed strictly for extensive site-mapping of glycans is GlycoPeptide Finder (GP Finder).278 Instead of searching data based on specific or hemispecific enzymatic digestion, GP Finder can be used to annotate nonspecifically digested glycopeptides using enzymes such as pronase E by calculating all possibilities of peptide sequences after digestion. With nonspecific digestion, glycans connected to variable lengths of peptides are observed. Although the lengths of peptides generated in this way vary, digestion time can be regulated to control the variability of peptide lengths. In data analysis, glycopeptide candidates are filtered based on accurate mass-to-charge values, fragmentations, and diagnostic oxonium ions. A decoy data library generated by adding 11 Da residue to theoretical glycan compositions was used to evaluate the false discovery rate. This software has been applied to glycosylation mapping of bovine pancreatic ribonuclease (RNaseB) with a single glycosylation site, a mixture of three known glycoproteins with multiple sites, and several unknown glycoproteins in very-low-density lipoprotein (vLDL) nanoparticles.278
A more recently developed software for glycoproteomic analysis is pGlyco, which takes advantage of the features of orbitrap MS with a new algorithm to reduce the FDR of glycopeptide identification.332,333 The software was applied to standard glycoprotein mixtures and mouse tissue samples using an Orbitrap Fusion instrument. In the analysis, HCD-MS/MS data were collected. Glycopeptide precursor ions were selected for further MS/MS or MS3 analysis when the diagnostic ion, 138.055, was detected in HCD tandem MS spectra (Figure 24). The CID tandem MS data contain mainly glycan fragments, and the structure of glycan was inferred using both HCD- and CID-MS/MS data. The MS3 spectrum of Y1 ion, the peptide backbone with one HexNAc residue, contained peptide backbone fragments to yield the peptide sequence. The identified glycan structures were aligned with the peptide sequences based on retention times, generating a list of glycopeptides. The FDR was estimated by a novel spectrum-based decoy method where a decoy spectrum of the target glycopeptide was generated by modifying each peak with a random mass of 1–30 Da and matched to an experimental spectrum. With pGlyco, over 300 unique glycopeptides were identified in a mixture of six standard glycoproteins. Subsequently, approximately 10k nonredundant glycopeptides from 955 glycoproteins in mouse tissues were identified.
Figure 24.
Overall flowchart of data collection for pGlyco 2.0 analysis. Reprinted with permission from ref 332. Under Creative Commons CC BY license (https://creativecommons.org/licenses/by/4.0/).
Earlier attempts for automated analysis include GlycoMaster DB,334 SweetNET,335 gFinder,336 and Integrated GlycoProteomeAnalyzer337 and the oldest GlycoX.15 These software were developed for specific applications, while all lacked a method to properly determine the false positive rates.
In addition to these tools, databases of glycans and glycoproteins/glycopeptides have been developed as references for glycomic and glycoproteomic studies. Some provide tentative structure of glycans, such as GlyTouCan,338 Carbohydrate Structure Database (CSDB),339 and UniCorn.340 GlyTouCan provides structures together with compositions and topologies of glycans, and accession numbers are assigned to glycans, making it more searching friendly. CSDB contains NMR spectra and other properties of both natural and derivatized glycans in plants; UniCorn provides theoretical structures of N-glycans that are derived by cleaving glycans with enzymes in different pathways of biosynthesis. There are also database related to specific research areas. For example, SugarBindDB is a database that covers the interactions between glycans and proteins such as human pathogens and adhesins.341 For researchers learning about glycans using ion mobility mass spectrometry, GlycoMob would be useful in the way of providing CCS values, masses, and fragments of glycans.161 Glycoprotein databases such as Unipep and dbOGAP include information on N-glycosylated proteins together with peptides and O-GlcNAcylated proteins, respectively.342
6. TOP-DOWN GLYCOPROTEOMICS
In comparison to the previously described strategies for the analysis of released glycans and glycopeptides using bottom-up proteomics approaches, top-down proteomics, or the analysis of intact proteins, has not yet found wide utility for glycoproteins. The reason is in part due to the relatively recent introduction of high resolution mass spectrometry (e.g., FTICR-MS, Orbitrap, and HR-TOF343,344), which is a requirement for top-down.345 Advancements in top-down proteomics have recently been reviewed extensively.346
One of the most prominent advantages of top-down glycoproteomics is that the information is obtained on the whole protein so that full coverage of the protein can be achieved and almost all of the glycosylation sites can be identified. Meanwhile, these can hardly be achieved in bottom-up glycoproteomics due to difficulties with digestion, unfavorable conditions for HPLC separation, and MS ionization. The availability of the information on the whole protein has additional consequences in that specific protein variants or proteoforms,347 which originate from the same gene but have undergone different processing and carry different PTMs (not only glycosylation but also phosphorylation, acetylation, and other PTMs), can be individually identified. With the recently increasing interest in personalized medicine,348 which requires knowledge of an individual’s phenotype, the characterization of individual proteoforms could yield highly valuable information for diagnosis and treatment.
While this approach may be potentially powerful, the application of top-down proteomics for the characterization of protein glycosylation is limited because its use is constrained by several drawbacks. One of the most important limitation is the high complexity of the mass spectra obtained, which include not only the extensive charge envelope but also the complexity related with the glycan heterogeneity. Furthermore, the sensitivity in top-down proteomics is limited compared to bottom-up for several reasons. Peptides are small compared to intact proteins and are more effectively separated with C18 stationary phases. While separation and enrichment may be achieved for proteins using C4 or C8 material,213,349 proteins are much less resolved, thus reducing the analytical sensitivity. Consequently, peptides from biological mixtures are often more easily analyzed, while the analysis of complex mixtures via top down is much more challenging. Thus, antibody-based enrichment of the target protein is typically required prior to top-down analysis.
Another limitation is the sheer size of the intact protein, which can affect tandem MS. The precursor signal can be divided over a vast number of fragments, resulting in the reduction of the sensitivity of the approach. Although this limitation can be ameliorated by increasing acquisition times, the typical requirement of more lengthy analysis time further limits multiplexing capabilities. The sensitivity can be further affected by the large number of vibrational modes. A large protein requiring significantly greater energy during tandem MS can accept more energy while simultaneously follows more numerous fragmentation pathways after activation. In contrast to peptides and glycopeptides which can often be sufficiently analyzed by CID, ETD or ECD are typically necessary for protein and glycoprotein fragmentation. The dissociation efficiencies of these techniques are generally much lower than CID due to the strong dependence on precursor charge density. The problem is compounded in glycoproteins, which are often in a low-charge state, resulting in lower fragment signal intensities and reduced analytical sensitivity.
Even with these major constraints, the method is finding early applications in the characterization of monoclonal antibodies (mABs).350 mABs are often available in larger quantities and are extensively purified as part of the manufacturing process. Further thorough characterization is required prior to FDA approval. The most common representation carry a single site of glycosylation in the Fc part of the protein,351 thus limiting the complexity due to glycan heterogeneity. A recent example is the study by Yang et al. with the analysis of three IgG4-Δhinge mutants at the intact protein and the released glycan levels.352 In general, the predominant glycoforms detected by the top-down approach were in good agreement with the analysis of released glycans, although specific analysis of the released glycans yielded considerably more glycoforms.
CID and to a lesser extend ETD and ECD still yielded insufficient fragmentation information to completely characterize glycoproteins in top-down workflows. However, novel fragmentation strategies that are likely to be more beneficial for the analysis of protein glycosylation in a top-down manner have emerged. Ultraviolet photodissociation (UVPD) was introduced for the so-called middle-down analysis of a therapeutic monoclonal antibody (Figure 25).73 In this analysis, restricted enzymatic digestion is employed to generate large peptides (~3–20 kDa) that are still amenable to high resolution LC-MS/MS analysis. It was shown that the rapid and high energy activation obtained through UVPD resulted in efficient cleavages of amino acid residues while maintaining the integrity of the more labile glycans, thereby allowing the confident localization of the glycan moieties. While this technique has been shown to provide very good protein coverage in top-down proteomics as well,353 it has not yet been employed for analysis of large intact glycoproteins.
Figure 25.
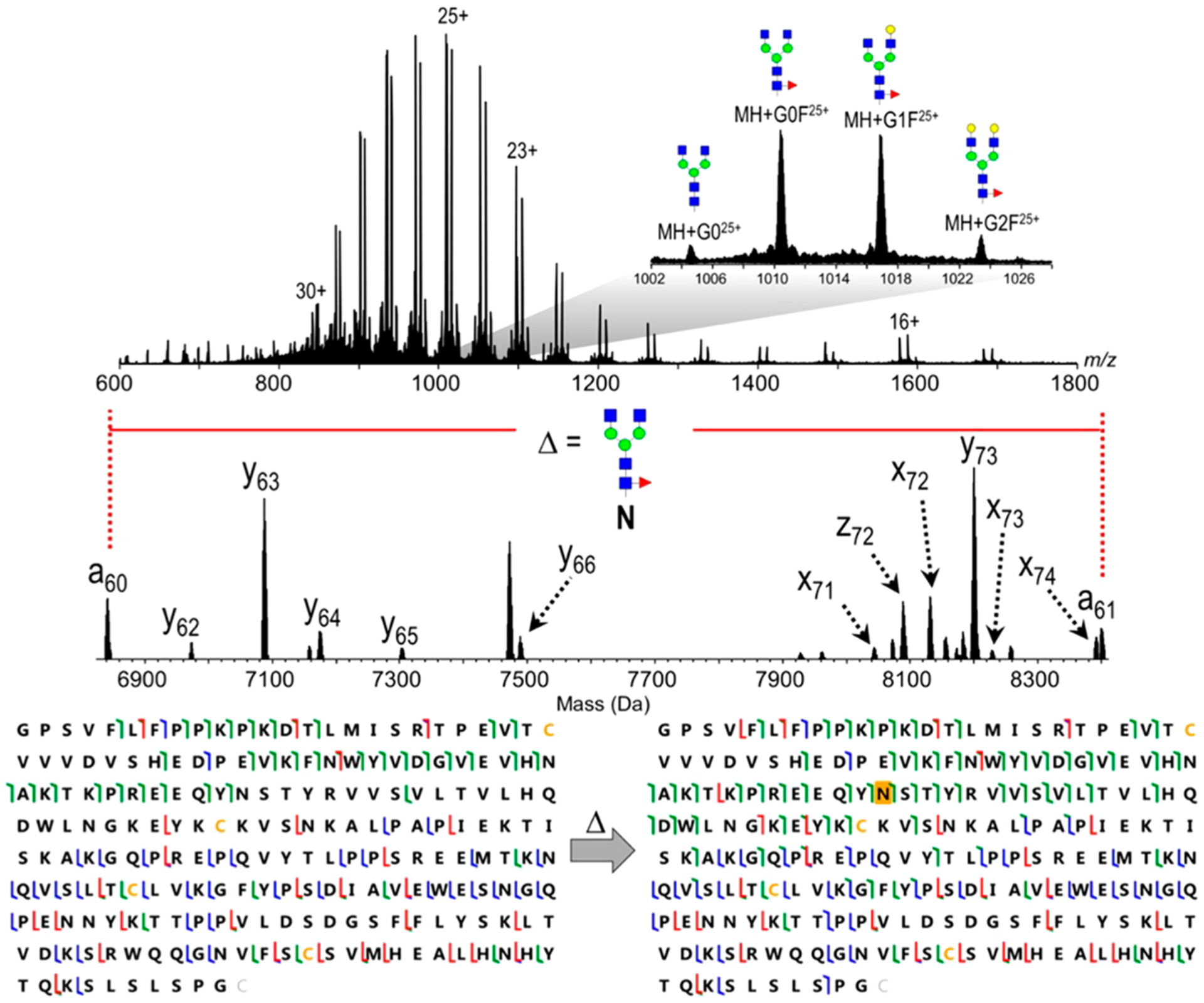
UVPD for middle-down analysis of a therapeutic monoclonal antibody. Reprinted with permission from ref 73. Copyright 2016 American Chemical Society.
The ETD and ECD fragmentation techniques suffer from a phenomenon called nondissociative electron capture or transfer, particularly for larger molecules such as intact (glyco)-proteins.354,355 Here, peptide or protein backbone cleavage occurs, but the product ions are held together by extensive intramolecular bonds, thus interfering with the generation of sequence information. Frese et al. introduced a combination of CID (or HCD) with ETD (EThcD), whereby a supplemental energy is applied to all ions formed by ETD to generate more informative spectra.356 In a comparative study using top-down proteomics, it was shown that EThcD provided increased protein sequence coverage and facilitates greater phosphorylation site identification compared to ETD.357 So far, EThcD has proven to be an attractive technology for glycopeptides identification,358 but the fragmentation technology has still not yet successfully been applied to intact glycoprotein analysis.
The application of additional energetic activation in ECD, recently called “activated ion ECD” (AI-ECD) has been shown to increase the ECD product ion yield and has successfully been applied in top-down proteomics. The technique was shown to be beneficial for the analysis of IgA O-glycopeptides359 and was likely to provide increased glycoprotein information compared to CID and ECD. Besides AI-ECD, there are attempts for the implementation of AI-ETD using infrared photoactivation in top-down proteomics.360,361 Riley et al. showed that the implementation of this technique on a quadrupole-orbitrap-linear ion trap hybrid mass spectrometer (Orbitrap Fusion Lumos) resulted in nearly complete sequence coverages of intact proteins.362,363 In the analysis of smaller proteins, AI-ETD was shown to outperform EThcD and provides at least similar protein coverage compared to UVPD. Because of the combination of the vibrational activation (through the IR laser) with the electron transfer driven activation, it is anticipated that AI-ETD will also provide more informative fragmentation in top-down glycoproteomics.
The previous examples were all performed using electrospray ionization. However, top-down glycoproteomics may also be employed using MALDI ionization. Nicolardi et al. conducted top-down glycoproteomics using MALDI-FTICR for the characterization of glycoforms of apolipoprotein C-III.364 In this study, ultrahigh resolution MALDI-FTICR MS was employed to characterize the glycosylation profiles of apolipoprotein C-III from 96 serum samples. The differences between the samples were assessed using MALDI-FTICR MS, while the identification of the different proteoforms was performed with ESI-FTICR MS with CID fragmentation.
Although the above-described methods covered mainly the application of direct infusion or MALDI ionization to the analysis of intact glycoproteins, hyphenation of the mass spectrometry technique with liquid chromatography or capillary electrophoresis has also been applied. Specifically, Van Scherpenzeel et al. have reported a platform in which C8 reverse-phase HPLC was combined with HR-TOF-MS for the characterization of glycosylation profiles from intact transferrin.213 The method was applied for the identification of patients with congenital glycosylation disorders based on the different top-down glycosylation profiles. Moreover, it provided potentially a sensitive technique for monitoring current and future therapies. A similar approach was applied for the in-depth characterization of two monoclonal antibodies.349 Using this technology, mean CVs of 2.2% and 3.7% were obtained for intra- and interassay variability, respectively, showing that a top-down strategy can be adopted for quality control of the production process of mAb biopharmaceuticals.
Capillary electrophoresis using coated capillaries helps avoiding protein binding and may be useful for separating intact glycoproteins prior to MS analysis. One of the early examples includes CE-TOF-MS analysis of erythropoietin (EPO), which carries three N-glycosylation sites and one O-glycosylation site.365 A similar method was used to assess the glycosylation of transferrin,366 and more recently a 2D-CE method was employed for the detailed MS characterization of mAb charge variants.367
Prostate specific antigen (PSA) is currently applied as a marker for prostate cancer, however, suffering from limited specificity.368 Recent reports indicate that PSA glycosylation may increase its specificity for prostate cancer,369 thus sparking increased interest for the characterization of PSA glycosylation. The association of biomolecular research facilities (ABRF) recently conducted a comparative glycomics study using a PSA standard from semen,370 and the top-down glycoproteomics was performed using HR-TOF-MS and orbitrap-MS, showing the potential application of the top-down glycoproteomics on the analysis of an individual glycoprotein with limited complexity. It was also concluded that the top-down glycoproteomics is promising for the analysis of smaller glycoproteins with a single glycosylation site. Although the detection of minor N-glycans yielded poor reproducibility, consistent identifications of all major and most intermediate abundance N-glycans were achieved. For more complex samples or larger glycoproteins, further evaluation on this technique is necessary.
7. QUANTITATION METHODS
The key to the eventual utility and understanding of glycosylation in biology and disease diagnosis lies in the ability to quantitate protein glycosylation. Quantification of protein glycosylation may be performed either at the relative or at the absolute level. Moreover, it may be determined at the glycan, the glycopeptide, or the glycoprotein levels, either in the forms of mixtures or a single glycoprotein. There has a been recent debate as to what strategy could provide the most useful results,371–373 with arguments being made specifically for the relative quantification of protein glycosylation,372 which for discovery purposes may be easier to develop. However, relative quantification does not allow for identification of changes in total glycosylation and is generally not accepted for translation to clinical diagnostics.373 Similarly, as we previously indicated for identification, the quantification at the glycan level may be performed with more glycan structural identifications than the quantification at the glycopeptide level. However, when just glycans are quantified, information on the protein and site of attachment is lost, which is important to determine biological significance.371 Therefore, the quantitative strategy (relative vs absolute quantitation and at which level) should be tailored toward the hypothesis in question and the expected results.
Similarly, protein and glycosylation quantitations may be performed at different levels of confidence depending on the required application. For example, if one wishes to assess whether a glycosylation pattern can differentiate healthy cells from cancer-derived cells in a discovery experiment, the precision requirements are different from a clinical test where the glycosylation on a specific glycopeptide can detect a certain disease. In relative quantitation, the ion abundances are directly compared between the two groups of samples, and only relative differences expressed in fold changes are reported. However, in absolute quantitation, absolute concentration is required and important, for example, for clinicians to make diagnosis. Typically, a targeted mass spectrometry approach is used in the latter, while nontargeted, full spectrum approaches are used in the first analysis. Clearly, there is a large gap in terms of performance between the relative quantification in the first method and absolute quantification in the clinical test. To bridge this gap, performance criteria have been developed for the translation of proteomics tests from discovery methods (tier 3) to clinical tests (tier 1).374 Similar criteria are applicable for the development and subsequent translation of methods for glycosylation analysis.
7.1. Quantitation of Glycan Profiles
An individual’s glycosylation profile is dependent on the actual glycan structures present on the proteins but also on the (relative) quantitation of the proteins carrying the glycosylation. Initially, the focus of quantitative glycosylation analysis has been on the analysis of the released glycans from individual glycoproteins,375 a subproteome,376 or the entire glycoproteome377,378 of an organism. In these studies, N- and/or O-glycans are released using PNGase F or chemical means and assessed for their compositional, structural and quantitative changes. However, a major disadvantage is the lack of information regarding the sites of glycosylation, which hampers linking differential glycosylation results to biological function.
Quantitative glycomics may be performed without labeling or relatively to labeled glycans. Song et al. employed a label-free approach using nanoLC-Chip-Q-TOF-MS in combination with an in-house serum glycan library to simultaneously identify and quantify over 170 N-glycans from human serum.9 The relative abundances of individual glycan structures were obtained (Figure 26). Similar label-free approaches have been successfully applied toward the identification of candidate markers for a large number of health and disease states.332,377,379–383 However, the information on the glycans quantified using these approaches is only as good as its qualitative properties. Recent focus has especially been on improving the quantitation of sialic acid containing glycans and linkage specificity of sialic acids (α(2,3) vs α(2,6)-linked residues). Traditionally, permethylation has been used as a method to neutralize sialic acids and improve their stability in MS,103 but variation in the number of methyl groups attached hampers this approach. More recent approaches enhance the analysis of sialic acid containing peptides by the selective modification of the sialic acid carboxyl group.384 Using this same approach, linkage specific derivatization strategies were developed for sialic acids.109 This derivatization approach might enable faster (relative) quantitative analysis of linkage specific differences, such as those identified on prostate specific antigen.369
Figure 26.
Relative abundances of human serum N-glycans averaged for nine individual sera. Reprinted with permission from ref 9. Copyright 2015 American Chemical Society.
To further enhance the relative quantitation of glycans, stable isotope enriched or more recently also isobaric labels have been introduced. These labels have recently been reviewed,385 and the reader is referred there for a complete overview. Isotopic labeling strategies based on permethylation,386,387 reductive amination using aminobenzamide384,388,389 or alanine,189,390–392 glycan reduction,393 and 18O incorporation by PNGaseF393–395 have been reported for N-glycans. Similarly, a stable isotope labeled PMP labeling method was developed for O-glycans.98,396 Reactions with tertiary amines,397,398 13CH3I/12CH2DI permethylation,6 and most recently quaternary amines399 were used to introduce isobaric tags. While these labeling strategies are valuable, they only allow for relative quantitation compared to other samples: ratios between the labels are being compared as representatives of the relative abundances of their reflecting glycans. As with all chemical modifications, the accuracy of the relative quantitation is highly dependent on the variability of the incorporation of the quantifying label.
An encouraging recent development is the synthesis of stable isotope labeled glycans (Figure 27).400 By using the chemoenzymatic synthesis, a library of biantennary 13C-labeled N-glycans with a mass increment of 8 Da was developed. The labeled glycans were quantified in absolute terms using NMR spectroscopy prior to their use as internal standards for the absolute quantitation of glycans derived from a monoclonal antibody by MALDI-TOF MS. The CVs obtained for the quantitation of eight glycans were below 14%. As pointed out by the authors, the complete liberation of N-glycans from the proteins by PNGaseF is a prerequisite for absolute quantitation using this method. It is anticipated that enlargement of the library will enable accurate quantitation of glycans in complex biological mixtures, which is required for the clinical application of glycans as biomarkers.
Figure 27.
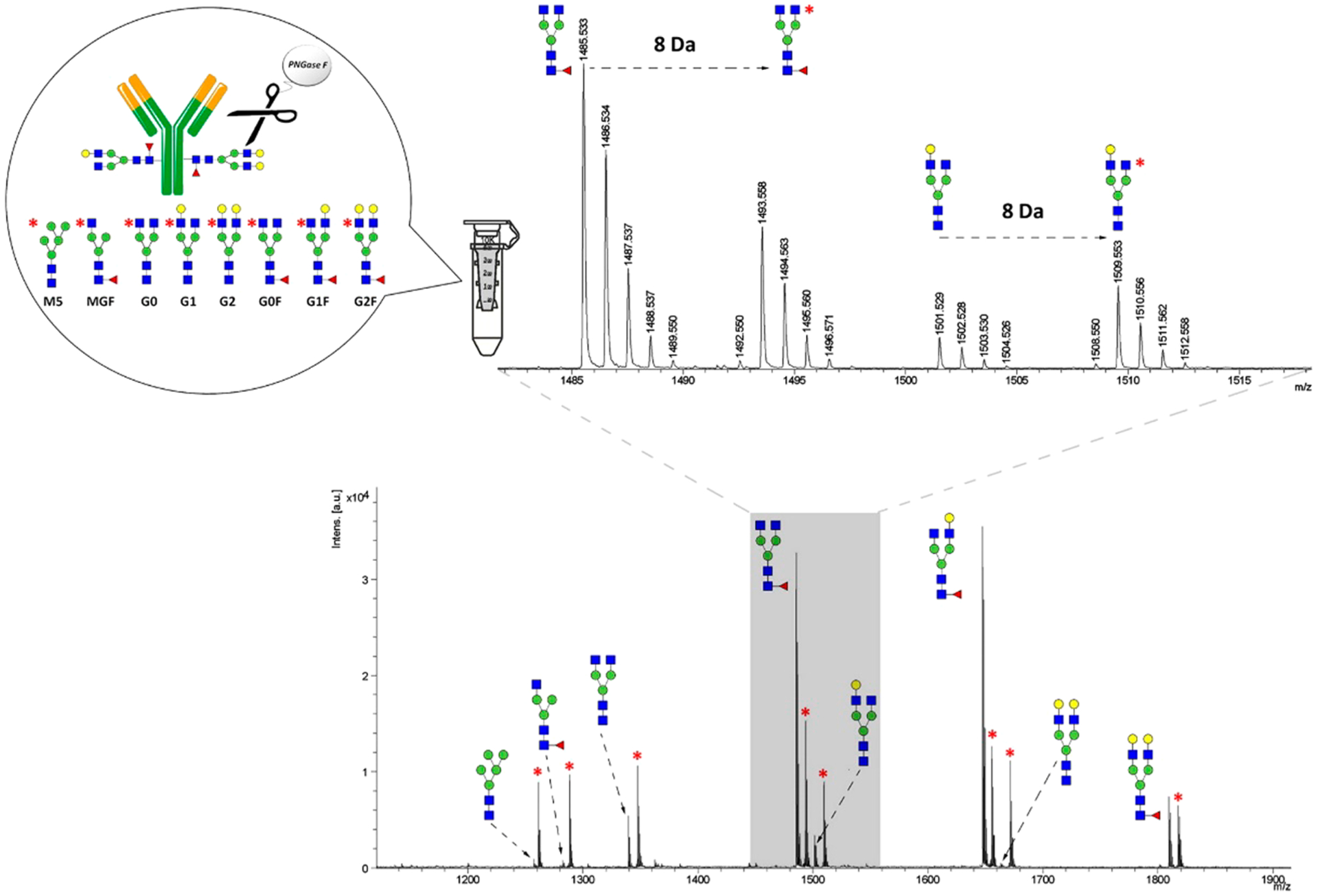
MALDI-TOF MS quantitation of IgG1 glycans using biantennary 13C-labeled N-glycans as internal standards (marked with an asterisk). Reprinted with permission from ref 400. Copyright 2015 American Chemical Society.
7.2. Quantitation of Glycosylation Site Occupancy
As stated previously, the decoration of proteins with N- or O-glycosylation is an enzymatic process that is dependent on the enzyme, glycosylation site, and substrate availability. Glycoprotein site occupancy may therefore influence its function. Quantification of glycosylation site occupancy may therefore reveal disease processes, and methods to do so were recently reviewed.401
The strategy that has categorically been used for the identification of differentially occupied glycoproteins is the analysis of deglycosylated peptides. In one approach, the glycans on the glycoproteins are irreversibly bound to beads, upon which the proteins are digested.402 Nonglycosylated peptides are washed away, and the deglycosylated peptides are then released from the beads using PNGaseF, thus removing the full glycan. Typically, this strategy has been employed in complicated samples such as tissue samples or other body fluids.210,403 Recently, automated methods for the identification and label-free relative quantitation of site occupancy have been developed using data independent strategies such as SWATH404 and SWAT, a targeted DIA approach.405 In a comparative study, SWAT outperformed SWATH for the quantitation of peptides in linearity and sensitivity, and more deglycosylated peptide/peptide pairs could be quantified to assess the occupancy of the glycosylation site. The authors indicated that this was mainly due to increased specificity.405
The incorporation of an isotopic label could improve the quantitation of glycosylation site occupancy. A strategy using 18O labeled Asp has been introduced for the potential diagnosis of CDG type-I patients406 and was shown to differentiate patients with different levels of severity of type-I CDG. More recently, a targeted PRM based method for the quantitation of site occupancy of 43 sites of glycosylation was developed according to performance criteria established by the Clinical Proteomic Tumor Analysis Consortium (CPTAC).210 Heavy stable isotope labeled peptides which incorporated 13C and 15N isotopes at the C-terminal K or R position of each tryptic peptide were used as internal standards, and site occupancy could be quantified with total assay CVs below 5%.
Importantly, the spontaneous deamidation of asparagine has been reported to occur during sample preparation.407,408 Therefore, the strategies where PNGaseF is used for de-N-glycosylation may result in incorrect quantities due to spontaneous deamidation of asparagine residues, which may overestimate the glycosylation occupancy.408 An alternative strategy has recently been reported where the unglycosylated portion of the glycoprotein is measured relative to the protein concentration.409 By including SIL peptides for both the unglycosylated peptide as well as the peptide used for protein quantitation, the effects of deamidation are compensated, and by using this strategy, the site occupancy of fetuin was reported to be significantly lower than with a PNGaseF based deamidation strategy. A similar strategy was used to assess the site occupancy in a fusion protein.410 Another strategy to perform glyan site occupancy is to use Endo H and/or Endo D exoglycosidases to deglycosyate glycoproteins. This approach performs partial deglycosylation while leaving single or small glycan moiety attached to the protein. Cao et al. used Endo H and PNGase F to quantify site occupancy in HIV-1 envelope glycoproteins.411 With this strategy, different masses were obtained for glycopeptides with high mannose, complex, and no glycan.
Also, absolute quantitation of site occupancy is determined by using MRM. In summary, glycoprotein is proteolytically digested prior to deglycosylation with PNGase F. The signal intensity of the standard containing deglycosylated and nonglycosylated peptides were used to calculate the site occupancy. Yang et al. used this method to obtained site occupancy of recombinant monoclonal antibody drugs.21
7.3. Quantitation of Glycopeptides
The identification of protein- and site-specific changes in glycosylation provides a much more targeted approach compared to the analysis of all glycans from (a subset of) the glycoproteome.371 The recent advances in the identification of glycopeptides have also opened up new opportunities for the site- and protein-specific quantitation. However, the quantitative analysis of protein- and site-specific glycosylation is only recently gaining attention. Strategies for glycopeptides quantitation can be label-free or relative to labeled standards and may be data-independent or targeted. The earlier reports of glycopeptide quantitation comprise label-free methods, where glycoproteins are isolated and glycopeptide extracted ion signal intensities are normalized either to one glycoform412,413 or to the sum of all glycoforms.414 While the strategy is most often applied to the analysis of N-glycopeptides, it has also been applied to O-glycopeptides.413,415 In a recent interlaboratory study on the identification and quantitation of glycopeptides from α−1-acid glycoprotein, the interlaboratory variability of label-free relative quantitation based on total glycopeptide abundance for the 10 major glycopeptides was shown to be <25%.416 Recent advances in the technology and software have significantly increased the number of glycopeptides (both N-glycopeptides332 and O-glycopeptides299) identified in complicated mixtures such as tissue or biofluids. It is anticipated that such techniques will soon also allow for label-free quantitation based on extracted ion chromatograms.
In an alternative strategy, different lots of human chorionic gonadotropin (HCG) were compared for their glycosylation pattern by using tandem mass tag (TMT 10-plex) labeling,417 which is traditionally used in comparative proteomics studies. By alternating HCD/ETD/CID fragmentation, 1000 N- and O-glycopeptides could be identified in at least one of the HCG samples. Of these, 167 glycopeptides were significantly differentially expressed based on reporter signal intensities of the TMT tags. While such an approach is only suitable due to the three-way fragmentation, the study shows that traditional proteomics TMT labeling is suitable for relative quantitation of glycopeptides in less complex mixtures.
7.3.1. Targeted Methods for Glycopeptide Quantitation.
Targeted mass spectrometry techniques, specifically multiple reaction monitoring (MRM) on triple quadruple instruments418 and parallel reaction monitoring (PRM) on orbitraps,419 have been widely adopted in the proteomics community for the quantitative evaluation of proteins in clinical specimens. The application of targeted MS strategies in combination with stable isotope labeled (SIL) peptides420 is now starting to be adopted for quantitation of proteins in clinical chemistry laboratories.421,422 The application of targeted methods for (relative) glycopeptide quantitation is still in its infancy and was recently reviewed.423,424 It could be concluded that MRM in combination with glycoprotein enrichment improves sensitivity and specificity of the quantitation of minor glycoforms, specifically after optimization of the instrument settings.
In targeted mass spectrometry approaches, the fragmentation behavior of the analyte is of great importance. Currently, the fragmentation of glycopeptides in targeted approaches is typically performed using CID. It is well established that the glycosidic bonds are more labile in CID than peptide bonds, resulting in fragmentation spectra dominated by B-type oxonium ions (glycan fragments) and potentially low amounts of fragments of the glycosylated peptide (y-type ions) (Figure 6).38,425 This is different for oligomannose type glycopeptides, where CID fragmentation results in highly intense y-type ions, and B-ions are of much lower intensity.20
Therefore, initial studies toward the quantitation of glycopeptides employed the use of B-type oxonium ions (m/z 204, 274, 292, 366, 512, and/or 657). While these fragments are common to all glycopeptides, and thus do not provide high specificity, it was shown by Hong et al. that this strategy was suitable for the accurate quantitation of glycopeptides from IgG, IgA, and IgM directly from serum,20,38 and later, Ruhaak et al. expanded the method with six other abundant glycoproteins (Figure 28).426–429 A similar approach was used to quantify glycopeptides from human milk.430 Similarly, B-type oxonium ions were applied for the quantitation of glycopeptides from haptoglobin isolated from serum.431 In an alternative approach, more specific Y1 fragments were monitored to quantify glycopeptides from mouse serum.432 More recently, a strategy in which Y0 fragments are monitored was proposed, because the Y1 fragments contain the innermost GlcNAc, which may be fucosylated, thus resulting in Y1+F fragments.433 These fragment ions have also been used in DIA approaches for glycopeptides.434,435
Figure 28.
LC-MRM MS chromatograms for the relative quantitation of glycopeptides from abundant serum proteins. Both peptides (top) and glycopeptides can be monitored. Adapted from ref 427. Copyright 2018 American Chemical Society.
However, similarly to quantitation of glycans, a reference is required to allow for the comparison of quantitative results between samples. Here, several approaches have been employed. One could argue to assess the glycosylation profile independent of the protein concentration; therefore, quantitation relative to the protein amount, as determined by the measurement of a nonglycosylated peptide, was employed. Using this strategy, differential glycosylation patterns could be identified in serum samples from ovarian cancer patients compared to healthy controls426 as well as in samples from liver disease.431,436 Alternatively, ratios between glycopeptides were used to identify altered expression of glycopeptides in patients with colorectal cancer and hepatocellular carcinoma,437 and results were comparable to label-free MALDI-FTICR quantitation.
Recently, the use of isotopically labeled internal standards for the quantitation of glycopeptides from mABs was reported.433 Tryptic digestion was performed in either H216O or H218O to induce a 2 Da mass difference, and glycopeptides were then analyzed using targeted mass spectrometry on a high-resolution instrument (triple TOF). Indeed, this is a promising strategy, but it has to be mentioned that a mass shift of 2 Da is limited and as such requires the use of high resolution instruments. The application of targeted methods for quantitation of glycopeptides is a promising strategy. However, for accurate absolute quantification and adoption in clinical chemistry tests, stable isotope labeled glycopeptides will be necessary.
7.3.2. Importance of Enzymatic Digestion.
As with the quantitation of proteins,374,438 the stable and complete enzymatic digestion of glycoproteins is of the highest importance for quantitation of protein glycosylation by means of its glycopeptides. While in protein quantitation a choice for an ideal peptide can typically be made, this is not the case for glycopeptides, where the specific peptide with the glycosylation site needs to be quantified. Specifically, the glycosylation may hamper or resist proteolytic digestion.439 Recently, the preferential tryptic cleavage of high mannose, hybrid, α(2,3)-sialylated, and bisected glycoforms on IgG was observed over the other most abundant neutral, fucosylated glycoforms.440 This was most profoundly seen in native protein, but was not completely diminished by protein denaturation. Furthermore, mucins, which are highly O-glycosylated, have been reported to strongly resist proteolytic cleavage, which may even be part of their biological function.441 Therefore, the efficiency of proteolytic cleavage, as well as glycopeptide degradation, needs to be taken into account when employing glycopeptides to quantify protein glycosylation.
Similarly, the variations in ionization efficiencies and fragmentation behavior between different glycopeptides are concerns for label-free relative quantitation. While the ionization efficiencies seem to be rather similar between neutral and sialylated glycopeptides in positive mode ESI-MS, MALDI based analysis may be somewhat more problematic as signals of sialylated glycans may be reduced due to ion suppression or in-source decay. Furthermore, when using MRM or PRM, fragments may form more or less easily, for example, fragment m/z 366 will form twice as much from a glycopeptide carrying a biantennary nonsialylated glycan than from a glycopeptide carrying a monoantennary glycan.38 Therefore, care needs to be taken when representing relative abundances based on mass spectrometry results.
7.4. Quantitation of Glycoproteins
The quantitation of intact glycoproteins is still in its infancy, although it is anticipated that methods currently being developed for the quantitation of intact proteins will be equally applicable to intact glycoproteins. Because it is still not known what the specific effects of glycans on the ionization efficiency of glycoproteins are, the quantitation of the different glycoforms based solely on ion abundances may be limited. This approach cannot yet be applied to complicated mixtures, but approaches where glycoproteins are captured using antibodies, such as that currently applied to transferrin and apolipoprotein C-III442,443 using mass spectrometry based immunoassays (MSIA),444 have shown encouraging results. The technique is promising and may have more immediate applications with both individual glycoproteins and enriched fractions.
8. TOWARD RAPID-THROUGHPUT ANALYSIS
Although structural analyses of glycans and oligosaccharides remain challenging, there have been notable efforts in developing rapid-throughput methods. Rapid-throughput structural elucidation is likely not possible for oligosaccharides or any class of biomolecules, however, rapid identification can make rapid-throughput glycomics possible. This method has been used to study human milk oligosaccharides and childhood stunting. Totten et al. employed an HMO library created by Wu et al.11,445 containing HPLC retention times and accurate masses to profile with quantiation HMOs from human milk samples.446 They found that stunted children had mothers milk that were lower in fucosylated, sialylated, and total oligosaccharides compared to nonstunted subjects.447 The analysis monitored over 300 structures in each run.
To observe cancer or disease specific changes in glycans and glycoproteins for discovery of biomarkers, large-sample-size studies are also needed, which further necessitates rapid-throughput quantitative methods for glycomics and glycoproteomics. Untargeted glycoproteomic mapping for large sample sets is still a challenge due to the lack of robust data processing software to identify and quantify glycopeptides concurrently in a biological sample. Although stable isotope labeling methods introduced above such as TMT 10-plex have been applied to the quantitation of glycopeptides, it could only be performed in 10-sample sets.448 Rapid-throughput analysis can be performed with MRM in a targeted effort by quantitating glycosylation with site specificity in proteins. The speed and robustness of this methodology make it ideal for large sample sizes. It has been implemented to monitor site-specific glycosylation in serum proteins for ovarian cancer biomarker discovery in a study involving 168 subjects. A biomarker panel containing 11 glycoproteins was discovered and proved to increase the accuracy of ovarian cancer prediction when combined with CA-125, the most widely used biomarker for ovarian cancer.426,427 This MRM method for quantitation of immunoglobulin glycopeptides also provides an ideal tool to study autoimmune-associated glycan alterations for autoimmune biomarker discovery.449
While methods for rapid and global glycoroteomic analysis need further improvements, rapid-throughput methods for released glycan analysis employing various platforms have been developed and employed to discover and validate disease biomarkers. MALDI-MS is one of the most widely used methods for rapid analysis and has recently been applied to the discovery of glycan biomarkers for various diseases such as gastric cancer450 and hepatocellular carcinoma451 with over 200 subjects. Furthermore, with emerging high-throughput permethylation and derivatization methods, the stability and ionization efficiency of sialylated glycans can be greatly improved.109,452 UPLC with fluorescence detection is another technology for the analysis of the glycome in a rapid-throughput manner. Pucic et al. combined this method with a novel 96-well plate IgG isolation platform and applied it to characterize 2-AB-labeled N-glycans released from isolated IgG from plasma of 2298 individuals.453 High variability in the 24 chromatographically separated glycans was observed between individuals. For simultaneous quantitation and in-depth structural characterization of glycans in complicated biological samples, however, high-throughput LC-MS methods are necessary. Ruhaak et al. demonstrated that chip-based nanoLC-TOF-MS is a highly stable and repeatable platform for large-scale serum N-glycan studies.39 This platform was applied to an epithelial ovarian cancer cohort with approximately 300 samples379 and a nonsmall cell lung cancer cohort with over 600 samples454 for glycan biomarker discovery.
9. CONCLUSIONS AND FUTURE DIRECTIONS
New analytical methods and techniques are advancing glycomic and glycoproteomic analyses at a rapid rate. The next step for glycomics and glycoproteomics is to achieve rapid-throughput analysis with quantitation. Glycomic analysis has initially focused on obtaining monosaccharide composition. When used with the known biological synthesis, it could provide putative structures particularly for N-glycans, which is sufficient for determining whether a compound was fucosylated, sialylated, or mainly a high mannose type. However, it yielded little information regarding the sequences or the linkages of glycans. For complete structures, chromatographic separation is necessary to profile components of the glycome. However, de novo structural analysis of oligosaccharides is not feasible in a rapid-throughput manner. In this regard, comprehensive structural analysis may take the same path as proteomics and metabolomics, that is, creating a reference library of structures and identity elements for each entry. Toward this end, the creation of structural libraries with annotated structures and identifiers such as chromatographic retention times and tandem MS spectra would advance this process considerably.
Glycoproteomic analysis can now provide simultaneous identification of proteins and characterization of the associated glycoforms in an automated manner. As the bioinformatic software improves, characterization of site heterogeneity will improve, providing more sites and more glycoforms. The more challenging aspect will be the structural analysis of glycans associated with specific sites. This should be the target of future efforts.
The analytical methods are nonetheless sufficiently advanced to solve a host of biological problems in glycobiology. What is most welcomed is for those with little expertise in separation or mass spectrometry to take advantage of these tools and solve problems in diseases and fundamental biology. It is perhaps in this last challenge, where the greatest obstacles will be.
ACKNOWLEDGMENTS
Funding provided by the National Institutes of Health (R01HD061923, RO1AT008759, and R01GM049077) is gratefully acknowledged.
ABBREVIATIONS
- 2-AA
2-aminobenzoic acid
- 2-AB
2-aminobenzamide
- AI-ECD
activated ion ECD
- CE
capillary electrophoresis
- CID
collision induced dissociation
- ECD
electron capture dissociation
- EED
electron excitation dissociation
- ETD
electron transfer dissociation
- EThcD
electron-transfer/higher-energy collision dissociation
- ERLIC
electrostatic repulsion hydrophilic interaction chromatography
- ESI
electrospray ionization
- FTICR
Fourier transform ion cyclotron resonance
- HILIC
hydrophilic interaction liquid chromatography
- HPLC
high performance liquid chromatography
- HR
high resolution
- IRMPD
infrared multiphoton dissociation
- LAC
lectin affinity chromatography
- mAB
monoclonal antibody
- MALDI
matrix-assisted laser desorption/ionization
- MRM
multiple reaction monitoring
- MS
mass spectrometry
- PNGase
F peptide-N-glycosidase F
- PGC
porous graphitized carbon
- QQQ
triple quadrupole
- RPLC
reversed-phase liquid chromatography
- TOF
time-of-flight
- UPLC
ultrahigh performance liquid chromatography
- UVPD
ultraviolet photodissociation
Biographies
L. Renee Ruhaak is an assistant professor in the Department of Clinical Chemistry and Laboratory Medicine at the Leiden University Medical Center (LUMC). She obtained her Ph.D. at the LUMC, which was followed by a postdoctoral appointment at the University of California, Davis, and an assistant professor position at MD Anderson Cancer Center. Her main interest is in the application of mass spectrometry for the discovery, validation, and translation of (glycosylation-based) biomarkers for health and disease states.
Gege Xu studied chemistry at Peking University in China, where she received her B.S. degree in 2013. She is currently completing her Ph.D. research at the University of California, Davis, under the supervision of Dr. Carlito B. Lebrilla. Her studies focus on developing qualitative and quantitative methods for the analysis of dietary saccharides and cell surface glycans and glycoproteins using LC-MS/MS.
Qiongyu Li graduated from Hong Kong Polytechnic University in 2014 with a B.S. in chemical technology. Currently, she is a Ph.D. candidate at the University of California, Davis, where her research focuses on the analysis of glycoproteins for biomarker discovery and the method development of click chemistry involved mass spectrometry for studying the role of membrane glycosylation in cell-cell interaction.
Elisha Goonatilleke received her B.S. in 2013 and continued with her Ph.D. at the University of California, Davis. Currently, she is a graduate student in the group of Dr. Carlito B. Lebrilla. Her field of study is in analytical chemistry with an emphasis on mass spectrometry. She focuses on the identification, characterization, and quantification of glycoproteins and oligosaccharides in human milk.
Carlito B. Lebrilla completed his B.S. from University of California, Irvine, and his Ph.D. from University of California, Berkeley. He was a NATO-NSF and an Alexander von Humboldt postdoctoral fellow with Prof. Helmut Schwarz at the Technical University in Berlin. He was also a UC President’s postdoctoral fellow at the University of California, Irvine. He has been on the faculty at UC Davis since 1987 and is currently a Distinguished Professor in the Chemistry Department and in Biochemistry and Molecular Medicine. He is on the editorial board of several mass spectrometry journals. His research focus is on bioanalytical chemistry with an emphasis on oligosaccharides and glycoconjugates.
Footnotes
The authors declare no competing financial interest.
REFERENCES
- (1).Akan I; Olivier-Van Stichelen S; Bond MR; Hanover JA Nutrient-driven O-GlcNAc in proteostasis and neurodegeneration. J. Neurochem 2018, 144, 7–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (2).Spiro RG Protein glycosylation: nature, distribution, enzymatic formation, and disease implications of glycopeptide bonds. Glycobiology 2002, 12, 43R–56R. [DOI] [PubMed] [Google Scholar]
- (3).Ly M; Leach FE 3rd; Laremore TN; Toida T; Amster IJ; Linhardt RJ The proteoglycan bikunin has a defined sequence. Nat. Chem. Biol 2011, 7, 827–833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (4).Ma JF; Hart GW O-GlcNAc profiling: from proteins to proteomes. Clin. Proteomics 2014, 11, 8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (5).Staples GO; Zaia J Analysis of Glycosaminoglycans Using Mass Spectrometry. Curr. Proteomics 2011, 8, 325–336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (6).Laine RA A calculation of all possible oligosaccharide isomers both branched and linear yields 1.05 × 10(12) structures for a reducing hexasaccharide: the Isomer Barrier to development of single-method saccharide sequencing or synthesis systems. Glycobiology 1994, 4, 759–767. [DOI] [PubMed] [Google Scholar]
- (7).German JB; Freeman SL; Lebrilla CB; Mills DA Human milk oligosaccharides: evolution, structures and bioselectivity as substrates for intestinal bacteria. Nestle Nutr Workshop Ser. Pediatr Program 2008, 62, 205–218 discussion 218–222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (8).Kronewitter SR; An HJ; de Leoz ML; Lebrilla CB; Miyamoto S; Leiserowitz GS The development of retrosynthetic glycan libraries to profile and classify the human serum N-linked glycome. Proteomics 2009, 9, 2986–2994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (9).Song T; Aldredge D; Lebrilla CB A Method for In-Depth Structural Annotation of Human Serum Glycans That Yields Biological Variations. Anal. Chem 2015, 87, 7754–7762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (10).Wu S; Grimm R; German JB; Lebrilla CB Annotation and structural analysis of sialylated human milk oligosaccharides. J. Proteome Res 2011, 10, 856–868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (11).Wu S; Tao N; German JB; Grimm R; Lebrilla CB Development of an annotated library of neutral human milk oligosaccharides. J. Proteome Res 2010, 9, 4138–4151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (12).Huang J; Guerrero A; Parker E; Strum JS; Smilowitz JT; German JB; Lebrilla CB Site-specific glycosylation of secretory immunoglobulin A from human colostrum. J. Proteome Res 2015, 14, 1335–1349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (13).Omtvedt LA; Royle L; Husby G; Sletten K; Radcliffe CM; Harvey DJ; Dwek RA; Rudd PM Glycan analysis of monoclonal antibodies secreted in deposition disorders indicates that subsets of plasma cells differentially process IgG glycans. Arthritis Rheum. 2006, 54, 3433–3440. [DOI] [PubMed] [Google Scholar]
- (14).Ruhaak LR; Barkauskas DA; Torres J; Cooke CL; Wu LD; Stroble C; Ozcan S; Williams CC; Camorlinga M; Rocke DM; et al. The Serum Immunoglobulin G Glycosylation Signature of Gastric Cancer. EuPa Open Proteomics 2015, 6, 1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (15).An HJ; Tillinghast JS; Woodruff DL; Rocke DM; Lebrilla CB A new computer program (GlycoX) to determine simultaneously the glycosylation sites and oligosaccharide heterogeneity of glycoproteins. J. Proteome Res 2006, 5, 2800–2808. [DOI] [PubMed] [Google Scholar]
- (16).Moremen KW; Tiemeyer M; Nairn AV Vertebrate protein glycosylation: diversity, synthesis and function. Nat. Rev. Mol. Cell Biol 2012, 13, 448–462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (17).Schachter H The joys of HexNAc. The synthesis and function of N-and O-glycan branches. Glycoconjugate J. 2000, 17, 465–483. [DOI] [PubMed] [Google Scholar]
- (18).You X; Qin H; Ye M Recent advances in methods for the analysis of protein o-glycosylation at proteome level. J. Sep. Sci 2018, 41, 248–261. [DOI] [PubMed] [Google Scholar]
- (19).Cao L; Qu Y; Zhang ZR; Wang Z; Prytkova I; Wu S Intact glycopeptide characterization using mass spectrometry. Expert Rev. Proteomics 2016, 13, 513–522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (20).Hong Q; Ruhaak LR; Stroble C; Parker E; Huang J; Maverakis E; Lebrilla CB A Method for Comprehensive Glycosite-Mapping and Direct Quantitation of Serum Glycoproteins. J. Proteome Res 2015, 14, 5179–5192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (21).Yang N; Goonatilleke E; Park D; Song T; Fan G; Lebrilla CB Quantitation of Site-Specific Glycosylation in Manufactured Recombinant Monoclonal Antibody Drugs. Anal. Chem 2016, 88, 7091–7100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (22).James P Protein identification in the post-genome era: the rapid rise of proteomics. Q. Rev. Biophys 1997, 30, 279–331. [DOI] [PubMed] [Google Scholar]
- (23).Harvey DJ Analysis of carbohydrates and glycoconjugates by matrix-assisted laser desorption/ionization mass spectrometry: An update covering the period 1999–2000. Mass Spectrom. Rev 2006, 25, 595–662. [DOI] [PubMed] [Google Scholar]
- (24).Harvey DJ Analysis of carbohydrates and glycoconjugates by matrix-assisted laser desorption/ionization mass spectrometry: An update for 2003–2004. Mass Spectrom. Rev 2009, 28, 273–361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (25).Harvey DJ Analysis of carbohydrates and glycoconjugates by matrix-assisted laser desorption/ionization mass spectrometry: an update for the period 2005–2006. Mass Spectrom. Rev 2011, 30, 1–100. [DOI] [PubMed] [Google Scholar]
- (26).Harvey DJ Analysis of carbohydrates and glycoconjugates by matrix-assisted laser desorption/ionization mass spectrometry: an update for 2007–2008. Mass Spectrom. Rev 2012, 31, 183–311. [DOI] [PubMed] [Google Scholar]
- (27).Harvey DJ Analysis of carbohydrates and glycoconjugates by matrix-assisted laser desorption/ionization mass spectrometry: an update for 2009–2010. Mass Spectrom. Rev 2015, 34, 268–422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (28).Mulagapati S; Koppolu V; Raju TS Decoding of O-Linked Glycosylation by Mass Spectrometry. Biochemistry 2017, 56, 1218–1226. [DOI] [PubMed] [Google Scholar]
- (29).Banazadeh A; Veillon L; Wooding KM; Zabet-Moghaddam M; Mechref Y Recent advances in mass spectrometric analysis of glycoproteins. Electrophoresis 2017, 38, 162–189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (30).Kailemia MJ; Ruhaak LR; Lebrilla CB; Amster IJ Oligosaccharide analysis by mass spectrometry: a review of recent developments. Anal. Chem 2014, 86, 196–212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (31).Palaniappan KK; Bertozzi CR Chemical Glycoproteomics. Chem. Rev 2016, 116, 14277–14306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (32).Xu G; Davis JC; Goonatilleke E; Smilowitz JT; German JB; Lebrilla CB Absolute Quantitation of Human Milk Oligosaccharides Reveals Phenotypic Variations during Lactation. J. Nutr 2017, 147, 117–124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (33).Zhang H; Wang Z; Stupak J; Ghribi O; Geiger JD; Liu QY; Li J Targeted glycomics by selected reaction monitoring for highly sensitive glycan compositional analysis. Proteomics 2012, 12, 2510–2522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (34).Park Y; Lebrilla CB Application of Fourier transform ion cyclotron resonance mass spectrometry to oligosaccharides. Mass Spectrom. Rev 2005, 24, 232–264. [DOI] [PubMed] [Google Scholar]
- (35).An HJ; Lebrilla CB A glycomics approach to the discovery of potential cancer biomarkers. Methods Mol. Biol 2010, 600, 199–213. [DOI] [PubMed] [Google Scholar]
- (36).Totten SM; Feasley CL; Bermudez A; Pitteri SJ Parallel Comparison of N-Linked Glycopeptide Enrichment Techniques Reveals Extensive Glycoproteomic Analysis of Plasma Enabled by SAX-ERLIC. J. Proteome Res 2017, 16, 1249–1260. [DOI] [PubMed] [Google Scholar]
- (37).Wang X; Emmett MR; Marshall AG Liquid chromatography electrospray ionization Fourier transform ion cyclotron resonance mass spectrometric characterization of N-linked glycans and glycopeptides. Anal. Chem 2010, 82, 6542–6548. [DOI] [PubMed] [Google Scholar]
- (38).Hong Q; Lebrilla CB; Miyamoto S; Ruhaak LR Absolute quantitation of immunoglobulin G and its glycoforms using multiple reaction monitoring. Anal. Chem 2013, 85, 8585–8593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (39).Ruhaak LR; Taylor SL; Miyamoto S; Kelly K; Leiserowitz GS; Gandara D; Lebrilla CB; Kim K Chip-based nLC-TOF-MS is a highly stable technology for large-scale high-throughput analyses. Anal. Bioanal. Chem 2013, 405, 4953–4958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (40).Hong Q; Ruhaak LR; Totten SM; Smilowitz JT; German JB; Lebrilla CB Label-free absolute quantitation of oligosaccharides using multiple reaction monitoring. Anal. Chem 2014, 86, 2640–2647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (41).Penn SG; Cancilla MT; Lebrilla CB Collision-induced dissociation of branched oligosaccharide ions with analysis and calculation of relative dissociation thresholds. Anal. Chem 1996, 68, 2331–2339. [DOI] [PubMed] [Google Scholar]
- (42).Ngoka LC; Gal JF; Lebrilla CB Effects of Cations and Charge Types on the Metastable Decay-Rates of Oligosaccharides. Anal. Chem 1994, 66, 692–698. [DOI] [PubMed] [Google Scholar]
- (43).Spengler B; Kirsch D; Kaufmann R; Cotter RJ Metastable Decay of Peptides and Proteins in Matrix-Assisted Laser-Desorption Mass-Spectrometry. Rapid Commun. Mass Spectrom 1991, 5, 198–202. [Google Scholar]
- (44).Zhang JH; LaMotte LT; Dodds ED; Lebrilla CB Atmospheric pressure MALDI Fourier transform mass spectrometry of labile oligosaccharides. Anal. Chem 2005, 77, 4429–4438. [DOI] [PubMed] [Google Scholar]
- (45).An HJ; Miyamoto S; Lancaster KS; Kirmiz C; Li B; Lam KS; Leiserowitz GS; Lebrilla CB Profiling of glycans in serum for the discovery of potential biomarkers for ovarian cancer. J. Proteome Res 2006, 5, 1626–1635. [DOI] [PubMed] [Google Scholar]
- (46).Budnik BA; Lee RS; Steen JAJ Global methods for protein glycosylation analysis by mass spectrometry. Biochim. Biophys. Acta, Proteins Proteomics 2006, 1764, 1870–1880. [DOI] [PubMed] [Google Scholar]
- (47).Dongre AR; Wysocki VH Linkage Position Determination of Lithium-Cationized Disaccharides by Surface-Induced Dissociation Tandem Mass-Spectrometry. Org. Mass Spectrom 1994, 29, 700–702. [Google Scholar]
- (48).An HJ; Lebrilla CB Structure elucidation of native N- and O-linked glycans by tandem mass spectrometry (tutorial). Mass Spectrom. Rev 2011, 30, 560–578. [DOI] [PubMed] [Google Scholar]
- (49).Wuhrer M; Deelder AM; van der Burgt YE Mass spectrometric glycan rearrangements. Mass Spectrom. Rev 2011, 30, 664–680. [DOI] [PubMed] [Google Scholar]
- (50).Dell A Preparation and desorption mass spectrometry of permethyl and peracetyl derivatives of oligosaccharides. Methods Enzymol. 1990, 193, 647–660. [DOI] [PubMed] [Google Scholar]
- (51).Russell SC; Czerwieniec G; Lebrilla C; Steele P; Riot V; Coffee K; Frank M; Gard EE Achieving high detection sensitivity (14 zmol) of biomolecular ions in bioaerosol mass spectrometry. Anal. Chem 2005, 77, 4734–4741. [DOI] [PubMed] [Google Scholar]
- (52).Prien JM; Ashline DJ; Lapadula AJ; Zhang H; Reinhold VN The High Mannose Glycans from Bovine Ribonuclease B Isomer Characterization by Ion Trap MS. J. Am. Soc. Mass Spectrom 2009, 20, 539–556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (53).Seipert RR; Dodds ED; Clowers BH; Beecroft SM; German JB; Lebrilla CB Factors that influence fragmentation behavior of N-linked glycopeptide ions. Anal. Chem 2008, 80, 3684–3692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (54).Seipert RR; Dodds ED; Lebrilla CB Exploiting Differential Dissociation Chemistries of O-Linked Glycopeptide Ions for the Localization of Mucin-Type Protein Glycosylation. J. Proteome Res 2009, 8, 493–501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (55).Haag AM Mass Analyzers and Mass Spectrometers. Adv. Exp. Med. Biol 2016, 919, 157–169. [DOI] [PubMed] [Google Scholar]
- (56).Sobott F; Watt SJ; Smith J; Edelmann MJ; Kramer HB; Kessler BM Comparison of CID Versus ETD Based MS/MS Fragmentation for the Analysis of Protein Ubiquitination. J. Am. Soc. Mass Spectrom 2009, 20, 1652–1659. [DOI] [PubMed] [Google Scholar]
- (57).Mechref Y Use of CID/ETD Mass Spectrometry to Analyze Glycopeptides. Curr. Protoc. Protein Sci 2012, 12, 12.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (58).Alley WR; Mechref Y; Novotny MV Characterization of glycopeptides by combining collision-induced dissociation and electron-transfer dissociation mass spectrometry data. Rapid Commun. Mass Spectrom 2009, 23, 161–170. [DOI] [PubMed] [Google Scholar]
- (59).Ford KL; Zeng W; Heazlewood JL; Bacic A Characterization of protein N-glycosylation by tandem mass spectrometry using complementary fragmentation techniques. Front. Plant Sci 2015, 6, 674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (60).Windwarder M; Yelland T; Djordjevic S; Altmann F Detailed characterization of the O-linked glycosylation of the neuropilin-1 c/MAM-domain. Glycoconjugate J. 2016, 33, 387–397. [DOI] [PubMed] [Google Scholar]
- (61).Moh ESX; Lin CH; Thaysen-Andersen M; Packer NH Site-Specific N-Glycosylation of Recombinant Pentameric and Hexameric Human IgM. J. Am. Soc. Mass Spectrom 2016, 27, 1143–1155. [DOI] [PubMed] [Google Scholar]
- (62).Frese CK; Altelaar AFM; van den Toorn H; Nolting D; Griep-Raming J; Heck AJR; Mohammed S Toward Full Peptide Sequence Coverage by Dual Fragmentation Combining Electron-Transfer and Higher-Energy Collision Dissociation Tandem Mass Spectrometry. Anal. Chem 2012, 84, 9668–9673. [DOI] [PubMed] [Google Scholar]
- (63).Parker BL; Thaysen-Andersen M; Fazakerley DJ; Holliday M; Packer NH; James DE Terminal galactosylation and sialylation switching on membrane glycoproteins upon TNF-alpha-induced insulin resistance in adipocytes. Mol. Cell. Proteomics 2016, 15, 141–153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (64).Yu Q; Wang B; Chen Z; Urabe G; Glover MS; Shi X; Guo L-W; Kent KC; Li L Electron-Transfer/Higher-Energy Collision Dissociation (EThcD)-Enabled Intact Glycopeptide/Glycoproteome Characterization. J. Am. Soc. Mass Spectrom 2017, 28, 1751–1764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (65).Håkansson K; Cooper HJ; Emmett MR; Costello CE; Marshall AG; Nilsson CL Electron capture dissociation and infrared multiphoton dissociation MS/MS of an N-glycosylated tryptic peptide to yield complementary sequence information. Anal. Chem 2001, 73, 4530–4536. [DOI] [PubMed] [Google Scholar]
- (66).Manri N; Satake H; Kaneko A; Hirabayashi A; Baba T; Sakamoto T Glycopeptide Identification Using Liquid-Chromatography-Compatible Hot Electron Capture Dissociation in a Radio-Frequency-Quadrupole Ion Trap. Anal. Chem 2013, 85, 2056–2063. [DOI] [PubMed] [Google Scholar]
- (67).Yu X; Jiang Y; Chen Y; Huang Y; Costello CE; Lin C Detailed Glycan Structural Characterization by Electronic Excitation Dissociation. Anal. Chem 2013, 85, 10017–10021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (68).Brodbelt JS Photodissociation mass spectrometry: new tools for characterization of biological molecules. Chem. Soc. Rev 2014, 43, 2757–2783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (69).Zhang L; Reilly JP Extracting Both Peptide Sequence and Glycan Structural Information by 157 nm Photodissociation of N-Linked Glycopeptides. J. Proteome Res 2009, 8, 734–742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (70).Madsen JA; Ko BJ; Xu H; Iwashkiw JA; Robotham SA; Shaw JB; Feldman MF; Brodbelt JS Concurrent automated sequencing of the glycan and peptide portions of O-linked glycopeptide anions by ultraviolet photodissociation mass spectrometry. Anal. Chem 2013, 85, 9253–9261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (71).Ko BJ; Brodbelt JS Comparison of Glycopeptide Fragmentation by Collision Induced Dissociation and Ultraviolet Photodissociation. Int. J. Mass Spectrom 2015, 377, 385–392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (72).Ko BJ; Brodbelt JS 193 nm Ultraviolet Photodissociation of Deprotonated Sialylated Oligosaccharides. Anal. Chem 2011, 83, 8192–8200. [DOI] [PubMed] [Google Scholar]
- (73).Cotham VC; Brodbelt JS Characterization of Therapeutic Monoclonal Antibodies at the Subunit-Level using Middle-Down 193 nm Ultraviolet Photodissociation. Anal. Chem 2016, 88, 4004–4013. [DOI] [PubMed] [Google Scholar]
- (74).Ji Y; Xiong Z; Huang G; Liu J; Zhang Z; Liu Z; Ou J; Ye M; Zou H Efficient enrichment of glycopeptides using metal-organic frameworks by hydrophilic interaction chromatography. Analyst 2014, 139, 4987–4993. [DOI] [PubMed] [Google Scholar]
- (75).Sandoval WN; Arellano F; Arnott D; Raab H; Vandlen R; Lill JR Rapid removal of N-linked oligosaccharides using microwave assisted enzyme catalyzed deglycosylation. Int. J. Mass Spectrom 2007, 259, 117–123. [Google Scholar]
- (76).Ren X; Bai H; Pan Y; Tong W; Qin P; Yan H; Deng S; Zhong R; Qin W; Qian X A graphene oxide-based immobilized PNGase F reagent for highly efficient N-glycan release and MALDI-TOF MS profiling. Anal. Methods 2014, 6, 2518–2525. [Google Scholar]
- (77).Tretter V; Altmann F; Marz L Peptide-N4-(N-Acetyl-Beta-Glucosaminyl)Asparagine Amidase-F Cannot Release Glycans with Fucose Attached Alpha-1-]3 to the Asparagine-Linked N-Acetylglucosamine Residue. Eur. J. Biochem 1991, 199, 647–652. [DOI] [PubMed] [Google Scholar]
- (78).Wang T; Cai ZP; Gu XQ; Ma HY; Du YM; Huang K; Voglmeir J; Liu L Discovery and characterization of a novel extremely acidic bacterial N-glycanase with combined advantages of PNGase F and A. Biosci. Rep 2014, 34, 673–684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (79).Maley F; Trimble RB; Tarentino AL; Plummer TH Characterization of Glycoproteins and Their Associated Oligosaccharides through the Use of Endoglycosidases. Anal. Biochem 1989, 180, 195–204. [DOI] [PubMed] [Google Scholar]
- (80).Brooks MM; Savage AV The substrate specificity of the enzyme endo-alpha-N-acetyl-D-galactosaminidase from Diplococcus pneumonia. Glycoconjugate J. 1997, 14, 183–190. [DOI] [PubMed] [Google Scholar]
- (81).Kozak RP; Royle L; Gardner RA; Bondt A; Fernandes DL; Wuhrer M Improved nonreductive O-glycan release by hydrazinolysis with ethylenediaminetetraacetic acid addition. Anal. Biochem 2014, 453, 29–37. [DOI] [PubMed] [Google Scholar]
- (82).Merry AH; Neville DCA; Royle L; Matthews B; Harvey DJ; Dwek RA; Rudd PM Recovery of intact 2-aminobenzamide-labeled O-glycans released from glycoproteins by hydrazinolysis. Anal. Biochem 2002, 304, 91–99. [DOI] [PubMed] [Google Scholar]
- (83).Song X; Ju H; Lasanajak Y; Kudelka MR; Smith DF; Cummings RD Oxidative release of natural glycans for functional glycomics. Nat. Methods 2016, 13, 528–534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (84).Goso Y; Sugaya T; Ishihara K; Kurihara M Comparison of Methods to Release Mucin-Type O-Glycans for Glycomic Analysis. Anal. Chem 2017, 89, 8870–8876. [DOI] [PubMed] [Google Scholar]
- (85).Ruhaak LR; Zauner G; Huhn C; Bruggink C; Deelder AM; Wuhrer M Glycan labeling strategies and their use in identification and quantification. Anal. Bioanal. Chem 2010, 397, 3457–3481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (86).Pabst M; Kolarich D; Poltl G; Dalik T; Lubec G; Hofinger A; Altmann F Comparison of fluorescent labels for oligosaccharides and introduction of a new postlabeling purification method. Anal. Biochem 2009, 384, 263–273. [DOI] [PubMed] [Google Scholar]
- (87).Anumula KR Advances in fluorescence derivatization methods for high-performance liquid chromatographic analysis of glycoprotein carbohydrates. Anal. Biochem 2006, 350, 1–23. [DOI] [PubMed] [Google Scholar]
- (88).Honda S; Akao E; Suzuki S; Okuda M; Kakehi K; Nakamura J High-Performance Liquid-Chromatography of Reducing Carbohydrates as Strongly Ultraviolet-Absorbing and Electrochemically Sensitive 1-Phenyl-3-Methyl-5-Pyrazolone Derivatives. Anal. Biochem 1989, 180, 351–357. [DOI] [PubMed] [Google Scholar]
- (89).Kakehi K; Suzuki S; Honda S; Lee YC Precolumn labeling of reducing carbohydrates with 1-(p-methoxy)phenyl-3-methyl-5-pyrazolone: analysis of neutral and sialic acid-containing oligosaccharides found in glycoproteins. Anal. Biochem 1991, 199, 256–268. [DOI] [PubMed] [Google Scholar]
- (90).Gil GC; Kim YG; Kim BG A relative and absolute quantification of neutral N-linked oligosaccharides using modification with carboxymethyl trimethylammonium hydrazide and matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Anal. Biochem 2008, 379, 45–59. [DOI] [PubMed] [Google Scholar]
- (91).Jang KS; Kim YG; Gil GC; Park SH; Kim BG Mass spectrometric quantification of neutral and sialylated N-glycans from a recombinant therapeutic glycoprotein produced in the two Chinese hamster ovary cell lines. Anal. Biochem 2009, 386, 228–236. [DOI] [PubMed] [Google Scholar]
- (92).Jiao J; Yang L; Zhang Y; Lu H Hydrazinonicotinic acid derivatization for selective ionization and improved glycan structure characterization by MALDI-MS. Analyst 2015, 140, 5475–5480. [DOI] [PubMed] [Google Scholar]
- (93).Zhao MZ; Zhang YW; Yuan F; Deng Y; Liu JX; Zhou YL; Zhang XX Hydrazino-s-triazine based labelling reagents for highly sensitive glycan analysis via liquid chromatography-electrospray mass spectrometry. Talanta 2015, 144, 992–997. [DOI] [PubMed] [Google Scholar]
- (94).Zhang Y; Peng Y; Bin Z; Wang H; Lu H Highly specific purification of N-glycans using phosphate-based derivatization as an affinity tag in combination with Ti(4+)-SPE enrichment for mass spectrometric analysis. Anal. Chim. Acta 2016, 934, 145–151. [DOI] [PubMed] [Google Scholar]
- (95).Li L; Jiao J; Cai Y; Zhang Y; Lu H Fluorinated carbon tag derivatization combined with fluorous solid-phase extraction: a new method for the highly sensitive and selective mass spectrometric analysis of glycans. Anal. Chem 2015, 87, 5125–5131. [DOI] [PubMed] [Google Scholar]
- (96).Jiang K; Zhu H; Xiao C; Liu D; Edmunds G; Wen L; Ma C; Li J; Wang PG Solid-phase reductive amination for glycomic analysis. Anal. Chim. Acta 2017, 962, 32–40. [DOI] [PubMed] [Google Scholar]
- (97).Wang C; Fan W; Zhang P; Wang Z; Huang L One-pot nonreductive O-glycan release and labeling with 1-phenyl-3-methyl-5-pyrazolone followed by ESI-MS analysis. Proteomics 2011, 11, 4229–4242. [DOI] [PubMed] [Google Scholar]
- (98).Wang C; Zhang P; Jin W; Li L; Qiang S; Zhang Y; Huang L; Wang Z Quantitative O-glycomics based on improvement of the one-pot method for nonreductive O-glycan release and simultaneous stable isotope labeling with 1-(d0/d5)phenyl-3-methyl-5-pyrazolone followed by mass spectrometric analysis. J. Proteomics 2017, 150, 18–30. [DOI] [PubMed] [Google Scholar]
- (99).Yang S; Hoti N; Yang W; Liu Y; Chen L; Li S; Zhang H Simultaneous analyses of N-linked and O-linked glycans of ovarian cancer cells using solid-phase chemoenzymatic method. Clin. Proteomics 2017, 14, 3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (100).Yang S; Hu Y; Sokoll L; Zhang H Simultaneous quantification of N- and O-glycans using a solid-phase method. Nat. Protoc 2017, 12, 1229–1244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (101).Ciucanu I; Kerek F A Simple and Rapid Method for the Permethylation of Carbohydrates. Carbohydr. Res 1984, 131, 209–217. [Google Scholar]
- (102).Ciucanu I; Costello CE Elimination of oxidative degradation during the per-O-methylation of carbohydrates. J. Am. Chem. Soc 2003, 125, 16213–16219. [DOI] [PubMed] [Google Scholar]
- (103).Kang P; Mechref Y; Klouckova I; Novotny MV Solid-phase permethylation of glycans for mass spectrometric analysis. Rapid Commun. Mass Spectrom 2005, 19, 3421–3428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (104).Shubhakar A; Kozak RP; Reiding KR; Royle L; Spencer DI; Fernandes DL; Wuhrer M Automated High-Throughput Permethylation for Glycosylation Analysis of Biologics Using MALDI-TOF-MS. Anal. Chem 2016, 88, 8562–8569. [DOI] [PubMed] [Google Scholar]
- (105).Hu Y; Borges CR A spin column-free approach to sodium hydroxide-based glycan permethylation. Analyst 2017, 142, 2748–2759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (106).Wheeler SF; Domann P; Harvey DJ Derivatization of sialic acids for stabilization in matrix-assisted laser desorption/ionization mass spectrometry and concomitant differentiation of alpha(2–> 3)- and alpha(2–> 6)-isomers. Rapid Commun. Mass Spectrom 2009, 23, 303–312. [DOI] [PubMed] [Google Scholar]
- (107).Alley WR; Novotny MV Glycomic Analysis of Sialic Acid Linkages in Glycans Derived from Blood Serum Glycoproteins. J. Proteome Res 2010, 9, 3062–3072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (108).Tousi F; Bones J; Hancock WS; Hincapie M Differential chemical derivatization integrated with chromatographic separation for analysis of isomeric sialylated N-glycans: a nano-hydrophilic interaction liquid chromatography-MS platform. Anal. Chem 2013, 85, 8421–8428. [DOI] [PubMed] [Google Scholar]
- (109).Reiding KR; Blank D; Kuijper DM; Deelder AM; Wuhrer M High-throughput profiling of protein N-glycosylation by MALDI-TOF-MS employing linkage-specific sialic acid esterification. Anal. Chem 2014, 86, 5784–5793. [DOI] [PubMed] [Google Scholar]
- (110).de Haan N; Reiding KR; Haberger M; Reusch D; Falck D; Wuhrer M Linkage-specific sialic acid derivatization for MALDI-TOF-MS profiling of IgG glycopeptides. Anal. Chem 2015, 87, 8284–8291. [DOI] [PubMed] [Google Scholar]
- (111).Yang S; Jankowska E; Kosikova M; Xie H; Cipollo J Solid-Phase Chemical Modification for Sialic Acid Linkage Analysis: Application to Glycoproteins of Host Cells Used in Influenza Virus Propagation. Anal. Chem 2017, 89, 9508–9517. [DOI] [PubMed] [Google Scholar]
- (112).Bladergroen MR; Reiding KR; Hipgrave Ederveen ALH; Vreeker GCM; Clerc F; Holst S; Bondt A; Wuhrer M; van der Burgt YEM Automation of High-Throughput Mass Spectrometry-Based Plasma N-Glycome Analysis with Linkage-Specific Sialic Acid Esterification. J. Proteome Res 2015, 14, 4080–4086. [DOI] [PubMed] [Google Scholar]
- (113).Li H; Gao W; Feng X; Liu BF; Liu X MALDI-MS analysis of sialylated N-glycan linkage isomers using solid-phase two step derivatization method. Anal. Chim. Acta 2016, 924, 77–85. [DOI] [PubMed] [Google Scholar]
- (114).Jiang K; Zhu H; Li L; Guo Y; Gashash E; Ma C; Sun X; Li J; Zhang L; Wang PG Sialic acid linkage-specific permethylation for improved profiling of protein glycosylation by MALDI-TOF MS. Anal. Chim. Acta 2017, 981, 53–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (115).Nishikaze T; Tsumoto H; Sekiya S; Iwamoto S; Miura Y; Tanaka K Differentiation of Sialyl Linkage Isomers by One-Pot Sialic Acid Derivatization for Mass Spectrometry-Based Glycan Profiling. Anal. Chem 2017, 89, 2353–2360. [DOI] [PubMed] [Google Scholar]
- (116).Walker SH; Carlisle BC; Muddiman DC Systematic comparison of reverse phase and hydrophilic interaction liquid chromatography platforms for the analysis of N-linked glycans. Anal. Chem 2012, 84, 8198–8206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (117).Alley WR; Madera M; Mechref Y; Novotny MV Chip-based Reversed-phase Liquid Chromatography-Mass Spectrometry of Permethylated N-Linked Glycans: A Potential Methodology for Cancer-biomarker Discovery. Anal. Chem 2010, 82, 5095–5106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (118).Hu Y; Shihab T; Zhou S; Wooding K; Mechref Y LC-MS/MS of permethylated N-glycans derived from model and human blood serum glycoproteins. Electrophoresis 2016, 37, 1498–1505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (119).Ritamo I; Rabina J; Natunen S; Valmu L Nanoscale reversed-phase liquid chromatography-mass spectrometry of permethylated N-glycans. Anal. Bioanal. Chem 2013, 405, 2469–2480. [DOI] [PubMed] [Google Scholar]
- (120).Vreeker GC; Wuhrer M Reversed-phase separation methods for glycan analysis. Anal. Bioanal. Chem 2017, 409, 359–378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (121).Bereman MS; Williams TI; Muddiman DC Development of a nanoLC LTQ Orbitrap Mass Spectrometric Method for Profiling Glycans Derived from Plasma from Healthy, Benign Tumor Control, and Epithelial Ovarian Cancer Patients. Anal. Chem 2009, 81, 1130–1136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (122).Bereman MS; Young DD; Deiters A; Muddiman DC Development of a Robust and High Throughput Method for Profiling N-Linked Glycans Derived from Plasma Glycoproteins by NanoLC-FTICR Mass Spectrometry. J. Proteome Res 2009, 8, 3764–3770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (123).Lam MP; Siu SO; Lau E; Mao X; Sun HZ; Chiu PC; Yeung WS; Cox DM; Chu IK Online coupling of reverse-phase and hydrophilic interaction liquid chromatography for protein and glycoprotein characterization. Anal. Bioanal. Chem 2010, 398, 791–804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (124).Ahn J; Bones J; Yu YQ; Rudd PM; Gilar M Separation of 2-aminobenzamide labeled glycans using hydrophilic interaction chromatography columns packed with 1.7μm sorbent. J. Chromatogr. B: Anal. Technol. Biomed. Life Sci 2010, 878, 403–408. [DOI] [PubMed] [Google Scholar]
- (125).Saldova R; Asadi Shehni A; Haakensen VD; Steinfeld I; Hilliard M; Kifer I; Helland A; Yakhini Z; Borresen-Dale AL; Rudd PM Association of N-glycosylation with breast carcinoma and systemic features using high-resolution quantitative UPLC. J. Proteome Res 2014, 13, 2314–2327. [DOI] [PubMed] [Google Scholar]
- (126).Doherty M; Bones J; McLoughlin N; Telford JE; Harmon B; DeFelippis MR; Rudd PM An automated robotic platform for rapid profiling oligosaccharide analysis of monoclonal antibodies directly from cell culture. Anal. Biochem 2013, 442, 10–18. [DOI] [PubMed] [Google Scholar]
- (127).Zacharias LG; Hartmann AK; Song EH; Zhao JF; Zhu R; Mirzaei P; Mechref Y HILIC and ERLIC Enrichment of Glycopeptides Derived from Breast and Brain Cancer Cells. J. Proteome Res 2016, 15, 3624–3634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (128).Adamczyk B; Tharmalingam-Jaikaran T; Schomberg M; Szekrenyes A; Kelly RM; Karlsson NG; Guttman A; Rudd PM Comparison of separation techniques for the elucidation of IgG N-glycans pooled from healthy mammalian species. Carbohydr. Res 2014, 389, 174–185. [DOI] [PubMed] [Google Scholar]
- (129).Mauko L; Lacher NA; Pelzing M; Nordborg A; Haddad PR; Hilder EF Comparison of ZIC-HILIC and graphitized carbon-based analytical approaches combined with exoglycosidase digestions for analysis of glycans from monoclonal antibodies. J. Chromatogr. B: Anal. Technol. Biomed. Life Sci 2012, 911, 93–104. [DOI] [PubMed] [Google Scholar]
- (130).Palmisano G; Larsen MR; Packer NH; Thaysen-Andersen M Structural analysis of glycoprotein sialylation - part II: LC-MS based detection. RSC Adv. 2013, 3, 22706–22726. [Google Scholar]
- (131).Hua S; An HJ; Ozcan S; Ro GS; Soares S; DeVere-White R; Lebrilla CB Comprehensive native glycan profiling with isomer separation and quantitation for the discovery of cancer biomarkers. Analyst 2011, 136, 3663–3671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (132).Hua S; Williams CC; Dimapasoc LM; Ro GS; Ozcan S; Miyamoto S; Lebrilla CB; An HJ; Leiserowitz GS Isomer-specific chromatographic profiling yields highly sensitive and specific potential N-glycan biomarkers for epithelial ovarian cancer. J. Chromatogr. A 2013, 1279, 58–67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (133).Anugraham M; Jacob F; Nixdorf S; Everest-Dass AV; Heinzelmann-Schwarz V; Packer NH Specific glycosylation of membrane proteins in epithelial ovarian cancer cell lines: glycan structures reflect gene expression and DNA methylation status. Mol. Cell. Proteomics 2014, 13, 2213–2232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (134).Park D; Brune KA; Mitra A; Marusina AI; Maverakis E; Lebrilla CB Characteristic Changes in Cell Surface Glycosylation Accompany Intestinal Epithelial Cell (IEC) Differentiation: High Mannose Structures Dominate the Cell Surface Glycome of Undifferentiated Enterocytes. Mol. Cell. Proteomics 2015, 14, 2910–2921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (135).Nwosu CC; Aldredge DL; Lee H; Lerno LA; Zivkovic AM; German JB; Lebrilla CB Comparison of the Human and Bovine Milk N-Glycome via High-Performance Microfluidic Chip Liquid Chromatography and Tandem Mass Spectrometry. J. Proteome Res 2012, 11, 2912–2924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (136).Kronewitter SR; Marginean I; Cox JT; Zhao R; Hagler CD; Shukla AK; Carlson TS; Adkins JN; Camp DG II; Moore RJ; et al. Polysialylated N-glycans identified in human serum through combined developments in sample preparation, separations, and electrospray ionization-mass spectrometry. Anal. Chem 2014, 86, 8700–8710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (137).Mechref Y; Novotny MV Glycomic analysis by capillary electrophoresis-mass spectrometry. Mass Spectrom. Rev 2009, 28, 207–222. [DOI] [PubMed] [Google Scholar]
- (138).Guttman A; Kerekgyarto M; Jarvas G Effect of Separation Temperature on Structure Specific Glycan Migration in Capillary Electrophoresis. Anal. Chem 2015, 87, 11630–11634. [DOI] [PubMed] [Google Scholar]
- (139).Szigeti M; Guttman A High-Resolution Glycan Analysis by Temperature Gradient Capillary Electrophoresis. Anal. Chem 2017, 89, 2201–2204. [DOI] [PubMed] [Google Scholar]
- (140).Feng HT; Su M; Rifai FN; Li P; Li SF Parallel analysis and orthogonal identification of N-glycans with different capillary electrophoresis mechanisms. Anal. Chim. Acta 2017, 953, 79–86. [DOI] [PubMed] [Google Scholar]
- (141).Mechref Y Analysis of glycans derived from glycoconjugates by capillary electrophoresis-mass spectrometry. Electrophoresis 2011, 32, 3467–3481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (142).Lapainis T; Rubakhin SS; Sweedler JV Capillary electrophoresis with electrospray ionization mass spectrometric detection for single-cell metabolomics. Anal. Chem 2009, 81, 5858–5864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (143).Smith RD; Barinaga CJ; Udseth HR Improved Electrospray Ionization Interface for Capillary Zone Electrophoresis - Mass-Spectrometry. Anal. Chem 1988, 60, 1948–1952. [Google Scholar]
- (144).Olivares JA; Nguyen NT; Yonker CR; Smith RD Online Mass-Spectrometric Detection for Capillary Zone Electrophoresis. Anal. Chem 1987, 59, 1230–1232. [Google Scholar]
- (145).Chang YZ; Her GR Sheathless capillary electrophoresis/electrospray mass spectrometry using a carbon-coated fused silica capillary. Anal. Chem 2000, 72, 626–630. [DOI] [PubMed] [Google Scholar]
- (146).Maxwell EJ; Zhong X; Zhang H; van Zeijl N; Chen DD Decoupling CE and ESI for a more robust interface with MS. Electrophoresis 2010, 31, 1130–1137. [DOI] [PubMed] [Google Scholar]
- (147).Jayo RG; Thaysen-Andersen M; Lindenburg PW; Haselberg R; Hankemeier T; Ramautar R; Chen DD Simple capillary electrophoresis-mass spectrometry method for complex glycan analysis using a flow-through microvial interface. Anal. Chem 2014, 86, 6479–6486. [DOI] [PubMed] [Google Scholar]
- (148).Mellors JS; Gorbounov V; Ramsey RS; Ramsey JM Fully integrated glass microfluidic device for performing high-efficiency capillary electrophoresis and electrospray ionization mass spectrometry. Anal. Chem 2008, 80, 6881–6887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (149).Khatri K; Klein JA; Haserick JR; Leon DR; Costello CE; McComb ME; Zaia J Microfluidic Capillary Electrophoresis-Mass Spectrometry for Analysis of Monosaccharides, Oligosaccharides, and Glycopeptides. Anal. Chem 2017, 89, 6645–6655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (150).Gray CJ; Thomas B; Upton R; Migas LG; Eyers CE; Barran PE; Flitsch SL Applications of ion mobility mass spectrometry for high throughput, high resolution glycan analysis. Biochim. Biophys. Acta, Gen. Subj 2016, 1860, 1688–1709. [DOI] [PubMed] [Google Scholar]
- (151).Hofmann J; Pagel K Glycan Analysis by Ion Mobility-Mass Spectrometry. Angew. Chem., Int. Ed 2017, 56, 8342–8349. [DOI] [PubMed] [Google Scholar]
- (152).Clowers BH; Dwivedi P; Steiner WE; Hill HH Jr.; Bendiak B Separation of sodiated isobaric disaccharides and trisaccharides using electrospray ionization-atmospheric pressure ion mobility-time of flight mass spectrometry. J. Am. Soc. Mass Spectrom 2005, 16, 660–669. [DOI] [PubMed] [Google Scholar]
- (153).Gabryelski W; Froese KL Rapid and sensitive differentiation of anomers, linkage, and position isomers of disaccharides using High-Field Asymmetric Waveform Ion Mobility Spectrometry (FAIMS). J. Am. Soc. Mass Spectrom 2003, 14, 265–277. [DOI] [PubMed] [Google Scholar]
- (154).Lu H; Zhang Y; Yang P Advancements in mass spectrometry-based glycoproteomics and glycomics. Natl. Sci. Rev 2016, 3, 345–364. [Google Scholar]
- (155).Dwivedi P; Bendiak B; Clowers BH; Hill HH Jr Rapid resolution of carbohydrate isomers by electrospray ionization ambient pressure ion mobility spectrometry-time-of-flight mass spectrometry (ESI-APIMS-TOFMS). J. Am. Soc. Mass Spectrom 2007, 18, 1163–1175. [DOI] [PubMed] [Google Scholar]
- (156).Fenn LS; McLean JA Structural resolution of carbohydrate positional and structural isomers based on gas-phase ion mobility-mass spectrometry. Phys. Chem. Chem. Phys 2011, 13, 2196–2205. [DOI] [PubMed] [Google Scholar]
- (157).Isailovic D; Kurulugama RT; Plasencia MD; Stokes ST; Kyselova Z; Goldman R; Mechref Y; Novotny MV; Clemmer DE Profiling of human serum glycans associated with liver cancer and cirrhosis by IMS-MS. J. Proteome Res 2008, 7, 1109–1117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (158).Plasencia MD; Isailovic D; Merenbloom SI; Mechref Y; Clemmer DE Resolving and assigning N-linked glycan structural isomers from ovalbumin by IMS-MS. J. Am. Soc. Mass Spectrom 2008, 19, 1706–1715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (159).Zhu F; Lee S; Valentine SJ; Reilly JP; Clemmer DE Mannose7 glycan isomer characterization by IMS-MS/MS analysis. J. Am. Soc. Mass Spectrom 2012, 23, 2158–2166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (160).Pagel K; Harvey DJ Ion mobility-mass spectrometry of complex carbohydrates: collision cross sections of sodiated N-linked glycans. Anal. Chem 2013, 85, 5138–5145. [DOI] [PubMed] [Google Scholar]
- (161).Struwe WB; Pagel K; Benesch JLP; Harvey DJ; Campbell MP GlycoMob: an ion mobility-mass spectrometry collision cross section database for glycomics. Glycoconjugate J. 2016, 33, 399–404. [DOI] [PubMed] [Google Scholar]
- (162).Yamaguchi Y; Nishima W; Re S; Sugita Y Confident identification of isomeric N-glycan structures by combined ion mobility mass spectrometry and hydrophilic interaction liquid chromatography. Rapid Commun. Mass Spectrom 2012, 26, 2877–2884. [DOI] [PubMed] [Google Scholar]
- (163).Lareau NM; May JC; McLean JA Non-derivatized glycan analysis by reverse phase liquid chromatography and ion mobility-mass spectrometry. Analyst 2015, 140, 3335–3338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (164).Harvey DJ; Scarff CA; Edgeworth M; Crispin M; Scanlan CN; Sobott F; Allman S; Baruah K; Pritchard L; Scrivens JH Travelling wave ion mobility and negative ion fragmentation for the structural determination of N-linked glycans. Electrophoresis 2013, 34, 2368–2378. [DOI] [PubMed] [Google Scholar]
- (165).Gaye MM; Valentine SJ; Hu Y; Mirjankar N; Hammoud ZT; Mechref Y; Lavine BK; Clemmer DE Ion mobility-mass spectrometry analysis of serum N-linked glycans from esophageal adenocarcinoma phenotypes. J. Proteome Res 2012, 11, 6102–6110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (166).Seo Y; Schenauer MR; Leary JA Biologically Relevant Metal-Cation Binding Induces Conformational Changes in Heparin Oligosaccharides as Measured by Ion Mobility Mass Spectrometry. Int. J. Mass Spectrom 2011, 303, 191–198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (167).Zhu F; Glover MS; Shi H; Trinidad JC; Clemmer DE Populations of metal-glycan structures influence MS fragmentation patterns. J. Am. Soc. Mass Spectrom 2015, 26, 25–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (168).Huang Y; Dodds ED Ion mobility studies of carbohydrates as group I adducts: isomer specific collisional cross section dependence on metal ion radius. Anal. Chem 2013, 85, 9728–9735. [DOI] [PubMed] [Google Scholar]
- (169).Huang Y; Dodds ED Ion-neutral collisional cross sections of carbohydrate isomers as divalent cation adducts and their electron transfer products. Analyst 2015, 140, 6912–6921. [DOI] [PubMed] [Google Scholar]
- (170).Morrison KA; Bendiak BK; Clowers BH Enhanced Mixture Separations of Metal Adducted Tetrasaccharides Using Frequency Encoded Ion Mobility Separations and Tandem Mass Spectrometry. J. Am. Soc. Mass Spectrom 2017, 28, 664–677. [DOI] [PubMed] [Google Scholar]
- (171).Zheng X; Zhang X; Schocker NS; Renslow RS; Orton DJ; Khamsi J; Ashmus RA; Almeida IC; Tang K; Costello CE; et al. Enhancing glycan isomer separations with metal ions and positive and negative polarity ion mobility spectrometry-mass spectrometry analyses. Anal. Bioanal. Chem 2017, 409, 467–476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (172).Struwe WB; Benesch JL; Harvey DJ; Pagel K Collision cross sections of high-mannose N-glycans in commonly observed adduct states-identification of gas-phase conformers unique to [M-H] (−) ions. Analyst 2015, 140, 6799–6803. [DOI] [PubMed] [Google Scholar]
- (173).McDonnell LA; Heeren RM Imaging mass spectrometry. Mass Spectrom. Rev 2007, 26, 606–643. [DOI] [PubMed] [Google Scholar]
- (174).Cornett DS; Reyzer ML; Chaurand P; Caprioli RM MALDI imaging mass spectrometry: molecular snapshots of bio-chemical systems. Nat. Methods 2007, 4, 828–833. [DOI] [PubMed] [Google Scholar]
- (175).Powers TW; Jones EE; Betesh LR; Romano PR; Gao P; Copland JA; Mehta AS; Drake RR Matrix assisted laser desorption ionization imaging mass spectrometry workflow for spatial profiling analysis of N-linked glycan expression in tissues. Anal. Chem 2013, 85, 9799–9806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (176).Drake RR; Powers TW; Jones EE; Bruner E; Mehta AS; Angel PM MALDI Mass Spectrometry Imaging of N-Linked Glycans in Cancer Tissues. Adv. Cancer Res 2017, 134, 85–116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (177).Powers T; Holst S; Wuhrer M; Mehta A; Drake R Two-Dimensional N-Glycan Distribution Mapping of Hepatocellular Carcinoma Tissues by MALDI-Imaging Mass Spectrometry. Biomolecules 2015, 5, 2554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (178).Powers TW; Neely BA; Shao Y; Tang H; Troyer DA; Mehta AS; Haab BB; Drake RR MALDI Imaging Mass Spectrometry Profiling of N-Glycans in Formalin-Fixed Paraffin Embedded Clinical Tissue Blocks and Tissue Microarrays. PLoS One 2014, 9, e106255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (179).Holst S; Heijs B; de Haan N; van Zeijl RJM; Briaire-de Bruijn IH; van Pelt GW; Mehta AS; Angel PM; Mesker WE; Tollenaar RA; et al. Linkage-Specific in Situ Sialic Acid Derivatization for N-Glycan Mass Spectrometry Imaging of Formalin-Fixed Paraffin-Embedded Tissues. Anal. Chem 2016, 88, 5904–5913. [DOI] [PubMed] [Google Scholar]
- (180).Heijs B; Holst S; Briaire-de Bruijn IH; van Pelt GW; de Ru AH; van Veelen PA; Drake RR; Mehta AS; Mesker WE; Tollenaar RA; et al. Multimodal Mass Spectrometry Imaging of N-Glycans and Proteins from the Same Tissue Section. Anal. Chem 2016, 88, 7745–7753. [DOI] [PubMed] [Google Scholar]
- (181).Everest-Dass AV; Briggs MT; Kaur G; Oehler MK; Hoffmann P; Packer NH N-Glycan MALDI Imaging Mass Spectrometry on Formalin-Fixed Paraffin-Embedded Tissue Enables the Delineation of Ovarian Cancer Tissues. Mol. Cell. Proteomics 2016, 15, 3003–3016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (182).Nwosu C; Yau HK; Becht S Assignment of Core versus Antenna Fucosylation Types in Protein N-Glycosylation via Procainamide Labeling and Tandem Mass Spectrometry. Anal. Chem 2015, 87, 5905–5913. [DOI] [PubMed] [Google Scholar]
- (183).Zhang JH; Schubothe K; Li BS; Russell S; Lebrilla CB Infrared multiphoton dissociation of O-linked mucin-type oligosaccharides. Anal. Chem 2005, 77, 208–214. [DOI] [PubMed] [Google Scholar]
- (184).Lancaster KS; An HJ; Li BS; Lebrilla CB Interrogation of N-linked oligosaccharides using infrared multiphoton dissociation in FT-ICR mass spectrometry. Anal. Chem 2006, 78, 4990–4997. [DOI] [PubMed] [Google Scholar]
- (185).Williams JP; Pringle S; Richardson K; Gethings L; Vissers JP; De Cecco M; Houel S; Chakraborty AB; Yu YQ; Chen W; et al. Characterisation of glycoproteins using a quadrupole time-of-flight mass spectrometer configured for electron transfer dissociation. Rapid Commun. Mass Spectrom 2013, 27, 2383–2390. [DOI] [PubMed] [Google Scholar]
- (186).Huang Y; Yu X; Mao Y; Costello CE; Zaia J; Lin C De novo sequencing of heparan sulfate oligosaccharides by electron-activated dissociation. Anal. Chem 2013, 85, 11979–11986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (187).Zhou S; Dong X; Veillon L; Huang Y; Mechref Y LC-MS/MS analysis of permethylated N-glycans facilitating isomeric characterization. Anal. Bioanal. Chem 2017, 409, 453–466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (188).Wheeler SF; Harvey DJ Negative ion mass spectrometry of sialylated carbohydrates: Discrimination of N-acetylneuraminic acid linkages by MALDI-TOF and ESI-TOF mass spectrometry. Anal. Chem 2000, 72, 5027–5039. [DOI] [PubMed] [Google Scholar]
- (189).Michael C; Rizzi AM Tandem mass spectrometry of isomeric aniline-labeled N-glycans separated on porous graphitic carbon: Revealing the attachment position of terminal sialic acids and structures of neutral glycans. Rapid Commun. Mass Spectrom 2015, 29, 1268–1278. [DOI] [PubMed] [Google Scholar]
- (190).Harvey DJ; Mattu TS; Wormald MR; Royle L; Dwek RA; Rudd PM ″Internal residue loss″: Rearrangements occurring during the fragmentation of carbohydrates derivatized at the reducing terminus. Anal. Chem 2002, 74, 734–740. [DOI] [PubMed] [Google Scholar]
- (191).Fellenberg M; Behnken HN; Nagel T; Wiegandt A; Baerenfaenger M; Meyer B Glycan analysis: scope and limitations of different techniques-a case for integrated use of LC-MS(/MS) and NMR techniques. Anal. Bioanal. Chem 2013, 405, 7291–7305. [DOI] [PubMed] [Google Scholar]
- (192).Wiegandt A; Meyer B Unambiguous characterization of N-glycans of monoclonal antibody cetuximab by integration of LC-MS/MS and (1)H NMR spectroscopy. Anal. Chem 2014, 86, 4807–4814. [DOI] [PubMed] [Google Scholar]
- (193).Smith DF; Cummings RD Application of microarrays for deciphering the structure and function of the human glycome. Mol. Cell. Proteomics 2013, 12, 902–912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (194).Guttman A Multistructure sequencing of N-linked fetuin glycans by capillary gel electrophoresis and enzyme matrix digestion. Electrophoresis 1997, 18, 1136–1141. [DOI] [PubMed] [Google Scholar]
- (195).Guttman A; Ulfelder KW Exoglycosidase matrix-mediated sequencing of a complex glycan pool by capillary electrophoresis. J. Chromatogr. A 1997, 781, 547–554. [DOI] [PubMed] [Google Scholar]
- (196).Royle L; Radcliffe CM; Dwek RA; Rudd PM Detailed structural analysis of N-glycans released from glycoproteins in SDS-PAGE gel bands using HPLC combined with exoglycosidase array digestions. Methods Mol. Biol 2006, 347, 125–143. [DOI] [PubMed] [Google Scholar]
- (197).Takegawa Y; Deguchi K; Nakagawa H; Nishimura S Structural analysis of an N-glycan with ″beta1–4 bisecting branch″ from human serum IgG by negative-ion MSn spectral matching and exoglycosidase digestion. Anal. Chem 2005, 77, 6062–6068. [DOI] [PubMed] [Google Scholar]
- (198).Campbell MP; Royle L; Radcliffe CM; Dwek RA; Rudd PM GlycoBase and autoGU: tools for HPLC-based glycan analysis. Bioinformatics 2008, 24, 1214–1216. [DOI] [PubMed] [Google Scholar]
- (199).Campbell MP; Royle L; Rudd PM GlycoBase and autoGU: resources for interpreting HPLC-glycan data. Methods Mol. Biol 2015, 1273, 17–28. [DOI] [PubMed] [Google Scholar]
- (200).Aldredge D; An HJ; Tang N; Waddell K; Lebrilla CB Annotation of a serum N-glycan library for rapid identification of structures. J. Proteome Res 2012, 11, 1958–1968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (201).Abrahams JL; Campbell MP; Packer NH Building a PGC-LC-MS N-glycan retention library and elution mapping resource. Glycoconjugate J. 2017, 9793–4. [DOI] [PubMed] [Google Scholar]
- (202).Kolarich D; Rapp E; Struwe WB; Haslam SM; Zaia J; McBride R; Agravat S; Campbell MP; Kato M; Ranzinger R; et al. The minimum information required for a glycomics experiment (MIRAGE) project: improving the standards for reporting mass-spectrometry-based glycoanalytic data. Mol. Cell. Proteomics 2013, 12, 991–995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (203).York WS; Agravat S; Aoki-Kinoshita KF; McBride R; Campbell MP; Costello CE; Dell A; Feizi T; Haslam SM; Karlsson N; et al. MIRAGE: the minimum information required for a glycomics experiment. Glycobiology 2014, 24, 402–406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (204).Struwe WB; Agravat S; Aoki-Kinoshita KF; Campbell MP; Costello CE; Dell A; Feizi T; Haslam SM; Karlsson NG; Khoo KH; Kolarich D; Liu Y; McBride R; Novotny MV; Packer NH; Paulson JC; Rapp E; Ranzinger R; Rudd PM; Smith DF; Tiemeyer M; Wells L; York WS; Zaia J; Kettner C The minimum information required for a glycomics experiment (MIRAGE) project: sample preparation guidelines for reliable reporting of glycomics datasets. Glycobiology 2016, 26, 907–910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (205).Cummings RD; Pierce JM The challenge and promise of glycomics. Chem. Biol 2014, 21, 1–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (206).Zhang H; Yi EC; Li XJ; Mallick P; Kelly-Spratt KS; Masselon CD; Camp DG 2nd; Smith RD; Kemp CJ; Aebersold R High throughput quantitative analysis of serum proteins using glycopeptide capture and liquid chromatography mass spectrometry. Mol. Cell. Proteomics 2005, 4, 144–155. [DOI] [PubMed] [Google Scholar]
- (207).Ahn YH; Kim JY; Yoo JS Quantitative mass spectrometric analysis of glycoproteins combined with enrichment methods. Mass Spectrom. Rev 2015, 34, 148–165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (208).Ongay S; Boichenko A; Govorukhina N; Bischoff R Glycopeptide enrichment and separation for protein glycosylation analysis. J. Sep. Sci 2012, 35, 2341–2372. [DOI] [PubMed] [Google Scholar]
- (209).Chen CC; Su WC; Huang BY; Chen YJ; Tai HC; Obena RP Interaction modes and approaches to glycopeptide and glycoprotein enrichment. Analyst 2014, 139, 688–704. [DOI] [PubMed] [Google Scholar]
- (210).Thomas SN; Harlan R; Chen J; Aiyetan P; Liu Y; Sokoll LJ; Aebersold R; Chan DW; Zhang H Multiplexed Targeted Mass Spectrometry-Based Assays for the Quantification of N-Linked Glycosite-Containing Peptides in Serum. Anal. Chem 2015, 87, 10830–10838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (211).Li H; Popp R; Borchers CH Affinity-mass spectrometric technologies for quantitative proteomics in biological fluids. TrAC, Trends Anal. Chem 2017, 90, 80–88. [Google Scholar]
- (212).Gbormittah FO; Bones J; Hincapie M; Tousi F; Hancock WS; Iliopoulos O Clusterin Glycopeptide Variant Characterization Reveals Significant Site-Specific Glycan Changes in the Plasma of Clear Cell Renal Cell Carcinoma. J. Proteome Res 2015, 14, 2425–2436. [DOI] [PubMed] [Google Scholar]
- (213).van Scherpenzeel M; Steenbergen G; Morava E; Wevers RA; Lefeber DJ High-resolution mass spectrometry glycoprofiling of intact transferrin for diagnosis and subtype identification in the congenital disorders of glycosylation. Transl. Res 2015, 166, 639.e1–649.e1. [DOI] [PubMed] [Google Scholar]
- (214).Takahashi S; Sugiyama T; Shimomura M; Kamada Y; Fujita K; Nonomura N; Miyoshi E; Nakano M Site-specific and linkage analyses of fucosylated N-glycans on haptoglobin in sera of patients with various types of cancer: possible implication for the differential diagnosis of cancer. Glycoconjugate J. 2016, 33, 471–482. [DOI] [PubMed] [Google Scholar]
- (215).Kontro H; Joenvaara S; Haglund C; Renkonen R Comparison of sialylated N-glycopeptide levels in serum of pancreatic cancer patients, acute pancreatitis patients, and healthy controls. Proteomics 2014, 14, 1713–1723. [DOI] [PubMed] [Google Scholar]
- (216).Yin H; Tan Z; Wu J; Zhu J; Shedden KA; Marrero J; Lubman DM Mass-Selected Site-Specific Core-Fucosylation of Serum Proteins in Hepatocellular Carcinoma. J. Proteome Res 2015, 14, 4876–4884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (217).Tan Z; Yin H; Nie S; Lin Z; Zhu J; Ruffin MT; Anderson MA; Simeone DM; Lubman DM Large-scale identification of core-fucosylated glycopeptide sites in pancreatic cancer serum using mass spectrometry. J. Proteome Res 2015, 14, 1968–1978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (218).Drake PM; Schilling B; Niles RK; Braten M; Johansen E; Liu H; Lerch M; Sorensen DJ; Li B; Allen S; et al. A lectin affinity workflow targeting glycosite-specific, cancer-related carbohydrate structures in trypsin-digested human plasma. Anal. Biochem 2011, 408, 71–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (219).Zeng Z; Hincapie M; Pitteri SJ; Hanash S; Schalkwijk J; Hogan JM; Wang H; Hancock WS A Proteomics Platform Combining Depletion, Multi-lectin Affinity Chromatography (M-LAC), and Isoelectric Focusing to Study the Breast Cancer Proteome. Anal. Chem 2011, 83, 4845–4854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (220).Gbormittah FO; Lee LY; Taylor K; Hancock WS; Iliopoulos O Comparative Studies of the Proteome, Glycoproteome, and N-Glycome of Clear Cell Renal Cell Carcinoma Plasma before and after Curative Nephrectomy. J. Proteome Res 2014, 13, 4889–4900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (221).Totten SM; Kullolli M; Pitteri SJ Multi-Lectin Affinity Chromatography for Separation, Identification, and Quantitation of Intact Protein Glycoforms in Complex Biological Mixtures. Methods Mol. Biol 2017, 1550, 99–113. [DOI] [PubMed] [Google Scholar]
- (222).Zhang C; Ye Z; Xue P; Shu Q; Zhou Y; Ji Y; Fu Y; Wang J; Yang F Evaluation of Different N-Glycopeptide Enrichment Methods for N-Glycosylation Sites Mapping in Mouse Brain. J. Proteome Res 2016, 15, 2960–2968. [DOI] [PubMed] [Google Scholar]
- (223).Syed P; Gidwani K; Kekki H; Leivo J; Pettersson K; Lamminmaki U Role of lectin microarrays in cancer diagnosis. Proteomics 2016, 16, 1257–1265. [DOI] [PubMed] [Google Scholar]
- (224).Gray CJ; Sanchez-Ruiz A; Sardzikova I; Ahmed YA; Miller RL; Reyes Martinez JE; Pallister E; Huang K; Both P; Hartmann M; Roberts HN;Šardzík R; Mandal S; Turnbull JE; Eyers CE; Flitsch SL; et al. Label-Free Discovery Array Platform for the Characterization of Glycan Binding Proteins and Glycoproteins. Anal. Chem 2017, 89, 4444–4451. [DOI] [PubMed] [Google Scholar]
- (225).Beloqui A; Calvo J; Serna S; Yan S; Wilson IBH; Martin-Lomas M; Reichardt NC Analysis of Microarrays by MALDI-TOF MS. Angew. Chem., Int. Ed 2013, 52, 7477–7481. [DOI] [PubMed] [Google Scholar]
- (226).Dong L; Feng S; Li S; Song P; Wang J Preparation of Concanavalin A-Chelating Magnetic Nanoparticles for Selective Enrichment of Glycoproteins. Anal. Chem 2015, 87, 6849–6853. [DOI] [PubMed] [Google Scholar]
- (227).Li YG; Wen T; Zhu MZ; Li LX; Wei J; Wu XL; Guo MZ; Liu SP; Zhao HY; Xia SY; et al. Glycoproteomic analysis of tissues from patients with colon cancer using lectin microarrays and nanoLC-MS/MS. Mol. BioSyst 2013, 9, 1877–1887. [DOI] [PubMed] [Google Scholar]
- (228).Caragata M; Shah AK; Schulz BL; Hill MM; Punyadeera C Enrichment and identification of glycoproteins in human saliva using lectin magnetic bead arrays. Anal. Biochem 2016, 497, 76–82. [DOI] [PubMed] [Google Scholar]
- (229).Loo D; Jones A; Hill MM Lectin Magnetic Bead Array for Biomarker Discovery. J. Proteome Res 2010, 9, 5496–5500. [DOI] [PubMed] [Google Scholar]
- (230).Shah AK; Cao KAL; Choi E; Chen D; Gautier B; Nancarrow D; Whiteman DC; Saunders NA; Barbour AP; Joshi V; Hill MM; et al. Serum Glycoprotein Biomarker Discovery and Qualification Pipeline Reveals Novel Diagnostic Biomarker Candidates for Esophageal Adenocarcinoma. Mol. Cell. Proteomics 2015, 14, 3023–3039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (231).Kuo CW; Wu IL; Hsiao HH; Khoo KH Rapid glycopeptide enrichment and N-glycosylation site mapping strategies based on amine-functionalized magnetic nanoparticles. Anal. Bioanal. Chem 2012, 402, 2765–2776. [DOI] [PubMed] [Google Scholar]
- (232).Thaysen-Andersen M; Mysling S; Hojrup P Site-Specific Glycoprofiling of N-Linked Glycopeptides Using MALDI-TOF MS: Strong Correlation between Signal Strength and Glycoform Quantities. Anal. Chem 2009, 81, 3933–3943. [DOI] [PubMed] [Google Scholar]
- (233).Zhao Y; Yu L; Guo Z; Li X; Liang X Reversed-phase depletion coupled with hydrophilic affinity enrichment for the selective isolation of N-linked glycopeptides by using Click OEG-CD matrix. Anal. Bioanal. Chem 2011, 399, 3359–3365. [DOI] [PubMed] [Google Scholar]
- (234).Yu L; Li X; Guo Z; Zhang X; Liang X Hydrophilic interaction chromatography based enrichment of glycopeptides by using click maltose: a matrix with high selectivity and glycosylation heterogeneity coverage. Chem. - Eur. J 2009, 15, 12618–12626. [DOI] [PubMed] [Google Scholar]
- (235).Bi C; Zhao Y; Shen L; Zhang K; He X; Chen L; Zhang Y Click Synthesis of Hydrophilic Maltose-Functionalized Iron Oxide Magnetic Nanoparticles Based on Dopamine Anchors for Highly Selective Enrichment of Glycopeptides. ACS Appl. Mater. Interfaces 2015, 7, 24670–24678. [DOI] [PubMed] [Google Scholar]
- (236).Shen A; Guo Z; Yu L; Cao L; Liang X A novel zwitterionic HILIC stationary phase based on ″thiol-ene″ click chemistry between cysteine and vinyl silica. Chem. Commun. (Cambridge, U. K.) 2011, 47, 4550–4552. [DOI] [PubMed] [Google Scholar]
- (237).Wohlgemuth J; Karas M; Eichhorn T; Hendriks R; Andrecht S Quantitative site-specific analysis of protein glycosylation by LC-MS using different glycopeptide-enrichment strategies. Anal. Biochem 2009, 395, 178–188. [DOI] [PubMed] [Google Scholar]
- (238).Calvano CD; Zambonin CG; Jensen ON Assessment of lectin and HILIC based enrichment protocols for characterization of serum glycoproteins by mass spectrometry. J. Proteomics 2008, 71, 304–317. [DOI] [PubMed] [Google Scholar]
- (239).Mysling S; Palmisano G; Hojrup P; Thaysen-Andersen M Utilizing Ion-Pairing Hydrophilic Interaction Chromatography Solid Phase Extraction for Efficient Glycopeptide Enrichment in Glycoproteomics. Anal. Chem 2010, 82, 5598–5609. [DOI] [PubMed] [Google Scholar]
- (240).Snovida SI; Bodnar ED; Viner R; Saba J; Perreault H A simple cellulose column procedure for selective enrichment of glycopeptides and characterization by nano LC coupled with electron-transfer and high-energy collisional-dissociation tandem mass spectrometry. Carbohydr. Res 2010, 345, 792–801. [DOI] [PubMed] [Google Scholar]
- (241).Selman MH; Hemayatkar M; Deelder AM; Wuhrer M Cotton HILIC SPE microtips for microscale purification and enrichment of glycans and glycopeptides. Anal. Chem 2011, 83, 2492–2499. [DOI] [PubMed] [Google Scholar]
- (242).Zhang Y; Go EP; Desaire H Maximizing coverage of glycosylation heterogeneity in MALDI-MS analysis of glycoproteins with up to 27 glycosylation sites. Anal. Chem 2008, 80, 3144–3158. [DOI] [PubMed] [Google Scholar]
- (243).Wada Y; Tajiri M; Yoshida S Hydrophilic affinity isolation and MALDI multiple-stage tandem mass spectrometry of glycopeptides for glycoproteomics. Anal. Chem 2004, 76, 6560–6565. [DOI] [PubMed] [Google Scholar]
- (244).Zauner G; Koeleman CA; Deelder AM; Wuhrer M Protein glycosylation analysis by HILIC-LC-MS of Proteinase K-generated N- and O-glycopeptides. J. Sep. Sci 2010, 33, 903–910. [DOI] [PubMed] [Google Scholar]
- (245).Alpert AJ Electrostatic repulsion hydrophilic interaction chromatography for isocratic separation of charged solutes and selective isolation of phosphopeptides. Anal. Chem 2008, 80, 62–76. [DOI] [PubMed] [Google Scholar]
- (246).Cheow ESH; Sim KH; de Kleijn D; Lee CN; Sorokin V; Sze SK Simultaneous Enrichment of Plasma Soluble and Extracellular Vesicular Glycoproteins Using Prolonged Ultracentrifugation-Electrostatic Repulsion-hydrophilic Interaction Chromatography (PUC-ERLIC) Approach. Mol. Cell. Proteomics 2015, 14, 1657–1671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (247).Cao L; Yu L; Guo Z; Li X; Xue X; Liang X Application of a strong anion exchange material in electrostatic repulsion-hydrophilic interaction chromatography for selective enrichment of glycopeptides. J. Chromatogr. A 2013, 1299, 18–24. [DOI] [PubMed] [Google Scholar]
- (248).Zhang H; Guo T; Li X; Datta A; Park JE; Yang J; Lim SK; Tam JP; Sze SK Simultaneous characterization of glyco- and phosphoproteomes of mouse brain membrane proteome with electrostatic repulsion hydrophilic interaction chromatography. Mol. Cell. Proteomics 2010, 9, 635–647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (249).Fang C; Xiong Z; Qin H; Huang G; Liu J; Ye M; Feng S; Zou H One-pot synthesis of magnetic colloidal nanocrystal clusters coated with chitosan for selective enrichment of glycopeptides. Anal. Chim. Acta 2014, 841, 99–105. [DOI] [PubMed] [Google Scholar]
- (250).Jiang B; Liang Y; Wu Q; Jiang H; Yang K; Zhang L; Liang Z; Peng X; Zhang Y New GO-PEI-Au-L-Cys ZIC-HILIC composites: synthesis and selective enrichment of glycopeptides. Nanoscale 2014, 6, 5616–5619. [DOI] [PubMed] [Google Scholar]
- (251).Palmisano G; Lendal SE; Engholm-Keller K; Leth-Larsen R; Parker BL; Larsen MR Selective enrichment of sialic acid-containing glycopeptides using titanium dioxide chromatography with analysis by HILIC and mass spectrometry. Nat. Protoc 2010, 5, 1974–1982. [DOI] [PubMed] [Google Scholar]
- (252).Zhang Y; Giboulot A; Zivy M; Valot B; Jamet E; Albenne C Combining various strategies to increase the coverage of the plant cell wall glycoproteome. Phytochemistry 2011, 72, 1109–1123. [DOI] [PubMed] [Google Scholar]
- (253).Zhang Y; Kuang M; Zhang L; Yang P; Lu H An accessible protocol for solid-phase extraction of N-linked glycopeptides through reductive amination by amine-functionalized magnetic nanoparticles. Anal. Chem 2013, 85, 5535–5541. [DOI] [PubMed] [Google Scholar]
- (254).Xu YW; Wu ZX; Zhang LJ; Lu HJ; Yang PY; Webley PA; Zhao DY Highly Specific Enrichment of Glycopeptides Using Boronic Acid-Functionalized Mesoporous Silica. Anal. Chem 2009, 81, 503–508. [DOI] [PubMed] [Google Scholar]
- (255).Liu L; Zhang Y; Zhang L; Yan G; Yao J; Yang P; Lu H Highly specific revelation of rat serum glycopeptidome by boronic acid-functionalized mesoporous silica. Anal. Chim. Acta 2012, 753, 64–72. [DOI] [PubMed] [Google Scholar]
- (256).Zhou W; Yao N; Yao G; Deng C; Zhang X; Yang P Facile synthesis of aminophenylboronic acid-functionalized magnetic nanoparticles for selective separation of glycopeptides and glycoproteins. Chem. Commun. (Cambridge, U. K.) 2008, 5577–5579. [DOI] [PubMed] [Google Scholar]
- (257).Lin Z-A; Zheng J-N; Lin F; Zhang L; Cai Z; Chen G-N Synthesis of magnetic nanoparticles with immobilized aminophenylboronic acid for selective capture of glycoproteins. J. Mater. Chem 2011, 21, 518–524. [Google Scholar]
- (258).Ma R; Hu J; Cai Z; Ju H Facile synthesis of boronic acid-functionalized magnetic carbon nanotubes for highly specific enrichment of glycopeptides. Nanoscale 2014, 6, 3150–3156. [DOI] [PubMed] [Google Scholar]
- (259).Wang J; Wang Y; Gao M; Zhang X; Yang P Multilayer Hydrophilic Poly(phenol-formaldehyde resin)-Coated Magnetic Graphene for Boronic Acid Immobilization as a Novel Matrix for Glycoproteome Analysis. ACS Appl. Mater. Interfaces 2015, 7, 16011–16017. [DOI] [PubMed] [Google Scholar]
- (260).Shi Z; Pu L; Guo Y; Fu Z; Zhao W; Zhu Y; Wu J; Wang F Boronic Acid-Modified Magnetic Fe3O4@mTiO2Microspheres for Highly Sensitive and Selective Enrichment of N-Glycopeptides in Amniotic Fluid. Sci. Rep 2017, 7, 4603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (261).Chen WX; Smeekens JM; Wu RH A Universal Chemical Enrichment Method for Mapping the Yeast N-glycoproteome by Mass Spectrometry (MS). Mol. Cell. Proteomics 2014, 13, 1563–1572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (262).Smeekens JM; Xiao HP; Wu RH Global Analysis of Secreted Proteins and Glycoproteins in Saccharomyces cerevisiae. J. Proteome Res 2017, 16, 1039–1049. [DOI] [PubMed] [Google Scholar]
- (263).Wang H; Bie Z; Lü C; Liu Z Magnetic nanoparticles with dendrimer-assisted boronate avidity for the selective enrichment of trace glycoproteins. Chem. Sci 2013, 4 4298430310 1039/c3sc51623g [Google Scholar]
- (264).Hanson SR; Hsu TL; Weerapana E; Kishikawa K; Simon GM; Cravatt BF; Wong CH Tailored glycoproteomics and glycan site mapping using saccharide-selective bioorthogonal probes. J. Am. Chem. Soc 2007, 129, 7266–7267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (265).Yang L; Nyalwidhe JO; Guo S; Drake RR; Semmes OJ Targeted identification of metastasis-associated cell-surface sialoglycoproteins in prostate cancer. Mol. Cell. Proteomics 2011, 10, M110 007294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (266).Smeekens JM; Chen W; Wu R Mass spectrometric analysis of the cell surface N-glycoproteome by combining metabolic labeling and click chemistry. J. Am. Soc. Mass Spectrom 2015, 26, 604–614. [DOI] [PubMed] [Google Scholar]
- (267).Spiciarich DR; Nolley R; Maund SL; Purcell SC; Herschel J; Iavarone AT; Peehl DM; Bertozzi CR Bioorthogonal Labeling of Human Prostate Cancer Tissue Slice Cultures for Glycoproteomics. Angew. Chem., Int. Ed 2017, 56, 8992–8997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (268).Xiao H; Tang GX; Wu R Site-Specific Quantification of Surface N-Glycoproteins in Statin-Treated Liver Cells. Anal. Chem 2016, 88, 3324–3332. [DOI] [PubMed] [Google Scholar]
- (269).Tarasova IA; Masselon CD; Gorshkov AV; Gorshkov MV Predictive chromatography of peptides and proteins as a complementary tool for proteomics. Analyst 2016, 141, 4816–4832. [DOI] [PubMed] [Google Scholar]
- (270).Kozlik P; Goldman R; Sanda M Study of structure-dependent chromatographic behavior of glycopeptides using reversed phase nanoLC. Electrophoresis 2017, 38, 2193–2199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (271).Ozohanics O; Turiak L; Puerta A; Vekey K; Drahos L High-performance liquid chromatography coupled to mass spectrometry methodology for analyzing site-specific N-glycosylation patterns. J. Chromatogr. A 2012, 1259, 200–212. [DOI] [PubMed] [Google Scholar]
- (272).Zhao Y; Szeto SS; Kong RP; Law CH; Li G; Quan Q; Zhang Z; Wang Y; Chu IK Online two-dimensional porous graphitic carbon/reversed phase liquid chromatography platform applied to shotgun proteomics and glycoproteomics. Anal. Chem 2014, 86, 12172–12179. [DOI] [PubMed] [Google Scholar]
- (273).Wohlgemuth J; Karas M; Jiang W; Hendriks R; Andrecht S Enhanced glyco-profiling by specific glycopeptide enrichment and complementary monolithic nano-LC (ZIC-HILIC/RP18e)/ESI-MS analysis. J. Sep. Sci 2010, 33, 880–890. [DOI] [PubMed] [Google Scholar]
- (274).Stavenhagen K; Plomp R; Wuhrer M Site-Specific Protein N- and O-Glycosylation Analysis by a C18-Porous Graphitized Carbon-Liquid Chromatography-Electrospray Ionization Mass Spectrometry Approach Using Pronase Treated Glycopeptides. Anal. Chem 2015, 87, 11691–11699. [DOI] [PubMed] [Google Scholar]
- (275).An HJ; Peavy TR; Hedrick JL; Lebrilla CB Determination of N-glycosylation sites and site heterogeneity in glycoproteins. Anal. Chem 2003, 75, 5628–5637. [DOI] [PubMed] [Google Scholar]
- (276).Wuhrer M; Koeleman CAM; Hokke CH; Deelder AM Protein glycosylation analyzed by normal-phase nano-liquid chromatography-mass spectrometry of glycopeptides. Anal. Chem 2005, 77, 886–894. [DOI] [PubMed] [Google Scholar]
- (277).Hua S; Nwosu CC; Strum JS; Seipert RR; An HJ; Zivkovic AM; German JB; Lebrilla CB Site-specific protein glycosylation analysis with glycan isomer differentiation. Anal. Bioanal. Chem 2012, 403, 1291–1302. [DOI] [PubMed] [Google Scholar]
- (278).Strum JS; Nwosu CC; Hua S; Kronewitter SR; Seipert RR; Bachelor RJ; An HJ; Lebrilla CB Automated Assignments of N- and O-Site Specific Glycosylation with Extensive Glycan Heterogeneity of Glycoprotein Mixtures. Anal. Chem 2013, 85, 5666–5675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (279).Nwosu CC; Seipert RR; Strum JS; Hua SS; An HJ; Zivkovic AM; German BJ; Lebrilla CB Simultaneous and extensive site-specific N- and O-glycosylation analysis in protein mixtures. J. Proteome Res 2011, 10, 2612–2624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (280).Huang Y; Nie Y; Boyes B; Orlando R Resolving Isomeric Glycopeptide Glycoforms with Hydrophilic Interaction Chromatography (HILIC). J. Biomol. Technol 2016, 27, 98–104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (281).Mao X; Chu IK; Lin B A sheath-flow nanoelectrospray interface of microchip electrophoresis MS for glycoprotein and glycopeptide analysis. Electrophoresis 2006, 27, 5059–5067. [DOI] [PubMed] [Google Scholar]
- (282).Amon S; Plematl A; Rizzi A Capillary zone electrophoresis of glycopeptides under controlled electroosmotic flow conditions coupled to electrospray and matrix-assisted laser desorption/ionization mass spectrometry. Electrophoresis 2006, 27, 1209–1219. [DOI] [PubMed] [Google Scholar]
- (283).Gimenez E; Ramos-Hernan R; Benavente F; Barbosa J; Sanz-Nebot V Analysis of recombinant human erythropoietin glycopeptides by capillary electrophoresis electrospray-time of flight-mass spectrometry. Anal. Chim. Acta 2012, 709, 81–90. [DOI] [PubMed] [Google Scholar]
- (284).Barroso A; Gimenez E; Benavente F; Barbosa J; Sanz-Nebot V Analysis of human transferrin glycopeptides by capillary electrophoresis and capillary liquid chromatography-mass spectrometry. Application to diagnosis of alcohol dependence. Anal. Chim. Acta 2013, 804, 167–175. [DOI] [PubMed] [Google Scholar]
- (285).Mancera-Arteu M; Gimenez E; Benavente F; Barbosa J; Sanz-Nebot V Analysis of O-Glycopeptides by Acetone Enrichment and Capillary Electrophoresis-Mass Spectrometry. J. Proteome Res 2017, 16, 4166–4176. [DOI] [PubMed] [Google Scholar]
- (286).Barroso A; Gimenez E; Benavente F; Barbosa J; Sanz-Nebot V Modelling the electrophoretic migration behaviour of peptides and glycopeptides from glycoprotein digests in capillary electrophoresis-mass spectrometry. Anal. Chim. Acta 2015, 854, 169–177. [DOI] [PubMed] [Google Scholar]
- (287).Kammeijer GS; Kohler I; Jansen BC; Hensbergen PJ; Mayboroda OA; Falck D; Wuhrer M Dopant Enriched Nitrogen Gas Combined with Sheathless Capillary Electrophoresis-Electrospray Ionization-Mass Spectrometry for Improved Sensitivity and Repeatability in Glycopeptide Analysis. Anal. Chem 2016, 88, 5849–5856. [DOI] [PubMed] [Google Scholar]
- (288).Both P; Green AP; Gray CJ; Sardzik R; Voglmeir J; Fontana C; Austeri M; Rejzek M; Richardson D; Field RA; et al. Discrimination of epimeric glycans and glycopeptides using IM-MS and its potential for carbohydrate sequencing. Nat. Chem 2014, 6, 65–74. [DOI] [PubMed] [Google Scholar]
- (289).Creese AJ; Cooper HJ Separation and identification of isomeric glycopeptides by high field asymmetric waveform ion mobility spectrometry. Anal. Chem 2012, 84, 2597–2601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (290).Hinneburg H; Hofmann J; Struwe WB; Thader A; Altmann F; Varon Silva D; Seeberger PH; Pagel K; Kolarich D Distinguishing N-acetylneuraminic acid linkage isomers on glycopeptides by ion mobility-mass spectrometry. Chem. Commun. (Cambridge, U. K.) 2016, 52, 4381–4384. [DOI] [PubMed] [Google Scholar]
- (291).Sarbu M; Zhu F; Peter-Katalinic J; Clemmer DE; Zamfir AD Application of ion mobility tandem mass spectrometry to compositional and structural analysis of glycopeptides extracted from the urine of a patient diagnosed with Schindler disease. Rapid Commun. Mass Spectrom 2015, 29, 1929–1937. [DOI] [PubMed] [Google Scholar]
- (292).Zhu FF; Trinidad JC; Clemmer DE Glycopeptide Site Heterogeneity and Structural Diversity Determined by Combined Lectin Affinity Chromatography/IMS/CID/MS Techniques. J. Am. Soc. Mass Spectrom 2015, 26, 1092–1102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (293).Guttman M; Lee KK Site-Specific Mapping of Sialic Acid Linkage Isomers by Ion Mobility Spectrometry. Anal. Chem 2016, 88, 5212–5217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (294).Wuhrer M; Catalina MI; Deelder AM; Hokke CH Glycoproteomics based on tandem mass spectrometry of glycopeptides. J. Chromatogr. B: Anal. Technol. Biomed. Life Sci 2007, 849, 115–128. [DOI] [PubMed] [Google Scholar]
- (295).Imre T; Schlosser G; Pocsfalvi G; Siciliano R; Molnar-Szollosi E; Kremmer T; Malorni A; Vekey K Glycosylation site analysis of human alpha-1-acid glycoprotein (AGP) by capillary liquid chromatography-electrospray mass spectrometry. J. Mass Spectrom 2005, 40, 1472–1483. [DOI] [PubMed] [Google Scholar]
- (296).Bezouska K; Sklenar J; Novak P; Halada P; Havlicek V; Kraus M; Ticha M; Jonakova V Determination of the complete covalent structure of the major glycoform of DQH sperm surface protein, a novel trypsin-resistant boar seminal plasma O-glycoprotein related to pB1 protein. Protein Sci. 1999, 8, 1551–1556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (297).Larsen MR; Hojrup P; Roepstorff P Characterization of gel-separated glycoproteins using two-step proteolytic digestion combined with sequential microcolumns and mass spectrometry. Mol. Cell. Proteomics 2005, 4, 107–119. [DOI] [PubMed] [Google Scholar]
- (298).Froehlich JW; Barboza M; Chu C; Lerno LA Jr.; Clowers BH; Zivkovic AM; German JB; Lebrilla CB Nano-LC-MS/MS of glycopeptides produced by nonspecific proteolysis enables rapid and extensive site-specific glycosylation determination. Anal. Chem 2011, 83, 5541–5547. [DOI] [PubMed] [Google Scholar]
- (299).Zhang Y; Xie X; Zhao X; Tian F; Lv J; Ying W; Qian X Systems analysis of singly and multiply O-glycosylated peptides in the human serum glycoproteome via EThcD and HCD mass spectrometry. J. Proteomics 2018, 170, 14–27. [DOI] [PubMed] [Google Scholar]
- (300).Kaji H; Saito H; Yamauchi Y; Shinkawa T; Taoka M; Hirabayashi J; Kasai K; Takahashi N; Isobe T Lectin affinity capture, isotope-coded tagging and mass spectrometry to identify N-linked glycoproteins. Nat. Biotechnol 2003, 21, 667–672. [DOI] [PubMed] [Google Scholar]
- (301).Tarentino AL; Plummer TH, Jr Enzymatic deglycosylation of asparagine-linked glycans: purification, properties, and specificity of oligosaccharide-cleaving enzymes from Flavobacterium meningosepticum. Methods Enzymol. 1994, 230, 44–57. [DOI] [PubMed] [Google Scholar]
- (302).Go EP; Hewawasam G; Liao HX; Chen H; Ping LH; Anderson JA; Hua DC; Haynes BF; Desaire H Characterization of glycosylation profiles of HIV-1 transmitted/founder envelopes by mass spectrometry. J. Virol 2011, 85, 8270–8284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (303).Hagglund P; Bunkenborg J; Elortza F; Jensen ON; Roepstorff P A new strategy for identification of N-glycosylated proteins and unambiguous assignment of their glycosylation sites using HILIC enrichment and partial deglycosylation. J. Proteome Res 2004, 3, 556–566. [DOI] [PubMed] [Google Scholar]
- (304).Chen W; Smeekens JM; Wu R Comprehensive analysis of protein N-glycosylation sites by combining chemical deglycosylation with LC-MS. J. Proteome Res 2014, 13, 1466–1473. [DOI] [PubMed] [Google Scholar]
- (305).Yang S; Chen LJ; Chan DW; Li QK; Zhang H Protein signatures of molecular pathways in non-small cell lung carcinoma (NSCLC): comparison of glycoproteomics and global proteomics. Clin. Proteomics 2017, 14, 31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (306).Al-wajeeh AS; Alsayrafi M; Harvey TM; Latiff ABA; Ismail SM; Salhimi SM; Saghir SAM; Khalid IA Identification of Glycobiomarker Candidates for Breast Cancer Using LTQ-Orbitrap Fusion Technique. Int. J. Pharmacol 2017, 13, 425–437. [Google Scholar]
- (307).Sawanobori A; Moriwaki K; Takamatsu S; Kamada Y; Miyoshi E A glycoproteomic approach to identify novel glycomarkers for cancer stem cells. Proteomics 2016, 16, 3073–3080. [DOI] [PubMed] [Google Scholar]
- (308).Frenkel-Pinter M; Shmueli MD; Raz C; Yanku M; Zilberzwige S; Gazit E; Segal D Interplay between protein glycosylation pathways in Alzheimer’s disease. Sci. Adv 2017, 3, e1601576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (309).Bai X; Li DY; Zhu J; Guan YD; Zhang QY; Chi LL From individual proteins to proteomic samples: characterization of O-glycosylation sites in human chorionic gonadotropin and human-plasma proteins. Anal. Bioanal. Chem 2015, 407, 1857–1869. [DOI] [PubMed] [Google Scholar]
- (310).DeCoux A; Tian Y; DeLeon-Pennell KY; Nguyen NT; de Castro Bras LE; Flynn ER; Cannon PL; Griswold ME; Jin YF; Puskarich MA; Jones AE; Lindsey ML; et al. Plasma Glycoproteomics Reveals Sepsis Outcomes Linked to Distinct Proteins in Common Pathways. Crit. Care Med 2015, 43, 2049–2058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (311).Huang JQ; Kast J Quantitative Glycoproteomic Analysis Identifies Platelet-Induced Increase of Monocyte Adhesion via the Up-Regulation of Very Late Antigen 5. J. Proteome Res 2015, 14, 3015–3026. [DOI] [PubMed] [Google Scholar]
- (312).Jia XW; Chen J; Sun SS; Yang WM; Yang S; Shah P; Hoti N; Veltri B; Zhang H Detection of aggressive prostate cancer associated glycoproteins in urine using glycoproteomics and mass spectrometry. Proteomics 2016, 16, 2989–2996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (313).Norton P; Comunale MA; Herrera H; Wang MJ; Houser J; Wimmerova M; Romano PR; Mehta A Development and application of a novel recombinant Aleuria aurantia lectin with enhanced core fucose binding for identification of glycoprotein biomarkers of hepatocellular carcinoma. Proteomics 2016, 16, 3126–3136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (314).Woo CM; Felix A; Byrd WE; Zuegel DK; Ishihara M; Azadi P; Iavarone AT; Pitteri SJ; Bertozzi CR Development of IsoTaG, a Chemical Glycoproteomics Technique for Profiling Intact N- and O-Glycopeptides from Whole Cell Proteomes. J. Proteome Res 2017, 16, 1706–1718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (315).Yu ZX; Zhao XY; Tian F; Zhao Y; Zhang Y; Huang Y; Qian XH; Ying WT Sequential fragment ion filtering and endoglycosidase-assisted identification of intact glycopeptides. Anal. Bioanal. Chem 2017, 409, 3077–3087. [DOI] [PubMed] [Google Scholar]
- (316).Noro E; Togayachi A; Sato T; Tomioka A; Fujita M; Sukegawa M; Suzuki N; Kaji H; Narimatsu H Large-Scale Identification of N-Glycan Glycoproteins Carrying Lewis x and Site-Specific N-Glycan Alterations in Fut9 Knockout Mice. J. Proteome Res 2015, 14, 3823–3834. [DOI] [PubMed] [Google Scholar]
- (317).Wang JX; Cunningham R; Zetterberg H; Asthana S; Carlsson C; Okonkwo O; Li LJ Label-free quantitative comparison of cerebrospinal fluid glycoproteins and endogenous peptides in subjects with Alzheimer’s disease, mild cognitive impairment, and healthy individuals. Proteomics: Clin. Appl 2016, 10, 1225–1241. [DOI] [PubMed] [Google Scholar]
- (318).Yang WM; Jackson B; Zhang H Identification of glycoproteins associated with HIV latently infected cells using quantitative glycoproteomics. Proteomics 2016, 16, 1872–1880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (319).Zacchi LF; Schulz BL SWATH-MS Glycoproteomics Reveals Consequences of Defects in the Glycosylation Machinery. Mol. Cell. Proteomics 2016, 15, 2435–2447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (320).Fang P; Wang XJ; Xue Y; Liu MQ; Zeng WF; Zhang Y; Zhang L; Gao X; Yan GQ; Yao J; Shen H-L; Yang P-Y In-depth mapping of the mouse brain N-glycoproteome reveals widespread N-glycosylation of diverse brain proteins. Oncotarget 2016, 7, 38796–38809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (321).Goyallon A; Cholet S; Chapelle M; Junot C; Fenaille F Evaluation of a combined glycomics and glycoproteomics approach for studying the major glycoproteins present in biofluids: Application to cerebrospinal fluid. Rapid Commun. Mass Spectrom 2015, 29, 461–473. [DOI] [PubMed] [Google Scholar]
- (322).Stadlmann J; Taubenschmid J; Wenzel D; Gattinger A; Durnberger G; Dusberger F; Elling U; Mach L; Mechtler K; Penninger JM Comparative glycoproteomics of stem cells identifies new players in ricin toxicity. Nature 2017, 549, 538–542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (323).Choi NY; Hwang H; Ji ES; Park GW; Lee JY; Lee HK; Kim JY; Yoo JS Direct analysis of site-specific N-glycopeptides of serological proteins in dried blood spot samples. Anal. Bioanal. Chem 2017, 409, 4971–4981. [DOI] [PubMed] [Google Scholar]
- (324).Ruhaak LR; Miyamoto S; Kelly K; Lebrilla CB N-Glycan Profiling of Dried Blood Spots. Anal. Chem 2012, 84, 396–402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (325).Ji YL; Wei SH; Hou JJ; Zhang CQ; Xue P; Wang JF; Chen XL; Guo XJ; Yang FQ Integrated proteomic and N-glycoproteomic analyses of doxorubicin sensitive and resistant ovarian cancer cells reveal glycoprotein alteration in protein abundance and glycosylation. Oncotarget 2017, 8, 13413–13427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (326).Li QK; Shah P; Tian Y; Hu YW; Roden RBS; Zhang H; Chan DW An integrated proteomic and glycoproteomic approach uncovers differences in glycosylation occupancy from benign and malignant epithelial ovarian tumors. Clin. Proteomics 2017, 14, 16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (327).Liu TH; Liu DH; Liu RQ; Jiang HC; Yan GQ; Li W; Sun L; Zhang S; Liu YK; Guo K Discovering potential serological biomarker for chronic Hepatitis B Virus-related hepatocellular carcinoma in Chinese population by MAL-associated serum glycoproteomics analysis. Sci. Rep 2017, 7, 38918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (328).Khatri K; Klein JA; Zaia J Use of an informed search space maximizes confidence of site-specific assignment of glycoprotein glycosylation. Anal. Bioanal. Chem 2017, 409, 607–618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (329).Hu H; Khatri K; Klein J; Leymarie N; Zaia J A review of methods for interpretation of glycopeptide tandem mass spectral data. Glycoconjugate J. 2016, 33, 285–296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (330).Kumozaki S; Sato K; Sakakibara Y A Machine Learning Based Approach to de novo Sequencing of Glycans from Tandem Mass Spectrometry Spectrum. IEEE/ACM Trans. Comput. Biol. Bioinf 2015, 12, 1267–1274. [DOI] [PubMed] [Google Scholar]
- (331).Bern M; Kil YJ; Becker C Byonic: advanced peptide and protein identification software. Curr. Protoc. Bioinformatics 2012, 13.20.11–13.20.14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (332).Liu MQ; Zeng WF; Fang P; Cao WQ; Liu C; Yan GQ; Zhang Y; Peng C; Wu JQ; Zhang XJ; Tu HJ; Chi H; Sun RX; Cao Y; Dong MQ; Jiang BY; Huang JM; Shen HL; Wong CCL; He SM; Yang PY pGlyco 2.0 enables precision N-glycoproteomics with comprehensive quality control and one-step mass spectrometry for intact glycopeptide identification. Nat. Commun 2017, 8, 438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (333).Zeng W-F; Liu M-Q; Zhang Y; Wu J-Q; Fang P; Peng C; Nie A; Yan G; Cao W; Liu C; Chi H; Sun RX; Wong CCL; He SM; Yang P pGlyco: a pipeline for the identification of intact N-glycopeptides by using HCD-and CID-MS/MS and MS3. Sci. Rep 2016, 6, 25102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (334).He L; Xin L; Shan BZ; Lajoie GA; Ma B GlycoMaster DB: Software To Assist the Automated Identification of N-Linked Glycopeptides by Tandem Mass Spectrometry. J. Proteome Res 2014, 13, 3881–3895. [DOI] [PubMed] [Google Scholar]
- (335).Nasir W; Toledo AG; Noborn F; Nilsson J; Wang MX; Bandeira N; Larson G SweetNET: A Bioinformatics Workflow for Glycopeptide MS/MS Spectral Analysis. J. Proteome Res 2016, 15, 2826–2840. [DOI] [PubMed] [Google Scholar]
- (336).Kim JW; Hwang H; Lim JS; Lee HJ; Jeong SK; Yoo JS; Paik YK gFinder: A Web-Based Bioinformatics Tool for the Analysis of N-Glycopeptides. J. Proteome Res 2016, 15, 4116–4125. [DOI] [PubMed] [Google Scholar]
- (337).Park GW; Kim JY; Hwang H; Lee JY; Ahn YH; Lee HK; Ji ES; Kim KH; Jeong HK; Yun KN; Kim YS; Ko JH; An HJ; Kim JH; Paik YK; Yoo JS Integrated GlycoProteome Analyzer (I-GPA) for Automated Identification and Quantitation of Site-Specific N-Glycosylation. Sci. Rep 2016, 6, 21175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (338).Aoki-Kinoshita K; Agravat S; Aoki NP; Arpinar S; Cummings RD; Fujita A; Fujita N; Hart GM; Haslam SM; Kawasaki T; et al. GlyTouCan 1.0-The international glycan structure repository. Nucleic Acids Res. 2016, 44, D1237–D1242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (339).Egorova KS; Kalinchuk NA; Knirel YA; Toukach PV Carbohydrate Structure Database (CSDB): new features. Russ. Chem. Bull 2015, 64, 1205–1210. [Google Scholar]
- (340).Akune Y; Lin CH; Abrahams JL; Zhang JY; Packer NH; Aoki-Kinoshita KF; Campbell MP Comprehensive analysis of the N-glycan biosynthetic pathway using bioinformatics to generate UniCorn: A theoretical N-glycan structure database. Carbohydr. Res 2016, 431, 56–63. [DOI] [PubMed] [Google Scholar]
- (341).Mariethoz J; Khatib K; Alocci D; Campbell MP; Karlsson NG; Packer NH; Mullen EH; Lisacek F SugarBindDB, a resource of glycan-mediated host-pathogen interactions. Nucleic Acids Res. 2016, 44, D1243–D1250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (342).Baycin Hizal DB; Wolozny D; Colao J; Jacobson E; Tian Y; Krag SS; Betenbaugh MJ; Zhang H Glycoproteomic and glycomic databases. Clin. Proteomics 2014, 11, 15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (343).Zubarev RA; Makarov A Orbitrap mass spectrometry. Anal. Chem 2013, 85, 5288–5296. [DOI] [PubMed] [Google Scholar]
- (344).Marshall AG; Hendrickson CL; Jackson GS Fourier transform ion cyclotron resonance mass spectrometry: A primer. Mass Spectrom. Rev 1998, 17, 1–35. [DOI] [PubMed] [Google Scholar]
- (345).Bogdanov B; Smith RD Proteomics by FTICR mass spectrometry: top down and bottom up. Mass Spectrom. Rev 2005, 24, 168–200. [DOI] [PubMed] [Google Scholar]
- (346).Toby TK; Fornelli L; Kelleher NL Progress in Top-Down Proteomics and the Analysis of Proteoforms. Annu. Rev. Anal. Chem 2016, 9, 499–519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (347).Smith LM; Kelleher NL Consortium for Top Down, P. Proteoform: a single term describing protein complexity. Nat. Methods 2013, 10, 186–187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (348).Hood L Systems biology and p4 medicine: past, present, and future. Rambam Maimonides Med. J 2013, 4, e0012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (349).Jacobs JF; Wevers RA; Lefeber DJ; van Scherpenzeel M Fast, robust and high-resolution glycosylation profiling of intact monoclonal IgG antibodies using nanoLC-chip-QTOF. Clin. Chim. Acta 2016, 461, 90–97. [DOI] [PubMed] [Google Scholar]
- (350).Zhang Z; Pan H; Chen X Mass spectrometry for structural characterization of therapeutic antibodies. Mass Spectrom. Rev 2009, 28, 147–176. [DOI] [PubMed] [Google Scholar]
- (351).Huhn C; Selman MH; Ruhaak LR; Deelder AM; Wuhrer M IgG glycosylation analysis. Proteomics 2009, 9, 882–913. [DOI] [PubMed] [Google Scholar]
- (352).Yang Y; Wang G; Song T; Lebrilla CB; Heck AJR Resolving the micro-heterogeneity and structural integrity of monoclonal antibodies by hybrid mass spectrometric approaches. MAbs 2017, 9, 638–645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (353).Shaw JB; Li W; Holden DD; Zhang Y; Griep-Raming J; Fellers RT; Early BP; Thomas PM; Kelleher NL; Brodbelt JS Complete protein characterization using top-down mass spectrometry and ultraviolet photodissociation. J. Am. Chem. Soc 2013, 135, 12646–12651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (354).Good DM; Wirtala M; McAlister GC; Coon JJ Performance characteristics of electron transfer dissociation mass spectrometry. Mol. Cell. Proteomics 2007, 6, 1942–1951. [DOI] [PubMed] [Google Scholar]
- (355).Swaney DL; McAlister GC; Wirtala M; Schwartz JC; Syka JE; Coon JJ Supplemental activation method for high-efficiency electron-transfer dissociation of doubly protonated peptide precursors. Anal. Chem 2007, 79, 477–485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (356).Frese CK; Altelaar AF; van den Toorn H; Nolting D; Griep-Raming J; Heck AJ; Mohammed S Toward full peptide sequence coverage by dual fragmentation combining electron-transfer and higher-energy collision dissociation tandem mass spectrometry. Anal. Chem 2012, 84, 9668–9673. [DOI] [PubMed] [Google Scholar]
- (357).Brunner AM; Lossl P; Liu F; Huguet R; Mullen C; Yamashita M; Zabrouskov V; Makarov A; Altelaar AF; Heck AJ Benchmarking multiple fragmentation methods on an orbitrap fusion for top-down phospho-proteoform characterization. Anal. Chem 2015, 87, 4152–4158. [DOI] [PubMed] [Google Scholar]
- (358).Yu Q; Wang B; Chen Z; Urabe G; Glover MS; Shi X; Guo LW; Kent KC; Li L Electron-Transfer/Higher-Energy Collision Dissociation (EThcD)-Enabled Intact Glycopeptide/Glycoproteome Characterization. J. Am. Soc. Mass Spectrom 2017, 28, 1751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (359).Takahashi K; Wall SB; Suzuki H; Smith A. D. t.; Hall S; Poulsen K; Kilian M; Mobley JA; Julian BA; Mestecky J; et al. Clustered O-glycans of IgA1: defining macro- and microheterogeneity by use of electron capture/transfer dissociation. Mol. Cell. Proteomics 2010, 9, 2545–2557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (360).Riley NM; Westphall MS; Coon JJ Activated Ion Electron Transfer Dissociation for Improved Fragmentation of Intact Proteins. Anal. Chem 2015, 87, 7109–7116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (361).Riley NM; Westphall MS; Hebert AS; Coon JJ Implementation of Activated Ion Electron Transfer Dissociation on a Quadrupole-Orbitrap-Linear Ion Trap Hybrid Mass Spectrometer. Anal. Chem 2017, 89, 6358–6366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (362).Riley NM; Westphall MS; Coon JJ Activated Ion-Electron Transfer Dissociation Enables Comprehensive Top-Down Protein Fragmentation. J. Proteome Res 2017, 16, 2653–2659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (363).Riley NM; Westphall MS; Coon JJ Sequencing Larger Intact Proteins (30–70 kDa) with Activated Ion Electron Transfer Dissociation. J. Am. Soc. Mass Spectrom 2018, 29, 140–149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (364).Nicolardi S; van der Burgt YE; Dragan I; Hensbergen PJ; Deelder AM Identification of new apolipoprotein-CIII glycoforms with ultrahigh resolution MALDI-FTICR mass spectrometry of human sera. J. Proteome Res 2013, 12, 2260–2268. [DOI] [PubMed] [Google Scholar]
- (365).Balaguer E; Demelbauer U; Pelzing M; Sanz-Nebot V; Barbosa J; Neususs C Glycoform characterization of erythropoietin combining glycan and intact protein analysis by capillary electrophoresis - electrospray - time-of-flight mass spectrometry. Electrophoresis 2006, 27, 2638–2650. [DOI] [PubMed] [Google Scholar]
- (366).Sanz-Nebot V; Balaguer E; Benavente F; Neususs C; Barbosa J Characterization of transferrin glycoforms in human serum by CE-UV and CE-ESI-MS. Electrophoresis 2007, 28, 1949–1957. [DOI] [PubMed] [Google Scholar]
- (367).Jooss K; Huhner J; Kiessig S; Moritz B; Neususs C Two-dimensional capillary zone electrophoresis-mass spectrometry for the characterization of intact monoclonal antibody charge variants, including deamidation products. Anal. Bioanal. Chem 2017, 409, 6057–6067. [DOI] [PubMed] [Google Scholar]
- (368).The PSA position. Nature 2011, 478, 286, 10.1038/478286a [DOI] [PubMed] [Google Scholar]
- (369).Yoneyama T; Ohyama C; Hatakeyama S; Narita S; Habuchi T; Koie T; Mori K; Hidari KI; Yamaguchi M; Suzuki T; et al. Measurement of aberrant glycosylation of prostate specific antigen can improve specificity in early detection of prostate cancer. Biochem. Biophys. Res. Commun 2014, 448, 390–396. [DOI] [PubMed] [Google Scholar]
- (370).Leymarie N; Griffin PJ; Jonscher K; Kolarich D; Orlando R; McComb M; Zaia J; Aguilan J; Alley WR; Altmann F; et al. Interlaboratory Study on Differential Analysis of Protein Glycosylation by Mass Spectrometry: The ABRF Glycoprotein Research Multi-Institutional Study 2012. Mol. Cell. Proteomics 2013, 12, 2935–2951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (371).Schumacher KN; Dodds ED A case for protein-level and site-level specificity in glycoproteomic studies of disease. Glycoconjugate J. 2016, 33, 377–385. [DOI] [PubMed] [Google Scholar]
- (372).Moh ES; Thaysen-Andersen M; Packer NH Relative versus absolute quantitation in disease glycomics. Proteomics: Clin. Appl 2015, 9, 368–382. [DOI] [PubMed] [Google Scholar]
- (373).Etxebarria J; Reichardt NC Methods for the absolute quantification of N-glycan biomarkers. Biochim. Biophys. Acta, Gen. Subj 2016, 1860, 1676–1687. [DOI] [PubMed] [Google Scholar]
- (374).Carr SA; Abbatiello SE; Ackermann BL; Borchers C; Domon B; Deutsch EW; Grant RP; Hoofnagle AN; Huttenhain R; Koomen JM; et al. Targeted peptide measurements in biology and medicine: best practices for mass spectrometry-based assay development using a fit-for-purpose approach. Mol. Cell. Proteomics 2014, 13, 907–917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (375).Ruhaak LR; Koeleman CA; Uh HW; Stam JC; van Heemst D; Maier AB; Houwing-Duistermaat JJ; Hensbergen PJ; Slagboom PE; Deelder AM; Wuhrer M; et al. Targeted biomarker discovery by high throughput glycosylation profiling of human plasma alpha1-antitrypsin and immunoglobulin a. PLoS One 2013, 8, e73082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (376).Ruhaak LR; Taylor SL; Stroble C; Nguyen UT; Parker EA; Song T; Lebrilla CB; Rom WN; Pass H; Kim K; Kelly K; Miyamoto S; et al. Differential N-Glycosylation Patterns in Lung Adenocarcinoma Tissue. J. Proteome Res 2015, 14, 4538–4549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (377).Reiding KR; Ruhaak LR; Uh HW; El Bouhaddani S; van den Akker EB; Plomp R; McDonnell LA; Houwing-Duistermaat JJ; Slagboom PE; Beekman M; et al. Human Plasma N-glycosylation as Analyzed by Matrix-Assisted Laser Desorption/Ionization-Fourier Transform Ion Cyclotron Resonance-MS Associates with Markers of Inflammation and Metabolic Health. Mol. Cell. Proteomics 2017, 16, 228–242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (378).Kyselova Z; Mechref Y; Kang P; Goetz JA; Dobrolecki LE; Sledge GW; Schnaper L; Hickey RJ; Malkas LH; Novotny MV Breast cancer diagnosis and prognosis through quantitative measurements of serum glycan profiles. Clin. Chem 2008, 54, 1166–1175. [DOI] [PubMed] [Google Scholar]
- (379).Kim K; Ruhaak LR; Nguyen UT; Taylor SL; Dimapasoc L; Williams C; Stroble C; Ozcan S; Miyamoto S; Lebrilla CB; et al. Evaluation of Glycomic Profiling as a Diagnostic Biomarker for Epithelial Ovarian Cancer. Cancer Epidemiol., Biomarkers Prev 2014, 23, 611–621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (380).Ruhaak LR; Miyamoto S; Lebrilla CB Developments in the Identification of Glycan Biomarkers for the Detection of Cancer. Mol. Cell. Proteomics 2013, 12, 846–855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (381).Borelli V; Vanhooren V; Lonardi E; Reiding KR; Capri M; Libert C; Garagnani P; Salvioli S; Franceschi C; Wuhrer M Plasma N-Glycome Signature of Down Syndrome. J. Proteome Res 2015, 14, 4232–4245. [DOI] [PubMed] [Google Scholar]
- (382).Veillon L; Fakih C; Abou-El-Hassan H; Kobeissy F; Mechref Y Glycosylation Changes in Brain Cancer. ACS Chem. Neurosci 2018, 9, 51–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (383).Hecht ES; Scholl EH; Walker SH; Taylor AD; Cliby WA; Motsinger-Reif AA; Muddiman DC Relative Quantification and Higher-Order Modeling of the Plasma Glycan Cancer Burden Ratio in Ovarian Cancer Case-Control Samples. J. Proteome Res 2015, 14, 4394–4401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (384).Zhou H; Warren PG; Froehlich JW; Lee RS Dual modifications strategy to quantify neutral and sialylated N-glycans simultaneously by MALDI-MS. Anal. Chem 2014, 86, 6277–6284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (385).Zhou S; Veillon L; Dong X; Huang Y; Mechref Y Direct comparison of derivatization strategies for LC-MS/MS analysis of N-glycans. Analyst 2017, 142, 4446–4455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (386).Alvarez-Manilla G; Warren NL; Abney T; Atwood J; Azadi P; York WS; Pierce M; Orlando R Tools for glycomics: relative quantitation of glycans by isotopic permethylation using (CH3I)-C-13. Glycobiology 2007, 17, 677–687. [DOI] [PubMed] [Google Scholar]
- (387).Kang P; Mechref Y; Kyselova Z; Goetz JA; Novotny MV Comparative glycomic mapping through quantitative permethylation and stable-isotope labeling. Anal. Chem 2007, 79, 6064–6073. [DOI] [PubMed] [Google Scholar]
- (388).Bowman MJ; Zaia J Tags for the stable isotopic labeling of carbohydrates and quantitative analysis by mass spectrometry. Anal. Chem 2007, 79, 5777–5784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (389).Prien JM; Prater BD; Qin Q; Cockrill SL Mass spectrometric-based stable isotopic 2-aminobenzoic acid glycan mapping for rapid glycan screening of biotherapeutics. Anal. Chem 2010, 82, 1498–1508. [DOI] [PubMed] [Google Scholar]
- (390).Xia B; Feasley CL; Sachdev GP; Smith DF; Cummings RD Glycan reductive isotope labeling for quantitative glycomics. Anal. Biochem 2009, 387, 162–170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (391).Mancera-Arteu M; Gimenez E; Barbosa J; Sanz-Nebot V Identification and characterization of isomeric N-glycans of human alfa-acid-glycoprotein by stable isotope labelling and ZIC-HILIC-MS in combination with exoglycosidase digestion. Anal. Chim. Acta 2016, 940, 92–103. [DOI] [PubMed] [Google Scholar]
- (392).Gimenez E; Sanz-Nebot V; Rizzi A Relative quantitation of glycosylation variants by stable isotope labeling of enzymatically released N-glycans using [12C]/[13C] aniline and ZIC-HILIC-ESI-TOF-MS. Anal. Bioanal. Chem 2013, 405, 7307–7319. [DOI] [PubMed] [Google Scholar]
- (393).Cao W; Zhang W; Huang J; Jiang B; Zhang L; Yang P Glycan reducing end dual isotopic labeling (GREDIL) for mass spectrometry-based quantitative N-glycomics. Chem. Commun. (Cambridge, U. K.) 2015, 51, 13603–13606. [DOI] [PubMed] [Google Scholar]
- (394).Zhang W; Cao W; Huang J; Wang H; Wang J; Xie C; Yang P PNGase F-mediated incorporation of 18O into glycans for relative glycan quantitation. Analyst 2015, 140, 1082–1089. [DOI] [PubMed] [Google Scholar]
- (395).Zhang W; Wang H; Tang H; Yang P Endoglycosidase-mediated incorporation of 18O into glycans for relative glycan quantitation. Anal. Chem 2011, 83, 4975–4981. [DOI] [PubMed] [Google Scholar]
- (396).Zhang P; Zhang Y; Xue X; Wang C; Wang Z; Huang L Relative quantitation of glycans using stable isotopic labels 1-(d0/d5) phenyl-3-methyl-5-pyrazolone by mass spectrometry. Anal. Biochem 2011, 418, 1–9. [DOI] [PubMed] [Google Scholar]
- (397).Hahne H; Neubert P; Kuhn K; Etienne C; Bomgarden R; Rogers JC; Kuster B Carbonyl-Reactive Tandem Mass Tags for the Proteome-Wide Quantification of N-Linked Glycans. Anal. Chem 2012, 84, 3716–3724. [DOI] [PubMed] [Google Scholar]
- (398).Yang S; Yuan W; Yang WM; Zhou JY; Harlan R; Edwards J; Li SW; Zhang H Glycan Analysis by Isobaric Aldehyde Reactive Tags and Mass Spectrometry. Anal. Chem 2013, 85, 8188–8195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (399).Yang S; Wang M; Chen L; Yin B; Song G; Turko IV; Phinney KW; Betenbaugh MJ; Zhang H; Li S QUANTITY: An Isobaric Tag for Quantitative Glycomics. Sci. Rep 2015, 5, 17585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (400).Echeverria B; Etxebarria J; Ruiz N; Hernandez A; Calvo J; Haberger M; Reusch D; Reichardt NC Chemo-Enzymatic Synthesis of (13)C Labeled Complex N-Glycans As Internal Standards for the Absolute Glycan Quantification by Mass Spectrometry. Anal. Chem 2015, 87, 11460–11467. [DOI] [PubMed] [Google Scholar]
- (401).Ji Y; Wei S; Hou J; Zhang C; Xue P; Wang J; Chen X; Guo X; Yang F Integrated proteomic and N-glycoproteomic analyses of doxorubicin sensitive and resistant ovarian cancer cells reveal glycoprotein alteration in protein abundance and glycosylation. Oncotarget 2017, 8, 13413–13427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (402).Zhang H; Li XJ; Martin DB; Aebersold R Identification and quantification of N-linked glycoproteins using hydrazide chemistry, stable isotope labeling and mass spectrometry. Nat. Biotechnol 2003, 21, 660–666. [DOI] [PubMed] [Google Scholar]
- (403).Surinova S; Choi M; Tao S; Schuffler PJ; Chang CY; Clough T; Vyslouzil K; Khoylou M; Srovnal J; Liu Y; et al. Prediction of colorectal cancer diagnosis based on circulating plasma proteins. EMBO Mol. Med 2015, 7, 1166–1178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (404).Xu Y; Bailey UM; Schulz BL Automated measurement of site-specific N-glycosylation occupancy with SWATH-MS. Proteomics 2015, 15, 2177–2186. [DOI] [PubMed] [Google Scholar]
- (405).Yeo KY; Chrysanthopoulos PK; Nouwens AS; Marcellin E; Schulz BL High-performance targeted mass spectrometry with precision data-independent acquisition reveals site-specific glycosylation macroheterogeneity. Anal. Biochem 2016, 510, 106–113. [DOI] [PubMed] [Google Scholar]
- (406).Hulsmeier AJ; Paesold-Burda P; Hennet T N-glycosylation site occupancy in serum glycoproteins using multiple reaction monitoring liquid chromatography-mass spectrometry. Mol. Cell. Proteomics 2007, 6, 2132–2138. [DOI] [PubMed] [Google Scholar]
- (407).Wright HT; Urry DW Nonenzymatic deamidation of asparaginyl and glutaminyl residues in proteins. Crit. Rev. Biochem. Mol. Biol 1991, 26, 1–52. [DOI] [PubMed] [Google Scholar]
- (408).Palmisano G; Melo-Braga MN; Engholm-Keller K; Parker BL; Larsen MR Chemical deamidation: a common pitfall in large-scale N-linked glycoproteomic mass spectrometry-based analyses. J. Proteome Res 2012, 11, 1949–1957. [DOI] [PubMed] [Google Scholar]
- (409).Zhu Z; Go EP; Desaire H Absolute quantitation of glycosylation site occupancy using isotopically labeled standards and LC-MS. J. Am. Soc. Mass Spectrom 2014, 25, 1012–1017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (410).Bongers J; Devincentis J; Fu J; Huang P; Kirkley DH; Leister K; Liu P; Ludwig R; Rumney K; Tao L; et al. Characterization of glycosylation sites for a recombinant IgG1 monoclonal antibody and a CTLA4-Ig fusion protein by liquid chromatography-mass spectrometry peptide mapping. J. Chromatogr. A 2011, 1218, 8140–8149. [DOI] [PubMed] [Google Scholar]
- (411).Cao L; Diedrich JK; Kulp DW; Pauthner M; He L; Park SR; Sok D; Su CY; Delahunty CM; Menis S; et al. Global site-specific N-glycosylation analysis of HIV envelope glycoprotein. Nat. Commun 2017, 8, 14954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (412).Selman MH; McDonnell LA; Palmblad M; Ruhaak LR; Deelder AM; Wuhrer M Immunoglobulin G glycopeptide profiling by matrix-assisted laser desorption ionization Fourier transform ion cyclotron resonance mass spectrometry. Anal. Chem 2010, 82, 1073–1081. [DOI] [PubMed] [Google Scholar]
- (413).Bondt A; Nicolardi S; Jansen BC; Stavenhagen K; Blank D; Kammeijer GS; Kozak RP; Fernandes DL; Hensbergen PJ; Hazes JM; van der Burgt JEM; Dolhain RJEM; Wuhrer M; et al. Longitudinal monitoring of immunoglobulin A glycosylation during pregnancy by simultaneous MALDI-FTICR-MS analysis of N- and O-glycopeptides. Sci. Rep 2016, 6, 27955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (414).Rebecchi KR; Wenke JL; Go EP; Desaire H Label-free quantitation: a new glycoproteomics approach. J. Am. Soc. Mass Spectrom 2009, 20, 1048–1059. [DOI] [PubMed] [Google Scholar]
- (415).Jiang J; Tian F; Cai Y; Qian X; Costello CE; Ying W Site-specific qualitative and quantitative analysis of the N- and O-glycoforms in recombinant human erythropoietin. Anal. Bioanal. Chem 2014, 406, 6265–6274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (416).Lee JY; Lee HK; Park GW; Hwang H; Jeong HK; Yun KN; Ji ES; Kim KH; Kim JS; Kim JW; et al. Characterization of Site-Specific N-Glycopeptide Isoforms of alpha-1-Acid Glycoprotein from an Interlaboratory Study Using LC-MS/MS. J. Proteome Res 2016, 15, 4146–4164. [DOI] [PubMed] [Google Scholar]
- (417).Zhu H; Qiu C; Ruth AC; Keire DA; Ye H A LC-MS All-in-One Workflow for Site-Specific Location, Identification and Quantification of N-/O-Glycosylation in Human Chorionic Gonadotropin Drug Products. AAPS J. 2017, 19, 846–855. [DOI] [PubMed] [Google Scholar]
- (418).Lange V; Picotti P; Domon B; Aebersold R Selected reaction monitoring for quantitative proteomics: a tutorial. Mol. Syst. Biol 2008, 4, 222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (419).Gallien S; Duriez E; Crone C; Kellmann M; Moehring T; Domon B Targeted proteomic quantification on quadrupole-orbitrap mass spectrometer. Mol. Cell. Proteomics 2012, 11, 1709–1723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (420).Desiderio DM; Kai M Preparation of stable isotope-incorporated peptide internal standards for field desorption mass spectrometry quantification of peptides in biologic tissue. Biomed. Mass Spectrom 1983, 10, 471–479. [DOI] [PubMed] [Google Scholar]
- (421).van den Broek I; Romijn FP; Nouta J; van der Laarse A; Drijfhout JW; Smit NP; van der Burgt YE; Cobbaert CM Automated Multiplex LC-MS/MS Assay for Quantifying Serum Apolipoproteins A-I, B, C-I, C-II, C-III, and E with Qualitative Apolipoprotein E Phenotyping. Clin. Chem 2016, 62, 188–197. [DOI] [PubMed] [Google Scholar]
- (422).Kushnir MM; Rockwood AL; Roberts WL; Abraham D; Hoofnagle AN; Meikle AW Measurement of thyroglobulin by liquid chromatography-tandem mass spectrometry in serum and plasma in the presence of antithyroglobulin autoantibodies. Clin. Chem 2013, 59, 982–990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (423).Goldman R; Sanda M Targeted methods for quantitative analysis of protein glycosylation. Proteomics: Clin. Appl 2015, 9, 17–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (424).Ruhaak LR; Lebrilla CB Applications of Multiple Reaction Monitoring to Clinical Glycomics. Chromatographia 2015, 78, 335–342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (425).Nilsson J Liquid chromatography-tandem mass spectrometry-based fragmentation analysis of glycopeptides. Glycoconjugate J. 2016, 33, 261–272. [DOI] [PubMed] [Google Scholar]
- (426).Ruhaak LR; Kim K; Stroble C; Taylor SL; Hong Q; Miyamoto S; Lebrilla CB; Leiserowitz G Protein-Specific Differential Glycosylation of Immunoglobulins in Serum of Ovarian Cancer Patients. J. Proteome Res 2016, 15, 1002–1010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (427).Miyamoto S; Stroble CD; Taylor S; Hong QT; Lebrilla CB; Leiserowitz GS; Kim K; Ruhaak LR Multiple Reaction Monitoring for the Quantitation of Serum Protein Glycosylation Profiles: Application to Ovarian Cancer. J. Proteome Res 2018, 17, 222–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (428).Ruhaak LR The Use of Multiple Reaction Monitoring on QQQ-MS for the Analysis of Protein- and Site-Specific Glycosylation Patterns in Serum. Methods Mol. Biol 2017, 1503, 63–82. [DOI] [PubMed] [Google Scholar]
- (429).Miyamoto S; Stroble CD; Taylor S; Hong Q; Lebrilla CB; Leiserowitz GS; Kim K; Ruhaak LR Multiple Reaction Monitoring for the quantitation of serum protein glycosylation profiles: Application to Ovarian Cancer. J. Proteome Res 2018, 17, 222–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (430).Huang J; Kailemia MJ; Goonatilleke E; Parker EA; Hong Q; Sabia R; Smilowitz JT; German JB; Lebrilla CB Quantitation of human milk proteins and their glycoforms using multiple reaction monitoring (MRM). Anal. Bioanal. Chem 2017, 409, 589–606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (431).Sanda M; Pompach P; Brnakova Z; Wu J; Makambi K; Goldman R Quantitative liquid chromatography-mass spectrometry-multiple reaction monitoring (LC-MS-MRM) analysis of site-specific glycoforms of haptoglobin in liver disease. Mol. Cell. Proteomics 2013, 12, 1294–1305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (432).Kurogochi M; Matsushista T; Amano M; Furukawa J; Shinohara Y; Aoshima M; Nishimura S Sialic acid-focused quantitative mouse serum glycoproteomics by multiple reaction monitoring assay. Mol. Cell. Proteomics 2010, 9, 2354–2368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (433).Srikanth J; Agalyadevi R; Babu P Targeted, Site-specific quantitation of N- and O-glycopeptides using 18O-labeling and product ion based mass spectrometry. Glycoconjugate J. 2017, 34, 95–105. [DOI] [PubMed] [Google Scholar]
- (434).Pan KT; Chen CC; Urlaub H; Khoo KH Adapting Data-Independent Acquisition for Mass Spectrometry-Based Protein Site-Specific N-Glycosylation Analysis. Anal. Chem 2017, 89, 4532–4539. [DOI] [PubMed] [Google Scholar]
- (435).Sanda M; Goldman R Data Independent Analysis of IgG Glycoforms in Samples of Unfractionated Human Plasma. Anal. Chem 2016, 88, 10118–10125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (436).Yuan W; Sanda M; Wu J; Koomen J; Goldman R Quantitative analysis of immunoglobulin subclasses and subclass specific glycosylation by LC-MS-MRM in liver disease. J. Proteomics 2015, 116, 24–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (437).Darebna P; Novak P; Kucera R; Topolcan O; Sanda M; Goldman R; Pompach P Changes in the expression of N- and O-glycopeptides in patients with colorectal cancer and hepatocellular carcinoma quantified by full-MS scan FT-ICR and multiple reaction monitoring. J. Proteomics 2017, 153, 44–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (438).Shuford CM; Walters JJ; Holland PM; Sreenivasan U; Askari N; Ray K; Grant RP Absolute Protein Quantification by Mass Spectrometry: Not as Simple as Advertised. Anal. Chem 2017, 89, 7406–7415. [DOI] [PubMed] [Google Scholar]
- (439).Deshpande N; Jensen PH; Packer NH; Kolarich D GlycoSpectrumScan: fishing glycopeptides from MS spectra of protease digests of human colostrum sIgA. J. Proteome Res 2010, 9, 1063–1075. [DOI] [PubMed] [Google Scholar]
- (440).Falck D; Jansen BC; Plomp R; Reusch D; Haberger M; Wuhrer M Glycoforms of Immunoglobulin G Based Biopharmaceuticals Are Differentially Cleaved by Trypsin Due to the Glycoform Influence on Higher-Order Structure. J. Proteome Res 2015, 14, 4019–4028. [DOI] [PubMed] [Google Scholar]
- (441).van der Post S; Subramani DB; Backstrom M; Johansson ME; Vester-Christensen MB; Mandel U; Bennett EP; Clausen H; Dahlen G; Sroka A; et al. Site-specific O-glycosylation on the MUC2 mucin protein inhibits cleavage by the Porphyromonas gingivalis secreted cysteine protease (RgpB). J. Biol. Chem 2013, 288, 14636–14646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (442).Nedelkov D Human proteoforms as new targets for clinical mass spectrometry protein tests. Expert Rev. Proteomics 2017, 14, 691–699. [DOI] [PubMed] [Google Scholar]
- (443).Trenchevska O; Schaab MR; Nelson RW; Nedelkov D Development of multiplex mass spectrometric immunoassay for detection and quantification of apolipoproteins C-I, C-II, C-III and their proteoforms. Methods 2015, 81, 86–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (444).Nelson RW; Krone JR; Bieber AL; Williams P Mass spectrometric immunoassay. Anal. Chem 1995, 67, 1153–1158. [DOI] [PubMed] [Google Scholar]
- (445).Wu SA; Grimm R; German JB; Lebrilla CB Annotation and Structural Analysis of Sialylated Human Milk Oligosaccharides. J. Proteome Res 2011, 10, 856–868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (446).Totten SM; Wu LD; Parker EA; Davis JCC; Hua S; Stroble C; Ruhaak LR; Smilowitz JT; German JB; Lebrilla CB Rapid-throughput glycomics applied to human milk oligosaccharide profiling for large human studies. Anal. Bioanal. Chem 2014, 406, 7925–7935. [DOI] [PubMed] [Google Scholar]
- (447).Charbonneau MR; O’Donnell D; Blanton LV; Totten SM; Davis JCC; Barratt MJ; Cheng JY; Guruge J; Talcott M; Bain JR; et al. Sialylated Milk Oligosaccharides Promote Microbiota-Dependent Growth in Models of Infant Undernutrition. Cell 2016, 164, 859–871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (448).Zhu HB; Qiu C; Ruth AC; Keire DA; Ye HP A LC-MS All-in-One Workflow for Site-Specific Location, Identification and Quantification of N-/O-Glycosylation in Human Chorionic Gonadotropin Drug Products. AAPS J. 2017, 19, 846–855. [DOI] [PubMed] [Google Scholar]
- (449).Maverakis E; Kim K; Shimoda M; Gershwin ME; Patel F; Wilken R; Raychaudhuri S; Ruhaak LR; Lebrilla CB Glycans in the immune system and The Altered Glycan Theory of Autoimmunity: A critical review. J. Autoimmun 2015, 57, 1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (450).Qin R; Zhao J; Qin W; Zhang Z; Zhao R; Han J; Yang Y; Li L; Wang X; Ren S; et al. Discovery of Non-invasive Glycan Biomarkers for Detection and Surveillance of Gastric Cancer. J. Cancer 2017, 8, 1908–1916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (451).Qin Y; Zhong Y; Ma T; Zhang J; Yang G; Guan F; Li Z; Li B A pilot study of salivary N-glycome in HBV-induced chronic hepatitis, cirrhosis, and hepatocellular carcinoma. Glycoconjugate J. 2017, 34, 523–535. [DOI] [PubMed] [Google Scholar]
- (452).Jeong HJ; Kim YG; Yang YH; Kim BG High-Throughput Quantitative Analysis of Total N-Glycans by Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry. Anal. Chem 2012, 84, 3453–3460. [DOI] [PubMed] [Google Scholar]
- (453).Pucic M; Knezevic A; Vidic J; Adamczyk B; Novokmet M; Polasek O; Gornik O; Supraha-Goreta S; Wormald MR; Redzic I; Campbell H; Wright A; Hastie ND; Wilson JF; Rudan I; Wuhrer M; Rudd PM; Josić D; Lauc G High Throughput Isolation and Glycosylation Analysis of IgG-Variability and Heritability of the IgG Glycome in Three Isolated Human Populations. Mol. Cell. Proteomics 2011, 10, 222–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (454).Ruhaak LR; Stroble C; Dai J; Barnett M; Taguchi A; Goodman GE; Miyamoto S; Gandara D; Feng Z; Lebrilla CB; et al. Serum Glycans as Risk Markers for Non-Small Cell Lung Cancer. Cancer Prev. Res 2016, 9, 317–323. [DOI] [PMC free article] [PubMed] [Google Scholar]