

Abstract
Plant food biofortification is recently receiving remarkable attention, as it aims to increase the intake of minerals, vitamins, or antioxidants, crucial for their contribution to the general human health status and disease prevention. In this context, the study of the plant’s secondary metabolites, such as polyphenols, plays a pivotal role for the development of a new generation of plant crops, compensating, at least in part, the low nutritional quality of Western diets with a higher quality of dietary sources. Due to the prevalent immunomodulatory activity at the intestinal level, polyphenols represent a nutritionally relevant class of plant secondary metabolites. In this review, we focus on the antioxidant and anti-inflammatory properties of different classes of polyphenols with a specific attention to their potential in the prevention of intestinal pathological processes. We also discuss the latest biotechnology strategies and new advances of genomic techniques as a helpful tool for polyphenols biofortification and the development of novel, healthy dietary alternatives that can contribute to the prevention of inflammatory bowel diseases.
Keywords: plant polyphenols, gut health, biotechnology, biofortification
1. The Polyphenols: Key Specialized Metabolites for Plants
Polyphenols are secondary metabolites, widespread in many fruits, vegetables and plant byproducts commonly consumed in the human diet. Examples of dietary sources rich in polyphenols include berries (i.e., blueberries, blackberries, black currants, etc.), apples, citrus species, grapes, coffee, tea, cocoa or vegetable crops, such as onions, carrots, artichokes, solanaceous species, and cruciferous plants (Table 1) [1]. Other less common under-utilized species rich in polyphenols include, for example, figs, white crowberry, glasswort, and gooseberries, which are recently attracting more attention as alternative dietary patterns enriched in healthy beneficial phytochemicals (Table 1).
Table 1.
Main polyphenol classes, content in dietary sources and detection methods. TCPA: total content of phenolic acids; TCF: total content of flavonoids; TCA, total content of anthocyanidins; TCS: total content of stilbenes; TPC: total polyphenol content.
| Polyphenol Class | Dietary Source and Content | Method of Determination | Reference |
|---|---|---|---|
|
Carrots (TCPA: 0.3–18.8 g·kg−1 DW; 0.5 g·kg−1 FW) |
Spectrophotometric (Folin-Ciocalteau)/ RP-HPLC DAD |
[13,14] |
| Artichokes (TCPA: 3.14–3.89 g·kg−1 DW) |
RP-HPLC DAD | [15,16] | |
| Coffee (TCPA: 5 g·kg−1 coffee pulp) |
RP-HPLC UV/VIS | [17] | |
| White crowberry (TPC: 16.140 g GAE·kg−1 DW) |
Spectrophotometric (Folin-Ciocalteau) |
[18] | |
| Glasswort (TPC: 290.5 mg GAE·kg−1 FW) |
Spectrophotometric (Folin-Ciocalteau) |
[19] | |
| Lettuce (TPC: 127–187 mg GAE·kg−1 FW) |
Spectrophotometric/HPLC-DAD- ESI/MSn |
[20] | |
|
Onions (flavonols: 81–93 mg·kg−1 FW) |
RP-HPLC PDA | [21] |
| Leek (flavonols: 160 mg·kg−1 FW) |
RP-HPLC PDA | [21] | |
| Tomato (flavonols: 68–105 mg·kg−1 FW) |
RP-HPLC PDA | [21] | |
| Broccoli (flavonols: 2 mg·kg−1 FW; 6.75 g·kg−1 FW) |
Spectrophotometric RP-HPLC UV |
[22,23] | |
| Kale (flavonols: 115 mg·kg−1 FW) |
RP-HPLC PDA | [21] | |
| Cauliflower (flavonols: 37.53 g·kg−1 FW) |
RP-HPLC UV | [23] | |
| Soybean (flavones: 9.4 mg·kg−1 FW) |
RP-HPLC PDA | [21] | |
| Soybean (isoflavones: 0.5–7.5 mg·kg−1 FW) |
RP-HPLC DAD | [24] | |
| Citrus fruit (flavanones: 98–4694 mg·kg−1 FW) |
RP-HPLC PDA | [25] | |
| Apricots (flavan-3-ols: 250 mg·kg−1 FW) |
RP-HPLC PDA | [21] | |
| Green Tea (brew) (TCF: 8.30 g QE·kg−1 DW; flavan-3-ols: 350–441 mg·100−1 mL; 12.81–43.65 g·kg−1 DW; TPC: 68.3–85.9 mg·100−1 mL) |
RP-HPLC DAD/ LC-ESI-Q-TOF-MS/ Spectrophotometric (Folin-Ciocalteau) |
[26,27,28] | |
| Black Tea (brew) (TCF: 7 g QE·kg−1 DW) |
RP-HPLC PDA/ Spectrophotometric |
[26] | |
| Apples (flavonols: 300–344 mg·kg−1 FW) |
RP-HPLC PDA | [21] | |
| Blueberries (flavonols: 172–327 mg·kg−1 FW) |
HPLC/ESI-MS | [29] | |
| Grapes (flavonols: 21–322 mg·kg−1 FW) |
HPLC/ESI-MS | [29] | |
| Cocoa (flavan-3-ols: 45–730 mg·kg−1 DW; (flavonols: 0.3–42 mg·kg−1 DW) |
LC-DAD, LC-MS | [30] | |
| Figs (flavan-3-ols: 29.6–130.2 g·kg−1 DW; (flavonols: 79.9–21.78 g·kg−1 DW) |
UPLC-PDA-FL | [31] | |
| Gooseberry (TCF: 3.04–6.99 g RE·kg−1 DW; 345.0–3726.5 mg·kg−1 DW) |
RP-HPLC DAD/ Spectrophotometric |
[32] | |
| Currant (TCF: 8.13–15.62 g RE·kg−1)c |
RP-HPLC DAD/ Spectrophotometric |
[32] | |
|
Red cabbage (TCA: 40–750 mg·kg−1 FW) |
HPLC-MS/MS | [33] |
| Black carrots (TCA: 14–177 mg·kg−1 FW) |
RP-HPLC PDA/HPLC/ESI-MS/ | [34] | |
| Blueberries (TCA: 1435–8227 mg·kg−1 FW) |
Spectrophotometric HPLC/ESI-MS |
[29] | |
| Blackberries (TCA: 2–4 g·kg−1 FW) |
HPLC/ESI-MS | [29,35] | |
| Grapes (TCA: 390–7900 mg·kg−1 FW) |
HPLC/ESI-MS | [29] | |
| Beetroot (TCA: 0.23–0.77 g Mv3G·kg−1 FW) |
Spectrophotometric (Folin-Ciocalteau) |
[36] | |
| Strawberries (TCA: 180 mg Mv3G·kg−1 FW) |
Spectrophotometric (Folin-Ciocalteau) |
[37] | |
| Figs (TCA: 4–1220 mg·kg−1 DW) |
UPLC-PDA-FL | [31] | |
| Gooseberry (TCA: 0.3–686.8 mg COG·kg−1 DW) |
RP-HPLC DAD/ Spectrophotometric |
[32] | |
| Currant (TCA: 603.4–1407.1 mg COG·kg−1 DW) |
RP-HPLC DAD/ Spectrophotometric |
[32] | |
|
Grapes (TCS: 0.1–1.7 g·kg−1 FW) |
RP-HPLC UV | [38] |
| Red currant (TCS: 15 mg·kg−1 FW) |
Capillary electrophoresis | [39] | |
| Blueberries (TCS: 4 mg·kg−1 FW) |
LC-MS | [39] | |
| Peanuts (TCS: 0.01–15 mg·kg−1 FW) |
RP-HPLC DAD | [40] |
Polyphenols are one of the most widespread and studied group of plant secondary metabolites, mainly for their contribution to the nutritional value of plant food-based diets and their benefits for human health. The consumption of foods and beverages enriched in polyphenols have been associated to the prevention of human diseases related to cellular oxidative stress [2]. Beside the well-known antioxidant activities, polyphenols have also investigated for anti-inflammatory properties at molecular level and protection against aging and chronic diseases, such as inflammatory bowel diseases [2,3].
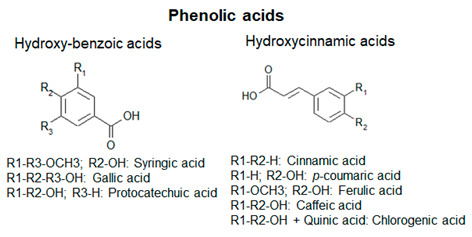
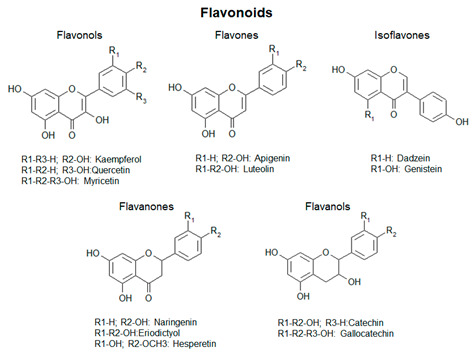
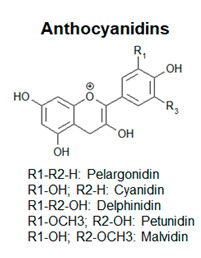
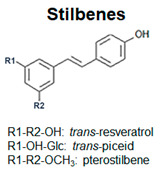
Plant polyphenols show a wide structural and chemical variability, among different families of plant kingdom or within the same family [4]. This structural diversity suggests that they do not play the same function: the different biological activities are influenced by the amounts and the nature of the lateral moieties linked to the basic structural backbone, composed by variously hydroxylated or decorated aromatic rings. The presence of hydroxyl groups, for example, confers to polyphenols the capacity to participate in oxidation reactions, which take part in several physiological processes [5]. Some polyphenols are pigmented (i.e., anthocyanins), therefore, being involved in the attraction of pollinators and seed-dispersers [6]. Other polyphenols are involved in the processes of cellular growth (i.e., hydroxycinnamic acids, lignins). Phenolic acids are directly involved in the defense towards different stresses, since they contribute to lignification of wounded tissues and have antimicrobial activities [7]. Flavonoids, one of the most common and studied classes of polyphenols, function as sunscreens and protect from UV exposure [8,9].
Polyphenols are synthesized in plants via phenylpropanoid pathway (Figure 1), through a finely orchestrated sequence of biochemical steps, coordinated by different structural and regulatory genes. Hence, the variability in the quantity and type of polyphenols found in a specific plant species derives from the activation of these genes resulting from the genetic background and plant environmental adaptation [10,11].
Figure 1.

Schematic representation of the general polyphenol biosynthetic pathway, with the main classes of polyphenols. PAL, Phenylalanine ammonia lyase; C4H, cinnamic acid 4-hydroxylase; C3H, p-coumarate 3-hydroxylase; HCT, hydroxycinnamoyl CoA shikimate/quinate hydroxycinnamoyl transferase; HQT, hydroxycinnamoyl CoA quinate hydroxycinnamoyl transferase; 4CL, 4-coumarate:coenzyme A ligase; STS, stilbene synthase; CHS, chalcone synthase; CHI, chalcone isomerase; IFS, isoflavone synthase; FNS, flavone synthase; F3H, flavanone-3-hydroxylase; FLS, flavonol synthase; DFR, dihydroflavonol reductase; LAR, leucoanthocyanidin reductase; ANS, anthocyanidin synthase; ARS, anthocyanidin reductase; GTs, glycosyltransferases.
Based on the number of aromatic rings and the structural elements linked to these basic rings, polyphenols can be distinguished in different classes: phenolic acids, including hydroxybenzoic and hydroxycinnamic acids; flavonoids, including flavonols, flavanones, isoflavones, flavanols; anthocyanins; stilbenes [12] (Figure 1; Table 1).
2. Plant Polyphenols and Gut Health: A Focus on the Antioxidant and Anti-Inflammatory Properties
Plant polyphenols commonly assumed with fruits and vegetables in human diet have been extensively associated to healthy beneficial effects, ranging from the antiaging, anti-tumoral, or anti-diabetic effects, to a positive impact on non-communicable diseases, such as cardiovascular or neurodegenerative diseases [1,41]. Several studies have also evidenced a beneficial impact of polyphenols for a healthy intestinal status, contributing to the prevention and protection against gut pathological processes, which often evolve in both acute and chronic states of inflammation leading to inflammatory bowel diseases (IBDs). Such pathologic conditions concur to stimulate the immune system at gut and systemic level [42]. In this context, nutritional strategies, including dietary anti-inflammatory compounds, could be beneficial at least in the prevention of these diseases [43,44].
Polyphenols are one of the most abundant antioxidant class in human diet [45]. A first reason explaining polyphenols antioxidant activities lies on their chemical structure and essentially on the presence of phenolic groups, particularly cathecol or galloyl groups [46]. Most of polyphenols are reducing agents and together with ascorbic acid, carotenoids and vitamin E, contribute to cell protection from tissue oxidative stress and damage [47]. Being reducing agents, polyphenols can exert a direct antioxidant activity on free radicals and Reactive Oxygen Species (ROS). Besides this “scavenging” role, polyphenols can have direct effects on oxidation of membrane lipids and the modulation of different enzymes [47,48], such as glutathione peroxidase and superoxide dismutase [41,48]. Through direct interaction with proteins involved in signal transduction, polyphenols can influence the cell redox status and trigger a series of redox-dependent reactions [45]. For example, Moskaug and co-workers [49] showed that flavonoids are able to increase the expression of γ-glutamylcisteine synthase (a rate-limiting enzyme in the synthesis of glutathione), with a concomitant increase of glutathione. As glutathione is implicated in several physiological and pathological cellular processes, polyphenols-mediated regulation of glutathione concentration can be one mechanism by which diet can influence oxidative related diseases [49].
Polyphenols can also impact on immune responses and intracellular signaling pathways, especially at intestinal level [3,50,51,52]. The effects on the inflammatory responses involve both cellular (macrophages, dendritic cells, lymphocytes) and molecular (cytokines, interleukins) mediators of inflammation. In vitro and in vivo studies have clearly demonstrated the anti-inflammatory effects of polyphenols (see Table 2), and their metabolites deriving from the processes of biotransformation at gut microbiota level [53,54,55,56].
Table 2.
Examples of in vitro and in vivo studies used to evaluate polyphenols antioxidant and anti-inflammatory activities at intestinal level.
| Plant Sources and Class of Polyphenols | In Vitro/In Vivo Model | Type of Administration/ Treatment |
Biological Activity | Reference |
|---|---|---|---|---|
| White grape skin (phenolic acids, proanthocyanidin, catechin, quercetin) |
HT-29 cells | Methanolic extract | Restored stress-related GSH reduction by polyphenols in intestinal cells | [84] |
| Apple (catechins, chlorogenic acid) | MKN 28 cells; Male Wistar rats |
Methanolic extract; polyphenol administration by drinking water or gavage |
Prevention of oxidative injury in gastric epithelial cells and gastric mucosa | [85] |
| Apple (flavonoids, phenolic acids) | HT-29 cells; CaCo-2 cells | Cider, apple juice | High preventive antioxidant capacity, decreased cellular reactive oxygen species, reduced oxidative cell damage | [86] |
| Grape pomace (phenolic acids, procyanidins) | IPEC-1 cells; TOPIG hybrid pigs | Aqueous extract; dietary administration of grape pomace | Decrease of lipid peroxidation in duodenum and colon and increase of the total antioxidant status | [87] |
| Pomegranate (ellagitannins) | Liposome model (large unilamellar vesicles, LUVs) | Pomegranate juice | Inhibition of the lipid peroxidation | [88] |
| Blueberries (anthocyanins) | CaCo-2 cells | Methanolic extracts | Reduced cellular oxidative stress | [89] |
| Red wine (tannins; anthocyanins) | HT-29 cells | Extract from red wine | Reduced iNOS and COX-2 levels, modulation of the NFkB signaling pathway | [90] |
| Grapeseeds (flavonoids) | CaCo-2 cells | Ethanolic extract | Reduced NfKB transactivation and TNFα transcripts levels | [91] |
| Dried peel of apple (flavonols, catechins, procyanidins) |
CaCo-2/15 cells | Crude extract and purified polyphenolic fraction | TNFα, IL6, E2 prostaglandin, COX2, NFkB down-regulation | [92] |
| Purple engineered tomato (flavonoids, anthocyanins) | CEC cells | Methanolic extracts | Inhibition of the pro-inflammatory cytokines | [93] |
| Green tea (polyphenols) | BALB/c mice with DSS-induced colitis | Dietary administration in chow diet | Reduction of TNFα and GSH levels | [94] |
| Strawberries (anthocyanins) |
Wistar rats with ethanol-induced gastric lesions | Oral administration of raw extract | Reduction of the ulcerative index; reduction of the gastric lipid peroxidation | [95] |
| Apple polyphenols | C57/BL6 mice with DSS-induced colitis | Administration of polyphenol mix in drinking water or oral gavage | Reduced levels of mediators of inflammation (TNFα, IFNγ, IL1β, IL6, IL17, IL22) | [96] |
| Green tea polyphenols (epigallocatechin-3-gallate) | C57/BL6 mice with DSS-induced colitis | Administration of polyphenol mix via oral gavage | Reduction of tissue damage and neutrophiles accumulation; increased levels of antioxidant enzymes | [97] |
| Grape juice | Wistar rats with TNBS-induced colitis | Administration in drinking water | Reduced inflammatory activity | [98] |
| Grape pomace | Wistar rats with DSS-induced colitis | Administration in chow diet | Reduced tissue damage and pro-inflammatory cytokines levels | [99] |
| Cocoa polyphenols | BALB/c mice with DSS-induced colitis | Administration in chow diet | Reduction of tumoral incidence, partially limited activation of the IL-6/STAT3 pathway | [100] |
| Bronze engineered tomato (flavonols, anthocyanins, stilbenes) | C57/BL6 mice with DSS-induced colitis | Administration in chow diet | Reduced inflammatory symptoms; beneficial changes in gut microbiota composition, reduced pro-inflammatory cytokines levels | [73] |
| Red and white grape skin (flavonols, anthocyanins, stilbenes) | C57/BL6 mice with DSS-induced colitis | Administration in chow diet | Reduced inflammatory symptoms; reduced pro-inflammatory cytokines levels | [73] |
| Bronze engineered tomato (flavonols, anthocyanins, stilbenes) | Winnie mice spontaneously developing colitis | Administration in chow diet | Changes in gut microbiota composition; reduced pro-inflammatory cytokines levels | [74] |
A first anti-inflammatory mechanism concerns the modulation of mediators of inflammation through the phospholipase A2 and cyclo-oxygenase 2 (COX-2) enzymatic activities. Polyphenols have been reported to reduce COX-2 activity and relative mRNAs in different cell lines [57,58]. A second mechanism involves the modulation of cytokines. For example, flavonoids are able to impair the expression of the interleukins IL-1β, IL-6, IL-8, and tumor necrosis factor (TNF) [51,53]. Other signaling pathways modulated by polyphenols participate to the inflammatory responses, proliferation and apoptosis, as in the case of the NFkB signaling pathway (Nuclear Factor kappa-light-chain enhancer of activated B cells [59]), and the MAP kinases pathway (Mitogen-Activated Protein kinase; [3,60]). In fact, the convergence of oxidative stress, inflammatory stimuli and their amplification by inflammatory cells and cytokines, culminate with the IkB kinases (IkKs) activation and the consequent IkB (cytoplasmic inhibitor of kB) phosphorylation.
NFkB, which is normally retained by IkB in the inactive form, is released after IkB phosphorylation, and once translocated to the nucleus, it induces the transcription of inflammatory genes (e.g., COX-2, TNFα, inducible nitric oxide synthase - iNOS). Polyphenols like epigallocatechin gallate, quercetin and anthocyanins have been showed to inhibit IkK phosphorylation, with consequent inhibition of the NFkB activation [56,61].
In the case of the MAPKs signaling pathway, polyphenols like anthocyanins can modulate the MAPK phosphorylation (p38MAPK, ERK, JNK) or the activity of transcription factors targeted by MAPKs (e.g., STAT-1, STAT-3) [3,61].
Another important mechanism concerns the nitric oxide (NO) production at the vascular level. NO is an essential component for vascular health maintenance and is involved in the inflammatory response triggered by free radicals. Some studies suggest that polyphenols inhibit NO release, suppressing the expression of endothelial nitric oxide synthetase (eNOS) [62,63]. Polyphenols can also promote a vascular remodeling activity and inhibit the extracellular matrix-degrading enzymes, i.e., the matrix metalloproteinase-2 and -9 [64,65].
Yet, an emerging and recently proposed anti-inflammatory mechanism is related to the cellular iron homeostasis and metabolism of dendritic cells (DCs), one of the major players of the intestinal adaptive immune response. The balance between the iron intracellular efflux and its extracellular transport (import/export) has been associated to DCs activation [66]. Iron uptake is a crucial response for immune cells against invading bacteria, whereas a physiological status of tolerance and tissue repair favors iron degradation and release, mediated by heme-oxygenase 1 (Hmox-1) and Ferroportin-1 [67]. The administration of iron-sequestrating quercetin, for example, induces a cascade of events starting with a strong up-regulation of Hmox-1 and Ferroportin-1 in DCs, resulting in iron release in the extracellular compartment and a reduction of cytoplasmic iron content [66]. Iron-depleted immune cells fail to produce inflammatory mediators while supporting the release of secretory leukoprotease inhibitor (Slpi) [68,69].
The preventive role of polyphenols on gut inflammation has been further confirmed using in vivo models. To date, several studies on rodent models of experimentally-induced IBDs, have indicated that dietary polyphenols can be effective in the prevention of these idiopathic diseases [70]. For example, the oral supplementation of dried bilberries reduced the pathological signs of inflammation, including IFNγ and TNF production by mesenteric lymph node cells, in a Balb/c mouse model of acute colitis induced by dextran sodium sulfate (DSS) [71]. The administration of pomegranate extract and its ellagic acid rich fraction has been described with anti-ulcerative effects and amelioration of the inflammatory condition in DSS-induced colitis [72]. In the same murine model, the administration of white or red grape-based diets, as well as of an engineered polyphenol-enriched tomato diet, has been useful to reduce inflammation symptoms and decrease the pro-inflammatory IL-6 and TNF [73]. Confirming results have been described in a spontaneously developing colitis mouse model (Winnie mice), with preventive roles under both homeostatic or inflammatory conditions [74].
Together with a direct anti-inflammatory role displayed by polyphenols, an increasing amount of literature is more recently exploring the possibility that they can favor the growth of beneficial microbial communities [75,76,77]. Polyphenol-induced changes on the gut microbiota have been described either at phylum and genera levels, modifications that could be taken into consideration in the treatment of intestinal dysbiosis, which can trigger or exacerbate a chronic inflammation status. For example, the dietary administration of blueberries, blackcurrants and other sources containing polyphenols, have been reported to beneficially impact on the Firmicutes/Bacteroidetes ratio in murine models [73,74,78]. The promotion of the growth of probiotic genera, such as Bifidobacterium or Lactobacillus, has been observed following anthocyanins-enriched diets [73,79]. On the other hand, antimicrobial effects and a reduction in the growth of Bacteroides, Prevotella, Oscillospira, Blautia genera and Clostridium spp. (these latest often associated to pathological conditions) have also been described following polyphenol administration [53,78,80,81,82]. However, an important aspect that should be considered is the bioavailability and the consequent bio-efficacy of polyphenols, which is largely influenced by the physicochemical properties of polyphenols, the modifications they undergo at gut level, the composition and interactions with complex food matrices and the biotransformation processes exerted by microbial groups, converting polyphenols in more simple, absorbable metabolites [83]. The bio-efficacy of microbiota metabolized forms of polyphenols have also been studied [53,54], highlighting that these forms can also display relevant biological properties [75].
Taken together all these findings indicate an overall bioactive potential of polyphenols useful for the prevention of inflammatory conditions at the gut level and point the central issue related to their content in the main fruits and vegetables commonly consumed by Wstern people.
3. Biofortification Strategies for the Improvement of Polyphenol Content in Plant Food
Daily intake of polyphenols is considered suboptimal in many Western countries. Del Bo’ and co-workers [101] suggested a total polyphenol intake of about 900 mg/day, considering literature studies on polyphenol intake assessment using various quantification methodologies and some polyphenols databases sources (e.g., USDA, Phenol-Explorer). A large variation in polyphenols uptake was also observed according different dietary patterns (e.g., Mediterranean diet, Western diet), lifestyle, educational and economic levels, significantly affecting the general eating habits.
Therefore, people are asked to shift their nutritional habits towards a larger use of fruits and vegetables. However, as indicated in Table 1, many species where higher amounts of polyphenols have been reported are not largely consumed or available throughout all the year. Furthermore, a large amount of bioactive compounds are lost after cooking, food processing phases or other common practices frequently undertaken before ingestion (i.e., peel removal). Therefore, the increase in the content of healthy beneficial plant secondary metabolites is mandatory to address the nutritional security issue to a large part of world population.
Biofortification refers to a number of different strategies aimed at the nutritional improvement of food crops, spanning from the agronomic practices and conventional breeding, to the modern biotechnological tools [102] (Table 3). The agronomic methods for biofortification include the application of nutrients to improve the content in minerals, vitamins or bioactive compounds. The typical example of plant biofortification is the improvement of iron, zinc, selenium or iodine content in soil or water or the application of essential micronutrient-based fertilizers to boost the nutritional value of commercial crops [102]. However, in recent years, the definition of biofortification has undergone an extension of its meaning, thus including not only the micronutrient improvement, but also the enhancement of crop nutritional content by including several classes of bioactive compounds and other plant secondary metabolites [103,104,105]. Some agronomical strategies, including the employment of conventional/organic farming methods or reduction of nitrogen fertilization, have been reported to impact on the content of health-promoting secondary metabolites, such as polyphenols [106,107]. Organic farming has been showed positively influencing the levels of polyphenols in vegetables, as in the case of phenolic acids and flavonoids in eggplants [108], cabbage [109] or lettuce [110]. However, the effects of different agronomic management practices on polyphenols levels in edible parts of horticultural crops are not easily predictable [111]. In fact, both conventional or organic farming could concur to nitrogen supply, which is an important parameter influencing either primary and secondary metabolites biosynthesis. However, in the case of polyphenols limited effects have been reported in some cases [112]. In others, a high nitrogen supply in the soil negatively impacted on the flavonoid accumulation in broccoli, basil and tomato [110], or on the anthocyanins content in grapevine berries (Table 3) [113].
Table 3.
Schematic summary of common biofortification methods for the improvement of polyphenol content in plant foods.
| Biofortification Method | Pros | Cons | |
|---|---|---|---|
| Conventional approaches | Agronomic practices/ fertilization strategies |
|
|
| Exposition to stresses (e.g., light-UV treatments) |
|
|
|
| Conventional breeding programs |
|
|
|
| Biotechnological approaches | Transgenic methods |
|
|
| Genome editing technologies |
|
|
|
Environmental factors and application of stresses can also affect polyphenol accumulation and provide an alternative way for biofortification. Light intensity or UV exposure under a controlled environment can stimulate anthocyanins or flavonoids production, since the biosynthetic pathway can be modulated by light response, as in the case of turnip seedlings [114], dark-purple tea plants [115], or red leaf lettuce (Table 3) [116].
Conventional breeding strategies are the most common way to select agronomic traits carrying nutritional features of consumers’ interest. The genetic improvement can be achieved either by selection of plants with desirable traits for seed and vegetative propagation, or by crossing closely related individuals to produce new hybrids [117]. In both cases, the genetic variability and availability is a necessary condition for the success of the breeding strategy to improve nutritional characteristics. Hence, the selection process starts from the knowledge of the main genetic traits responsible for the synthesis of specific phytonutrients, since a large quantitative variation may occur between cultivated and wild species. In this way, it is possible the introgression of the specific traits from wild to domesticated species. An example of such strategy is represented by the Sun Black tomato variety, which can accumulate anthocyanins in the fruit skin, due to the presence of an introgressed dominant Aft (Anthocyanin fruit) gene together with the atv (atroviolacea fruit) gene [118,119]. Another example is represented by an eggplant variety derived from some introgression lines (S. integrifolium, S. aethiopicum, S. sodomaeum), which showed enhanced nutritional properties and high levels of polyphenols [120].
Furthermore, new advances in molecular biology, genomics and metabolomics have brought a further contribute in the assisted selection and breeding, and the detection and mapping of the major quantitative trait loci (QTL) is greatly promising for this scope [117].
The biotechnological approach has the advantage to select and directly introduce genetic traits of interest without the strong limitation of compatible genetic background, as in the case of conventional breeding [104]. This approach includes the classical transgenic modifications and the novel breeding technologies (NBTs), which are now rapidly emerging and proposing as the flywheel for the next generation of functional foods. In both cases, a specific plan of metabolic engineering is required, based on a deep scientific knowledge of plant metabolisms, besides the possibility to easily transform and regenerate the desired plant species [121].
Genetic engineering has already showed its potential addressing nutritional requirements, in specific plant organs (i.e., seed, vegetative tissues, fruit) and combining multiple introduced genetic traits without complex and long-term breeding programs (Table 3) [104]. One of the most popular strategies of metabolic engineering include the use of key structural genes of the polyphenol pathway, aiming mostly their over-expression, to stimulate the new biochemical steps that lead to the accumulation of new classes of polyphenols. This strategy resulted in polyphenol content enhancement [122,123,124], even though in some cases with very modest effects [125]. On the other hand, the modulation of polyphenol pathway through the use of regulatory genes encoding transcription factors revealed to be a more effective strategy for polyphenol enhancement in edible crops. Examples of this successfully approach have been reported in apple [126], tomato [127,128], or potato [129].
A second generation of metabolic engineered plant foods include the combination of structural and regulatory genes, a multilevel engineering strategy which led, in some cases, to a high accumulation of polyphenols, enhanced the nutritional potential of relevant crops to reach health benefits [74,130]. Yet, some examples come from tomato, in which the contextual enrichment of high levels of phenolic acids, flavonols, stilbenes, isoflavones or anthocyanins has been realized [74,130]. Anyway, the single or multilevel strategies also allowed to compare the healthy beneficial effects of different classes of phytochemicals within the same food matrix which is an important pre-requisite for the development of tailored crops for specific nutritional requirements.
4. The Genome Editing: A Step Forward in Plant Metabolic Engineering
Despite the metabolic engineering approach has offered a new and exciting way of thinking the plant nutritional quality, conventional breeding still remains the most widely used method for crop improvement, although it is labor-intensive and usually takes several years for the screening and identification of genotypes with desirable genetic traits. On the other hand, the use of genetically modified (GM) crops is still largely debated for health and environmental safety concerns, which have greatly restricted the GM use to a small number of available crops (Table 3). More recently, the so-called new breeding technologies (NBTs), which include the genome editing ones, have emerged as very powerful tools for precise modifications of desired genetic traits. The genome editing mediated by transcription activator-like effector nucleases (TALENs) and zinc-finger nucleases (ZFNs) have represented the initial pioneers, although recently the clustered regularly interspaced short palindromic repeats (CRISPR)/Cas system has mainly been preferred and utilized.
In all cases, sequence-specific nucleases (SSNs) recognize target genome sequences and generate double-stranded breaks (DSBs). The endocellular systems (the error-prone non-homologous end joining, NHEJ, or the homologous recombination, HR) repair the DSBs, leading to deletions or insertions, thus causing gene knockout or, in some cases, replacement [131]. Although the NHEJ is the primary and most efficient mechanism of repair in plant cells [132], HR can permit a precise genome repair by using customized template sequences flanked by homologous ends or arms compatible with the DSBs’ site [133].
In the case of zinc finger nucleases (ZFNs), the technology is based on chimeric proteins, composed of an artificial zinc finger DNA binding domain at the N-terminal region and a FokI DNA cleavage domain at the C-terminal region. The zinc finger DNA binding domain can be modified to specifically target the genomic sequence of interest. In fact, since each zinc finger protein recognizes three tandem nucleotides, the manipulation in the order of more zinc fingers enable the recognition and binding to specific sequences. Therefore, the assembly of ZFNs involves the modular design and linking of zinc fingers in a sequence that allow the recognition of the DNA target sequence [134].
TALENs-mediated genome editing is instead based on sequence-specific nucleases consisting of transcription activator-like effectors fused to the catalytic domain of the FokI endonuclease. The DNA-binding domain in TALE monomers is composed of a central repeat domain (CRD) that directs DNA binding [135]. The CRD is formed by tandem repeats, each binding a single nucleotide in the target sequence, a one-to-one pairing that enables TALENs to target the specific sequence [131].
However, recent advances highlighted how the CRISPR/Cas systems can represent a more versatile tool for genome editing in comparison with ZFNs or TALENS [136]. This system takes advantage of a Cas9 endonuclease, composed by a recognition functional domain and a nuclease domain, a guide RNA, formed by the complex of non-coding RNA elements (crRNA) and small trans-encoded RNA elements (i.e., tracrRNA) [137,138,139], which directs the Cas9 endonuclease to the target sequence along with the PAM (protospacer adjacent motif) site. To date, several Cas9 variants are also available, such as Cas9 nickase (nCas9), which is derived from a mutation in native Cas9 domains and has the ability to induce nicks in the genome [135], or dead Cas9 (dCas9), which is basically a catalytically inactive Cas9 [137,140].
dCas9 represents a novel tool for the genetic manipulation through transcriptional alteration since the expression levels of a specific gene can be altered by assembling dCas9 with transcriptional regulators that are recruited to the promoter region. In a similar manner, dCas9 can be fused to epigenetic modulators, such as methylation and deacetylation enzymes. Therefore, CRISPR/dCas9 systems can also be a useful tool for epigenome editing, modulation of chromatin topology and live-cell chromatin imaging in plants. However, suitable transcriptional modulators, accurate study of the target sites, appropriate sgRNA construct design and delivery of the CRISPR/Cas9 system are important requirements for the success of this application [141].
In addition to these Cas9 variants, other CRISPR/Cas systems are now rapidly developing, which might overcome some CRISPR/Cas9 limitations [142]. For example, CRISPR/Cas12 (CRISPR/Cpf1) might enhance the editing efficiency (especially of insertions at the target site) [140,142], CRISPR/Cas13 has an efficiency comparable to RNAi systems [143], or CRISPR/Cas14a, a “miniature” nuclease able to cleave a single-strand DNA, are now being considered for applications in plant species [142,144].
In general, the limitations of the CRISPR/Cas9 system include the availability of suitable plant transformation/regeneration systems. In fact, the most common method of plant transformation is so far mediated by Agrobacterium strains, although new delivery systems suitable for protoplasts or biolistic approaches have also been proposed. DNA-free in vitro systems that consist in the direct delivery of a ribonucleoprotein complex, formed by the Cas9 protein and the guide RNA has been recently proposed [127,136,145]. However, these types of delivery show a variable efficiency in different plant species [132], a reason that pushes the necessity of an accurate planning of the plant transformation step.
5. Genome Editing Technologies and Nutritional Aspects: A Focus on Polyphenols Biosynthesis Modification
Gene knockouts or insertion mutants produced by genome editing technologies can be useful tools for crop nutritional improvement. The modifications can impact either on anti-nutritional factors or on nutritionally/organoleptic relevant compounds, thus finally influencing the health-promoting properties of food products.
For example, TALENs have been used in potato tubers to knock out the vacuolar invertase gene (Vinv), resulting in low levels of undesirable reducing sugars in tubers [146]. A new rice line has been created targeting TALENs on the OsBADH2 gene, responsible for the biosynthesis of 2-acetyl-1-pyrroline, with the result of restoring the content of this compound, an important component of the fragrance of natural rice mutants [136,147]. TALENs has also been used to knock out the sterol side chain reductase 2 (SSR2) gene, responsible of non-desirable steroidal glycoalkaloids, with a consequent reduction in the levels of chaconine and solanine in new potato lines [148]. TALENs have also been employed for the improvement of starch metabolism in potato tubers [149] or to target the SlANT1 gene, involved in the anthocyanin biosynthesis in tomato [150].
CRISPR/Cas9 was used to simultaneously target conserved domains in the α-gliadin gene family in wheat and obtain new low-gluten lines [151]. Nutritional quality has also been improved by lowering the content in antinutrient compounds, as the reduction of phytic acid content by inducing mutations on OsITPK6 gene in rice grain [152], or reducing anti-nutritional proteins by editing the α-amylase/trypsin inhibitor genes in durum wheat [153].
Improved quality by using CRISPR/Cas9 system has been achieved in Camelina sativa seeds producing high-oleic acid levels [154,155], in Brassica napus, by editing the fatty acid desaturase 2 gene and improving oleic acid levels [156], in tomatoes, improving the γ-aminobutyric acid content by using a multiplexed CRISPR/Cas9 system on five key genes (PDS, GABATP1, GABA-TP2, GABA-TP3, CAT9, SSADH genes: phytoene desaturase, pyruvate-dependent GABA-transaminase 1, pyruvate-dependent GABA-transaminase 2, pyruvate-dependent GABA-transaminase 3, cationic aminoacid transporter 9 and succinate semialdehyde dehydrogenase genes, respectively; [157,158]), or in lycopene-enriched tomatoes by targeting SGR1, LCY-E, Blc, LCY-B1 and LCY-B2 genes (stay-green 1, lycopene δ-cyclase, beta-lycopene cyclase, lycopene β-cyclase 1 and 2, respectively) [159].
CRISPR/Cas9 system has been studied and tested also for polyphenol biosynthesis, in some cases to test the blockage at some levels of the pathway, that can give an easy colored proof-of-concept of its correct functioning, and eventually to provide a demonstration of the application of the system [160,161,162] (Table 4). For example, CRISPR/Cas9 targeting the carrot flavanone-3-hydroxylase (F3H) gene have been tested for blocking the anthocyanin biosynthesis in a model of purple-colored calli [163], torenia [161], petunia [164] and black rice [165].
Table 4.
Examples of genome editing systems for the study, blockage, enrichment or preservation of the polyphenol content in plant models or crop species.
| Model or Species | Target Gene | Genome Editing System | Effect | References |
|---|---|---|---|---|
| Japanese morning glory (Ipomoea [Pharbitis] nil) |
Dihydroflavonol- 4-reductase-B (InDFR-B) |
CRISPR/Cas9 | White flower mutants | [160] |
| Purple-colored calli of carrot (Daucus carota L.) | Flavanone-3-hydroxylase (DcF3H) | CRISPR/Cas9 | Block of anthocyanin biosynthesis, resulting in non-purple-colored calli | [163] |
| Torenia plant (Torenia fourneri L.) |
Flavanone-3-hydroxylase (TfF3H) |
CRISPR/Cas9 | Block of anthocyanin biosynthesis, resulting in pale blue flowers | [160] |
| Petunia (Petunia hybrida cv. Madness Midnight) |
Flavanone-3-hydroxylase (PhF3H) |
CRISPR/Cas9 | Modified, pale purplish flower color | [164] |
| Sage (Salvia miltiorrhiza) hairy roots | Rosmarinic acid synthase (SmRAS) | CRISPR/Cas9 | Decreased content of phenolic acids, including rosmarinic acid and lithospermic acid | [173] |
| Black rice (Oryza sativa L. cv Heugseonchal and Sinmyungh) |
Flavanone-3-hydroxylase, Dihydroflavonol-
4-reductase, Leucoanthocyanidins dioxygenase (OsF3′H, OsDFR and OsLDOX) |
CRISPR/Cas9 | Reduction of anthocyanin accumulation in mutant lines | [165] |
| Japanese gentian (cv ‘Albireo’, Gentiana triflora × Gentiana scabra) |
Anthocyanin- related glutathione S-transferase (GST) | CRISPR/Cas9 | Reduction of anthocyanin content | [162] |
| Tomato (Solanum lycopersicum L.) |
Dihydroflavonol-
4-reductase (SlDFR) |
CRISPR/Cas9 | Reduction of anthocyanin content in seedlings | [170] |
| Pomegranate (Punica granatum L.) hairy roots |
UDP-dependent glycosyltransferases (PgUGT84A23 and PgUGT84A24) |
CRISPR/Cas9 | Changes in galloyl-glucose conjugates and derivatives | [174] |
| Rapeseed (Brassica napus L.) | TRANSPARENT TESTA 8 (BnTT8) | CRISPR/Cas9 | Yellow seeds with improved oil and protein contents, block of pro-anthocyanidin deposition in seed coat | [168] |
| Tomato (Solanum lycopersicum L.) | Anthocyanin mutant 1 (SlANT1) | TALENs CRISPR/Cas9 CRISPR/Cpf1 |
Anthocyanin accumulation in the whole tomato plant | [150,169] |
| Tomato (Solanum lycopersicum L.) | R2R3MYB transcription factor 12 (SlMYB12) | CRISPR/Cas9 | Accumulation of flavonoids in pink-colored tomato fruits | [166] |
| Tomato (Solanum lycopersicum L.) | Anthocyanin 2 (SlAN2) | CRISPR/Cas9 | Anthocyanin pigmentation in tomato fruits | [167] |
| Grapevine (Vitis vinifera L. rootstock 101-14) | Trans-Acting Small-interfering locus4 (VvTAS4) and R2R3MYB transcription factor A5/6/7 (VvMYBA5/6/7) | CRISPR/Cas9 | Lack of visible pigment phenotypes in edited plants | [175] |
| Mushroom (Agaricus bisporus) Potato (Solanum tuberosum L.) |
Polyphenol oxidase (PPO) | CRISPR/Cas9 | Reduced fruit browning | [171,172] |
In the specific case of polyphenol content improvement, the CRISPR/Cas9 system has been employed to reduce/knockout the function of negative regulators of the phenylpropanoid/flavonoid pathway or vice-versa by trying to enhance the function of genes expression activators implicated in polyphenol biosynthesis. As an example, the regulatory SlMYB12 gene knocking out have led to an accumulation of flavonoids in pink-colored tomato fruits [166]. SlAN2 tomato mutants, exhibiting anthocyanin pigmentation and accumulation similarly to the ‘Indigo Rose’ variety, has been generated using CRISPR/Cas9 targeted on the gene sequence encoding the AN2 MYB transcription factor [167]. CRISPR/Cas9 has also been employed to induce mutations on the regulatory BnTT8 gene. The mutants showed a suppressed phenylpropanoid/flavonoid pathway and pro-anthocyanidin deposition in the seed coat and a desirable trait of yellow-seeds [168]. On the other hand, Čermák and co-workers [150] have delivered a CRISPR/Cas9 together with a geminivirus-based replicon system to target the SlANT1 gene and introduce upstream to its sequence a constitutive promoter, exploiting the HR mechanism of repair and thus developing new edited tomato lines that accumulate anthocyanins in the whole plants, including the fruits. Furthermore, in this study, the targeted DNA insertions were observed in segregating population with purple phenotypes, confirming the hereditability of the new mutated alleles. A similar approach was employed using CRISPR/Cpf1 in tomato to enhance the expression of the SlANT1 gene and resulting in anthocyanin-pigmented tomato plants [169], with similar patterns of segregation in the progeny as observed by Čermák and co-workers [150]. In another work, the HR mechanism of repair has been exploited to repristinate the anthocyanin pigmentation in tomato seedlings following a CRISPR/Cas9-induced deletion on the SlDFR target gene [170].
Other studies targeted the polyphenol oxidase (PPO), one of the polyphenols degrading-enzymes in fruit and vegetable, resulting in non-browning mushrooms and potatoes [171,172].
All these examples represent new and stimulating approaches to preserve or improve the nutritional value of crop species (Table 4).
6. Conclusions and Perspectives
In this review, we focused on the benefits that polyphenols bring to gut health, with special attention to their antioxidant and anti-inflammatory activities that can influence the immune-modulatory response at the basis of many intestinal diseases. Nutritional interventions for the prevention of these pathological processes represent an important hallmark for preventive nutrition. However, daily polyphenol intake is considered suboptimal for most people living in Western countries. Requests to change the nutritional habits and increase the consumption of fruits and vegetables are multiplying to address this goal and in general the nutritional security issue for Western countries populations. In this context, two possible strategies for polyphenols daily intake increase can be utilized. The first refers to the (re)discovery of under-utilized niche species that accumulate high levels of secondary metabolites in edible parts of the plant. Some examples are listed in Table 1. However, robust characterization of the main classes of secondary metabolites still lacks for many of these species and future efforts have to be undertaken to fill this gap. A second interesting strategy to increase the content of healthy components in major crops refers to all the biofortification strategies so far developed. Table 3 of this review reports about all the main traditional or innovative techniques to address this goal with limitations or strength points for each one. Among the conventional ones, breeding techniques require a long time for the selection of specific genetic traits of interest and their introgression in cultivated varieties. Agronomic practices can influence either yield and quality of crops, but results so far obtained on the polyphenols content look controversial. Stress application can be an exciting option, but it requires great attention to select those impacting the least possible on the final yield and quality of fresh products.
Among biofortification strategies more recently proposed and tested for polyphenols improvement, the multi-level metabolic engineering approach is an elegant strategy to finely regulate and induce new target genes, but restrictive regulatory policies in some countries (e.g., Europe) and a limited public acceptance have so far greatly limited their success.
New breeding technologies are emerging as possible alternatives to overcome the limitations of GMOs. However, given the wide versatility of genome-editing applications, it will be essential to select the most suitable systems for each plant species and intervention type (gene knockout/insertion). However, despite the many technical challenges (e.g., efficiency of the genome editing systems, efficiency of plant transformation, selection of transgenic-free mutant lines) still to be addressed, other important issues regarding the general public’s acceptance and the adoption of more permissive regulatory policies must be considered and settled before the wide application of these techniques in agriculture and improved crop varieties onto the market.
Acknowledgments
The authors acknowledge EU-H2020 CropBooster-P project (grant agreement 817690) and PlantEd–COST Action (Genome editing in plants; CA18111).
Author Contributions
A.S. (Aurelia Scarano), M.C. and A.S. (Angelo Santino) wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This work was in part funded by the Apulia region SICURA project (KC3U5Y1) and CNR-DiSBA project NutrAge (project nr. 7022).
Conflicts of Interest
The authors declare no conflict of interest.
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Koch W. Dietary Polyphenols—Important Non-Nutrients in the Prevention of Chronic Noncommunicable Diseases. A Systematic Review. Nutrients. 2019;11:1039. doi: 10.3390/nu11051039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Yao L.H., Jiang Y.M., Shi J., Tomás-Barberán F.A., Datta N., Singanusong R., Chen S.S. Flavonoids in food and their health benefits. Plants Foods Hum. Nutr. 2004;59:113–122. doi: 10.1007/s11130-004-0049-7. [DOI] [PubMed] [Google Scholar]
- 3.Romier B., Schneider Y.J., Larondelle Y., During A. Dietary polyphenols can modulate the intestinal inflammatory response. Nutr. Rev. 2009;67:363–378. doi: 10.1111/j.1753-4887.2009.00210.x. [DOI] [PubMed] [Google Scholar]
- 4.Mouradov A., Spangenberg G. Flavonoids: A metabolic network mediating plants adaptation to their real estate. Front. Plant Sci. 2014;5:620. doi: 10.3389/fpls.2014.00620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Heim K.E., Tagliaferro A.R., Bobilya D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002;13:572–584. doi: 10.1016/S0955-2863(02)00208-5. [DOI] [PubMed] [Google Scholar]
- 6.Zhang Y., Butelli E., Martin C. Engineering anthocyanin biosynthesis in plants. Curr. Opin. Plant Biol. 2014;19:81–90. doi: 10.1016/j.pbi.2014.05.011. [DOI] [PubMed] [Google Scholar]
- 7.Pandey K.B., Rizvi S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell Longev. 2009;2:270–278. doi: 10.4161/oxim.2.5.9498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Chong J., Poutaraud A., Hugueney P. Metabolism and roles of stilbenes in plants. Plant Sci. 2009;177:143–155. doi: 10.1016/j.plantsci.2009.05.012. [DOI] [Google Scholar]
- 9.Tohge T., Fernie A.R. Leveraging natural variance towards enhanced understanding of phytochemical sunscreens. Trends Plant Sci. 2017;2:308–315. doi: 10.1016/j.tplants.2017.01.003. [DOI] [PubMed] [Google Scholar]
- 10.Andrè M.C., Oufir M., Hoffmann L., Hausman J.-F., Rogez H., Larondelle Y., Evers D. Influence of environment and genotype on polyphenol compounds and in vitro antioxidant capacity of native Andean potatoes (Solanum tuberosum L.) J. Food Comp. Anal. 2009;22:517–524. doi: 10.1016/j.jfca.2008.11.010. [DOI] [Google Scholar]
- 11.Aruna C.R., Ratnavathi C.V., Suguna M., Ranga B., Kumar P.P., Annapurna A., Bahadure D.M., Toapi V.A. Genetic variability and GxE interactions for total polyphenol content and antioxidant activity in white and red sorghums (Sorghum bicolor) Plant Breed. 2020;139:119–130. doi: 10.1111/pbr.12756. [DOI] [Google Scholar]
- 12.Li A., Li S., Zhang Y.J., Xu X.R., Chen Y.M., Li H.B. Resources and biological activities of natural polyphenols. Nutrients. 2014;6:6020–6047. doi: 10.3390/nu6126020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Sun T., Simon P.W., Tanumihardjo S.A. Antioxidant phytochemicals and antioxidant capacity of biofortified carrots (Daucus carota L.) of various colors. J. Agric. Food Chem. 2009;57:4142. doi: 10.1021/jf9001044. [DOI] [PubMed] [Google Scholar]
- 14.López-Gámez G., Elez-Martinez P., Soliva-Fortuny R., Martin-Belloso O. Pulsed electric fields affect endogenous enzyme activities, respiration and biosynthesis of phenolic compounds in carrots. Postharvest Biol. Technol. 2020;168:111284. doi: 10.1016/j.postharvbio.2020.111284. [DOI] [Google Scholar]
- 15.Schütz K., Kammerer D., Carle R., Schieber A. Identification and Quantification of Caffeoylquinic Acids and Flavonoids from Artichoke (Cynara scolymus L.) Heads, Juice, and Pomace by HPLC-DAD-ESI/MSn. J. Agric. Food Chem. 2004;52:4090–4096. doi: 10.1021/jf049625x. [DOI] [PubMed] [Google Scholar]
- 16.D’Antuono I., Carola A., Sena L.M., Linsalata V., Cardinali A., Logrieco A.F., Colucci M.G., Apone F. Artichoke polyphenols produce skin anti-age effects by improving endothelial cell integrity and functionality. Molecules. 2018;23:2729. doi: 10.3390/molecules23112729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Torres-Mancera M.T., Cordova-López J., Rodríguez-Serrano G., Roussos S., Ramírez-Coronel M.A., Favela-Torres E., Saucedo-Castañeda G. Enzymatic extraction of hydroxycinnamic acids from coffee pulp. Food Technol. Biotechnol. 2011;49:369–373. [Google Scholar]
- 18.Andrade S., Guiné R.P.F., Gonçalves F. Evaluation of phenolic compounds, antioxidant activity and bioaccessibility in white crowberry (Corema album) J. Food Meas. Char. 2017;11:1936–1946. doi: 10.1007/s11694-017-9576-4. [DOI] [Google Scholar]
- 19.Barroca M.J., Guiné R.P.F., Amado A.M., Ressureição S., da Silva A.M.G.M., Marques M.P.M., de Carvalho L.A.E.B. The drying process of Sarcocornia perennis: Impact on nutritional and physico-chemical properties. J. Food Sci. Technol. 2020;57:4443–4458. doi: 10.1007/s13197-020-04482-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Riga P., Benedicto L., Gil-Izquierdo A., Collado J., Ferreres F., Medina S. Diffuse light affects the contents of vitamin C, phenolic compounds and free amino acids in lettuce plants. Food Chem. 2018;272:227–234. doi: 10.1016/j.foodchem.2018.08.051. [DOI] [PubMed] [Google Scholar]
- 21.Cao J., Chen W., Zhang Y., Zhang Y., Zhao X. Content of selected flavonoids in 100 edible vegetables and fruits. Food Sci. Technol. Res. 2010;16:395–402. doi: 10.3136/fstr.16.395. [DOI] [Google Scholar]
- 22.Koh E., Wimalasiri K.M.S., Chassy A.W., Mitchell A.E. Content of ascorbic acid, quercetin, kaempferol and total phenolics in commercial broccoli. J. Food Comp. Anal. 2009;22:637–643. doi: 10.1016/j.jfca.2009.01.019. [DOI] [Google Scholar]
- 23.Reis L.C.R.D., de Olivera V.R., Hagen M.K., Jablonski A., Flôres S.H., de Oliveira Rios A. Carotenoids, flavonoids, chlorophylls, phenolic compounds and antioxidant activity in fresh and cooked broccoli (Brassica oleracea var. Avenger) and cauliflower (Brassica oleracea var. Alphina F1) LWT Food Sci. Technol. 2015;63:177–183. doi: 10.1016/j.lwt.2015.03.089. [DOI] [Google Scholar]
- 24.Zhang J., Ge Y., Han F., Li B., Yan S., Sun J., Wang L. Isoflavone content of soybean cultivars from maturity group 0 to VI grown in northern and southern China. J. Am. Oil. Chem. Soc. 2014;91:1019–1028. doi: 10.1007/s11746-014-2440-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Nogata Y., Sakamoto K., Shiratsuchi H., Ishii T., Yano M., Ohta H. Flavonoid composition of fruit tissues of citrus species. Biosci. Biotechnol. Biochem. 2006;70:178–192. doi: 10.1271/bbb.70.178. [DOI] [PubMed] [Google Scholar]
- 26.Pereira V.P., Knor F.J., Vellosa J.C.R., Beltrame F.L. Determination of phenolic compounds and antioxidant activity of green, black and white teas of Camellia sinensis (L.,) Kuntze, Theaceae. Rev. Bras. Plantas Med. Camp. 2014;16:490–498. doi: 10.1590/1983-084X/13_061. [DOI] [Google Scholar]
- 27.Koch W., Kukula-Koch W., Komsta Ł., Marzec Z., Szwerc W., Głowniak K. Green tea quality evaluation based on its catechins and metals composition in combination with chemometric analysis. Molecules. 2018;23:1689. doi: 10.3390/molecules23071689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Huang W.-Y., Lin Y.-R., Ho R.-F., Liu H.-Y., Lin Y.-S. Effects of water solutions on extracting green tea leaves. Sci. World J. 2013;2013:368650. doi: 10.1155/2013/368350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Cho M.J., Howard L.R., Prior R.L., Clark J.R. Flavonoid glycosides and antioxidant capacity of various blackberry, blueberry and red grape genotypes determined by high-performance liquid chromatography/mass spectrometry. J. Sci. Food Agric. 2004;84:1771–1782. doi: 10.1002/jsfa.1885. [DOI] [Google Scholar]
- 30.Andrés-Lacueva C., Monagas M., Khan N., Izquierdo-Pulido M., Urpi-Sarda M., Permanyer J., Lamuela-Raventós R.M. Flavanol and flavonol contents of cocoa powder roducts: Influence of the manufacturing process. J. Agric. Food Chem. 2008;56:3111–3117. doi: 10.1021/jf0728754. [DOI] [PubMed] [Google Scholar]
- 31.Wojdyło A., Nowicka P., Carbonell-Barrachina A.A., Hernández F. Phenolic compounds, antioxidant and antidiabetic activity of different cultivars of Ficus carica L. fruits. J. Funct. Foods. 2016;25:421–432. doi: 10.1016/j.jff.2016.06.015. [DOI] [Google Scholar]
- 32.Orsavova J., Hlaváčová I., Mlček J., Snopek l., Misurcova L. Contribution of phenolic compounds, ascorbic acid and vitamin E to antioxidant activity of currant (Ribes L.) and gooseberry (Ribes uva-crispa L.) fruits. Food Chem. 2019;284:323–333. doi: 10.1016/j.foodchem.2019.01.072. [DOI] [PubMed] [Google Scholar]
- 33.Scalzo R.L., Genna A., Branca F., Chedin M., Chassaigne H. Anthocyanin composition of cauliflower (Brassica oleracea L. var. botrytis) and cabbage (B. oleracea L. var. capitata) and its stability in relation to thermal treatments. Food Chem. 2008;107:136–144. doi: 10.1016/j.foodchem.2007.07.072. [DOI] [Google Scholar]
- 34.Montilla E.C., Arzaba M.R., Hillebrand S., Winterhalter P. Anthocyanin composition of black carrot (Daucus carota ssp. sativus var. atrorubens Alef.) cultivars Antonina, Beta Sweet, Deep Purple, and Purple Haze. J. Agric. Food Chem. 2011;59:3385–3390. doi: 10.1021/jf104724k. [DOI] [PubMed] [Google Scholar]
- 35.Manach C., Scalbert A., Morand C., Rémésy C., Jiménez L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004;79:727–747. doi: 10.1093/ajcn/79.5.727. [DOI] [PubMed] [Google Scholar]
- 36.Guiné R.P.F., Gonçalves F., Lerat C., El-Idrissi T., Rodrigo E., Correira P.R., Gonçalves J.C. Extraction of Phenolic Compounds with Antioxidant Activity from Beetroot (Beta Vulgaris L.) Curr. Nutr. Food Sci. 2018;14:350–357. doi: 10.2174/1573401313666170609102336. [DOI] [Google Scholar]
- 37.Guiné R.P.F., dos Reis Correia P.M., Ferrão A.C., Gonçalves F., Lerat C., El-Idrissi T., Rodrigo E. Evaluation of phenolic and antioxidant properties of strawberry as a function of extraction conditions. Braz. J. Food Technol. 2020;23:e2019142. doi: 10.1590/1981-6723.14219. [DOI] [Google Scholar]
- 38.Vincenzi S., Tomasi D., Gaiotti F., Lovat L., Giacosa S., Torchio F., Río Segade S., Rolle L. Comparative study of the resveratrol content of twenty-one Italian red grape varieties. S. Afr. J. Enol. Vitic. 2013;34:30–35. doi: 10.21548/34-1-1078. [DOI] [Google Scholar]
- 39.Błaszczyk A., Sady S., Sielicka M. The stilbene profile in edible berries. Phytochem. Rev. 2019;18:37–67. doi: 10.1007/s11101-018-9580-2. [DOI] [Google Scholar]
- 40.Hasan M.M., Cha M., Bajpai V.K., Baek K.-H. Production of a major stilbene phytoalexin, resveratrol in peanut (Arachis hypogaea) and peanut products: A mini review. Rev. Environ. Sci. Biotechnol. 2013;12:209–221. doi: 10.1007/s11157-012-9294-7. [DOI] [Google Scholar]
- 41.Han X., Shen T., Lou H. Dietary polyphenols and their biological significance. Int. J. Mol. Sci. 2007;8:950–988. doi: 10.3390/i8090950. [DOI] [Google Scholar]
- 42.Wallace K.L., Zheng L.-B., Kanazawa Y., Shih D.Q. Immunopathology of inflammatory bowel disease. World J. Gastroenterol. 2014;20:6–21. doi: 10.3748/wjg.v20.i1.6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Al Mijiam M., Lim B.O. Diets, functional foods, and nutraceuticals as alternative therapies for inflammatory bowel disease: Present status and future trends. World J. Gastroenterol. 2018;24:2673–2685. doi: 10.3748/wjg.v24.i25.2673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kaulmann A., Bohn T. Bioactivity of polyphenols: Preventive and adjuvant strategies toward reducing inflammatory bowel disease—Promises, perspectives, and pitfalls. Ox. Med. Cell Longev. 2016;9346470:1–29. doi: 10.1155/2016/9346470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Scalbert A., Johnson I.T., Saltmarsh M. Polyphenols: Antioxidants and beyond. Am. J. Clin. Nutr. 2005;81:215S–217S. doi: 10.1093/ajcn/81.1.215S. [DOI] [PubMed] [Google Scholar]
- 46.Scarano A., Santino A. The plant polyphenol metabolism as functional architecture and its nutritional exploitation. Nutr. Food Sci. Int. J. 2019;8:26–30. doi: 10.19081/NFSIJ.2019.08.555731. [DOI] [Google Scholar]
- 47.Martínez V., Mitjans M., Vinardell M.P. Cytoprotective effects of polyphenols against oxidative damage. In: Watson R.R., Preedy V.R., Sherma Z., editors. Polyphenols in Human Health and Disease. Academic Press; Cambridge, MA, USA: 2014. pp. 275–288. [DOI] [Google Scholar]
- 48.Xu D., Hu M.-J., Wang Y.-Q., Cui Y.-L. Antioxidant activities of quercetin and its complexes for medicinal application. Molecules. 2019;24:1123. doi: 10.3390/molecules24061123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Moskaug J.Ø., Carlsen H., Myhrstad M.C.W., Blomhoff R. Polyphenols and glutathione synthesis regulation. Am. J. Clin. Nutr. 2005;81:277S–283S. doi: 10.1093/ajcn/81.1.277S. [DOI] [PubMed] [Google Scholar]
- 50.Espley R.V., Butts C.A., Laing W.A., Martell S., Smith H., McGhie T.K., Zhang J., Paturi G., Hedderley D., Bovy A., et al. Dietary flavonoids from modified apple reduce inflammation markers and modulate gut microbiota in mice. J. Nutr. 2014;144:146–154. doi: 10.3945/jn.113.182659. [DOI] [PubMed] [Google Scholar]
- 51.Cavalcanti E., Vadrucci E., Delvecchio F.R., Addabbo F., Bettini S., Liou R., Monsurrò V., Huang A.Y.C., Pizzaro T.T., Santino A., et al. Administration of reconstituted polyphenol oil bodies efficiently suppresses dendritic cell inflammatory pathways and acute intestinal inflammation. PLoS ONE. 2013;9:e88898. doi: 10.1371/journal.pone.0088898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Delvecchio F.R., Vadrucci E., Cavalcanti E., De Santis S., Vacca M., Myers J., Frederick A., Bianco G., Huang A., Monsurrò V., et al. Polyphenol Administration Impairs T cells Proliferation by Imprinting a Distinct Dendritic Cell Maturational Profile. Eur. J. Immunol. 2015;45:2638–2649. doi: 10.1002/eji.201545679. [DOI] [PubMed] [Google Scholar]
- 53.Larrosa M., Yanez-Gascon M.J., Selma V.M., González-Sarrías A., Toti S., Cerón J.J., Tomás-Barberán F., Dolara P., Espín J.C. Effect of a low dose of dietary resveratrol on colon microbiota, inflammation and tissue damage in a DSS-induced colitis rat model. J. Agric. Food Chem. 2009;57:2211–2220. doi: 10.1021/jf803638d. [DOI] [PubMed] [Google Scholar]
- 54.Monagas M., Khan N., Andrés-Lacueva C., Urpí-Sardá M., Vázquez-Agell M., Lamuela-Raventós R.M., Estruch R. Dihydroxylated phenolics acids derived from microbial metabolism reduce lipopolysaccharide-stimulated cytokine secretion by human peripheral blood mononuclear cells. Br. J. Nutr. 2009;102:201–206. doi: 10.1017/S0007114508162110. [DOI] [PubMed] [Google Scholar]
- 55.Wang D., Ho L., Faith J., Ono K., Janle E.M., Lachcik P.J., Cooper B.R., Jannasch A.H., D’Arcy B.R., Williams B.A., et al. Role of intestinal microbiota in the generation of polyphenol derived phenolic acid mediated attenuation of Alzheimer’s disease β-amiloid oligomerization. Mol. Nutr. Food Res. 2015;59:1025–1040. doi: 10.1002/mnfr.201400544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Santino A., Scarano A., De Santis S., De Benedictis M., Giovinazzo G., Chieppa M. Gut microbiota modulation and anti-inflammatory properties of dietary polyphenols in IBD: New and consolidated perspectives. Curr. Pharm. Des. 2017;23:2344–2351. doi: 10.2174/1381612823666170207145420. [DOI] [PubMed] [Google Scholar]
- 57.D’Introno A., Paradiso A., Scoditti E., D’Amico L., De Paolis A., Carluccio M.A., Nicoletti N., De Gara L., Santino A., Giovinazzo G. Antioxidant and anti-inflammatory properties of tomato fruit synthesizing different amounts of stilbenes. Plant Biotech. J. 2009;7:422–429. doi: 10.1111/j.1467-7652.2009.00409.x. [DOI] [PubMed] [Google Scholar]
- 58.Serra D., Paixão J., Nunes C., Dinis T.C.P., Almeida L.M. Cyanidin-3-glucoside suppresses cytokine-induced inflammatory response in human intestinal cells: Comparison with 5- aminosalicylic acid. PLoS ONE. 2013;8:e73001-10. doi: 10.1371/journal.pone.0073001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Karin M., Ben-Neriah Y. Phosphorylation meets ubiquitination: The control of NF-kappaB activity. Annu. Rev. Immunol. 2000;18:621–663. doi: 10.1146/annurev.immunol.18.1.621. [DOI] [PubMed] [Google Scholar]
- 60.Soobrattee M.A., Neergheen V.S., Luximon-Ramma A., Aruoma O.I., Bahorun T. Phenolics as potential antioxidant therapeutic agents: Mechanism and actions. Mutat. Res. Fundam. Mol. Mech. Mutagenesis. 2005;579:200–213. doi: 10.1016/j.mrfmmm.2005.03.023. [DOI] [PubMed] [Google Scholar]
- 61.Vendrame S., Klimis-Zacas D. Anti-inflammatory effect of anthocyanins via modulation of nuclear factor-kB and mitogen-activated protein kinase signaling cascades. Nutr. Rev. 2015;73:348–358. doi: 10.1093/nutrit/nuu066. [DOI] [PubMed] [Google Scholar]
- 62.Rajapakse A.G., Yepuri G., Carvas J.M., Stein S., Matter C.M., Scerri I., Ruffieux J., Montani J.-P., Ming X.-F., Yang Z. Hyperactive S6K1 mediates oxidative stress and endothelial dysfunction in aging: Inhibition by resveratrol. PLoS ONE. 2001;6:e19237. doi: 10.1371/journal.pone.0019237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Furuuchi R., Shimizu I., Yoshida Y., Hayashi Y., Ikegami R., Suda M., Katsuumi G., Wakasugi T., Nakao M., Minamino T. Boysenberry polyphenol inhibits endothelial dysfunction and improves vascular health. PLoS ONE. 2018;13:e0202051. doi: 10.1371/journal.pone.0202051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Calabriso N., Massaro M., Scoditti E., Pellegrino M., Ingrosso I., Giovinazzo G., Carluccio M.A. Red grape skin polyphenols blunt matrix metalloproteinase-2 and -9 activity and expression in cell models of vascular inflammation: Protective role in degenerative and inflammatory diseases. Molecules. 2016;21:1147. doi: 10.3390/molecules21091147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Calabriso N., Scoditti E., Massaro M., Pellegrino M., Storelli C., Ingrosso I., Giovinazzo G., Carluccio M.A. Multiple anti-inflammatory and anti-atherosclerotic properties of red wine polyphenolic extracts: Different role of hydroxycinnamic acids, flavonols and stilbenes on endothelial inflammatory gene expression. Eur. J. Nutr. 2016;55:477–489. doi: 10.1007/s00394-015-0865-6. [DOI] [PubMed] [Google Scholar]
- 66.Galleggiante V., De Santis S., Cavalcanti E., Scarano A., De Benedictis M., Serino G., Caruso M.L., Mastronardi M., Pinto A., Campiglia P., et al. Dendritic cells modulate iron homeostasis and inflammatory abilities following quercetin exposure. Curr. Pharm. Des. 2017;23:2139–2146. doi: 10.2174/1381612823666170112125355. [DOI] [PubMed] [Google Scholar]
- 67.Chieppa M., Giannelli G. Immune cells and microbiota response to iron starvation. Front. Med. 2018;5:109. doi: 10.3389/fmed.2018.00109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.De Santis S., Kunde D.A., Serino G., Galleggiante V., Caruso M.L., Mastronardi M., Cavalcanti E., Ranson N., Pinto A., Campiglia P., et al. Secretory leukoprotease inhibitor is required for efficient quercetin-mediated suppression of TNFα secretion. Oncotarget. 2016:7. doi: 10.18632/oncotarget.12415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.De Santis S., Galleggiante V., Scandiffo L., Liso M., Sommella E., Sobolewski A., Spilotro V., Pinto A., Campiglia P., Serino G., et al. Secretory leukoprotease inhibitor (Slpi) expression is required for educating murine dendritic cells inflammatory response following quercetin exposure. Nutrients. 2017;9:706. doi: 10.3390/nu9070706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Martin D.A., Bolling B.W. A review of the efficacy of dietary polyphenols in experimental models of inflammatory bowel diseases. Food Funct. 2015;6:1773–1786. doi: 10.1039/C5FO00202H. [DOI] [PubMed] [Google Scholar]
- 71.Piberger H., Oehme A., Hofmann C., Dreiseitel A., Sand P.G., Obermeier F., Schoelmerich J., Schreier P., Krammer G., Rogler G. Billberries and their anthocyanins ameliorate experimental colitis. Mol. Nutr. Food Res. 2011;55:1724–1729. doi: 10.1002/mnfr.201100380. [DOI] [PubMed] [Google Scholar]
- 72.Singh K., Jaggi A.S., Singh S. Exploring the ameliorative potential of Punica granatum in dextran sulfate sodium induced ulcerative colitis in mice. Phytother. Res. 2009;23:1565–1574. doi: 10.1002/ptr.2822. [DOI] [PubMed] [Google Scholar]
- 73.Scarano A., Butelli E., De Santis S., Cavalcanti E., Hill L., De Angelis M., Giovinazzo G., Chieppa M., Martin C., Santino A. Combined dietary anthocyanins, flavonols, and stilbenoids alleviate inflammatory bowel disease symptoms in mice. Front. Nutr. 2018;4:75. doi: 10.3389/fnut.2017.00075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Liso M., De Santis S., Scarano A., Verna G., Dicarlo M., Galleggiante V., Campiglia P., Mastronardi M., Lippolis A., Vacca M., et al. Bronze-tomatoes enriched diet affects the intestinal microbiome in homeostatic and inflammatory conditions. Nutrients. 2018;10:1862. doi: 10.3390/nu10121862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Cardona F., Andrés-Lacueva C., Tulipani S., Tinahones F.J., Queipo-Ortuño M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013;24:1415–1422. doi: 10.1016/j.jnutbio.2013.05.001. [DOI] [PubMed] [Google Scholar]
- 76.Liso M., De Santis S., Verna G., Dicarlo M., Calasso M., Santino A., Gigante I., Eri R., Raveenthiraraj S., Sobolewski A., et al. A specific mutation in Muc2 determines early dysbiosis in colitis-prone Winnie mice. Inflamm. Bowel Dis. 2019;26 doi: 10.1093/ibd/izz279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Singh A.K., Cabral C., Kumar R., Ganguly R., Rana H.K., Gupta A., Lauro M.R., Carbone C., Reis F., Pandey A.K. Beneficial effects of dietary polyphenols on gut microbiota and strategies to improve delivery efficiency. Nutrients. 2019;11:2216. doi: 10.3390/nu11092216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Lavefve L., Howard L.R., Carbonero F. Berry polyphenols metabolism and impact on human gut microbiota and health. Food Funct. 2020;11:45. doi: 10.1039/C9FO01634A. [DOI] [PubMed] [Google Scholar]
- 79.Vendrame S., Guglielmetti S., Riso P., Arioli S., Klimis-Zacas D., Porrini M. Six-week consumption of a wild blueberry powder drink increases bifidobacterial in the human gut. J. Agric. Food Chem. 2011;59:12815–12820. doi: 10.1021/jf2028686. [DOI] [PubMed] [Google Scholar]
- 80.Dolara P., Luceri C., De Filippo C., Femia A.P., Giovannelli L., Caderni G., Cecchini C., Silvi S., Orpianesi C., Cresci A. Red wine polyphenols influence carcinogenesis, intestinal microflora, oxidative damage and gene expression profiles of colonic mucosa in F344 rats. Mutat. Res. 2005;591:237–246. doi: 10.1016/j.mrfmmm.2005.04.022. [DOI] [PubMed] [Google Scholar]
- 81.Queipo-Ortuño M.I., Boto-Ordóñez M., Murri M., Gomez-Zumaquero J.M., Clemente-Postigo M., Estruch R., Cardona Diaz F., Andrés-Lacueva C., Tinahones F.J. Influence of red wine polyphenols and ethanol on the gut microbiota ecology and biochemical biomarkers. Am. J. Clin. Nutr. 2012;95:1323–1324. doi: 10.3945/ajcn.111.027847. [DOI] [PubMed] [Google Scholar]
- 82.Hidalgo M., Oruna-Concha M.J., Kolida S., Walton G.E., Kallithraka S., Spencer J.P.E., de Pascual-Teresa S. Metabolism of anthocyanins by human gut microflora and their influence on gut bacteria growth. J. Agric. Food Chem. 2012;60:3882–3890. doi: 10.1021/jf3002153. [DOI] [PubMed] [Google Scholar]
- 83.Ozdal T., Sela D.A., Xiao J., Boyacioglu D., Chen F., Capanoglu E. The reciprocal interactions between polyphenols and gut microbiota and effects on bioaccessibility. Nutrients. 2016;8:78. doi: 10.3390/nu8020078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Garbetta A., Nicassio L., D’Antuono I., Cardinali A., Linsalata V., Attolico G., Minervini F. Influence of in vitro digestion process on polyphenolic profile of skin grape (cv. Italia) and on antioxidant activity in basal or stressed conditions of human intestinal cell line (HT-29) Food Res. Int. 2018;106:878–884. doi: 10.1016/j.foodres.2018.01.072. [DOI] [PubMed] [Google Scholar]
- 85.Graziani G., D’Argenio G., Tuccillo C., Loguercio C., Ritieni A., Morisco F., Del Vecchio Blanco C., Fogliano V., Romano M. Apple polyphenol extracts prevent damage to human gastric epithelial cells in vitro and to rat gastric mucosa in vivo. Gut. 2005;54:193–200. doi: 10.1136/gut.2004.046292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Schaefer S., Baum M., Eisenbrand G., Dietrich H., Will F., Janzowki C. Polyphenolic apple juice extracts and their major constituents reduce oxidative damage in human colon cell lines. Mol. Nutr. Food Res. 2006;50:24–33. doi: 10.1002/mnfr.200500136. [DOI] [PubMed] [Google Scholar]
- 87.Chedea V.S., Palade L.M., Marin D.E., Pelmus R.S., Habeanu M., Rotar M.C., Gras M.A., Pistol G.C., Taranu I. Intestinal absorption and antioxidant activity of grape pomace polyphenols. Nutrients. 2018;10:588. doi: 10.3390/nu10050588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Seeram N.P., Adams L.S., Henning S.M., Niu Y., Zhang Y., Nair M.G., Heber D. In vitro antiproliferative, apoptotic and antioxidant activities of punicalagin, ellagic acid and a total pomegranate tannin extract are enhanced in combination with other polyphenols as found in pomegranate juice. J. Nutr. Biochem. 2005;16:360–367. doi: 10.1016/j.jnutbio.2005.01.006. [DOI] [PubMed] [Google Scholar]
- 89.Bornsek S.M., Ziberna L., Polak T., Vanzo A., Ulrih N.P., Abram V., Tramer F., Passamonti S. Bilberry and blueberry anthocyanins act as powerful intracellular antioxidants in mammalian cells. Food Chem. 2012;134:1878–1884. doi: 10.1016/j.foodchem.2012.03.092. [DOI] [PubMed] [Google Scholar]
- 90.Nunes C., Ferreira E., Freitas V., Almeida L., Barbosa R.M., Laranjinha J. Intestinal anti-inflammatory activity of red wine extract: Unveiling the mechanisms in colonic epithelial cells. Food Funct. 2013;4:373–383. doi: 10.1039/C2FO30233K. [DOI] [PubMed] [Google Scholar]
- 91.Gessner D.K., Ringseis R., Siebers M., Keller J., Kloster J., Wen G., Eder K. Inhibition of the pro-inflammatory NF-kB pathway by a grape seed and grape marc meal extract in intestinal epithelial cells. J. Anim. Physiol. Anim. Nutr. 2012;96:1074–1083. doi: 10.1111/j.1439-0396.2011.01222.x. [DOI] [PubMed] [Google Scholar]
- 92.Denis M.C., Furtos A., Dudonné S., Montoudis A., Garofalo C., Desjardins Y., Delvin E., Levy E. Apple peel polyphenols and their beneficial actions on oxidative stress and inflammation. PLoS ONE. 2013;8:e53725. doi: 10.1371/journal.pone.0053725. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 93.Tomlinson M.L., Butelli E., Martin C., Carding S.R. Flavonoids from engineered tomatoes inhibit gut barrier pro-inflammatory cytokines and chemokines, via SAPK/JNK and p38 MAPK pathways. Front. Nutr. 2017;4:61. doi: 10.3389/fnut.2017.00061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Oz H.S., Chen T.S., McClain C.J., de Villiers W.J. Antioxidants as novel therapy in a murine model of colitis. J. Nutr. Biochem. 2005;16:27–304. doi: 10.1016/j.jnutbio.2004.09.007. [DOI] [PubMed] [Google Scholar]
- 95.Alvarez-Suarez J.M., Dekanski D., Ristic S., Radonjic N.V., Petronijevic N.D., Giampieri F., Astolfi P., Gonzalez-Paramas A.M., Santos-Buelga C., Tulipani S., et al. Strawberry polyphenols attenuate ethanol-induced gastric lesions in rats by activation of antioxidant enzymes and attenuation of MDA increase. PLoS ONE. 2011;6:e25878. doi: 10.1371/journal.pone.0025878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Skyberg J.A., Robison A., Golden S., Rollins M.F., Callis G., Huarte E., Kochetkova I., Jutila M.A., Pascual D.W. Apple polyphenols require T cells to ameliorate dextran sulfate sodium-induced colitis and dampen proinflammatory cytokine expression. J. Leucoc. Biol. 2011;90:1043–1054. doi: 10.1189/jlb.0311168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Brückner M., Westphal S., Domschke W., Kucharzik T., Lugering A. Green tea polyphenol epigallocatechin-3-gallate shows therapeutic antioxidative effects in a murine model of colitis. J. Chrons. Colitis. 2012;6:226–235. doi: 10.1016/j.crohns.2011.08.012. [DOI] [PubMed] [Google Scholar]
- 98.Paiotti A.P., Neto R.A., Marchi P., Silva R.M., Pazine V.L., Noguti J., Pastrelo M.M., Gollucke A.P., Miszputen S.J., Ribeiro D.A. The anti-inflammatory potential of phenolic compounds in grape juice concentrate (G8000TM) on 2,4,6-trinitrobenzene sulphonic acid-induced colitis. Br. J. Nutr. 2013;110:973–980. doi: 10.1017/S000711451300007X. [DOI] [PubMed] [Google Scholar]
- 99.Boussenna A., Cholet J., Goncalves-Mendes N., Joubert-Zakeyh J., Fraisse D., Vasson M.P., Texier O., Felgines C. Polyphenol-rich grape pomace extracts protect against dextran sulfate sodium-induced colitis in rats. J. Sci. Food Agric. 2016;96:1260–1268. doi: 10.1002/jsfa.7214. [DOI] [PubMed] [Google Scholar]
- 100.Saadatdoust Z., Pandurangan A.K., Ananda Sadagopan S.K., Esa N.M., Ismail A., Mustafa M.R. Dietary cocoa inhibits colitis associated cancer: A crucial involvement of the IL-6/STAT3 pathway. J. Nutr. Biochem. 2015;26:1547–1558. doi: 10.1016/j.jnutbio.2015.07.024. [DOI] [PubMed] [Google Scholar]
- 101.Del Bo’ C., Bernardi S., Marino M., Porrini M., Tucci M., Guglielmetti S., Cherubini A., Carrieri B., Kirkup B., Kroon P., et al. Systematic review on polyphenol intake and health outcomes: Is there sufficient evidence to define a health-promoting polyphenol-rich dietary pattern? Nutrients. 2019;11:1355. doi: 10.3390/nu11061355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Garg M., Sharma N., Sharma S., Kapoor P., Kumar A., Chunduri V., Arora P. Biofortified Crops Generated by Breeding, Agronomy, and Transgenic Approaches Are Improving Lives of Millions of People around the World. Front. Nutr. 2018;5:12. doi: 10.3389/fnut.2018.00012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Zhu C., Naqvi S., Gomez-Galera S., Pelacho A.M., Capell T., Christou P. Transgenic strategies for the nutritional enhancement of plants. Trends Plant Sci. 2007;12:1360S–1385S. doi: 10.1016/j.tplants.2007.09.007. [DOI] [PubMed] [Google Scholar]
- 104.Zhu C., Sanahuja G., Yuan D., Farré G., Arjó G., Berman J., Zorilla-López U., Banakar R., Bai C., Pérez-Massot E., et al. Biofortification of plants with altered antioxidant content and composition: Genetic engineering strategies. Plant Biotechnol. J. 2013;11:129–141. doi: 10.1111/j.1467-7652.2012.00740.x. [DOI] [PubMed] [Google Scholar]
- 105.Martin C., Li J. Medicine is not health care, food is health care: Plant metabolic engineering, diet and human health. New Phytol. 2017;216:699–719. doi: 10.1111/nph.14730. [DOI] [PubMed] [Google Scholar]
- 106.Nguyen P.M., Niemeyer E.D. Effects of nitrogen fertilization on the phenolic composition and antioxidant properties of basil (Ocinum basilicum L.) J. Agric. Food Chem. 2008;56:8685–8691. doi: 10.1021/jf801485u. [DOI] [PubMed] [Google Scholar]
- 107.Fortier E., Desjardins Y., Tremblay N., Bèlec C., Côtè M. Influence of irrigation and nitrogen fertilization on broccoli polyphenolics concentration. Acta Hortic. 2010;856:55–62. doi: 10.17660/ActaHortic.2010.856.6. [DOI] [Google Scholar]
- 108.Singh A.P., Luthria D., Wilson T., Vorsa N., Singh V., Banuelos G., Pasakdee S. Polyphenols content and antioxidant capacity of eggplant pulp. Food Chem. 2009;114:955–961. doi: 10.1016/j.foodchem.2008.10.048. [DOI] [Google Scholar]
- 109.Sousa C., Valentão P., Rangel J., Pereira J.A., Ferreres F., Seambra R.M., Andrade P.B. Influence of two fertilization regimens on the amounts of organic acids and phenolic compounds of Tronchuda Cabbage (Brassica oleracea L. Var. costata DC) J. Agric. Food Chem. 2005;53:9128–9132. doi: 10.1021/jf051445f. [DOI] [PubMed] [Google Scholar]
- 110.Heimler D., Vignolini P., Arfaioli P., Isolani L., Romani A. Conventional, organic and biodynamic farming: Differences in polyphenol content and antioxidant activity of Batavia lettuce. J. Sci. Food Agric. 2012;92:551–556. doi: 10.1002/jsfa.4605. [DOI] [PubMed] [Google Scholar]
- 111.Heimler D., Romani A., Ieri F. Plant polyphenol content, soil fertilization and agricultural management: A review. Eur. Food Res. Technol. 2017;243:1107–1115. doi: 10.1007/s00217-016-2826-6. [DOI] [Google Scholar]
- 112.Stefanson A.L., Tsao R., Liu R., Duizer L., Bakovic M., Martin R.C. Effect of variety, soil fertility status and agronomic treatments on carrot mineral and phytochemical composition and consumer liking of flavor traits. J. Sci. Food Agric. 2019;99:5457–5474. doi: 10.1002/jsfa.9807. [DOI] [PubMed] [Google Scholar]
- 113.Hilbert G., Soyer J.P., Molot C., Giraudon J., Milin S., Gaudillere J.P. Effects of nitrogen supply on must quality and anthocyanin accumulation in berries of cv. Merlot. Vitis. 2003;42:69–76. [Google Scholar]
- 114.Wang Y., Zhou B., Sun M., Li Y., Kawabata S. UV-A Light Induces Anthocyanin Biosynthesis in a Manner Distinct from Synergistic Blue + UV-B Light and UV-A/Blue Light Responses in Different Parts of the Hypocotyls in Turnip Seedlings. Plant Cell Physiol. 2012;53:1470–1480. doi: 10.1093/pcp/pcs088. [DOI] [PubMed] [Google Scholar]
- 115.Li W., Tan L., Zou Y., Tan X., Huang J., Chen E., Tang Q. The effects of ultraviolet A/b treatments on anthocyanins accumulation and gene expression in dark-purple tea cultivar ‘Ziyan’ (Camellia sinensis) Molecules. 2020;25:354. doi: 10.3390/molecules25020354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Goto E., Hayashi K., Furuyama S., Hikosaka S., Ishigami Y. Effect of UV light on phytochemical accumulation and expression of anthocyanin biosynthesis genes in red leaf lettuce. Acta Hortic. 2016;1134:179–186. doi: 10.17660/ActaHortic.2016.1134.24. [DOI] [Google Scholar]
- 117.Kaushik P., Andújar I., Vilanova S., Plazas M., Gramazio P., Herraiz F.J., Brar N.S., Prohens J. Breeding vegetables with increased content in bioactive phenolic acids. Molecules. 2015;20:18464–18481. doi: 10.3390/molecules201018464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Mazzucato A., Willems D., Bernini R., Picarella M.E., Santangelo E., Ruiu F., Tilesi F., Soressi G.P. Novel phenotype related to the breeding of purple-fruited tomatoes and effect of peel extracts on human cancer cell proliferation. Plant Physiol. Biochem. 2013;72:125–133. doi: 10.1016/j.plaphy.2013.05.012. [DOI] [PubMed] [Google Scholar]
- 119.Blando F., Berland H., Maiorano G., Durante M., Mazzucato A., Picarella M.E., Nicoletti I., Gerardi C., Mita G., Andersen Ø.M. Nutraceutical characterization of anthocyanin-rich fruits produced by “Sun Black” tomato line. Front. Nutr. 2019;6:133. doi: 10.3389/fnut.2019.00133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Mennella G., Rotino G.L., Fibiani M., D’Alessandro A., Francese G., Toppino L., Cavallanti F., Acciarri N., Lo Scalzo R. Characterization of Health-Related Compounds in Eggplant (Solanum Melongena L.) Lines Derived From Introgression of Allied Species. J. Agric. Food Chem. 2010;58:7597–7603. doi: 10.1021/jf101004z. [DOI] [PubMed] [Google Scholar]
- 121.Scarano A., Chieppa M., Santino A. Looking at flavonoid biodiversity in horticultural crops: A colored mine with nutritional benefits. Plants. 2018;7:98. doi: 10.3390/plants7040098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Muir S.R., Collins G.J., Robinson S., Hughes S., Bovy A., Ric De Vos C.H., van Tunen A.J., Verhoeyen M.E. Overexpression of petunia chalcone isomerase in tomato results in fruit containing increased levels of flavonols. Nat. Biotechnol. 2001;19:470–474. doi: 10.1038/88150. [DOI] [PubMed] [Google Scholar]
- 123.Giovinazzo G., D’Amico L., Paradiso A., Bollini R., Sparvoli F., De Gara L. Antioxidant metabolite profiles in tomato fruit constitutively expressing the grapevine stilbene synthase gene. Plant Biotech. J. 2005;3:57–69. doi: 10.1111/j.1467-7652.2004.00099.x. [DOI] [PubMed] [Google Scholar]
- 124.Wei Q., Wang Q.Y., Feng Z.-H., Wang B., Zhang Y.-F., Yang Q. Increased accumulation of anthocyanins in transgenic potato tubers by overexpressing the 3GT gene. Plant Biotechnol. Rep. 2012;6:69–75. doi: 10.1007/s11816-011-0201-4. [DOI] [Google Scholar]
- 125.Niggeweg R., Michael A.J., Martin C. Engineering plants with increased levels of the antioxidant chlorogenic acid. Nat. Biotech. 2004;22:746–754. doi: 10.1038/nbt966. [DOI] [PubMed] [Google Scholar]
- 126.Espley R.V., Hellens R.P., Putterill J., Stevenson D.E., Kutty-Amma S., Allan A.C. Red colouration in apple fruit is due to the activity of the MYB transcription factor, MdMYB10. Plant J. 2007;49:414–427. doi: 10.1111/j.1365-313X.2006.02964.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Luo J., Butelli E., Hill L., Parr A., Niggeweg R., Bailey P., Weisshaar B., Martin C. AtMYB12 regulates caffeoyl quinic acid and flavonol synthesis in tomato: Expression in fruit results in very high levels of both types of polyphenol. Plant J. 2008;56:316–326. doi: 10.1111/j.1365-313X.2008.03597.x. [DOI] [PubMed] [Google Scholar]
- 128.Butelli E., Titta L., Giorgio M., Mock H.P., Peterek S., Schijlen E.G., Hall R.D., Bovy A.G., Luo J., Martin C. Enrichment of tomato fruit with health-promoting anthocyanins by expression of selected transcription factors. Nat. Biotechnol. 2008;26:1301–1308. doi: 10.1038/nbt.1506. [DOI] [PubMed] [Google Scholar]
- 129.Jung C.S., Griffiths H.M., De Jong D.M., Cheng S., Bodis M., Kim T.S., De Jong W.S. The potato developer (D) locus encodes an R2R3 MYB transcription factor that regulates expression of multiple anthocyanin structural genes in tuber skin. Theor. Appl. Genet. 2009;120:45–57. doi: 10.1007/s00122-009-1158-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Zhang Y., Butelli E., Alseekh S., Tohge T., Rallapalli G., Luo J., Kawar P.G., Hill L., Santino A., Fernie A.R., et al. Multi-level engineering facilitates the production of phenylpropanoid compounds in tomato. Nat. Commun. 2015;6:8635. doi: 10.1038/ncomms9635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Xu J., Hua K., Lang Z. Genome editing for horticultural crop improvement. Hortic. Res. 2019;6:113. doi: 10.1038/s41438-019-0196-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Van Eck J. Applying gene editing to tailor precise genetic modifications in plants. J. Biol. Chem. 2020;295:13267–13276. doi: 10.1074/jbc.REV120.010850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Sedeek K.E.M., Mahas A., Mahfouz M. Plant genome engineering for targeted improvement of crop traits. Front. Plant Sci. 2019;10:114. doi: 10.3389/fpls.2019.00114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Kamburova V.S., Nikitina E.V., Shermatov S.E., Buriev Z.T., Kumpatla S.P., Emani C., Abdurakhmonov I.Y. Genome editing in plants: An overview of tools and applications. Int. J. Agron. 2017;2017:1–15. doi: 10.1155/2017/7315351. [DOI] [Google Scholar]
- 135.Zhang Y., Massel K., Godwin I.D., Gao C. Applications and potential of genome editing in crop improvement. Genome Biol. 2018;19:210. doi: 10.1186/s13059-018-1586-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Chen K., Wang Y., Zhang R., Zhang H., Gao C. CRISPR/Cas9 genome editing and precision plant breeding in agriculture. Ann. Rev. Plant Biol. 2019;70:667–697. doi: 10.1146/annurev-arplant-050718-100049. [DOI] [PubMed] [Google Scholar]
- 137.Khatodia S., Bhatotia K., Passricha N., Khurana S.M.P., Tuteja N. The CRISPR/Cas genome-editing tool: Application in improvement of crops. Front. Plant Sci. 2016;7:506. doi: 10.3389/fpls.2016.00506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Wang T., Zhang H., Zhu H. CRISPR technology is revolutioning the improvement of tomato and other fruit crops. Hortic. Res. 2019;6:77. doi: 10.1038/s41438-019-0159-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.El-Mounadi K., Morales-Floriano M.L., Garcia-Ruiz H. Principles, applications, and biosafety of plant genome editing using CRISPR/Cas9. Front. Plant Sci. 2020;11:56. doi: 10.3389/fpls.2020.00056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Vats S., Kumawat S., Kumar V., Patil G.B., Joshi T., Sonah H., Sharma T.R., Deshmukh R. Genome editing in plants: Exploration of technological advancements and challenges. Cells. 2019;8:1386. doi: 10.3390/cells8111386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Moradpour M., Abulah S.N.A. CRISPR/dCas9 platforms in plants: Strategies and applications beyond genome editing. Plant Biotechnol. J. 2020;18:32–44. doi: 10.1111/pbi.13232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Manghwar H., Lindsey K., Zhang X., Jin S. CRISPR/Cas System: Recent advances and future prospects for genome editing. Trends Plant Sci. 2019;24:1102–1125. doi: 10.1016/j.tplants.2019.09.006. [DOI] [PubMed] [Google Scholar]
- 143.Abudayyeh O.O., Gootenberg J.S., Essletzbichler P., Han S., Joung J., Belanto J.J., Verdine V., Cox D.B.T., Kellner M.J., Regev A., et al. RNA targeting with CRISPR-Cas13a. Nature. 2017;550:280–284. doi: 10.1038/nature24049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Ghogare R., Williamson-Benavides B., Ramírez-Torres F., Dhingra A. CRISPR-associated nucleases: The Dawn of a new age of efficient crop improvement. Transgenic Res. 2020;29:1–35. doi: 10.1007/s11248-019-00181-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Malnoy M., Viola R., Jung M.H., Koo O.-J., Kim S., Kim J.-S., Velasco R., Kanchiswamy C.K. DNA-free genetically edited grapevine and apple protoplast using CRISPR/Cas9 ribonucleoproteins. Front. Plant Sci. 2016;7:1904. doi: 10.3389/fpls.2016.01904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Clasen B.M., Stoddard T.J., Luo S., Demorest Z.L., Li J., Cedrone F., Tibebu R., Davison S., Ray E.E., Daulhac A., et al. Improving cold storage and processing traits in potato through targeted gene knockout. Plant Biotechnol. J. 2015;14:169–176. doi: 10.1111/pbi.12370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Shan Q., Zhang Y., Chen K., Zhang K., Gao C. Creation of fragrant rice by targeted knockout of the OsBADH2 gene using TALEN technology. Plant Biotechnol. J. 2015;13:791–800. doi: 10.1111/pbi.12312. [DOI] [PubMed] [Google Scholar]
- 148.Yasumoto S., Umemoto N., Lee H.J., Nakayasu M., Sawai S., Sakuma T., Yamamoto T., Mizutani M., Saito K., Muranaka T. Efficient genome engineering using Platinum TALEN in potato. Plant Biotechnol. Tokyo. 2019;36:167–173. doi: 10.5511/plantbiotechnology.19.0805a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Andersson M., Turesson H., Nicolia A., Fält A.-S., Samuelsson M., Hofvander P. Efficient targeted multiallelic mutagenesis in tetraploid potato (Solanum tuberosum) by transient CRISPR-Cas9 expression in protoplasts. Plant Cell Rep. 2017;36:117–128. doi: 10.1007/s00299-016-2062-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Čermák T., Baltes N.J., Čegan R., Zhang Y., Voytas D.F. High-frequency, precise modification of the tomato genome. Genome Biol. 2015;16:232. doi: 10.1186/s13059-015-0796-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Sanchez-Leon S., Gil-Humanes J., Ozuna C.V., Gimenez M.J., Sousa C., Voytas D.F., Barro F. Low-gluten, nontransgenic wheat engineered with CRISPR/Cas9. Plant Biotechnol. J. 2018;16:902–910. doi: 10.1111/pbi.12837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Jiang M., Liu Y., Liu Y., Tan Y., Huang J., Shu Q. Mutation of Inositol 1,3,4-trisphosphate 5/6-kinase6 Impairs Plant Growth and Phytic Acid Synthesis in Rice. Plants. 2019;8:114. doi: 10.3390/plants8050114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Camerlengo F., Frittelli A., Sparks C., Doherty A., Martignago D., Larrè C., Lupi R., Sestili F., Masci S. CRISPR-Cas9 Multiplex Editing of the α-Amylase/Trypsin Inhibitor Genes to Reduce Allergen Proteins in Durum Wheat. Front. Sustain. Food Syst. 2020;4:104. doi: 10.3389/fsufs.2020.00104. [DOI] [Google Scholar]
- 154.Jiang W.Z., Henry I.M., Lynagh P.G., Comai L., Cahoon E.B., Weeks D.P. Significant enhancement of fatty acid composition in seeds of the allohexaploid, Camelina sativa, using CRISPR/Cas9 gene editing. Plant Biotechnol. J. 2017;15:648–657. doi: 10.1111/pbi.12663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Morineau C., Bellec Y., Tellier F., Gissot L., Kelemen Z., Nogué F., Faure J.-D. Selective gene dosage by CRISPRCas9 genome editing in hexaploid Camelina sativa. Plant Biotechnol. J. 2017;15:729–739. doi: 10.1111/pbi.12671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Okuzaki A., Ogawa T., Koizuka C., Kaneko K., Inaba M., Imamura J., Koizuka N. CRISPR/Cas9-mediated genome editing of the fatty acid desaturase 2 gene in Brassica napus. Plant Physiol. Biochem. 2018;131:63–69. doi: 10.1016/j.plaphy.2018.04.025. [DOI] [PubMed] [Google Scholar]
- 157.Li R., Li R., Li X., Fu D., Zhu B., Tian H., Luo Y., Zhu H. Multiplexed CRISPR/Cas9-mediated metabolic engineering of γ-aminobutyric acid levels in Solanum lycopersicum. Plant Biotechnol. J. 2018;16:415–427. doi: 10.1111/pbi.12781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Nonaka S., Arai C., Takayama M., Matsukura C., Ezura H. Efficient increase of γ-aminobutyric acid (GABA) content in tomato fruits by targeted mutagenesis. Sci. Rep. 2017;7:7057. doi: 10.1038/s41598-017-06400-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Li X., Wang Y., Chen S., Tian H., Fu D., Zhu B., Luo Y., Zhu H. Lycopene is enriched in tomato fruit by CRISPR/Cas9-mediated multiplex genome editing. Front. Plant Sci. 2018;9:559. doi: 10.3389/fpls.2018.00559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Watanabe K., Kobayashi A., Endo M., Sage-Ono K., Toki S., Ono M. CRISPR/Cas9-mediated mutagenesis of the dihydroflavonol-4-reductase-B (DFR-B) locus in the Japanese morning glory Ipomea (Pharbitis) nil. Sci. Rep. 2017;7:10028. doi: 10.1038/s41598-017-10715-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Nishihara M., Higuchi A., Watanabe A., Tasaki K. Application of the CRISPR/Cas9 system for modification of flower color in Torenia fournieri. BMC Plant Biol. 2018;18:331. doi: 10.1186/s12870-018-1539-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Tasaki K., Yoshida M., Nakajima M., Higuchi A., Watanabe A., Nishihara M. Molecular characterization of an anthocyanin-related glutathione S-transferase gene in Japanese gentian with CRISPR/Cas9 system. BMC Plant Biol. 2020;20:370. doi: 10.1186/s12870-020-02565-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Klimek-Chodacka M., Oleszkiewicz T., Lowder L.G., Qi Y., Baranski R. Efficient CRISPR/Cas9-based genome editing in carrot cells. Plant Cell Rep. 2018;37:575–586. doi: 10.1007/s00299-018-2252-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Yu J., Tu L., Subburaj S., Bae S., Lee G.-J. Simultaneous targeting of duplicated genes in Petunia protoplast for flower color modification via CRISPR/Cas9 ribonucleoproteins. Plant Cell Rep. 2020 doi: 10.1007/s00299-020-02593-1. [DOI] [PubMed] [Google Scholar]
- 165.Jung Y.J., Lee H.J., Kim J.H., Kim D.H., Kim D.H., Kim H.K., Cho Y.-G., Bae S., Kang K.K. CRISPR/Cas9-targeted mutagenesis of F3′H, DFR and LDOX, genes related to anthocyanin biosynthesis in black rice (Oryza sativa L.) Plant Biotechnol. Rep. 2019;13:521–531. doi: 10.1007/s11816-019-00579-4. [DOI] [Google Scholar]
- 166.Deng L., Wang H., Sun C., Li Q., Jiang H., Minmin D., Li C.-B., Li C. Efficient generation of pink-fruited tomatoes using CRISPR/Cas9 system. J. Gen. Genom. 2017;45:51. doi: 10.1016/j.jgg.2017.10.002. [DOI] [PubMed] [Google Scholar]
- 167.Zhi J., Liu X., Li D., Huang Y., Yan S., Cao B., Qiu Z. CRISPR/Cas9-mediated SlAN2 mutants reveal various regulatory models of anthocyanin biosynthesis in tomato plant. Plant Cell Rep. 2020;39:799–809. doi: 10.1007/s00299-020-02531-1. [DOI] [PubMed] [Google Scholar]
- 168.Zhai Y., Yu K., Cai S., Hu L., Amoo O., Xu L., Yang Y., Ma B., Jiao Y., Zhang C., et al. Targeted mutagenesis of BnTT8 homologs controls yellow seed coat development for effective oil production in Brassica napus L. Plant Biotechnol. J. 2020;18:1153–1168. doi: 10.1111/pbi.13281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Van Vu T., Sivankalyani V., Kim E.-J., Doan D.T.H., Tran M.T., Kim J., Sung Y.W., Park M., Kang Y.J., Kim J.-Y. Highly efficient homology-directed repair using CRISPR/Cpf1-geminiviral replicon in tomato. Plant Biotechnol. J. 2020;18:2133–2143. doi: 10.1111/pbi.13373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Danilo B., Perrot L., Botton E., Nogué F., Mazier M. The DFR locus: A smart landing pad for targeted transgene insertion in tomato. PLoS ONE. 2018;13:e0208395. doi: 10.1371/journal.pone.0208395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Waltz E. Gene-edited CRISPR mushroom escapes US regulation. Nature. 2016;532:293. doi: 10.1038/nature.2016.19754. [DOI] [PubMed] [Google Scholar]
- 172.González M.N., Massa G.A., Andersson M., Turesson H., Olsson N., Fält A.-S., Storani L., Oneto C.A.D., Hofvander P., Feingold S.E. Reduced Enzymatic Browning in Potato Tubers by Specific Editing of a Polyphenol Oxidase Gene via Ribonucleoprotein Complexes Delivery of the CRISPR/Cas9 System. Front. Plant Sci. 2020;10:1649. doi: 10.3389/fpls.2019.01649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Zhou Z., Tan H., Li Q., Chen J., Gao S., Wang Y., Chen W., Zhang L. CRISPR/Cas9-mediated efficient targeted mutagenesis of RAS in Salvia miltiorrhiza. Phytochemistry. 2018;148:63–70. doi: 10.1016/j.phytochem.2018.01.015. [DOI] [PubMed] [Google Scholar]
- 174.Chang L., Wu S., Tian L. Effective genome editing and identification of a regiospecific gallic acid 4-O-glycosyltransferase in pomegranate (Punica granatum L.) Hortic. Res. 2019;6:123. doi: 10.1038/s41438-019-0206-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Sunitha S., Rock C. CRISPR/Cas9-mediated targeted mutagenesis of TAS4 and MYBA7 loci in grapevine rootstock 101-14. Transgenic Res. 2020;29:355–367. doi: 10.1007/s11248-020-00196-w. [DOI] [PMC free article] [PubMed] [Google Scholar]