

Abstract
Alzheimer’s disease (AD), Parkinson’s disease (PD), and amyotrophic lateral sclerosis (ALS) are neurodegenerative disorders characterized by a progressive degeneration of the central or peripheral nervous systems. A central role of the RNA metabolism has emerged in these diseases, concerning mRNAs processing and non-coding RNAs biogenesis. We aimed to identify possible common grounds or differences in the dysregulated pathways of AD, PD, and ALS. To do so, we performed RNA-seq analysis to investigate the deregulation of both coding and long non-coding RNAs (lncRNAs) in ALS, AD, and PD patients and controls (CTRL) in peripheral blood mononuclear cells (PBMCs). A total of 293 differentially expressed (DE) lncRNAs and 87 mRNAs were found in ALS patients. In AD patients a total of 23 DE genes emerged, 19 protein coding genes and four lncRNAs. Through Kyoto Encyclopedia of Genes and Genomes (KEGG) and Gene Ontology (GO) analyses, we found common affected pathways and biological processes in ALS and AD. In PD patients only five genes were found to be DE. Our data brought to light the importance of lncRNAs and mRNAs regulation in three principal neurodegenerative disorders, offering starting points for new investigations on deregulated pathogenic mechanisms.
Keywords: Alzheimer’s disease, Parkinson’s disease, amyotrophic lateral sclerosis, gene expression, long non-coding RNAs
1. Introduction
Aging-related neurodegenerative diseases, such as Alzheimer’s disease (AD), Parkinson’s disease (PD), and amyotrophic lateral sclerosis (ALS), among others, are characterized by the progressive degeneration of the structure and function of the central or peripheral nervous systems [1]. The onset of these diseases is growing as a consequence of the increased aging population [2] and it is thus necessary to fully decipher the molecular bases underlining these pathological conditions [3]. The progressive loss of motor neurons (MNs) is the primary feature of ALS, however its etiology remains unclear. ALS can be either familial (~10% of cases) or sporadic, and MN degeneration may be caused by common pathways leading to identical clinical appearance. Among the pathological hallmarks of this disease, of great relevance is the nuclear deficiency of RNA-binding proteins (RBPs) such as TDP-43 and FUS [4]. Amyloid plaques and neurofibrillary tangles (NFTs) are the main features of AD, along with other molecular alterations concerning neuropil threading, dystrophic neurites, neuroinflammation, and cerebral amyloid angiopathy [5]. The misfunctioning of these processes leads to neurodegeneration and thus macroscopic atrophy of the CNS. PD is a neurodegenerative disease characterized by the degeneration of dopaminergic neurons in the substantia nigra pars compacta (SNpc), which ultimately leads to impaired movements control [6].
Several common features of neurodegeneration have been studied and associated to the molecular pathogenesis of AD, PD, and ALS, and these include neuroinflammation, autophagy, and oxidative stress [7,8]. Moreover, pathogenic mutations, amyloid precursor proteins, superoxide dismutase, and DNA and RNA binding proteins are important contributing factors to these neurodegenerative diseases [9]. Although many common features have been found and studied, other emergent mechanisms remain unclear. The newly discovered role of RNA metabolism in these disorders is the object of recent studies [10] and still needs further investigation. Interestingly, recent evidences suggest that there are some common molecular events happening during the development and progression of these neurodegenerative disorders, which could be specifically driven by RNA perturbation [11]. Indeed, the disruption of RNA stability may play an important role in these pathologies. Alterations in RNA turnover have been identified in several disease pathways, including RNA sequestration in stress granules or foci, RNA transport, exosomes, alternative splicing, and retrotransposons [12]. As the importance of RNA metabolism is gaining more and more relevance, the role of both small and long noncoding RNAs has also been investigated. MicroRNAs directly interact with partially complementary target sites located in the 3′ UTR of target mRNAs and modulate their expression by repressing their translation into proteins [13]. Their involvement in the central nervous system (CNS) homeostasis, and their dysfunction in neurodegenerative disease are objects of intense study [14,15]. Lately, long noncoding RNAs were also found to be involved in neurodegenerative-related processes. For example, the antisense NEAT1_2 was reported to be upregulated in conjunction with increased paraspeckle formation in the spinal motor neurons of patients with ALS [16] and its overexpression in Huntington’s disease was described as a defensive mechanism against cell injury [17]. Its deregulation was also displayed in PD, where its function is controversial. In fact, NEAT1_2 is reported to exert a protective role both when up-regulated or down-regulated. In the first case, it is described as a provider of drug-inducible neuroprotection from oxidative stress [18], and when down-regulated it seems to suppress disease’s progression [19]. Indeed, lncRNAs are just now starting to be characterized, as can be seen by the number of conflicting studies concerning their role in disease pathogenesis.
When focusing on the molecular pathogenesis of neurodegenerative diseases, the role of RNA metabolism has been the object of recent investigations [20,21,22], but many RNA driven mechanisms remain unknown. Indeed, further studies are needed in order to unravel the role of aberrant gene expression in sporadic ALS (SALS), AD, and PD. There is also currently no evidence in the literature which focuses on analyzing the common or divergent deregulated transcriptional profile in these three disorders. Thanks to RNA sequencing (RNA-seq) a huge variety of RNA species are detectable, including mRNA, non-coding RNA, transcript isoforms and splice variants [23]. Through a meta-analysis and validation of gene expression in different groups of patients, the identification of disease-associated genes that may play a significant role in several diseases’ pathophysiology is possible.
In this paper, we report a whole transcriptome profiling of both long non-coding and coding RNAs in peripheral blood mononuclear cells (PBMCs) of unmutated SALS) patients [24], unmutated sporadic AD patients, unmutated sporadic PD patients, and matched controls. PBMCs were chosen as they have been reported to be source of protein and nucleic acid biomarkers in neurodegenerative disorders, also allowing for their use as cellular models for these diseases [24,25,26,27]. After transcriptome analysis, we focused on the identification of possible crossroads or deviations in the dysregulated pathways of AD, PD, and ALS. Our results pointed out a different degree of RNA metabolism and dysregulation involvement in AD, PD, and ALS, highlighting a different role for lncRNAs and mRNAs regulation in the most relevant neurodegenerative disorders, offering an interesting starting point for future investigations on the pathogenic mechanisms involved.
2. Results
2.1. RNA-Seq Differentially Expressed mRNAs and lncRNAs
Throughout the whole work, coding and non-coding genes were considered differentially expressed and retained for further analysis when they presented a |log2(disease sample/healthy control)| ≥ 1 and a FDR ≤ 0.1. The number of DE genes found was different in the three groups, as was their biotype. A summary of these results is represented in Table 1. The first evidence is relative to the number of DE transcripts among the different diseases. In fact, in SALS the total number of affected genes is 380, while in AD it is 25 and in PD only five. It is also important to highlight that, in both SALS and PD, the majority of DE genes belongs to non-protein coding class, but in AD only four transcripts out of 23 do not encode for proteins. By analyzing the DE genes lists (Supplementary Table S1), none were shared amongst the three conditions, but further bioinformatics and literature searches allowed us to identify some common features that will be highlighted in Section 2.3.2.
Table 1.
Number of statistically significant differentially expressed mRNAs and lncRNAs in PBMCs from SALS, AD and PD patients, in terms of up-regulated transcripts, down-regulated transcripts and total.
| SALS | AD | PD | ||||
|---|---|---|---|---|---|---|
| mRNA | lncRNA | mRNA | lncRNA | mRNA | lncRNA | |
| Up-regulated | 57 | 183 | 8 | 3 | 0 | 1 |
| Down-regulated | 30 | 110 | 11 | 1 | 1 | 3 |
| Total | 87 | 293 | 19 | 4 | 1 | 4 |
2.1.1. Amyotrophic Lateral Sclerosis
In SALS patients, RNA-seq data reported 380 differentially expressed (DE) genes, 293 of which were lncRNAs (183 up-regulated and 110 down-regulated genes). Specifically, 184 out of 293 were reported as antisense, 81 out of 293 as lincRNAs, while the remaining 28 were classified as processed transcripts or intronic sense RNAs. Concerning coding genes, 87 differentially expressed mRNAs were identified, 30 of which were down-regulated whereas 57 were up-regulated. Heat-maps separately representing the expression levels of all dysregulated mRNAs and lncRNAs in SALS and healthy subjects are represented in Figure 1A,B. Different expression profiles in SALS and healthy controls can be visibly distinguished. The most deregulated coding genes were KIAA2013, HDAC1 and MYCBP. KIAA2013 is an uncharacterized protein and HDAC1 is an histone deacetylase whose aberrant expression has already been linked to ALS [28]. Interestingly, MYCBP is the binding protein of MYC, an important oncogene well characterized in cancer [29].
Figure 1.

Expression profiles of differently expressed genes in SALS, AD, PD and healthy controls. In panel (A), SALS differentially expressed mRNAs are shown, while in panel (B) SALS differentially expressed lncRNAs are shown. 10 sALS and 3 CTRLs were used for this analysis. In panel (C) and (D) both mRNAs and lncRNAs DE respectively in AD and PD are shown, given the lower amount of DE genes. For both AD and PD heatmaps, 6 patients’ samples and 6 CTRLs were used. All comparisons are given between the disease state and the control samples. We considered as differentially expressed only genes showing |log2(disease sample/healthy donor)| ≥ 1 and a False Discovery Rate ≤ 0.1.
When considering the top 10 of DE lncRNAs, our data showed an interesting deregulation of antisense (AS) RNAs related to genes involved in transcription regulation pathways such as ZEB1-AS and ZBTB11-AS. Also, XXbac-BPG252P9.10 is described as the antisense transcript of IER3, involved in transcription. Interestingly, some of the sense genes regulated by the DE AS lncRNAs in SALS are already linked to neurodegenerative disease, such as UBXN7-AS [30], ATG10-AS38 [31], and ADORA2A-AS [32].
2.1.2. Alzheimer’s Disease
In AD patients, a total of 23 DE genes has emerged, 19 of which were protein coding genes (eight up-regulated and 11 down-regulated) and four of which were non-coding genes, with three up-regulated lincRNAs and one down-regulated small nucleolar RNA. As the total number of DE in AD is limited, the heat-map (Figure 1C) shows the expression rate of both coding and non-coding genes in AD population versus matched healthy controls. Despite the low number of deregulated genes, there is a neat separation between the two groups, indicating a disease-specific gene expression pattern.
Amongst the most deregulated, we found coding genes already associated to AD such as TLN1 [33], and four nuclear pseudogenes (MTRNR2L6, MTRNR2L1, MTRNR2L10, and MTRNR2L8) of the mitochondrial MT-RNR2 gene [34]. VCL was also upregulated. This gene encodes for a protein involved in a pathway associated to Aβ toxicity [35] and associates to TLN1 [36].
When looking at the deregulated lncRNAs, three up-regulated lincRNAs were found: CH507-513H4.4, CH507-513H4.6, and CH507-513H4.3. These are novel transcripts, similar to YY1 associated myogenesis RNA 1 (YAM1), and they are reported as AD associated in LncRNADisease v2.0 Database [37].
2.1.3. Parkinson’s Disease
In PD patients, only five genes were found to be DE, four of which were down-regulated (one protein coding, one small nucleolar RNA, one sense intronic transcript, and one lincRNA) and one of which was an up-regulated lincRNA. The heat-map reporting DE mRNAs and lncRNAs is shown in Figure 1D. The protein coding gene TBC1D3 has been associated to generation of basal neural progenitors [38]. The lncRNAs found in PD patients are currently not correlated to specific pathways.
2.2. Validation of Deregulated Coding and Non-Coding Genes
We selected specific mRNAs and lncRNAs for further qPCR validation, and two replicates per sample were performed. The validated DE RNAs were selected considering their fold change (FC), their previous description as “known”, antisense, and processed transcripts, and a balance between up- and down-regulated transcripts.
Transcripts validated in SALS are reported in Figure 2. Those validated in AD are reported in Figure 3 and PD validated ones are shown in Figure 4. Given the relevance of DE genes found in sALS patients, we decided to expand qPCR validation on a larger cohort of sporadic patients: 30 SALS and 30 healthy controls PBMCs underwent lncRNAs confirmation, while 10 SALS and 10 healthy controls PBMCs underwent mRNAs validation.
Figure 2.

Differentially expressed transcripts verified by Real Time PCR in PBMCs from a larger cohort of SALS and CTRLs (N = 30 for lncRNAs and N = 10 for coding RNAs. * p < 0.05, ** p < 0.001 vs. CTR).
Figure 3.

Differentially expressed transcripts verified by Real Time PCR in PBMC from AD and CTRs (N = 6).
Figure 4.

Differentially expressed transcripts verified by Real Time PCR in PBMC from PD and CTRs (N = 6).
2.3. mRNA Pathway Analysis
2.3.1. Amyotrophic Lateral Sclerosis
KEGG pathway analysis for the totality of DE mRNAs includes evidences of cancer-related pathways, longevity pathway and atherosclerosis (Figure 5). The GO biological processes enriched terms are related to intracellular transport, Reactive Oxygen Species (ROS) response, and regulation of transcription (Figure 6A).
Figure 5.

Analysis of pathways performed with KEGG using DE genes in SALS group compared to healthy controls. We only show top 10 KEGG terms. The significance of the specific gene-set term is represented by the length of the bar. The significance of the term is indicated by the brightness of the bar’s color (the brighter, the more significant).
Figure 6.
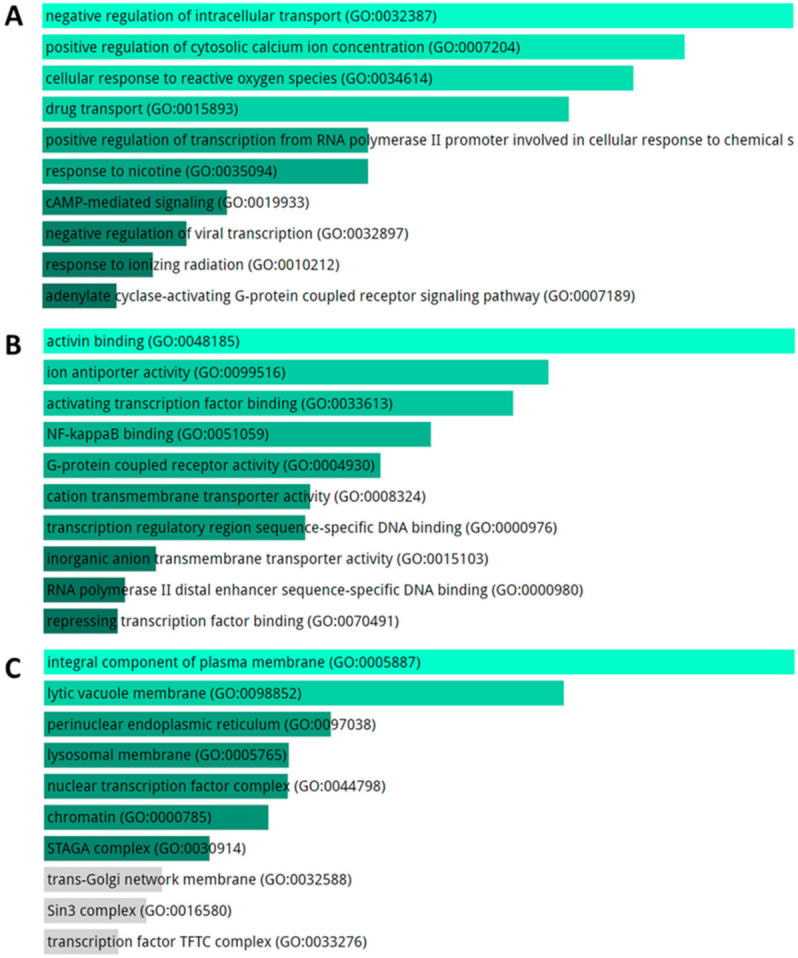
GO analysis for DE genes in SALS patients compared to healthy controls. TOP10 enriched GO terms for biological process (A), molecular function (B) and cellular component (C) The length of the bar represents the significance of that specific gene-set or term. The brighter the color, the more significant that term is, and the grey color refers to non-significant terms.
Activating binding, transcription factor activity, NF-kappaB binding and activating transcription factor binding, and DNA binding were the most enriched GO terms in the Molecular Function database. (Figure 6B). Moreover, integral components of plasma membrane and chromatin resulted as enriched GO terms for cellular component (Figure 6C).
2.3.2. Alzheimer’s Disease
KEGG pathways enriched by DE genes highlighted terms related to ribosome, focal adhesion, and atherosclerosis (Figure 7). The GO enriched terms for biological process are represented by exocytosis and protein targeting membrane (Figure 8A). GO molecular function analysis resulted in ribosome and focal adhesion (Figure 8B). With respect to cellular components, the most enriched GO concern phosphorylase and oxidase activity (Figure 8C).
Figure 7.

Pathways analysis performed with KEGG using DE genes in AD group compared to healthy controls. We report the top 10 KEGG terms. The significance of the specific gene-set term is represented by the length of the bar. The significance of the term is indicated by the brightness of the bar’s color (the brighter, the more significant).
Figure 8.
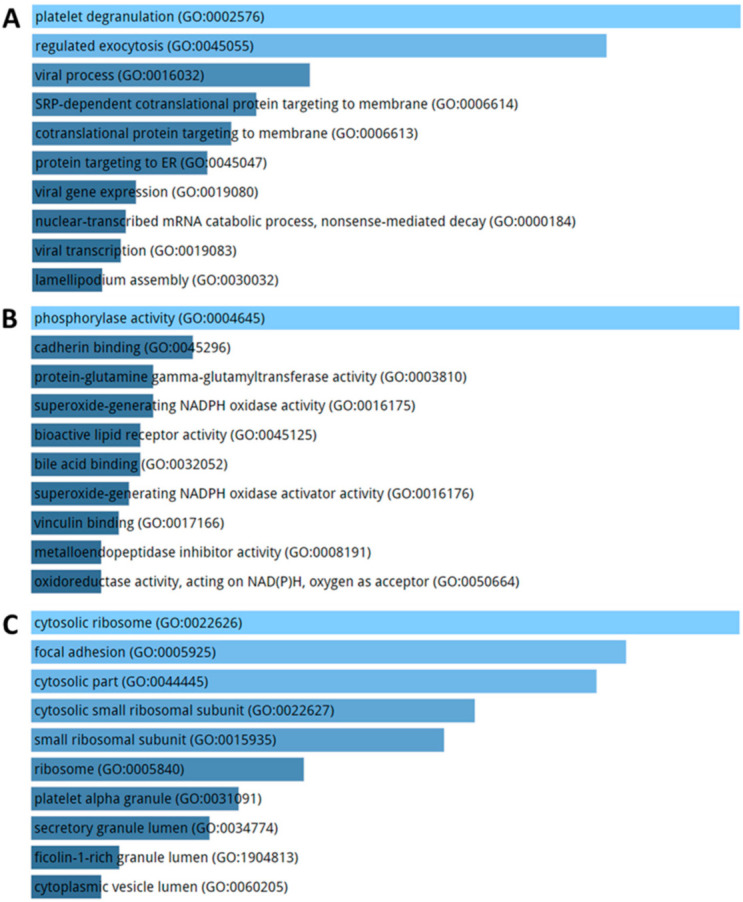
Biological process (A), molecular function (B) and cellular component (C) enrichment analysis performed on DE genes in AD patients compared to healthy controls. The significance of the specific gene-set term is represented by the length of the bar. The significance of the term is indicated by the brightness of the bar’s color (the brighter, the more significant).
KEGG pathway and GO terms enrichment analysis for DEs in SALS and AD patients compared to healthy controls has been performed for DE genes [39]. We did not conduce this analysis for PD because of the low number of resulting DE. Despite the fact that no common DE genes were found in the three cohorts of patients (Figure 9), some shared features related to KEGG and GO were reported (Table 2).
Figure 9.
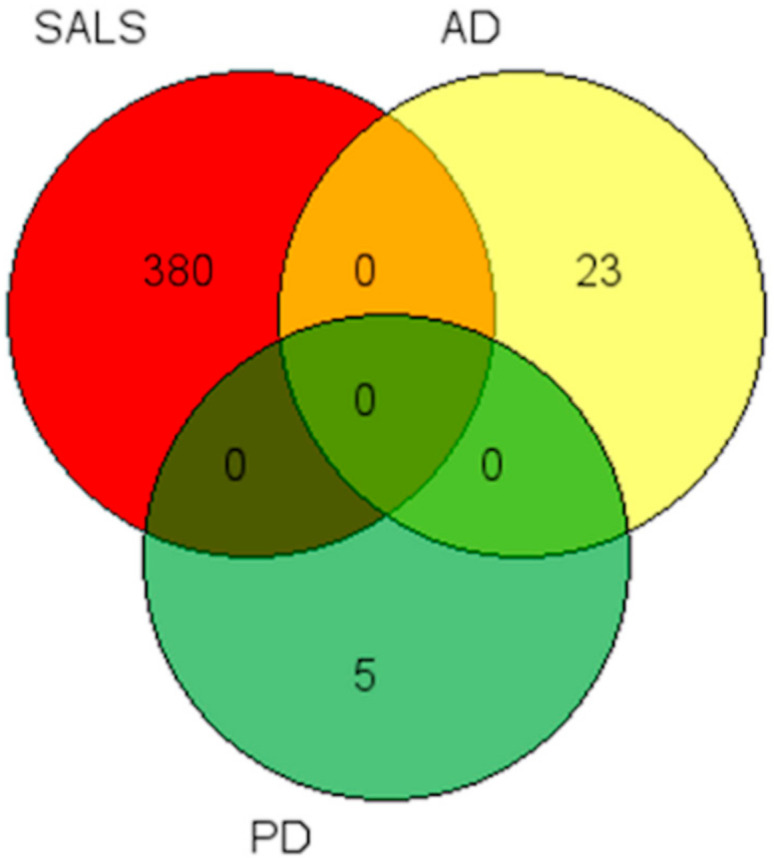
Venn diagram shows differentially expressed genes in common among the three conditions. No common deregulated gene emerged from DE analysis. In red the number of DE genes in SALS, in yellow the number of DE genes on AD and in green the number of DE genes in PD.
Table 2.
Shared enriched KEGG or GO terms among SALS and AD patients. In the first column the term is reported; in the columns relative to each disease the genes with the highest fold change involved in each term are reported, in green those up-regulated, in red those down-regulated.
| KEGG/GO Term | SALS | AD |
|---|---|---|
| Dilated cardiomyopathy (DCM) | ADCY9 PLN |
ITGA2B |
| Complement and coagulation cascades | A2M F2RL2 |
F13A1 |
| Fluid shear stress and atherosclerosis | RELA ACVR2A NFE2L2 |
NCF1 ITGA2B |
| Cellular response to reactive oxygen species | MPV17L RELA NFE2L2 |
NCF1 |
| Protein localization cell surface | SMURF1 | VCL |
3. Discussion
Transcriptome analysis enables one to envision the complete biological context of disease pathogenesis by studying the gene expression profile. Moreover, the emergent role of various classes of regulatory non-coding RNAs (ncRNAs) in the onset of several diseases, such as cancer and neurodegenerative disorders, is becoming increasingly recognized. We performed a full profiling, via an RNA-Seq approach, of the lncRNAs and mRNAs in human PBMCs from SALS patients, AD patients, PD patients, and healthy controls in order to understand the coding and ncRNAs driven epitranscriptomics effect on the pathogenesis of these diseases.
Our work consisted in a deep screening of both coding and non-coding RNAs in ALS, AD and PD patients with the aim of describing the transcriptome alterations present in these diseases and assessing whether a shared deregulation was present in coding/noncoding RNAs.
Even if we did not find any common DE genes in the three cohorts of patients, some shared feature related to KEGG and GO terms were reported in SALS and AD, but not in PD as the number of DE genes made impossible the realization of this analysis. KEGG analysis revealed “dilated cardiomyopathy”, “complement and coagulation cascade” and “fluid shear stress” as deregulated pathways in both diseases. Interestingly, the specific genes involved in these pathways are different, and moreover it is possible to nota that in SALS the genes are both up-regulated and down-regulated, whereas in AD these pathways pertain only up-regulated genes. A similar situation is also found in Gene Ontology analysis, where we found an enrichment of “cellular response to reactive oxygen species” and “protein localization to cell surface”. In SALS group, the response to ROS is again represented by two up-regulated genes and one down-regulation of a gene.
Even if it was not possible to find overlapping terms between PD and the other two diseases, it is interesting to note that, amongst the KEGG pathways related to AD downregulated DE RNAs, the term “Parkinson’s disease” is present. Indeed, NDUFV2, down-regulated in AD patients, has been addressed as a genetic variation promoting a mild form of Parkinsonism with a prognosis similar to that of idiopathic PD [40].
The common pathways affected in both ALS and AD that emerged from KEGG analysis are extremely relevant in neurodegeneration pathophysiology. Indeed, “dilated cardiomyopathy” has already been associated to heart failure in both AD, because of Aβ amyloid accumulation in the heart [41], and ALS, because of sympathetic hyperactivity that leads to cardiac death [42], which is the second cause of death in these patients. Also, “complement and coagulation cascades” resulted from DE genes in both groups of patients. These two cascades are known to be regulated by a tight balance between protective and toxic effects and under molecular stress typical of neurodegeneration the signaling of these processes might be compromised promoting cellular damage [43]. It is also relevant to mention that one of the resulting cellular processes in GO analysis was “cellular response to reactive oxygen species”. This is highly relevant since the oxidative stress is one of the most studied molecular features of neurodegenerative diseases [44,45].
The crucial result that emerged from our RNA-seq analysis is the one concerning the small number of DE genes found in PD PBMCs. Despite the fact that an important deregulation of noncoding genes in CNS is observed [46], it has already been reported that, in peripheral blood, these RNAs are present but do not change their expression when compared to healthy controls [47,48]. In this group, we found the TBC1D3 gene to be downregulated. This transcript is not related to any PD molecular alteration, but it has been addressed as a promoter of the generation of basal neural progenitors and inductor of cortical folding in mice [38]. The only up-regulated gene was SCARNA2 (small Cajal body-specific RNA 2), a lincRNA which is independently transcribed [49]. Two other ncRNAs were found down-regulated, namely RP1-29C18.9 and RP1-29C18.8, but there is nothing reported in literature concerning their potential implication in PD.
Our data highlighted the different grade of RNA metabolism involvement in Alzheimer’s disease, amyotrophic lateral sclerosis, and Parkinson’s disease. Specifically, we found a great difference in the amount of differentially expressed genes in the three conditions, meaning that a milder transcriptional machinery involvement is present in AD and PD compared to ALS. Despite this imbalance, some molecular features common amongst them are present.
Considering our results, it emerged that a more extensive study of coding and noncoding genes modulation is needed in order to exploit novel mechanisms involved in the molecular pathogenesis of ALS, with the aim of identifying possible therapeutic targets.
4. Materials and Methods
4.1. Study Subjects
Briefly, 10 SALS, 6 AD, and 6 PD patients and 14 age- and sex-matched healthy controls (CTR) were recruited after obtaining their written informed consent (Table 3). All the subjects were deep-sequenced and included in Real Time PCR experiments. ALS, AD and PD patients underwent clinical and neurologic examination at IRCCS Mondino Foundation (Pavia, Italy). ALS patients were diagnosed following the El Escorial criteria [50]. AD diagnosis was made according to the National Institute of Neurological and Communicative Disorders and Stroke and the AD and Related Disorders Association (NINCDS-ADRDA) criteria [51]. For PD, patients Movement Disorder Society (MDS) clinical diagnostic criteria were used [52]. All patients were analyzed to exclude any mutations in causative genes by whole exome sequencing. The control subjects were recruited at the Transfusional Service and Centre of Transplantation Immunology, Foundation San Matteo, IRCCS (Pavia, Italy). The study protocol to obtain PBMCs from patients and controls was approved by the Ethical Committee of the IRCCS Mondino Foundation (Pavia, Italy), (Codes 20200045392; n°20170001758; n°2020042334). Before being enrolled, the subjects participating in the study signed an informed consent form (Code 375/04—version 07/01/2004). All experiments were performed in accordance with relevant guidelines and regulations.
Table 3.
Baseline characteristics of subjects recruited for this study.
| SALS | AD | PD | CTRL | |
|---|---|---|---|---|
| n = 10 | n = 6 | n = 6 | n = 14 | |
| Age (M ± SD) | 66.6 ± 10.1 | 78.2 ± 8.0 | 62.7 ± 6.7 | 53.2 ± 8.3 |
| Sex | ||||
| Males n (%) | 45% | 34% | 83% | 64% |
| Females n (%) | 55% | 66% | 17% | 32% |
4.2. Isolation of Human Peripheral Blood Mononuclear Cells and RNA Extraction Sequencing
PBMCs were prepared by centrifugation layering peripheral blood on Ficoll-Histopaque (density = 1.077) and centrifuged at 950 g for 30 min. After isolation, cell viability was assessed by a trypan blue exclusion test. Total RNA was isolated by Trizol reagent (Life Science Technologies, Monza, Italy) according to manufacturer’s instructions. Quantification and quality control of RNAs were performed using a Nanodrop ND-100 Spectrophotometer (Nanodrop Technologies, Wilmington, USA) and a 2100 Bioanalyzer (Agilent RNA 6000 Nano Kit, Waldbronn, Germany); RNAs with a 260:280 ratio of ≥1.5 and an RNA integrity number of ≥8 were selected for deep sequencing.
4.3. Libraries Preparation for RNA-Seq and Bioinformatic Data Analysis
Sequencing libraries of SALS and AD patients and matched controls were prepared with the Illumina TruSeq Stranded RNA Library Prep kit, version 2, Protocol D, using 500 ng total RNA (Illumina, San Diego, USA). For PD patients and controls libraries it has been used the SENSE Total RNA-Seq Library Prep Kit (Lexogen, Vienna, Austria), starting from 200 ng total RNA. Quality of sequencing libraries was assessed by 2100 Bioanalyzer with a DNA1000 assay and DNA High Sensitivity assay (Agilent, Waldbronn, Germany).
RNA processing was carried out using Illumina NextSeq 500 Sequencing (Illumina, San Diego, USA). FastQ files obtained from raw sequencing reads produced by Illumina NextSeq sequencer were generated via llumina bcl2 fastq2(Version2.17.1.14-http://support.illumina.com/downloads/bcl-2fastq-conversion-software-v217.htm). STAR/RSEM software was used for computing gene and transcript intensities [53] using Gencode Release 19 (GRCh37.p13) as a reference, using the “stranded” option. Number of transcripts having at least 10 counts in the three diseases are reported in Supplementary Table S1. R package EBSeq was used for differential expression analysis of mRNA was performed using [54], because of its higher performance in identifying isoforms differential expression [55], while for long non-coding RNAs the R package DESeq.2 was employed [56]. Coding and non-coding genes were considered differentially expressed and retained for further analysis with |log2(disease sample/healthy control)| ≥ 1 and a FDR ≤ 0.1. This choice is motivated by the decision to maximize the sensitivity of this analysis, in order to perform a massive screening. RNA sequencing data are available in GEO repository (GSE106443; GSE161199).
4.4. Pathway Analysis
Coding genes underwent gene enrichment analysis [39]. To do so, we used Gene Ontology 2018 (GO) for biological processes, cellular components, and molecular function and KEGG 2019 (Kyoto Encyclopedia of Genes and Genomes http://www.genome.ad.jp/kegg) for pathway analysis via the enrichR web tool [57,58].
4.5. Real Time PCR
Primer 3.0 was used to design PCR oligonucleotides specifically targeting and amplifying mRNA, avoiding non-specific amplification products. Moreover, primers were chosen so as to not overlap with antisense sequences (primers upon request). An iScript™ cDNA Synthesis Kit (BioRad, Richmond, CA, USA) was used for preparing total cDNAs. Real time PCR (qPCR) reactions were performed with SYBR Green SuperMix (BioRad, Richmond, CA, USA), using 1 μL of cDNA template (or water control). Cycle threshold (Ct) values were normalized against those determined for GAPDH. Determination of fold-expression differences relative to healthy controls were carried out using the 2ΔΔCt method. Student’s t-test was used for the determination of significance of gene expression changes relative to controls using Prism GraphPad 5.02 software (GraphPad Software, San Diego, CA, USA).
Acknowledgments
F.R. would like to acknowledge and thank the Fondazione Fratelli Confalonieri for financial support during her Ph.D.
Abbreviations
| SALS | Sporadic Amyotrophic Lateral Sclerosis |
| AD | Alzheimer’s disease |
| PD | Parkinson’s disease |
| CTRL | Control |
| lncRNAs | Long non-coding RNAs |
| DE | Differentially expressed |
Supplementary Materials
Supplementary materials can be found at https://www.mdpi.com/1422-0067/21/24/9500/s1.
Author Contributions
M.G. and C.P. experiments execution, drafting the manuscript and figures; M.B. experiments execution; O.P. acquisition and analysis of data; F.R. experiments execution; S.C. revision of the manuscript and analysis of data; A.C., B.M. and L.D. patients and controls recruitment.; S.Z. bioinformatics analysis, C.C. design of the study, revision of the manuscript; S.G. design of the study, analysis of data, and drafting the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
Project was partially funded by Fondazione Regionale Ricerca Biomedica (TRANS-ALS)-(2015-0023), by the Italian Ministry of Health (Ricerca Corrente 2018–2020), GR-2016-02361552 Italian Ministry of Health, EuroNanoMed III JTC 2018 and Italian Ministry of Health, Cariplo Foundation (Italy)—[FrailBioTrack 2017-0557], Agenzia Italiana del Farmaco (2016-02364678).
Conflicts of Interest
The authors declare no conflict of interest. S.Z. has shares of enGenome, an italian bioinformatics company The funders and enGenome had no role in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Hussain R., Zubair H., Pursell S., Shahab M. Neurodegenerative Diseases: Regenerative Mechanisms and Novel Therapeutic Approaches. Brain Sci. 2018;8 doi: 10.3390/brainsci8090177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Wyss-Coray T. Ageing, neurodegeneration and brain rejuvenation. Nature. 2016;539:180–186. doi: 10.1038/nature20411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Jeong S. Molecular and Cellular Basis of Neurodegeneration in Alzheimer’s Disease. Mol. Cells. 2017;40:613–620. doi: 10.14348/molcells.2017.0096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Polymenidou M., Lagier-Tourenne C., Hutt K.R., Bennett C.F., Cleveland D.W., Yeo G.W. Misregulated RNA processing in amyotrophic lateral sclerosis. Brain Res. 2012;1462:3–15. doi: 10.1016/j.brainres.2012.02.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Serrano-Pozo A., Frosch M.P., Masliah E., Hyman B.T. Neuropathological alterations in Alzheimer disease. Cold Spring Harb. Perspect. Med. 2011;1:a006189. doi: 10.1101/cshperspect.a006189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kalia L.V., Lang A.E. Parkinson’s disease. Lancet. 2015;386:896–912. doi: 10.1016/S0140-6736(14)61393-3. [DOI] [PubMed] [Google Scholar]
- 7.Molteni M., Rossetti C. Neurodegenerative diseases: The immunological perspective. J. Neuroimmunol. 2017;313:109–115. doi: 10.1016/j.jneuroim.2017.11.002. [DOI] [PubMed] [Google Scholar]
- 8.Kiriyama Y., Nochi H. The Function of Autophagy in Neurodegenerative Diseases. Int. J. Mol. Sci. 2015;16:26797–26812. doi: 10.3390/ijms161125990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.García J.C., Bustos R.H. The Genetic Diagnosis of Neurodegenerative Diseases and Therapeutic Perspectives. Brain Sci. 2018;8 doi: 10.3390/brainsci8120222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Liu E.Y., Cali C.P., Lee E.B. RNA metabolism in neurodegenerative disease. Dis. Model. Mech. 2017;10:509–518. doi: 10.1242/dmm.028613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Alaqeel A.M., Abou Al-Shaar H., Shariff R.K., Albakr A. The role of RNA metabolism in neurological diseases. Balk. J. Med. Genet. 2015;18:5–14. doi: 10.1515/bjmg-2015-0080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Weskamp K., Barmada S.J. RNA Degradation in Neurodegenerative Disease. Adv. Neurobiol. 2018;20:103–142. doi: 10.1007/978-3-319-89689-2_5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Huntzinger E., Izaurralde E. Gene silencing by microRNAs: Contributions of translational repression and mRNA decay. Nat. Rev. Genet. 2011;12:99–110. doi: 10.1038/nrg2936. [DOI] [PubMed] [Google Scholar]
- 14.Figueroa-Romero C., Hur J., Lunn J.S., Paez-Colasante X., Bender D.E., Yung R., Sakowski S.A., Feldman E.L. Expression of microRNAs in human post-mortem amyotrophic lateral sclerosis spinal cords provides insight into disease mechanisms. Mol. Cell. Neurosci. 2016;71:34–45. doi: 10.1016/j.mcn.2015.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chen Y., Wei Q., Chen X., Li C., Cao B., Ou R., Hadano S., Shang H.F. Aberration of miRNAs Expression in Leukocytes from Sporadic Amyotrophic Lateral Sclerosis. Front. Mol. Neurosci. 2016;9:69. doi: 10.3389/fnmol.2016.00069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Nishimoto Y., Nakagawa S., Hirose T., Okano H.J., Takao M., Shibata S., Suyama S., Kuwako K., Imai T., Murayama S., et al. The long non-coding RNA nuclear-enriched abundant transcript 1_2 induces paraspeckle formation in the motor neuron during the early phase of amyotrophic lateral sclerosis. Mol. Brain. 2013;6:31. doi: 10.1186/1756-6606-6-31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Sunwoo J.S., Lee S.T., Im W., Lee M., Byun J.I., Jung K.H., Park K.I., Jung K.Y., Lee S.K., Chu K., et al. Altered Expression of the Long Noncoding RNA NEAT1 in Huntington’s Disease. Mol. Neurobiol. 2017;54:1577–1586. doi: 10.1007/s12035-016-9928-9. [DOI] [PubMed] [Google Scholar]
- 18.Simchovitz A., Hanan M., Niederhoffer N., Madrer N., Yayon N., Bennett E.R., Greenberg D.S., Kadener S., Soreq H. NEAT1 is overexpressed in Parkinson’s disease substantia nigra and confers drug-inducible neuroprotection from oxidative stress. FASEB J. 2019;33:11223–11234. doi: 10.1096/fj.201900830R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Sun Q., Zhang Y., Wang S., Yang F., Cai H., Xing Y., Chen Z., Chen J. NEAT1 Decreasing Suppresses Parkinson’s Disease Progression via Acting as miR-1301-3p Sponge. J. Mol. Neurosci. 2020 doi: 10.1007/s12031-020-01660-2. [DOI] [PubMed] [Google Scholar]
- 20.Kapeli K., Martinez F.J., Yeo G.W. Genetic mutations in RNA-binding proteins and their roles in ALS. Hum. Genet. 2017;136:1193–1214. doi: 10.1007/s00439-017-1830-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Millan M.J. Linking deregulation of non-coding RNA to the core pathophysiology of Alzheimer’s disease: An integrative review. Prog. Neurobiol. 2017;156:1–68. doi: 10.1016/j.pneurobio.2017.03.004. [DOI] [PubMed] [Google Scholar]
- 22.Singh A., Sen D. MicroRNAs in Parkinson’s disease. Exp. Brain Res. 2017;235:2359–2374. doi: 10.1007/s00221-017-4989-1. [DOI] [PubMed] [Google Scholar]
- 23.Byron S.A., van Keuren-Jensen K.R., Engelthaler D.M., Carpten J.D., Craig D.W. Translating RNA sequencing into clinical diagnostics: Opportunities and challenges. Nat. Rev. Genet. 2016;17:257–271. doi: 10.1038/nrg.2016.10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Gagliardi S., Zucca S., Pandini C., Diamanti L., Bordoni M., Sproviero D., Arigoni M., Olivero M., Pansarasa O., Ceroni M., et al. Long non-coding and coding RNAs characterization in Peripheral Blood Mononuclear Cells and Spinal Cord from Amyotrophic Lateral Sclerosis patients. Sci. Rep. 2018;8:2378. doi: 10.1038/s41598-018-20679-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Luotti S., Pasetto L., Porcu L., Torri V., Elezgarai S.R., Pantalone S., Filareti M., Corbo M., Lunetta C., Mora G., et al. Diagnostic and prognostic values of PBMC proteins in amyotrophic lateral sclerosis. Neurobiol. Dis. 2020;139:104815. doi: 10.1016/j.nbd.2020.104815. [DOI] [PubMed] [Google Scholar]
- 26.Mosallaei M., Ehtesham N., Rahimirad S., Saghi M., Vatandoost N., Khosravi S. PBMCs: A new source of diagnostic and prognostic biomarkers. Arch. Physiol. Biochem. 2020:1–7. doi: 10.1080/13813455.2020.1752257. [DOI] [PubMed] [Google Scholar]
- 27.Gagliardi S., Franco V., Sorrentino S., Zucca S., Pandini C., Rota P., Bernuzzi S., Costa A., Sinforiani E., Pansarasa O., et al. Curcumin and Novel Synthetic Analogs in Cell-Based Studies of Alzheimer’s Disease. Front. Pharmacol. 2018;9:1404. doi: 10.3389/fphar.2018.01404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lazo-Gómez R., Ramírez-Jarquín U.N., Tovar-Y-Romo L.B., Tapia R. Histone deacetylases and their role in motor neuron degeneration. Front. Cell. Neurosci. 2013;7:243. doi: 10.3389/fncel.2013.00243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kim T., Cui R., Jeon Y.J., Fadda P., Alder H., Croce C.M. MYC-repressed long noncoding RNAs antagonize MYC-induced cell proliferation and cell cycle progression. Oncotarget. 2015;6:18780–18789. doi: 10.18632/oncotarget.3909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Guo X., Qi X. VCP cooperates with UBXD1 to degrade mitochondrial outer membrane protein MCL1 in model of Huntington’s disease. Biochim. Biophys. Acta Mol. Basis Dis. 2017;1863:552–559. doi: 10.1016/j.bbadis.2016.11.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lee J.K., Shin J.H., Lee J.E., Choi E.J. Role of autophagy in the pathogenesis of amyotrophic lateral sclerosis. Biochim. Biophys. Acta. 2015;1852:2517–2524. doi: 10.1016/j.bbadis.2015.08.005. [DOI] [PubMed] [Google Scholar]
- 32.Villar-Menéndez I., Porta S., Buira S.P., Pereira-Veiga T., Díaz-Sánchez S., Albasanz J.L., Ferrer I., Martín M., Barrachina M. Increased striatal adenosine A2A receptor levels is an early event in Parkinson’s disease-related pathology and it is potentially regulated by miR-34b. Neurobiol. Dis. 2014;69:206–214. doi: 10.1016/j.nbd.2014.05.030. [DOI] [PubMed] [Google Scholar]
- 33.Castillo E., Leon J., Mazzei G., Abolhassani N., Haruyama N., Saito T., Saido T., Hokama M., Iwaki T., Ohara T., et al. Comparative profiling of cortical gene expression in Alzheimer’s disease patients and mouse models demonstrates a link between amyloidosis and neuroinflammation. Sci. Rep. 2017;7:17762. doi: 10.1038/s41598-017-17999-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Hashimoto Y., Niikura T., Tajima H., Yasukawa T., Sudo H., Ito Y., Kita Y., Kawasumi M., Kouyama K., Doyu M., et al. A rescue factor abolishing neuronal cell death by a wide spectrum of familial Alzheimer’s disease genes and Abeta. Proc. Natl. Acad. Sci. USA. 2001;98:6336–6341. doi: 10.1073/pnas.101133498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Grace E.A., Busciglio J. Aberrant activation of focal adhesion proteins mediates fibrillar amyloid beta-induced neuronal dystrophy. J. Neurosci. 2003;23:493–502. doi: 10.1523/JNEUROSCI.23-02-00493.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Calderwood D.A., Campbell I.D., Critchley D.R. Talins and kindlins: Partners in integrin-mediated adhesion. Nat. Rev. Mol. Cell Biol. 2013;14:503–517. doi: 10.1038/nrm3624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Bao Z., Yang Z., Huang Z., Zhou Y., Cui Q., Dong D. LncRNADisease 2.0: An updated database of long non-coding RNA-associated diseases. Nucleic Acids Res. 2019;47:D1034–D1037. doi: 10.1093/nar/gky905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ju X.C., Hou Q.Q., Sheng A.L., Wu K.Y., Zhou Y., Jin Y., Wen T., Yang Z., Wang X., Luo Z.G. The hominoid-specific gene TBC1D3 promotes generation of basal neural progenitors and induces cortical folding in mice. Elife. 2016;5 doi: 10.7554/eLife.18197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Subramanian A., Tamayo P., Mootha V.K., Mukherjee S., Ebert B.L., Gillette M.A., Paulovich A., Pomeroy S.L., Golub T.R., Lander E.S., et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA. 2005;102:15545–15550. doi: 10.1073/pnas.0506580102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Nishioka K., Vilariño-Güell C., Cobb S.A., Kachergus J.M., Ross O.A., Hentati E., Hentati F., Farrer M.J. Genetic variation of the mitochondrial complex I subunit NDUFV2 and Parkinson’s disease. Parkinsonism Relat. Disord. 2010;16:686–687. doi: 10.1016/j.parkreldis.2010.09.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Troncone L., Luciani M., Coggins M., Wilker E.H., Ho C.Y., Codispoti K.E., Frosch M.P., Kayed R., Del Monte F. Aβ Amyloid Pathology Affects the Hearts of Patients With Alzheimer’s Disease: Mind the Heart. J. Am. Coll. Cardiol. 2016;68:2395–2407. doi: 10.1016/j.jacc.2016.08.073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Gdynia H.J., Kurt A., Endruhn S., Ludolph A.C., Sperfeld A.D. Cardiomyopathy in motor neuron diseases. J. Neurol. Neurosurg. Psychiatry. 2006;77:671–673. doi: 10.1136/jnnp.2005.078600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Orsini F., de Blasio D., Zangari R., Zanier E.R., de Simoni M.G. Versatility of the complement system in neuroinflammation, neurodegeneration and brain homeostasis. Front. Cell. Neurosci. 2014;8:380. doi: 10.3389/fncel.2014.00380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Dasuri K., Zhang L., Keller J.N. Oxidative stress, neurodegeneration, and the balance of protein degradation and protein synthesis. Free. Radic. Biol. Med. 2013;62:170–185. doi: 10.1016/j.freeradbiomed.2012.09.016. [DOI] [PubMed] [Google Scholar]
- 45.Kim G.H., Kim J.E., Rhie S.J., Yoon S. The Role of Oxidative Stress in Neurodegenerative Diseases. Exp. Neurobiol. 2015;24:325–340. doi: 10.5607/en.2015.24.4.325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Wei C.W., Luo T., Zou S.S., Wu A.S. The Role of Long Noncoding RNAs in Central Nervous System and Neurodegenerative Diseases. Front. Behav. Neurosci. 2018;12:175. doi: 10.3389/fnbeh.2018.00175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Soreq L., Guffanti A., Salomonis N., Simchovitz A., Israel Z., Bergman H., Soreq H. Long non-coding RNA and alternative splicing modulations in Parkinson’s leukocytes identified by RNA sequencing. PLoS Comput. Biol. 2014;10:e1003517. doi: 10.1371/journal.pcbi.1003517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Elkouris M., Kouroupi G., Vourvoukelis A., Papagiannakis N., Kaltezioti V., Matsas R., Stefanis L., Xilouri M., Politis P.K. Long Non-coding RNAs Associated With Neurodegeneration-Linked Genes Are Reduced in Parkinson’s Disease Patients. Front. Cell. Neurosci. 2019;13:58. doi: 10.3389/fncel.2019.00058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Gérard M.A., Myslinski E., Chylak N., Baudrey S., Krol A., Carbon P. The scaRNA2 is produced by an independent transcription unit and its processing is directed by the encoding region. Nucleic Acids Res. 2010;38:370–381. doi: 10.1093/nar/gkp988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Brooks B.R. El Escorial World Federation of Neurology criteria for the diagnosis of amyotrophic lateral sclerosis. Subcommittee on Motor Neuron Diseases/Amyotrophic Lateral Sclerosis of the World Federation of Neurology Research Group on Neuromuscular Diseases and the El Escorial “Clinical limits of amyotrophic lateral sclerosis” workshop contributors. J. Neurol. Sci. 1994;124:96–107. doi: 10.1016/0022-510x(94)90191-0. [DOI] [PubMed] [Google Scholar]
- 51.McKhann G., Drachman D., Folstein M., Katzman R., Price D., Stadlan E.M. Clinical diagnosis of Alzheimer’s disease: Report of the NINCDS-ADRDA Work Group under the auspices of Department of Health and Human Services Task Force on Alzheimer’s Disease. Neurology. 1984;34:939–944. doi: 10.1212/WNL.34.7.939. [DOI] [PubMed] [Google Scholar]
- 52.Postuma R.B., Berg D., Stern M., Poewe W., Olanow C.W., Oertel W., Obeso J., Marek K., Litvan I., Lang A.E., et al. MDS clinical diagnostic criteria for Parkinson’s disease. Mov. Disord. 2015;30:1591–1601. doi: 10.1002/mds.26424. [DOI] [PubMed] [Google Scholar]
- 53.Li B., Dewey C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011;12:323. doi: 10.1186/1471-2105-12-323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Leng N., Dawson J.A., Thomson J.A., Ruotti V., Rissman A.I., Smits B.M., Haag J.D., Gould M.N., Stewart R.M., Kendziorski C. EBSeq: An empirical Bayes hierarchical model for inference in RNA-seq experiments. Bioinformatics. 2013;29:1035–1043. doi: 10.1093/bioinformatics/btt087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Carrara M., Lum J., Cordero F., Beccuti M., Poidinger M., Donatelli S., Calogero R.A., Zolezzi F. Alternative splicing detection workflow needs a careful combination of sample prep and bioinformatics analysis. BMC Bioinform. 2015;16(Suppl. 9):S2. doi: 10.1186/1471-2105-16-S9-S2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Love M.I., Huber W., Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014;15:550. doi: 10.1186/s13059-014-0550-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Kuleshov M.V., Jones M.R., Rouillard A.D., Fernandez N.F., Duan Q., Wang Z., Koplev S., Jenkins S.L., Jagodnik K.M., Lachmann A., et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016;44:W90–W97. doi: 10.1093/nar/gkw377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Chen E.Y., Tan C.M., Kou Y., Duan Q., Wang Z., Meirelles G.V., Clark N.R., Ma’ayan A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013;14:128. doi: 10.1186/1471-2105-14-128. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.