

Abstract
Resistance (host capacity to reduce parasite burden) and tolerance (host capacity to reduce impact on its health for a given parasite burden) manifest two different lines of defense. Tolerance can be independent from resistance, traded off against it, or the two can be positively correlated because of redundancy in underlying (immune) processes. We here tested whether this coupling between tolerance and resistance could differ upon infection with closely related parasite species. We tested this in experimental infections with two parasite species of the genus Eimeria. We measured proxies for resistance (the (inverse of) number of parasite transmission stages (oocysts) per gram of feces at the day of maximal shedding) and tolerance (the slope of maximum relative weight loss compared to day of infection on number of oocysts per gram of feces at the day of maximal shedding for each host strain) in four inbred mouse strains and four groups of F1 hybrids belonging to two mouse subspecies, Mus musculus domesticus and Mus musculus musculus. We found a negative correlation between resistance and tolerance against Eimeria falciformis, while the two are uncoupled against Eimeria ferrisi. We conclude that resistance and tolerance against the first parasite species might be traded off, but evolve more independently in different mouse genotypes against the latter. We argue that evolution of the host immune defenses can be studied largely irrespective of parasite isolates if resistance–tolerance coupling is absent or weak (E. ferrisi) but host–parasite coevolution is more likely observable and best studied in a system with negatively correlated tolerance and resistance (E. falciformis).
Keywords: coevolution, Eimeria, resistance, tolerance
We tested whether closely related parasite species could show different coupling between tolerance and resistance and found a trade‐off between resistance and tolerance to one, Eimeria falciformis, but not to its close relative Eimeria ferrisi. The framework of resistance–tolerance coupling allows to prioritize research questions to be addressed with different parasites: broad questions of relevance for the host species as a whole with parasites showing no coupling, questions of local adaptation and host–parasite co‐evolution with parasites showing coupling.
1. INTRODUCTION
Host defense mechanisms evolve to alleviate the detrimental effect of parasites. They can be categorized into two components: resistance and tolerance (Råberg et al., 2009). Resistance is the ability of a host to reduce parasite burden, resulting from defense against parasite infection or proliferation early after infection (Schmid‐Hempel, 2013). The negative effect of resistance on parasite fitness can lead to antagonistic coevolution. According to theoretical models, fluctuating host and parasite genotypes arise, and balancing selection maintains resistance alleles polymorphic (Boots et al., 2008; Roy & Kirchner, 2000). Resistance has been the classical "catch all" measure for host‐parasite systems, but recently it has been shown to be incomplete, especially with respect to potential fitness effects on the host (Kutzer & Armitage, 2016; Råberg et al., 2009).
Disease tolerance (not to be confused from "immunological tolerance," unresponsiveness to self‐antigens; Medzhitov et al., 2012) is the ability of the host to limit the impact of parasite on its fitness (Kutzer & Armitage, 2016; Råberg et al., 2009; Vale & Little, 2012). By potentially providing a longer‐living niche, this defense mechanism improves, or at least does not deteriorate, the fitness of the parasite. Tolerance alleles are thus predicted by theoretical models to evolve to fixation due to positive feedback loops (Boots et al., 2008; Restif & Koella, 2004; Roy & Kirchner, 2000). From a mechanistic perspective, tolerance alleviates direct or indirect damage (e.g., excessive immune response underlying resistance against parasites, called immunopathology; Graham et al., 2005) caused by parasites (Råberg et al., 2009). Tolerance mechanisms include modulation of inflammatory response (Ayres & Schneider, 2012), tissue repair (stress response, damage repair, and cellular regeneration mechanisms; Soares et al., 2017), and compensation of parasite‐induced damage by increase of reproductive effort (Baucom & Roode, 2011). Even in the absence of parasite infection, the maintenance of tolerance mechanisms can be detrimental to other functions, ultimately affecting host fitness (Råberg et al., 2009; Stowe et al., 2000). The resulting costs of resistance and tolerance determine the optimal (steady state and infection inducible) extent of both immune defenses (Sheldon & Verhulst, 1996).
Resistance and tolerance can be positively associated if they involve the same metabolic pathway, as was shown in the plant model Arabidopsis thaliana in response against herbivory (Mesa et al., 2017). In animals, genetic association studies of resistance and tolerance of Drosophila melanogaster against the bacterium Providencia rettgeri have shown positively correlated genetic effects, as the same loci were associated with changes of both traits in the same direction (Howick & Lazzaro, 2017).
Nevertheless, resistance and tolerance can also be genetically and physiologically independent, involving different proximate mechanisms. Lack of correlation between both defenses was shown for example in monarch butterflies (Danaus plexippus) infected by the protozoan parasite Ophryocystis elektroscirrha. This study found genetic variation in resistance between butterflies families, but a fixed tolerance (Lefèvre et al., 2010). Similarly, no correlation could be found between resistance and tolerance for the fish Leuciscus burdigalensis in response to infection with its parasite Tracheliastes polycolpus. The authors explain the decoupling of both defenses by the fact that, in this system, tolerance likely involves wound repair rather than immune regulation, making resistance and tolerance mechanisms independent (Mazé‐Guilmo et al., 2014).
In other systems, resistance and tolerance have been found negatively correlated. For example, inbred laboratory mouse strains lose weight upon infection with Plasmodium chabaudi. The extent of this impact on host health is negatively correlated with the peak number of parasites found in the blood (Råberg et al., 2007), meaning that mouse strains with higher resistance present lower tolerance. Similarly, infections of sea trout (Salmo trutta trutta) and Atlantic salmon (Salmo salar) with the trematode Diplostomum pseudospathaceum showed that resistance and tolerance were negatively correlated when assessing mean levels of both traits in different host populations (Klemme & Karvonen, 2016). This is interpreted as a result of trade‐off between resistance and tolerance (Råberg et al., 2009; Restif & Koella, 2004; Sheldon & Verhulst, 1996).
We have seen that depending on the system studied, resistance and tolerance can be (a) uncoupled (independent), (b) positively correlated (involving same genes and mechanisms), or (c) negatively correlated (traded off). Theoretical models show that coupling between resistance and tolerance (or absence thereof) could depend not only on the host but also on the parasite (Carval & Ferriere, 2010). Here we tested this hypothesis. More precisely, we asked whether there could be differences in the resistance–tolerance coupling upon infection of one host type with two closely related parasite species. To answer this question, we infected four inbred mouse strains and four groups of F1 hybrids representative of two house mouse subspecies, Mus musculus domesticus and Mus musculus musculus, with two parasite isolates representative of two naturally occurring parasite species, the protozoan parasites Eimeria ferrisi and Eimeria falciformis (Jarquín‐Díaz et al., 2019). Eimeria spp. are monoxenous parasites that expand asexually and reproduce sexually in intestinal epithelial cells, leading to malabsorption of nutrients, tissue damage, and weight loss (Chapman et al., 2013). The evolutionary history of these different Eimeria species in the two house mouse subspecies is unknown and it is unclear whether subspecies‐specific adaptation exists in one or the other. We tested if coupling between resistance and tolerance differs between both parasite species and discussed the implication for parasite–host coevolution.
2. MATERIAL AND METHODS
2.1. Parasite isolates
The three parasite isolates used in this study were isolated from feces of three different M. m. domesticus/M. m. musculus hybrid mice captured in Brandenburg, Germany, in 2016 (capture permit No. 2347/35/2014). The parasite isolates belong to both the most prevalent Eimeria species in this area, namely E. ferrisi (isolate Brandenburg64) and E. falciformis (isolate Brandenburg88)(Jarquín‐Díaz et al., 2019). Isolate Brandenburg64 was isolated in a 92% M. m. domesticus individual (hybrid index (HI) = 0.08: Proportion of M. m. musculus alleles in a set of 14 diagnostic markers, see Balard et al. (2020)) and isolate Brandenburg88 in a 80% M. m. domesticus (HI = 0.2). Prepatency and the peak day of parasite shedding for these isolates were estimated during infection in NMRI laboratory mice (Al‐khlifeh et al., 2019) which were also used for serial passaging of the isolates. Previous to the experiment, the isolates had been passaged, respectively, 3 and 4 times in NMRI laboratory mice. Parasite infective forms (oocysts) were recovered by flotation in saturated NaCl solution followed by washing and observation under light microscope (following the protocol described in Clerc et al. (2019)) and stored at room temperature in 1 ml of 2% potassium dichromate for a maximum of 2 months before infection of the wild‐derived mice. Oocysts were allowed to sporulate 10 days before infection in a water bath at 30°C.
2.2. Mouse groups
We used four wild‐derived inbred mouse strains from which we generated four groups of F1 hybrids. Hybrids between M. m. domesticus and M. m. musculus are used in the present study solely to increase statistical power for comparisons among strains (such as resistance–tolerance correlations). In the future, analyses of a hybrid effect (Balard et al., 2020) could investigate tolerance and resistance employing a larger panel of such hybrid strains allowing statistical analysis of an outbreeding effect. Two parental strains represented M. m. domesticus: SCHUNT (Locality: Schweben, Hessen, Germany [N: 5°0 26′, E: 9°36′] (Martincová et al., 2019)) and STRA (Locality: Straas, Bavaria, Germany [N: 50°11′, E: 11°46′] (Piálek et al., 2008), and two derived from M. m. musculus: BUSNA (Locality: Buškovice, Bohemia, Czech Republic [N: 5°0 14′, E: 1°3 22′] (Piálek et al., 2008)) and PWD (Locality: Kunratice, Bohemia, Czech Republic [N: 5°0 01′, E: 14 2°9′] (Gregorová & Forejt, 2000)). These four strains were fully inbred, that is, passing more than 20 generations of brother–sister mating. The four groups of F1 hybrids consisted of two intrasubspecific hybrids (SCHUNTxSTRA and PWDxBUSNA) and two intersubspecific hybrids (STRAxBUSNA and SCHUNTxPWD) (Figure 1). Age of the mice at the time of infection ranged between 5.6 and 21.4 weeks, with the mean for each eight mouse group ranging between 10.5 and 14.7 weeks. All mouse strains and F1 hybrids were obtained from the Institute of Vertebrate Biology of the Czech Academy of Sciences in Studenec (license number 61974/2017‐MZE‐17214; for further details on strains see https://housemice.cz/en).
FIGURE 1.
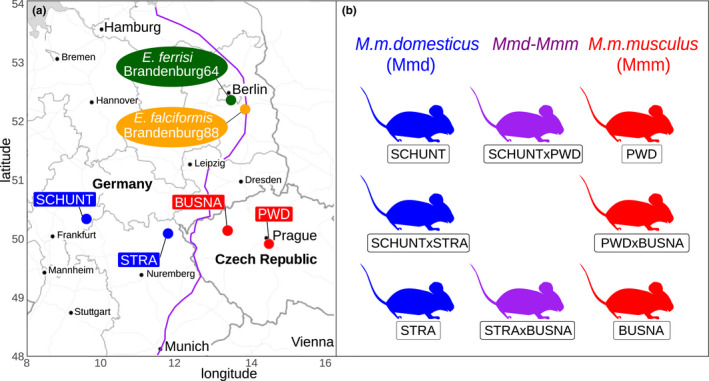
Parasite isolates and mouse wild‐derived strains. (a) Map showing locations at which mice were collected for breeding of mouse strains and isolation of parasites. The purple line is an estimation of the center of the house mouse hybrid zone betweenMus musculus domesticusandMus musculus musculusbased on sampling and genotyping of mice in this area (Balard et al., 2020; Ďureje et al., 2012; Macholán et al., 2019). (b) The eight mouse groups (parents and F1s) used in our experimental infections
Parasites of the Eimeria genus are known to induce host immune protection against reinfection (Rose et al., 1992; Smith & Hayday, 2000). To ensure that our mice were Eimeria‐naive, mouse fecal samples were tested before infection for the presence of Eimeria spp. oocysts by flotation in saturated NaCl solution followed by washing and observation under light microscope.
2.3. Experimental infection
Mice were kept in individual cages during infection. Water and food (SNIFF, Rat/Mouse maintenance feed 10 mm) were provided ad libitum supplemented with 1 g of sunflower and barley seeds per day. Mice were orally infected with 150 sporulated oocysts of one Eimeria isolate suspended in 100 μl phosphate‐buffered saline (PBS) and monitored daily until their sacrifice by cervical dislocation at time of regression of infection (reduction of oocyst output). Individuals presenting severe health deficiency and/or a weight loss approaching 18% relative to their starting weight were sacrificed earlier at defined humane end points (experiment license Reg. 0431/17). Weight was recorded and feces collected on a daily basis. Fecal pellets were collected every day from each individual cage and suspended in 2% potassium dichromate. Parasite oocysts were recovered using NaCl flotation (see above).
All individuals were negative for Eimeria at the beginning of our experiment (before infection of first batch, as described in the next paragraph). In total, 143 mice were infected. Mice were randomly allocated to experimental groups ensuring homogeneous distribution of ages and sexes between groups. Our experiments were conducted in four (partially overlapping) consecutive batches for logistical reasons. The first two batches were infected with E. ferrisi isolates (Brandenburg64), the third and fourth by one E. ferrisi isolate (Brandenburg64) and one E. falciformis isolate (Brandenburg88). Our experimental design is summarized in Table 1 (chronology of experimental batches can be scrutinized in Appendix S1).
TABLE 1.
Infection experiment design
| Host | Parasite | ||
|---|---|---|---|
| Mouse strains | Mouse subspecies | Eimeria ferrisi Brandenburg64 | Eimeria falciformis Brandenburg88 |
| SCHUNT | F0 M. m. domesticus | 14 (6M/8F) | 6 (3M/3F) |
| STRA | F0 M. m. domesticus | 15 (8M/7F) | 7 (4M/3F) |
| SCHUNTxSTRA | F1 M. m. domesticus | 6 (2M/4F) | 8 (5M/3F) |
| STRAxBUSNA | F1 hybrid | 8 (5M/3F) | 8 (3M/5F) |
| SCHUNTxPWD | F1 hybrid | 8 (3M/5F) | 6 (4M/2F) |
| PWDxBUSNA | F1 M. m. musculus | 9 (4M/5F) | 7 (4M/3F) |
| BUSNA | F0 M. m. musculus | 14 (8M/6F) | 7 (3M/4F) |
| PWD | F0 M. m. musculus | 13 (10M/3F) | 7 (1M/6F) |
Nematode infection is common in breeding facilities (Baker, 1998) and could interact with Eimeria (Clerc et al., 2019). We surveyed for their presence and nematode eggs (Syphacia sp. and Aspiculuris sp.) were observed in flotated feces of mice belonging to all genotypes before the experiment. Despite treatment of the first infection batch of mice (B1, 12 mice) with anthelminthics (Profender®, Bayer AG) following the protocol of Mehlhorn et al. (2005), nematodes were still detected with PCR (following the protocol of (Floyd et al., 2005)) in randomly sampled fecal samples a week later. We therefore decided not to treat mice of the following infection batches. Moreover, we observed Eimeria oocysts in the feces of 28 mice belonging to the last experimental batch (batch B4) at the day of infection, likely due to cross‐contamination between batches. For following statistical analyses, we considered along with the full data set (N = 143) a conservative data set in which cross‐contaminated animals and animals treated by anthelminthic were removed (N = 103). Results obtained on the conservative data set can be found in Appendix S2 and S3. Despite differences in significance due to a lower statistical power, the main conclusions of our analyses were consistent with those obtained on the main data set.
2.4. Statistical analyses
2.4.1. Choice of proxies for resistance, impact of parasite on host and tolerance
As resistance is the capacity of a host to reduce its parasite burden, it is usually estimated by the inverse of infection intensity (Råberg et al., 2009). Prepatency (the time to shedding of infectious stages, so‐called oocysts) is longer for E. falciformis (7 days) than for E. ferrisi (5 days) (Al‐khlifeh et al., 2019). Therefore, as a proxy of (inverse of) resistance, we used the number of oocysts per gram of feces (OPG) at the day of maximal shedding. Using Spearman's nonparametric rank correlation test, we found this measurement to be tightly correlated with the sum of oocysts shed throughout the experiment (Spearman's ρ = 0.93, N = 168, p < 0.001). Due to the aggregation characteristic of parasites (Shaw & Dobson, 1995), the appropriate distribution for maximum number of OPG was found to be the negative binomial distribution. This was confirmed based on log‐likelihood, AIC criteria, and goodness‐of‐fits plots (density, CDF, Q‐Q, P‐P plots; R packages MASS (Venables & Ripley, 2002) and fitdistrplus (Delignette‐Muller & Dutang, 2015)). We confirmed the fit of our models by assessing the uniformity of the distribution of model residuals.
Both parasite species provoke inflammation, cellular infiltration, enteric lesions, diarrhea, and ultimately weight loss (Al‐khlifeh et al., 2019; Ankrom et al., 1975; Ehret et al., 2017; Schito et al., 1996). Therefore, the impact of parasites on host health was measured as the maximum relative weight loss compared to day 0 (body weight measured at the start of the experimental infection). For mice sacrificed at humane end points before the end of the experiment, the last weight of the living animal was used. This weight (loss) can be expected to be a very conservative estimate for our analyses (rendering tolerance conservatively low for these animals, which might have lost more weight if not sacrificed).
Tolerance is usually defined as a reaction norm, that is, the regression slope of host fitness (or health condition if that is the parameter of interest) on infection intensity per host genotype (Råberg et al., 2009; Simms, 2000). Thus, tolerance was assessed as the slope of maximum relative weight loss compared to day 0 on number of OPG at the day of maximal shedding, within each mouse group and for each parasite isolate. A steep slope indicates a low tolerance (high weight lost for a given parasite burden).
2.4.2. Statistical comparison of resistance, impact on health, and tolerance in E. ferrisi and E. falciformis
The comparison between E. ferrisi and E. falciformis was performed using, respectively, the isolates Brandenburg64 and Brandenburg88 with which we infected all our eight mouse groups (see Table 1). Maximum OPG and relative weight loss were modeled separately as a response of mouse group, parasite isolate, and their interaction. We used a negative binomial generalized linear model for maximum OPG, and a linear model for relative weight loss. Tolerance was assessed by modeling relative weight loss as a response of maximum OPG interacting with mouse group, parasite isolate, and the interaction of the two latter. As each mouse was controlled against itself at the start of the experiment, before losing weight, or shedding parasites, we performed a linear regression with null intercept. To test the significance of the marginal contribution of each parameter to the full model, each parameter was removed from the full model, and the difference between full and reduced model was assessed using likelihood ratio tests (G).
For each of our models that showed a significant interaction term, we also asked within each parasite isolate if the response differed between mouse groups using likelihood ratio tests (G) as described above. In the case of a nonsignificant interaction term, we performed post hoc tests corrected for multiple testing (Tukey honest significant differences (HSD)) to compare within all pairwise comparisons between groups (parasite isolate–mouse strain).
Of note, four mice infected with E. falciformis isolate Brandenburg88 did not shed any oocysts as death occurred at or one day before the peak of oocysts shedding in other mice. For this reason, we modeled maximum OPG when mice infected with this parasite were included using a zero‐inflated negative binomial (ZINB) generalized linear model, after verifying that it provided a better fit than the simple negative binomial based on log‐likelihood and AIC criteria.
2.4.3. Test of coupling between resistance and tolerance
We tested coupling between resistance and tolerance for E. ferrisi and E. falciformis using the isolates Brandenburg64 and Brandenburg88 and our eight mouse groups. To test such coupling, one can assess the strength of correlation between measure of resistance and measure of tolerance (Råberg et al., 2007). Of note, tolerance (in absolute value) is measured as the slope α of the linear regression of parasite load (x) on maximum relative weight loss (y) of equation y = α x + β (α being the slope and β the intercept, 0 in our case). Therefore, tolerance is expressed as α = y/x − β/x. As x and y/x are by definition not independent, testing the correlation between resistance and tolerance can lead to spurious correlation (Brett, 2004). To alleviate the dangers of this statistical artifact, we additionally tested differences in resistance, impact on health, and tolerance between mouse groups separately (as described before, see 2.1, 2.4.2) and also the underlying correlation between mean parasite load (x) and mean relative weight loss (y). We use the terminology "coupling" (between resistance and tolerance) to describe genotype‐level correlation between tolerance and resistance additionally supported by the absence of positive correlation between health effect and resistance. Correlations were tested using Spearman's rank correlation.
After testing the resistance–tolerance coupling separately in both parasites, we tested the statistical difference in the relationship between (a) health effect and resistance and (b) tolerance and resistance in the two Eimeria species infections. To achieve this aim, we used the mean values predicted by our three models (see 2.1, 2.4.2) for each eight mouse groups to perform first a linear regression of the mean predicted relative weight loss as a response of the mean predicted OPG, parasite isolate, and their interaction, and second a linear regression of the mean predicted tolerance value as a response of the mean predicted OPG, parasite isolate, and their interaction. The significance of the marginal contribution of each parameter to the full model was assessed by removing each parameter from the full model, and the difference between full and reduced model was assessed using likelihood ratio tests (G).
All analyses were performed using R version 3.5.2 (R Development Core Team, 2013) (negative binomial: function glm.nb from R package MASS (Venables & Ripley, 2002); ZIBN: function zeroinfl from R package pscl (Jackman, 2020; Zeileis et al., 2008); linear model: function lm from R core package stats; mean and 95% confidence intervals: function ggpredict from R package ggeffect (Lüdecke, 2018)). Graphics were produced using the R package ggplot2 (Wickham, 2016) and compiled using the free software inkscape (https://inkscape.org).
3. RESULTS
3.1. General
Parasites of all isolates successfully infected all mouse groups (at the exception of 5 individuals infected with the E. falciformis isolate Brandenburg88 that died or had to be sacrificed due to a strong weight loss before the peak of shedding for this parasite), meaning that no "qualitative infection resistance" (sensu (Gandon & Michalakis, 2000)) was detected. For E. ferrisi isolate Brandenburg64, the prepatent period was 5 days postinfection (dpi) and the median day of maximal oocyst shedding was 6 dpi (standard deviation SD = 0.9). The median day of maximum weight loss was 5 dpi for both isolates (SD = 1.7). For E. falciformis isolate, Brandenburg88 prepatency was 7 dpi, median day of maximal shedding was 8 dpi (SD = 1.3), and median day of maximal weight loss was 9 dpi (SD = 1.6)(Figure 2). Of note a considerable number of mice infected with this isolate (13 out of 56 = 23%) died or had to be sacrificed at humane end points less than 3 days after the oocysts shedding peak for the group, all belonging to M. m. musculus subspecies (PWD, BUSNA, or their F1 PWDxBUSNA; 5 died at dpi 8, 5 at dpi 9, 3 at dpi 10). E. falciformis isolate Brandenburg88 was more lethal for the M. m. musculus mice strains than for the other strains (,p < 0.001; Table 2).
FIGURE 2.
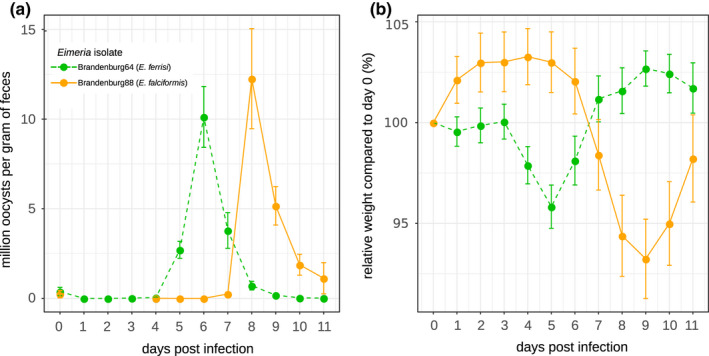
Parasite density (a) and host relative weight (b) duringEimeriainfection. Parasite density is calculated as number of oocysts detected (in millions) per gram of feces, and host relative weight is calculated as the percentage of weight compared to day 0. Mean and 95% CI are plotted for each parasite isolate. All mouse groups are pooled for each parasite isolate
TABLE 2.
Contingency table: number of mice and status at dpi 11 for each mouse group upon infection with Eimeria falciformis isolate Brandenburg88
| Mouse | Status at dpi 11 | ||
|---|---|---|---|
| Subspecies | Group | Alive | Dead |
| Mmd | SCHUNT | 6 | 0 |
| Mmd | STRA | 7 | 0 |
| Mmd | SCHUNTxSTRA | 8 | 0 |
| Mmd‐Mmm | STRAxBUSNA | 8 | 0 |
| Mmd‐Mmm | SCHUNTxPWD | 6 | 0 |
| Mmm | PWDxBUSNA | 4 | 3 |
| Mmm | BUSNA | 3 | 4 |
| Mmm | PWD | 1 | 6 |
| Total | 43 | 13 | |
3.2. Comparison of resistance–tolerance coupling between E. ferrisi and E. falciformis
3.2.1. Differences in resistance and tolerance between mouse groups depends on the parasite
Considering all mice infected with either E. ferrisi isolate Brandenburg 64 and E. falciformis isolate Brandenburg 88, we found our proxy for (inverse of) resistance (maximum number of OPG) to be statistically different between mouse groups, parasite isolates, and their interaction (LRT: mouse groups: G = 55.5, df = 28, p < 0.01; parasite isolates: G = 40.5, df = 16, p < 0.001; interaction: G = 27.9, df = 14, p = 0.015). Results were similar for our proxy for tolerance (LRT: mouse groups: G = 28.4, df = 14, p = 0.01; parasite isolates: G = 20.1 df = 8, p = 0.01; interaction: G = 18.8, df = 7, p < 0.01). Our proxy for impact on weight (maximum relative weight loss) was significantly different between mouse groups and parasite isolates, but not for their interaction (LRT: mouse groups: G = 44.9, df = 14, p < 0.001; parasite isolates: G = 33, df = 8, p < 0.001; interaction: G = 7.5, df = 7, p = 0.38). For the latter model, impact on weight, post hoc tests showed that the only statistical differences between two mouse groups within a parasite infection were found in E. falciformis infection, between PWD and STRA (Tukey HSD test, p‐value = 0.02), PWD and STRAxBUSNA (Tukey HSD test, p‐value = 0.03), and PWD and SCHUNTxPWD (Tukey HSD test, p‐value = 0.02). No difference was found within one mouse group between the two parasite isolates at the 0.05 significance threshold.
We found that the mean predicted number of OPG varies with the mean predicted relative weight loss (LRT: G = 10, df = 2, p < 0.01) and differs between both parasites (LRT: G = 8.9, df = 2, p = 0.012), and more importantly we found a significant interaction term (LRT: G = 8.3, df = 1, p < 0.01). This means that the relationship between mean health effect and mean resistance differs between the two Eimeria species infections. Then, we performed a linear regression of the mean predicted tolerance for each eight mouse groups as a response of the mean predicted OPG, parasite isolate, and their interaction. In this case, we found that the mean number of OPG varies along with tolerance (LRT: G = 8.5, df = 2, p = 0.01) but does not statistically differ between both parasites (LRT: G = 1.1, df = 2, p = 0.57), and the interaction term was not found significant (LRT: G = 0.03, df = 1, p = 0.86). In this respect, the correlation between resistance and tolerance was not found to significantly differ between both parasites. Following these results, we looked at the coupling of resistance and tolerance within each of the two isolates.
3.2.2. Resistance and tolerance to E. ferrisi isolate Brandenburg64 are uncoupled
We tested coupling between resistance and tolerance for E. ferrisi isolate Brandenburg64 in our eight mouse groups. First, we tested whether our proxies for resistance and tolerance were different between the mouse groups. We found the maximum number of OPG to be statistically different between mouse groups (LRT: G = 26.6, df = 7, p < 0.001; Figure 3a). Tolerance was not found to significantly differ between mouse groups for this parasite isolate (LRT: G = 6.8, df = 7, p = 0.45; Figure 3b).
FIGURE 3.
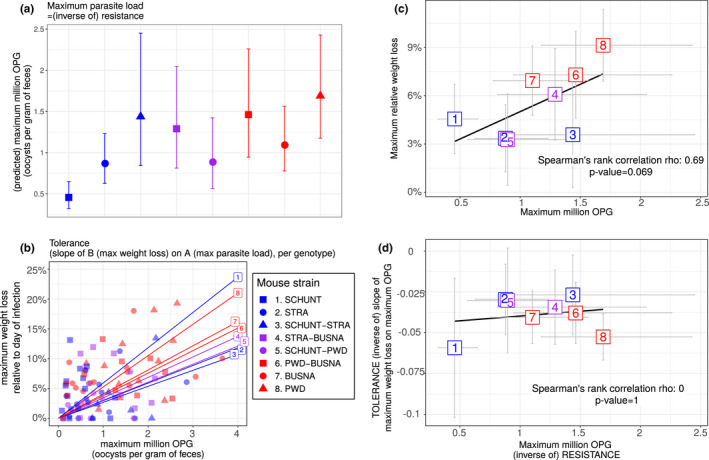
No indication of resistance–tolerance coupling forEimeria ferrisiisolate Brandenburg64. Colors represent mouse subspecies (blue:Mus musculus domesticus, red:Mus musculus musculus, purple: Mmd‐Mmm). Left side: comparison of maximum oocysts per gram of feces used as a proxy for (inverse of) resistance (a) and tolerance (b) between mouse groups estimated by the slope of the linear regression with null intercept modeling maximum relative weight loss as a response of maximum oocysts per gram of feces, a steep slope corresponding to a low tolerance. Maximum number of OPG differs between mouse groups, but tolerance is similar. Right side: nonsignificant positive correlation between mean maximum oocysts per gram of feces and mean relative weight loss (c) and absence of correlation between maximum oocysts per gram of feces used as a proxy for (inverse of) resistance and tolerance (d); grey error bars represent 95% confidence intervals. Our results do not support coupling between resistance and toleranceEimeria ferrisiisolate Brandenburg64
We found a nonsignificant positive correlation between resistance (inverse of maximum number of OPG) and impact on health (maximum weight loss) (Spearman's ρ = 0.69, p = 0.07, N = 8; Figure 3c). Moreover, we did not find a correlation between resistance (inverse of maximum number of OPG) and tolerance (inverse of slope of maximum weight loss on maximum OPG) (Spearman's ρ = 0, p = 1, N = 8; Figure 3d).
In conclusion, we did not find indications of resistance–tolerance coupling for E. ferrisi isolate Brandenburg64, the different mouse groups infected by this parasite presenting a similar level of tolerance while showing an effect of quantitative resistance on health.
3.2.3. Coupling between resistance and tolerance to E. falciformis
We then tested coupling between resistance and tolerance for E. falciformis isolate Brandenburg88 in our eight mouse groups. First, we tested if our proxies for resistance and tolerance were different between the mouse groups. We found the maximum number of OPG to be statistically different between mouse groups (LRT: G = 28.6, df = 14, p = 0.012; Figure 4a). Contrary to our results on E. ferrisi isolate Brandenburg64, the tolerance slopes for E. falciformis isolate Brandenburg88 were different between mouse groups (LRT: G = 13.9, df = 7, p = 0.05; Figure 4b).
FIGURE 4.
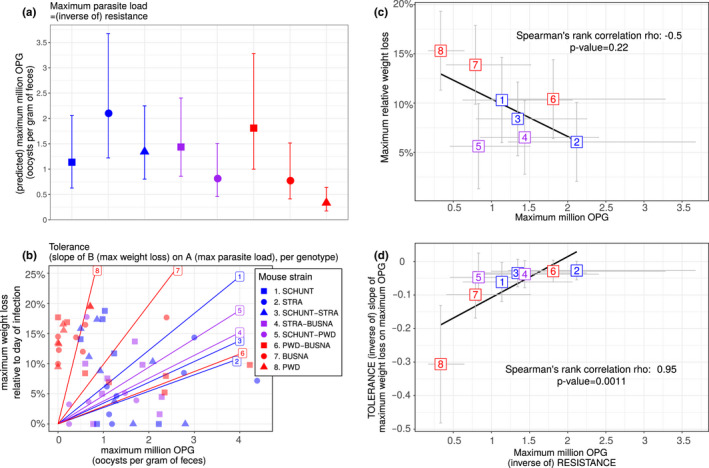
Coupling between resistance and tolerance forEimeria falciformisisolate Brandenburg88. Colors represent mouse subspecies (blue:Mus musculus domesticus, red:Mus musculus musculus, purple: Mmd‐Mmm). Left side: comparison of maximum oocysts per gram of feces used as a proxy for (inverse of) resistance (a) and tolerance between mouse groups estimated by the slope of the linear regression with null intercept modeling maximum relative weight loss as a response of maximum oocysts per gram of feces, a steep slope corresponding to a low tolerance (b). Maximum number of OPG and tolerance differ between mouse groups. Right side: nonsignificant negative correlation between mean maximum oocysts per gram of feces and mean relative weight loss (c) and strong positive correlation between maximum oocysts per gram of feces used as a proxy for inverse of resistance and tolerance (corresponding to a negative correlation between resistance and tolerance) (d); grey error bars represent 95% confidence intervals. Our results support coupling between resistance and toleranceEimeria falciformisisolate Brandenburg88
We detected a strong negative correlation between (inverse of) resistance (maximum number of OPG) and tolerance (inverse of slope of maximum weight loss on maximum OPG) (Spearman's ρ = −0.95, p = 0.001; Figure 4d). This result was robust to the exclusion of the extreme point corresponding to mouse strain PWD (point 8 in Figure 4d; Spearman's ρ = −0.93, p < 0.01).
We conclude that this correlation is unlikely a statistical artifact, as (a) mouse groups present statistically different values of resistance and tolerance (see 2.1) and (b) we found a (nonsignificant) negative correlation between resistance (inverse of maximum number of OPG) and impact on health (maximum weight loss) (Spearman's ρ = −0.5, p = 0.22; Figure 4c), indicating that mouse groups losing more weight also shed less parasites.
We conclude that our results indicate the presence of negative resistance–tolerance coupling for E. falciformis isolate Brandenburg88.
4. DISCUSSION
In this study, we assessed resistance and tolerance to two closely related parasites, E. ferrisi and E. falciformis, in four mouse strains and their intra‐ and intersubspecific hybrids. Understanding this coupling has two major implications:
From a practical "measurement" perspective, we can ask whether tolerance can be predicted from resistance, as the latter is easier to measure (e.g., in field sampling). Many studies assess the impact of parasites on host fitness based on resistance. If, as we found in the present study, resistance and tolerance are decoupled, this can be misleading. In our host system, the house mice, for example, it has been shown that hybrids between M. m. domesticus and M. m. musculus are more resistant to parasites (Baird et al., 2012; Balard et al., 2020), including Eimeria, but tolerance could not be measured under natural conditions (Balard et al., 2020). The effect of parasites on host fitness in the evolution of the house mouse hybrid zone is thus still rather ambiguous (Baird & Goüy de Bellocq, 2019). We show that careful distinction between parasite species is necessary when analyzing parasite–host interaction (see also Jarquín‐Díaz et al., 2019) and that it is indispensable to measure both resistance and tolerance in Eimeria infections of house mice.
In this work, we used the concept of tolerance as used originally in the plant literature and later on transferred to animal studies (Fineblum & Rausher, 1995). This concept of tolerance can be criticized, as it links tolerance mathematically to resistance. Nevertheless, we argue that this view is biologically meaningful considering resistance and tolerance as a double defense system, one step limiting the parasite multiplication, the other limiting the impact of this multiplication on fitness‐related traits. To limit the possibility of statistical artifact, our approach did not only consist in calculating correlations between resistance and tolerance, but also in testing differences in resistance, impact on health and tolerance. Of note, a positive correlation between mean health effect and mean resistance of each host strains could indicate some host strains having few parasites–few effects on health, and others more parasites–more effects on health; this configuration would limit the possibility of detecting an actual resistance–tolerance trade‐off by lack of a full range of resistance values. For this reason, our approach consisted in testing the "coupling" between resistance and tolerance, that is, (a) a genotype‐level correlation between tolerance and resistance additionally supported by (b) the absence of positive correlation between health effect and resistance. We argue that this additional step increases the confidence in the presence of a biologically meaningful negative correlation between resistance and tolerance, likely implying a trade‐off.
Differences between parasite species could explain the evolution of different strategies: E. ferrisi commits to sexual reproduction after a relatively short time with few cycles of asexual expansion (Al‐khlifeh et al., 2019; Ankrom et al., 1975), while E. falciformis has a relatively longer life cycle (Al‐khlifeh et al., 2019; Haberkorn, 1970). As E. ferrisi infections do not reach extremely high intensities, high tolerance might be the optimal strategy for both house mouse subspecies. Resistance could then evolve relatively freely without any major impact of the parasite on the hosts’ health. In the case of E. falciformis, the long life cycle might lead to high tissue load. Tissue damage is observed during sexual reproduction for this parasite (Ehret et al., 2017) and might mean that a certain level of resistance is required. On the other hand, immunopathology has been observed in advanced E. falciformis infections (Stange et al., 2012). These intrinsic characteristics of E. falciformis might lead to multiple different optima for resistance and tolerance, leading to a trade‐off.
More generally, from an evolutionary perspective, coupling between resistance and tolerance might help determine whether coevolution between host and parasite can be expected: A host‐parasite system in which one finds negative coupling between tolerance and resistance would be an especially promising system for studies of host–parasite coevolution. Indeed, coevolution in host–parasite systems is often assumed but rarely proven (Woolhouse et al., 2002). Janzen (1980) notes that not all parasite–host systems are coevolving. The presence of efficient host defenses against a given parasite is not necessarily produced in response to this parasite specifically and the parasite does not necessarily respond specifically. In the mouse‐E. ferrisi system, where resistance and tolerance are decoupled, host and parasite fitness might be decoupled as a result, making host–parasite coevolution less likely. In the mouse–E. falciformis system, we found a negative coupling between tolerance and resistance, making coevolution between host and parasite more likely.
In conclusion, we show that the coupling between resistance and tolerance can differ between closely related parasite species and we argue that this trait of a host–parasite system determines the questions to be best approached with a particular parasite.
CONFLICT OF INTEREST
This work is original and has not been published elsewhere, nor is it currently under consideration for publication elsewhere, we have no conflicts of interest to disclose, its submission for publication has been approved by all relevant authors and institutions, all persons entitled to authorship have been so named, and all authors have seen and agreed to the submitted version of the manuscript.
AUTHOR CONTRIBUTIONS
Alice Balard: Conceptualization (equal); data curation (equal); formal analysis (equal); investigation (equal); methodology (equal); project administration (equal); software (equal); validation (equal); visualization (equal); writing – original draft (equal); writing – review and editing (equal). Víctor Hugo Jarquín‐Díaz: Investigation (equal). Jenny Jost: Investigation (equal). Vivian Mittné: Formal analysis (equal); investigation (equal). Francisca Böhning: Formal analysis (equal); investigation (equal). Ľudovít Ďureje: Resources (equal). Jaroslav Piálek: Conceptualization (equal); resources (equal); writing – review and editing (equal). Emanuel Heitlinger: Conceptualization (equal); data curation (equal); funding acquisition (equal); project administration (equal); supervision (equal); writing – review and editing (equal).
Open Research Badges
This article has earned an Open Materials Badge for making publicly available the components of the research methodology needed to reproduce the reported procedure and analysis. All materials are available at https://doi.org/10.5281/zenodo.4122009.
Supporting information
Appendix S1
Appendix S2
Appendix S3
Supplementary Material
ACKNOWLEDGMENTS
This work was funded by the German Research Foundation (DFG) Grant [HE 7320/1‐1] to EH. VHJ is an associated student of GRK 2046 funded by the DFG. The maintenance of wild‐derived strains was supported by the CAS within the program of the Strategy AV 21 and the Czech Science Foundation (project 16‐23773S) to JP. Open access funding enabled and organized by ProjektDEAL.
Balard A, Jarquín‐Díaz VH, Jost J, et al. Coupling between tolerance and resistance for two related Eimeria parasite species. Ecol Evol. 2020;10:13938–13948. 10.1002/ece3.6986
DATA AVAILABILITY STATEMENT
Code and full data: Zenodo https://doi.org/10.5281/zenodo.4122009.
REFERENCES
- Al‐khlifeh, E. , Balard, A. , Jarquín‐Díaz, V. H. , Weyrich, A. , Wibbelt, G. , & Heitlinger, E. (2019). Eimeria falciformis BayerHaberkorn1970 and novel wild derived isolates from house mice: Differences in parasite lifecycle, pathogenicity and host immune reactions, bioRxiv, 611277 10.1101/611277 [DOI] [Google Scholar]
- Ankrom, S. L. , Chobotar, B. , & Ernst, J. V. (1975). Life cycle of Eimeria ferrisi Levine & Ivens, 1965 in the mouse, Mus musculus . The Journal of Protozoology, 22, 317–323. 10.1111/j.1550-7408.1975.tb05177.x [DOI] [Google Scholar]
- Ayres, J. S. , & Schneider, D. S. (2012). Tolerance of infections. Annual Review of Immunology, 30, 271–294. 10.1146/annurev-immunol-020711-075030 [DOI] [PubMed] [Google Scholar]
- Baird, S. J. E. , & Goüy de Bellocq, J. (2019). Shifting paradigms for studying parasitism in hybridising hosts: Response to Theodosopoulos, Hund, and Taylor. Trends in Ecology & Evolution, 34, 387–389. 10.1016/j.tree.2019.01.011 [DOI] [PubMed] [Google Scholar]
- Baird, S. J. E. , Ribas, A. , Macholán, M. , Albrecht, T. , Piálek, J. , & Goüy de Bellocq, J. (2012). Where are the wormy mice? A reexamination of hybrid parasitism in the European house mouse hybrid zone. Evolution, 66, 2757–2772. 10.1111/j.1558-5646.2012.01633.x [DOI] [PubMed] [Google Scholar]
- Balard, A. , Jarquín‐Díaz, V. H. , Jost, J. , Martincová, I., Ďureje, Ľ. , Piálek, J. , Macholán, M. , de Bellocq, J. G. , Baird, S. J. E. , & Heitlinger, E. (2020). Intensity of infection with intracellular Eimeria spp. and pinworms is reduced in hybrid mice compared to parental subspecies. Journal of Evolutionary Biology, 33, 435–448. 10.1111/jeb.13578 [DOI] [PubMed] [Google Scholar]
- Baker, D. G. (1998). Natural pathogens of laboratory mice, rats, and rabbits and their effects on research. Clinical Microbiology Reviews, 11(2), 231–266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baucom, R. S. , & de Roode, J. C. (2011). Ecological immunology and tolerance in plants and animals. Functional Ecology, 25, 18–28. 10.1111/j.1365-2435.2010.01742.x [DOI] [Google Scholar]
- Boots, M. , Best, A. , Miller, M. R. , & White, A. (2008). The role of ecological feedbacks in the evolution of host defence: What does theory tell us? Philosophical Transactions of the Royal Society B: Biological Sciences, 364, 27–36. 10.1098/rstb.2008.0160 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brett, M. T. (2004). When is a correlation between non‐independent variables “spurious”? Oikos, 105, 647–656. 10.1111/j.0030-1299.2004.12777.x [DOI] [Google Scholar]
- Carval, D. , & Ferriere, R. (2010). A unified model for the coevolution of resistance, tolerance, and virulence. Evolution, 64, 2988–3009. 10.1111/j.1558-5646.2010.01035.x [DOI] [PubMed] [Google Scholar]
- Chapman, H. D. , Barta, J. R. , Blake, D. , Gruber, A. , Jenkins, M. , Smith, N. C. , Suo, X. , & Tomley, F. M. (2013). Chapter two – A selective review of advances in coccidiosis research. Advances in Parasitology, 83, 93–171. 10.1016/B978-0-12-407705-8.00002-1 [DOI] [PubMed] [Google Scholar]
- Clerc, M. , Fenton, A. , Babayan, S. A. , & Pedersen, A. B. (2019). Parasitic nematodes simultaneously suppress and benefit from coccidian coinfection in their natural mouse host. Parasitology, 146, 1096–1106. 10.1017/S0031182019000192 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delignette‐Muller, M. L. , & Dutang, C. (2015). Fitdistrplus: An r package for fitting distributions. Journal of Statistical Software, 64, 1–34. 10.18637/jss.v064.i04 [DOI] [Google Scholar]
- Ďureje, Ľ. , Macholán, M. , Baird, S. J. E. , & Piálek, J. (2012). The mouse hybrid zone in Central Europe: From morphology to molecules. Journal of Vertebrate Biology, 61, 308–318. 10.25225/fozo.v61.i3.a13.2012 [DOI] [Google Scholar]
- Ehret, T. , Spork, S. , Dieterich, C. , Lucius, R. , & Heitlinger, E. (2017). Dual RNA‐seq reveals no plastic transcriptional response of the coccidian parasite Eimeria falciformis to host immune defenses. BMC Genomics, 18, 686 10.1186/s12864-017-4095-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fineblum, W. L. , & Rausher, M. D. (1995). Tradeoff between resistance and tolerance to herbivore damage in a morning glory. Nature, 377, 517–520. 10.1038/377517a0 [DOI] [Google Scholar]
- Floyd, R. M. , Rogers, A. D. , Lambshead, P. J. D. , & Smith, C. R. (2005). Nematode‐specific PCR primers for the 18S small subunit rRNA gene. Molecular Ecology Notes, 5, 611–612. 10.1111/j.1471-8286.2005.01009.x [DOI] [Google Scholar]
- Gandon, S. , & Michalakis, Y. (2000). Evolution of parasite virulence against qualitative or quantitative host resistance. Proceedings of the Royal Society of London. Series B: Biological Sciences, 267, 985–990. 10.1098/rspb.2000.1100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graham, A. L. , Allen, J. E. , & Read, A. F. (2005). Evolutionary causes and consequences of immunopathology. Annual Review of Ecology, Evolution, and Systematics, 36, 373–397. 10.1146/annurev.ecolsys.36.102003.152622 [DOI] [Google Scholar]
- Gregorová, S. , & Forejt, J. (2000). PWD/Ph and PWK/Ph inbred mouse strains of Mus m. musculus subspecies–a valuable resource of phenotypic variations and genomic polymorphisms. Folia Biologica, 46, 31–41. [PubMed] [Google Scholar]
- Haberkorn, A. (1970). Die Entwicklung von Eimeria falciformis (Eimer 1870) in der weißen Maus (Mus musculus). Zeitschrift Für Parasitenkunde, 34, 49–67. 10.1007/BF00629179 [DOI] [Google Scholar]
- Howick, V. M. , & Lazzaro, B. P. (2017). The genetic architecture of defence as resistance to and tolerance of bacterial infection in Drosophila melanogaster . Molecular Ecology, 26, 1533–1546. 10.1111/mec.14017 [DOI] [PubMed] [Google Scholar]
- Jackman, S. (2020). pscl: Classes and methods for R developed in the political science computational laboratory. United States Studies Centre, University of Sydney, Sydney, New South Wales, Australia; R package version 1.5.5, https://github.com/atahk/pscl/ [Google Scholar]
- Janzen, D. H. (1980). When is it coevolution? Evolution, 34, 611–612. 10.1111/j.1558-5646.1980.tb04849.x [DOI] [PubMed] [Google Scholar]
- Jarquín‐Díaz, V. H. , Balard, A. , Jost, J. , Kraft, J. , Dikmen, M. N. , Kvičerová, J. , & Heitlinger, E. (2019). Detection and quantification of house mouse Eimeria at the species level – Challenges and solutions for the assessment of coccidia in wildlife. International Journal for Parasitology: Parasites and Wildlife, 10, 29–40. 10.1016/j.ijppaw.2019.07.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klemme, I. , & Karvonen, A. (2016). Vertebrate defense against parasites: Interactions between avoidance, resistance, and tolerance. Ecology and Evolution, 7, 561–571. 10.1002/ece3.2645 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kutzer, M. A. M. , & Armitage, S. A. O. (2016). Maximising fitness in the face of parasites: A review of host tolerance. Zoology, 119, 281–289. 10.1016/j.zool.2016.05.011 [DOI] [PubMed] [Google Scholar]
- Lefèvre, T. , Williams, A. J. , & de Roode, J. C. (2010). Genetic variation in resistance, but not tolerance, to a protozoan parasite in the monarch butterfly. Proceedings of the Royal Society B: Biological Sciences, 278, 751–759. 10.1098/rspb.2010.1479 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lüdecke, D. (2018). Ggeffects: Tidy data frames of marginal effects from regression models. Journal of Open Source Software, 3, 772 10.21105/joss.00772 [DOI] [Google Scholar]
- Macholán, M. , Baird, S. J. E. , Fornůsková, A. , & Martincová, I. , Rubík, P. , Ďureje, Ľ. , Heitlinger, E. , & Piálek, J. (2019). Widespread introgression of the Mus musculus musculus Y chromosome in Central Europe. bioRxiv. 10.1101/2019.12.23.887471 [DOI] [Google Scholar]
- Mehlhorn, H. , Schmahl, G. , Frese, M. , Mevissen, I. , Harder, A. , & Krieger, K. (2005). Effects of a combinations of emodepside and praziquantel on parasites of reptiles and rodents. Parasitol Res, 97, S65–S69. 10.1007/s00436-005-1446-z [DOI] [PubMed] [Google Scholar]
- Martincová, I. , Ďureje, Ľ. , Kreisinger, J. , Macholán, M. , & Piálek, J. (2019). Phenotypic effects of the Y chromosome are variable and structured in hybrids among house mouse recombinant lines. Ecology and Evolution, 9, 6124–6137. 10.1002/ece3.5196 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazé‐Guilmo, E. , Loot, G. , Páez, D. J. , Lefèvre, T. , & Blanchet, S. (2014). Heritable variation in host tolerance and resistance inferred from a wild host–parasite system. Proceedings of the Royal Society B: Biological Sciences, 281, 20132567 10.1098/rspb.2013.2567 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Medzhitov, R. , Schneider, D. S. , & Soares, M. P. (2012). Disease tolerance as a defense strategy. Science, 335, 936–941. 10.1126/science.1214935 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mesa, J. M. , Scholes, D. R. , Juvik, J. A. , & Paige, K. N. (2017). Molecular constraints on resistance–tolerance trade‐offs. Ecology, 98, 2528–2537. 10.1002/ecy.1948 [DOI] [PubMed] [Google Scholar]
- Piálek, J. , Vyskočilová, M. , Bímová, B. , Havelková, D. , Piálková, J. , Dufková, P. , Bencová, V. , Ďureje, L. , Albrecht, T. , Hauffe, H. C. , Macholán, M. , Munclinger, P. , Storchová, R. , Zajícová, A. , Holáň, V. , Gregorová, S. , & Forejt, J. (2008). Development of unique house mouse resources suitable for evolutionary studies of speciation. Journal of Heredity, 99, 34–44. 10.1093/jhered/esm083 [DOI] [PubMed] [Google Scholar]
- R Development Core Team (2013). R: A language and environment for statistical computing. R Foundation for Statistical Computing; Retrieved from http://www.R‐project.org/ [Google Scholar]
- Råberg, L. , Graham, A. L. , & Read, A. F. (2009). Decomposing health: Tolerance and resistance to parasites in animals. Philosophical Transactions of the Royal Society B: Biological Sciences, 364, 37–49. 10.1098/rstb.2008.0184 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Råberg, L. , Sim, D. , & Read, A. F. (2007). Disentangling genetic variation for resistance and tolerance to infectious diseases in animals. Science, 318, 812–814. 10.1126/science.1148526 [DOI] [PubMed] [Google Scholar]
- Restif, O. , & Koella, J. C. (2004). Concurrent evolution of resistance and tolerance to pathogens. The American Naturalist, 164, E90–E102. 10.1086/423713 [DOI] [PubMed] [Google Scholar]
- Rose, M. E. , Hesketh, P. , & Wakelin, D. (1992). Immune control of murine coccidiosis: CD4+ and CD8+ T lymphocytes contribute differentially in resistance to primary and secondary infections. Parasitology, 105, 349–354. 10.1017/S0031182000074515 [DOI] [PubMed] [Google Scholar]
- Roy, B. A. , & Kirchner, J. W. (2000). Evolutionary dynamics of pathogen resistance and tolerance. Evolution, 54, 51–63. 10.1111/j.0014-3820.2000.tb00007.x [DOI] [PubMed] [Google Scholar]
- Schito, M. L. , Barta, J. R. , & Chobotar, B. (1996). Comparison of four murine Eimeria species in immunocompetent and immunodeficient mice. The Journal of Parasitology, 82, 255–262. 10.2307/3284157 [DOI] [PubMed] [Google Scholar]
- Schmid‐Hempel, P. (2013). Evolutionary parasitology: The integrated study of infections, immunology, ecology, and genetics. Oxford University Press; 10.1093/acprof:oso/9780199229482.001.0001 [DOI] [Google Scholar]
- Shaw, D. J. , & Dobson, A. P. (1995). Patterns of macroparasite abundance and aggregation in wildlife populations: A quantitative review. Parasitology, 111, S111–S133. 10.1017/S0031182000075855 [DOI] [PubMed] [Google Scholar]
- Sheldon, B. C. , & Verhulst, S. (1996). Ecological immunology: Costly parasite defences and trade‐offs in evolutionary ecology. Trends in Ecology & Evolution, 11, 317–321. 10.1016/0169-5347(96)10039-2 [DOI] [PubMed] [Google Scholar]
- Simms, E. L. (2000). Defining tolerance as a norm of reaction. Evolutionary Ecology, 14, 563–570. 10.1023/a:1010956716539 [DOI] [Google Scholar]
- Smith, A. L. , & Hayday, A. C. (2000). Genetic Dissection of primary and secondary responses to a widespread natural pathogen of the gut, Eimeria vermiformis . Infection and Immunity, 68, 6273–6280. 10.1128/IAI.68.11.6273-6280.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soares, M. P. , Teixeira, L. , & Moita, L. F. (2017). Disease tolerance and immunity in host protection against infection. Nature Reviews Immunology, 17, 83–96. 10.1038/nri.2016.136 [DOI] [PubMed] [Google Scholar]
- Stange, J. , Hepworth, M. R. , Rausch, S. , Zajic, L. , Kühl, A. A. , Uyttenhove, C. , Renauld, J.‐C. , Hartmann, S. , & Lucius, R. (2012). IL‐22 mediates host defense against an intestinal intracellular parasite in the absence of IFN‐γ at the cost of Th17‐driven immunopathology. Journal of Immunology, 188, 2410–2418. 10.4049/jimmunol.1102062 [DOI] [PubMed] [Google Scholar]
- Stowe, K. , Marquis, R. , Hochwender, C. , & Simms, E. L. (2000). The evolutionary ecology of tolerance to consumer damage. Annual Review of Ecology, Evolution, and Systematics, 31, 565–595. 10.1146/annurev.ecolsys.31.1.565 [DOI] [Google Scholar]
- Vale, P. F. , & Little, T. J. (2012). Fecundity compensation and tolerance to a sterilizing pathogen in Daphnia . Journal of Evolutionary Biology, 25, 1888–1896. 10.1111/j.1420-9101.2012.02579.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Venables, W. N. , & Ripley, B. D. (2002). Modern applied statistics with S (4th ed.). Springer; 10.1007/978-0-387-21706-2 [DOI] [Google Scholar]
- Wickham, H. (2016). Ggplot2: Elegant graphics for data analysis (2nd ed.). Springer; 10.1007/978-0-387-98141-3 [DOI] [Google Scholar]
- Woolhouse, M. E. J. , Webster, J. P. , Domingo, E. , Charlesworth, B. , & Levin, B. R. (2002). Biological and biomedical implications of the co‐evolution of pathogens and their hosts. Nature Genetics, 32, 569–577. 10.1038/ng1202-569 [DOI] [PubMed] [Google Scholar]
- Zeileis, A. , Kleiber, C. , & Jackman, S. (2008). Regression models for count data in R. Journal of Statistical Software, 27(8). http://www.jstatsoft.org/v27/i08/ [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Appendix S1
Appendix S2
Appendix S3
Supplementary Material
Data Availability Statement
Code and full data: Zenodo https://doi.org/10.5281/zenodo.4122009.