

Abstract
Adenomyosis remains an enigmatic disease in the clinical and research communities. The high prevalence, diversity of morphological and symptomatic presentations, array of potential etiological explanations, and variable response to existing interventions suggest that different subgroups of patients with distinguishable mechanistic drivers of disease may exist. These factors, combined with the weak links to genetic predisposition, make the entire spectrum of the human condition challenging to model in animals. Here, after an overview of current approaches, a vision for applying physiomimetic modeling to adenomyosis is presented. Physiomimetics combines a system's biology analysis of patient populations to generate hypotheses about mechanistic bases for stratification with in vitro patient avatars to test these hypotheses. A substantial foundation for three-dimensional (3D) tissue engineering of adenomyosis lesions exists in several disparate areas: epithelial organoid technology; synthetic biomaterials matrices for epithelial–stromal coculture; smooth muscle 3D tissue engineering; and microvascular tissue engineering. These approaches can potentially be combined with microfluidic platform technologies to model the lesion microenvironment and can potentially be coupled to other microorgan systems to examine systemic effects. In vitro patient-derived models are constructed to answer specific questions leading to target identification and validation in a manner that informs preclinical research and ultimately clinical trial design.
Keywords: adenomyosis, organoids, models of adenomyosis, tissue engineering, microfluidic device
Deconstructing Adenomyosis: What to Model?
In constructing any in vitro disease model, the first question is “what to model.” This question is often relatively easily answered for diseases with germ line genetic mutations and clearly defined clinical phenotypes directly linked to the molecular mutation, such as cystic fibrosis. In such cases, high throughput phenotypic screens in cell lines, based on correcting a known molecular defect, have been remarkably effective in identifying blockbuster clinical therapies. 1 Adenomyosis is arguably at the opposite end of the spectrum.
First, the clinical landscape is convoluted: adenomyosis is over 10 times more prevalent than most single-gene defect diseases; symptoms overlap and are comorbid with myriad other diseases and conditions; there is no consensus on the definition of “disease” nor a clearly defined set of metrics to diagnose it and assess response to therapy; and diagnostic bias toward older multiparous patients clouds true incidence. 2 3 4 5 Patients with pain, heavy menstrual bleeding, and/or infertility may have adenomyosis—or something else entirely—as an underlying cause. 6 About 40% of patients with symptoms, but no indication of adenomyosis on magnetic resonance imaging (MRI) or ultrasound, are found to have adenomyosis lesions upon histopathological assessment. 7 Symptom-targeting therapies for adenomyosis, like those for endometriosis, 8 work remarkably well in some patients, but are not at all effective or tolerated in others. Moreover, there is neither reliable manner to predict who will respond nor alternative therapies (other than developing coping mechanisms around eternal suffering) for those who do not. 9 10 As such, adenomyosis is not one disease, and it is unlikely there is one model that captures all salient features for all patients.
Second, the etiologies—given the prevalence and vast diversity in patient presentations, there is likely more than one—are obscure: adenomyosis appears to arise from a complex web of gene–environment interactions that may start in utero ; it might be triggered perinatally via seeding of the myometrium by cells that escape the endometrium; it might arise later in life due to uterine mechanical injury; or it might arise due to invasion of the myometrium by endometrium injured by an inflammatory insult. 2 4 11 12 13 This diversity of possible etiologies, along with the diversity of clinical presentations, underscores a shortcoming in how the disease research has traditionally been approached: adenomyosis is “a” disease. Yet, every patient is different. While there are not likely millions of individual diseases, there are likely multiple different constellations of molecular networks—immune networks, invasion networks, and metabolic networks—that go awry to instigate symptomatic and phenotypic appearance of ectopic endometrium in the myometrium, giving rise to groups of patients who could potentially be stratified molecularly as has been proposed for endometriosis. 8 9 14 15 16 17 Animal models—where strain variation, genetic perturbations, and targeted interventions to modulate specific pathways can be deployed to probe potential contributions to the adenomyotic etiology—offer insights into where to look for such mechanistic stratification in humans.
This last point brings us to the third big challenge with modeling adenomyosis—the relative lack of deep clinical and pathological phenotyping to guide patient stratification into tractable and mechanistically targetable subgroups. 5 Such clinical phenotyping and disease staging/classification is intertwined with therapeutic development through impacts on reimbursements for treatment 18 and evaluation of efficacy of various forms of treatment for clusters of patients with well-defined characteristics, as is done most thoroughly for cancer. 19 20 In this way, adenomyosis is the sister, though not precisely the twin, of endometriosis, which is inadequately staged based on lesion number, size, and location. 8 Like endometriosis, adenomyosis is currently only definitively diagnosed via surgical intervention (usually, a hysterectomy) and histopathological characterization. However, adenomyosis, as one of several pathologies that contribute to abnormal uterine bleeding (AUB), is included in the PALM-COEIN system for classifying such disorders, as first described in 2011 6 and updated in 2018. 21 PALM-COEIN serves as a crucial tool for patient–clinician–researcher communication; hence, it is a step toward personalized medicine for the patients suffering from AUB as a symptom of adenomyosis.
The state of personalized medicine for adenomyosis is still distant from as compared with cancer and other diseases. In this era of molecular stratification, oncologists routinely tailor treatments to a combination of physical features in addition to molecular biosignatures linked to disease mechanism and prognosis, sometimes using patient-derived organoids or tissue models to test drug sensitivity. 22 Mechanistic markers also inform the development of preclinical in vitro models, providing scientists an essential connection to the translational medicine. Such approaches are nascent only for endometriosis and even more primitive for adenomyosis, in part because somatic mutations, which are highly informative in cancer, are at best weakly associated with ectopic endometrial diseases. Hence, classifications based on protein or metabolic network states (or possibly epigenetics), complemented with clinical phenotypic data incorporating patient symptoms and life-long prognosis, 17 23 24 are therefore an appealing route to classification; such approaches will likely yield constellations of “molecular signatures” of disease state, rather than one single indicator of “diseased, or not.” 14 An exemplary model for how other noncancer immune-mediated disease communities have spurred such phenotyping is the Juvenile Diabetes Research Foundation-sponsored Network for Pancreatic Organ Donors with Diabetes (nPOD) program. 25 Application of programs like these would significantly improve our understanding of adenomyotic disease.
Finally, a fourth challenge is which stage(s) of disease to model. Modeling the features of disease as they present in diagnosed patients, who often have advanced varieties of disease progression, is arguably essential for identifying molecular targets and developing new therapies to treat these patients, and even more so for judicious prescription of existing therapies. Modeling proposed etiologies may yield preventive measures, or early treatment options, presuming a diagnostic for nascent stage disease, can be defined. The features of such models—and the throughputs required for getting useful information out of them—are somewhat but not entirely overlapping. Patients with advanced disease may also have nascent progressing disease, for example. Precise definition of the clinical phenotype, along with mechanistic hypotheses about the features of disease being studied, as they relate to the clinic , is an essential first step in designing, and then in implementing, in vitro models of adenomyosis.
A Roadmap of Phenomena to Model
How do we define what to model? Adenomyosis studies generally lack the kind of deep clinical phenotyping that links molecular and cellular pathological findings to patient symptoms, comorbidities, and treatment responses. However, several commonly observed pathological phenotypes can be modeled now using in vitro models, with a longer-term goal of more specialized patient avatars focusing on specific subgroups. The overall vision for the role of in vitro models is illustrated in Fig. 1 . Briefly, we envision that there are a relatively small number of subsets of patients who are phenomenologically similar, but distinct enough from each other that different therapeutic approaches are needed. Tissues and body fluids harvested from patients are used on the one hand for “omics” analysis—which can include genomics, proteomics, metabolomics—of molecular/cellular networks that enable stratification of patients into groups with data-informed machine learning methods, 14 based at least in part on mechanistic hypotheses that may distinguish the groups. Mechanistic stratification offers a path to target identification and experimental hypothesis testing. On the other hand, tissue banks created from the patients are used to create patient avatars that can be used to test hypotheses about mechanistic stratification and targets, with the goal of informing translational research and employing personalized therapies ( Fig. 1 ). In vitro models can thus potentially inform directions in clinical phenotyping, in an iterative fashion.
Fig. 1 Physiomimetic approach for developing targeted therapies for adenomyosis.
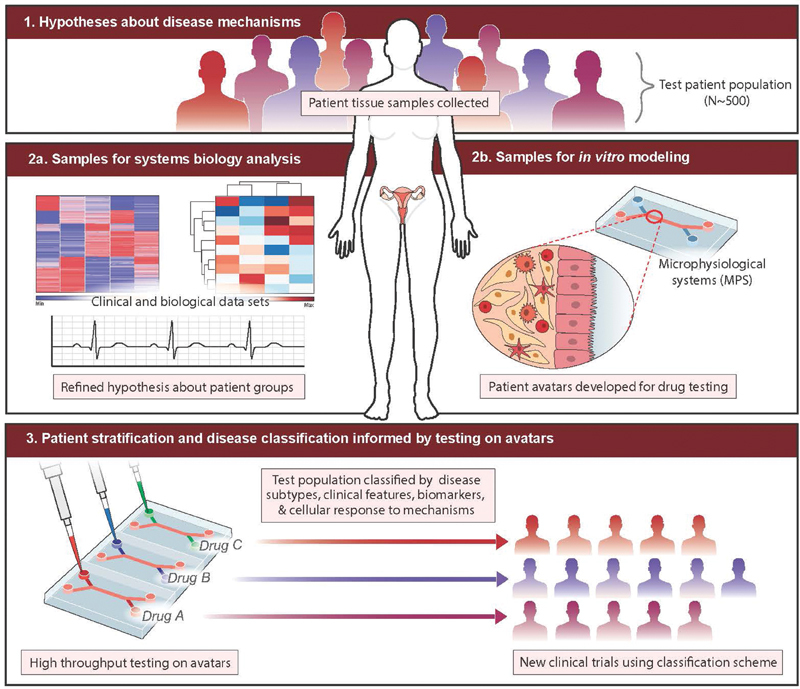
. Hypotheses regarding different mechanisms of disease that may be operative in patient subgroups are tested with tissue and fluid samples from a large patient population containing the subgroups (1); analysis of tissue and fluid samples to refine hypotheses about mechanism (2a and 3) are performed in tandem with development of cell banks and construction of patient avatars (2b). Mechanistic hypothesis about patient subgroups and interventions can then be tested in patient avatars representing the subgroups to define stratified clinical trials, or inform more judicious use of existing therapies.
Conceptualizing Models of Adenomyosis
“Physiomimetics” is the process of conceptualizing a complex physiological process to define essential features, then building in vitro models that capture the most relevant aspects of physiology in ways that can ultimately yield clinically actionable outcomes. Physiomimetic models encompass a range of experimental complexities, from relatively standard culture of individual cell types up through complex three-dimensional (3D) microfluidic models, driven by biological questions. We start here by conceptualizing established adenomyosis lesion phenotypes and the dynamic behaviors implicated in their pathophysiology, as the etiology of nascent adenomyotic lesions is still obscure, and there is great need to treat established adenomyotic lesions. Furthermore, the mechanisms contributing to disease progression may be operative to some extent in disease onset. We focus on diffuse adenomyosis lesions, as cystic adenomyosis lesions can in some instances be treated surgically. 26 Conceptually, here we define adenomyotic lesion as the ectopic presence of lesion-initiating cells —endometrial epithelial glands and stroma—surrounded by the lesion microenvironment comprising myometrial muscle cells, nerves, vasculature, and immune cells with associated extracellular matrix (ECM; Fig. 2 ).
Fig. 2.
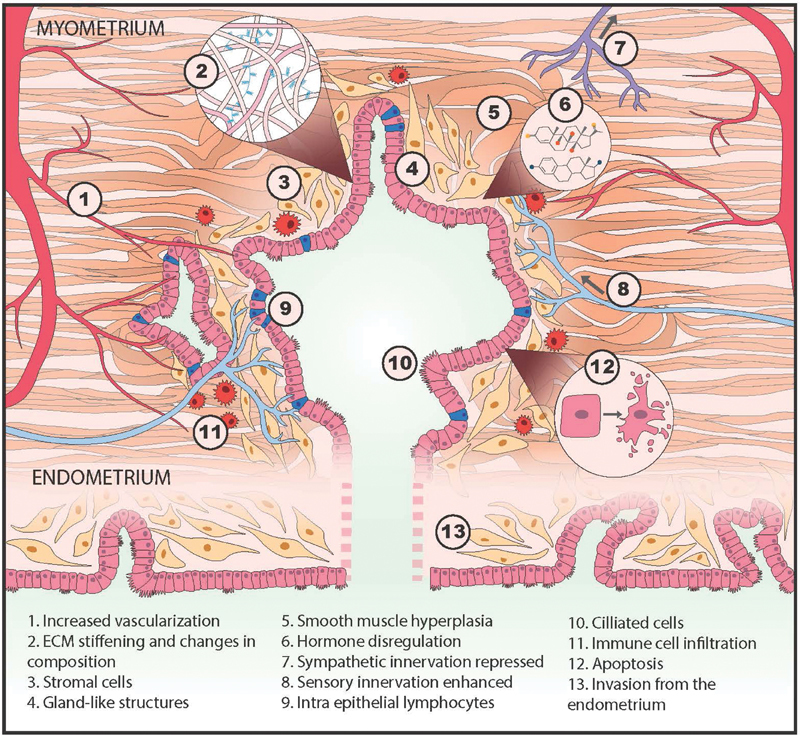
Conceptualization of an adenomyosis lesion, showing the biological components and pathological processes to consider in building an in vitro model.
Dynamic behaviors of lesion-initiating cells that are implicated in pathologies include (1) invasion into surrounding microenvironment, potentially seeding new, distant lesions; (2) proliferation , which may drive lesion growth, invasion, or feed into a hormone-driven cycle of cell death and proliferation; and (3) production of inflammatory cytokines, chemokines, proteases, and other modulators of signaling and remodeling of the local microenvironment 27 28 ( Fig. 2 ). The dynamic responses of the microenvironment associated with pathologies include (1) smooth muscle hyperplasia of local myometrial cells (or recruitment of fibroblasts or mesenchymal stem cells that acquire that phenotype 29 ); (2) influx and activation of immune cells; (3) recruitment of vasculature and modulation of vascular permeability, possibly with local bleeding and clotting; (4) enhancement of sensory innervation and regression of sympathetic innervation; and (5) stiffening of the local microenvironment, via matrix deposition and cellular changes. Cells within lesions respond to local cues, including mechanical stimulation from myometrial contractions, and to systemic cues, notably sex steroids, but are also modulated by nutrition, systemic inflammation, stress hormones, and other factors. These responses play out in an orchestrated fashion, as each cell type expresses a unique repertoire of receptors for the cues, including estrogen and progesterone receptors. 30 31 32 Many of the same phenomena are the subject of substantial investigation in carcinomas, but the spotlight on dynamic changes spurred by cyclic variation in sex steroids is omnipresent in adenomyosis. After a review of the known features of the myometrium and its changes in adenomyosis, we then consider how each of these dynamic phenomena can be modeled in vitro.
The Uterus in Adenomyosis
The presence of adenomyosis in the myometrium induces changes throughout the uterus in ways that suggest interesting hypotheses that can be tested using well-designed in vitro models. The myometrium is composed of the inner myometrium (or junctional zone, JZ), characterized as having a relatively greater cell density with relatively less ECM, and the outer myometrium, which is more ECM-rich and has a lower cell density. 33 On a cellular level, the myometrium comprises layers of smooth muscle cells (SMCs) arranged into fibular bundles approximately 0.3 mm in diameter, interwoven with blood vessels and connective, lymphatic, and nerve tissue. 33 34 These layers of SMC bundles are oriented in different directions around the uterus to enable complex contractile patterns that drive fluid flow in the nonpregnant uterus. 33 In adenomyosis, these contractile functions can become dysregulated. 35 36 Like the endometrium, the inner myometrium/JZ exhibits molecular and cellular changes in response to cyclic hormones through the menstrual cycle—such as tissue thickening and modulation of estrogen and progesterone receptor expression—but the outer myometrium appears less sensitive to menstrual cycle hormonal changes at both the gross anatomical level and the molecular level. 37 38 39 How these differences translate into the propensity for adenomyosis lesions to drive symptoms is unknown, but it is a potential facet that could be modeled in vitro. The healthy human myometrium is typically well delineated from the endometrium, but this clear delineation is blurred in adenomyosis, suggesting invasion of cells from the endometrium into the myometrium, attributed by one theory to microtraumas at the endometrial–myometrial interface during uterine peristalsis. 11 38 40 41 42 43
Changes in ECM architecture and composition, as well as smooth muscle hyperplasia of the region immediately surrounding lesions, characteristically affect multiple cell types and may involve chemical and mechanical cues. Patients with adenomyosis have been shown to have increased levels of matrix metalloproteinases (MMP)-2 and -9, suggesting abnormal matrix remodeling may be present in these patients. 44 Inflammation may be driven by repeated local bleeding in the lesion, 45 which may further enhance contractile phenotypes that in turn drive fibrogenic processes. 46
Endometrial cells in adenomyotic tissue exhibit many aberrant behaviors, including apparent resistance to progesterone, epithelial–mesenchymal transitions, decreased apoptosis regardless of menstrual phase, and other processes. 11 Although adenomyosis-targeting therapeutic approaches are intended to suppress aberrant proliferation of endometrial-derived tissues by targeting proestrogenic pathways or inducing progesterone action, 11 these approaches fail in some patients, a finding consistent with dysregulation of specific hormone receptors ESR1, ESR2, and PGR isoforms A and B in adenomyosis. 47 48 The downstream consequences of estrogenic signaling and acquired progesterone resistance that may contribute to the etiology of disease are still not well understood, and in vitro models offer an opportunity to parse the individual mechanisms.
Adenomyosis is associated with both pain and heavy menstrual bleeding. Several studies have reported an increased sensory nerve fiber density in both ectopic lesions and eutopic endometrium in women with endometriosis , along with reduction in sympathetic nerves, suggesting a possible diagnostic criterion. 49 50 51 52 Nerve fibers are increased in the fibrotic endometriosis nodules in the rectovaginal space and greater nerve fiber infiltration of lesions is found in patients with greater pain. 53 54 55 Increased nerve fiber density is also associated with an increase in macrophages, 56 with some evidence of estrogen modulation of the crosstalk. 57 Similar analysis of nerve fiber and macrophage density in adenomyosis is relatively scarce, but one study examined nerve fiber density in the endometrium and myometrium of women with adenomyosis and fibroids, and found no difference. 58
The immune environment in the uterus is dynamic, with dramatic increases in the number of macrophages, uterine natural killer (NK) cells, and other immune components during the secretory phase. The extent to which the increased number of immune cells arises from in situ proliferation of tissue-resident cells or from recruitment of circulating monocytes is still unclear. 59 60 61 Furthermore, whether CD45+ intraepithelial lymphocytes (IELs) that are characteristically observed in the epithelial layer in adenomyosis lesions 62 derive from locally recruited cells (and are therefore part of the microenvironment) or derive from IELs present in the epithelial layer of the eutopic endometrium (and are therefore lesion-initiating cells) is unknown. The crosstalk between nerves and macrophages or other immune cells may contribute to pain and/or bleeding. An intriguing clinical finding, albeit from a small study, examined macrophage density in the uteri of 54 patients undergoing hysterectomy for adenomyosis due to pain as the primary symptom, with a comparison group of 20 women undergoing hysterectomy for adenomyosis with heavy menstrual bleeding as the primary symptom, 63 where adenomyosis was tentatively diagnosed by both transvaginal ultrasound and MRI prior to surgery. Greater leukocyte infiltration was seen in both the myometrium and endometrium of patients with pain, and groups of macrophages were seen near lesions and interspersed in the muscle in the patients with pain; in both groups, there were macrophages around blood vessels, as macrophages are known to regulate vessel permeability. 64 Although the detailed interactions between the vasculature and adenomyotic lesions are not fully appreciated in the establishment and progression of adenomyosis, increased microvessel density has been observed in adenomyosis, 65 66 and a recent meta-analysis of studies examining vascular morphology and marker expression in the myometrium and endometrium suggests increased angiogenesis in the endometrium of women with adenomyosis. 67 Early studies that explored immune responses in adenomyosis showed that the disruption of the JZ at the endometrial–myometrial interface may result from the abnormal aggregation of macrophages in the myometrium, similar to the increased number of macrophages previously observed in endometriosis patients. 68 Finally, intriguing data on sex hormone responsiveness of mast cells, which are among the most common immune cell types in the myometrium and implicated in the pain associated with interstitial cystitis, may contribute to adenomyosis symptoms. 69 These observations suggest that recapitulating immune–epithelial–neuronal interactions in vitro using patient-specific cells may yield insights between symptomology and molecular and cellular behaviors characteristic of distinct patient populations.
Interestingly, the eutopic endometrium of patients with adenomyosis confer dysregulated angiogenesis, innervation, immune cell presence, and gene and protein expression that suggest greater invasive and survival potential at ectopic sites and contribution to symptoms. 70 71 72 73 74 Global changes in the myometrium outside the peri-lesion environment, including smooth muscle hypertrophy, increased stiffness, and immune cell infiltration, 38 75 raise questions about whether these global changes precede, or are the result of, lesion establishment—a hypothesis that could potentially be tested in vitro.
Designing, Building, and Interpreting In Vitro Models
The growing evidence that animal models do not capture essential features of human diseases involving chronic inflammation, sex steroid signaling, or other complex, molecular phenomena is driving development of sophisticated in vitro models that capture complex tissue architecture, and even dynamic perfusion of microvascular networks, using human patient-derived cells. While these approaches are just now emerging, especially driven by applications in cancer, cardiovascular, and liver diseases, they offer promise for modeling adenomyosis. In this section, we review basic approaches to tissue engineering of the uterine environment, including discussion of cell sourcing and 3D model design. Then, in the next section, we turn to modeling specific phenotypic facets of adenomyosis including invasion, vascular and systemic interactions, mechanics, and innervation, drawing from impactful work in disparate tissue systems.
Basic Approaches in Tissue Engineering of the Endometrium and Myometrium
The functions of the uterus arise from the integrated actions of epithelia, stromal fibroblasts, myometrial smooth muscle, immune cells, and endothelial cells, which are each directly and indirectly responsive to variations in sex hormones. Thus, many responses of various cell types to hormone changes are indirect and governed by dynamic communications with other cells. The endometrial epithelial response to progesterone, for example, is governed by signaling proteins produced by the progesterone-responsive stromal fibroblasts, which in turn respond to endothelial-derived signals to enhance the decidualization process. 31 76 While parsing responses of individual cell types to hormonal and other perturbations in vitro is an essential first step to understanding the integrated responses, building models that capture the dynamic interlinked hormone responsiveness in vitro is ultimately essential for illuminating complex phenomena like adenomyosis.
Cell Sources: Explants, Cell Lines, Primary Cells
For decades, explant cultures of the endometrium and myometrium were the only external window into the functions of the human uterus in vitro 36 77 78 79 80 81 and, until recent advances in primary cell culture, they remained preferred models for analysis of complex primary tissue functions. 31 81 82 Endometrial explants in the absence of ECM quickly deteriorate, 83 but can undergo outgrowth and remodeling when embedded in a fibrin matrix. 84 For the endometrium—the presumed instigator of adenomyosis lesions—definition of protocols for isolating and culturing epithelial cells and stromal fibroblasts from endometrial biopsies 85 enabled mechanistic studies on patient-derived samples, allowing diverse healthy and diseased donors to be combined in different ways to parse contributions of each cell type to health and disease. Primary cells remain the gold standard for mechanistic disease studies, but permanent epithelial 86 87 and stromal 88 89 90 cell lines that exhibit many (but not all) tissue-specific hormone-responsive features, and some disease states, have been routinely used for protocol development, pilot studies, and investigations of general features of endometrial cell behavior. The carcinoma-derived Ishikawa cell line is most commonly used for “normal” epithelia, 91 92 93 the h-TERT-immortalized endometriosis-derived “12Z” line is used for invasive endometrial epithelia, 15 87 94 and the h-TERT-immortalized stromal cell line “tHESC” 89 91 is used for endometrial stromal cells. (Caution: cell line contamination was reported for the endometrial epithelial cell line “HES” 86 ; hence, routine SNP profiling of cell lines is recommended for these, as it is with all cell lines, to validate provenance.) While cell lines offer convenience and relatively good reproducibility, including from laboratory to laboratory (subject to the typical cell line drift), they have some shortcomings for reproducing primary cell behaviors, 91 including greatly skewed cytokine and growth factor production profiles. 95 Pilot studies with cell lines can, however, offer insights that can be investigated further in more difficult-to-culture primary cells. 15
The majority of studies on primary endometrial cells focus on stromal cells, as they can readily be expanded in standard culture and frozen to create tissue banks, allowing repeated studies from the same donor within limits of passage number. 96 97 For years, however, investigators were limited by the lack of robust protocols for expansion and cryopreservation of endometrial epithelial cells, with few laboratories reporting success. 98 The landscape changed dramatically in 2017 with publication of the first two studies describing robust expansion and cryopreservation of human endometrial epithelial cells as organoids ( Fig. 3a ), with retention of hormone responsiveness, cellular architecture, polarity, heterogeneity, and other functions, 99 100 using modifications of protocols first described by Clevers and coworkers for expansion of human intestinal epithelia. 101 In the Clevers approach, epithelial cells are either dispersed as individual cells or fragments and cultured in Matrigel, a basement membrane-rich isolate of a tumor cell line propagated in mice. 101 While Matrigel had long been used as an ECM for 3D culture of endometrial glands 99 102 and many other types of epithelial cells, enabling remarkable retention of tissue phenotype, Clevers defined a cocktail of growth factors and other medium supplements that spur dramatic proliferation of the stem cell compartment, allowing the cells in an initial biopsy to expand orders of magnitude in culture and withstand cryopreservation, while preserving their ability to differentiate into proper tissue phenotypes upon a change in medium composition to remove stem cell cues. 101 This organoid approach has also been applied to derive continuously propagated organoid cultures from ectopic endometrium and endometrial tumors. 103
Fig. 3 Development of 3D in vitro models using synthetic hydrogels and endometrial epithelial organoids technologies.
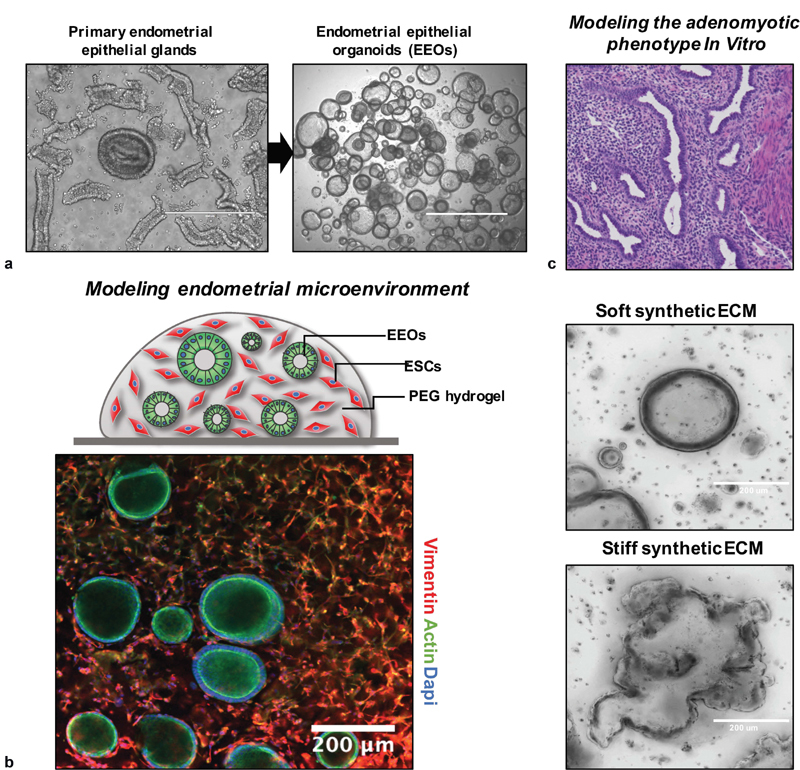
. ( a ) Endometrial epithelial organoids (EEOs) promote the culture, expansion, and propagation of epithelial cells using 3D hydrogel systems. EEOs retain epithelial structure, heterogeneity, and function of the native tissue glands. ( b ) Bio-labile, synthetic extracellular matrices (ECMs) can be designed to establish EEOs cultures and cocultures that include additional cell populations, such as endometrial stromal cells (ESCs). ( c ) Polyethylene glycol (PEG)-derived hydrogels are fully defined, modular, and can be tuned by modifying their molecular and biophysical properties to mimic key features of the adenomyotic and endometriotic phenotype.
An additional advance essential to building and characterizing physiologically relevant 3D models is the identification of both mesenchymal and epithelial stem and progenitor cell compartments within the human endometrium. 96 104 105 106 Intriguingly, recent data suggest that stem or at least progenitor cells are present in the endometrial functional layer and luminal regions of the endometrium, as evidenced by in situ hybridization for expression of the epithelial stem cell marker LGR5 . 107 The tools to identify putative stem and progenitor cells are valuable in assessing the phenotypic states of cells that are used to initiate models, and to assess their phenotypes under long-term culture conditions, especially to test hypotheses regarding possible contributions of these stem cells to ectopic adenomyosis lesions. 108 Related to efforts to identify and characterize functions of stem and progenitor cells in postnatal endometrium are efforts to create uterine tissue de novo from induced pluripotent stem cells (iPSCs), which could potentially advance patient-specific disease modeling as they have for other diseases. Encouragingly, a protocol for creation of human stromal cells with responses to progesterone characteristic of endometrial stromal cells undergoing decidualization in vitro has been developed, 109 providing a foundation for future efforts to reproduce other uterine tissues.
To model the pathogenesis of adenomyosis, it is important to consider its ectopic environment in addition to the lesion-initiating cells. The myometrium comprises several cell types, but the major focus in in vitro models is on myometrial SMCs, which have different properties in the inner (JZ) and outer myometrium and express different receptor and signaling repertoires. 33 37 110 Protocols for deriving and culturing primary SMCs developed for many other tissues (vascular, intestinal, etc.) facilitated development of robust protocols for isolation and culture of myometrial SMCs, which are commonly obtained from hysterectomy or caesarean section specimens. Cell origin (geographic location, pregnant or nonpregnant uterus) is an important consideration in defining primary cell source for myometrium. Permanent myometrial cell lines are also widely used for circumscribed studies such as screening drug actions. 111 112 113
Finally, all studies with patient-derived cells must carefully consider the donor profiles in design and interpretation of experiments. Age and parity may influence donor cell phenotype. Unlike endometriosis lesions, which can be collected from even very young patients, almost all myometrial tissue, along with adenomyosis lesion tissue, is derived from hysterectomy specimens. Although protocols for resecting and biopsying adenomyotic tissue have been described, 7 26 114 these procedures are not widely practiced. Thus, significant barriers exist in creating tissue banks from young patients who suffer from adenomyosis.
General Approaches to 3D Tissue Engineering of Endometrium and Myometrium Using Natural and Synthetic ECMs
An enduring challenge at present is to define suitable conditions to sustain long-term cocultures of multiple endometrial cell types that each has their own ECM and growth factor requirements. Matrigel comprises a mix of proteins and proteoglycans typical of the epithelial basement membrane, which filters proteins from the stromal compartment that requires a type I collagen and fibronectin-rich ECM with a very different composition of proteoglycans and associated matrix-bound growth factors. 115 The first 3D primary cell endometrial coculture model, tailored for studying blastocyst implantation, addressed this by embedding stromal fibroblasts in a collagen gel, mimicking features of the stromal matrix, which was coated by a thin layer of Matrigel onto which the epithelial cells were seeded, resulting in a confluent well-differentiated epithelia monolayer comprising ciliated and secretory (glandular) epithelia. 116 Versions of this model using cell lines, with stromal cells in collagen topped with Matrigel, were adapted to mimic the upregulation of MMPs and accompanying matrix degradation following progesterone withdrawal as a trigger of menstruation 28 117 as well as some features of the menstrual response to hormones, and other shorter-term aspects of stromal epithelial communication. 91 118 In a simpler version, in which stromal cells were embedded in Matrigel with epithelial cells on top, epithelial cells underwent 3D morphogenesis and proliferated, but stromal cells were relatively nonproliferative within the gel, 119 suggesting the need for cell-type–tailored ECM. An alternate approach, adapted from a successful clinical matrix for healing full-thickness (dermal–epidermal) wounds in skin, is to seed stromal cells in a porous collagen matrix fabricated by freezing and lyophilizing a type I collagen solution, and overlaying the matrix with epithelial cells, which produce their own basement membrane. 120
These models employing natural ECMs have shortcomings, in part because there are no “one-size-fits-all” ECMs for both epithelia and stroma (and potentially other cell types), and also because natural proteins are impure (Matrigel has myriad growth factors); are substantially variable from lot-to-lot; are relatively rapidly degraded by cells; and are subject to variation in structural and mechanical properties depending on how quickly or slowly the gels are polymerized. 115 121 Variation is further exacerbated as some investigators use atelocollagen, which yields very different outcomes than whole collagen. 122 These shortcomings have prompted a sustained effort in the biomaterials community to create synthetic alternatives to Matrigel and collagen. 123 124 A popular approach exploits the relative inertness of poly(ethylene glycol) (PEG), which is commercially available in multi-arm star/branched configurations activated with cell-compatible reactive groups, to create modular cell-encapsulating hydrogels crosslinked with short matrix metalloprotease (MMP)-sensitive peptides and incorporating synthetic integrin-binding motifs. 121 123 124 125 126 Other types of synthetic gel ECMs based entirely on synthetic proteins or semisynthetic gels based on modified hyaluronic acid are also widely used. 124 127 The local and bulk mechanical modulus, permeability, degradation properties, and biochemical recognition motifs—all of which have been correlated to cell responses 115 121 125 —can be tuned independently to match the needs of individual cells and tissues.
Recently, a locally responsive, “one-size-fits-all” PEG-based modular synthetic ECM was developed specifically for coculture of human endometrial epithelial and stromal cells, incorporating synthetic integrin-binding peptides for both cell types, and synthetic peptides that capture and sequester the different ECM proteins produced by epithelial and stromal cells. 92 95 121 A version of this synthetic ECM also supports expansion of human endometrial organoids 121 and formation of networks by human microvascular endothelial cells, 125 thus providing a foundation for engineering complex 3D cocultures ( Fig. 3b ). This matrix can be dissolved by a microbial enzyme to release cells and local cell–cell signaling molecules for analysis. 92 121 A relatively unsolved problem, however, is formulation of culture media compositions that support multiple different cell types. Solution of this problem may require finer specification and validation of “physiological” cell phenotypes, particularly for stromal cells which are often grown in serum-containing media optimized for maximal cell proliferation rates, rather than specific tissue functions.
Tissue engineering of the myometrium has benefitted extensively from the intense activity in using 3D models to illuminate arterial smooth muscle pathophysiology and regenerate blood vessels, where the roles of basement membrane, 3D architecture, cell density, and biomechanical stimulation on cell and tissue phenotype have been analyzed. 128 Understanding and modulating contractions in the pregnant uterus, to intervene therapeutically in preterm birth, has been the major application focus of in vitro 3D myometrial explant and tissue-engineered models. 36 111 112 113 129 130 131 132 Tissue-engineered models employing myometrial SMCs embedded in 3D collagen gels, adapted from successful models for vascular applications, offer advantages over explants by enabling a reproducible cell source from a tissue bank to be employed in multiple experiments, 131 and providing control over which cell types are present, to parse contributions of various immune cell populations, for example. An innovation involving an ECM-free magnetic cell aggregation 112 has been adapted to the myometrial contraction assays, but approaches involving synthetic ECMs are still only nascent for other SMC applications. 133
To summarize this section, the essential cell-based foundation for creating complex, 3D models of myometrial–endometrial interactions has been established through a combination of advances in basic methods of cell culture, combined with technologies for controlling the ECM microenvironment with synthetic biomaterials ( Fig. 3c ). These tools can be further combined with microfluidic devices to model pathologic phenotypes in a physiomimetic sense.
Modeling the Dynamic Processes of the Adenomyotic Microenvironment
The fundamental tools for building 3D tissue models are the constituent cells, scaffolds, matrices, platforms, and soluble cues that facilitate reconstruction or morphogenesis of physiologically relevant mimics of tissues. These fundamental tools are then deployed in specialized ways to model specific dynamic processes and phenotypes. Here, we describe the processes illustrated in Fig. 2 , and draw from applications in other disease research areas to illustrate how fundamental tissue engineering tools are often combined with other devices to further control elements of the culture microenvironment, such as mechanical stimulation, innervation, or fluid flow.
Invasion and Survival
Whether adenomyosis initially arises from a true invasion of the myometrium from the endometrium or through transport of endometrial cells into the myometrium by other means is debated, but once present, the displaced endometrial cells form lesions that grow and invade the surrounding tissue as the lesion enlarges. Cancer, wound healing, immune response to infection, and myriad other pathophysiological behaviors—including endometriosis and adenomyosis—have motivated a vast array of mechanistic experimental studies and biophysical models of cell migration and invasion. 15 122 134 135 136 Although various authors often interchange the terms migration and invasion, here we will refer to migration as a phenomenon of movement along a two-dimensional (2D) surface (e.g., endothelial cells moving across a denuded region of a vessel) or a quasi-2D surface (highly porous 3D matrix such as a large-pore transwell membrane), where no ECM degradation is required for movement. In contrast, we define invasion as a 3D process where breakdown of matrix or cell–cell junctions is required.
Migrating and invading cells inherently integrate an array of individual molecular processes to generate biophysical forces resulting in cell movement, which may be random or influenced by chemotactic (chemical-based), durotactic (matrix stiffness-based), or haptotactic (adhesion-based) gradients. 134 136 This integration can result in nonintuitive outcomes, necessitating careful attention to quantitative experimental parameters and metrics. For example, experimentally, cell migration speeds exhibit maxima for intermediate values of ECM-coating densities, across many different cell types and ECM coatings, as predicted by a biophysical model describing the balance between cell-matrix adhesion forces and cell-generated contractile forces. 134 Growth factors, which modulate both cell-matrix adhesion and cell contractile force generation, can appear to increase or decrease cell speed for a given ECM-coating density, depending on which side of the biphasic curve is operative for the chosen condition. 137 3D invasion assays inherently integrate more complex processes. The diversity in types of 3D movement, together with cell-mediated matrix degradation, results in different biophysical phenomena governance compared with that of 2D migration assays. 122 138
Although there are numerous studies focusing on migration and invasion of cells derived from eutopic or ectopic tissue in patients with endometriosis, 15 139 studies on adenomyosis are more limited and focused on stromal cells. 136 140 141 Consideration of how the individual molecular components in the extracellular environment (e.g., adhesion molecules, growth factors, porosity and stiffness of the matrix, matrix remodeling) and intracellular environment (e.g., integrin–cytoskeletal links, actin–myosin contractile forces, signaling pathways governing protein–protein associations) are integrated biophysically provides insight into why literature reports comparing adenomyotic and healthy cell migration and invasion are conflicting. 136 When eutopic endometrial stromal cells from adenomyosis patients and controls were compared in an assay format involving migration/invasion of cells across a thin porous membrane coated with Matrigel for 24 hours, no differences between the two groups were observed. 140 However, when eutopic endometrial stromal cells from patients with adenomyosis were compared with controls in an assay that involved invasion for 10 days into a 1 cm-thick slab of type I collagen gel, adenomyotic cells invaded further than controls; invasion of both control and adenomyotic cells was further enhanced when myometrial cells from control donors were included in the gel, and even further enhanced again if the donor myometrial cells were from an adenomyosis patient. 141 Furthermore, this same assay format revealed that estradiol and tamoxifen drive additional increases in the invasion depth of cells in each condition, while progesterone blunts the effects. 142 The 24-hour Matrigel invasion assay may not have captured differences between control and diseased populations, due to a combination of the relatively short assay time, use of an epithelial basement membrane-type matrix (Matrigel) rather than a stromal-like matrix (collagen I), and the potential for the many growth factors in Matrigel to stimulate chemokinesis (random motility) in a way that drove maximal invasion and obscured the differences. Still, whether these endometrial cells would invade a true smooth muscle structure, and if so, whether the factors regulating such invasion are the same as those regulating migration in ECM, is an interesting question for future models to address.
The enduring challenge, as noted at the outset of this article, is whether new targets for therapeutics are revealed, and if so, whether drugs that treat them (as well as existing drugs) can be circumscribed to certain groups of patients who might be identified by accessible clinical tests. Answering these related questions, especially in a manner that captures the multiple phenotypes leading to symptomatic pain and bleeding in adenomyosis, is still at an early stage of physiomimetic analysis for any type of therapeutic. With respect to identifying new targets, a study employing a high-throughput 3D invasion assay for the 12Z endometriotic cell line in collagen gels that examined invasion as a function of growth factor/cytokine stimulation, intracellular kinase signaling pathway response, and proteolytic shedding of cell surface receptors and growth factors 15 might be considered relevant for adenomyosis, as the 12Z line likely shares features with adenomyotic epithelia. As noted earlier, experiments with cell lines offer insights that can be followed up with primary cells. In the study, a Bayesian analysis linking the recursive protease-growth factor-kinase signaling loops revealed potential vulnerabilities in the network involving, among others, JUN kinase (JNK), and this prediction was experimentally verified with inhibitors. 15 Interestingly, a JNK inhibitor was also identified as a driver of inflammation in a separate study of peritoneal fluid in patients with surgically verified endometriosis, where a subset of patients exhibited a constellation of signature cytokines. 14 Two preclinical studies indicated effectiveness of a JNK inhibitor against endometriosis, 143 144 yet a clinical study in a nonstratified patient population (NCT01630252, NCT01631981) failed to meet endpoints. (None of the patients in either study were evaluated for possible adenomyosis, which is often comorbid with endometriosis and may contribute to symptoms and which may also respond to therapies for endometriosis.) Further exploration of this hypothesis with the types of models now being developed that include multicellular compartments and robust metrics may provide insights into how to design a clinical trial to treat subsets of patients with certain characteristics.
Adenomyosis invasion phenocopies features of carcinoma invasion into smooth muscle. Although the overwhelming majority of in vitro invasion studies have investigated invasion into ECM-rich tissues, the conceptual framework for building 3D invasion models in muscle has been mapped out over the past decade by observations in clinical specimens and intravital microscopy of carcinoma invasion in muscle in mouse models. 135 Epithelial invasion into muscle is associated with changes in multiple facets of cell cytoskeletal and mechanosensing behaviors, driving a preference for stiff environments. 135 Cells can invade into muscle as individual cells, or as chains, clusters, or collective strands, following the collagenous matrix between bundles or migrating along individual SMCs. 135 Features of collective cell invasion have been observed in deep infiltrating endometriosis lesions in the rectocervical space, 55 suggesting the possible invasion from adenomyotic lesions emanating from the cervical region of the uterus. The role of the regular smooth muscle contractions in the invasion process is still speculative, as there are consequences for mechanical damage, as well as promoting mitosis. 135 Hence, the development of models that experimentally recapitulate these multiple features is the goal of adenomyosis physiomimetic modeling.
Vascularization and Immune Interactions
With a vision toward building an adenomyosis lesion model that incorporates large-scale features of the lesion microenvironment, including a vascular bed, creation of a microvascular network becomes essential for survival and homeostasis of the engineered tissue. First, the microvascular system in the uterus, as in other organs, is essential for providing oxygen and nutrients. The dimensions of 3D in vitro models are inherently limited by diffusion of oxygen and nutrients to approximately 0.2 mm for metabolically active tissue like liver and muscle, and slightly larger for relatively acellular connective tissues such as dermis. 115 Second, the microvascular barrier also regulates trafficking of immune cells (and possibly circulating stem cells) and plays essential roles in tissue homeostasis through paracrine signaling to tissues, serving as the nidus for commencing decidualization of the endometrium. 145 Vascular function is modulated by systemic and local levels of sex hormones and inflammatory cues. 146 147 In vitro, cultures of endometrial stromal and epithelial cells in the presence of estrogen regulate angiogenesis, 148 and signals from flow-activated endothelial cells enhance hormone response in endometrial stromal cells. 145
Encouragingly, a fusion of tissue engineering and microfluidic cell culture technologies has spurred development of devices and protocols that could be applied to modeling adenomyosis lesions. Microfluidic devices with planar cultures that probe endothelial–stromal–immune interactions have already been applied to study the role of environmental chemicals on facets of endometriosis phenotypes. 149 Microfluidic and mesofluidic devices incorporating 3D tissue structures that can be perfused continuously with culture media, imaged, and sampled in situ are endemic tools in the emerging field of “microphysiological systems (MPSs)”—in vitro representations of complex physiological phenomena involving multiple cell types and dynamic behaviors in 3D. A crucial first step was to define protocols for the creation of stable, perfusable microvascular networks in a collagen or fibrin gel situated between two channels perfused with cell culture media ( Fig. 4a ). 150 151 152 Stable microvessels from endothelial cells and fibroblasts seeded in the central gel region ( Fig. 4b ), enabling phenomena involving trafficking of immune ( Fig. 4c ) and tumor cells across the endothelial barrier to be observed in detail ( Fig. 4d ). 153 These foundational approaches have now been widely adapted to support applications as diverse as in vitro tumor and islet microvascularization and formation of functional blood–brain barriers. 154 155 156 157 These studies establish principles for how a microvascularized endometrial–stromal lesion could be modeled; indeed, the authors are currently leveraging them to build multicellular microvascularized models of endometriosis lesions, employing primary patient-derived endometrial epithelial, stromal, and immune cells for analysis of how microvascular permeability and immune cell recruitment are regulated as a function of circulating sex steroids. A model of vascularized muscle developed for analyzing fibrosis and muscle damage 158 could potentially be adapted for creating a model of vascularized adenomyosis in the host tissue. One significant technical challenge relevant for modeling reproductive tissues is that the material most commonly used for fabricating microfluidic devices, polydimethylsiloxane (PDMS), absorbs lipophilic compounds like estrogen and progesterone, making it almost impossible to control their concentrations and thus tissue exposure. 159 Among fabrication materials, PDMS is uniquely permeable to oxygen—a limiting nutrient in most cultures; hence, significant design changes to provide oxygenation are required in transitioning to more suitable materials, but these shifts are gradually occurring as commercial chips based on thermoplastics are coming into use. 160
Fig. 4 Microfluidic model of immune and tumor cell trafficking between the microvasculature and tissues.

. ( a ) A microfluidic device comprising a central tissue-containing channel, flanked by two channels for flow of culture medium, is inoculated with a mixture of fibrin containing endothelial and stromal cells. Over the initial few days, cells undergo morphogenesis to form perfusable microvascular networks which are stable for weeks in culture as shown in ( b ) for a day 23 culture (actin stain). ( c ) Immune cells (green) can be perfused through the microvasculature (red) to model peripheral cell recruitment. ( d ) The dynamic cell-level phenomena in the devices can be imagined by confocal or two-photon microscopy to observe phenomena such as neutrophil–tumor cell interactions in the extravasation of tumor cells through the vascular wall into tissue. (Images from Zhang et al, 110 permission is in progress.)
Biomechanical Stimulation
It is now well recognized that the mechanical compliance of the ECM microenvironment—“stiffness” or “softness”—dramatically influences the phenotypes of epithelial cells, stem cells, and other cell types. 161 Matrices that are relatively stiff compared with the normal mechanical properties that a cell experiences can drive transition of fibroblasts to myofibroblasts and epithelial cells to a tumor-like phenotype. 161 Similarly, tumor cells accustomed to a stiff environment may exhibit poor survival and growth when placed into a more normal mechanical environment—a factor that may contribute to the difficulty in growing cells from endometriosis and adenomyosis lesions. These adaptations occur through a compendium of mechanical signaling pathways that are integrated in part by the intracellular translocation of transcriptional regulators YAP and TAZ to the nucleus, allowing differences in mechanical signaling between two different tissue environments in vivo to be inferred from immunohistochemistry. 162 An elegant in vivo – in vitro mechanical signaling study involving the myometrium first demonstrated links between the mechanical properties of uterine leiomyomas (fibroids) relative to surrounding myometrium and the corresponding relative amounts of nuclear YAP/TAZ (via immunohistochemistry), and then showed that isolated myometrial SMCs or fibroid cells cultured in 2D on a set of synthetic hydrogel substrates with systematically varied mechanical stiffness recapitulated the trends in nuclear YAP/TAZ seen for each cell type in vivo, along with enhanced ECM deposition by fibroid cells as seen in vivo. 162 This in vivo – in vitro correspondence suggests that the in vitro model may be a useful proxy for analyzing potential therapeutic interventions into mechanical signaling pathways. Other behaviors, such as invasion/survival, may require a 3D environment to capture these in vivo behaviors. Toward this goal, we have observed that normal human endometrial epithelial cells, which exhibit a prototypical spherical organoid morphology when cultured in soft synthetic hydrogels, 121 adopt a lesion-like morphology with invasive protrusions and epithelial-mesenchymal transition (EMT)-like cellular morphology when cultured in stiff synthetic hydrogels (Gnecco et al, unpublished data Aug, 15, 2020; Fig. 3c ). These type of reductionist models may help parse the relative contributions of inflammatory cues, hormones, and other microenvironment factors reported in adenomyosis lesions. However, the myometrium is not just a static microenvironment. The nonpregnant myometrium undergoes constant contractions to move fluid, in ways that are disrupted in adenomyosis. 35 36 The propensity of endometrial cells to form endometriosis lesions in smooth muscle of the intestinal tract and adenomyosis lesions in the myometrium, and to similarly be associated with hyperplastic smooth muscle (or myofibroblasts) in other ectopic contexts such as the bladder, ureter, rectocervical space, and relatively absent from the mesentery and omentum, suggests an interesting tropism for microenvironments that may offer mechanical stimulation.
Although the ability of drugs and hormones to alter contractile phenotype of myometrial cells has been used as a screen in explant and 3D culture assays, 81 113 129 and to compare general contractile abilities of normal and adenomyotic myometrium, 36 extension to more complex questions of etiology and interaction with endometrium under dynamic conditions is only nascent. Dynamic mechanical stimulation of human uterine cells in 2D has shown that endometrial stromal cells can acquire a contractile phenotype 163 and that human myometrial cells exhibit dramatic shifts in the phosphoproteome under stretch. 164 A study designed to illuminate the possible effects of myometrial contractions on endometrial phenotype used a novel microfluidic reactor to coculture an endometrial cell line on a layer of myometrial cells, in a manner that exposed the coculture to dynamic peristalsis that generated shear flow in the fluid impinging on the endometrial epithelial cell layer. 165 This proof-of-principle study performed with cell lines showed morphological changes in the epithelial barrier in response to the mechanical forces, 165 and provides a foundation for next-generation experiments with primary cells and a more complex tissue architecture, building on the well-developed observations about tissue engineering of vascular smooth muscle under pulsatile mechanical stimulation. 166
Innervation
In addition to the well-known animal dorsal root ganglia sources of neurons, the efficient derivation of genetically diverse peripheral sensory neurons from human cells has provided novel avenues for investigating and culturing human nerve fibers in vitro 167 168 and for building models for neuromotor action and pain. 167 While models of the endometrium have not yet been explored, microfluidic models of enteric nerve–epithelial interactions in the intestine are exemplary of the approaches. 169 Similarly, a microfluidic model of neuronal activation of vascularized skeletal muscle contraction, which allows quantitative analysis of axonal growth, muscle maturations, and contraction, provides a template for design of a similar device for the myometrium. 170 Ultimately, these tools might be combined to build models of innervated, vascularized adenomyosis lesions, using patient-specific samples to understand the manifestation of debilitating menorrhea.
Systemic and Organ–Organ interactions
Finally, adenomyosis and other chronic inflammatory diseases both exert and respond to systemic effects, potentially including those emanating from the gut microbiome. A growing number of microfluidic and mesofluidic models are being developed to connect MPSs representing multiple different organ systems (e.g., gut, liver, heart, brain) in a continuously communicating fluidic network for extended (weeks) culture periods, using fabrication materials that avoid the problems of PDMS. 171 172 173 A new pumping technology driven by integrating microfluidic pumps that are safe for immune cells onto the platform has been applied to examine the interplay between tissue-resident cells in the gut and liver and circulating immune cells in response to short-chain fatty acid metabolites produced in the colon, revealing paradoxical responses. 173 Such technologies might be used in the future to examine chronic cell trafficking between the vasculature and lesions in a single-MPS model of adenomyosis as a function of hormone cycles, or, more ambitiously, the interplay between the bone marrow–adenomyosis lesion axis with respect to circulating cells and factors, given the interplay between bone marrow–derived cells and wound healing/lesion phenomena. 174
Data-Driven Analysis and Predictive Modeling of Human Responses
A premise of the physiomimetics approach ( Fig. 1 ) is that patients can first be stratified on the basis of a combination of clinical phenotypes and molecular network analysis, to generate hypotheses for mechanistically distinguishable subgroups. In vitro tissue-engineered models of these subgroups can then be constructed using well-characterized patient-derived specimens and perturbed. Finally, multi-omic measurements combined with phenotypic metrics can be interpreted to drive identification and validation of therapies for patient subgroups, thus providing a foundation for stratified clinical trials. Molecular stratification of endometrium-derived diseases is still in early stages. A meta-analysis of genome-wide association studies for endometriosis showed stronger reproducibility across eight of nine disease-associated loci for patients with Stage III/IV disease compared with Stage I, 175 suggesting there may be clues to processes that amplify lesion characteristics. However, the loci implicate genes widely expressed throughout the body; thus, one could reasonably construct hypotheses around gut permeability influencing systemic immune function, for example, as a possible contributing mechanism. While systems-level physiomimetic modeling of complex systemic immune reactions involving gut permeability are emerging, 173 these are not the most likely place to begin to parse adenomyosis. Similarly, transcriptomic changes in endometrial biopsies are more pronounced in severe versus mild endometriosis, 176 suggesting several possible pathways for intervention. However, without other measurements of lesion tissue context and associated symptom characteristics, these datasets provide incomplete directions on designing physiomimetic models to test mechanisms.
These types of analyses might be paired, though, with phenotypic assays representing dynamic lesion properties. For example, the fine-grained in vitro analysis of pancreatic tumor organoid invasion characteristics, used for parsing mechanistic relationships between pancreatic cancer mutation subtypes and survival, 177 might provide phenotypic discrimination among patient subgroups, which could in turn lead to additional molecular phenotyping based on hypotheses regarding mechanisms governing the phenotypes. Even the assay used for the pancreatic tumors, however, revealed paradoxical relationships with survival 177 —suggesting an assay incorporating more complex tumor–stroma interactions could be more revealing. A piece of a physiomimetic puzzle for endometriosis, involving a bioinformatics prediction that JNK would govern cytokine release from peritoneal macrophages, which was then confirmed in vitro with patient samples, 14 may also have implications for adenomyosis, as macrophages are also involved. A physiomimetic model incorporating patient macrophages along with lesions may reveal disease-related phenotypic characteristics that could be modulated with a JNK inhibitor. At a systems level, the effects of potential new therapeutics can also be assessed using multi-MPS systems including liver and the immune system, incorporating features of liver metabolism and other metabolic and systemic transformations, using multi-omics to parse complex system responses. 172 173 These technologies are developing quickly and may greatly enhance physiomimetic modeling of gynecology diseases in general once resources to clinically phenotype patients and build corresponding tissue banks are identified.
Conclusions and Future Directions
The enormous spectrum of symptoms, histological/morphological appearances, and associated comorbidities that occur in patients with adenomyosis suggest that there may be distinct subsets of patients who could be targeted with personalized therapies—if the rules for defining these patient populations and approaches for developing the targeted therapies are established and implemented in an integrated fashion. The complexity of adenomyosis requires a physiomimetic approach: parsing the phenomena that may contribute using computational modeling approaches and then building a physiological model that provides information that translates back into the clinic. The fundamental in vitro tissue engineering approaches necessary to create physiomimetic models have largely been established: methods to conceptually and computationally characterize the disease to generate hypotheses about patient stratification approaches; methods to create tissue banks of relevant cell types derived from carefully phenotyped patients; 3D synthetic matrices to engineer microenvironments in a reproducible way; microfluidic platforms to control vascular and nerve interactions with 3D tissues; and analytical outputs using functional and molecular assays. The future of modeling adenomyosis lesion complexity arguably depends on how well these tools become democratized, whether large enough patient populations can be pooled to define the bedside-to-bench-to-bedside paradigm of physiomimetic modeling, and whether standardized methods can be defined and implemented for access to patient myometrial/adenomyotic tissue in younger patients who undergo fertility-sparing procedures. We are optimistic that the roadmap described here will spur multidisciplinary teams to hasten development and implementation of better treatments for patients.
Acknowledgments
We thank Hilary Critchley, Stacey Missmer, and Doug Lauffenburger for critical reading of the manuscript. This work was supported by the John and Karine Begg Foundation, the Manton Foundation, the National Science Foundation, and NIH U01 EB029132.
Funding Statement
Funding National Institutes of Health http://dx.doi.org/10.13039/100000002 EB029132 National Science Foundation http://dx.doi.org/10.13039/100000001
Footnotes
Conflict of Interest None declared.
References
- 1.Lopes-Pacheco M. CFTR modulators: the changing face of cystic fibrosis in the era of precision medicine. Front Pharmacol. 2020;10:1662. doi: 10.3389/fphar.2019.01662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Benagiano G, Habiba M, Brosens I. The pathophysiology of uterine adenomyosis: an update. Fertil Steril. 2012;98(03):572–579. doi: 10.1016/j.fertnstert.2012.06.044. [DOI] [PubMed] [Google Scholar]
- 3.Chapron C, Tosti C, Marcellin L. Relationship between the magnetic resonance imaging appearance of adenomyosis and endometriosis phenotypes. Hum Reprod. 2017;32(07):1393–1401. doi: 10.1093/humrep/dex088. [DOI] [PubMed] [Google Scholar]
- 4.García-Solares J, Donnez J, Donnez O, Dolmans M M. Pathogenesis of uterine adenomyosis: invagination or metaplasia? Fertil Steril. 2018;109(03):371–379. doi: 10.1016/j.fertnstert.2017.12.030. [DOI] [PubMed] [Google Scholar]
- 5.Habiba M, Gordts S, Bazot M, Brosens I, Benagiano G. Exploring the challenges for a new classification of adenomyosis. Reprod Biomed Online. 2020;40(04):569–581. doi: 10.1016/j.rbmo.2020.01.017. [DOI] [PubMed] [Google Scholar]
- 6.FIGO Working Group on Menstrual Disorders . Munro M G, Critchley H O, Broder M S, Fraser I S. FIGO classification system (PALM-COEIN) for causes of abnormal uterine bleeding in nongravid women of reproductive age. Int J Gynaecol Obstet. 2011;113(01):3–13. doi: 10.1016/j.ijgo.2010.11.011. [DOI] [PubMed] [Google Scholar]
- 7.Tellum T, Qvigstad E, Skovholt E K, Lieng M. In vivo adenomyosis tissue sampling using a transvaginal ultrasound-guided core biopsy technique for research purposes: safety, feasibility, and effectiveness. J Minim Invasive Gynecol. 2019;26(07):1357–1362. doi: 10.1016/j.jmig.2019.02.002. [DOI] [PubMed] [Google Scholar]
- 8.Zondervan K T, Becker C M, Koga K, Missmer S A, Taylor R N, Viganò P. Endometriosis. Nat Rev Dis Primers. 2018;4(01):9. doi: 10.1038/s41572-018-0008-5. [DOI] [PubMed] [Google Scholar]
- 9.Koninckx P R, Ussia A, Adamyan L, Wattiez A, Gomel V, Martin D C. Heterogeneity of endometriosis lesions requires individualisation of diagnosis and treatment and a different approach to research and evidence based medicine. Facts Views Vis ObGyn. 2019;11(01):57–61. [PMC free article] [PubMed] [Google Scholar]
- 10.Osuga Y, Fujimoto-Okabe H, Hagino A. Evaluation of the efficacy and safety of dienogest in the treatment of painful symptoms in patients with adenomyosis: a randomized, double-blind, multicenter, placebo-controlled study. Fertil Steril. 2017;108(04):673–678. doi: 10.1016/j.fertnstert.2017.07.021. [DOI] [PubMed] [Google Scholar]
- 11.Vannuccini S, Tosti C, Carmona F. Pathogenesis of adenomyosis: an update on molecular mechanisms. Reprod Biomed Online. 2017;35(05):592–601. doi: 10.1016/j.rbmo.2017.06.016. [DOI] [PubMed] [Google Scholar]
- 12.Inoue S, Hirota Y, Ueno T. Uterine adenomyosis is an oligoclonal disorder associated with KRAS mutations. Nat Commun. 2019;10(01):5785. doi: 10.1038/s41467-019-13708-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Shaked S, Jaffa A J, Grisaru D, Elad D. Uterine peristalsis-induced stresses within the uterine wall may sprout adenomyosis. Biomech Model Mechanobiol. 2015;14(03):437–444. doi: 10.1007/s10237-014-0614-4. [DOI] [PubMed] [Google Scholar]
- 14.Beste M T, Pfäffle-Doyle N, Prentice E A. Molecular network analysis of endometriosis reveals a role for c-Jun-regulated macrophage activation. Sci Transl Med. 2014;6(222):222ra16. doi: 10.1126/scitranslmed.3007988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Miller M A, Meyer A S, Beste M T. ADAM-10 and -17 regulate endometriotic cell migration via concerted ligand and receptor shedding feedback on kinase signaling. Proc Natl Acad Sci U S A. 2013;110(22):E2074–E2083. doi: 10.1073/pnas.1222387110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Shafrir A L, Missmer S A. Towards subtypes - deep endometriosis oestrogen receptor-α expression. Nat Rev Endocrinol. 2020;16(10):541–542. doi: 10.1038/s41574-020-0394-0. [DOI] [PubMed] [Google Scholar]
- 17.Zondervan K T, Rahmioglu N, Morris A P. Beyond endometriosis genome-wide association study: from genomics to phenomics to the patient. Semin Reprod Med. 2016;34(04):242–254. doi: 10.1055/s-0036-1585408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Conklin J E, Lieberman J V, Barnes C A, Louis D Z. Disease staging: implications for hospital reimbursement and management. Health Care Financ Rev. 1984;1984:13–22. [PMC free article] [PubMed] [Google Scholar]
- 19.Gonnella J S, Hornbrook M C, Louis D Z. Staging of disease. A case-mix measurement. JAMA. 1984;251(05):637–644. [PubMed] [Google Scholar]
- 20.Gonella J, Louis D Z, Guozum M VE, Callahan C A, Barnes C A.Disease stagingMI: Ann Arbor;2001927 [Google Scholar]
- 21.FIGO Menstrual Disorders Committee . Munro M G, Critchley H OD, Fraser I S. The two FIGO systems for normal and abnormal uterine bleeding symptoms and classification of causes of abnormal uterine bleeding in the reproductive years: 2018 revisions. Int J Gynaecol Obstet. 2018;143(03):393–408. doi: 10.1002/ijgo.12666. [DOI] [PubMed] [Google Scholar]
- 22.Driehuis E, van Hoeck A, Moore K. Pancreatic cancer organoids recapitulate disease and allow personalized drug screening. Proc Natl Acad Sci U S A. 2019;116(52):26580–26590. doi: 10.1073/pnas.1911273116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.WERF EPHect Working Group . Vitonis A F, Vincent K, Rahmioglu N. World Endometriosis Research Foundation Endometriosis Phenome and Biobanking Harmonization Project: II. Clinical and covariate phenotype data collection in endometriosis research. Fertil Steril. 2014;102(05):1223–1232. doi: 10.1016/j.fertnstert.2014.07.1244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.WERF EPHect Working Group . Rahmioglu N, Fassbender A, Vitonis A F. World Endometriosis Research Foundation Endometriosis Phenome and Biobanking Harmonization Project: III. Fluid biospecimen collection, processing, and storage in endometriosis research. Fertil Steril. 2014;102(05):1233–1243. doi: 10.1016/j.fertnstert.2014.07.1208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Pugliese A, Yang M, Kusmarteva I. The Juvenile Diabetes Research Foundation Network for Pancreatic Organ Donors with Diabetes (nPOD) program: goals, operational model and emerging findings. Pediatr Diabetes. 2014;15(01):1–9. doi: 10.1111/pedi.12097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gordts S, Campo R, Brosens I. Hysteroscopic diagnosis and excision of myometrial cystic adenomyosis. Gynecol Surg. 2014;11(04):273–278. doi: 10.1007/s10397-014-0861-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Critchley H O, Jones R L, Lea R G. Role of inflammatory mediators in human endometrium during progesterone withdrawal and early pregnancy. J Clin Endocrinol Metab. 1999;84(01):240–248. doi: 10.1210/jcem.84.1.5380. [DOI] [PubMed] [Google Scholar]
- 28.Critchley H OD, Maybin J A, Armstrong G M, Williams A RW. Physiology of the endometrium and regulation of menstruation. Physiol Rev. 2020;100(03):1149–1179. doi: 10.1152/physrev.00031.2019. [DOI] [PubMed] [Google Scholar]
- 29.Vigano P, Candiani M, Monno A, Giacomini E, Vercellini P, Somigliana E. Time to redefine endometriosis including its pro-fibrotic nature. Hum Reprod. 2018;33(03):347–352. doi: 10.1093/humrep/dex354. [DOI] [PubMed] [Google Scholar]
- 30.Thiruchelvam U, Dransfield I, Saunders P T, Critchley H O. The importance of the macrophage within the human endometrium. J Leukoc Biol. 2013;93(02):217–225. doi: 10.1189/jlb.0712327. [DOI] [PubMed] [Google Scholar]
- 31.Osteen K G, Rodgers W H, Gaire M, Hargrove J T, Gorstein F, Matrisian L M. Stromal-epithelial interaction mediates steroidal regulation of metalloproteinase expression in human endometrium. Proc Natl Acad Sci U S A. 1994;91(21):10129–10133. doi: 10.1073/pnas.91.21.10129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Henderson T A, Saunders P T, Moffett-King A, Groome N P, Critchley H O. Steroid receptor expression in uterine natural killer cells. J Clin Endocrinol Metab. 2003;88(01):440–449. doi: 10.1210/jc.2002-021174. [DOI] [PubMed] [Google Scholar]
- 33.Myers K M, Elad D.Biomechanics of the human uterusWiley Interdiscip Rev Syst Biol Med 2017;9(05): [DOI] [PubMed]
- 34.Aguilar H N, Mitchell B F. Physiological pathways and molecular mechanisms regulating uterine contractility. Hum Reprod Update. 2010;16(06):725–744. doi: 10.1093/humupd/dmq016. [DOI] [PubMed] [Google Scholar]
- 35.Mehasseb M K, Bell S C, Pringle J H, Habiba M A. Uterine adenomyosis is associated with ultrastructural features of altered contractility in the inner myometrium. Fertil Steril. 2010;93(07):2130–2136. doi: 10.1016/j.fertnstert.2009.01.097. [DOI] [PubMed] [Google Scholar]
- 36.Guo S W, Mao X, Ma Q, Liu X. Dysmenorrhea and its severity are associated with increased uterine contractility and overexpression of oxytocin receptor (OTR) in women with symptomatic adenomyosis. Fertil Steril. 2013;99(01):231–240. doi: 10.1016/j.fertnstert.2012.08.038. [DOI] [PubMed] [Google Scholar]
- 37.Brosens J J, Barker F G, de Souza N M. Myometrial zonal differentiation and uterine junctional zone hyperplasia in the non-pregnant uterus. Hum Reprod Update. 1998;4(05):496–502. doi: 10.1093/humupd/4.5.496. [DOI] [PubMed] [Google Scholar]
- 38.Benagiano G, Brosens I, Habiba M. Structural and molecular features of the endomyometrium in endometriosis and adenomyosis. Hum Reprod Update. 2014;20(03):386–402. doi: 10.1093/humupd/dmt052. [DOI] [PubMed] [Google Scholar]
- 39.Strauss J F, Lessey B A.The structure, function, and evaluation of the female reproductive tractIn: Yen & Jaffe's Reproductive Endocrinology, Elsevier, NY.2009191–233. [Google Scholar]
- 40.Leyendecker G, Wildt L. A new concept of endometriosis and adenomyosis: tissue injury and repair (TIAR) Horm Mol Biol Clin Investig. 2011;5(02):125–142. doi: 10.1515/HMBCI.2011.002. [DOI] [PubMed] [Google Scholar]
- 41.Ibrahim M G, Chiantera V, Frangini S.Ultramicro-trauma in the endometrial-myometrial junctional zone and pale cell migration in adenomyosis Fertil Steril 2015104061475–830., 3 [DOI] [PubMed] [Google Scholar]
- 42.Zhang Q, Duan J, Liu X, Guo S W. Platelets drive smooth muscle metaplasia and fibrogenesis in endometriosis through epithelial-mesenchymal transition and fibroblast-to-myofibroblast transdifferentiation. Mol Cell Endocrinol. 2016;428:1–16. doi: 10.1016/j.mce.2016.03.015. [DOI] [PubMed] [Google Scholar]
- 43.Kobayashi H, Kishi Y, Matsubara S. Mechanisms underlying adenomyosis-related fibrogenesis. Gynecol Obstet Invest. 2020;85(01):1–12. doi: 10.1159/000502822. [DOI] [PubMed] [Google Scholar]
- 44.Li T, Li Y G, Pu D M. Matrix metalloproteinase-2 and -9 expression correlated with angiogenesis in human adenomyosis. Gynecol Obstet Invest. 2006;62(04):229–235. doi: 10.1159/000094426. [DOI] [PubMed] [Google Scholar]
- 45.Guo S W. Fibrogenesis resulting from cyclic bleeding: the Holy Grail of the natural history of ectopic endometrium. Hum Reprod. 2018;33(03):353–356. doi: 10.1093/humrep/dey015. [DOI] [PubMed] [Google Scholar]
- 46.Li H, Yu Y, Shi Y. HoxA13 stimulates myometrial cells to secrete IL-1β and enhance the expression of contraction-associated proteins. Endocrinology. 2016;157(05):2129–2139. doi: 10.1210/en.2015-2005. [DOI] [PubMed] [Google Scholar]
- 47.Mehasseb M K, Panchal R, Taylor A H, Brown L, Bell S C, Habiba M.Estrogen and progesterone receptor isoform distribution through the menstrual cycle in uteri with and without adenomyosis Fertil Steril 201195072228–2235., 2235.e1 [DOI] [PubMed] [Google Scholar]
- 48.Jichan Nie, Xishi Liu, Guo S W. Promoter hypermethylation of progesterone receptor isoform B (PR-B) in adenomyosis and its rectification by a histone deacetylase inhibitor and a demethylation agent. Reprod Sci. 2010;17(11):995–1005. doi: 10.1177/1933719110377118. [DOI] [PubMed] [Google Scholar]
- 49.Scheerer C, Frangini S, Chiantera V, Mechsner S. Reduced sympathetic innervation in endometriosis is associated to Semaphorin 3C and 3F expression. Mol Neurobiol. 2017;54(07):5131–5141. doi: 10.1007/s12035-016-0058-1. [DOI] [PubMed] [Google Scholar]
- 50.Wang G, Tokushige N, Fraser I S. Nerve fibers and menstrual cycle in peritoneal endometriosis. Fertil Steril. 2011;95(08):2772–2774. doi: 10.1016/j.fertnstert.2011.01.150. [DOI] [PubMed] [Google Scholar]
- 51.Al-Jefout M, Dezarnaulds G, Cooper M. Diagnosis of endometriosis by detection of nerve fibres in an endometrial biopsy: a double blind study. Hum Reprod. 2009;24(12):3019–3024. doi: 10.1093/humrep/dep275. [DOI] [PubMed] [Google Scholar]
- 52.Browne A S, Yu J, Huang R P, Francisco A M, Sidell N, Taylor R N. Proteomic identification of neurotrophins in the eutopic endometrium of women with endometriosis. Fertil Steril. 2012;98(03):713–719. doi: 10.1016/j.fertnstert.2012.05.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Anaf V, Simon P, El Nakadi I. Relationship between endometriotic foci and nerves in rectovaginal endometriotic nodules. Hum Reprod. 2000;15(08):1744–1750. doi: 10.1093/humrep/15.8.1744. [DOI] [PubMed] [Google Scholar]
- 54.Mechsner S, Kaiser A, Kopf A, Gericke C, Ebert A, Bartley J. A pilot study to evaluate the clinical relevance of endometriosis-associated nerve fibers in peritoneal endometriotic lesions. Fertil Steril. 2009;92(06):1856–1861. doi: 10.1016/j.fertnstert.2008.09.006. [DOI] [PubMed] [Google Scholar]
- 55.García-Solares J, Dolmans M M, Squifflet J L, Donnez J, Donnez O. Invasion of human deep nodular endometriotic lesions is associated with collective cell migration and nerve development. Fertil Steril. 2018;110(07):1318–1327. doi: 10.1016/j.fertnstert.2018.08.016. [DOI] [PubMed] [Google Scholar]
- 56.Tran L V, Tokushige N, Berbic M, Markham R, Fraser I S. Macrophages and nerve fibres in peritoneal endometriosis. Hum Reprod. 2009;24(04):835–841. doi: 10.1093/humrep/den483. [DOI] [PubMed] [Google Scholar]
- 57.Greaves E, Temp J, Esnal-Zufiurre A, Mechsner S, Horne A W, Saunders P T. Estradiol is a critical mediator of macrophage-nerve cross talk in peritoneal endometriosis. Am J Pathol. 2015;185(08):2286–2297. doi: 10.1016/j.ajpath.2015.04.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Zhang X, Lu B, Huang X, Xu H, Zhou C, Lin J. Innervation of endometrium and myometrium in women with painful adenomyosis and uterine fibroids. Fertil Steril. 2010;94(02):730–737. doi: 10.1016/j.fertnstert.2009.03.026. [DOI] [PubMed] [Google Scholar]
- 59.Vacca P, Vitale C, Montaldo E. CD34+ hematopoietic precursors are present in human decidua and differentiate into natural killer cells upon interaction with stromal cells. Proc Natl Acad Sci U S A. 2011;108(06):2402–2407. doi: 10.1073/pnas.1016257108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Armstrong G M, Maybin J A, Murray A A. Endometrial apoptosis and neutrophil infiltration during menstruation exhibits spatial and temporal dynamics that are recapitulated in a mouse model. Sci Rep. 2017;7(01):17416. doi: 10.1038/s41598-017-17565-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Russell P, Anderson L, Lieberman D.The distribution of immune cells and macrophages in the endometrium of women with recurrent reproductive failure I: Techniques J Reprod Immunol 201191(1-2):90–102. [DOI] [PubMed] [Google Scholar]
- 62.Bulmer J N, Jones R K, Searle R F.Intraepithelial leukocytes in endometriosis and adenomyosis: comparison of eutopic and ectopic endometrium with normal endometrium Hum Reprod 199813(1O):2910–2915. [DOI] [PubMed] [Google Scholar]
- 63.Orazov M R, Radzinsky V E, Nosenko E N, Khamoshina M B, Dukhin A O, Lebedeva M G. Immune-inflammatory predictors of the pelvic pain syndrome associated with adenomyosis. Gynecol Endocrinol. 2017;33 01:44–46. doi: 10.1080/09513590.2017.1399696. [DOI] [PubMed] [Google Scholar]
- 64.He H, Mack J J, Güç E. Perivascular macrophages limit permeability. Arterioscler Thromb Vasc Biol. 2016;36(11):2203–2212. doi: 10.1161/ATVBAHA.116.307592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Schindl M, Birner P, Obermair A, Kiesel L, Wenzl R. Increased microvessel density in adenomyosis uteri. Fertil Steril. 2001;75(01):131–135. doi: 10.1016/s0015-0282(00)01642-3. [DOI] [PubMed] [Google Scholar]
- 66.Liu X, Shen M, Qi Q, Zhang H, Guo S W. Corroborating evidence for platelet-induced epithelial-mesenchymal transition and fibroblast-to-myofibroblast transdifferentiation in the development of adenomyosis. Hum Reprod. 2016;31(04):734–749. doi: 10.1093/humrep/dew018. [DOI] [PubMed] [Google Scholar]
- 67.Harmsen M J, Wong C FC, Mijatovic V. Role of angiogenesis in adenomyosis-associated abnormal uterine bleeding and subfertility: a systematic review. Hum Reprod Update. 2019;25(05):647–671. doi: 10.1093/humupd/dmz024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Ota H, Igarashi S, Hatazawa J, Tanaka T. Is adenomyosis an immune disease? Hum Reprod Update. 1998;4(04):360–367. doi: 10.1093/humupd/4.4.360. [DOI] [PubMed] [Google Scholar]
- 69.De Leo B, Esnal-Zufiaurre A, Collins F, Critchley H OD, Saunders P TK. Immunoprofiling of human uterine mast cells identifies three phenotypes and expression of ERβ and glucocorticoid receptor. F1000 Res. 2017;6:667. doi: 10.12688/f1000research.11432.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Benagiano G, Brosens I. The endometrium in adenomyosis. Womens Health (Lond) 2012;8(03):301–312. doi: 10.2217/whe.12.8. [DOI] [PubMed] [Google Scholar]
- 71.Tremellen K P, Russell P. The distribution of immune cells and macrophages in the endometrium of women with recurrent reproductive failure. II: adenomyosis and macrophages. J Reprod Immunol. 2012;93(01):58–63. doi: 10.1016/j.jri.2011.12.001. [DOI] [PubMed] [Google Scholar]
- 72.Ota H, Tanaka T. Stromal vascularization in the endometrium during adenomyosis. Microsc Res Tech. 2003;60(04):445–449. doi: 10.1002/jemt.10282. [DOI] [PubMed] [Google Scholar]
- 73.Ota H, Igarashi S, Hatazawa J, Tanaka T. Immunohistochemical assessment of superoxide dismutase expression in the endometrium in endometriosis and adenomyosis. Fertil Steril. 1999;72(01):129–134. doi: 10.1016/s0015-0282(99)00152-1. [DOI] [PubMed] [Google Scholar]
- 74.Herndon C N, Aghajanova L, Balayan S. Global transcriptome abnormalities of the eutopic endometrium from women with adenomyosis. Reprod Sci. 2016;23(10):1289–1303. doi: 10.1177/1933719116650758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Acar S, Millar E, Mitkova M, Mitkov V. Value of ultrasound shear wave elastography in the diagnosis of adenomyosis. Ultrasound. 2016;24(04):205–213. doi: 10.1177/1742271X16673677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Al-Sabbagh M, Lam E W, Brosens J J. Mechanisms of endometrial progesterone resistance. Mol Cell Endocrinol. 2012;358(02):208–215. doi: 10.1016/j.mce.2011.10.035. [DOI] [PubMed] [Google Scholar]
- 77.Hellweg G, Shaka J A. Endometrial granulocytes; tissue culture studies of endometrium and decidua with special attention to the endometrial granulocytes. Obstet Gynecol. 1959;13(05):519–529. [PubMed] [Google Scholar]
- 78.Kawaguchi K, Fujii S, Konishi I, Okamura H, Mori T. Ultrastructural study of cultured smooth muscle cells from uterine leiomyoma and myometrium under the influence of sex steroids. Gynecol Oncol. 1985;21(01):32–41. doi: 10.1016/0090-8258(85)90229-x. [DOI] [PubMed] [Google Scholar]
- 79.Gellersen B, Bonhoff A, Hunt N, Bohnet H G. Decidual-type prolactin expression by the human myometrium. Endocrinology. 1991;129(01):158–168. doi: 10.1210/endo-129-1-158. [DOI] [PubMed] [Google Scholar]
- 80.Heidari Kani M H, Chan E-C, Young R C, Butler T, Smith R, Paul J W. 3D cell culturing and possibilities for myometrial tissue engineering. Ann Biomed Eng. 2017;45(07):1746–1757. doi: 10.1007/s10439-016-1749-5. [DOI] [PubMed] [Google Scholar]
- 81.Arrowsmith S, Keov P, Muttenthaler M, Gruber C W. Contractility measurements of human uterine smooth muscle to aid drug development. J Vis Exp. 2018;(131):56639. doi: 10.3791/56639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Gaide Chevronnay H P, Lemoine P, Courtoy P J, Marbaix E, Henriet P. Ovarian steroids, mitogen-activated protein kinases, and/or aspartic proteinases cooperate to control endometrial remodeling by regulating gene expression in the stroma and glands. Endocrinology. 2010;151(09):4515–4526. doi: 10.1210/en.2009-1398. [DOI] [PubMed] [Google Scholar]
- 83.Schäfer W R, Fischer L, Roth K. Critical evaluation of human endometrial explants as an ex vivo model system: a molecular approach. Mol Hum Reprod. 2011;17(04):255–265. doi: 10.1093/molehr/gaq095. [DOI] [PubMed] [Google Scholar]
- 84.Fasciani A, Bocci G, Xu J. Three-dimensional in vitro culture of endometrial explants mimics the early stages of endometriosis. Fertil Steril. 2003;80(05):1137–1143. doi: 10.1016/s0015-0282(03)02164-2. [DOI] [PubMed] [Google Scholar]
- 85.Osteen K G, Hill G A, Hargrove J T, Gorstein F. Development of a method to isolate and culture highly purified populations of stromal and epithelial cells from human endometrial biopsy specimens. Fertil Steril. 1989;52(06):965–972. doi: 10.1016/s0015-0282(16)53160-4. [DOI] [PubMed] [Google Scholar]
- 86.Korch C, Spillman M A, Jackson T A. DNA profiling analysis of endometrial and ovarian cell lines reveals misidentification, redundancy and contamination. Gynecol Oncol. 2012;127(01):241–248. doi: 10.1016/j.ygyno.2012.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Banu S K, Lee J, Starzinski-Powitz A, Arosh J A. Gene expression profiles and functional characterization of human immortalized endometriotic epithelial and stromal cells. Fertil Steril. 2008;90(04):972–987. doi: 10.1016/j.fertnstert.2007.07.1358. [DOI] [PubMed] [Google Scholar]
- 88.Samalecos A, Reimann K, Wittmann S. Characterization of a novel telomerase-immortalized human endometrial stromal cell line, St-T1b. Reprod Biol Endocrinol. 2009;7:76. doi: 10.1186/1477-7827-7-76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Krikun G, Mor G, Alvero A. A novel immortalized human endometrial stromal cell line with normal progestational response. Endocrinology. 2004;145(05):2291–2296. doi: 10.1210/en.2003-1606. [DOI] [PubMed] [Google Scholar]
- 90.Wu J, Taylor R N, Sidell N. Retinoic acid regulates gap junction intercellular communication in human endometrial stromal cells through modulation of the phosphorylation status of connexin 43. J Cell Physiol. 2013;228(04):903–910. doi: 10.1002/jcp.24241. [DOI] [PubMed] [Google Scholar]
- 91.Schutte S C, James C O, Sidell N, Taylor R N. Tissue-engineered endometrial model for the study of cell-cell interactions. Reprod Sci. 2015;22(03):308–315. doi: 10.1177/1933719114542008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Valdez J, Cook C D, Ahrens C C. On-demand dissolution of modular, synthetic extracellular matrix reveals local epithelial-stromal communication networks. Biomaterials. 2017;130:90–103. doi: 10.1016/j.biomaterials.2017.03.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Arnold J T, Lessey B A, Seppälä M, Kaufman D G. Effect of normal endometrial stroma on growth and differentiation in Ishikawa endometrial adenocarcinoma cells. Cancer Res. 2002;62(01):79–88. [PubMed] [Google Scholar]
- 94.Zeitvogel A, Baumann R, Starzinski-Powitz A. Identification of an invasive, N-cadherin-expressing epithelial cell type in endometriosis using a new cell culture model. Am J Pathol. 2001;159(05):1839–1852. doi: 10.1016/S0002-9440(10)63030-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Cook C D, Hill A S, Guo M. Local remodeling of synthetic extracellular matrix microenvironments by co-cultured endometrial epithelial and stromal cells enables long-term dynamic physiological function. Integr Biol. 2017;9(04):271–289. doi: 10.1039/c6ib00245e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Barragan F, Irwin J C, Balayan S. Human endometrial fibroblasts derived from mesenchymal progenitors inherit progesterone resistance and acquire an inflammatory phenotype in the endometrial niche in endometriosis. Biol Reprod. 2016;94(05):118. doi: 10.1095/biolreprod.115.136010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.von Wolff M, Stieger S, Lumpp K, Bücking J, Strowitzki T, Thaler C J. Endometrial interleukin-6 in vitro is not regulated directly by female steroid hormones, but by pro-inflammatory cytokines and hypoxia. Mol Hum Reprod. 2002;8(12):1096–1102. doi: 10.1093/molehr/8.12.1096. [DOI] [PubMed] [Google Scholar]
- 98.Chen J C, Hoffman J R, Arora R. Cryopreservation and recovery of human endometrial epithelial cells with high viability, purity, and functional fidelity. Fertil Steril. 2016;105(02):501–100. doi: 10.1016/j.fertnstert.2015.10.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Turco M Y, Gardner L, Hughes J. Long-term, hormone-responsive organoid cultures of human endometrium in a chemically defined medium. Nat Cell Biol. 2017;19(05):568–577. doi: 10.1038/ncb3516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Boretto M, Cox B, Noben M. Development of organoids from mouse and human endometrium showing endometrial epithelium physiology and long-term expandability. Development. 2017;144(10):1775–1786. doi: 10.1242/dev.148478. [DOI] [PubMed] [Google Scholar]
- 101.Sato T, Vries R G, Snippert H J.Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche Nature 2009459(7244):262–265. [DOI] [PubMed] [Google Scholar]
- 102.Valentijn A J, Saretzki G, Tempest N, Critchley H O, Hapangama D K. Human endometrial epithelial telomerase is important for epithelial proliferation and glandular formation with potential implications in endometriosis. Hum Reprod. 2015;30(12):2816–2828. doi: 10.1093/humrep/dev267. [DOI] [PubMed] [Google Scholar]
- 103.Boretto M, Maenhoudt N, Luo X. Patient-derived organoids from endometrial disease capture clinical heterogeneity and are amenable to drug screening. Nat Cell Biol. 2019;21(08):1041–1051. doi: 10.1038/s41556-019-0360-z. [DOI] [PubMed] [Google Scholar]
- 104.Hapangama D K, Drury J, Da Silva L. Abnormally located SSEA1+/SOX9+ endometrial epithelial cells with a basalis-like phenotype in the eutopic functionalis layer may play a role in the pathogenesis of endometriosis. Hum Reprod. 2019;34(01):56–68. doi: 10.1093/humrep/dey336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Gargett C E, Schwab K E, Zillwood R M, Nguyen H P, Wu D. Isolation and culture of epithelial progenitors and mesenchymal stem cells from human endometrium. Biol Reprod. 2009;80(06):1136–1145. doi: 10.1095/biolreprod.108.075226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Tempest N, Maclean A, Hapangama D K. Endometrial stem cell markers: current concepts and unresolved questions. Int J Mol Sci. 2018;19(10):E3240. doi: 10.3390/ijms19103240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Tempest N, Baker A M, Wright N A, Hapangama D K. Does human endometrial LGR5 gene expression suggest the existence of another hormonally regulated epithelial stem cell niche? Hum Reprod. 2018;33(06):1052–1062. doi: 10.1093/humrep/dey083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Valentijn A J, Palial K, Al-Lamee H. SSEA-1 isolates human endometrial basal glandular epithelial cells: phenotypic and functional characterization and implications in the pathogenesis of endometriosis. Hum Reprod. 2013;28(10):2695–2708. doi: 10.1093/humrep/det285. [DOI] [PubMed] [Google Scholar]
- 109.Miyazaki K, Dyson M T, Coon V J S. Generation of progesterone-responsive endometrial stromal fibroblasts from human induced pluripotent stem cells: role of the WNT/CTNNB1 pathway. Stem Cell Reports. 2018;11(05):1136–1155. doi: 10.1016/j.stemcr.2018.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Zhang Y, Zhou L, Li T C, Duan H, Yu P, Wang H Y. Ultrastructural features of endometrial-myometrial interface and its alteration in adenomyosis. Int J Clin Exp Pathol. 2014;7(04):1469–1477. [PMC free article] [PubMed] [Google Scholar]
- 111.Hutchinson J L, Rajagopal S P, Yuan M, Norman J E. Lipopolysaccharide promotes contraction of uterine myocytes via activation of Rho/ROCK signaling pathways. FASEB J. 2014;28(01):94–105. doi: 10.1096/fj.13-237040. [DOI] [PubMed] [Google Scholar]
- 112.Souza G R, Tseng H, Gage J A. Magnetically bioprinted human myometrial 3D cell rings as a model for uterine contractility. Int J Mol Sci. 2017;18(04):E683. doi: 10.3390/ijms18040683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Vaes R DW, van den Berk L, Boonen B, van Dijk D PJ, Olde Damink S WM, Rensen S S. A novel human cell culture model to study visceral smooth muscle phenotypic modulation in health and disease. Am J Physiol Cell Physiol. 2018;315(04):C598–C607. doi: 10.1152/ajpcell.00167.2017. [DOI] [PubMed] [Google Scholar]
- 114.Osada H. Uterine adenomyosis and adenomyoma: the surgical approach. Fertil Steril. 2018;109(03):406–417. doi: 10.1016/j.fertnstert.2018.01.032. [DOI] [PubMed] [Google Scholar]
- 115.Griffith L G, Swartz M A. Capturing complex 3D tissue physiology in vitro. Nat Rev Mol Cell Biol. 2006;7(03):211–224. doi: 10.1038/nrm1858. [DOI] [PubMed] [Google Scholar]
- 116.Bentin-Ley U, Pedersen B, Lindenberg S, Larsen J F, Hamberger L, Horn T. Isolation and culture of human endometrial cells in a three-dimensional culture system. J Reprod Fertil. 1994;101(02):327–332. doi: 10.1530/jrf.0.1010327. [DOI] [PubMed] [Google Scholar]
- 117.Kelly R W, King A E, Critchley H O. Cytokine control in human endometrium. Reproduction. 2001;121(01):3–19. doi: 10.1530/rep.0.1210003. [DOI] [PubMed] [Google Scholar]
- 118.Schutte S C, Taylor R N. A tissue-engineered human endometrial stroma that responds to cues for secretory differentiation, decidualization, and menstruation. Fertil Steril. 2012;97(04):997–1003. doi: 10.1016/j.fertnstert.2012.01.098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Arnold J T, Kaufman D G, Seppälä M, Lessey B A. Endometrial stromal cells regulate epithelial cell growth in vitro: a new co-culture model. Hum Reprod. 2001;16(05):836–845. doi: 10.1093/humrep/16.5.836. [DOI] [PubMed] [Google Scholar]
- 120.Abbas Y, Brunel L G, Hollinshead M S. Generation of a three-dimensional collagen scaffold-based model of the human endometrium. Interface Focus. 2020;10(02):2.0190079E7. doi: 10.1098/rsfs.2019.0079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Hernandez-Gordillo V, Kassis T, Lampejo A. Fully synthetic matrices for in vitro culture of primary human intestinal enteroids and endometrial organoids. Biomaterials. 2020;254:120125. doi: 10.1016/j.biomaterials.2020.120125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Wolf K, Alexander S, Schacht V. Collagen-based cell migration models in vitro and in vivo. Semin Cell Dev Biol. 2009;20(08):931–941. doi: 10.1016/j.semcdb.2009.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Lutolf M P, Hubbell J A. Synthesis and physicochemical characterization of end-linked poly(ethylene glycol)-co-peptide hydrogels formed by Michael-type addition. Biomacromolecules. 2003;4(03):713–722. doi: 10.1021/bm025744e. [DOI] [PubMed] [Google Scholar]
- 124.Caliari S R, Burdick J A. A practical guide to hydrogels for cell culture. Nat Methods. 2016;13(05):405–414. doi: 10.1038/nmeth.3839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Brown A, He H, Trumper E, Valdez J, Hammond P, Griffith L G. Engineering PEG-based hydrogels to foster efficient endothelial network formation in free-swelling and confined microenvironments. Biomaterials. 2020;243:119921. doi: 10.1016/j.biomaterials.2020.119921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Kyburz K A, Anseth K S. Synthetic mimics of the extracellular matrix: how simple is complex enough? Ann Biomed Eng. 2015;43(03):489–500. doi: 10.1007/s10439-015-1297-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.DiMarco R L, Dewi R E, Bernal G, Kuo C, Heilshorn S C. Protein-engineered scaffolds for in vitro 3D culture of primary adult intestinal organoids. Biomater Sci. 2015;3(10):1376–1385. doi: 10.1039/c5bm00108k. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Stegemann J P, Hong H, Nerem R M. Mechanical, biochemical, and extracellular matrix effects on vascular smooth muscle cell phenotype. J Appl Physiol (1985) 2005;98(06):2321–2327. doi: 10.1152/japplphysiol.01114.2004. [DOI] [PubMed] [Google Scholar]
- 129.Dallot E, Pouchelet M, Gouhier N, Cabrol D, Ferré F, Breuiller-Fouché M. Contraction of cultured human uterine smooth muscle cells after stimulation with endothelin-1. Biol Reprod. 2003;68(03):937–942. doi: 10.1095/biolreprod.102.008367. [DOI] [PubMed] [Google Scholar]
- 130.Rajagopal S P, Hutchinson J L, Dorward D A, Rossi A G, Norman J E. Crosstalk between monocytes and myometrial smooth muscle in culture generates synergistic pro-inflammatory cytokine production and enhances myocyte contraction, with effects opposed by progesterone. Mol Hum Reprod. 2015;21(08):672–686. doi: 10.1093/molehr/gav027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Devost D, Zingg H H. Novel in vitro system for functional assessment of oxytocin action. Am J Physiol Endocrinol Metab. 2007;292(01):E1–E6. doi: 10.1152/ajpendo.00529.2005. [DOI] [PubMed] [Google Scholar]
- 132.Wendremaire M, Hadi T, Pezze M. Macrophage-induced reactive oxygen species promote myometrial contraction and labor-associated mechanisms. Biol Reprod. 2020;102(06):1326–1339. doi: 10.1093/biolre/ioaa032. [DOI] [PubMed] [Google Scholar]
- 133.Peyton S R, Raub C B, Keschrumrus V P, Putnam A J. The use of poly(ethylene glycol) hydrogels to investigate the impact of ECM chemistry and mechanics on smooth muscle cells. Biomaterials. 2006;27(28):4881–4893. doi: 10.1016/j.biomaterials.2006.05.012. [DOI] [PubMed] [Google Scholar]
- 134.Lauffenburger D A, Horwitz A F. Cell migration: a physically integrated molecular process. Cell. 1996;84(03):359–369. doi: 10.1016/s0092-8674(00)81280-5. [DOI] [PubMed] [Google Scholar]
- 135.Beunk L, Brown K, Nagtegaal I, Friedl P, Wolf K. Cancer invasion into musculature: mechanics, molecules and implications. Semin Cell Dev Biol. 2019;93:36–45. doi: 10.1016/j.semcdb.2018.07.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Weimar C H, Macklon N S, Post Uiterweer E D, Brosens J J, Gellersen B. The motile and invasive capacity of human endometrial stromal cells: implications for normal and impaired reproductive function. Hum Reprod Update. 2013;19(05):542–557. doi: 10.1093/humupd/dmt025. [DOI] [PubMed] [Google Scholar]
- 137.Maheshwari G, Brown G, Lauffenburger D A, Wells A, Griffith L G.Cell adhesion and motility depend on nanoscale RGD clustering J Cell Sci 2000113(Pt 10):1677–1686. [DOI] [PubMed] [Google Scholar]
- 138.Zaman M H, Trapani L M, Sieminski A L. Migration of tumor cells in 3D matrices is governed by matrix stiffness along with cell-matrix adhesion and proteolysis. Proc Natl Acad Sci U S A. 2006;103(29):10889–10894. doi: 10.1073/pnas.0604460103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Tan C W, Lee Y H, Tan H H. CD26/DPPIV down-regulation in endometrial stromal cell migration in endometriosis. Fertil Steril. 2014;102(01):167–1.77E11. doi: 10.1016/j.fertnstert.2014.04.001. [DOI] [PubMed] [Google Scholar]
- 140.Yang J-H, Wu M Y, Chen M J, Chen S U, Yang Y S, Ho H N. Increased matrix metalloproteinase-2 and tissue inhibitor of metalloproteinase-1 secretion but unaffected invasiveness of endometrial stromal cells in adenomyosis. Fertil Steril. 2009;91(05):2193–2198. doi: 10.1016/j.fertnstert.2008.05.090. [DOI] [PubMed] [Google Scholar]
- 141.Mehasseb M K, Taylor A H, Pringle J H, Bell S C, Habiba M. Enhanced invasion of stromal cells from adenomyosis in a three-dimensional coculture model is augmented by the presence of myocytes from affected uteri. Fertil Steril. 2010;94(07):2547–2551. doi: 10.1016/j.fertnstert.2010.04.016. [DOI] [PubMed] [Google Scholar]
- 142.Taylor A H, Kalathy V, Habiba M. Estradiol and tamoxifen enhance invasion of endometrial stromal cells in a three-dimensional coculture model of adenomyosis. Fertil Steril. 2014;101(01):288–293. doi: 10.1016/j.fertnstert.2013.09.042. [DOI] [PubMed] [Google Scholar]
- 143.Palmer S S, Altan M, Denis D. Bentamapimod (JNK inhibitor AS602801) induces regression of endometriotic lesions in animal models. Reprod Sci. 2016;23(01):11–23. doi: 10.1177/1933719115600553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Hussein M, Chai D C, Kyama C M. c-Jun NH2-terminal kinase inhibitor bentamapimod reduces induced endometriosis in baboons: an assessor-blind placebo-controlled randomized study. Fertil Steril. 2016;105(03):815–8.24E7. doi: 10.1016/j.fertnstert.2015.11.022. [DOI] [PubMed] [Google Scholar]
- 145.Gnecco J S, Ding T, Smith C, Lu J, Bruner-Tran K L, Osteen K G. Hemodynamic forces enhance decidualization via endothelial-derived prostaglandin E2 and prostacyclin in a microfluidic model of the human endometrium. Hum Reprod. 2019;34(04):702–714. doi: 10.1093/humrep/dez003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Goddard L M, Murphy T J, Org T. Progesterone receptor in the vascular endothelium triggers physiological uterine permeability preimplantation. Cell. 2014;156(03):549–562. doi: 10.1016/j.cell.2013.12.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Maggioli E, McArthur S, Mauro C. Estrogen protects the blood-brain barrier from inflammation-induced disruption and increased lymphocyte trafficking. Brain Behav Immun. 2016;51:212–222. doi: 10.1016/j.bbi.2015.08.020. [DOI] [PubMed] [Google Scholar]
- 148.Pence J C, Clancy K BH, Harley B AC. Proangiogenic activity of endometrial epithelial and stromal cells in response to estradiol in gelatin hydrogels. Adv Biosyst. 2017;1(09):1.700056E6. doi: 10.1002/adbi.201700056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Gnecco J S, Pensabene V, Li D J. Compartmentalized culture of perivascular stroma and endothelial cells in a microfluidic model of the human endometrium. Ann Biomed Eng. 2017;45(07):1758–1769. doi: 10.1007/s10439-017-1797-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Zervantonakis I K, Hughes-Alford S K, Charest J L, Condeelis J S, Gertler F B, Kamm R D. Three-dimensional microfluidic model for tumor cell intravasation and endothelial barrier function. Proc Natl Acad Sci U S A. 2012;109(34):13515–13520. doi: 10.1073/pnas.1210182109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Hsu Y H, Moya M L, Hughes C C, George S C, Lee A P. A microfluidic platform for generating large-scale nearly identical human microphysiological vascularized tissue arrays. Lab Chip. 2013;13(15):2990–2998. doi: 10.1039/c3lc50424g. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Chen M B, Whisler J A, Fröse J, Yu C, Shin Y, Kamm R D. On-chip human microvasculature assay for visualization and quantification of tumor cell extravasation dynamics. Nat Protoc. 2017;12(05):865–880. doi: 10.1038/nprot.2017.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Chen M B, Hajal C, Benjamin D C. Inflamed neutrophils sequestered at entrapped tumor cells via chemotactic confinement promote tumor cell extravasation. Proc Natl Acad Sci U S A. 2018;115(27):7022–7027. doi: 10.1073/pnas.1715932115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Rambøl M H, Han E, Niklason L E.Microvessel network formation and interactions with pancreatic islets in three-dimensional chip cultures Tissue Eng Part A 202026(9-10):556–568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Aref A R, Campisi M, Ivanova E. 3D microfluidic ex vivo culture of organotypic tumor spheroids to model immune checkpoint blockade. Lab Chip. 2018;18(20):3129–3143. doi: 10.1039/c8lc00322j. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Campisi M, Shin Y, Osaki T, Hajal C, Chiono V, Kamm R D. 3D self-organized microvascular model of the human blood-brain barrier with endothelial cells, pericytes and astrocytes. Biomaterials. 2018;180:117–129. doi: 10.1016/j.biomaterials.2018.07.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Sobrino A, Phan D T, Datta R. 3D microtumors in vitro supported by perfused vascular networks. Sci Rep. 2016;6:31589. doi: 10.1038/srep31589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Bersini S, Gilardi M, Ugolini G S. Engineering an environment for the study of fibrosis: a 3D human muscle model with endothelium specificity and endomysium. Cell Rep. 2018;25(13):3858–3.868E7. doi: 10.1016/j.celrep.2018.11.092. [DOI] [PubMed] [Google Scholar]
- 159.Regehr K J, Domenech M, Koepsel J T. Biological implications of polydimethylsiloxane-based microfluidic cell culture. Lab Chip. 2009;9(15):2132–2139. doi: 10.1039/b903043c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Ivanova E, Kuraguchi M, Xu M. Use of ex vivo patient-derived tumor organotypic spheroids to identify combination therapies for HER2 mutant non-small cell lung cancer . Clin Cancer Res. 2020;26(10):2393–2403. doi: 10.1158/1078-0432.CCR-19-1844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Gilbert P M, Weaver V M. Cellular adaptation to biomechanical stress across length scales in tissue homeostasis and disease. Semin Cell Dev Biol. 2017;67:141–152. doi: 10.1016/j.semcdb.2016.09.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Purdy M P, Ducharme M, Haak A J. YAP/TAZ are activated by mechanical and hormonal stimuli in myometrium and exhibit increased baseline activation in uterine fibroids. Reprod Sci. 2020;27(04):1074–1085. doi: 10.1007/s43032-019-00106-4. [DOI] [PubMed] [Google Scholar]
- 163.Kim J, Ushida T, Montagne K. Acquired contractile ability in human endometrial stromal cells by passive loading of cyclic tensile stretch. Sci Rep. 2020;10(01):9014. doi: 10.1038/s41598-020-65884-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Copley Salem C, Ulrich C, Quilici D, Schlauch K, Buxton I LO, Burkin H. Mechanical strain induced phospho-proteomic signaling in uterine smooth muscle cells. J Biomech. 2018;73:99–107. doi: 10.1016/j.jbiomech.2018.03.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Elad D, Zaretsky U, Kuperman T. Tissue engineered endometrial barrier exposed to peristaltic flow shear stresses. APL Bioeng. 2020;4(02):26107. doi: 10.1063/5.0001994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Huang A H, Niklason L E. Engineering biological-based vascular grafts using a pulsatile bioreactor. J Vis Exp. 2011;(52):2646. doi: 10.3791/2646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Wainger B J, Buttermore E D, Oliveira J T. Modeling pain in vitro using nociceptor neurons reprogrammed from fibroblasts. Nat Neurosci. 2015;18(01):17–24. doi: 10.1038/nn.3886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Lemke K A, Aghayee A, Ashton R S. Deriving, regenerating, and engineering CNS tissues using human pluripotent stem cells. Curr Opin Biotechnol. 2017;47:36–42. doi: 10.1016/j.copbio.2017.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Puzan M, Hosic S, Ghio C, Koppes A. Enteric nervous system regulation of intestinal stem cell differentiation and epithelial monolayer function. Sci Rep. 2018;8(01):6313. doi: 10.1038/s41598-018-24768-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Osaki T, Sivathanu V, Kamm R D. Crosstalk between developing vasculature and optogenetically engineered skeletal muscle improves muscle contraction and angiogenesis. Biomaterials. 2018;156:65–76. doi: 10.1016/j.biomaterials.2017.11.041. [DOI] [PubMed] [Google Scholar]
- 171.Oleaga C, Bernabini C, Smith A S. Multi-organ toxicity demonstration in a functional human in vitro system composed of four organs. Sci Rep. 2016;6:20030. doi: 10.1038/srep20030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Edington C D, Chen W LK, Geishecker E. Interconnected microphysiological systems for quantitative biology and pharmacology studies. Sci Rep. 2018;8(01):4530. doi: 10.1038/s41598-018-22749-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Trapecar M, Communal C, Velazquez J. Gut-liver physiomimetics reveal paradoxical modulation of IBD-related inflammation by short-chain fatty acids. Cell Syst. 2020;10(03):223–2.39E11. doi: 10.1016/j.cels.2020.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Critchley H OD, Babayev E, Bulun S E.Menstruation: science and society Am J Obstet Gynecol 2020(epub ahead of print). 10.1016/j.ajog.2020.06.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Rahmioglu N, Nyholt D R, Morris A P, Missmer S A, Montgomery G W, Zondervan K T. Genetic variants underlying risk of endometriosis: insights from meta-analysis of eight genome-wide association and replication datasets. Hum Reprod Update. 2014;20(05):702–716. doi: 10.1093/humupd/dmu015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Aghajanova L, Giudice L C. Molecular evidence for differences in endometrium in severe versus mild endometriosis. Reprod Sci. 2011;18(03):229–251. doi: 10.1177/1933719110386241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Huang W, Navarro-Serer B, Jeong Y J. Pattern of invasion in human pancreatic cancer organoids is associated with loss of SMAD4 and clinical outcome. Cancer Res. 2020;80(13):2804–2817. doi: 10.1158/0008-5472.CAN-19-1523. [DOI] [PMC free article] [PubMed] [Google Scholar]