

An image of a pig painted at least 45,500 years ago on a cave wall in Sulawesi may be the earliest figurative rock art ever found.
Abstract
Indonesia harbors some of the oldest known surviving cave art. Previously, the earliest dated rock art from this region was a figurative painting of a Sulawesi warty pig (Sus celebensis). This image from Leang Bulu’ Sipong 4 in the limestone karsts of Maros-Pangkep, South Sulawesi, was created at least 43,900 years ago (43.9 ka) based on Uranium-series dating. Here, we report the Uranium-series dating of two figurative cave paintings of Sulawesi warty pigs recently discovered in the same karst area. The oldest, with a minimum age of 45.5 ka, is from Leang Tedongnge. The second image, from Leang Balangajia 1, dates to at least 32 ka. To our knowledge, the animal painting from Leang Tedongnge is the earliest known representational work of art in the world. There is no reason to suppose, however, that this early rock art is a unique example in Island Southeast Asia or the wider region.
INTRODUCTION
Sulawesi is the largest island (~174,000 km2) in Wallacea, a biogeographically distinct zone of oceanic islands situated between continental Asia and Australia (Fig. 1). Sulawesi has a long history of human occupation. The earliest archaeological evidence is from Talepu, a Middle Pleistocene site in the south of the island (1). The Talepu findings comprise in situ stone artifacts associated with fossils of extinct terrestrial megafauna (1). Dated to ~194 to 118 thousand years (ka), these artifacts may reflect initial colonization by an as-yet unidentified archaic hominin (1). It is not yet certain when anatomically modern humans (AMH) first colonized Sulawesi. AMH appear to have been established in mainland Southeast Asia (Sunda) by 73 to 63 ka (2). They were also possibly in Pleistocene Australia–New Guinea (Sahul) by 69 to 59 ka (3). There is some controversy, however, over the validity of the latter age estimates (4), which are based on recent excavations at Madjedbebe rock shelter in northern Australia (3). In some models of early human settlement in Sahul, the large Sulawesi landmass would have been the first “stop” on a series of ocean crossings through northern Wallacea to the western tip of New Guinea (5). If the oldest published dates for Madjedbebe (3) are acceptable, then Homo sapiens may have reached Sulawesi up to 69 to 59 ka.
Fig. 1. South Sulawesi study area.
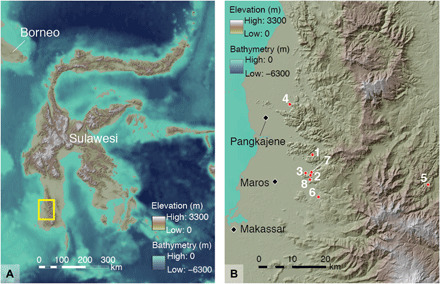
(A) The rock art sites under study are all situated in limestone karst areas in the island’s southwestern peninsula [see (B) for detail of area shown inside the rectangle]. (B) Locations of limestone caves or shelters discussed in the study: 1, Leang Tedongnge; 2, Leang Timpuseng; 3, Leang Barugayya 2; 4, Leang Bulu’ Sipong 4; 5, Gua Uhallie; 6, Leang Balangajia 1; 7, Leang Bulu Bettue; and 8, Leang Burung 2. Map data: Shuttle Radar Topography Mission 1 Arc-Second Global by NASA/NGS/USGS and GEBCO_2014 Grid version 20150318 (http://gebco.net). Base maps were created by M. Kottermair and A. Jalandoni.
Thus far, the earliest dated proxy evidence for AMH in Sulawesi consists of prehistoric rock art (6, 7). Rock art was first identified 70 years ago (8) in Maros-Pangkep, a ~450-km2 lowland “tower” karst region in the island’s southwestern peninsula (Fig. 1). Some 300 caves and shelters with parietal imagery have now been identified in this area. Two cave art sites are also known from the Bone karsts ~35 km to the east (Fig. 1). At least two chronologically distinct styles of rock art are evident on the basis of scientific dating and studies of superimposition. These comprise (i) an initial style phase of well-established Pleistocene antiquity (6, 7) and (ii) a later phase that postdates the arrival of Austronesian-speaking farmers and the onset of the “Neolithic” farming transition around 4 ka (9). Where no sequence of superimposition is evident, the former is distinguishable from the latter in terms of subject matter, technique, and preservation (6, 10).
The pre-Austronesian rock art is characterized by hand stencils and figurative animal paintings. In most instances, animal images were executed in a single color (typically red or purple/mulberry) using brushwork and/or fingertips. The artists used somewhat simplified forms of outline representation to depict animals in profile. The animal outlines are typically infilled with irregular patterns of painted lines and dashes rather than recognizable anatomical detail; solid infill is documented, but rare. Most discernible animal images appear to represent the island’s largest endemic land mammals: suids and dwarf bovids (anoas, Bubalus sp.) (11). Images of suids are strongly dominant: Thus far, some 73 separate parietal motifs representing suids or suid-like figures (81.1% of animal representations) have been identified in Maros-Pangkep and Bone. Most seem to portray Sus celebensis (Sulawesi warty pig), a small (40 to 85 kg), short-legged pig with characteristic facial warts (12–15). S. celebensis is still extant (12–15). Although an old Sulawesi endemic, this species was widely translocated within Indonesia in late prehistory (14).
In total, five figurative paintings of animals from Maros-Pangkep have previously been dated to the Late Pleistocene using Uranium-series (U-series) isotope analysis of associated calcium carbonate deposits (coralloid speleothems) (6, 7). The earliest dated images occur on the same rock art panel at the limestone cave of Leang Bulu’ Sipong 4 (Fig. 1) (7). The 4.5-m-wide panel at this site consists of a scene portraying several human-like figures apparently hunting Sulawesi warty pigs and anoas (7). U-series isotope analysis was undertaken on four coralloid speleothems from three separate animal images (7). A Sulawesi warty pig image has a minimum age of 43.9 ka, while two anoa images have minimum ages of 41 and 40.9 ka, respectively (7). At another site, Leang Timpuseng, a figurative depiction of a suid was dated to at least 35.4 ka (6). Moreover, at Leang Barugayya 2, a painting of an unidentified suid-like animal has a minimum age of 35.7 ka (6). U-series dating of hand stencils (N = 12) from various sites in Maros-Pangkep also yielded minimum ages of between 39.9 and 17.4 ka (6). Notably, excavations at two rock art sites in the Maros district, Leang Bulu Bettue (16) and Leang Burung 2 (17) (Fig. 1), have revealed in situ archaeological evidence for Late Pleistocene pigment processing. This includes used ochre fragments at both sites and, at Leang Bulu Bettue, ochre-stained stone artifacts deposited around 40 ka (16). At the latter site, excavated examples of figurative and nonrepresentational artworks (“portable art”) also date to ~24 to 16 ka (18). Evidence for rock art production of a broadly similar antiquity is found in karst areas in the adjacent island of Borneo: At Lubang Jeriji Saléh, a high-level limestone cave in Kalimantan, U-series dating indicates a minimum age of 40 ka for a figurative painting of a Bornean banteng (Bos javanicus lowi) (19).
In the present study, we describe the recent discovery and U-series dating of two Late Pleistocene figurative paintings of Sulawesi warty pigs in Maros-Pangkep. These parietal artworks are from two newly identified cave art sites: Leang Tedongnge and Leang Balangajia 1. We describe the animal images at these localities and report the U-series dating results on associated speleothems.
RESULTS
Materials and methods
We undertook U-series isotope analysis of small coralloid speleothems that had formed on two figurative paintings of suids at Leang Tedongnge and Leang Balangajia 1, as well as a superimposed hand stencil at the latter site. One of the suid images (Figs. 2 and 3) is located at Leang Tedongnge, a limestone cave in a secluded karst valley on the border of the Maros and Pangkep karst districts (Fig. 1). The other suid image (Fig. 4) is from the limestone cave of Leang Balangajia 1 in the southern Maros district of Simbang (Fig. 1). We discovered these previously unknown rock art sites in 2017 and 2018, respectively. Figures 2 to 4 provide site descriptions and additional information pertaining to the context of the prehistoric artworks at both localities.
Fig. 2. Figurative paintings of pigs at Leang Tedongnge.
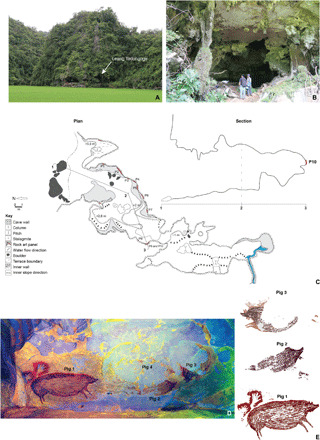
(A and B) Leang Tedongnge cave. The cave is located at the foot of a limestone karst hill (A); the cave mouth entrance is shown in (B). (C) Plan and section of Leang Tedongnge site. Rock art panel 10 (P10) is located on a ledge toward the rear of the cave and features at least three large figurative paintings of pigs. (D and E) Details of rock art panel 10 showing pigs 1 to 3. Dimensions: pig 1 (136 cm by 54 cm), pig 2 (125 cm by 53 cm), and pig 3 (138 cm by 71 cm). A fourth animal figure (probably a depiction of a pig and hence provisionally denoted pig 4) was present but is too heavily exfoliated for analysis. (D) Stitched panorama view of panel 10 after enhancing with the Decorrelation Stretch (DStretch) computer program (44) (channel setting: _lds; LDS colorspace). (E) Tracings of pigs 1 to 3. Pig 3 was painted using two different colors; possibly, but not certainly, with a time lapse between these painting episodes. Photo credits: A. A. Oktaviana, ARKENAS/Griffith University.
Fig. 3. Dated pig figure (pig 1) in the rock art panel at Leang Tedongnge.

(A) Digital tracing of the rock art panel (P10). (B) Pig 1. (C) Detail of the head area of pig 1 [enhanced using the Decorrelation Stretch (DStretch) computer program (44); channel setting: _ac_lab; AC, auto contrast and LAB color space]. HC, head crest; FW, preorbital facial warts. Pig 1 displays an unidentified morphological feature, a pair of teat-like protuberances in the lower neck area; these are highlighted by white arrows in (C). Photo credits: A. A. Oktaviana, ARKENAS/Griffith University.
Fig. 4. Dated painting of a pig at Leang Balangajia 1.
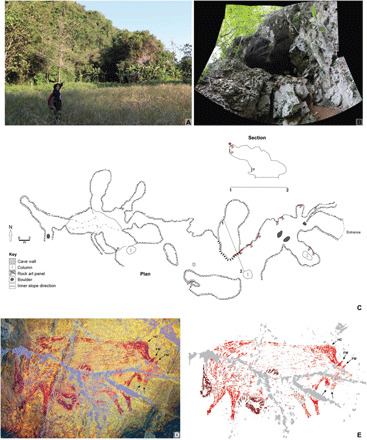
(A and B) Leang Balangajia 1 cave. The cave is located in the upper karst network of a limestone hill (A); the cave entrance [concealed by dense vegetation in (A)] is shown in the photostitched panorama in (B). (C) Plan and section of Leang Balangajia 1 (the red star shows the location of the dated pig figure). (D and E) Dated artwork, a large red outline profile painting of a pig. (D) Photograph enhanced using the Decorrelation Stretch (DStretch) computer program (44) (channel setting: _ac_cb_lds; AC; CB, auto image color balance; and LDS color space). (E) Tracing (gray shading indicates exfoliated areas of the panel). The pig figure is 187 cm in length and 110 cm in height. FW, preorbital facial warts, a diagnostic characteristic of S. celebensis; HC, head crest. Four hand stencils (darker in color) are superimposed on the pig. The pig figure displays an unidentified morphological feature, a pair of hairy teat-like protuberances in the lower neck area [highlighted by arrows in (D) and (C)]. Photo credits: A. A. Oktaviana, ARKENAS/Griffith University.
The suid images at Leang Tedongnge (Figs. 2 and 3) and Leang Balangajia 1 (Fig. 4) were both executed in red or dark red/purplish (mulberry) mineral pigments (ochre). In each instance, the animal depiction consists of a complete body outline of a suid shown in profile view. Both suid figures are represented in immobile or static positions. The pictorial outlines are infilled with irregular patterns of painted lines and dashes. The prehistoric artists did not clearly portray any primary sexual characteristics (e.g., genitalia and mammary glands) in the outline representations of the suids; however, secondary sexual characteristics may have been depicted (see below).
At Leang Tedongnge, the dated suid figure (pig 1) is positioned on the rear wall of the cave (Fig. 2). Measuring 136 cm by 54 cm, this motif is associated with two hand stencils situated above and close to the hindquarters of the suid representation. At least two or three other suid figures (pigs 2 and 3) are located on the same panel (Figs. 2 and 3). In contrast to pig 1, which is relatively complete, the latter images are only partially preserved owing to exfoliation of the cave wall “canvas” on which they were painted. Clearly visible, however, are the heads of two of the suids. The animal figures seem to have been depicted confronting each other. The dated pig figure is positioned to the left of pigs 2 and 3 and faces toward them. A fourth animal image, almost completely obliterated by exfoliation of the rock surface, is located on the same panel above pigs 2 and 3. The arrangement of the figures is suggestive, in our view, of a narrative composition or scene in the modern Western sense (20). We infer that this rock art panel may have been intended to portray an episode of social interaction between at least three, and possibly four, individual suids.
At Leang Balangajia 1, the large red pig figure was painted on the ceiling of a small side chamber. This suid motif measures 187 cm in length and 110 cm in height (Fig. 4). There are possibly at least two other figurative animal motifs on the wall and ceiling of the chamber. They are very poorly preserved, however, and largely indistinct; hence, we do not consider them further here. Four hand stencils are superimposed on top of the large red pig figure (Fig. 4).
U-series isotope dating at Leang Tedongnge
We conducted U-series isotope analysis on a small coralloid speleothem (sample LTed3) overlying the complete figurative pig painting (pig 1) at Leang Tedongnge (Fig. 5). In the field, we observed that the coralloid speleothem (~12 mm2) overlaid the red pigment associated with one of the rear feet of pig 1 on the main rock art panel (Fig. 5, A to C). This speleothem was too small in size to remove with a rotary tool. Hence, we used a small chisel to prise it from the cave wall surface. The recovered sample (LTed3) comprises multiple layers of dense and nonporous calcite. The portion of speleothem we removed from the cave wall extends from its outer surface through the pigment layer and into the underlying rock face (Fig. 5, D and E). After we had removed it from the cave wall, we noted that the portion of the underlying paint layer previously covered by the coralloid speleothem had come away with the latter—that is, the rock art pigment was affixed to the base of the sampled speleothem. In the laboratory, we micro-excavated LTed3 in arbitrary “spits” that extended over the entire surface of the speleothem. This produced a series of four aliquots measuring less than 1 mm in thickness (Fig. 5, F to G). We observed the red pigment layer corresponding to the artwork across the entire length of the sample. The pigment layer was also clearly visible in the rear portion of the sample that was partially covered by translucent calcite from the cave wall (Fig. 5E). In total, we obtained three U-series age determinations for the LTed3 speleothem (Table 1). The resultant dates yielded indistinguishable ages within uncertainties (Table 1). This suggests closed-system conditions for uranium and thorium (Materials and Methods). The results of U-series dating indicate a minimum age of 45.5 ka for the large figurative image of a suid at Leang Tedongnge (Fig. 3 and Materials and Methods).
Fig. 5. U-series dating of the pig figure (pig 1) at Leang Tedongnge.
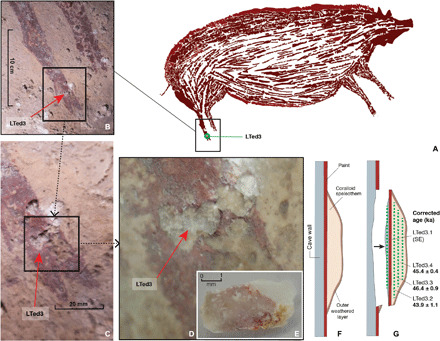
(A) Sample LTed3 comprises a coralloid speleothem overlying the figure’s rear foot. (B to D) Sample LTed3 overlying red pigment of the foot. (E) Underside of sample LTed3 after removal from the rock art panel; clearly visible is the red pigment of the paint layer adhering to the base of the coralloid speleothem. (F to G) Schematic diagrams of sample removal and dating. (F) Sampled coralloid speleothem (LTed3) formed on top of the paint layer corresponding to the rear foot of the pig. (G) When removed, the sampled coralloid speleothem (LTed3) took the underlying paint layer with it affixed to the base (see Materials and Methods). U-series isotope analysis was conducted on four micromilled subsamples (green dotted lines); one aliquot was collected from immediately below the pigment layer (LTed3.1), and the remaining three aliquots (LTed3.2, LTed3.3, and LTed3.4) came from progressively higher levels above (i.e., on top of) the pigment layer. SE, secular equilibrium. Uncertainties reported at 2σ. Our results indicate a minimum U-series age of 45.5 ka for this figurative image of a suid at Leang Tedongnge. Photo credits: M. Aubert, Griffith University.
Table 1. U-series dating results.
U-series dating results for Leang Tedongnge (LTed3) and Leang Balangajia 1 (LBLGJ1 and LBLGJ2). U-Th isotope ratios are activity ratios. ppm, parts per million.
| Sample | Description |
Sample weight (mg) |
238U (ppm) |
230Th/232Th | 230Th/238U | 234U/238U |
Uncorrected age (ka) |
±2σ (ka) |
Corrected age (ka) |
±2σ (ka) |
Corrected initial 234U/238U |
| LTed3.4 | Sequence of aliquots |
1.6 | 13.3 | 355 | 0.1841 ± 0.0010 | 0.5665 ± 0.0011 | 45.53 | 0.36 | 45.44 | 0.36 | 0.5070 ± 0.0016 |
| LTed3.3 | Sequence of aliquots |
1.5 | 2.1 | 57 | 0.1878 ± 0.0025 | 0.5645 ± 0.0018 | 46.99 | 0.86 | 46.40 | 0.94 | 0.5022 ± 0.0028 |
| LTed3.2 | Overlies animal figure |
1.5 | 2.1 | 37 | 0.1836 ± 0.0029 | 0.5717 ± 0.0018 | 44.74 | 0.96 | 43.88 | 1.10 | 0.5133 ± 0.0030 |
| LTed3.1 | Underlies animal figure |
1.1 | 1.4 | 137 | 0.9229 ± 0.0075 | 0.9247 ± 0.0027 | SE | ||||
| LBLGJ1.1 | Sequence of aliquots |
5.6 | 2.37 | 354 | 0.2848 ± 0.0015 | 0.982 ± 0.0011 | 37.39 | 0.24 | 37.32 | 0.24 | 0.9800 ± 0.0013 |
| LBLGJ1.2 | Sequence of aliquots |
5.6 | 3.03 | 587 | 0.2348 ± 0.0008 | 0.9819 ± 0.0005 | 29.83 | 0.11 | 29.79 | 0.11 | 0.9803 ± 0.0006 |
| LBLGJ1.3 | Overlies animal figure |
5.7 | 2.17 | 613 | 0.2318 ± 0.0010 | 0.9791 ± 0.0011 | 29.49 | 0.15 | 29.45 | 0.15 | 0.9773 ± 0.0012 |
| LBLGJ1.4 | Underlies animal figure |
5.7 | 1.71 | 301 | 0.5173 ± 0.0018 | 0.9787 ± 0.0012 | 82.29 | 0.45 | 82.13 | 0.46 | 0.9731 ± 0.0016 |
| LBLGJ2.1 | Sequence of aliquots |
6.0 | 2.44 | 460 | 0.2376 ± 0.0010 | 0.9733 ± 0.0011 | 30.56 | 0.16 | 30.51 | 0.16 | 0.9709 ± 0.0012 |
| LBLGJ2.2 | Sequence of aliquots |
5.6 | 1.81 | 355 | 0.2373 ± 0.0011 | 0.9710 ± 0.0012 | 30.60 | 0.17 | 30.54 | 0.17 | 0.9684 ± 0.0013 |
| LBLGJ2.3 | Sequence of aliquots |
5.2 | 1.66 | 476 | 0.2262 ± 0.0015 | 0.9697 ± 0.0014 | 29.01 | 0.22 | 28.96 | 0.22 | 0.9671 ± 0.0016 |
| LBLGJ2.4 | Overlies hand stencil |
5.3 | 1.63 | 360 | 0.2497 ± 0.0015 | 0.9736 ± 0.0010 | 32.36 | 0.23 | 32.29 | 0.24 | 0.9711 ± 0.0011 |
| LBLGJ2.5 | Underlies hand stencil |
1.0 | 1.52 | 173 | 0.4743 ± 0.0033 | 0.9756 ± 0.0030 | 72.87 | 0.81 | 72.61 | 0.82 | 0.97 ± 0.0038 |
U-series isotope dating at Leang Balangajia 1
We conducted U-series isotope analysis on two small coralloid speleothems (samples LBLGJ1 and LBLGJ2) overlying the large red pig painting and a hand stencil, respectively (Fig. 6). As we have previously noted, the hand stencil is superimposed on top of the pig figure; therefore, LBLGJ2 also provides a minimum age for the underlying pig figure. We removed samples LBLGJ1 (~150 mm2) and LBLGJ2 (~100 mm2) from the rock art panel using a battery-operated rotary tool equipped with a diamond saw blade. In the laboratory, we micro-excavated each sample in arbitrary spits extending over the surface of each speleothem, creating a series of four (LBLGJ1) and five (LBLGJ2) aliquots per sample measuring less than 1 mm in thickness. The resultant sections consist of two continuous microstratigraphic profiles extending from the outer surface of each speleothem through the pigment layers and into the underlying rock face. The samples overlying both the pig figure and the hand stencil comprise multiple layers of dense and nonporous calcite. In the case of both sampled coralloids, we clearly observed the pigment layer corresponding to each artwork (the red pig figure and the hand stencil) across the entire length of each sample. In total, we obtained nine U-series age determinations for the red pig figure and the hand stencil overlying it (Table 1 and Fig. 6, D and E). Sample LBLGJ1 yielded an older age at the surface of the sample. Because this aliquot (LBLGJ1.1) includes parts of the weathered and friable crust of the speleothem, we interpret this older age as being caused by partial leaching of uranium at the surface. Samples LBLGJ1.2 and LBLGJ1.3 show a slightly reversed age profile (<100 years). We attribute this to individual aliquots sampling a series of concentric growth layers and/or “mounds” of widely varying ages (Materials and Methods) (7). The oldest minimum U-series age for the red pig figure is 29.7 ka (LBLGJ1.2). The maximum age for this painting (82.6 ka) is provided by sample LBLGJ1.4. Sample LBLGJ2 displays an age profile typical of closed-system conditions for uranium and thorium, except for sample LBLGJ2.3, which has a younger age (~2000 years). We also interpret this anomaly as a by-product of our microsampling strategy and the convoluted process of coralloid speleothem formation (Materials and Methods). The internal growth structure revealed in the cross section (Fig. 6E) is obviously more complex, implying a high probability of cross-growth layers mixing in different proportions among different aliquots during the microsampling process. We therefore consider that the oldest minimum U-series date of 32 ka given by subsample LBLGJ2.4 represents the minimum age of the hand stencil, whereas the maximum age for this particular artwork (73.4 ka) is provided by the U-series date for underlying sample LBLGJ2.5. Because the hand stencil is superimposed on top of the large red pig painting, we are able to bracket the age of this figurative animal depiction to between 73.4 and 32 ka.
Fig. 6. U-series dating of the red pig figure at Leang Balangajia 1.

U-series isotope analysis was conducted on two small coralloid speleothems (samples LBLGJ1 and LBLGJ2) overlying the large red pig painting (A) and an overlying purple/mulberry hand stencil (A), respectively. The photograph in (A) has been enhanced using the Decorrelation Stretch (DStretch) computer program (44) (channel setting: _ac_cb_lds; AC, CB, and LDS color space). Samples LBLGJ1 (~150 mm2) and LBLGJ2 (~100 mm2) are shown in situ in (B) and (C), respectively; cross sections with U-series dating results are shown in (D) and (E). Subsamples LBLGJ1.4 (D) and LBLGJ2.5 (E) were both collected below the pigment lines and thus provide maximum ages for the artworks. Uncertainties reported at 2σ. Our dating results indicate that the suid figure was painted between 73.4 and 32 ka. Photo credits: (A) A. A. Oktaviana, ARKENAS/Griffith University; (B to E) M. Aubert, Griffith University.
Identity of the suids depicted in the rock art
Various identifying features of the suid images painted at Leang Tedongnge and Leang Balangajia 1 allow us to conclude that these animal representations are artistic depictions of S. celebensis. The artists did not obviously portray any discernible anatomical details within the suid profiles, such as eyes and ears, muscle tone, pelage markings and coloration, snout bands, canine teeth (tusks), and so on (although it is possible that an eye may be represented on the Leang Balangajia 1 pig; Fig. 4). Nevertheless, we contend that two diagnostic traits of S. celebensis (12–15) are identifiable in the pictorial outlines (Figs. 2 to 4): (i) “spiky” head crests, represented by a row of short dashed lines in the crown and upper back area; and (ii) preorbital facial warts, represented by two conspicuous, horn-like protrusions depicted side by side in the upper snout area.
The first anatomical trait is clearly portrayed and easily distinguished on both pig figures. With regard to the second trait, the protuberances on top of the snouts superficially resemble ears. Even allowing for artistic license, however, these forms are positioned too far along the snout and are too large, in our opinion, to plausibly represent ears (S. celebensis has small ears that are located well behind the eyes) (12–15). They also do not seem to be stylized or distorted representations of canine teeth, which, in S. celebensis, project laterally from the mouth (12–15). We surmise that these features are most consistent with the preorbital facial warts of Sulawesi warty pigs. These prominent warts grow in pairs in front of the eyes and constitute one of the most distinguishing morphological characteristics of S. celebensis (12–15). Facial warts are most pronounced on males, growing larger (and more striking) with age (12–15). We therefore consider that the dated suid motifs from Leang Tedongnge and Leang Balangajia 1 may be depictions of adult male Sulawesi warty pigs. It should be noted that the artists portrayed preorbital warts in the so-named twisted perspective. This is a common method of graphical representation in prehistoric art that entails using a single outline profile image of an animal to depict how it appears to an onlooker when observed from different viewpoints (21).
The dated pig images are, in our view, inconsistent with the external morphology of the only other still-extant species of endemic Sulawesi suid: the babirusa (Babyrousa sp.) (13, 22). The babirusa has unique anatomical characteristics including, in adult males, ornate maxillary canines that instead of pointing downward grow in a spiral posteriorly over the snout (13, 22). Previously, the dated Late Pleistocene suid painting at Leang Timpuseng has been interpreted as a figurative depiction of a female babirusa (6). It is also possible, however, that this image portrays a young female S. celebensis or a subadult male of this species. Celebochoerus spp. was the only other suid taxon that inhabited Sulawesi in pre-Neolithic times (1). However, this now-extinct lineage of “giant” pigs is known only from the Early to Middle Pleistocene fossil record (1); there is no evidence that AMH overlapped in time with these archaic and robust suids in Sulawesi.
The dated suid images from Leang Tedongnge and Leang Balangajia 1 that we identify as S. celebensis were both depicted with an unfamiliar anatomical feature: a pair of protuberances resembling teat-like appendages in the lower neck (Figs. 2 to 4). In the case of the Leang Balangajia 1 pig, the forms depicted seem to be hairy or bristly. They therefore give the impression of being tassel-like in appearance. This unidentified morphological character is evident on the other two suid figures depicted on the same rock art panel at Leang Tedongnge—we also regard these animal motifs as clear representations of S. celebensis (Figs. 2 and 3). Similar forms are evident on two undated large irregular infill outline paintings of S. celebensis at Gua Uhallie in Bone (Fig. 1). We consider possible explanations for this unidentified character in a forthcoming study.
DISCUSSION
In sum, U-series dating results provide respective minimum ages of 45.5 and 32 ka for the S. celebensis images at Leang Tedongnge and Leang Balangajia 1. The image from Leang Tedongnge, with a firmly established minimum age of 45.5 ka, would now appear to be the earliest known dated artwork in Sulawesi. It also represents the oldest reported indication for the presence of AMH on the island and perhaps in the wider Wallacean region (see below). We infer a similar minimum age estimate for the as-yet undated suid motifs visible on the rock art panel at Leang Tedongnge. As noted, together with the dated figure, these suid images seem to constitute a single narrative composition or scene—perhaps a depiction of social interaction between Sulawesi warty pigs. Furthermore, as far as we have been able to ascertain, the securely dated painting of a Sulawesi warty pig at Leang Tedongnge would now seem to be the world’s oldest surviving representational image of an animal. In addition, this dated depiction of an endemic Wallacean suid may also constitute the most ancient figurative artwork known to archaeology.
Concerning the latter, it has recently been claimed that nonfigurative rock art in Spain dates to at least 65 ka and was created by Neanderthals (23); however, the empirical evidence used to support this proposition, dating and otherwise, has been vigorously contested (24–27). As noted, archaeological excavations in South Sulawesi have recovered stone artifacts dating to ~194 to 118 ka (1). These implements are assumed to reflect early colonization of the island by an archaic and now-extinct hominin, not H. sapiens (1). The notion that AMH could have been established at such an early time in Sulawesi or elsewhere in Wallacea, or the wider region, is contentious, but not a priori impossible. On the basis of the fossil evidence, it is now contended that our species had emerged in Africa by 300 ka (28). Moreover, some archaeological and genomic data imply that H. sapiens was established in eastern Asia by 120 ka (29). AMH were also possibly in northernmost Sunda at around the same time (30). On the basis of the presently available evidence, we are unable to definitively conclude that the dated figurative rock art depiction from Leang Tedongnge is the handiwork of cognitively “modern” members of our species. However, this seems to be the most likely explanation given the sophistication of this early representational artwork and the fact that figurative depiction has so far only been attributed to AMH everywhere else in the world.
If so, the dated pig image from Leang Tedongnge would appear to provide some of the earliest evidence, if not the earliest, for the presence of our species in Wallacea. The minimum age of this artwork is compatible with the earliest established indications of AMH from excavated deposits in the Lesser Sunda islands, which formerly provided the oldest archaeological evidence for H. sapiens in Wallacea (~44.6 ka cal BP) (31, 32). Hence, dating results for the Leang Tedongnge painting underline the view (6, 7, 16) that representational art, including figurative animal art and depictions of narrative scenes (7), was a key part of the cultural repertoire of the first AMH populations to cross from Sunda into Wallacea—the gateway to the continent of Australia.
MATERIALS AND METHODS
Experimental design
The coralloid speleothem samples collected in this study formed from thin films of water on cave surfaces over a long period of time. When precipitated from saturated solutions, calcium carbonate usually contains small amounts of soluble uranium (238U and 234U), which eventually decay to 230Th. The latter is essentially insoluble in cave waters and will not precipitate with the calcium carbonate. This produces disequilibrium in the decay chain where all isotopes in the series are no longer decaying at the same rate. Subsequently, 238U and 234U decay to 230Th until secular equilibrium is reached. Because the decay rates are known, the precise measurement of these isotopes allows calculation of the age of the carbonate formation (33).
Methods
U-series dating was carried out using a Nu Plasma multicollector inductively coupled plasma mass spectrometer (MC-ICP-MS) in the Radiogenic Isotope Facility at the School of Earth and Environmental Sciences, University of Queensland, following chemical treatment procedures and MC-ICP-MS analytical protocols that have been previously described (34–36). Powdered subsamples weighing about 1 to 6 mg were spiked with a mixed 229Th-233U tracer and then completely dissolved in concentrated HNO3. After digestion, each sample was treated with H2O2 to decompose trace amounts of organic matters (if any) and to facilitate complete sample-tracer homogenization. U and Th were separated using conventional anion exchange column chemistry using Bio-Rad AG 1-X8 resin. After stripping the matrix from the column using double-distilled 7N HNO3 as eluent, 3 ml of a 2% HNO3 solution mixed with trace amount of Hydrofluoric acid (HF) was used to elute both U and Th into a 3.5-ml precleaned test tube, ready for MC-ICP-MS analyses, without the need for further drying and remixing. After column chemistry, the U-Th mixed solution was injected into the MC-ICP-MS through a DSN-100 desolvation nebulizer system with an uptake rate of around ~0.06 ml per minute. U-Th isotopic ratio measurement was performed on the MC-ICP-MS using a detector configuration to allow simultaneous measurements of both U and Th (36, 37).
It is common for secondary calcium carbonate to be contaminated by detrital materials, such as windblown or waterborne sediments, a process that can lead to U-series ages that are erroneously older than the true age of the sample. This is owing to the preexisting 230Th present in the detrital components, which is broadly analogous to the radiocarbon marine reservoir effect. As the detrital/initial 230Th cannot be physically separated from the radiogenic 230Th for measurement, its contribution to the calculated 230Th age of the sample is often corrected for using an assumed 230Th/232Th activity ratio in the detrital component. Given that the detrital component within a cave is often composed of windblown or waterborne sediments chemically approaching average continental crust, the mean bulk Earth or upper continental crustal value of 232Th/238U = 3.8, corresponding to an 230Th/232Th activity ratio of 0.825, with an arbitrarily assigned uncertainty of 50%, has been commonly assumed for detrital/initial 230Th corrections (31). In this regard, the degree of detrital contamination may be reflected by the measured 230Th/232Th activity ratio in a sample, with a higher value (such as >20) indicating a relatively small or insignificant effect on the calculated age and a lower value (<20) indicating that the correction on the age will be significant (33). Because 232Th in the sample is largely present in the detrital fraction and plays no part in the decay chain of uranium, the detrital 230Th in a sample with a measured 230Th/232Th activity ratio >20 would make up only <0.825/20 = ~4.1% of the total 230Th in the sample.
Sometimes, the assumed 230Th/232Th activity ratio of 0.825 (±50%) for the detrital component may not cover all situations. If the actual 230Th/232Th activity ratio in the detrital component significantly deviates from this assumed range, then the detrital correction scheme may introduce significant bias, especially to samples with 230Th/232Th activity ratio <20. In such situations, the 230Th/232Th activity ratio in the detrital component can be obtained through direct measurement of sediments associated with speleothems (2, 23, 38) or computed using isochron methods or stratigraphical constraints (39). In our case, our samples were relatively pure with the 230Th/232Th activity ratio ranging from 37 to 613 for individual aliquots. Corrections for detrital components were therefore calculated, assuming the bulk Earth values mentioned above.
A conceivable problem with the U-series dating method is that calcium carbonate accretions can behave as an open system for uranium, whereby the element can be leached out of the accretions or remobilized [see, e.g., (40, 41)]. In such instances, the calculated ages will be too old because the dating method relies on the accurate measurement of uranium versus its decay product 230Th. In this study, this problem was tackled by avoiding porous samples and measuring four to five aliquots from every sample. The ages of these subsamples were mostly in chronological order or yielded indistinguishable ages within uncertainties, confirming the integrity of the dated coralloids. If uranium had partially leached out of the samples, then a reverse age profile would have been evident (that is, the ages would have gotten older toward the surface). Samples LBLGJ1 and LBLGJ2 each display a single minor outlier (Table 1). We attribute these inversions to the calcite deposits having accumulated in ring-like formations (e.g., as with onions) rather than roughly flat and parallel to the pigment layer (as in typical flowstones) (40). Alternatively, some coralloid speleothems have complex internal morphologies that reflect their origin as aggregates of a cluster of cylindrical, mound-like calcite structures (40), leaving overhanging features with gaps between older material that are infilled by carbonate materials of younger age. Because the microsampling procedure involves collecting material from an arbitrary depth above the pigment layer, as opposed to sampling individual laminae, the resultant U-series age could—in some instances—be an average of the older mound material and the younger infill. It would not be possible in either case to estimate the proportion of each carbonate component that contributes to the calculated age. In summary, owing to the small size of the dated coralloids LBLGJ1 and LBLGJ2 and the arbitrary nature of the micro-excavated spits, it is likely that individual aliquots average out a series of concentric growth layers and/or mounds of widely varying ages, thus explaining the presence of minor outliers in the dating sequences. Because the individual age estimates presented in this study all represent an average of multiple layers of varying ages, the true minimum age of the underlying artwork is possibly older than that reported.
Statistical analysis
The 230Th/238U and 234U/238U activity ratios of the samples were calculated using the decay constants given in (42). U-Th ages were calculated using the Isoplot/Ex 3.75 Program (43). Here, minimum ages are quoted as measured ages (unrounded) minus 2 s and maximum ages as measured ages (unrounded) plus 2 s rounded to one decimal place.
Acknowledgments
For authorizing the research, we thank Indonesia’s State Ministry of Research and Technology (RISTEK), as well as the Director of ARKENAS, I. Made Geria; Head of Balai Arkeologi Sulawesi Selatan, I. Mahmud; and Head of Balai Pelestarian Cagar Budaya Makassar, L. Aksa. Field assistance was provided by S. S. Budi, Sirajuddin, Darfin, F. A. Syahdar, A. M. Saiful, Suryatman, Hamrullah, and Asri. W. Zhou, F. Liu, and Y. Feng helped with the U-series dating. We acknowledge input and advice from the following authorities on S. celebensis and other wild suids: C. Groves, A. A. Macdonald, A. Haris, J. Burton, E. Meijaard, G. Larson, P. Piper, and G. D. van den Bergh. M. Zeder, P. Veth, B. David, P. S. C. Taçon, W. Feeney, M. W. Moore, I. Davidson, and I. Lilley are thanked for their comments on earlier drafts of the paper. A. Duli, I. Sumantri, S. O’Connor, J. Fox, H. Brooks, M. Kottermair, and A. Jalandoni are acknowledged for material contributions or discussions that improved the paper. Funding: The research was funded by the Australian Research Council fellowships awarded to A.B. (FT160100119) and M.A. (FT170100025), along with financial support from Griffith University. Author contributions: A.B. conceived and led the project with M.A. and scientific counterparts, B.H., P.H.S., and M.R. (ARKENAS), as well as Balai Pelestarian Cagar Budaya (BPCB) Makassar cultural heritage partner, R.L. As part of A.B.’s collaboration with ARKENAS, B.B. led the survey team that discovered Leang Tedongnge and conducted the initial documentation. Also under the auspices of this project, A.A.O. led the survey team that found the rock art at Leang Balangajia 1. Parietal imagery at these sites was recorded and analyzed in the field by A.A.O., who also produced the digital tracings of all reported motifs. M.A. identified and collected the coralloid speleothem samples and conducted the subsampling procedure at Griffith University. S.A. and I.S. provided technical and logistical contributions to the research. All in-field sampling involving rock art was carried out under the direct supervision of R.L. J.-x.Z. conducted the U-series dating at the University of Queensland. M.A. and J.-x.Z. analyzed and interpreted the U-series data and discussed and approved correction factors and other methodological details. A.B. and M.A. wrote the paper with contributions from various authors. M.A. and J.-x.Z. prepared the methods. A.B. produced and/or designed the figures. All authors reviewed and edited the paper. Competing interests: The authors declare that they have no competing interests. Data and materials availability: All data needed to evaluate the conclusions in the paper are present in the paper. Data from the descriptive analysis and dating of the rock art can be provided by the ARKENAS/Griffith University pending scientific review and, where applicable, a completed material transfer agreement. Requests for the data should be submitted to the corresponding authors. Additional data related to this paper may be requested from the authors.
REFERENCES AND NOTES
- 1.van den Bergh G. D., Li B., Brumm A., Grün R., Yurnaldi D., Moore M. W., Kurniawan I., Setiawan R., Aziz F., Roberts R. G., Suyono M. S., Setiabudi E., Morwood M. J., Earliest hominin occupation of Sulawesi, Indonesia. Nature 529, 208–211 (2016). [DOI] [PubMed] [Google Scholar]
- 2.Westaway K. E., Louys J., Due Awe R., Morwood M. J., Price G. J., Zhao J.-x., Aubert M., Joannes-Boyau R., Smith T. M., Skinner M. M., Compton T., Bailey R. M., van den Bergh G. D., de Vos J., Pike A. W. G., Stringer C., Saptomo E. W., Rizal Y., Zaim J., Santoso W. D., Trihascaryo A., Kinsley L., Sulistyanto B., An early modern human presence in Sumatra 73,000–63,000 years ago. Nature 548, 322–325 (2017). [DOI] [PubMed] [Google Scholar]
- 3.Clarkson C., Jacobs Z., Marwick B., Fullagar R., Wallis L., Smith M., Roberts R. G., Hayes E., Lowe K., Carah X., Florin S. A., McNeil J., Cox D., Arnold L. J., Hua Q., Huntley J., Brand H. E. A., Manne T., Fairbairn A., Shulmeister J., Lyle L., Salinas M., Page M., Connell K., Park G., Norman K., Murphy T., Pardoe C., Human occupation of northern Australia by 65,000 years ago. Nature 547, 306–310 (2017). [DOI] [PubMed] [Google Scholar]
- 4.O’Connell J. F., Allen J., Williams M. A. J., Williams A. N., Turney C. S. M., Spooner N. A., Kamminga J., Brown G., Cooper A., When did Homo sapiens first reach Southeast Asia and Sahul? Proc. Natl. Acad. Sci. U.S.A. 115( 34), 8482–8490 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kealy S., Louys J., O’Connor S., Least-cost pathway models indicate northern human dispersal from Sunda to Sahul. J. Hum. Evol. 125, 59–70 (2018). [DOI] [PubMed] [Google Scholar]
- 6.Aubert M., Brumm A., Ramli M., Sutikna T., Saptomo E. W., Hakim B., Morwood M. J., van den Bergh G. D., Kinsley L., Dosseto A., Pleistocene cave art from Sulawesi, Indonesia, Indonesia. Nature 514, 223–227 (2014). [DOI] [PubMed] [Google Scholar]
- 7.Aubert M., Lebe R., Oktaviana A. A., Tang M., Burhan B., Hamrullah, Jusdi A., Abdullah, Hakim B., Zhao J.-x., Geria I. M., Sulistyarto P. H., Sardi R., Brumm A., Earliest hunting scene in prehistoric art. Nature 576, 442–445 (2019). [DOI] [PubMed] [Google Scholar]
- 8.van Heekeren H. R., Rock-paintings and other prehistoric discoveries near Maros (South West Celebes). Laporan Tahunan Dinas Purbakala 1950, 22–35 (1952). [Google Scholar]
- 9.P. Bellwood, First Islanders (Wiley, 2017). [Google Scholar]
- 10.Y. Eriawati, Lukisan di Gua-Gua Karst Maros–Pangkep, Sulawesi Selatan: Gambaran Penghuni dan Matapencahariannya (Indonesian Ministry of Cultural Media Development, 2003).
- 11.Burton J. A., Hedges S., Mustari A. H., The taxonomic status, distribution and conservation of the lowland anoa Bubalus depressicornis and mountain anoa Bubalus quarlesi. Mammal Rev. 35, 25–50 (2005). [Google Scholar]
- 12.J. A. Burton, A. H. Mustari, I. S. Rejeki, Sulawesi warty pig Sus celebensis (Muller and Schlegel, 1843), in Ecology, Conservation and Management of Wild Pigs and Peccaries (Cambridge Univ. Press, 2018), pp. 184–192. [Google Scholar]
- 13.C. Groves, Ancestors for the Pigs: Taxonomy and Phylogeny of the Genus Sus (Australian National University, 1981). [Google Scholar]
- 14.C. P. Groves, P. Grubb, The Eurasian suids Sus and Babyrousa, in Pigs, Peccaries, and Hippos (International Union for Conservation of Nature and Natural Resources, 1993), pp. 107–111. [Google Scholar]
- 15.A. A. Macdonald, The Sulawesi warty pig (Sus celebensis), in Pigs, Peccaries, and Hippos (International Union for Conservation of Nature and Natural Resources, 1993), pp. 155–160.
- 16.Brumm A., Langley M. C., Moore M. W., Hakim B., Ramli M., Sumantri I., Burhan B., Saiful A. M., Siagian L., Suryatman, Sardi R., Jusdi A., Abdullah, Mubarak A. P., Hasliana, Hasrianti, Oktaviana A. A., Adhityatama S., van den Bergh G. D., Aubert M., Zhao J.-x., Huntley J., Li B., Roberts R. G., Saptomo E. W., Perston Y., Grün R., Early human symbolic behavior in the Late Pleistocene of Wallacea. Proc. Natl. Acad. Sci. U.S.A. 114, 4105–4110 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Brumm A., Hakim B., Ramli M., Aubert M., van den Bergh G. D., Li B., Burhan B., Saiful A. M., Siagian L., Sardi R., Jusdi A., Abdullah, Mubarak A. P., Moore M. W., Roberts R. G., Zhao J.-X., Gahan D. M., Jones B. G., Perston Y., Szabó K., Mahmud M. I., Westaway K., Jatmiko, Saptomo E. W., van der Kaars S., Grün R., Wood R., Dodson J., Morwood M. J., A reassessment of the early archaeological record at Leang Burung 2, a Late Pleistocene rock-shelter site on the Indonesian island of Sulawesi. PLOS ONE 13, e0193025 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Langley M. C., Hakim B., Oktaviana A. A., Burhan B., Sumantri I., Sulistyarto P. H., Lebe R., McGahan D., Brumm A., Portable art from Pleistocene Sulawesi. Nat. Hum. Behav. 4, 597–602 (2020). [DOI] [PubMed] [Google Scholar]
- 19.Aubert M., Setiawan P., Oktaviana A. A., Brumm A., Sulistyarto P. H., Saptomo E. W., Istiawan B., Ma’rifat T. A., Wahyuono V. N., Atmoko F. T., Zhao J.-x., Huntley J., Taçon P. S. C., Howard D. L., Brand H. E. A., Palaeolithic cave art in Borneo. Nature 564, 254–257 (2018). [DOI] [PubMed] [Google Scholar]
- 20.Azéma M., Rivère F., Animation in Palaeolithic art: A pre-echo of cinema. Antiquity 86, 316–324 (2012). [Google Scholar]
- 21.Dobrez L., Dobrez P., Rock art animals in profile: Visual recognition and the principles of canonical form. Rock Art Res. 30, 75–90 (2013). [Google Scholar]
- 22.Groves C. P., Notes on the systematics of babyrousa (artiodactyla, suidae). Zoologische Mededelingen 55, 29–46 (1980). [Google Scholar]
- 23.Hoffmann D. L., Standish C. D., García-Diez M., Pettitt P. B., Milton J. A., Zilhão J., Alcolea-González J. J., Cantalejo-Duarte P., Collado H., de Balbín R., Lorblanchet M., Ramos-Muñoz J., Weniger G.-C., Pike A. W. G., U–Th dating of carbonate crusts reveals Neandertal origin of Iberian cave art. Science 359, 912–915 (2018). [DOI] [PubMed] [Google Scholar]
- 24.Aubert M., Brumm A., Huntley J., Early dates for “Neanderthal cave art” may be wrong. J. Hum. Evol. 125, 215–217 (2018). [DOI] [PubMed] [Google Scholar]
- 25.Pearce D. G., Bonneau A., Trouble on the dating scene. Nat. Ecol. Evol. 2, 925–926 (2018). [DOI] [PubMed] [Google Scholar]
- 26.Pons-Branchu E., Sanchidrián J. L., Fontugne M., Medina-Alcaide M. Á., Quiles A., Thil F., Valladas H., U-series dating at Nerja cave reveal open system. Questioning the Neanderthal origin of Spanish rock art. J. Archaeol. Sci. 117, 105120 (2020). [Google Scholar]
- 27.White R., Bosinski G., Bourrillon R., Clottes J., Conkey M. W., Rodriguez S. C., Cortés-Sánchez M., de la Rasilla Vives M., Delluc B., Delluc G., Floss V. F. H., Foucher P., Fritz C., Fuentes O., Garate D., Gómez J. G., González-Morales M. R., González-Pumariega Solis M., Groenen M., Jaubert J., Martinez-Aguirre M. A., Alcaide M.-Á. M., Abadia O. M., Peredo R. O., Paillet-Man-Estier E., Pailleta P., Petrognani S., Pigeaud R., Pinçon G., Plassard F., López S. R., Vilá O. R., Robert E., Ruiz-Redondo A., López J. F. R., Juan-Foucher C. S., Torti J. L. S., Sauvet G., Simón-Vallejo M. D., Tosello G., Utrilla P., Vialou D., Willis M. D., Still no archaeological evidence that Neanderthals created Iberian cave art. J. Hum. Evol. 144, 102640 (2020). [DOI] [PubMed] [Google Scholar]
- 28.Hublin J.-J., Ben-Ncer A., Bailey S. E., Freidline S. E., Neubauer S., Skinner M. M., Bergmann I., Le Cabec A., Benazzi S., Harvati K., Gunz P., New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens, Morocco and the pan-African origin of Homo sapiens. Nature 546, 289–292 (2017). [DOI] [PubMed] [Google Scholar]
- 29.Liu W., Martinón-Torres M., Cai Y.-j., Xing S., Tong H.-w., Pei S.-w., Sier M. J., Wu X.-h., Edwards R. L., Cheng H., Li Y.-y., Yang X.-x., Bermúdez de Castro J. M., Wu X.-j., The earliest unequivocally modern humans in southern China. Nature 526, 696–699 (2015). [DOI] [PubMed] [Google Scholar]
- 30.Pagani L., Lawson D. J., Jagoda E., Mörseburg A., Eriksson A., Mitt M., Clemente F., Hudjashov G., DeGiorgio M., Saag L., Wal J. D., Mägi A. C. R., Sayres M. A. W., Kaewert S., Inchley C., Scheib C. L., Järve M., Karmin M., Jacobs G. S., Antao T., Iliescu F. M., Kushniarevic A., Ayub Q., Tyler-Smith C., Xue Y., Yunusbayev B., Tambets K., Mallick C. B., Saag L., Pocheshkhova E., Andriadze G., Muller C., Westaway M. C., Lambert D. M., Zoraqi G., Turdikulova S., Dalimova D., Sabitov Z., Sultana G. N. N., Lachance J., Tishkoff S., Momynaliev K., Isakova J., Damba L. D., Gubina M., Nymadawa P., Evseeva I., Atramentova L., Utevska O., Ricaut F.-X., Brucato N., Sudoyo H., Letellier T., Cox M. P., Barashkov N. A., Škaro V., Mulahasanovic L., Primorac D., Sahakyan H., Mormina M., Eichstaedt C. A., Lichman D. V., Abdullah S., Chaubey G., Wee J. T. S., Mihailov E., Karunas A., Litvinov S., Khusainova R., Ekomasova N., Akhmetova V., Khidiyatova I., Marjanović D., Yepiskoposyan L., Behar D. M., Balanovska E., Metspalu A., Derenko M., Malyarchuk B., Voevod M., Fedorova S. A., Osipova L. P., Lahr M. M., Gerbault P., Leavesley M., Migliano A. B., Petraglia M., Balanovsky O., Khusnutdinova E. K., Metspalu E., Thomas M. G., Manica A., Nielsen R., Villems R., Willerslev E., Kivisild T., Metspalu M., Genomic analyses inform on migration events during the peopling of Eurasia. Nature 538, 238–242 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hawkins S., O’Connor S., Maloney T. R., Litster M., Kealy S., Fenner J. N., Aplin K., Boulanger C., Brockwell S., Willan R., Piotto E., Louys J., Oldest human occupation of Wallacea at Laili Cave, Timor-Leste, shows broad-spectrum foraging responses to late Pleistocene environments. Quat. Sci. Rev. 171, 58–72 (2017). [Google Scholar]
- 32.O’Connor S., Louys J., Kealy S., Samper Carro S. C., Hominin dispersal and settlement east of Huxley’s Line: The role of sea level changes, island size, and subsistence behavior. Curr. Anthropol. 58 ( Suppl. 17), (2017). [Google Scholar]
- 33.B. Bourdon, G. M. Henderson, C. C. Lundstrom, S. P. Turner, Uranium-series Geochemistry (Mineralogical Society of America, 2003).
- 34.Zhao J.-x., Yu K. F., Feng Y. X., High-precision 238U–234U–230Th disequilibrium dating of the recent past - a review. Quat. Geochronol. 4, 423–433 (2009). [Google Scholar]
- 35.Clark T. R., Zhao J.-x., Feng Y.-x., Done T. J., Jupiter S., Lough J., Pandolfi J. M., Spatial variability of initial 230Th/232Th in modern Porites from the inshore region of the Great Barrier Reef. Geochim. Cosmochim. Acta 78, 99–118 (2012). [Google Scholar]
- 36.Clark T. R., Zhao J.-x., Roff G., Feng Y.-x., Done T. J., Nothdurft L. D., Pandolfi J. M., Discerning the timing and cause of historical mortality events in modern Porites from the Great Barrier Reef. Geochim. Cosmochim. Acta 138, 57–80 (2014). [Google Scholar]
- 37.Zhou H. Y., Zhao J.-x., Wang Q., Feng Y. X., Tang J., Speleothem-derived Asian summer monsoon variations in Central China, 54-46 ka. J. Quat. Sci. 26, 781–790 (2011). [Google Scholar]
- 38.St Pierre E., Zhao J.-x., Reed E., Expanding the utility of uranium-series dating of speleothems for archaeological and palaeontological applications. J. Archaeol. Sci. 36, 1416–1423 (2009). [Google Scholar]
- 39.Hellstrom J., U–Th dating of speleothems with high initial 230Th using stratigraphical constraint. Quat. Geochronol. 1, 289–295 (2006). [Google Scholar]
- 40.Plagnes V., Causse C., Fontugne M., Valladas H., Chazine J.-M., Fage L.-H., Cross dating (Th/U-14C) of calcite covering prehistoric paintings in Borneo. Quatern. Res. 60, 172–179 (2003). [Google Scholar]
- 41.Vanghi V., Frisia S., Borsato A., Genesis and microstratigraphy of calcite coralloids analysed by high resolution imaging and petrography. Sedim. Geol. 359, 16–28 (2017). [Google Scholar]
- 42.Cheng H., Edwards R. L., Hoff J., Richards D. A., Asmerom Y., The half-lives of uranium-234 and thorium-230. Chem. Geol. 169, 17–33 (2000). [Google Scholar]
- 43.K. R. Ludwig, User’s Manual for Isoplot 3.75. A Geochronological Toolkit for Microsoft Excel (Berkeley Geochronology Center, 2012).
- 44.J. Harman, Using decorrelation stretch to enhance rock art images (2005); www.dstretch.com/AlgorithmDescription.pdf.