

Abstract
Key message
The Bavarian MAGIC Wheat population, comprising 394 F6:8 recombinant inbred lines was phenotyped for Puccinia triticina resistance in multi-years’ field trials at three locations and in a controlled environment seedling test. Simple intervall mapping revealed 19 QTL, corresponding to 11 distinct chromosomal regions.
Abstract
The biotrophic rust fungus Puccinia triticina is one of the most important wheat pathogens with the potential to cause yield losses up to 70%. Growing resistant cultivars is the most cost-effective and environmentally friendly way to encounter this problem. The emergence of leaf rust races being virulent against common resistance genes increases the demand for wheat varieties with novel resistances. In the past decade, the use of complex experimental populations, like multiparent advanced generation intercross (MAGIC) populations, has risen and offers great advantages for mapping resistances. The genetic diversity of multiple parents, which has been recombined over several generations, leads to a broad phenotypic diversity, suitable for high-resolution mapping of quantitative traits. In this study, interval mapping was performed to map quantitative trait loci (QTL) for leaf rust resistance in the Bavarian MAGIC Wheat population, comprising 394 F6:8 recombinant inbred lines (RILs). Phenotypic evaluation of the RILs for adult plant resistance was carried out in field trials at three locations and two years, as well as in a controlled-environment seedling inoculation test. In total, interval mapping revealed 19 QTL, which corresponded to 11 distinct chromosomal regions controlling leaf rust resistance. Six of these regions may represent putative new QTL. Due to the elite parental material, RILs identified to be resistant to leaf rust can be easily introduced in breeding programs.
Electronic supplementary material
The online version of this article (10.1007/s00122-020-03657-2) contains supplementary material, which is available to authorized users.
Introduction
With approximately 219 million hectares worldwide and 30% of global major cereal crop production in 2017, wheat (Triticum spp.) belongs to the most important crops for human nutrition (Braun et al. 2010; FAO 2019). Leaf rust, caused by the obligate biotrophic fungus Puccinia triticina Eriks., is nowadays the most destructive and prevalent rust pathogen in wheat (Kolmer 2005). Due to its adaptation to a wide range of different environments, leaf rust occurs in many wheat-producing areas of the temperate zone, causing yield losses up to 70% (Aktar-Uz-Zaman et al. 2017; Herrera-Foessel et al. 2006; Marasas et al. 2004). Although the application of fungicides helps to avoid yield losses, the deployment of resistant cultivars is the most effective, economic, and environmentally friendly approach to manage this disease. For wheat leaf rust, both qualitative and quantitative resistances are known. Seedling/qualitative resistance is monogenically inherited and only effective against a subset of races. Thus, it mainly follows the gene-for-gene concept, in which resistance depends on a specific genetic interaction between host-resistance genes and avirulence genes of the pathogen (Flor 1956, 1971). These major genes confer vertical resistance and tend to be expressed from seedling to adult plant stages. Genotypes carrying such resistances show a hypersensitive response or programmed cell death (Bolton et al. 2008). In contrast, quantitative resistance is based on minor genes encoding various resistance responses, which are not restricted to specific pathogen races. Quantitative resistances are effective at later growth stages and are therefore referred to as field resistance or adult plant resistance (APR, Krattinger and Keller 2016). To date, more than 80 resistance genes to leaf rust (Lr genes) have been identified in bread wheat, durum wheat, and diploid wheat species (Gill et al. 2019). While most of them show race-specific resistance at the seedling stage, genes like Lr12, Lr13, Lr22a/b, Lr34, Lr35, Lr37, Lr46, Lr67, Lr68, and Lr77 confer resistance at the adult plant stage (Dakouri et al. 2013; McIntosh et al. 2013, 2017).
The identification of such resistance genes as well as of quantitative trait loci (QTL) has been mainly based on biparental crosses (Huang et al. 2012). The weakness of such populations is the narrow genetic variation and the fact that genetic recombination is limited, which leads to a lower map resolution (Bandillo et al. 2013). Nowadays, high-throughput marker systems are available and genetic marker information is no longer limiting (Bayer et al. 2017; Chen et al. 2014; Cui et al. 2017; He et al. 2014; Mammadov et al. 2012), but the genetic variation present in respective populations (Asimit and Zeggini 2010; Gibson 2012). Thus, complex experimental populations such as nested association mapping (NAM, Yu et al. 2008) and multiparent advanced generation intercross (MAGIC) populations have been developed to detect QTL with a better reliability (Cavanagh et al. 2008). First multiparental intermated populations were exploited in mice (Churchill et al. 2004) and Drosophila melanogaster (King et al. 2012). In plants, MAGIC populations were first developed and described in studies regarding Arabidopsis thaliana (Cavanagh et al. 2008; Kover et al. 2009). These experimental designs involved multiple intercrosses of inbred founders for several generations to combine the genetic variation of all parental lines in the resulting progeny (Huang et al. 2012). MAGIC populations have been widely used to conduct QTL mapping in several crop species, such as rice (Bandillo et al. 2013), maize (Dell’Acqua et al. 2015), tomato (Pascual et al. 2015), faba bean (Sallam and Martsch 2015), sorghum (Ongom and Ejeta 2018), barley (Sannemann et al. 2015), and wheat (Gardner et al. 2016; Huang et al. 2012; Mackay et al. 2014; Milner et al. 2016; Sannemann et al. 2018). There are two clear advantages of using multiparental populations. First, based on the choice of founders, more traits of interest from each founder can be analyzed. Second, due to the higher genetic variability and recombination rate, QTL detection can be performed with increased precision and resolution (Bandillo et al. 2013; Cavanagh et al. 2008).
The Bavarian MAGIC Wheat population (BMWpop) is one of only two German MAGIC wheat populations, which are mainly based on adapted German elite cultivars. It captures 71.7% of the allelic diversity available in the German wheat breeding gene pool (Stadlmeier et al. 2018). These populations provide the potential to carry out genetic studies of important economical traits, such as plant height and resistance to powdery mildew (Sannemann et al. 2018; Stadlmeier et al. 2018). In addition, Stadlmeier et al. (2019) detected six, seven and nine QTL for resistance to important fungal pathogens, i.e., Blumeria graminis, Zymoseptoria tritici, and Pyrenophora tritici-repentis, respectively. The objectives of the current study were to (1) phenotype the BMW population for quantitative and qualitative leaf rust resistance in multi-environment field trials and an extensive seedling test and (2) genetically map QTL in order to develop closely linked molecular markers suitable for marker-assisted selection (MAS).
Material and methods
Plant material
The study is based on the multiparental BMW population comprising elite wheat cultivars (Stadlmeier et al. 2018). It consists of 394 diverse F6:8 recombinant inbred lines (RILs), which were derived from a simplified eight founder MAGIC mating design with additional eight-way intercrosses. The founders ‘Event’, ‘BAYB4535′, ‘Potenzial’, ‘Bussard’, ‘Firl3565’, ‘Format’, ‘Julius’ and ‘Ambition’ originated from German and Danish wheat breeders and were selected on the criteria of (1) variation for agronomic, quality and disease resistance traits, (2) originating from different breeding programs, and (3) being important cultivars in the respective baking quality group. More detailed information about the development and the genetics of the BMW population is provided by Stadlmeier et al. (2018).
Phenotypic assessment of leaf rust resistance in field
Five field trials were performed, each using a randomized incomplete block design with two replications at three locations in Germany: Quedlinburg (QLB, 51° 46′ 21.45″ N 11° 8′ 34.8″ E) in Saxony-Anhalt, Soellingen (SOE, 52° 5′ 45.506″ N 10° 55′ 41.711″ E) and Lenglern (LEN, 51° 35′ 47.53″ N 9° 51′ 39.118″ E) in Lower Saxony. The 394 RILs, the eight founders, and the susceptible standard ‘Schamane’ were evaluated in double rows under natural disease epidemics in SOE (2017 and 2018) and LEN (2018). In QLB entries were sown 2016/2017 and 2017/2018 in double rows of 1 m length with 30 plants per row and spacing of 0.2 m between rows. Additional infection stripes of susceptible varieties were arranged in regular intervals of every third plot. Growth regulator Medax® Top (BASF Agricultural Solutions, Germany, 1 L ha−1) was applied twice (BBCH31, BBCH37) to reduce plant height and lodging. No selective fungicides were used. To ensure uniform infestation, the infection stripes were artificially inoculated at the beginning of flowering using the highly virulent Puccinia triticina isolate 77WxR (Tab. S1). For this, a spore suspension of 10 mg uredospores in 100 ml Isopar M (ExxonMobil Chemical Company, USA) was applied in a total amount of 10 ml suspension per m2, using a hand-held spinning disc sprayer (Bromyard, U.K.). Phenotyping of the trials was carried out by scoring the average percentage of infected leaf area of the second and third youngest leaves in the two rows at two (SOE17, SOE18, LEN18), three (QLB18), and four (QLB17) subsequent dates according to Moll et al. (2010), starting at the time of clearly visible disease symptoms on the infection stripe or the susceptible standard, respectively. A time period of 1 to 2 weeks was chosen between the scorings.
Phenotypic assessment of leaf rust resistance in seedlings
All RILs, the parental lines, and the susceptible standard ‘Borenos’ were evaluated for resistance at seedling stage in a detached leaf assay (Douchkov et al. 2012). Seedlings were grown in 77-cell trays with mixed potting soil (Gebr. Patzer GmbH Co KG, Sinntal, Germany) using a randomized complete block design with five replications. Water agar (7 g L−1) containing 45 mg L−1 benzimidazole (Sigma-Aldrich Chemie GmbH, Taufkirchen, Germany), used to delay senescence of leaf segments, was dispensed in 4 × 10 mL aliquots into nonsterile four-well polystyrene plates (8 × 12 x 1 cm, Greiner Bio-One GmbH, Frickenhausen, Germany). Ten days after sowing, when the second leaf was developed, 2.5-cm sections were cut from the middle of the primary leaves and placed into the plates, keeping the randomization. White polytetrafluoroethylene frames (eMachineShop, Mahwah, USA) were used to fix the leaves. Inoculation was performed by an infection tower with three seconds swirling duration and three minutes of settling time (Melching 1967). Due to space restrictions, plates were divided into two infection groups per replication. Each group was inoculated with leaf rust isolate 77WxR using a mixture of 30 mg uredospores and white clay (1:1 w/w, VWR International GmbH, Darmstadt, Germany) after application of a 0.01% Tween 20 (Sigma-Aldrich) solution to support adhesion. For 24 h, the plates were covered by wet cotton paper to support spore germination in the dark and at high humidity. Inoculated leaf segments were subsequently incubated in greenhouse at night/day temperatures of 16 °C/18 °C with additional lighting (16 h/8 h day/night) for ten days. Quantitative scoring was conducted using a high-throughput phenotyping platform (Douchkov et al. 2012). Digital images with a resolution of 20 Megapixel and four wavelengths between 315 and 750 nm (UV, blue, green, and red) were taken automatically from every plate. Subsequently, the leaf area was calculated and compared to the area of uredospore pustules for analyzing the percentage of infected leaf area (Pi) using the software HawkSpex® (Fraunhofer IFF, Magdeburg, Germany). Additionally, all entries were visually evaluated for infection type (IT) using a 0−4 scale (McIntosh et al. 1995). To generate metric data, original IT data were converted to a 0 – 10 linear disease scale, modified according to Zhang et al. (2014) as follows: 0, 0;, − 1,1, + 1, − 2, 2, 2 + , − 3, 3, + 3 were coded as 0, 0.5, 1, 2, 3, 4, 5, 6, 7, 8 and 9, respectively. IT − 4 and 4 were coded as 10 and in case of special annotation code “C” for chlorosis, 0.5 was added to the linear scale.
Data analysis
The multiple scorings of the percentage of infected leaf area in field trials were taken to calculate the area under the disease progress curve (AUDPC) and the average ordinate (AO, Moll et al. 1996) for each RIL using the following equations:
where yi is the disease level at the ith observation, ti is the time at the ith observation, N is the total number of observations and T is the total observation time from the first to the last scoring date in days. Out of percentage of infected leaf area, AUDPC, and AO, only AO values were used for subsequent statistical analysis. Different year–location combinations of all trials were referred to as “environment”.
Analyses of all phenotypic data were carried out using proc mixed of the software package SAS 9.4 (SAS Institute Inc., NY, USA). In order to apply a mixed linear model, a log10 data transformation of the AO, IT, and Pi values was performed. The factors genotype, environment, and the genotype × environment interaction of field data were set as fix effects, while the design effects of replication and block were set as random. To obtain variance components for calculation of the broad sense heritability, all model parameters were set as random. Heritability was estimated on a progeny mean basis according to Hallauer et al. (2010).
For analyzing IT and Pi scores from seedling test the model:
was used, where yijk is the trait observation, µ is the overall mean, gi is the fixed effect of the genotype, rj is the fixed effect of the replication, lk is the random effect of the infection group nested in the replication and eijk is the random residual error. Variance components were obtained by fitting the genotype as random to calculate the repeatability as the ratio of the genotypic variance and the sum of the genotypic and the residual error variance divided by the number of replications. For each trait, least-square means (lsmeans) were calculated and used for subsequent QTL analysis.
QTL mapping
The BMW population and the parental lines were genotyped using the 15 K + 5 K Infinium® iSelect® array containing 17,267 single nucleotide polymorphism (SNP) markers (TraitGenetics, Germany). The preparation of genotypic data and the construction of the linkage map used for QTL mapping were described in detail by Stadlmeier et al. (2018). QTL mapping was performed using the R (× 32 3.2.5) package mpMap V2.0.2 (Huang and George 2011; R Core Team 2017). To conduct simple interval mapping (SIM), founder probabilities were calculated using the function ‘mpprob’. These give information about the probability of each locus that the observed genotype was inherited from one of the eight founders and are based on multipoint haplotype probabilities (Broman et al. 2003). To determine the parental origin of an allele, the threshold was set to 0.7. For SIM, a genome-wide significant threshold of α < 0.05 was calculated for each trait. The thresholds were obtained from permutation of phenotypic data with 1000 simulation runs (Churchill and Doerge 1994). QTL detection was performed using the function ‘mpIM’, implemented in the mpMap package (Huang and George 2011). Phenotypic variance explained by individual QTL and additive QTL effects were estimated separately using the categorical allele information of the founders. QTL support intervals were determined using the function ‘supportinterval’ of the mpMap package. A QTL support interval was defined as the map interval surrounding a QTL peak at a − log10(p) drop of one unit (Huang and George 2011).
In order to compare QTL identified in the present study with previously described QTL, overlapping QTL based on the support interval was merged together. Databases of the Triticeae Toolbox (https://triticeaetoolbox.org/wheat/genotyping/marker_selection.php), GrainGenes (https://wheat.pw.usda.gov/GG3/), as well as CerealsDB (https://www.cerealsdb.uk.net/cerealgenomics/CerealsDB/axiom_download.php) were used to obtain marker information. Physical positions were received by nucleotide BLAST (BLAST-n) of the marker sequences against the reference sequence RefSeq v1.0 using the database of 10 + Genome Project (https://webblast.ipk-gatersleben.de/wheat_ten_genomes/). BLAST hits were considered significant if the percent identity was greater than 95%, and only the best hit was taken if multiple BLAST hits were detected (Gao et al. 2016). The start and end positions of peak marker sequences preceded by the chromosome name were taken to the URGI database to obtain functional gene annotations available from IWGSC (https://wheat-urgi.versailles.inra.fr/Seq-Repository/Annotations). Sequences of the closest related species, Triticum urartu (A-genome donor) and Aegilops tauschii (D-genome donor), were considered for the detection of orthologous genes.
Results
Phenotypic assessment
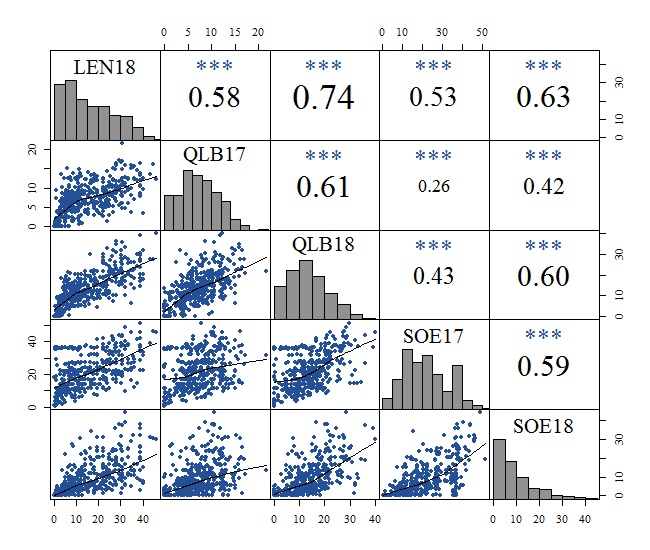
Leaf rust severity of field trials clearly varied between years and location, displaying in QLB 2017, SOE 2018, and LEN 2018 the lowest infestations of leaf rust (Fig. S1). Pearson correlation coefficient between the different environments ranged from 0.26 to 0.74 (P < 0.001). Nevertheless, after mixed model adjustment, a broad sense heritability (h2) of 0.83 was estimated (Table 1). The mean phenotypic distribution of AOs was slightly right-skewed and indicated a broad variability within the population (Fig. 1a), ranging between 0.2 and 34.8% (mean 13.5%) leaf area diseased. However, single maximal AO scores up to 63.8% were observed within the population (Table 1). The average performance of parental lines was evenly distributed, resulting in a nonsignificant difference (p < 0.05) from the progeny mean. Founders ‘BAYP4535′ and ‘Bussard’ were identified as the most resistant (4.5%) and most susceptible (22.9%) parental line to leaf rust, respectively. The analysis of variance showed significant differences concerning genotype, environment, and the interaction between genotype and environment (Table 2).
Table 1.
Descriptive statistics and heritability / repeatability for field trials (AO) and seedling test (IT and Pi)
| Traita | Mean founders | Mean population | Minb | Maxc | SE±d | CVe | h2/rep |
|---|---|---|---|---|---|---|---|
| AO [%] | 13.70 | 13.50 | 0 | 63.75 | 0.17 | 0.83 | 0.83f |
| IT [1–10] | 3.32 | 3.84 | 0 | 10.00 | 0.06 | 0.96 | 0.93g |
| Pi [%] | 8.06 | 8.47 | 0 | 57.73 | 0.18 | 0.72 | 0.91g |
a Average ordinate (AO), infection type (IT), infected leaf area (Pi)
b Minimum
c Maximum
d Standard error
e Coefficient of variance
f Broad-sense heritability (h2)
g Repeatability (rep)
Fig. 1.

Averaged phenotypic distribution of resistance to Puccinia triticina for field trials (A) and seedling test (B, C). Performance of each parental line is shown as vertical dashed line
Table 2.
Analysis of variance of log10-transformed data for leaf rust severity evaluated in field trials (AO) and seedling test (IT and Pi)
| Traita/factor | DFb | F value | P value |
|---|---|---|---|
| AO | |||
| Genotype | 402 | 18.98 | < 0.0001 |
| Environment | 4 | 16.05 | 0.0049 |
| Genotype × environment | 1605 | 2.39 | < 0.0001 |
| IT | |||
| Genotype | 402 | 17.69 | < 0.0001 |
| Replication | 4 | 0.94 | 0.5196 |
| Pi | |||
| Genotype | 402 | 16.63 | < 0.0001 |
| Replication | 4 | 6.66 | 0.0426 |
Significance level at P < 0.05
a Average ordinate (AO), infection type (IT), infected leaf area (Pi)
b Degrees of freedom
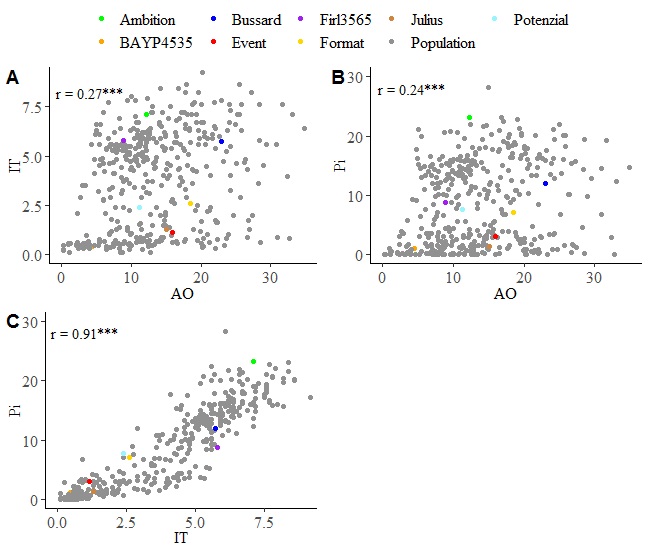
Scoring qualitative resistance in seedling test was performed twice—using an image analysis software to obtain the Pi and visually by assessing the IT (1–10). For both traits, phenotypic data revealed a large variability (Fig. 1b and c). The average IT ranged from 0.1 to 9.2 (mean 3.8). For Pi, the disease severity was on average between 0 and 28.3% (mean 8.5%). Phenotypic distributions of IT and Pi were slightly bimodal, with 131 and 185 RILs showing IT values smaller 2 (few areas with restricted sporulation) and Pi values below 5%, respectively. Maximal scores of 10 (IT) and 57.7% (Pi) were observed (Table 1). The population means of IT and Pi were not significantly different from the means of parental lines. According to the results of field trials, ‘BAYP4535’ and ‘Ambition’ were the most resistant and susceptible founders, respectively, in the seedling inoculation test. Pearson correlation displayed a high correlation coefficient between both traits (r = 0.91; Fig S2 C). The qualitative traits IT and Pi and the quantitative scoring of AO showed weak correlations of r = 0.27 and r = 0.24 (Fig S2 A and B). For both traits, a significant genotype effect was observed, while for Pi also a significance of replication was found. Repeatability of both traits was high with rep(IT) = 0.93 and rep(Pi) = 0.91 (Table 1). From the parental lines, only ‘BAYB4535 showed all stage resistance, whereas cv. ‘Event’, Format’, ‘Julius’, ‘Potenzial’ only showed resistance at seedling and ‘Firl3565’ at adult plant stage, respectively. In total, 68 genotypes in the population expressed all stage resistance, 92 genotypes showed resistance only at seedling stage and 44 genotypes were observed showing APR.
QTL mapping
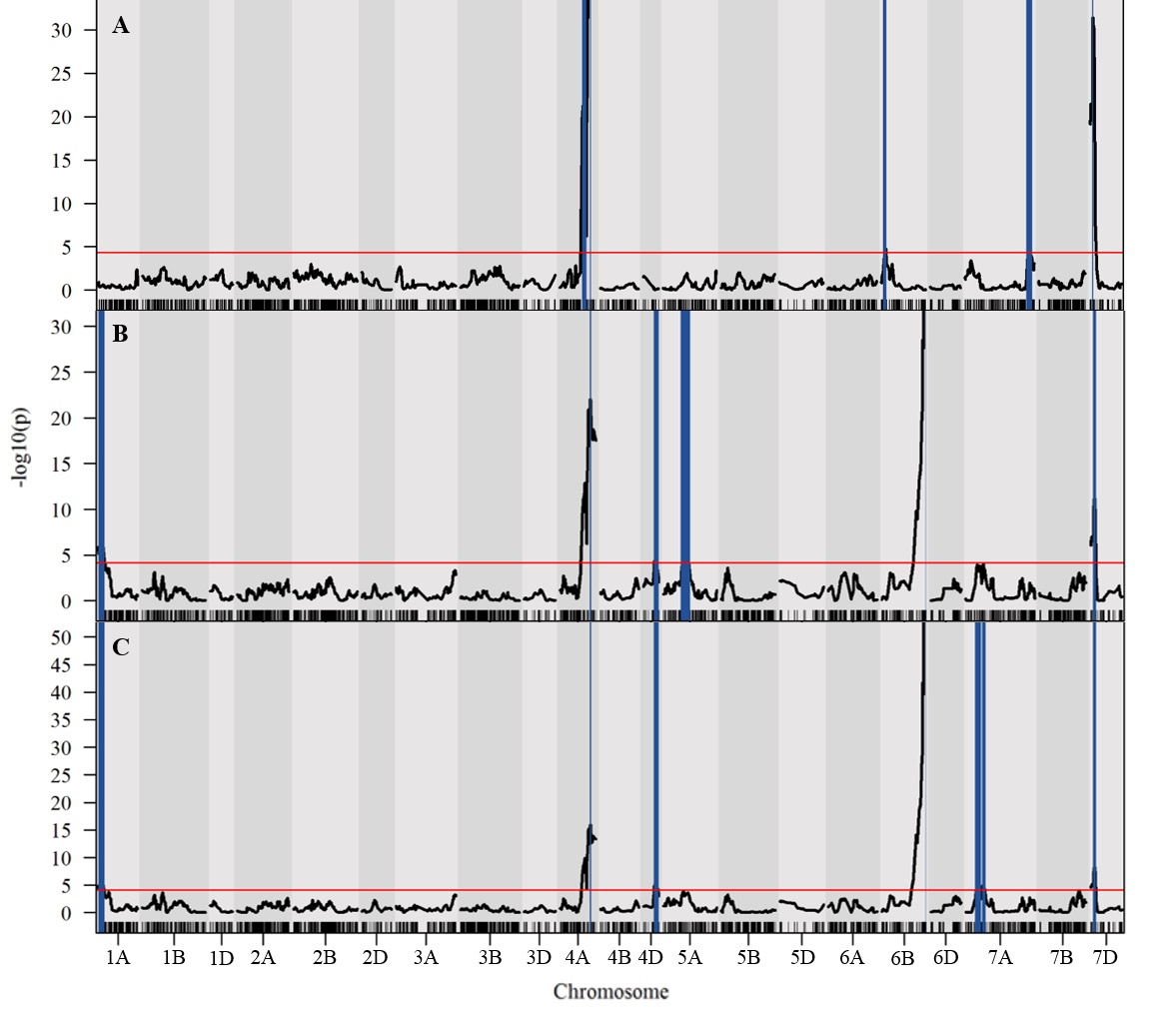
Overall, SIM revealed 19 QTL located on chromosomes 1A, 4A, 4D, 5A, 6B, 7A, and 7D. Hence, five QTL were detected based on field data and seven QTL for seedling resistance, each for IT and Pi values (Table 3, Tab. S2).
Table 3.
QTL for resistance to Puccinia triticina in the BMW population detected in field trials and seedling tests
| Trait | Chr.a | Pos.[cM]b | SI [cM]c | P value | R2d | No. Env.f | Eff (A)g | Eff (B)g | Eff (C)g | Eff (D)g | Eff (E)g | Eff (F)g | Eff G)g | Eff (H)g |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AO | 4A | 133 | 125–151 | 2.00E-22 | 0.31 | 4 | − 0.17 | − 3.12 | + 0.93 | − 1.21 | + 0.90 | + 0.88 | + 0.86 | + 0,83 |
| 4A | 172 | 170–174 | 2.52E-58 | 0.50 | 4 | + 0.94 | − 3.96 | − 0.13 | − 0.44 | + 2.04 | − 0.18 | + 1.94 | − 0,23 | |
| 6B | 22 | 10–30 | 1.49E-05 | 0.08 | 1 | + 0.16 | − 1.23 | + 1.70 | + 1.25 | − 1.32 | − 0.89 | − 1.16 | + 1.47 | |
| 7A | 368 | 346–379 | 1.52E-05 | 0.07 | 1 | − 0.16 | − 1.26 | + 1.10 | − 1.39 | − 1.42 | + 1.22 | + 0.94 | + 0.89 | |
| 7D | 18 | 15–19 | 3.68E-32 | 0.28 | 4 | na | − 3.16 | na | na | + 1.44 | + 0.94 | + 0.18 | + 0.58 | |
| IT | 1A | 28 | 0–34 | 1.55E-06 | 0.11 | na | + 0.76 | − 0.27 | na | − 0.75 | − 0.98 | − 0.67 | + 1.88 | |
| 4A | 170 | 168–174 | 8.79E-23 | 0.28 | 0.00 | − 2.57 | − 1.32 | + 1.59 | + 1.15 | − 1.10 | + 1.58 | + 1.12 | ||
| 4D | 69 | 59–86 | 2.57E-05 | 0.01 | na | + 0.01 | na | na | + 1.16 | na | na | -1.98 | ||
| 5A | 112 | 102–152 | 1.56E-05 | 0.08 | − 0.99 | + 0.21 | + 0.63 | + 0.06 | − 2.06 | + 0.67 | + 0.87 | + 0.60 | ||
| 5A | 139 | 99–152 | 3.31E-05 | 0.05 | − 0.26 | + 0.78 | + 1.29 | − 1.29 | − 1.44 | + 1.12 | − 1.29 | + 1.07 | ||
| 6B | 249 | 248–250 | 2.18E-55 | 0.01 | − 0.5 | na | na | na | na | na | na | + 0.5 | ||
| 7D | 22 | 15–30 | 6.14E-12 | 0.17 | na | − 1.84 | na | na | + 0.61 | + 0.61 | + 0.02 | + 0.61 | ||
| Pi | 1A | 26 | 0–34 | 8.11E-06 | 0.12 | na | + 0.60 | + 1.64 | na | − 1.35 | − 1.59 | − 1.27 | + 1.98 | |
| 4A | 171 | 168–174 | 1.11E-16 | 0.21 | + 0.33 | − 4.14 | − 1.33 | + 1.67 | + 1.47 | − 0.88 | + 1.47 | + 1.42 | ||
| 4D | 72 | 59–86 | 4.27E-06 | 0.09 | na | − 0.06 | na | na | + 1.6 | na | na | − 1.54 | ||
| 6B | 249 | 247–250 | 1.76E-91 | < .01 | − 0.52 | na | na | na | na | na | na | + 0.53 | ||
| 7A | 65 | 54–87 | 7.02E-06 | 0.05 | na | + 1.05 | + 2.15 | − 0.95 | − 0.13 | − 0.73 | − 1.41 | + 0.03 | ||
| 7A | 99 | 94–111 | 6.12E-06 | 0.08 | − 0.97 | + 1.38 | + 0.81 | + 0.08 | + 0.21 | + 0.23 | − 2.38 | + 0.65 | ||
| 7D | 22 | 15–30 | 5.64E-09 | 0.14 | na | − 2.78 | na | na | + 1.11 | + 1.11 | − 0.27 | + 0.85 |
a Chromosomal position of QTL
b Position of peak marker based on Stadlmeier et al. (2018)
c Support interval
d Proportion of phenotypic variance explained by a single QTL
f Number of single environments in which QTL was detected
g Additive effects ( ±) of the founders Event (A), BAYP4535 (B), Ambition (C), Firl3565 (D), Format (E), Potenzial (F), Bussard (G) and Julius (H) relative to the population mean. Shown values are back-transformed to the original trait scale
Founder effects were reported as not available (na) if none of the RILs reached the probability threshold of 0.7
The phenotypic variance (R2) explained by the individual QTL detected in field trials ranged between 8 and 50%, with support intervals (SI) from 4 to 33 cM. The two strongest QTL, explaining 31% and 50% of R2, were located on chromosome 4A with peak markers at 133 cM and 172 cM. The largest allelic effects of these QTL were contributed by ‘BAYP4535’, reducing disease severity by 3.1% and 4.0%, respectively. Another QTL detected on chromosome 7D (at 18 cM) explained 28% of the phenotypic variance with ‘BAYP4535′ as the most resistant founder, reducing infected leaf area by 3.2%. Remaining QTL on chromosomes 6B (at 22 cM) and 7A (at 368 cM) accounted for 8% and 7% of leaf rust variation. For these QTL, cv. ‘Format’ contributed the largest allelic effect reducing infected leaf area by 1.3% and 1.4%, respectively.
For IT, phenotypic variance explained by the seven QTL ranged from 1 to 28% with SIs ranging between 2 and 53 cM (Table 3). QTL on chromosomes 4A and 7D accounted for the highest R2 i.e. 28% and 17% with peak markers at 170 cM and 22 cM. The largest allelic effect of both QTL was contributed by ‘BAYP4535′, reducing disease severity by 2.6 and 1.8 scores, respectively. On chromosome 1A, one QTL was detected at 28 cM, explaining 11% of the phenotypic variance. A maximum effect of -1.0 score was detected for cv. ‘Potenzial’. Furthermore, two QTL were detected on chromosome 5A with 8% (at 112 cM) and 7% (at 139 cM) of the explained variance. SIs of these QTL ranged from 102 to 152 cM and from 99 to 152 cM, respectively. For both, ‘Format’ contributed the highest allelic effect (− 2.1 and − 1.4 scores). QTL located on chromosomes 4D (69 cM) and 6B (249 cM) explained only 1% of the phenotypic variance, each. By analyzing each environment separately, the two QTL on chromosomes 4A were also detected in LEN18, QLB17, QLB18 and SOE18, as well as LEN18, QLB18, SOE17 and SOE18, respectively. The QTL on chromosome 6B and 7D was detected in one (SOE18) and four (LEN18, QLB17, QLB18, SOE18) environments.
SIM of Pi values also revealed seven individual QTL with R2 ranging from less than 1% to 21%. The support intervals varied between 2 and 34 cM. QTL regions on chromosomes 1A, 4A, 4D, 6B and 7D overlapped with QTL regions detected for IT (Table 3). Nevertheless, smaller R2 of 21% (4A), 14% (7D) and < 1% (6B), as well as larger R2 of 12% (1A) and 9% (4D) were calculated for individual QTL. The maximal reducing effect of the QTL on chromosomes 1A, 4A, 4D, and 6B ranged between 0.5% and 4.1%, while for 7D, only the founder ‘BAYP4535′ showed a reducing allelic effect of 2.8%. Additionally, two QTL were detected on chromosome 7A at 65 cM and 99 cM, accounting for 5% and 8% of the phenotypic variance. SI ranged from 54 to 87 cM and from 94 to 111 cM, respectively. Founders ‘Firl3565′ and ‘Bussard’ contributed the largest allelic effect, reducing the disease severity by 0.9% and 2.4%.
Based on support intervals of 19 QTL, detected in total for the different traits, 11 main QTL were identified (Fig. S3, Table 4). In silico annotations of peak markers revealed seven genes with known functions partly involved in resistance. Hence, marker CAP8_c2448_355 on chromosome 1A referred to a DnaJ domain. A Protein kinase domain and a NB-ARC domain were identified for peak markers of QLr.jki-4A.1 and QLr.jki-4A.2 on chromosome 4A. Marker AX-95126745 on chromosome 4D and RAC875_c31670_389 on chromosome 5A referred to a cation/calcium exchanger 4 and ankyrin repeats, respectively. For peak markers of QLr.jki-7A.1 and QLr.jki-7A.1 on chromosome 7A, a pyridoxal-phosphate dependent enzyme and a sugar efflux transporter were annotated, respectively.
Table 4.
Main QTL for resistance to Puccinia triticina merged over all evaluated traits
| QTL | Chr.a | Peak marker | Pos.[cM]b | SI [cM]c | Pos.RefSeq [bp]d | Potential origin | Adjacent T.aestivum gene | Orthologous gene | Identity | Functional annotation | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Start | End | ||||||||||
| QLr.jki-1A.1 | 1A | CAP8_c2448_355 | 27.63 | 0–34 | 10069841 | 10069932 | Potenzial | TraesCS1A01G020600 | TRIUR3 04361e | 93.63 | DnaJ domain |
| RAC875_c57939_78 | 26.12 | 11571831 | 11571931 | TraesCS1A01G023400 | F775 01617f | 94.93 | |||||
| QLr.jki-4A.1 | 4A | Kukri_rep_c109167_89 | 133.99 | 125–151 | 634737614 | 634737686 | BAYP4535 | TraesCS4A01G361100 | TRIUR3 34719e | 99.78 | Protein kinase domain |
| F775 31833f | 99.28 | ||||||||||
| QLr.jki-4A.2 | 4A | BobWhite_c47168_598 | 171.04 | 168–174 | 726214891 | 726214991 | BAYP4535 | TraesCS4A01G461700 | NB-ARC domain | ||
| Excalibur_c46904_84 | 169.52 | 737340474 | 737340573 | TraesCS4A01G481400 | TRIUR3 03302e | 96.59 | |||||
| F775 10262f | 96.25 | ||||||||||
| QLr.jki-4D.1 | 4D | BS00023112_51 | 69.43 | 58–86 | 455763978 | 455764078 | Julius | TraesCS4D01G285000 | F775 08229f | 100.00 | |
| AX-95126745 | 71.96 | 464988433 | 464988533 | TraesCS4D01G294600 | F775 05351f | 99.69 | Cation/calcium exchanger 4 | ||||
| QLr.jki-5A.1 | 5A | IAAV2363 | 111.77 | 98–152 | 481901324 | 481901524 | Format | TraesCS5A01G271500 | F775 15669f | 96.01 | |
| RAC875_c31670_389 | 138.69 | 514094550 | 514094650 | TraesCS5A01G305200 | F775 21555f | 98.40 | Ankyrin repeats | ||||
| QLr.jki-6B.1 | 6B | AX-94557244 | 21.83 | 10–30 | 25914587 | 25914687 | Format | TraesCS6B01G041900 | |||
| QLr.jki-6B.2 | 6B | RAC875_c57692_88 | 249.34 | 247–250 | 712673112 | 712673182 | Event | TraesCS6B01G456500 | |||
| QLr.jki-7A.1 | 7A | BS00011330_51 | 64.66 | 54–87 | 63112744 | 63112844 | Firl3565 | TraesCS7A01G102800 | TRIUR3 02989e | 99.72 | Pyridoxal-phosphate dependent enzyme |
| F775 27910f | 95.24 | ||||||||||
| QLr.jki-7A.2 | 7A | wsnp_Ku_c26530_36497050 | 98.82 | 93–111 | 84772316 | 84772460 | Bussard | TRIUR3 06012e | 94.36 | ||
| QLr.jki-7A.3 | 7A | BS00011622_51 | 368.31 | 346–379 | 712309001 | 712309084 | Format | TraesCS7A01G533900 | TRIUR3 33918e | 88.99 | Sugar efflux transporter for intercellular exchange |
| F775 06947f | 87.19 | ||||||||||
| QLr.jki-7D.1 | 7D | AX-94930280 | 18.13 | 15–30 | 16119641 | 16119741 | BAYP4535 | TraesCS7D01G030600 | |||
| IACX11794 | 21.65 | 12470235 | 12470390 | TraesCS7D01G026100 | F775 15174f | 97.59 | |||||
a Chromosomal position of QTL
b Position of peak marker based onStadlmeier et al. (2018)
c Support interval
d Position of peak marker at the reference genome RefSeq v1.0
e Triticum urartu
f Aegilops tauschii
Discussion
Continuous evolution of leaf rust results in the emergence of new pathotypes virulent against single major resistance genes commonly present in cultivars. Many of these race specific Lr genes have been broken down in the past (Kolmer 2005; Serfling et al. 2013). Detection of effective leaf rust resistances is of essential importance to avoid rust epidemics. Therefore, experimental populations such as MAGIC populations provide powerful tools to discover, characterize, and deploy QTL for complex traits including resistances (Cavanagh et al. 2008). Out of 80 designated Lr genes, it was reported, that only Lr1, Lr3, Lr10, Lr13, Lr14a, Lr17b, Lr20, Lr24, Lr26, Lr34, and Lr37 were used individually or in combination in European varieties (Goyeau et al. 2006; Goyeau and Lannou 2011; Serfling et al. 2013). The BMW population emerged from crosses of eight elite parental lines originating from Germany and Denmark. Nevertheless, Stadlmeier et al. (2018) were able to show the potential of the BMW population to detect new QTL for resistance to powdery mildew, septoria tritici blotch, as well as tan spot, and in general the usefulness for further gene mapping studies (Stadlmeier et al. 2018, 2019).
In this study, phenotyping of 394 RILs from the BMW population resulted in a broad variability of resistance to Puccinia triticina. Despite an average correlation coefficient of 0.54 between the disease severities in five environments, a broad sense heritability of 0.83 was calculated which is in the range of previously published studies (Bemister et al. 2019; Gao et al. 2016; Zhang et al. 2017, 2019). This may hint to a quantitative inheritance due to QTL involved in slow rusting loci, which are characterized by relatively high heritabilities (Kolmer 1996). Phenotypic distribution for field trials was slightly right-skewed, while almost a bi-modal distribution was observed for both IT and Pi values in seedling test. This may give hint that mostly horizontal (quantitative) or vertical (qualitative) resistances were scored, respectively. Calculation of correlation between field data and seedling test results showed r values of 0.27 (IT) and 0.24 (Pi), which are in accordance with correlations reported by Gao et al. (2016). Different virulence/avirulence patterns of leaf rust races may be an explanation for these low correlations (Gao et al. 2016). While a single highly aggressive race, with many virulence genes was used for artificial inoculation for seedling tests and field trials in QLB, field trials in SOE and LEN were conducted under natural infection pressure.
Overall, simple interval mapping detected 19 QTL, which corresponded to 11 distinct chromosomal regions (Table 4, Fig. S3). QTLs identified using the LSmeans over all environments were also identified by analyzing the single environments separately. Out of the 11 distinct chromosomal regions three QTL were detected at the adult plant stage. Six QTL conferred seedling resistance and two were active in both growth stages, indicating the presence of effective all-stage leaf rust resistance genes. In total, the regions were located on wheat chromosomes 1A, 4A, 4D, 5A, 6B, 7A and 7D. Peak markers of QTL could be partially annotated to genes, known to be involved in quantitative resistances to leaf rust, e.g. sugar efflux transporters, DnaJ domain belonging to heat shock protein family (Bekh-Ochir et al. 2013), a protein kinase domain, a NB-ARC domain and a cation/calcium exchanger. Such genes show an increased expression during defense reactions in wheat-leaf rust (Sharma et al. 2018) and wheat-stripe rust interactions (Wang et al. 2020) and as response to environmental stresses.
In this study, QLr.jki-1A.1 on chromosome 1A is based on the evaluation of IT and Pi in seedling tests and is physically located in a region between 1.3 Mbp and 12.5 Mbp (Table 5). Pinto da Silva et al. (2018) reviewed 11 QTL described in hexaploid wheat located on chromosome 1A. Based on available physical marker positions, QLr.ccsu-1A.1 and QLr.cau-1AS identified in two different studies, were found to correspond to the region of QLr.jki-1A.1 (Du et al. 2015; Kumar et al. 2013). While QLr.ccsu-1A.1 is only 1.7 Mbp and 0.2 Mbp apart from our peak markers, the distance of the linked marker to QLr.cau-1AS is 2.4 Mbp and 3.9 Mbp, respectively (Tables 4, 5). Additionally, Elbasyoni et al. (2017) detected several marker-trait associations (MTAs) covering a region from 7.2 Mbp to 13.7 Mbp, which includes the region of QLr.jki-1A.1. Furthermore, the resistance gene Lr10, which is completely sequenced, is mapped at 12.6 Mbp, i.e. 2.5 Mbp and 1 Mbp apart from our peak marker (Table 4; Feuillet et al. 1997, 2003). Thus, and due to the fact that Lr10, Lr1, Lr3a and Lr20 are the most prevalent genes used worldwide, Lr10 is a promising candidate for the QTL aforementioned (Dakouri et al. 2013).
Table 5.
Comparison of physical positions of the QTL identified in the present study (bold) with those reported previously. Physical positions based on comparison of marker sequence data to the wheat reference genome (RefSeq1.0)
| QTL | Marker interval | Physical position [Mbp] | Genetic material | References |
|---|---|---|---|---|
| QLr.jki-1A.1 | IAAV3919–Tdurum_contig42479_3800 | 1.3–12.5 | BMW population (RILa) | Lr10? |
| QLr.ccsu-1A.1 | Xbarc263–Xcdo426 | 11.8–nab | Opata85 × W-7984 (RIL) | Kumar et al. (2013) |
| QLr.cau-1AS | gpw2246 | 7.7 | Luke × AQ24788-83 (RIL) | Du et al. (2015) |
| MTA | IWA3182–IWA7191 | 7.1–13.7 | Spring wheat collection | Elbasyoni et al. (2017) |
| Lr10 | 12.6 | Feuillet et al. (2003) | ||
| QLr.jki-4A.1 | AX-95253498–TA006348.0950 | 618.6–649.9 | BMW population (RIL) | |
| MTA | IWA2816 | 641.5 | Hexaploid Wheat Landraces | Kertho et al. (2015) |
| QLr.jki-4A.2 | Tdurum_contig75819_1220–Excalibur_c33542_113 | 712.9–na | BMW Population (RIL) | |
| 4A_t2 | BobWhite_c47168_289 | 726.2 | Elite spring wheat lines | Gao et al. (2016) |
| QLr.hebau-4AL | BobWhite_c15697_675–Excalibur_c2827_580 | 598.7–726.4 | Zhou8425B × Chinese Spring (RIL) | Zhang et al. (2017) |
| QLr.jki-4D.1 | AX-94793903–AX-94838884 | 130.9–479.7 | BMW population (RIL) | Novel? |
| QLr.fcu-4DL | Xgdm61–Xcfa2173 | na | TA4152-60 × ND495 (DHc) | Chu et al. (2009) |
| QLr.hebau-4DL | AX-110476142–AX-111092299 | 381.2–428.6 | Pingyuan50 × Mingxian169 | Zhang et al. (2019) |
| QLr.sfrs-4DL | Xglk302b–Xpsr1101a | na | Forno × Oberkulmer (RIL) | Messmer et al. (2000) |
| Lr67 | Xgwm165–Xgwm192 | 412.7 | RL6077 × Avocet (RIL) | Herrera-Foessel et al. (2011) |
| QLr.jki-5A.1 | AX-94732470–wsnp_Ex_c49211_53875600 | 444.6–na | BMW population | Novel? |
| QLr.cim-5AC | wPt-3187–wPt-7769 | Na–464.7 | Avocet-YrA × Kenya Kongoni (RIL) | Calvo-Salazar et al. (2015) |
| QLr.jki-6B.1 | AX-94739546–TA003005.0339 | 19.3–34.3 | BMW population | Novel? |
| QLr.caas-6BS.1 | Xcfd13–Xwmc487 | 34.2–36.5 | Bainong64 × Jingshuang16 (DH) | Ren et al. (2012) |
| QLr.wpt-6BS.2 | wPt2175 | nab | Winter wheat accessions | Gerard et al. (2018) |
| QLr.jki-6B.2 | wsnp_Ex_c54772_57528275–Excalibur_c29748_954 | 710.1–719.7 | BMW population | Lr3? |
| QLr.cim-6BL | 277,143–1,234,305 | 714.3–na | Bairds × Atred#1 (RIL) | Lan et al. (2017) |
| 6B_4 | BobWhite_c43263_180–BS00011795_51 | 718.9–720.6 | Elite spring wheat lines | Gao et al. (2016) |
| QLr.jki-7A.1 | BobWhite_rep_c58252_112–wsnp_BF473884A_Ta_1_3 | 54.9–71.1 | BMW population | Novel |
| QLr.jki-7A.2 | RAC875_c75528_355–BS00024786_51 | 79.6–na | BMW population | Novel? |
| QLr.stars-7AS1 | wsnp_Ex_c41150_48040078 | 78.4 | Winter wheat accessions | Li et al. (2016) |
| MTA | IWA7192 | 81.1 | Spring wheat collection | Elbasyoni et al. (2017) |
| Lr47 | 115 | Helguera et al. (2000) | ||
| QLr.jki-7A.3 | Tdurum_contig29240_206–wsnp_CAP11_c298_250917 | 702.4–724.1 | BMW Population | Lr20? |
| MTA | IWA4175 | 717.1 | Spring wheat accessions | Turner et al. 2017 |
| QLr.jki-7D.1 | TA016282.1180–AX-94883448 | na–29.4 | BMW Population | Novel |
| Lr34 | 47.4–51 | Krattinger et al. (2009) |
a Recombinant inbred line population
b marker information was not available or position could not be identified in the RefSeq v1.0
c Doubled haploid population
On chromosome 4A, two regions harboring leaf rust resistance were identified in this study (QLr.jki-4A.1, QLr.jki-4A.2, Table 4). To date, there are two Lr genes, Lr28 originating from Ae. speltoides and Lr30 from T. aestivum, and two QTL reported on chromosome 4A (Dyck and Kerber 1971; McIntosh et al. 2013; Pinto da Silva et al. 2018). Kertho et al. (2015) found one MTA at 641.5 Mbp, using the leaf rust race MCDL. Therefore, the marker is physically located within the region of QLr.jki-4A.1, but 6.8 Mbp apart from our peak marker. Due to the specific virulence pattern of the MCDL race, which is avirulent to Lr30, the MCDL-MTA might identify this Lr gene. However, to our knowledge, no mapping information for Lr30 is available to allow a more precise comparison between Lr30, the MCDL-MTA and QLr.jki-4A.1 detected in this study. Another significant MTA (4A_t2, Gao et al. 2016) was detected in the region of QLr.jki-4A.2, only 309 bp apart from the peak marker for this QTL (Table 4). 4A_t2 was mapped approximately at the position of the marker linked to Lr28 (Bipinraj et al. 2011). This may be a hint that QLr.jki-4A.2 also corresponds to Lr28, but further analyses have to be conducted. Furthermore, Zhang et al. (2017) reported a minor QTL for APR in Chinese Spring (QLr.hebau-4AL), which is physically located between 598.7 Mbp and 726.4 Mbp. This region includes both QTL on chromosome 4A detected in this study (Table 5).
In total, nine QTLs were detected on chromosome 4D so far, including the resistance gene Lr67/Yr46/Sr55 (Herrera-Foessel et al. 2011; McIntosh et al. 2013; Pinto da Silva et al. 2018). In this study, QLr.jki-4D.1 was detected for both IT and Pi in the seedling tests and mapped at the distal end of chromosome 4DL. Physically, it is located in a large interval from 130.9 Mbp to 479.7 Mbp (Table 5) with peak markers at 455.8 Mbp and 465 Mbp, respectively (Table 4). Chu et al. (2009) located a QTL (QLr.fcu-4DL) in douple-haploid population ‘TA4152-60 × ND495′, mapped at a similar position as Lr67, around 412.7 Mbp (Herrera-Foessel et al. 2011; Zhang et al. 2019). Another QTL on chromosome 4DL (QLr.hebau-4D) was located between 381.2 Mbp and 428.6 Mbp (Zhang et al. 2019). Considering the physical distances to our peak marker, it appears that QLr.jki-4D.1 is independent from QLr.fcu-4DL, QLr.hebau-4D, and Lr67 (Table 5). A higher similarity may exist with another QTL (QLr.sfrs-4DL) detected by Messmer et al. (2000). This QTL resulted in an APR and was mapped in the Swiss RIL population ‘Forno × Oberkulmer’ also at the distal end of chromosome 4DL. Since QLr.jki-4D.1 has only been detected at the seedling stage, QLr.sfrs-4DL also seems to be located in a different region and with the available data, it is not possible to further determine whether it corresponds to our regions.
On chromosome 5A one QTL (QLr.jki-5A.1) was detected in seedling tests for IT (Table 4). To our knowledge, on chromosome 5A there is no designated Lr gene and only two QTL (QLr.cim-5AC, QLr.cimmyt-5A) are known (Calvo-Salazar et al. 2015; Rosewarne et al. 2012). QLr.cimmyt-5A was mapped on the long arm of chromosome 5A, closely linked to Vrn-A1 at 587.0 Mbp (Rosewarne et al. 2012). QLr.cim-5AC was located in the centromeric region of chromosome 5A and flanked by markers wPt-7769 and wPt-3187, of which the latter is located at 464.7 Mbp (Table 5). When comparing the physical positions of these three QTL, it is more likely that QLr.jki-5A.1 corresponds to QLr.cim-5AC or is a novel QTL.
On chromosome 6B, two QTL were identified (QLr.jki-6B.1 and QLr.jki-6B.2) in the present study (Table 4). QLr.jki-6B.1 was mapped on the short arm of chromosome 6B, at 19.3—34.3 Mbp (Table 5). Up to now, 5 QTL have been described on chromosome 6BS, but only QLr.caas-6BS.1, derived from the wheat cultivar Bainong 64, was physically localized in the region between 32 and 34 Mbp (Gerard et al. 2018; Kankwatsa et al. 2017; Ren et al. 2012). Gerard et al. (2018) stated that another QTL (QLr.wpt-6BS.2) is genetically located in the same region as QLr.caas 6BS.1, whereas QLr.wpt-6BS.2 was mapped close to the centromere, a region clearly distinct from QLr.jki-6B.1 (Table 5). Therefore, further studies are required to confirm whether our QTL is located closely to these known QTL. The second QTL QLr.jki-6B.2 was mapped at the distal end of chromosome 6BL, within a small interval encompassing 247 cM to 250 cM (710 – 720 Mbp). Out of six QTL already detected on chromosome 6BL, two QTL (QLr.cim-6BL and 6B_4) were also located at the distal end of chromosome 6BL (Chu et al. 2009; Gao et al. 2016; Lan et al. 2017; Rosewarne et al. 2012; William et al. 2006). The DArTseq markers 1234305 and 2277143 flank QLr.cim-6BL detected by Lan et al. (2017). Marker 2277143 was converted into a diagnostic KASP marker, which is located at 714.3 Mbp, i.e. 1.6 Mbp distal from our peak marker of QLr.jki-6B.2 (Tables 4 , 5 ). The results of Lan et al. (2017) indicated uniqueness of QLr.cim-6BL, showing no relationship to other QTL on chromosome 6BL, as well as to Lr3a co-segregating with Xmwg798 (Sacco et al. 1998). However, the second known QTL 6B_4 was physically mapped between 718.9 Mbp and 720.6 Mbp, and appeared to be in high linkage disequilibrium with Lr3 (Gao et al. 2016). Regarding the similar physical regions, QLr.jki-6B.2 may correspond to QLr.cim-6BL and 6B_4, but further research is needed to come to a closer understanding of the relationship between these QTL and Lr3.
On chromosome 7A, the major resistance genes Lr20, forming a disease-resistance gene cluster with Pm1, and Lr47, which was transferred from chromosome 7S of Ae. speltoides have been reported (Dubcovsky et al. 1998; Neu et al. 2002). Additionally, three QTL on chromosome 7AL and several MTAs were detected (Pinto da Silva et al. 2018). In the present study, three QTL (QLr.jki-7A.1 to QLr.jki-7A.3) were identified on chromosome 7A. The first two QTL were detected for Pi in the seedling test and their support intervals were separated from each other by a map distance of 7.1 cM on chromosome 7AS. QLr.jki-7A.1 was physically mapped between 54.9 Mbp and 71.1 Mbp (Table 5). To our knowledge, no QTL have been reported in this region. Hence, QLr.jki-7A.1 might be a novel QTL. The second QTL (QLr.jki-7A.2) on chromosome 7AS was located between 93 and 111 cM. The peak marker was mapped at 84.8 Mbp (Table 4). To date, there are two MTAs from different studies detected in similar regions as QLr.jki-7A.2 (Elbasyoni et al. 2017; Li et al. 2016). The first MTA (QLr.stars-7AS1), associated with marker IWA3760 was mapped at 78.4 Mbp, hence, it appears that QLr.stars-7AS1 does not correspond to QLr.jki-7A.2. The second MTA (IWA7192) was detected by Elbasyoni et al. (2017) at 81.1 Mbp, and might be correspondent to the resistance gene Lr47. When comparing the physical position of a diagnostic marker for Lr47 (around 115 Mbp), both IWA7192, and QLr.jki-7A.2 seem to be different from this Lr gene (Helguera et al. 2000). Thus, QLr.jki-7A.2 is likely a novel locus involved in resistance to P. triticina.
The third QTL (QLr.jki-7A.3) determined in field trials during this study was mapped between 346 and 379 cM on chromosome 7AL. This translates to a large physical distance between 702.4 Mbp and 724.1 Mbp, with the peak marker at 712.3 Mbp (Tables 4, 5). Out of five known regions on chromosome 7AL involved in leaf rust resistance (Kankwatsa et al. 2017; Li et al. 2016; Lu et al. 2017; Tsilo et al. 2014), only the MTA detected by Turner et al. (2017) may be localized within the region of QLr.jki-7A.3. The associated marker IWA4175 was mapped at 717.1 Mbp, which is 4.8 Mbp apart from our peak marker. However, after Bonferroni correction, the marker was no longer significant (P < 0.1). The Lr gene Lr20 is genetically located in the distal part of chromosome 7AL (Neu et al. 2002), which may correspond to QLr.jki-7A.3. Based on the available data, investigations with diagnostic markers need to be conducted to gain further insights.
Finally, one QTL was detected on chromosome 7DS, based on phenotypic data from field trials and seedling test (Table 4). To date, out of 21 QTL reported on chromosome 7D, 19 correspond to the resistance gene Lr34, which confers race non-specific, partial, and slow rusting resistance to leaf rust (Lagudah et al. 2009; Pinto da Silva et al. 2018). Lr34 has been physically located at 47.4 Mbp (Krattinger et al. 2009). Thus, QLr.jki-7D.1 identified in our study does not correspond to the resistance gene Lr34 and the 19 QTL reported (Table 5). The remaining two QTL QLr.cim-7DS and QLr.hebau-7DS on chromosome 7DS, which were detected in the two RIL populations ‘Avocet-YrA × Francolin#1′ and ‘Shanghai3/Catbird × Naxos’, respectively, were located in different chromosome region (Lan et al. 2014; Zhou et al. 2014). Hence, QLr.jki-7D.1 seems to be a novel locus.
The objective of this study was to identify QTL for resistance to leaf rust, using the Bavarian MAGIC Wheat population. We identified 19 leaf rust resistance QTL that were confined to 11 distinct chromosomal regions. To date, more than 249 leaf rust resistance QTL and 200 MTAs were reported covering all 21 chromosomes of hexaploid wheat (Pinto da Silva et al. 2018). These regions were identified in several mapping populations using different genotyping methods. Because of the absence of information on physical positions for many of these QTL, it is difficult to unequivocally determine the identity of newly described QTL. In the present study, six putatively new QTL were identified on chromosomes 4D, 5A, 6B, 7A and 7D. SNP markers linked to these regions may be converted into KASP markers suitable for MAS in wheat breeding programs (Neelam et al. 2013; Rasheed et al. 2016). This will enable stacking of the detected resistance loci to breed new varieties with an improved resistance to leaf rust.
Electronic supplementary material
Below is the link to the electronic supplementary material.
{kind=link}
Fig. S1 Pearson correlation of leaf rust severity between different field trials. Diagonals are histograms of each environment (Lengern LEN 2018, Quedlinburg QLB 2017 2018, Söllingen SOE 2017 2018). *** denotes significance at α = 0.001. Lowess curves were adjusted to the data points with a smoothing range of 0.75, based on the ‘lowess’ function implemented in the R-based ‘stats’ package (JPEG 133 kb)
{kind=link}
Fig. S2 Pearson correlation (r) between averaged infection type (IT), infected leaf area (Pi) of seedling test and average ordinate (AO) of field trials (A, B), as well as correlation between IT and Pi (C). *** denotes significance at α = 0.001 (JPEG 78 kb)
{kind=link}
Fig. S3 Simple interval mapping of resistance to Puccinia triticina in field trials (A) and seedling test (B, C). The x axis shows the 21 wheat chromosomes. Positions are based on the genetic map, and the log10(p) values of each Marker are displayed on the y axis (black line). The red horizontal line represents the significance thresholds. SI of the significant QTL detected in this study are coloured in blue (JPEG 188 kb)
Tab. S1 List of virulences and avirulences of Puccinia triticina isolate 77WxR used in field trials and seedling test. Brackets indicate ambiguous results due to the differing symptom ratings between replications or moderate susceptibility (based on Zetzsche et al. 2019) (DOCX 13 kb)
Tab. S2 Complete information of the 19 QTL for leaf rust resistance in BMW population, evaluated in field trials (AO) and seedling test (IT and Pi) (XLSX 16 kb)
Acknowledgement
This research was supported by the Federal Ministry for Food and Agriculture (BMEL) (FKZ 2818200515). The authors deeply thank all project partners from Strube Research and Breun Saatzucht for conducting and scoring the field trials in Söllingen and Lenglern. We also gratefully thank Melanie Stadlmeier for providing the genotypic data and the genetic map of the BMWpop, as well as for helpful advice on the use of ‘mpMap’ package. The authors thank NIAB for provision of MAGIC analysis script.
Author contributions statement
LH and FO planned and managed the project. LH and MG provided and characterized all RILs. MG contributed in the analyses of the results. AS, FO, MG, LH, and VM contributed to the interpretation and discussion of the results. SR conducted the field screenings and seedling test, analyzed the data and wrote the manuscript.
Funding
Open Access funding provided by Projekt DEAL.
Compliance with ethical standards
Conflict of interest
The authors declare that there are no conflicts of interest in the reported research.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Sandra Rollar, Email: sandra.rollar@julius-kuehn.de.
Frank Ordon, Email: frank.ordon@julius-kuehn.de.
References
- Aktar-Uz-Zaman M, Tuhina-Khatun M, Hanafi MM, Sahebi M. Genetic analysis of rust resistance genes in global wheat cultivars: an overview. Biotechnol Biotechnol Equip. 2017 doi: 10.1080/13102818.2017.1304180. [DOI] [Google Scholar]
- Asimit J, Zeggini E. Rare variant association analysis methods for complex traits. Annu Rev Genet. 2010 doi: 10.1146/annurev-genet-102209-163421. [DOI] [PubMed] [Google Scholar]
- Bandillo N, Raghavan C, Muyco PA, Sevilla MAL, Lobina IT, Dilla-Ermita CJ, Tung C-W, McCouch S, Thomson M, Mauleon R, Singh RK, Gregorio G, Redoña E, Leung H. Multi-parent advanced generation inter-cross (MAGIC) populations in rice: progress and potential for genetics research and breeding. Rice (N Y) 2013 doi: 10.1186/1939-8433-6-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bayer MM, Rapazote-Flores P, Ganal M, Hedley PE, Macaulay M, Plieske J, Ramsay L, Russell J, Shaw PD, Thomas W, Waugh R. Development and evaluation of a barley 50k iSelect SNP array. Front Plant Sci. 2017 doi: 10.3389/fpls.2017.01792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bekh-Ochir D, Shimada S, Yamagami A, Kanda S, Ogawa K, Nakazawa M, Matsui M, Sakuta M, Osada H, Asami T, Nakano T. A novel mitochondrial DnaJ/Hsp40 family protein BIL2 promotes plant growth and resistance against environmental stress in brassinosteroid signaling. Planta. 2013;237:1509–1525. doi: 10.1007/s00425-013-1859-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bemister DH, Semagn K, Iqbal M, Randhawa H, Strelkov SE, Spaner DM. Mapping QTL associated with stripe rust, leaf rust, and leaf spotting in a Canadian spring wheat population. Crop Sci. 2019 doi: 10.2135/cropsci2018.05.0348. [DOI] [Google Scholar]
- Bipinraj A, Honrao B, Prashar M, Bhardwaj S, Rao S, Tamhankar S. Validation and identification of molecular markers linked to the leaf rust resistance gene Lr28 in wheat. J Appl Genet. 2011 doi: 10.1007/s13353-010-0026-9. [DOI] [PubMed] [Google Scholar]
- Bolton MD, Kolmer JA, Garvin DF. Wheat leaf rust caused by Puccinia triticina. Mol Plant Pathol. 2008 doi: 10.1111/j.1364-3703.2008.00487.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Braun HJ, Atlin G, Payne T. Multi-location testing as a tool to identify plant response to global climate change. In: Reynolds MP, editor. Climate change and crop production. Wallingford, Oxfordshire, UK, Cambridge, MA: CABI; 2010. pp. 115–138. [Google Scholar]
- Broman KW, Wu H, Sen S, Churchill GA. R/qtl: QTL mapping in experimental crosses. Bioinformatics. 2003 doi: 10.1093/bioinformatics/btg112. [DOI] [PubMed] [Google Scholar]
- Calvo-Salazar V, Singh RP, Huerta-Espino J, Cruz-Izquierdo S, Lobato-Ortiz R, Sandoval-Islas S, Vargas-Hernández M, German S, Silva P, Basnet BR, Lan CX, Herrera-Foessel SA. Genetic Analysis of Resistance to Leaf Rust and Yellow Rust in Spring Wheat Cultivar Kenya Kongoni. Plant Dis. 2015 doi: 10.1094/PDIS-07-14-0718-RE. [DOI] [PubMed] [Google Scholar]
- Cavanagh C, Morell M, Mackay I, Powell W. From mutations to MAGIC: resources for gene discovery, validation and delivery in crop plants. Curr Opin Plant Biol. 2008 doi: 10.1016/j.pbi.2008.01.002. [DOI] [PubMed] [Google Scholar]
- Chen H, Xie W, He H, Yu H, Chen W, Li J, Yu R, Yao Y, Zhang W, He Y, Tang X, Zhou F, Deng XW, Zhang Q. A high-density SNP genotyping array for rice biology and molecular breeding. Mol Plant. 2014 doi: 10.1093/mp/sst135. [DOI] [PubMed] [Google Scholar]
- Chu C-G, Friesen TL, Xu SS, Faris JD, Kolmer JA. Identification of novel QTLs for seedling and adult plant leaf rust resistance in a wheat doubled haploid population. Theor Appl Genet. 2009 doi: 10.1007/s00122-009-1035-0. [DOI] [PubMed] [Google Scholar]
- Churchill GA, Doerge RW. Empirical threshold values for quantitative trait mapping. Genetics. 1994;138:963–971. doi: 10.1093/genetics/138.3.963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Churchill GA, Airey DC, Allayee H, Angel JM, Attie AD, Beatty J, Beavis WD, Belknap JK, Bennett B, Berrettini W, Bleich A, Bogue M, Broman KW, Buck KJ, Buckler E, Burmeister M, Chesler EJ, Cheverud JM, Clapcote S, Cook MN, Cox RD, Crabbe JC, Crusio WE, Darvasi A, Deschepper CF, Doerge RW, Farber CR, Forejt J, Gaile D, Garlow SJ, Geiger H, Gershenfeld H, Gordon T, Gu J, Gu W, de Haan G, Hayes NL, Heller C, Himmelbauer H, Hitzemann R, Hunter K, Hsu H-C, Iraqi FA, Ivandic B, Jacob HJ, Jansen RC, Jepsen KJ, Johnson DK, Johnson TE, Kempermann G, Kendziorski C, Kotb M, Kooy RF, Llamas B, Lammert F, Lassalle J-M, Lowenstein PR, Lu L, Lusis A, Manly KF, Marcucio R, Matthews D, Medrano JF, Miller DR, Mittleman G, Mock BA, Mogil JS, Montagutelli X, Morahan G, Morris DG, Mott R, Nadeau JH, Nagase H, Nowakowski RS, O’Hara BF, Osadchuk AV, Page GP, Paigen B, Paigen K, Palmer AA, Pan H-J, Peltonen-Palotie L, Peirce J, Pomp D, Pravenec M, Prows DR, Qi Z, Reeves RH, Roder J, Rosen GD, Schadt EE, Schalkwyk LC, Seltzer Z’e, Shimomura K, Shou S, Sillanpää MJ, Siracusa LD, Snoeck H-W, Spearow JL, Svenson K, Tarantino LM, Threadgill D, Toth LA, Valdar W, de Villena FP-M, Warden C, Whatley S, Williams RW, Wiltshire T, Yi N, Zhang D, Zhang M, Zou F. The Collaborative Cross, a community resource for the genetic analysis of complex traits. Nat Genet. 2004 doi: 10.1038/ng1104-1133. [DOI] [PubMed] [Google Scholar]
- Cui F, Zhang N, Fan X-l, Zhang W, Zhao C-h, Yang L-j, Pan R-q, Chen M, Han J, Zhao X-q, Ji J, Tong Y-p, Zhang H-x, Jia J-z, Zhao G-y, Li J-m. Utilization of a wheat 660K SNP array-derived high-density genetic map for high-resolution mapping of a major QTL for kernel number. Sci Rep. 2017 doi: 10.1038/s41598-017-04028-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dakouri A, McCallum BD, Radovanovic N, Cloutier S. Molecular and phenotypic characterization of seedling and adult plant leaf rust resistance in a world wheat collection. Mol Breed. 2013 doi: 10.1007/s11032-013-9899-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dell’Acqua M, Gatti DM, Pea G, Cattonaro F, Coppens F, Magris G, Hlaing AL, Aung HH, Nelissen H, Baute J, Frascaroli E, Churchill GA, Inzé D, Morgante M, Pè ME. Genetic properties of the MAGIC maize population: a new platform for high definition QTL mapping in Zea mays. Genome Biol. 2015 doi: 10.1186/s13059-015-0716-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Douchkov D, Lück S, Baum T, Seiffert U, Schweizer P (2012) High-throughput phenomics platform for interactions of barley with fungal pathogens. In: Plant & Animal Genome (PAG-XX), Conference San Diego, Poster, P0115.
- Du Z, Che M, Li G, Chen J, Quan W, Guo Y, Wang Z, Ren J, Zhang H, Zhang Z. A QTL with major effect on reducing leaf rust severity on the short arm of chromosome 1A of wheat detected across different genetic backgrounds and diverse environments. Theor Appl Genet. 2015 doi: 10.1007/s00122-015-2533-x. [DOI] [PubMed] [Google Scholar]
- Dubcovsky J, Lukaszewski AJ, Echaide M, Antonelli EF, Porter DR. Molecular Characterization of Two Triticum speltoides Interstitial Translocations Carrying Leaf Rust and Greenbug Resistance Genes. Crop Sci. 1998 doi: 10.2135/cropsci1998.0011183X003800060040x. [DOI] [Google Scholar]
- Dyck PL, Kerber ER. Chromosome location of three genes for leaf rust resistance in common wheat. Can J Genet Cytol. 1971 doi: 10.1139/g71-072. [DOI] [Google Scholar]
- Elbasyoni I, El-Orabey W, Baenziger P, Eskridge K. Association mapping for leaf and stem rust resistance using worldwide spring wheat collection. AJOB. 2017 doi: 10.9734/AJOB/2017/38120. [DOI] [Google Scholar]
- Feuillet C, Schachermayr G, Keller B. Molecular cloning of a new receptor-like kinase gene encoded at the Lr10 disease resistance locus of wheat. Plant J. 1997 doi: 10.1046/j.1365-313x.1997.11010045.x. [DOI] [PubMed] [Google Scholar]
- Feuillet C, Travella S, Stein N, Albar L, Nublat A, Keller B. Map-based isolation of the leaf rust disease resistance gene Lr10 from the hexaploid wheat (Triticum aestivum L.) genome. Proc Natl Acad Sci USA. 2003 doi: 10.1073/pnas.2435133100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flor HH. The complementary genic systems in flax and flax rust. Adv Genet. 1956 doi: 10.1016/S0065-2660(08)60498-8. [DOI] [Google Scholar]
- Flor HH. Current status of the gene-for-gene concept. Annu Rev Phytopathol. 1971 doi: 10.1146/annurev.py.09.090171.001423. [DOI] [Google Scholar]
- Food and Agriculture Organization of the United Nations (FAO). FAOSTAT. Crops. Latest update: 18/01/2019. Accessed 19/06/2019. URL https://www.fao.org/faostat/en/#data/QC
- Gao L, Turner MK, Chao S, Kolmer J, Anderson JA. Genome wide association study of seedling and adult plant leaf rust resistance in elite spring wheat breeding lines. PLoS ONE. 2016 doi: 10.1371/journal.pone.0148671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardner KA, Wittern LM, Mackay IJ. A highly recombined, high-density, eight-founder wheat MAGIC map reveals extensive segregation distortion and genomic locations of introgression segments. Plant Biotechnol J. 2016 doi: 10.1111/pbi.12504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerard GS, Kobiljski B, Lohwasser U, Börner A, Simón MR. Genetic architecture of adult plant resistance to leaf rust in a wheat association mapping panel. Plant Pathol. 2018 doi: 10.1111/ppa.12761. [DOI] [Google Scholar]
- Gibson G. Rare and common variants: twenty arguments. Nat Rev Genet. 2012 doi: 10.1038/nrg3118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gill HS, Li C, Sidhu JS, Liu W, Wilson D, Bai G, Gill BS, Sehgal SK. Fine Mapping of the Wheat Leaf Rust Resistance Gene Lr42. Int J Mol Sci. 2019 doi: 10.3390/ijms20102445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goyeau H, Lannou C. Specific resistance to leaf rust expressed at the seedling stage in cultivars grown in France from 1983 to 2007. Euphytica. 2011 doi: 10.1007/s10681-010-0261-5. [DOI] [Google Scholar]
- Goyeau H, Park R, Schaeffer B, Lannou C. Distribution of pathotypes with regard to host cultivars in French wheat leaf rust populations. Phytopathology. 2006 doi: 10.1094/PHYTO-96-0264. [DOI] [PubMed] [Google Scholar]
- Hallauer AR, Carena MJ, Miranda Filho JB (2010) Quantitative genetics in maize breeding, [Second edition]. Handbook of plant breeding, vol 6. Springer, New York, Dordrecht, Heidelberg, London
- He J, Zhao X, Laroche A, Lu Z-X, Liu H, Li Z. Genotyping-by-sequencing (GBS), an ultimate marker-assisted selection (MAS) tool to accelerate plant breeding. Front Plant Sci. 2014 doi: 10.3389/fpls.2014.00484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helguera M, Khan IA, Dubcovsky J. Development of PCR markers for the wheat leaf rust resistance gene Lr47. Theor Appl Genet. 2000 doi: 10.1007/s001220051397. [DOI] [Google Scholar]
- Herrera-Foessel SA, Singh RP, Huerta-Espino J, Crossa J, Yuen J, Djurle A. Effect of leaf rust on grain yield and yield traits of durum wheats with race-specific and slow-rusting resistance to leaf rust. Plant Dis. 2006 doi: 10.1094/PD-90-1065. [DOI] [PubMed] [Google Scholar]
- Herrera-Foessel SA, Lagudah ES, Huerta-Espino J, Hayden MJ, Bariana HS, Singh D, Singh RP. New slow-rusting leaf rust and stripe rust resistance genes Lr67 and Yr46 in wheat are pleiotropic or closely linked. Theor Appl Genet. 2011 doi: 10.1007/s00122-010-1439-x. [DOI] [PubMed] [Google Scholar]
- Huang BE, George AW, Forrest KL, Kilian A, Hayden MJ, Morell MK, Cavanagh CR. A multiparent advanced generation inter-cross population for genetic analysis in wheat. Plant Biotechnol J. 2012 doi: 10.1111/j.1467-7652.2012.00702.x. [DOI] [PubMed] [Google Scholar]
- Huang BE, George AW. R/mpMap: a computational platform for the genetic analysis of multiparent recombinant inbred lines. Bioinformatics. 2011 doi: 10.1093/bioinformatics/btq719. [DOI] [PubMed] [Google Scholar]
- Kankwatsa P, Singh D, Thomson PC, Babiker EM, Bonman JM, Newcomb M, Park RF. Characterization and genome-wide association mapping of resistance to leaf rust, stem rust and stripe rust in a geographically diverse collection of spring wheat landraces. Mol Breed. 2017 doi: 10.1007/s11032-017-0707-8. [DOI] [Google Scholar]
- Kertho A, Mamidi S, Bonman JM, McClean PE, Acevedo M. Genome-wide association mapping for Resistance to Leaf and Stripe Rust in Winter-Habit Hexaploid Wheat Landraces. PLoS ONE. 2015 doi: 10.1371/journal.pone.0129580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- King EG, Macdonald SJ, Long AD. Properties and power of the Drosophila Synthetic Population Resource for the routine dissection of complex traits. Genetics. 2012 doi: 10.1534/genetics.112.138537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kolmer JA. Genetics of resistance to wheat leaf rust. Annu Rev Phytopathol. 1996 doi: 10.1146/annurev.phyto.34.1.435. [DOI] [PubMed] [Google Scholar]
- Kolmer JA. Tracking wheat rust on a continental scale. Curr Opin Plant Biol. 2005 doi: 10.1016/j.pbi.2005.05.001. [DOI] [PubMed] [Google Scholar]
- Kover PX, Valdar W, Trakalo J, Scarcelli N, Ehrenreich IM, Purugganan MD, Durrant C, Mott R. A multiparent advanced generation inter-cross to fine-map quantitative traits in Arabidopsis thaliana. PLoS Genet. 2009 doi: 10.1371/journal.pgen.1000551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krattinger SG, Keller B. Molecular genetics and evolution of disease resistance in cereals. New Phytol. 2016 doi: 10.1111/nph.14097. [DOI] [PubMed] [Google Scholar]
- Krattinger SG, Lagudah ES, Spielmeyer W, Singh RP, Huerta-Espino J, McFadden H, Bossolini E, Selter LL, Keller B. A putative ABC transporter confers durable resistance to multiple fungal pathogens in wheat. Science. 2009 doi: 10.1126/science.1166453. [DOI] [PubMed] [Google Scholar]
- Kumar A, Chhuneja P, Jain S, Kaur S, Balyan HS, Gupta PK. Mapping main effect QTL and epistatic interactions for leaf rust and yellow rust using high density ITMI linkage map. Aust J Crop Sci. 2013;7:492–499. [Google Scholar]
- Lagudah ES, Krattinger SG, Herrera-Foessel S, Singh RP, Huerta-Espino J, Spielmeyer W, Brown-Guedira G, Selter LL, Keller B. Gene-specific markers for the wheat gene Lr34/Yr18/Pm38 which confers resistance to multiple fungal pathogens. Theor Appl Genet. 2009 doi: 10.1007/s00122-009-1097-z. [DOI] [PubMed] [Google Scholar]
- Lan C, Rosewarne GM, Singh RP, Herrera-Foessel SA, Huerta-Espino J, Basnet BR, Zhang Y, Yang E. QTL characterization of resistance to leaf rust and stripe rust in the spring wheat line Francolin#1. Mol Breed. 2014 doi: 10.1007/s11032-014-0075-6. [DOI] [Google Scholar]
- Lan C, Basnet BR, Singh RP, Huerta-Espino J, Herrera-Foessel SA, Ren Y, Randhawa MS. Genetic analysis and mapping of adult plant resistance loci to leaf rust in durum wheat cultivar Bairds. Theor Appl Genet. 2017 doi: 10.1007/s00122-016-2839-3. [DOI] [PubMed] [Google Scholar]
- Li G, Xu X, Bai G, Carver BF, Hunger R, Bonman JM, Kolmer J, Dong H. Genome-wide association mapping reveals novel QTL for seedling leaf rust resistance in a worldwide collection of winter wheat. Plant Genome. 2016 doi: 10.3835/plantgenome2016.06.0051. [DOI] [PubMed] [Google Scholar]
- Lu Y, Bowden RL, Zhang G, Xu X, Fritz AK, Bai G. Quantitative Trait Loci for Slow-Rusting Resistance to Leaf Rust in Doubled-Haploid Wheat Population CI13227 × Lakin. Phytopathology. 2017 doi: 10.1094/PHYTO-09-16-0347-R. [DOI] [PubMed] [Google Scholar]
- Mackay IJ, Bansept-Basler P, Barber T, Bentley AR, Cockram J, Gosman N, Greenland AJ, Horsnell R, Howells R, O’Sullivan DM, Rose GA, Howell PJ (2014) An eight-parent multiparent advanced generation inter-cross population for winter-sown wheat: creation, properties, and validation. G3 (Bethesda) 4:1603–1610. 10.1534/g3.114.012963 [DOI] [PMC free article] [PubMed]
- Mammadov J, Aggarwal R, Buyyarapu R, Kumpatla S. SNP markers and their impact on plant breeding. Int J Plant Genomics. 2012 doi: 10.1155/2012/728398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marasas CN, Smale M, Singh RP (2004) The economic impact in developing countries of leaf rust resistance breeding in CIMMYT-related spring bread wheat. Economics Program Paper 04–01, DF, Mexico
- McIntosh RA, Wellings CR, Park RF (1995) Wheat rusts: An atlas of resistances genes. CSIRO Publishing
- McIntosh RA, Yamazaki Y, Dubcovsky J, Rogers J, Morris C, Somers DJ, Appels R, Devos KM (2013) Catalogue of gene symbols for wheat: 20013 supplement. https://shigen.nig.ac.jp/wheat/komugi/genes/download.jsp
- McIntosh RA, Dubcovsky J, Rogers WJ, Morris C, Xia XC (2017) Catalogue of gene symbols for wheat: 2017 supplement. https://shigen.nig.ac.jp/wheat/komugi/genes/macgene/supplement2017.pdf
- Melching JS (1967) Improved deposition of airborne urediospores of Puccinia graminis and P. striiformis on glass slides and on wheat leaves by use of a turntable. Phytopathology 57:647 (abstr.)
- Messmer MM, Seyfarth R, Keller M, Schachermayr G, Winzeler M, Zanetti S, Feuillet C, Keller B. Genetic analysis of durable leaf rust resistance in winter wheat. Theor Appl Genet. 2000 doi: 10.1007/s001220050055. [DOI] [Google Scholar]
- Milner SG, Maccaferri M, Huang BE, Mantovani P, Massi A, Frascaroli E, Tuberosa R, Salvi S. A multiparental cross population for mapping QTL for agronomic traits in durum wheat (Triticum turgidum ssp. durum) Plant Biotechnol J. 2016 doi: 10.1111/pbi.12424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moll E, Walther U, Flath K, Prochnow J, Sachs E (1996) Methodische Anleitung zur Bewertung der partiellen Resistenz und die SAS-Anwendung RESI. Mitteilungen aus der Biologischen Bundesanstalt für Land- und Forstwirtschaft:p. 4–6
- Moll E, Flath K, Tessenow I (2010) Assessment of resistance in cereal cultivars. Design and analysis of experiments using the SAS-application RESI 2. Berichte aus dem Julius-Kühn-Institut 154
- Neelam K, Brown-Guedira G, Huang L. Development and validation of a breeder-friendly KASPar marker for wheat leaf rust resistance locus Lr21. Mol Breed. 2013 doi: 10.1007/s11032-012-9773-0. [DOI] [Google Scholar]
- Neu C, Stein N, Keller B. Genetic mapping of the Lr20-Pm1 resistance locus reveals suppressed recombination on chromosome arm 7AL in hexaploid wheat. Genome. 2002 doi: 10.1139/g02-040. [DOI] [PubMed] [Google Scholar]
- Nover I, Lehmann CO. Resistenzeigenschaften im Gersten- und Weizensortiment Gatersleben. Die Kulturpflanze. 1967 doi: 10.1007/BF02095713. [DOI] [Google Scholar]
- Ongom PO, Ejeta G (2018) Mating design and genetic structure of a Multi-Parent Advanced Generation Intercross (MAGIC) population of sorghum (Sorghum bicolor (L.) Moench). G3 (Bethesda) 8:331–341. 10.1534/g3.117.300248 [DOI] [PMC free article] [PubMed]
- Pascual L, Desplat N, Huang BE, Desgroux A, Bruguier L, Bouchet J-P, Le QH, Chauchard B, Verschave P, Causse M. Potential of a tomato MAGIC population to decipher the genetic control of quantitative traits and detect causal variants in the resequencing era. Plant Biotechnol J. 2015 doi: 10.1111/pbi.12282. [DOI] [PubMed] [Google Scholar]
- Pinto da Silva GB, Zanella CM, Martinelli JA, Chaves MS, Hiebert CW, McCallum BD, Boyd LA. Quantitative trait loci conferring leaf rust resistance in hexaploid wheat. Phytopathology. 2018 doi: 10.1094/PHYTO-06-18-0208-RVW. [DOI] [PubMed] [Google Scholar]
- R Core Team (2017) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available at: https://www.R-project.org/
- Rasheed A, Wen W, Gao F, Zhai S, Jin H, Liu J, Guo Q, Zhang Y, Dreisigacker S, Xia X, He Z. Development and validation of KASP assays for genes underpinning key economic traits in bread wheat. Theor Appl Genet. 2016 doi: 10.1007/s00122-016-2743-x. [DOI] [PubMed] [Google Scholar]
- Ren Y, Li Z, He Z, Wu L, Bai B, Lan C, Wang C, Zhou G, Zhu H, Xia X. QTL mapping of adult-plant resistances to stripe rust and leaf rust in Chinese wheat cultivar Bainong 64. Theor Appl Genet. 2012 doi: 10.1007/s00122-012-1910-y. [DOI] [PubMed] [Google Scholar]
- Rosewarne GM, Singh RP, Huerta-Espino J, Herrera-Foessel SA, Forrest KL, Hayden MJ, Rebetzke GJ. Analysis of leaf and stripe rust severities reveals pathotype changes and multiple minor QTLs associated with resistance in an Avocet × Pastor wheat population. Theor Appl Genet. 2012 doi: 10.1007/s00122-012-1786-x. [DOI] [PubMed] [Google Scholar]
- Sacco F, Suárez EY, Naranjo T. Mapping of the leaf rust resistance gene Lr3 on chromosome 6B of Sinvalocho MA wheat. Genome. 1998 doi: 10.1139/g98-067. [DOI] [Google Scholar]
- Sallam A, Martsch R. Association mapping for frost tolerance using multi-parent advanced generation inter-cross (MAGIC) population in faba bean (Vicia faba L.) Genetica. 2015 doi: 10.1007/s10709-015-9848-z. [DOI] [PubMed] [Google Scholar]
- Sannemann W, Huang BE, Mathew B, Léon J. Multi-parent advanced generation inter-cross in barley: high-resolution quantitative trait locus mapping for flowering time as a proof of concept. Mol Breed. 2015 doi: 10.1007/s11032-015-0284-7. [DOI] [Google Scholar]
- Sannemann W, Lisker A, Maurer A, Léon J, Kazman E, Cöster H, Holzapfel J, Kempf H, Korzun V, Ebmeyer E, Pillen K. Adaptive selection of founder segments and epistatic control of plant height in the MAGIC winter wheat population WM-800. BMC Genomics. 2018 doi: 10.1186/s12864-018-4915-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Serfling A, Krämer I, Perovic D, Ordon F. Erweiterung der genetischen Basis der Resistenzdes Weizens (Triticum aestivum) gegen Braunrost (Puccinia triticina f. sp. tritici) Journal für Kulturpflanzen. 2013 doi: 10.5073/JFK.2013.07.02. [DOI] [Google Scholar]
- Sharma C, Saripalli G, Kumar S, Gautam T, Kumar A, Rani S, Jain N, Prasad P, Raghuvanshi S, Jain M, Sharma JB, Prabhu KV, Sharma PK, Balyan HS, Gupta PK. A study of transcriptome in leaf rust infected bread wheat involving seedling resistance gene Lr28. Funct Plant Biol. 2018 doi: 10.1071/FP17326. [DOI] [PubMed] [Google Scholar]
- Stadlmeier M, Hartl L, Mohler V. Usefulness of a multiparent advanced generation intercross population with a greatly reduced mating design for genetic studies in winter wheat. Assessment of resistance in cereal cultivars Design and analysis of experiments using the SAS-application RESI 2. Front Plant Sci. 2018 doi: 10.3389/fpls.2018.01825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stadlmeier M, Jørgensen LN, Corsi B, Cockram J, Hartl L, Mohler V. Genetic dissection of resistance to the three fungal plant pathogens Blumeria graminis, Zymoseptoria tritici, and Pyrenophora tritici-repentis using a multiparental winter wheat population. G3 (Bethesda) 2019 doi: 10.1534/g3.119.400068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsilo TJ, Kolmer JA, Anderson JA. Molecular mapping and improvement of leaf rust resistance in wheat breeding lines. Phytopathology. 2014 doi: 10.1094/PHYTO-10-13-0276-R. [DOI] [PubMed] [Google Scholar]
- Turner MK, Kolmer JA, Pumphrey MO, Bulli P, Chao S, Anderson JA. Association mapping of leaf rust resistance loci in a spring wheat core collection. Theor Appl Genet. 2017 doi: 10.1007/s00122-016-2815-y. [DOI] [PubMed] [Google Scholar]
- Wang H, Zou S, Li Y, Lin F, Tang D. An ankyrin-repeat and WRKY-domain-containing immune receptor confers stripe rust resistance in wheat. Nat Commun. 2020 doi: 10.1038/s41467-020-15139-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- William HM, Singh RP, Huerta-Espino J, Palacios G, Suenaga K. Characterization of genetic loci conferring adult plant resistance to leaf rust and stripe rust in spring wheat. Genome. 2006 doi: 10.1139/g06-052. [DOI] [PubMed] [Google Scholar]
- Yu J, Holland JB, McMullen MD, Buckler ES. Genetic design and statistical power of nested association mapping in maize. Genetics. 2008 doi: 10.1534/genetics.107.074245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zetzsche H, Serfling A, Ordon F. Breeding progress in seedling resistance against various races of stripe and leaf rust in european bread wheat. Crop Breed Genet Genom. 2019 doi: 10.20900/cbgg20190021. [DOI] [Google Scholar]
- Zhang D, Bowden RL, Yu J, Carver BF, Bai G. Association analysis of stem rust resistance in U.S. winter wheat. PLoS ONE. 2014 doi: 10.1371/journal.pone.0103747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang P, Yin G, Zhou Y, Qi A, Gao F, Xia X, He Z, Li Z, Liu D. QTL mapping of adult-plant resistance to leaf rust in the wheat cross Zhou 8425B/Chinese Spring using high-density SNP markers. Front Plant Sci. 2017 doi: 10.3389/fpls.2017.00793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang P, Lan C, Asad MA, Gebrewahid TW, Xia X, He Z, Li Z, Liu D. QTL mapping of adult-plant resistance to leaf rust in the Chinese landraces Pingyuan 50/Mingxian 169 using the wheat 55K SNP array. Mol Breed. 2019 doi: 10.1007/s11032-019-1004-5. [DOI] [Google Scholar]
- Zhou Y, Ren Y, Lillemo M, Yao Z, Zhang P, Xia X, He Z, Li Z, Liu D. QTL mapping of adult-plant resistance to leaf rust in a RIL population derived from a cross of wheat cultivars Shanghai 3/Catbird and Naxos. Theor Appl Genet. 2014 doi: 10.1007/s00122-014-2346-3. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Fig. S1 Pearson correlation of leaf rust severity between different field trials. Diagonals are histograms of each environment (Lengern LEN 2018, Quedlinburg QLB 2017 2018, Söllingen SOE 2017 2018). *** denotes significance at α = 0.001. Lowess curves were adjusted to the data points with a smoothing range of 0.75, based on the ‘lowess’ function implemented in the R-based ‘stats’ package (JPEG 133 kb)
Fig. S2 Pearson correlation (r) between averaged infection type (IT), infected leaf area (Pi) of seedling test and average ordinate (AO) of field trials (A, B), as well as correlation between IT and Pi (C). *** denotes significance at α = 0.001 (JPEG 78 kb)
Fig. S3 Simple interval mapping of resistance to Puccinia triticina in field trials (A) and seedling test (B, C). The x axis shows the 21 wheat chromosomes. Positions are based on the genetic map, and the log10(p) values of each Marker are displayed on the y axis (black line). The red horizontal line represents the significance thresholds. SI of the significant QTL detected in this study are coloured in blue (JPEG 188 kb)
Tab. S1 List of virulences and avirulences of Puccinia triticina isolate 77WxR used in field trials and seedling test. Brackets indicate ambiguous results due to the differing symptom ratings between replications or moderate susceptibility (based on Zetzsche et al. 2019) (DOCX 13 kb)
Tab. S2 Complete information of the 19 QTL for leaf rust resistance in BMW population, evaluated in field trials (AO) and seedling test (IT and Pi) (XLSX 16 kb)