

Abstract
Gut microorganisms are vital for many aspects of human health, and the commensal bacterium Akkermansia muciniphila has repeatedly been identified as a key component of intestinal microbiota. Reductions in A. muciniphila abundance are associated with increased prevalence of metabolic disorders such as obesity and type 2 diabetes. It was recently discovered that administration of A. muciniphila has beneficial effects and that these are not diminished, but rather enhanced after pasteurization. Pasteurized A. muciniphila is proposed for use as a food ingredient, and was therefore subjected to a nonclinical safety assessment, comprising genotoxicity assays (bacterial reverse mutation and in vitro mammalian cell micronucleus tests) and a 90‐day toxicity study. For the latter, Han Wistar rats were administered with the vehicle or pasteurized A. muciniphila at doses of 75, 375 or 1500 mg/kg body weight/day (equivalent to 4.8 × 109, 2.4 × 1010, or 9.6 × 1010 A. muciniphila cells/kg body weight/day) by oral gavage for 90 consecutive days. The study assessed potential effects on clinical observations (including detailed arena observations and a modified Irwin test), body weight, food and water consumption, clinical pathology, organ weights, and macroscopic and microscopic pathology. The results of both in vitro genotoxicity studies were negative. No test item‐related adverse effects were observed in the 90‐day study; therefore, 1500 mg/kg body weight/day (the highest dose tested, equivalent to 9.6 × 1010 A. muciniphila cells/kg body weight/day) was established as the no‐observed‐adverse‐effect‐level. These results support that pasteurized A. muciniphila is safe for use as a food ingredient.
Keywords: Akkermansia muciniphila; beneficial microorganism; food ingredient; gut microbiota; safety; subchronic toxicity, genotoxicity
Short abstract
Pasteurized A. muciniphila is proposed for use as a food ingredient and was therefore subjected to in vitro genotoxicity tests and a 90‐day oral (gavage) toxicity study in rats. There was no evidence of genotoxicity, and the highest dose tested in the 90‐day study (1500 mg/kg body weight/day, equivalent to 9.6 × 1010 A. muciniphila cells/kg body weight/day) was established as the no‐observed‐adverse‐effect‐level. These results support the safety of pasteurized A. muciniphila for use as a food ingredient.
Abbreviations
- AAALAC
Association for Assessment and Accreditation of Laboratory Animal Care
- EFSA
European Food Safety Authority
- FDA
Food and Drug Administration
- NOAEL
no‐observed‐adverse‐effect level
- OECD
Organization for Economic Co‐operation and Development
- T3
triiodothyronine
- T4
thyroxine
1. INTRODUCTION
Akkermansia muciniphila is an elliptical‐shaped human gut commensal bacterium that is nonmotile and does not form spores. It was isolated from the human gastrointestinal tract for the first time by Derrien and colleagues in 2004 (Derrien, Vaughan, Plugge, & de Vos, 2004). The relative abundance of A. muciniphila found in fecal samples derived from healthy humans ranges from 0.5% to 5% (Collado, Derrien, Isolauri, de Vos, & Salminen, 2007; Derrien, Collado, Ben‐Amor, Salminen, & de Vos, 2008). Primarily considered to reside in the large intestine, data suggest that the small intestine can also be colonized by several A. muciniphila cells (Geerlings, Kostopoulos, de Vos, & Belzer, 2018). A. muciniphila is present in the human gut from infancy (as young as 1 month old), and its abundance increases significantly during growth into adulthood. A. muciniphila cells have been detected in human breast milk (Collado, Laitinen, Salminen, & Isolauri, 2012), thus demonstrating A. muciniphila consumption from birth through infant consumption of breast milk. The capacity of A. muciniphila to use human milk oligosaccharides as a sole energy, carbon and nitrogen source, could explain its presence in breast milk and the breast tissue of lactating woman (Collado et al., 2012; Ottman, 2017). Human milk may act as a carrier for A. muciniphila in the transfer from mother to infant, explaining the presence of A. muciniphila in the infant intestine already early in life. Furthermore, another characteristic of A. muciniphila can also facilitate this transfer from mother to infant. Indeed, A. muciniphila has an active acid resistance system (Ottman, 2017). This system, together with the mild acidic conditions in the infant stomach during early life, could help A. muciniphila to reach and colonize the intestine in early life.
Recently, combined analysis of large global datasets (3984 subjects) revealed that the genus Akkermansia was present in 77.73% of the total assessed cohorts globally and in 81.81% of cohorts targeting Western populations (Falony et al., 2016). In a study comprising 249 subjects across various age groups, A. muciniphila was detected in eight of 50 fecal samples of 1‐month‐old infants (16%), in 36 of 50 children aged 6 months (72%), in 45 of 50 children at 12 months of age (90%), and in 96 to 100% of samples from adults and the elderly (Collado et al., 2007).
Inspired by the numerous observations that the relative levels of A. muciniphila decreased during obesity and metabolic disorders in mice and humans (for review: Cani & de Vos, 2017; Derrien, Belzer, & de Vos, 2017; Xu et al., 2020), further studies were conducted to investigate the causal link between A. muciniphila and improvements in metabolism. It was found that the administration of live A. muciniphila partly protected against diet‐induced obesity, diabetes and gut barrier dysfunction in mice (Everard et al., 2013). Indeed, the oral administration of live A. muciniphila was able to counteract the body weight gain, fat mass gain and glucose intolerance induced by high‐fat feeding in murine models. In addition, daily supplementation with live A. muciniphila was associated with a decrease of plasmatic level of lipopolysaccharides and an increase of mucus thickness and expression of antimicrobial peptide mRNA, which showed an overall improvement of the gut barrier function; a function usually altered in diet‐induced obesity mice models. Another key finding of this study was that the protective effects of A. muciniphila disappeared when the bacterium was heat‐killed by autoclaving (15 minutes at 121°C), which is a heating procedure that destroyed all constituents of the bacteria (Everard et al., 2013). As A. muciniphila is a gram‐negative bacterium, and hence a nonspore former, research was conducted to elucidate whether the removal of the protective effects of A. muciniphila observed after autoclaving would also be seen after pasteurization (30 minutes at 70°C), which is a milder heat‐inactivation method compared with autoclaving. In a study assessing the effects of pasteurized A. muciniphila on diet‐induced metabolic disorders in mice, not only was it found that pasteurization did not diminish the beneficial effects, but it unexpectedly enhanced the beneficial impact of A. muciniphila (Plovier et al., 2005). Since this discovery, the beneficial effects of pasteurized A. muciniphila in the prevention of metabolic disorders associated with obesity have been confirmed in mice (Depommier et al., 2020) and in humans (Depommier et al., 2019). Depommier et al. (2020) confirmed that daily administration of pasteurized A. muciniphila alleviates diet‐induced obesity in mice. Furthermore, Depommier et al. (2020) explored the mechanism of action underlaying the observed beneficial effects more thoroughly and found that administration of pasteurized A. muciniphila is able to impact the whole‐body energy metabolism by increasing energy expenditure, spontaneous physical activity and fecal energy excretion in a diet‐induced obesity mice model. A randomized, double‐blind, placebo‐controlled proof‐of‐concept study in overweight/obese insulin‐resistant human volunteers showed that the daily supplementation with pasteurized A. muciniphila at a dose of 1 × 1010 cells per day for 12 weeks improved several metabolic parameters such as insulin sensitivity, insulinemia, plasma total cholesterol, as well as relevant blood markers for liver dysfunction and inflammation independently of any caloric restriction or modification of the physical activity (Depommier et al., 2019). Recently, the beneficial effects of pasteurized A. muciniphila were also demonstrated in the context of colitis and colitis‐associated colorectal cancer (Wang et al., 2019). Oral administration of pasteurized A. muciniphila from 2 weeks before dextran sulfate sodium treatment and until death was associated with a reduction of disease severity (reduction of colon shortening and splenomegaly; attenuation of histological injuries in the colon). Furthermore, in a model of colitis‐associated colorectal cancer, the oral administration of pasteurized A. muciniphila was able to delay the tumor formation and decrease the tumor numbers and size. These results suggest that this decreased tumorigenesis could be mediated by an attenuation of DNA damage, cell apoptosis and abnormal proliferation.
Pasteurized A. muciniphila has now been developed for use as a food ingredient. As A. muciniphila is a highly prevalent intestinal microbe colonizing virtually all healthy subjects studied and is detected in breast milk, it can be argued that it has some history of consumption (Gómez‐Gallego, Pohl, Salminen, De Vos, & Kneifel, 2016). However, pasteurized A. muciniphila (produced by fermentation, pasteurization and freeze‐drying) is considered a novel food and therefore requires a thorough safety assessment before being approved as a food ingredient.
Although detected in breast milk, A. muciniphila is not absorbed into the systemic circulation to any significant extent, which is similar to other nonabsorbable compounds found in breast milk, such as human milk oligosaccharides, which are considered as nondigestible carbohydrates (EFSA, 2016). A. muciniphila is almost exclusively excreted in feces following oral administration. Preclinical and clinical studies have shown that A. muciniphila supplementation does not induce major changes in the overall gut microbiome (Depommier et al., 2019; Everard et al., 2013). The detection of A. muciniphila in fecal samples in these preclinical and clinical studies demonstrates that it is excreted in feces.
Pasteurization is not expected to change the profile of A. muciniphila; thus, pasteurized A. muciniphila is also expected to be nonabsorbable. Following the US Food and Drug Administration (FDA) and European Food Safety Authority (EFSA) guidances on toxicity testing required for safety assessment of a new nonabsorbable food ingredient (EFSA, 2012; US FDA Redbook, 2014), pasteurized A. muciniphila was subjected to nonclinical studies assessing the potential for genotoxicity (bacterial reverse mutation and in vitro micronucleus testa on mammalian cells) and subchronic toxicity (90‐day oral toxicity study in rats). The objective of this research article is to describe the methodology and results for these nonclinical studies conducted with pasteurized A. muciniphila, as part of a comprehensive assessment to demonstrate its safety for use as a food ingredient.
2. MATERIALS AND METHODS
2.1. Regulatory guidelines and quality assurance
The studies were performed at two of Citoxlab's Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC)‐accredited facilities (genotoxicity studies conducted at Citoxlab France and 90‐day study conducted at Citoxlab Denmark) and inspected by Quality Assurance Units in compliance with the Organization for Economic Co‐operation and Development (OECD) Principles of Good Laboratory Practice (OECD, 1998). All studies were conducted following OECD guidelines: the 90‐day oral gavage toxicity study was conducted using test methods described in OECD Test Guideline No. 408 (OECD, 2018), the bacterial reverse mutation test was conducted according to OECD Test Guideline No. 471 (OECD, 1997), and the in vitro micronucleus test on mammalian cells was conducted according to OECD Test Guideline No. 487 (OECD, 2016). The housing of the animals was in accordance with EU Directive 2010/63/EU of September 22, 2010 on the protection of animals used for scientific purposes (European Union, 2010).
2.2. Test materials
Pasteurized A. muciniphila (strain MucT [ATCC BAA‐835]) was supplied by A‐Mansia Biotech SA as an off‐white to beige homogeneous powder and contained at least 2.5 × 1010 A. muciniphila cells per gram of powder; viable cells represent ≤0.0001% of this total cell count. The vehicle for the 90‐day study (sterile phosphate‐buffered saline) was supplied by Sigma‐Aldrich. Water for injection (CDM Lavoisier) served as the vehicle and negative control for both genotoxicity studies. Positive controls used in the bacterial reverse mutation test were supplied by Sigma‐Aldrich (sodium azide, 9‐aminoacridine, 2‐anthramine, benzo[a]pyrene and 2‐nitrofluorene) and by Acros Organics BVBA (mitomycin C) and all except for mitomycin C (dissolved in water) were dissolved in dimethyl sulfoxide (Merck KGaA) before use. The five strains of Salmonella typhimurium TA1535, TA1537, TA98, TA100 and TA102 used in the bacterial reverse mutation test were supplied by Moltox or culture collections (Public Health England). Positive controls used in the in vitro micronucleus test on mammalian cells were supplied by Sigma‐Aldrich (colchicine and cyclophosphamide) and by Acros Organics BVBA (mitomycin C), and dissolved in water before use. Liver homogenate used to replicate metabolic activation (S9 mix) was supplied by Moltox and obtained from the liver of rats previously administered with 500 mg/kg Aroclor 1254 by intraperitoneal injection.
2.3. Genotoxicity
2.3.1. Bacterial reverse mutation test (Ames test)
The bacterial reverse mutation test was conducted using the plate incorporation and preincubation methods, as has been previously described (Ames, McCann, & Yamasaki, 1975; Maron & Ames, 1983). S. typhimurium strains TA98, TA100, TA102, TA1535 and TA1537 were exposed to pasteurized A. muciniphila at concentrations of up to the OECD Test Guideline No. 471 (OECD, 1997) maximum recommended concentration of 5000 μg/plate, in the absence or presence of external metabolic activation (S9 mix). The only exception was for the TA98 strain in the presence of the S9 mix, where the high dose was limited by cytotoxicity (highest tested concentration of 555.6 μg/plate using the direct plate incorporation method and 1666.7 μg/plate following the preincubation method).
For the direct plate incorporation method, 0.1 mL suspensions of pasteurized A. muciniphila were mixed with 0.5 mL of S9 mix or pH 7.4 phosphate buffer (depending on whether metabolic activation was required), 0.1 mL of a suspension of each bacterial strain and 2 mL of overlay agar containing Agar (BD Biosciences), NaCl (CDM Lavoisier), D(+) Biotine (Sigma‐Aldrich) and l‐Histidine (Sigma‐Aldrich). After rapid homogenization (a few minutes by vortex), the mixture was overlaid on to a Petri dish containing minimum medium (Trinova Biochem GmbH). The same quantities of test item suspensions (0.1 mL), S9 mix (0.5 mL) and bacterial suspensions (0.1 mL) were used for the preincubation method, but the mixture was then preincubated for 60 minutes at 37°C (under shaking) before overlay agar was added and the mixture was poured on to the surface of a minimum agar plate. For both methods, after 48‐72 hours of incubation at 37°C, the number of revertants per plate were scored for each strain and for each experimental point using an automatic counter (Sorcerer Automatic Colony Counter [Perceptive Instruments Ltd.] for the scoring of colonies and Ames Study Manager [Perceptive Instruments Ltd.] for the data management). Manual counting was used when required. Thinning of the bacterial lawn and the presence of precipitate were assessed visually with the naked eye.
A positive result for mutagenicity was defined as a reproducible two‐ or three‐fold increase in the number of revertant colonies, compared with vehicle controls, and/or a reproducible dose‐response relationship.
2.3.2. In vitro mammalian cell micronucleus test
The in vitro mammalian cell micronucleus test was conducted using the cytokinesis‐block method (Fenech, 1993; Fenech & Morley, 1985). For each experiment, cell cultures were prepared from whole blood samples (obtained from young, 18‐35‐year‐old, healthy, nonsmoking donors), which were added to 8 mL of culture medium containing phytohemagglutinin (a mitogen to stimulate the lymphocytes to divide; supplied by Oxoid). The cultures were incubated at 37°C for 44‐48 hours, before being centrifuged at 300 g for 10 minutes. Supernatants were discarded and cells were resuspended in fresh culture medium (supplemented with S9 mix when metabolic activation was required) and exposed to pasteurized A. muciniphila (suspended in water for injections) for 3 hours (with 24 hours recovery) in the absence or presence of S9 mix, and for 24 hours in the absence of S9 mix (without recovery). Cytochalasin B (dissolved in dimethyl sulfoxide) was added in to each culture to block cytokinesis. The concentrations selected for micronucleus analysis were 187.5, 375 and 750 μg/mL for the 3‐hour exposure period and 93.75, 187.5 and 375 μg/mL for the 24‐hour exposure periods (the selected high concentrations were the lowest concentrations showing precipitate in the culture medium at the end of the exposure periods, in accordance with OECD Test Guideline No. 487).
At harvest, the cells were collected by centrifugation (300 g for 10 minutes) and subjected to a hypotonic treatment to induce cell swelling (i.e., incubation of 3 minutes in 4 mL of 0.075 m potassium chloride prewarmed at 37°C). The cells were then fixed in a methanol/acetic acid mixture prepared by Citoxlab (both reagents were supplied by Carlo Erba Reagents S.A.S.). Following fixation, the cells were stored by refrigeration at 4°C for at least an overnight period, before undergoing two rounds of centrifugation (1250 g for 3 minutes) and removal of the supernatant. The resultant cells were spread on glass slides and stained for 7 minutes with 5% Giemsa (Merck KGaA) in water. The slides were coded to allow for blind scoring. Micronuclei were analyzed in 1000 binucleated cells per culture when possible (in total, 2000 binucleated cells per concentration level in all conditions). For statistical analysis, cell cultures exposed to the test item were compared with that of the vehicle control cell cultures using the χ2 test, in which P = .05 was used as the lowest level of significance. To assess the dose‐response trend, a linear regression was performed between the mean frequencies of micronucleated binucleated cells and the concentrations. This statistical analysis was performed using SAS (version 9.2).
A positive result for clastogenicity or aneugenicity was defined as a statistically significant increase in the frequency of micronucleated binucleated cells in comparison with the corresponding vehicle controls, obtained at one or more dose levels, associated with a dose‐response relationship, and for at least one concentration, the frequency of micronucleated binucleated cells of each replicate culture is above the corresponding vehicle historical range.
2.4. Subchronic toxicity
2.4.1. Animals and housing conditions
In total, 40 male and 40 female specific pathogen‐free Crl:WI (Han) rats were supplied by Charles River for the study. The animals were 5‐6 weeks old at the start of the acclimatization period (lasting 12 days), with males weighing 159‐191 g and females weighing 83‐110 g. The study took place in an animal room supplied with filtered air (ventilated to give 10 air changes per hour) and maintained at a temperature of 18‐24°C and relative humidity of 40%‐60%. A 12‐hour light/dark cycle was controlled automatically. The rats were kept in transparent polycarbonate cages (floor area: 1500 cm2 and height: 21 cm) with two or three animals per cage, and males and females were separated. The animals were provided with Aspen bedding 2HV (Tapvei Estonia OÜ), as well as environmental enrichment (Aspen Wood Wool; Tapvei Estonia OÜ), a wooden block (both supplied by Tapvei Estonia OÜ) and a red transparent Rat House (Tecniplast Gazzada S.a.r.l.). Pelleted rodent diet was available ad libitum; Altromin 1314 Fortified diet (Altromin Spezialfutter GmbH & Co. KG) was provided until day 49 of the dosing period, after which the diet was switched to Altromin 1324 Fortified diet (Altromin Spezialfutter GmbH & Co. KG) for adult animals. Domestic quality drinking water was freely available throughout the study.
2.4.2. Test item formulation, administration and dosing schedule
The test item was initially divided into vials, stored under nitrogen (to expel air and avoid oxidation) and frozen (−18°C or below) until use. On each day of dosing, the required number of vials were defrosted and the vehicle (sterile phosphate‐buffered saline previously diluted 1:9 with water) was added, before the vials were mixed with a vortex. The resultant suspensions were administered by oral gavage to groups of 10 males and 10 females at 75, 375 or 1500 mg/kg body weight/day (equivalent to 4.8 × 109, 2.4 × 1010 or 9.6 × 1010 A. muciniphila cells/kg body weight/day) at a dose volume of 5 mL/kg body weight, once daily for 90 (males) or 91 (females) days, until the day before necropsy. Animals in the vehicle control group (also comprising 10 males and 10 females) received the vehicle under the same conditions as animals in test item groups. The high dose of 1500 mg/kg body weight/day (equivalent to 9.6 × 1010 A. muciniphila cells/kg body weight/day) was selected to provide a margin of safety of at least 100‐fold compared with worst‐case exposure in foods; four‐ to five‐fold descending intervals selected for the mid and low doses, respectively, to demonstrate any dose‐related response in effects.
All animals were dosed within 4 hours of formulation preparation, which was within the stability period (4 hours when stored at room temperature protected from light and 6 hours when stored refrigerated) confirmed as part of a separate validation study (data not shown). A. muciniphila cell counts (conducted using flow cytometry) were within expected ranges for test item formulations prepared for use in weeks 1 and 13, and no A. muciniphila cells were detected in vehicle control formulations (data not shown), which demonstrates that formulations were accurately prepared.
2.4.3. Clinical examinations, body weights, and food and water consumption
All animals were observed at least once daily for changes in clinical condition, with standard arena observations that were more comprehensive and conducted once weekly. Once before the first dose administration (for all animals) and once in week 13 (for vehicle controls and high‐dose animals only) both eyes of each animal were examined with an indirect ophthalmoscope and a portable slit‐lamp microscope after application of tropicamide 1% solution (Mydriacyl; supplied by Alcon Universal Ltd.). On days 86 and 87 of dosing, all animals were subjected to a modified Irwin test. The Irwin test included a functional observation battery with assessment of sensory reactivity to different stimuli, grip strength and motor activity. The body weight of each animal was recorded on the first day of dosing and weekly thereafter, until necropsy. Food and water consumption were recorded twice weekly throughout the dosing period.
2.4.4. Hematology, coagulation and clinical chemistry
At the end of the dosing period (just before necropsy), blood samples were collected from all surviving animals for hematology, coagulation and clinical chemistry assessments. For hematology, 400 μL of blood was collected into K3 ethylenediaminetetraacetic acid tubes (Sarstedt AG & Co.) for analysis of the following parameters using the ABX Pentra DX120SPS (Horiba): hemoglobin, red blood cell count, reticulocyte count, hematocrit, mean cell volume, mean cell hemoglobin, mean cell hemoglobin concentration, white blood cell count, neutrophils, lymphocytes, eosinophils, basophils, monocytes and platelet count. Values for coagulation parameters (activated partial thromboplastin time, prothrombin time and fibrinogen) were determined (from 500 μL blood samples collected into tubes; Eppendorf AG) containing 50 μL citrate solution (129 mmol/L trisodium citrate prepared from trisodium citrate dihydrate 294.1 g/mol; Merck KGaA) using an ACL 9000 (Instrumentation Laboratories). For clinical chemistry, 750 μL of blood was collected into plain glass tubes for serum; values for alanine aminotransferase, aspartate aminotransferase, alkaline phosphatase, bilirubin, gamma‐glutamyl transferase, cholesterol (total, high‐density lipoprotein and low‐density lipoprotein), triglycerides, urea, creatinine, glucose, sodium, potassium, calcium, magnesium, inorganic phosphorus, chloride, total protein, albumin and albumin/globulin ratio were determined using a Cobas 6000 (Roche), values for triiodothyronine (T3) and thyroxine (T4) by liquid chromatography with tandem mass spectrometry and values for thyroid‐stimulating hormone using xMAP technology (Luminex; Bio‐Rad Laboratories).
2.4.5. Macroscopic examinations, organ weights and histopathology
On the day of necropsy (days 91 and 92 for males and females, respectively), all surviving animals were weighed, examined externally and (for all females) the stage of the estrus cycle was recorded by wet vaginal smears, before being killed by exposure to a steadily increasing concentration of CO2. A macroscopic examination was performed by opening the cranial, thoracic and abdominal cavities and observing the appearance of organs and tissues in situ. For animals in all groups, specified organs (adrenal glands, brain, epididymides, heart, kidney, liver, ovary, pituitary gland, prostate, spleen, testes, thymus and uterus) were weighed. A full list of organs and tissues (aorta, adrenal gland, bone marrow, bone from the femur, brain, caecum, colon, duodenum, epididymis, eyes, femorotibial joint, heart, ileum, jejunum, kidney, larynx, liver, lungs, lymph nodes [mandibular and mesenteric], mammary gland, esophagus, optic and sciatic nerves, ovaries, pancreas, parathyroid, pituitary, rectum, salivary glands [parotid, sublingual and submandibular], seminal vesicles, skeletal muscle, spinal cord [cervical, thoracic and lumbar], skin, spleen, stomach, testes, thymus, thyroid, tongue, trachea, ureter, urinary bladder, uterus and vagina) was examined microscopically for animals in the vehicle control and high‐dose groups only. All tissues were fixed in phosphate‐buffered neutral 4% formaldehyde (Hounissen), except for the eyes and testes, which were fixed in Modified Davidsons's fixative (prepared by Citoxlab from water, ethanol, formaldehyde and acetic acid).
After fixation, the organs and tissues from vehicle control and high‐dose animals were trimmed and representative specimens were taken for histological processing. The specimens were embedded in paraffin wax and cut at a nominal thickness of 5 μm, before being stained with hematoxylin and eosin and examined under a light microscope (Leica DM3000). All pathological findings were entered directly into Instem Provantis (version 9.3.0.0). Histological findings were graded on a five‐level scale (minimal, mild, moderate, marked and severe).
2.4.6. Statistical analysis
Data were processed to give group mean values and standard deviations where appropriate. Thereafter, each continuous variable was tested for homogeneity of variance with Levene's test. If the variance was homogeneous, analysis of variance was carried out for the variable. If any significant differences were detected, possible intergroup differences were assessed with Dunnett's test (comparing test item groups with a control group). If the variance was heterogeneous, each variable was tested for normality by the Shapiro‐Wilk method. In case of normal distribution, possible intergroup differences were identified with Student's t‐test. Otherwise, the possible intergroup differences were assessed by the Kruskal‐Wallis test. If any significant intergroup differences were detected, the subsequent identification of the groups were carried out with Wilcoxon rank‐sum test. Ranked type of data were analyzed with the Kruskal‐Wallis test. If any significant intergroup differences were detected, the subsequent identification of the groups were assessed with Wilcoxon rank‐sum test. For all tests, the level of significance was P < .05. Statistical analyses were conducted using SAS (version 9.3) and SAS/STAT (version 12.1).
3. RESULTS
3.1. Genotoxicity
3.1.1. Bacterial reverse mutation test (Ames test)
Mean revertant colony counts are provided in Table 1. An increase in the mean number of revertants was observed at 5000 μg/plate after short‐term exposure in the absence of S9 for the TA98 strain (experiment 2), and the mean value (41.3) was slightly outside the historical control range (10‐40); however, there was no evidence of a dose‐response. As this increase was not observed in the first experiment, a third experiment was conducted using TA98 only (under the same experimental conditions) to elucidate whether this increase was reproducible. In this repeat experiment, no noteworthy increases in the number of revertants was reported, demonstrating that the increase in the second experiment was not reproducible and was therefore biologically irrelevant. The overall results did not to show any biologically relevant increases in the mean number of revertant colonies following exposure to pasteurized A. muciniphila in any of the tested strains compared with corresponding vehicle controls, in the presence or absence of metabolic activation. The positive controls induced increases in mean revertant colony numbers of at least twice (or three times in the case of TA1535 and TA1537 strains) that of the concurrent vehicle controls (with metabolic activation where required), which demonstrated the sensitivity of the assay and metabolic activity of the S9 mix. The mean revertant colony counts for the vehicle controls were within the historical control ranges of the laboratory and there were at least five analyzable concentrations for each strain and test condition. These results confirmed the validity of the study and all criteria for a negative response were met.
TABLE 1.
Bacterial reverse mutation test with pasteurized A. muciniphila
| Concentration (μg/plate) | Revertant colonies per plate (mean ± standard deviation) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Without metabolic activation (–S9) | With metabolic activation (+S9) | |||||||||
| S. typhimurium | S. typhimurium | |||||||||
| TA98 | TA100 | TA102 | TA1535 | TA1537 | TA98 | TA100 | TA102 | TA1535 | TA1537 | |
| Plate incorporation assay (experiment 1) | ||||||||||
| 0 (water) | 14.3 ± 2.5 | 130.7 ± 15.6 | 455.7 ± 31.8 | 9.0 ± 3.6 | 7.7 ± 1.5 | 24.3 ± 5.5 | 114.3 ± 9.1 | 520.3 ± 56.1 | 11.0 ± 4.6 | 8.3 ± 3.8 |
| 2.29 | – | – | – | – | – | 20.0 ± 2.6 | – | – | – | – |
| 6.86 | – | – | – | – | – | 31.0 ± 5.0 | – | – | – | – |
| 20.6 | 16.3 ± 3.8 | 129.3 ± 11.7 | 501.0 ± 39.0 | 17.0 ± 0.0 | 4.0 ± 1.0 | 30.3 ± 8.1 | 116.0 ± 23.6 | 482.3 ± 34.8 | 11.0 ± 4.6 | 6.7 ± 2.3 |
| 61.7 | 17.3 ± 4.6 | 131.0 ± 14.7 | 396.3 ± 162.3 | 12.7 ± 3.2 | 6.3 ± 1.5 | 18.7 ± 4.5 | 115.3 ± 17.0 | 558.3 ± 60.7 | 12.3 ± 4.2 | 6.7 ± 2.3 |
| 185.2 | 13.0 ± 1.0 | 128.3 ± 17.2 | 354.0 ± 27.0 | 17.3 ± 2.1 | 10.0 ± 1.0 | 22.7 ± 2.3 | 117.7 ± 11.7 | 530.0 ± 25.4 | 10.3 ± 2.1 | 7.7 ± 3.5 |
| 555.6 | 19.0 ± 5.6 | 131.7 ± 11.1 | 479.3 ± 73.9 | 13.0 ± 5.0 | 9.3 ± 0.6 | 24.7 ± 4.0 | 132.3 ± 3.1 | 468.3 ± 60.3 | 13.3 ± 1.2 | 7.7 ± 2.3 |
| 1666.7 | 18.3 ± 4.0 | 119.0 ± 10.0 | 386.7 ± 46.3 | 14.7 ± 0.6 | 5.3 ± 4.0 | – | 118.0 ± 12.1 | 513.3 ± 28.9 | 12.3 ± 5.0 | 12.0 ± 7.2 |
| 5000 | 20.7 ± 2.1 | 169.0 ± 4.4 | 397.0 ± 75.4 | 17.7 ± 5.0 | 8.7 ± 0.6 | – | 153.0 ± 24.2 | 500.7 ± 60.0 | 16.0 ± 4.6 | 9.7 ± 3.5 |
| Positive control a | 109.3 ± 7.6 | 693.3 ± 27.3 | 1215.3 ± 77.9 | 607.3 ± 60.0 | 83.0 ± 11.3 | 1071.3 ± 30.7 | 913.3 ± 27.1 | 1060.7 ± 30.2 | 113.0 ± 25.5 | 93.0 ± 3.6 |
| Plate incorporation assay (experiment 2) | ||||||||||
| 0 (water) | 18.7 ± 4.5 | 101.0 ± 8.7 | 458.3 ± 34.6 | 6.0 ± 2.6 | 6.3 ± 1.2 | – | – | – | – | – |
| 61.7 | 18.0 ± 3.5 | 95.0 ± 6.0 | 444.3 ± 24.3 | 7.0 ± 2.6 | 8.7 ± 3.1 | – | ||||
| 185.2 | 18.3 ± 9.5 | 122.0 ± 15.4 | 439.0 ± 36.5 | 12.3 ± 3.2 | 6.3 ± 4.5 | – | – | – | – | – |
| 555.6 | 17.7 ± 2.9 | 117.7 ± 25.0 | 510.0 ± 38.0 | 12.3 ± 1.2 | 9.7 ± 2.5 | – | – | – | – | – |
| 1666.7 | 15.7 ± 2.9 | 142.0 ± 25.1 | 453.0 ± 20.9 | 13.7 ± 6.4 | 7.3 ± 3.1 | – | – | – | – | – |
| 5000 | 41.3 ± 4.7 | 136.7 ± 7.5 | 543.0 ± 80.9 | 11.3 ± 2.3 | 10.0 ± 1.7 | – | – | – | – | – |
| Positive control b | 95.7 ± 11.7 | 623.0 ± 34.4 | 1245.7 ± 78.8 | 677.3 ± 28.7 | 160.0 ± 60.9 | – | – | – | – | – |
| Plate incorporation assay (experiment 3) | ||||||||||
| 0 (water) | 14.3 ± 5.1 | – | – | – | – | – | – | – | – | – |
| 312.5 | 15.0 ± 6.2 | |||||||||
| 625 | 17.0 ± 5.0 | – | – | – | – | – | – | – | – | – |
| 1250 | 17.7 ± 2.5 | – | – | – | – | – | – | – | – | – |
| 2500 | 15.7 ± 0.6 | – | – | – | – | – | – | – | – | – |
| 3750 | 13.7 ± 3.2 | – | – | – | – | – | – | – | – | – |
| 5000 | 19.7 ± 8.7 | – | – | – | – | – | – | – | – | – |
| Positive control c | 79.3 ± 9.0 | – | – | – | – | – | – | – | – | – |
| Preincubation assay | ||||||||||
| 0 (water) | – | – | – | – | – | 24.3 ± 4.5 | 153.3 ± 25.0 | 413.7 ± 12.3 | 9.7 ± 0.6 | 5.7 ± 1.5 |
| 2.29 | – | – | – | – | – | – | – | – | – | – |
| 6.86 | – | – | – | – | – | 29.0 ± 3.5 | – | – | – | – |
| 20.6 | – | – | – | – | – | 23.3 ± 4.2 | – | – | – | – |
| 61.7 | – | – | – | – | – | 35.0 ± 2.0 | 161.0 ± 21.6 | 492.3 ± 23.6 | 9.0 ± 2.6 | 5.7 ± 2.1 |
| 185.2 | – | – | – | – | – | 28.7 ± 9.3 | 172.3 ± 13.3 | 451.7 ± 56.1 | 12.0 ± 5.2 | 12.7 ± 5.7 |
| 555.6 | – | – | – | – | – | 26.3 ± 2.3 | 181.7 ± 6.0 | 421.7 ± 22.3 | 9.3 ± 4.5 | 14.3 ± 4.2 |
| 1666.7 | – | – | – | – | – | 24.7 ± 5.1 | 173.3 ± 20.1 | 505.3 ± 36.2 | 13.7 ± 0.6 | 7.7 ± 2.1 |
| 5000 | – | – | – | – | – | – | 199.3 ± 11.0 | 446.3 ± 75.8 | 14.7 ± 5.0 | 12.7 ± 6.0 |
| Positive control d | – | – | – | – | 1453.7 ± 43.7 | 615.3 ± 38.6 | 918.7 ± 45.5 | 149.7 ± 10.4 | 75.7 ± 4.2 | |
–S9, without metabolic activation; +S9, with metabolic activation; 2‐AM, 2‐anthramine; 9‐AA, 9‐aminoacridine, 2‐NF, 2‐nitrofluorene; 4NQO, 4‐nitroquinoline‐1‐oxide; B[a]P, benzo[a]pyrene; MMC: mitomycin C; NaN3, sodium azide; S. typhimurium, Salmonella typhimurium.
Positive controls: –S9: TA98 = 0.5 μg/plate 2‐NF; TA100 and TA1535 = 1 μg/plate NaN3; TA1537 = 50 μg/plate 9‐AA; TA102 = 0.5 μg/plate MMC; +S9: TA98, TA1535 and TA1537 = 2 μg/plate 2‐AM; TA102 = 20 μg/plate 2‐AM; TA100 = 5 μg/plate B[a]P.
Positive controls –S9: TA98 = 0.5 μg/plate 2‐NF; TA100 and TA1535 = 1 μg/plate NaN3; TA1537 = 50 μg/plate 9‐AA; TA102 = 0.5 μg/plate MMC.
Positive controls –S9: TA98 = 0.5 μg/plate 2‐NF.
Positive controls +S9: TA98, TA1535 and TA1537 = 2 μg/plate 2‐AM; TA102 = 20 μg/plate 2‐AM; TA100 = 5 μg/plate B[a]P.
3.1.2. In vitro mammalian cell micronucleus test
Results for micronucleus analysis are provided in Table 2. Micronucleated binucleated cell values for vehicle control groups were within vehicle control historical ranges (data not shown) and the positive control compounds induced biologically relevant and statistically significant increases in the frequency of micronucleated binucleated cells under appropriate conditions, demonstrating the validity of the assay. Following the 3‐ and 24‐hour exposure periods, pasteurized A. muciniphila did not induce any statistically significant or dose‐related increases in the frequency of micronucleated binucleated cells, compared with vehicle controls. Moreover, the frequencies of micronucleated binucleated cells under all conditions after exposure to the test item were within the corresponding vehicle control historical ranges. These results met the criteria for a negative response.
TABLE 2.
In vitro mammalian cell micronucleus test with pasteurized A. muciniphila
| Concentration (μg/mL) | RI as mean % of control | Mean decrease in RI (%) | Frequency of micronucleated binucleated cells (per 1000) |
|---|---|---|---|
| 3‐h exposure –S9 with 24 h recovery | |||
| Negative control (water) | 3 | ||
| 187.5 | 96 | 4 | 2 |
| 375 | 101 | 0 | 1.5 |
| 750 | 94 | 6 | 1.0 |
| Positive control (COL 0.1 μg/mL) | 60 | 40 | 12.5 *** |
| 3‐h exposure +S9 with 24 h recovery | |||
| Negative control (water) | 2.5 | ||
| 187.5 | 103 | 0 | 2.5 |
| 375 | 103 | 0 | 2.0 |
| 750 | 98 | 2 | 2.0 |
| Positive control (CPA 6 μg/mL) | 74 | 26 | 9.5 ** |
| 24‐hour exposure –S9 without recovery | |||
| Negative control (water) | 3.0 | ||
| 93.75 | 103 | 0 | 4.0 |
| 187.5 | 100 | 0 | 3.0 |
| 375 | 99 | 1 | 2.0 |
| Positive control (MMC 0.1 μg/mL) | 81 | 19 | 16.5 ** |
–S9, without metabolic activation; +S9, with metabolic activation; COL, colchicine; CPA, cyclophosphamide; MMC, mitomycin C; RI, replication index.
Statistically significantly different from vehicle control (P < .01).
Statistically significantly different from vehicle control (P < .001).
3.2. Subchronic toxicity
3.2.1. Clinical examinations, body weights, and food and water consumption
There were no test item‐related deaths or clinical signs (data not shown). One female in the group given 1500 mg/kg body weight/day was killed on day 24 of dosing due to clinical signs, including reduced activity, periorbital tightening, slight to moderate piloerection and markedly forced abdominal respiration. Macroscopic findings (perforated esophagus, with cloudy, thick fluid found in the thoracic cavity) and microscopic findings (mild focal hemorrhage and neutrophilic inflammation of the submucosa of the esophagus) were consistent with a handling error during the dosing, rather than any effect of the test item. There were no ocular changes and no test item‐related findings observed during arena observations or in the modified Irwin test.
Body weights and food consumption throughout the dosing period are presented in Figures 1 and 2, respectively (water consumption data not shown). Animals given pasteurized A. muciniphila gained similar amounts of weight, ate similar amounts of food and drank similar amounts of water compared with vehicle controls.
FIGURE 1.
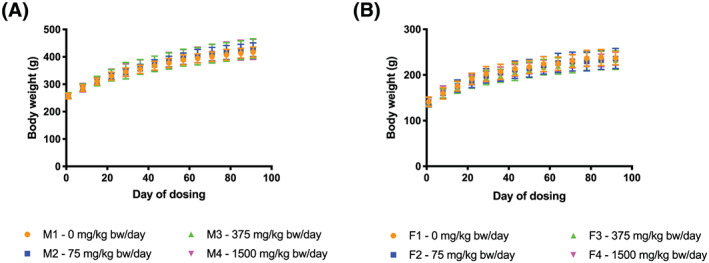
Body weights of rats during the 90‐day study. A, Male. B, Female. bw, body weight
FIGURE 2.
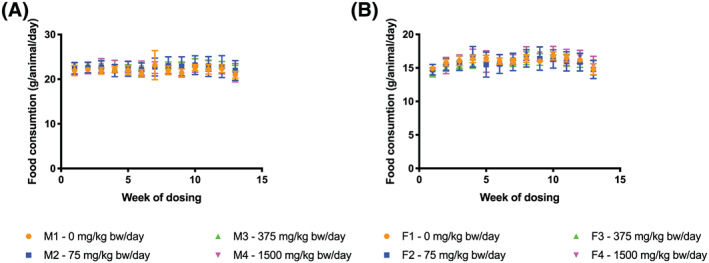
Food consumption of rats during the 90‐day study. A, Male. B, Female. bw, body weight
3.2.2. Hematology, coagulation and clinical chemistry
There were no test item‐related differences in values for hematology or coagulation parameters between pasteurized A. muciniphila‐dosed groups and controls (Table 3). Statistically significant increases in absolute neutrophil count and total white blood cell count were observed for males given 1500 mg/kg body weight/day (9.6 × 1010 cells/kg body weight/day), but these differences were not seen for females. All individual white blood cell count values for high‐dose males were within the historical control range (historical data not shown), whereas three vehicle control values were below the lower limit, indicating that the statistically significant increase for males given 1500 mg/kg body weight/day was likely attributable to control values being lower than expected for animals of this age and strain, rather than any effect of the test item. Furthermore, all but one of the individual absolute neutrophil values for high‐dose males were within the historical control range (one value [2.71 × 109/L] being only narrowly above the upper limit [2.70 × 109/L]), showing that these values also reflected normal biological variation. Statistically significant reductions in relative eosinophils for low‐ and mid‐dose males were clearly unrelated to the test item, as the differences were not observed in the high‐dose group.
TABLE 3.
Hematology and coagulation values for rats administered pasteurized A. muciniphila by oral gavage for 90 days
| Parameter | Hematology and coagulation values (mean ± standard deviation) | |||||||
|---|---|---|---|---|---|---|---|---|
| Males (n = 10 per group) | Females (n = 10 per group except group 4 with n = 9) | |||||||
| 0 (vehicle control) | Pasteurized A. muciniphila dose (mg/kg bw/day) | 0 (vehicle control) | Pasteurized A. muciniphila dose (mg/kg bw/day) | |||||
| 75 | 375 | 1500 | 75 | 375 | 1500 | |||
| WBC (×109/L) | 6.05 ± 1.70 | 7.07 ± 0.96 | 7.20 ± 0.78 | 7.77 ± 0.86 * | 5.18 ± 1.04 | 5.16 ± 0.81 | 5.20 ± 1.24 | 5.16 ± 0.92 |
| RBC (×1012/L) | 9.01 ± 0.35 | 8.95 ± 0.36 | 8.83 ± 0.25 | 9.10 ± 0.36 | 8.08 ± 0.27 | 8.06 ± 0.34 | 7.90 ± 0.47 | 8.10 ± 0.24 |
| Hb (mmol/L) | 10.59 ± 0.22 | 10.46 ± 0.25 | 10.24 ± 0.39 | 10.35 ± 0.23 | 9.80 ± 0.23 | 9.62 ± 0.54 | 9.59 ± 0.56 | 9.77 ± 0.17 |
| Hematocrit (L/L) | 0.47 ± 0.01 | 0.47 ± 0.01 | 0.46 ± 0.02 | 0.46 ± 0.01 | 0.44 ± 0.01 | 0.43 ± 0.01 | 0.43 ± 0.03 | 0.44 ± 0.01 |
| MCV (fL) | 52.3 ± 2.0 | 52.2 ± 2.0 | 52.3 ± 2.1 | 50.8 ± 1.2 | 54.0 ± 1.2 | 54.0 ± 1.3 | 55.0 ± 2.2 | 53.9 ± 1.5 |
| MCH (fmol) | 1.18 ± 0.05 | 1.17 ± 0.05 | 1.16 ± 0.05 | 1.14 ± 0.03 | 1.21 ± 0.03 | 1.19 ± 0.07 | 1.21 ± 0.06 | 1.21 ± 0.03 |
| MCHC (mmol/L) | 22.56 ± 0.27 | 22.40 ± 0.24 | 22.29 ± 0.36 | 22.45 ± 0.24 | 22.40 ± 0.53 | 22.14 ± 1.14 | 22.08 ± 0.79 | 22.37 ± 0.31 |
| Platelets (×109/L) | 533.40 ± 51.00 | 560.50 ± 60.10 | 542.50 ± 77.70 | 601.30 ± 32.60 | 592.30 ± 62.20 | 544.90 ± 112.10 | 563.90 ± 84.30 | 628.00 ± 56.60 |
| Retic (×1012/L) | 0.29 ± 0.03 | 0.28 ± 0.04 | 0.30 ± 0.06 | 0.29 ± 0.05 | 0.25 ± 0.04 | 0.27 ± 0.03 | 0.26 ± 0.03 | 0.28 ± 0.05 |
| Neutrophils (×109/L) | 1.22 ± 0.39 | 1.36 ± 0.26 | 1.43 ± 0.40 | 1.76 ± 0.54* | 1.00 ± 0.25 | 0.91 ± 0.25 | 0.93 ± 0.40 | 0.83 ± 0.25 |
| Lymphocytes (×109/L) | 4.62 ± 1.49 | 5.52 ± 0.83 | 5.60 ± 0.97 | 5.80 ± 0.50 | 4.04 ± 0.84 | 4.13 ± 0.70 | 4.16 ± 0.95 | 4.14 ± 0.78 |
| Monocytes (×109/L) | 0.008 ± 0.006 | 0.009 ± 0.006 | 0.009 ± 0.006 | 0.010 ± 0.008 | 0.004 ± 0.005 | 0.002 ± 0.004 | 0.004 ± 0.007 | 0.006 ± 0.005 |
| Eosinophils (×109/L) | 0.19 ± 0.08 | 0.15 ± 0.06 | 0.14 ± 0.04 | 0.19 ± 0.06 | 0.14 ± 0.06 | 0.12 ± 0.03 | 0.10 ± 0.05 | 0.17 ± 0.14 |
| Basophils (×109/L) | 0.008 ± 0.004 | 0.026 ± 0.047 | 0.014 ± 0.007 | 0.012 ± 0.006 | 0.009 ± 0.009 | 0.009 ± 0.006 | 0.009 ± 0.006 | 0.011 ± 0.006 |
| APTT (s) | 13.88 ± 1.73 | 12.53 ± 1.51 | 12.95 ± 1.84 | 13.05 ± 2.49 | 15.31 ± 1.16 | 16.53 ± 3.41 | 15.25 ± 1.21 | 14.92 ± 2.34 |
| PT (s) | 16.46 ± 0.67 | 16.14 ± 0.54 | 16.10 ± 0.58 | 16.50 ± 0.45 | 15.95 ± 0.71 | 16.14 ± 0.73 | 16.13 ± 0.51 | 16.13 ± 0.72 |
APTT, activated partial thromboplastin time; bw, body weight Hb, hemoglobin; MHC, mean corpuscular hemoglobin; MCHC, mean corpuscular hemoglobin concentration; MCV, mean corpuscular volume; n, number of animals; PT, prothrombin time; RBC, red blood cell count; Retic, absolute reticulocyte count; WBC, white blood cell count.
Statistically significantly different from vehicle controls (P < .05).
No biologically relevant or statistically significant differences in coagulation or blood chemistry parameters were observed between controls and test item groups (Table 4). Statistically significant increases in T3 and T4 for low‐dose males, and in T4 for males given 1500 mg/kg body weight/day, were considered unrelated to the test item as they were not associated with a dose response.
TABLE 4.
Clinical chemistry values for rats administered pasteurized A. muciniphila by oral gavage for 90 days
| Parameter | Clinical chemistry values (mean ± standard deviation) | |||||||
|---|---|---|---|---|---|---|---|---|
| Males (n = 10 per group) | Females (n = 10 per group except group 4 with n = 9) | |||||||
| 0 (vehicle control) | Pasteurized A. muciniphila dose (mg/kg bw/day) | 0 (vehicle control) | Pasteurized A. muciniphila dose (mg/kg bw/day) | |||||
| 75 | 375 | 1500 | 75 | 375 | 1500 | |||
| Sodium (mmol/L) | 134.68 ± 4.11 | 131.30 ± 5.87 | 132.3 ± 5.46 | 132.55 ± 3.04 | 132.76 ± 4.35 | 131.75 ± 6.10 | 132.80 ± 5.22 | 136.10 ± 1.85 |
| Potassium (mmol/L) | 6.71 ± 0.62 | 6.73 ± 1.02 | 7.32 ± 0.67 | 6.96 ± 1.15 | 6.21 ± 0.72 | 6.03 ± 0.83 | 5.79 ± 0.70 | 5.77 ± 0.62 |
| Chloride (mmol/L) | 95.95 ± 3.22 | 93.87 ± 3.93 | 94.89 ± 3.88 | 95.31 ± 1.80 | 95.26 ± 3.23 | 94.69 ± 4.44 | 95.36 ± 3.12 | 97.80 ± 1.08 |
| Calcium (mmol/L) | 2.49 ± 0.13 | 2.46 ± 0.12 | 2.47 ± 0.11 | 2.45 ± 0.07 | 2.47 ± 0.08 | 2.46 ± 0.20 | 2.44 ± 0.12 | 2.50 ± 0.05 |
| Magnesium (mmol/L) | 0.89 ± 0.05 | 0.91 ± 0.05 | 0.94 ± 0.08 | 0.91 ± 0.04 | 1.01 ± 0.04 | 1.03 ± 0.11 | 1.01 ± 0.13 | 0.97 ± 0.02 |
| Phosphorus (mmol/L) | 1.52 ± 0.13 | 1.39 ± 0.29 | 1.43 ± 0.09 | 1.43 ± 0.22 | 1.40 ± 0.27 | 1.38 ± 0.25 | 1.51 ± 0.42 | 1.48 ± 0.17 |
| ALP (μkat/L) | 2.18 ± 0.50 | 2.10 ± 0.32 | 2.23 ± 0.58 | 2.24 ± 0.62 | 1.50 ± 0.57 | 1.42 ± 0.39 | 1.29 ± 0.44 | 1.47 ± 0.45 |
| AST (μkat/L) | 1.30 ± 0.17 | 1.42 ± 0.25 | 1.42 ± 0.12 | 1.22 ± 0.16 | 1.35 ± 0.19 | 1.24 ± 0.15 | 1.17 ± 0.20 | 1.24 ± 0.14 |
| ALT (μkat/L) | 0.65 ± 0.13 | 0.61 ± 0.87 | 0.64 ± 0.11 | 0.59 ± 0.10 | 0.62 ± 0.12 | 0.64 ± 0.09 | 0.60 ± 0.12 | 0.62 ± 0.10 |
| Carbamide urea (mmol/L) | 7.14 ± 0.39 | 6.89 ± 0.57 | 7.52 ± 1.19 | 7.43 ± 0.26 | 7.60 ± 1.04 | 7.15 ± 0.46 | 7.11 ± 0.86 | 6.90 ± 0.78 |
| Creatinine (μmol/L) | 32.00 ± 3.70 | 29.30 ± 1.3 | 29.90 ± 2.9 | 28.70 ± 3.80 | 32.30 ± 1.90 | 31.20 ± 3.80 | 30.30 ± 3.40 | 32.00 ± 2.00 |
| Total protein (g/L) | 66.76 ± 4.93 | 66.92 ± 4.17 | 69.78 ± 4.61 | 66.87 ± 3.07 | 71.19 ± 3.53 | 70.77 ± 6.79 | 68.19 ± 4.87 | 69.43 ± 2.36 |
| Albumin (g/L) | 41.55 ± 1.65 | 42.11 ± 2.19 | 41.94 ± 3.21 | 41.54 ± 1.99 | 47.59 ± 2.65 | 45.93 ± 4.03 | 45.38 ± 3.03 | 47.58 ± 2.12 |
| A/G ratio | 1.67 ± 0.21 | 1.72 ± 0.24 | 1.51 ± 0.15 | 1.67 ± 0.27 | 23.60 ± 2.77 | 24.84 ± 3.52 | 22.81 ± 3.60 | 21.86 ± 2.29 |
| Triglyceride (mmol/L) | 1.51 ± 0.51 | 1.80 ± 0.50 | 1.99 ± 0.97 | 1.46 ± 0.58 | 1.21 ± 0.39 | 1.59 ± 0.55 | 1.02 ± 0.51 | 1.45 ± 0.60 |
| Total cholesterol (mmol/L) | 1.68 ± 0.19 | 1.59 ± 0.24 | 1.77 ± 0.25 | 1.67 ± 0.28 | 1.29 ± 0.22 | 1.31 ± 0.28 | 1.32 ± 0.37 | 1.40 ± 0.34 |
| Glucose (mmol/L) | 6.20 ± 0.73 | 6.02 ± 0.53 | 6.53 ± 1.02 | 6.54 ± 0.33 | 5.41 ± 0.57 | 5.49 ± 0.50 | 5.86 ± 0.84 | 5.62 ± 0.79 |
A/G ratio, albumin/globulin ratio; ALP, alkaline phospahtase; ALT, alanine aminotransferase; AST, aspartate aminotransferase; bw, body weight; n, number of animals.
3.2.3. Macroscopic examinations, organ weights and histopathology
Organ weights relative to body weight are provided in Table 5. There were no differences in organ weights between controls and pasteurized A. muciniphila‐dosed groups. There were also no test item‐related macroscopic or microscopic findings (data not shown). The only findings observed were incidental and generally consistent with changes encountered in rats of this age and strain kept under laboratory conditions (data not shown).
TABLE 5.
Organ weights relative to body weight for rats administered pasteurized A. muciniphila by oral gavage for 90 days
| Organ | Organ weights relative to body weight (mean ± standard deviation) | |||||||
|---|---|---|---|---|---|---|---|---|
| Males (n = 10 per group) | Females (n = 10 per group except group 4 with n = 9) | |||||||
| 0 (vehicle control) | Pasteurized A. muciniphila dose (mg/kg bw/day) | 0 (vehicle control) | Pasteurized A. muciniphila dose (mg/kg bw/day) | |||||
| 75 | 375 | 1500 | 75 | 375 | 1500 | |||
| Adrenals | 0.014 ± 0.002 | 0.013 ± 0.002 | 0.013 ± 0.002 | 0.013 ± 0.002 | 0.032 ± 0.005 | 0.032± 0.003 | 0.032 ± 0.004 | 0.032 ± 0.006 |
| Brain | 0.533 ± 0.031 | 0.539 ± 0.043 | 0.528 ± 0.037 | 0.523 ± 0.039 | 0.866 ± 0.058 | 0.860 ± 0.747 | 0.876 ± 0.078 | 0.874 ± 0.058 |
| Epididymides | 0.357 ± 0.060 | 0.352 ± 0.437 | 0.337 ± 0.042 | 0.337 ± 0.037 | NA | NA | NA | NA |
| Heart | 0.291 ± 0.015 | 0.295 ± 0.011 | 0.298 ± 0.016 | 0.292 ± 0.009 | 0.332 ± 0.018 | 0.330 ± 0.025 | 0.344 ± 0.022 | 0.350 ± 0.035 |
| Kidneys | 0.612 ± 0.041 | 0.640 ± 0.035 | 0.6319 ± 0.052 | 0.643 ± 0.041 | 0.679 ± 0.033 | 0.694 ± 0.026 | 0.681 ± 0.022 | 0.699 ± 0.032 |
| Liver | 3.097 ± 0.181 | 3.223 ± 0.216 | 3.218 ± 0.249 | 3.236 ± 0.266 | 3.256 ± 0.157 | 3.369 ± 0.205 | 3.26± 0.140 | 3.372 ± 0.188 |
| Ovaries | NA | NA | NA | NA | 0.044 ± 0.006 | 0.048 ± 0.008 | 0.049 ± 0.009 | 0.049 ± 0.008 |
| Pituitary | 0.002 ± 0.0002 | 0.002 ± 0.0004 | 0.0023 ± 0.0004 | 0.002 ± 0.0003 | 0.006 ± 0.0008 | 0.0052 ± 0.0011 | 0.005 ± 0.0004 | 0.006 ± 0.0004 |
| Prostate | 0.141 ± 0.029 | 0.148 ± 0.024 | 0.1342 ± 0.035 | 0.142 ± 0.027 | NA | NA | NA | NA |
| Spleen | 0.163 ± 0.029 | 0.158 ± 0.017 | 0.165 ± 0.018 | 0.161 ± 0.011 | 0.190 ± 0.029 | 0.198 ± 0.025 | 0.202 ± 0.028 | 0.210± 0.022 |
| Testes | 0.903 ± 0.093 | 0.897 ± 0.095 | 0.879 ± 0.081 | 0.913 ± 0.068 | NA | NA | NA | NA |
| Thymus | 0.068 ± 0.016 | 0.068 ± 0.017 | 0.061 ± 0.014 | 0.064 ± 0.017 | 0.133 ± 0.023 | 0.123 ± 0.020 | 0.129 ± 0.025 | 0.131 ± 0.022 |
| Uterus | NA | NA | NA | NA | 0.308 ± 0.112 | 0.364 ± 0.130 | 0.371± 0.117 | 0.393± 0.19409 |
bw, body weight; n, number of animals; NA, not applicable.
4. DISCUSSION
The majority of probiotics sold on the market mainly include microorganisms from the genera Lactobacillus and Bifidobacterium (Douillard & de Vos, 2014). However, much data support the interest of next generation beneficial bacteria in providing health benefit to humans (O'Toole, Marchesi, & Hill, 2017). The safety of these novel microbes for human consumption has to be evaluated carefully (Brodmann et al., 2017).
A. muciniphila is one of the most abundant single species in the human intestinal microbiota (0.5%‐5% of the total bacteria) but was only isolated and characterized in 2004 (Collado et al., 2007; Derrien et al., 2004). The bacterium is present in the human gut from infancy and cells have been detected in breast milk and its abundance increases significantly during growth into adulthood (Geerlings et al., 2018). Over the last 10 years, numerous studies have found that the bacterium A. muciniphila was positively associated with healthy gut lining, reduced metabolic disorders and decreased low‐grade inflammation (Cani & de Vos, 2017). In addition, the abundance of the bacterium is strongly increased upon gastric bypass or antidiabetic drug treatment such as metformin (Forslund et al., 2015; Liou et al., 2013). The administration of live A. muciniphila has been associated with several beneficial effects on cardiometabolic risk factors (Everard et al., 2013; Shin et al., 2013). It was only in 2017 that the strong metabolic effects of pasteurized A. muciniphila were described (Plovier et al., 2005) and then confirmed in other studies (Depommier et al., 2020; Wang et al., 2019). The beneficial effects of consuming microorganisms inactivated by pasteurization for 30 minutes at 70°C, a mild heat treatment limiting the denaturation of cellular components, have also been described in previous studies (Peng & Hsu, 2015; Sakai et al., 2017). Therefore, pasteurization of A. muciniphila, provides enhanced beneficial health effects (compared with live A. muciniphila) as well as increased stability and negligible risk of toxicity.
As A. muciniphila is part of nutrition in the early stages of life (Collado et al., 2012) and is a commensal gut bacterium constituting 3%‐5% of the gut microbiota, it would reasonably be expected to be safe for human consumption. Furthermore, the development of pasteurized A. muciniphila still reinforces this safety aspect, as the pasteurized bacterium is unable to replicate or to colonize the human gut. Other heat‐killed bacteria, even some bacteria close to pathogens from a taxonomic point of view, have already been recognized as safe by EFSA (2015, 2019). There are also previously published studies assessing the safety of A. muciniphila (Brodmann et al., 2017; Cozzolino et al., 2020; Gómez‐Gallego et al., 2016; Zhang, Li, Cheng, Buch, & Zhang, 2020); however, to the best of our knowledge, this is the first article reporting the findings of studies conducted with A. muciniphila according to the requirements of the US FDA and EFSA guidances on toxicity testing required for the safety assessment of a novel nonabsorbable food ingredient (EFSA, 2012; US FDA Redbook, 2014).
Pasteurized A. muciniphila has been demonstrated to have only beneficial effects for humans and administration of pasteurized A. muciniphila was not associated with any notable change to the overall pattern of gut microbiota and abundance of other bacterial species in humans and rodents (Depommier et al., 2019; Everard et al., 2013; Li, Lin, Vanhoutte, Woo, & Xu, 2016). A randomized, double‐blind, placebo‐controlled proof‐of‐concept study in overweight/obese insulin‐resistant volunteers showed that the daily supplementation with pasteurized A. muciniphila at a dose of 1 × 1010 cells/day for 12 weeks was well tolerated and improved several metabolic parameters (i.e., insulin sensitivity, insulinemia, plasma total cholesterol as well as relevant blood markers for liver dysfunction and inflammation) (Depommier et al., 2019). In addition, as pasteurized A. muciniphila has been heat‐treated, it will not colonize the intestine and is primarily excreted as dead bacteria in feces, as demonstrated previously using rRNA gene‐based detection methods (Depommier et al., 2019).
Despite there being no reported adverse effects associated with A. muciniphila in previously conducted studies, in addition to the high prevalence of A. muciniphila in the gastrointestinal tract of virtually all healthy humans and its presence in breast milk, there are no data available or any documented history of use as a food ingredient. Thus, given the vastly increasing amount of scientific research showing an interest to propose pasteurized A. muciniphila as a potential food ingredient, an appropriate toxicological evaluation was warranted.
The methodology for the safety assessment of pasteurized A. muciniphila follows the tiered approach to toxicity testing proposed by EFSA and the US FDA Redbook guidance on safety assessment of food ingredients (EFSA, 2012; US FDA Redbook, 2014). This approach involves assessment of potential genotoxicity using the bacterial reverse mutation and in vitro mammalian cell micronucleus tests, followed by a 90‐day oral toxicity study to assess the potential for subchronic toxicity.
The results of the in vitro genotoxicity tests demonstrated that pasteurized A. muciniphila is neither mutagenic (as assessed in the bacterial reverse mutation test) nor clastogenic or aneugenic (as assessed in the in vitro mammalian cell micronucleus test). In the 90‐day study, there were no test item‐related adverse effects on clinical observations, neurobehavioral assessments, body weight, food consumption, water consumption, clinical pathology, organ weights or histopathology. Statistically significant differences in leukocyte counts for pasteurized A. muciniphila‐administered groups compared with vehicle controls were minimal, limited to one gender and individual values were generally within historical control ranges, thus reflecting normal biological variation. Absence of any associated changes in organ weights, or of any corresponding macroscopic or histopathological findings, confirms that these differences were likely to be incidental, rather than a direct result of pasteurized A. muciniphila administration. Therefore, the no‐observed‐adverse‐effect level (NOAEL) for the 90‐day study was concluded to be 1500 mg/kg body weight/day (9.6 × 1010 cells/kg body weight/day), the highest dose tested. This NOAEL provides a safety factor of over 100‐fold when compared with the worst‐case anticipated exposure from use in foods; 100‐fold is considered the minimum required safety factor when applying animal experimentation data to humans (21 CFR 170.22; US FDA, 2000).
In conclusion, the negative in vitro genotoxicity results, combined with the absence of any test item‐related adverse effects in the 90‐day toxicity study, support the safety of pasteurized A. muciniphila for use as a food ingredient.
CONFLICT OF INTEREST
A‐Mansia Biotech SA supplied the test item and sponsored the studies reported in this research article. P.D.C. and W.M.dV. are co‐founders of A‐Mansia Biotech SA and owner of patents on the use of A. muciniphila and health. C.D. and H.P. are employees of A‐Mansia Biotech SA. K.R.P. is an employee of Intertek Health Sciences Inc., which has provided consultancy services to A‐Mansia Biotech SA.
ACKNOWLEDGMENTS
This work was supported by the Walloon Region (DG06, FIRST Spin‐Off grant from the Walloon Region Microbe4U no. 1410053). P.D.C. is a senior research associate at Fonds de la Recherche Scientifique (FRS‐FNRS). P.D.C. is a recipient of the PoC ERC grant 2016 (no. Microbes4U_713547)
Druart C, Plovier H, Van Hul M, et al. Toxicological safety evaluation of pasteurized Akkermansia muciniphila . J Appl Toxicol. 2021;41:276–290. 10.1002/jat.4044
REFERENCES
- Ames, B. N. , McCann, J. , & Yamasaki, E. (1975). Methods for detecting carcinogens and mutagens with the Salmonella/mammalian‐microsome mutagenicity test. Mutation Research, 31(6), 347–363. 10.1016/0165-1161(75)90046‐1. [DOI] [PubMed] [Google Scholar]
- Brodmann, T. , Endo, A. , Gueimonde, M. , Vinderola, G. , Kneifel, W. , de Vos, W. M. , … Gómez‐Gallego, C. (2017). Safety of novel microbes for human consumption: practical examples of assessment in the European Union. Frontiers in Microbiology, 8, 1725 10.3389/fmicb.2017.01725 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cani, P. D. , & de Vos, W. M. (2017). Next‐generation beneficial microbes: the case of Akkermansia muciniphila . Frontiers in Microbiology, 8, 1765 10.3389/fmicb.2017.01765 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collado, M. C. , Derrien, M. , Isolauri, E. , de Vos, W. M. , & Salminen, S. (2007). Intestinal integrity and Akkermansia muciniphila, a mucin‐degrading member of the intestinal microbiota present in infants, adults, and the elderly. Applied and Environmental Microbiology, 73, 7767–7770. 10.1128/AEM.01477-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collado, M. C. , Laitinen, K. , Salminen, S. , & Isolauri, E. (2012). Maternal weight and excessive weight gain during pregnancy modify the immunomodulatory potential of breast milk. Pediatric Research, 72, 77–85. 10.1038/pr.2012.42 [DOI] [PubMed] [Google Scholar]
- Cozzolino, A. , Vergalito, F. , Tremonte, P. , Iorizzo, M. , Lombardi, S. J. , Sorrentino, E. , … Succi, M. (2020). Preliminary evaluation of the safety and probiotic potential of Akkermansia muciniphila DSM 22959 in comparison with Lactobacillus rhamnosus GG. Microorganisms, 8, 189 10.3390/microorganisms8020189 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Depommier, C. , Everard, A. , Druart, C. , Plovier, H. , Van Hul, M. , Vieira‐Silva, S. , … Cani, P. D. (2019). Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: a proof‐of‐concept exploratory study. Nature Medicine, 25(7), 1096–1103. 10.1038/s41591-019-0495-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Depommier, C. , Van Hul, M. , Everard, A. , Delzenne, N. M. , De Vos, W. M. , & Cani, P. D. (2020). Pasteurized Akkermansia muciniphila increases whole‐body energy expenditure and fecal energy excretion in diet‐induced obese mice. Gut Microbes, 1–15 [Advance online publication]. 10.1080/19490976.2020.1737307 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Derrien, M. , Belzer, C. , & de Vos, W. M. (2017). Akkermansia muciniphila and its role in regulating host functions. Microbial Pathogenesis, 106, 171–181. 10.1016/j.micpath.2016.02.005 [DOI] [PubMed] [Google Scholar]
- Derrien, M. , Collado, M. C. , Ben‐Amor, K. , Salminen, S. , & de Vos, W. M. (2008). The Mucin degrader Akkermansia muciniphila is an abundant resident of the human intestinal tract. Applied and Environmental Microbiology, 74(5), 1646–1648. 10.1128/AEM.01226-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Derrien, M. , Vaughan, E. E. , Plugge, C. M. , & de Vos, W. M. (2004). Akkermansia muciniphila gen. nov., sp. nov., a human intestinal mucin‐degrading bacterium. International Journal of Systematic and Evolutionary Microbiology, 54, 1469–1476. 10.1099/ijs.0.02873-0 [DOI] [PubMed] [Google Scholar]
- Douillard, F. P. , & de Vos, W. M. (2014). Functional genomics of lactic acid bacteria: from food to health. Microbial Cell Factories, 13(Suppl 1), S8(21pp). 10.1186/1475-2859-13-s1-s8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA . (2012). Guidance for submission for food additive evaluations. EFSA Journal, 10(7), 2760 10.2903/j.efsa.2012.2760 [DOI] [Google Scholar]
- EFSA . (2015). Scientific Opinion on the safety of “heat‐treated milk products fermented with Bacteroides xylanisolvens DSM 23964” as a novel food. EFSA Journal, 13(1), 3956 10.2903/j.efsa.2015.3956 [DOI] [Google Scholar]
- EFSA . (2016). Guidance on the preparation and presentation of an application for authorisation of a novel food in the context of Regulation (EU) 2015/2283. EFSA Journal, 14(11), 4594 10.2903/j.efsa.2016.4594 [DOI] [Google Scholar]
- EFSA . (2019). Scientific Opinion on the safety of heat‐killed Mycobacterium setense manresensis as a novel food pursuant to Regulation (EU) 2015/2283. EFSA Journal, 17(11), 5824(13pp). 10.2903/j.efsa.2019.5824 [DOI] [PMC free article] [PubMed] [Google Scholar]
- European Union . (2010). Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes. Official Journal of the European Union, 53(L276), 33–79. Retrieved from: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A32010L0063 [Google Scholar]
- Everard, A. , Belzer, C. , Geurts, L. , Ouwerkerk, J. P. , Druart, C. , Bindels, L. B. , … Cani, P. D. (2013). Cross‐talk between Akkermansia muciniphila and intestinal epithelium controls diet‐induced obesity. Proceedings of the National Academy of Sciences of the United States of America, 110(22), 9066–9071. 10.1073/pnas.1219451110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falony, G. , Joossens, M. , Vieira‐Silva, S. , Wang, J. , Darzi, Y. , Faust, K. , … Raes, J. (2016). Population‐level analysis of gut microbiome variation. Science, 352(6285), 560–564. 10.1126/science.aad3503 [DOI] [PubMed] [Google Scholar]
- Fenech, M. (1993). The cytokinesis‐block micronucleus technique and its application to genotoxicity studies in human populations. Environmental Health Perspectives, 101(Suppl. 3), 101–107. 10.1289/ehp.93101s3101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fenech, M. , & Morley, A. A. (1985). Measurement of micronuclei in lymphocytes. Mutation Research, 147, 29–36. 10.1016/0165-1161(85)90015-9 [DOI] [PubMed] [Google Scholar]
- Forslund, K. , Hildebrand, F. , Nielsen, T. , Falony, G. , Le Chatelier, E. , Sunagawa, S. , … Pedersen, O. (2015). Disentangling type 2 diabetes and metformin treatment signatures in the human gut microbiota. Nature, 528(7581), 262–266. 10.1038/nature15766 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geerlings, S. Y. , Kostopoulos, I. , de Vos, W. M. , & Belzer, C. (2018). Akkermansia muciniphila in the human gastrointestinal tract: when, where, and how? Microorganisms, 6(3), 75 10.3390/microorganisms6030075 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gómez‐Gallego, C. , Pohl, S. , Salminen, S. , De Vos, W. M. , & Kneifel, W. (2016). Akkermansia muciniphila: a novel functional microbe with probiotic properties. Beneficial Microbes, 7, 571–584. 10.3920/BM2016.0009 [DOI] [PubMed] [Google Scholar]
- Li, J. , Lin, S. , Vanhoutte, P. M. , Woo, C. W. , & Xu, A. (2016). Akkermansia muciniphila protects against atherosclerosis by preventing metabolic endotoxemia‐induced inflammation in Apoe −/− mice. Circulation, 133, 2434–2446. 10.1161/CIRCULATIONAHA.115.019645 [DOI] [PubMed] [Google Scholar]
- Liou, A. P. , Paziuk, M. , Luevano, J. M. Jr. , Machineni, S. , Turnbaugh, P. J. , & Kaplan, L. M. (2013). Conserved shifts in the gut microbiota due to gastric bypass reduce host weight and adiposity. Science Translational Medicine, 5(178), 178ra41 10.1126/scitranslmed.3005687 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maron, D. M. , & Ames, B. N. (1983). Revised methods for the Salmonella mutagenicity test. Mutation Research, 113, 173–215. 10.1016/0165-1161(83)90010-9 [DOI] [PubMed] [Google Scholar]
- OECD . (1997). Bacterial reverse mutation test. In OECD guidelines for the testing of chemicals. OECD Guideline No. 471 (Updated & Adopted: 21 July 1997). Paris, France: Organisation for Economic Co‐operation and Development (OECD). Retrieved from: http://www.oecd-ilibrary.org/environment/test-no-471-bacterial-reverse-mutation-test_9789264071247-en;jsessionid=1ijgh8o34k0qd.delta
- OECD . (1998). OECD principles of good laboratory practice. Series on Principles of Good Laboratory Practice and Compliance Monitoring, No. 1 (ENV/MC/CHEM(98)17). Paris, France: Organisation for Economic Co‐Operation & Development (OECD), Environment Directorate, Chemicals Group and Management Committee. Retrieved from: http://www.oecd-ilibrary.org/environment/oecd-principles-on-goodlaboratory-practice_9789264078536-en
- OECD . (2016). In vitro mammalian cell micronucleus test. In OECD guidelines for the testing of chemicals. OECD Guideline No. 487 (Updated & Adopted: 29 July 2016). Paris, France: Organisation for Economic Cooperation and Development (OECD). Retrieved from: http://www.oecd-ilibrary.org/environment/test-no-487-in-vitro-mammalian-cellmicronucleus-test_9789264264861-en;jsessionid=58ip5ibjcf4kc.xoecd-live-02
- OECD . (2018). Repeated dose 90‐day oral toxicity study in rodents. In OECD guidelines for the testing of chemicals. OECD Guideline no 408 (Updated and Adopted 27 June 2018). Paris, France, Organisation for Economic Cooperation and Development (OECD). Retrieved from: https://www.oecd-ilibrary.org/environment/test-no-408-repeated-dose-90-day-oral-toxicity-study-in-rodents_9789264070707-en
- O'Toole, P. W. , Marchesi, J. R. , & Hill, C. (2017). Next‐generation probiotics: the spectrum from probiotics to live biotherapeutics. Nat Microbiol, 2, 17057. PMID: 28440276. 10.1038/nmicrobiol.2017.57 [DOI] [PubMed] [Google Scholar]
- Ottman, N. (2015). Host immunostimulation and substrate utilization of the gut symbiont Akkermansia muciniphila. PhD thesis. Wagenigen, The Netherlands: Wagenigen University. Retrieved from: https://library.wur.nl/WebQuery/wurpubs/490930
- Peng, G. C. , & Hsu, C. H. (2005). The efficacy and safety of heat‐killed Lactobacillus paracasei for treatment of perennial allergic rhinitis induced by house‐dust mite. Pediatric Allergy and Immunology, 16, 433–438. 10.1111/j.1399-3038.2005.00284.x [DOI] [PubMed] [Google Scholar]
- Plovier, H. , Everard, A. , Druart, C. , Depommier, C. , Van Hul, M. , Geurts, L. , … Cani, P. D. (2017). A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nature Medicine, 23, 107–113. 10.1038/nm.4236 [DOI] [PubMed] [Google Scholar]
- Sakai, T. , Taki, T. , Nakamoto, A. , Shuto, E. , Tsutsumi, R. , Toshimitsu, T. , … Ikegami, S. (2013). Lactobacillus plantarum OLL2712 regulates glucose metabolism in C57BL/6 mice fed a high‐fat diet. Journal of Nutritional Science and Vitaminology, 59, 144–147. 10.3177/jnsv.59.144 [DOI] [PubMed] [Google Scholar]
- Shin, N.‐R. , Lee, J.‐C. , Lee, H.‐Y. , Kim, M.‐S. , Whon, T. W. , Lee, M.‐S. , & Bae, J.‐W. (2014). An increase in the Akkermansia spp. population induced by metformin treatment improves glucose homeostasis in diet‐induced obese mice. Gut, 63, 727–735. 10.1136/gutjnl-2012-303839 [DOI] [PubMed] [Google Scholar]
- US FDA . (2000). Guidance for industry and other stakeholders: Toxicological principles for the safety assessment of food ingredients: Redbook 2000 [updated to July 2007]. Silver Spring, MD: US Food and Drug Administration (US FDA), Center for Food Safety and Applied Nutrition (CFSAN). Retrieved from: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/guidance-industry-and-other-stakeholders-toxicological-principles-safety-assessment-food-ingredients-0
- US FDA . (2019). Part 170—Food additives. §170.22—Safety factors to be considered. In US Code of Federal Regulations (CFR). Title 21: Food and drugs. Food and Drug Administration. Washington, DC: US Government Printing Office (GPO). Retrieved from: https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/CFRSearch.cfm?fr=170.22
- Wang, L. , Tang, L. , Feng, Y. , Zhao, S. , Han, M. , Zhang, C. , … Zhang, Z. (2020). A purified membrane protein from Akkermansia muciniphila or the pasteurised bacterium blunts colitis associated tumourigenesis by modulation of CD8(+) T cells in mice. Gut (online ahead of print—March 13, 2020). 10.1136/gutjnl-2019-320105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu, Y. , Wang, N. , Tan, H. Y. , Li, S. , Zhang, C. , & Feng, Y. (2020). Function of Akkermansia muciniphila in obesity: interactions with lipid metabolism, immune response and gut systems. Frontiers in Microbiology, 11, 219 10.3389/fmicb.2020.00219 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, T. , Li, Q. , Cheng, L. , Buch, H. , & Zhang, F. (2019). Akkermansia muciniphila is a promising probiotic. Microbial Biotechnology, 12, 1109–1125. 10.1111/1751-7915.13410 [DOI] [PMC free article] [PubMed] [Google Scholar]