

Abstract
Gastric cancer (GC) is a common cancer worldwide and its precise mechanism is largely unknown. The aim of the present study was to evaluate the expression levels of NOD-like receptor X1 (NLRX1), tumor necrosis factor receptor-associated factor 6 (TRAF6) and NF-κB in GC and normal gastric tissue samples to determine the association with the clinicopathological features of GC. GC and adjacent normal gastric tissues from 60 patients with GC were analyzed using immunohistochemistry and western blotting analysis. In addition, the association between NLRX1, TRAF6 and NF-κB expression levels were investigated by Spearman's correlation analysis. The results revealed that NLRX1 protein expression levels were downregulated in the GC tissues compared with the normal gastric tissues (P<0.05). Conversely, TRAF6 and NF-κB protein expression levels were upregulated in the GC tissues compared with the normal gastric tissues (P<0.05). A significant difference was identified between GC patients with high and low expression levels of each protein in the tumor size, vascular invasion, neural invasion, lymph node metastasis, differentiation, gross stage and clinical stage. In addition, a negative correlation was observed between NLRX1 and TRAF6, and NLRX1 and NF-κB expression levels, while a positive correlation was observed between TRAF6 and NF-κB expression levels. In conclusion, NLRX1 expression levels were discovered to be downregulated in GC tissues. The expression levels of NLRX1, TRAF6 and NF-κB were also significantly associated with the clinicopathological characteristics of GC, and the aforementioned results indicated that NLRX1 may be a biomarker in assessing GC.
Keywords: NOD-like receptor X1, tumor necrosis factor receptor-associated factor 6, gastric cancer, NF-κB, clinico-pathological characteristics
Introduction
Gastric cancer (GC) is a challenging disease for general surgeons to manage and is one of the most common types of cancer worldwide, and GC has the third leading mortality rate worldwide, causing 723,000 deaths every year (1,2). In developing countries, 70% of deaths result from GC compared with 40% in China (3). Changes in lifestyle and eating habits, for example unhealthy diet, may increase the possibility of the incidence of GC (4). GC is more prevalent in East Asia than other geographic areas (5). Surgery, including open surgery and minimally invasive surgery, is the main treatment option for GC; however, as <10% patients with GC in developing countries are diagnosed early, there is a poor survival rate (6,7). Thus, the mechanisms underlying GC require further investigation.
Previous studies have demonstrated that NF-κB served a crucial role during the progression of GC (8,9). The NF-κB complex is activated in the cytoplasm following the degradation of inhibitor (I)κB, and is then involved in the nuclear physiologic response (10). Tumor necrosis factor receptor-associated factor 6 (TRAF6), which belongs to the TRAF family (11), is an adaptor protein that has important roles in innate immune responses and is a participator in the activatory process of the NF-κB signaling pathway (12). In addition, Han et al (11) reported that TRAF6 promoted the invasion and metastasis of GC and was an index for GC prognosis; Sun et al (13) concluded that the expression levels of TRAF6 in the skeletal muscle of patients with GC were significantly upregulated; and Maeda et al (14) suggested that Helicobacter pylori (H. pylori) may lead to NF-κB activation through a TRAF6 intracellular signaling pathway, which to the authors' best knowledge, is the only report of a relationship between NF-κB and TRAF6 in GC.
The NOD-like receptor (NLR) family serves a crucial role in immune defense and inflammation (15,16). A new member of the NLR family member, NLRX1, has been identified as a protein localized to the membrane of the mitochondria (16,17). Previous studies have revealed that NLRX1 suppressed tumorigenesis by inhibiting NF-κB signaling (18,19); however, the role of NLRX1, the correlation between NLRX1, TRAF6 and NF-κB expression levels, and the relationship between NLRX1 and the clinicopathological characteristics of GC have not been established.
In the present study, the expression levels of NLRX1, TRAF6 and NF-κB in GC tissues were determined and the association between these three proteins and the clinicopathological characteristics of gastric adenocarcinoma (GA) were examined. The present research may offer insight into novel molecular mechanisms underlying GC, specifically GA.
Materials and methods
Patients studies
The present study included 60 patients (age range, 34-83 years; 51 males and 9 females), who were diagnosed with GA based on post-operative pathologic evaluations at The Third People's Hospital of Dalian (Dalian, China) between October 2017 and April 2019. All patient demographic and clinicopathological data were recorded (Table I). The patients were diagnosed with GA by gastroscopy. Patients that received pre-operative chemotherapy or radiotherapy were excluded from the present study. The tissues were obtained at the time of surgery and immediately stored at -80˚C. GA and adjacent normal gastric tissues (>5 cm from the tumor) were collected. Written informed consent regarding information and tissue samples were acquired from the patients. The present study was approved by the Ethics Committee of The Third People's Hospital of Dalian (approval no. 2017-KY-004).
Table I.
Demographic and clinicopathological characteristics of the patients.
| Clinicopathological features | |
|---|---|
| Age, years (mean ± SD) | 66.5±11.4 |
| Male, number (%) | 51(85) |
| Female, number (%) | 9(15) |
| Gross type, number | |
| Early GC | 8 |
| Advanced GC | 52 |
| Tumor size, number | |
| <5 cm | 29 |
| ≥5 cm | 31 |
| Differentiation, number | |
| Moderately-to-well | 33 |
| Poorly differentiated | 27 |
| Lymph node metastasis, number | |
| Positive | 31 |
| Negative | 29 |
| Vascular invasion, number | |
| Yes | 35 |
| No | 25 |
| Nerve invasion, number | |
| Yes | 26 |
| No | 34 |
| Depth of infiltration, number | |
| T1 or T2 | 18 |
| T3 or T4 | 42 |
| Clinical stage, number | |
| I/II | 34 |
| III/IV | 26 |
GC, gastric cancer; T, tumor.
The histologic grade was evaluated using the World Health Organization tumor classification: i) grade well if gland tissue is present, possibly including metaplasia; ii) grade poorly if highly irregular glands are indistinguishable; and iii) grade moderately if the condition is between the grade well and grade poorly classifications (20).
Immunohistochemistry (IHC)
IHC staining was performed to analyze the expression levels of NLRX1, TRAF6 and NF-κB in GC and normal gastric tissues. The tissues were fixed in 10% formalin for 24 h at room temperature, Subsequently, the tissues were embedded in paraffin and cut into 4-µm thick sections. Paraffin sections were placed in an incubator at 60˚C for 120 min. Xylene was used for deparaffinization and ethanol was used for rehydration at room temperature. Then the sections were subsequently incubated at 37˚C with 3% H2O2 for 10 min to block endogenous peroxidase activity. Antigen retrieval was performed by boiling the sections with 0.01 M citric acid buffer (pH 6.0) at 95˚C for 20 min and the sections were then blocked using goat serum (cat. no. SP-9000; OriGene Technologies, Inc.) at 37˚C for 15 min. Subsequent incubation was performed with primary antibodies against NLRX1 (1:100; cat. no. ABP57527; Abbkine Scientific Co., Ltd.), TRAF6 (1:100; cat. no. ABP52637; Abbkine Scientific Co., Ltd.) and NF-κB (1:100; cat. no. ABP51957; Abbkine Scientific Co., Ltd.) overnight in a humidified chamber at 4˚C. Negative controls were performed using PBS. Following the primary antibody incubation, the samples were then incubated with HRP-conjugated anti-rabbit secondary antibody (cat. no. SP-9000; OriGene Technologies, Inc.) at 37˚C for 15 min. Finally, DAB (cat. no. ZLI-9018; OriGene Technologies, Inc.) was added for 5 min and the tissue sections were counterstained with hematoxylin for 20 sec (both at room temperature).
IHC-stained slides were accessed by two pathologists who were blinded to the nature of the research using a light microscope (Nikon Eclipse Ni-E; Nikon Corporation; magnification, x400). The stained slides were evaluated semi-quantitatively and the intensity of staining was categorized into four levels as follows: No staining=0; weak staining=1; moderate staining=2; and strong staining=3. Positive stained cells were scored as follows: 1, 0-25; 2, 26-50; 3, 51-75; and 4, 76-100%. The final score was based on the above scores (score of positive stained cells multiplied by the intensity of staining) (21). There were four categories based on the final staining scores as follows: 0, -; 1-4, +; 5-8, ++; and 9-12, +++. The results were evaluated as negative and moderate for- and + (grouped as negative) and positive for ++ and +++ (21).
Western blotting
Total protein was extracted using RIPA lysis buffer, proteinase inhibitor, phosphatase inhibitors and PMSF (all from Beyotime Institute of Biotechnology). Proteins (30 µg; quantified using the BCA method) and the molecular weight marker were separated by 10% SDS-PAGE (cat. no. P0015A; Beyotime Institute of Biotechnology). The separated proteins were subsequently transferred onto PVDF membranes, which were blocked with 5% non-fat milk in 0.1% Tween-20 in TBS (TBST) for 1 h at room temperature, and incubated overnight at 4˚C with rabbit polyclonal anti-NLRX1 (1:1,000), anti-TRAF6 (1:1,000), anti-NF-κB (1:500) and anti-β-actin (1:1,000; cat. no. ABP57456; Abbkine Scientific Co., Ltd.) diluted in TBST. Following the primary antibody incubation, the membranes were washed three times with TBS-T for 10 min and incubated for 1 h at room temperature with an anti-rabbit IgG HRP-conjugated secondary antibody (1:5,000; cat. no. A21020; Abbkine Scientific Co., Ltd.). BeyoECL Plus (Beyotime Institute of Biotechnology) was used to visualize the protein bands. All results were analyzed using Gel-Pro Analyzer (version 4.0; Media Cybernetics, Inc.).
Statistical analysis
Data are presented as the mean ± SD of three experimental repeats. Statistical analyses were performed using SPSS 20.0 software (IBM Corp.) and GraphPad Prism 7.0 software (GraphPad Software, Inc.). A χ2 and Fisher's exact test were used to determine the associations between NLRX1, TRAF6 and NF-κB expression levels and the clinicopathological parameters. A paired Student's t-test was used for the western blotting analysis. Spearman's rank correlation was used for correlation analyses. P<0.05 was considered to indicate a statistically significant difference.
Results
Demographic and clinicopathological characteristics of the enrolled patients
Demographic information was collected; the average age of the patients was 66.5±11.4 years, and there were 51 males and 9 females in the present study. Clinicopathological characteristics were also collected, including age, sex, tumor size, histologic grade, lymph node metastasis, vascular invasion, nerve invasion, depth of infiltration (22) and clinical stage (Table I).
NLRX1, TRAF6 and NF-κB protein expression levels in GC and normal gastric tissues
To investigate the roles of NLRX1, TRAF6 and NF-κB in the progression of GC, IHC staining was performed to evaluate the changes in the protein expression levels. IHC staining of NLRX1 was markedly reduced in the GC tissues compared with the normal tissues (Fig. 1). In contrast, IHC staining of TRAF6 and NF-κB were markedly increased in the GC tissues compared with the normal tissues.
Figure 1.
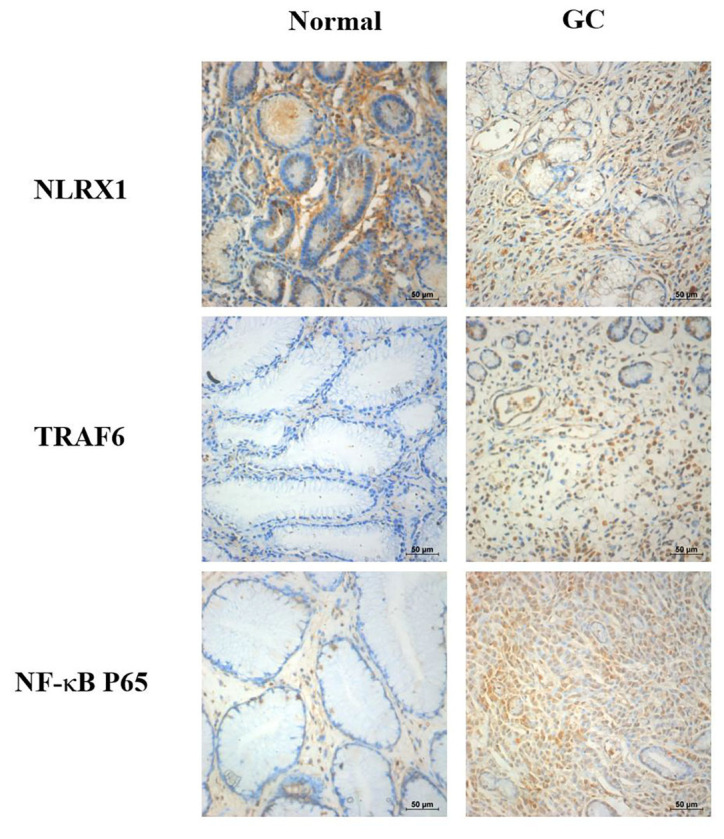
NLRX1, TRAF6 and NF-κB protein expression levels based on immunohistochemical assays. NLRX1 expression levels in the GC tissues were downregulated, while TRAF6 and NF-κB expression levels were upregulated in the GC tissues compared with the normal tissues. Scale bar, 50 µm. NLRX1, NOD-like receptor X1; TRAF6, tumor necrosis factor receptor-associated factor 6; GC, gastric cancer.
Association between NLRX1, TRAF6 and NF-κB expression levels and clinicopathological characteristics
NLRX1, TRAF6 and NF-κB IHC staining of GC samples and clinicopathological characteristics are presented in Table II. There were no significant differences identified between positive or negative expression levels of NLRX1, TRAF6 or NF-κB for either patient sex or age (≤65 or >65 years). However, significant differences were identified between positive or negative NLRX1, TRAF6 or NF-κB expression levels for tumor size (<5 cm or ≥5 cm), vascular invasion (yes or no), neural invasion (yes or no), lymph node metastasis (yes or no), differentiation (poorly or moderately-to-well differentiated), gross stage (early or advanced) and clinical stage (I/II or III/IV) (23).
Table II.
Association between NLRX1, TRAF6 and NF-κB expression levels and the clinicopathologic features of patients with gastric cancer.
| A, NLRX1 expression levels | ||||
|---|---|---|---|---|
| Clinicopathological variables | Number of patients | Positive expression | Negative expression | P-value |
| Age, years | 0.955 | |||
| ≤65 | 27 | 8 | 19 | |
| >65 | 33 | 10 | 23 | |
| Sex | 0.431 | |||
| Male | 51 | 14 | 37 | |
| Female | 9 | 4 | 5 | |
| Tumor size, cm | 0.012 | |||
| <5 | 28 | 13 | 15 | |
| ≥5 | 32 | 5 | 27 | |
| Vascular invasion | 0.010 | |||
| Yes | 35 | 6 | 29 | |
| No | 25 | 12 | 13 | |
| Neural invasion | <0.001 | |||
| Yes | 26 | 1 | 25 | |
| No | 34 | 17 | 17 | |
| Lymph node metastasis | 0.011 | |||
| Yes | 29 | 4 | 25 | |
| No | 31 | 14 | 17 | |
| Differentiation | 0.001 | |||
| Poorly | 27 | 2 | 25 | |
| Moderately-to-well | 33 | 16 | 17 | |
| Gross stage | 0.005 | |||
| Early | 8 | 6 | 2 | |
| Advanced | 52 | 11 | 41 | |
| Clinical stage | <0.001 | |||
| I/II | 34 | 17 | 17 | |
| III/IV | 26 | 1 | 25 | |
| B, TRAF6 expression levels | ||||
| Clinicopathological variables | Number of patients | Positive expression | Negative expression | P-value |
| Age, years | 0.180 | |||
| ≤65 | 27 | 17 | 10 | |
| >65 | 33 | 20 | 13 | |
| Sex | 0.284 | |||
| Male | 51 | 33 | 18 | |
| Female | 9 | 4 | 5 | |
| Tumor size, cm | <0.001 | |||
| <5 | 28 | 10 | 18 | |
| ≥5 | 32 | 27 | 5 | |
| Vascular invasion | <0.001 | |||
| Yes | 35 | 29 | 6 | |
| No | 25 | 8 | 17 | |
| Neural invasion | 0.002 | |||
| Yes | 26 | 22 | 4 | |
| No | 34 | 15 | 19 | |
| Lymph node metastasis | <0.001 | |||
| Yes | 29 | 25 | 4 | |
| No | 31 | 12 | 19 | |
| Differentiation | 0.001 | |||
| Poorly | 27 | 23 | 4 | |
| Moderately-to-well | 33 | 14 | 19 | |
| Gross stage | <0.001 | |||
| Early | 8 | 0 | 8 | |
| Advanced | 52 | 37 | 15 | |
| Clinical stage | 0.002 | |||
| I/II | 34 | 15 | 19 | |
| III/IV | 26 | 22 | 4 | |
| C, NF-κB expression levels | ||||
| Clinicopathological variables | Number of patients | Positive expression | Negative expression | P-value |
| Age, years | 0.957 | |||
| ≤65 | 27 | 17 | 10 | |
| >65 | 33 | 21 | 12 | |
| Sex | 0.111 | |||
| Male | 51 | 38 | 13 | |
| Female | 9 | 4 | 5 | |
| Tumor size, cm | 0.020 | |||
| <5 | 28 | 12 | 16 | |
| ≥5 | 32 | 26 | 6 | |
| Vascular invasion | 0.002 | |||
| Yes | 35 | 28 | 7 | |
| No | 25 | 10 | 15 | |
| Neural invasion | 0.017 | |||
| Yes | 26 | 21 | 5 | |
| No | 34 | 17 | 17 | |
| Lymph node metastasis | 0.007 | |||
| Yes | 29 | 24 | 5 | |
| No | 31 | 15 | 16 | |
| Differentiation | 0.015 | |||
| Poorly | 27 | 22 | 5 | |
| Moderately-to-well | 33 | 16 | 17 | |
| Gross stage | <0.001 | |||
| Early | 8 | 0 | 8 | |
| Advanced | 52 | 39 | 13 | |
| Clinical stage | 0.017 | |||
| I/II | 34 | 17 | 17 | |
| III/IV | 26 | 21 | 5 | |
NLRX1, NOD-like receptor X1; TRAF6, tumor necrosis factor receptor-associated factor 6.
NLRX1, TRAF6 and NF-κB expression levels based on western blotting
The results from the western blotting experiments revealed that NLRX1 expression levels were significantly downregulated in the GC tissues compared with the normal gastric tissues (Fig. 2). Conversely, TRAF6 and NF-κB expression levels were discovered to be significantly upregulated in the GC tissues compared with the normal tissues.
Figure 2.

NLRX1, TRAF6 and NF-κB protein expression levels were analyzed using western blotting. NLRX1 expression levels in the cancer tissues were significantly downregulated compared with the normal tissues, while TRAF6 and NF-κB expression levels in the GC tissues were significantly upregulated compared with the normal tissues. *P<0.05 vs. normal. NLRX1, NOD-like receptor X1; TRAF6, tumor necrosis factor receptor-associated factor 6; GC, gastric cancer.
Correlation analysis between NLRX1 and TRAF6, NLRX1 and NF-κB and TRAF6 and NF-κB expression levels
Spearman's rank correlation was used to determine the correlation between NLRX1 and TRAF6, NLRX1 and NF-κB and TRAF6 and NF-κB expression levels. A negative correlation was identified between NLRX1 and TRAF6 expression levels (Table III), NLRX1 and NF-κB protein expression levels (Table IV) and a positive correlation between TRAF6 and NF-κB protein expression levels in GC tissues (Table V).
Table III.
Correlation between NLRX1 and TRAF6 expression levels.
| NLRX1 expression | |||||||
|---|---|---|---|---|---|---|---|
| Staining score, (n) | - (23) | + (19) | ++ (14) | +++ (4) | Correlation coefficient | P-value | |
| TRAF6 expression | - (3) | - | - | 2 | 1 | rs=-0.635 | P<0.001 |
| + (20) | 2 | 7 | 8 | 3 | |||
| ++ (18) | 8 | 6 | 4 | - | |||
| +++ (19) | 13 | 6 | - | - | |||
NLRX1, NOD-like receptor X1; TRAF6, tumor necrosis factor receptor-associated factor 6.
Table IV.
Correlation between NLRX1 and NF-κB expression levels.
| NLRX1 expression | |||||||
|---|---|---|---|---|---|---|---|
| Staining score, (n) | - (23) | + (19) | ++ (14) | +++ (4) | Correlation coefficient | P-value | |
| NF-κB P65 expression | - (4) | - | 1 | 1 | 2 | rs=-0.530 | P<0.001 |
| + (18) | 3 | 6 | 8 | 1 | |||
| ++ (17) | 6 | 7 | 3 | 1 | |||
| +++ (21) | 14 | 5 | 2 | - | |||
NLRX1, NOD-like receptor X1.
Table V.
Correlation between TRAF6 and NF-κB expression levels.
| TRAF6 expression | |||||||
|---|---|---|---|---|---|---|---|
| Staining score, (n) | - (3) | + (20) | ++ (17) | +++ (20) | Correlation coefficient | P-value | |
| NF-κB P65 expression | - (4) | 1 | 3 | - | - | rs=0.781 | P<0.001 |
| + (18) | 2 | 14 | 2 | - | |||
| ++ (17) | - | 3 | 7 | 7 | |||
| +++ (21) | - | - | 8 | 13 | |||
TRAF6, tumor necrosis factor receptor-associated factor 6.
Discussion
The worldwide incidence of GC has decreased markedly in recent decades (24). Data from the 2014 Chinese National Cancer Center (NCCRC) have also revealed a decreased incidence of GA (25). Nevertheless, the poor prognosis among patients with GC remains a serious threat to global health. The incidence of GC varies by country and is 2-3-fold higher in males compared with females (26). In the present study, the male-to-female GC ratio was 51:9 (5.7-fold higher in males). The 2014 NCCRC reported a high incidence of GC in the 60-79-year group (27), which coincides with the age data (66.5±11.4 years) obtained in the present research.
NLRX1 is a negative regulator of antiviral immunity during the early stage of virus infection (16). With additional research involving NLRX1, other functions of NLRX1 have been discovered, such as its ability to regulate inflammation (28), metabolism (29) and development of histiocytic sarcoma (18). Previous studies have also reported that NLRX1 served as a tumor suppressor in colorectal cancer (30,31). For example, the deletion of NLRX1 in intestinal epithelial cells did not alter the architecture of the intestines, but there was an increased susceptibility among NLRX1-/- mice for developing colitis-associated colorectal cancer (30,31). Wang et al (32) also concluded that NLRX1 expression levels were downregulated in hepatocellular carcinoma tissues and that NLRX1 expression levels may be used as a prognostic marker in HCC hepatectomy. Castano-Rodriguez et al (33) reported that NLRX1 expression levels were downregulated in H. pylori-infected gastric tissues. However, the role of NLRX1 in GC has not been elucidated.
By examining IHC staining and western blotting of NLRX1 in GC and normal gastric tissues, the present study revealed that NLRX1 expression levels were downregulated in GC tissues, indicating that NLRX1 may be a tumor suppressor. The changes in NLRX1 expression levels were significantly associated with tumor size, vascular invasion, neural invasion, lymph node metastasis, differentiation, gross stage and clinical stage, which indicated that NLRX1 may serve as an index in assessing GC. However, no statistical differences were observed between NLRX1 expression levels and the age or sex of the patient. A previous study concluded that NLRX1 was a tumor suppressor in primary solid tumors of the breast (34). Hu et al (19) also reported that downregulated expression levels of NLRX1 were associated with liver cancer prognosis.
NF-κB is a pivotal mammalian transcription factor that was discovered to exert protumorigenic effects in liver (35), lung (36), breast (37) and prostate (38) cancer, in addition to in GC (39). In normal cells, NF-κB is located in the cytosol in the form of an inactive complex bound to IκBα. Once stimulated, IκBα is phosphorylated and separated from NF-κB for degradation (40). NF-κB has been demonstrated to remain in an active status in pancreatic cancer cells (41). The results of the present study revealed that NF-κB expression levels were upregulated in GC tissues through IHC staining and western blotting, which is consistent with the aforementioned studies.
microRNA-146a was discovered to upregulate NF-κB by targeting TRAF6 in human cervical cancer (42). In multiple myeloma, TRAF6 was discovered to mediate NF-κB activation and was suggested as a potential therapeutic target (43). Both of these studies illustrated that TRAF6 could regulate NF-κB as an upstream gene. However, to the best of our knowledge, the relationship between TRAF6 and NF-κB in GC remains to be clarified. It has also been reported that in response to lipopolysaccharide, NLRX1 interacted with TRAF6 and negatively regulated NF-κB activation in 293T cells (16,44). Allen et al (16) demonstrated that in response to viral infection, NLRX1 decreased the inflammatory responses by interacting with TRAF6.
TRAF6, as a TRAF protein family member, activates IκB kinase, thus resulting in the degradation of IκB and the activation of NF-κB (45,46). Therefore, a positive correlation is suggested to exist between TRAF6 and NF-κB. In the present study, a positive correlation was also discovered and TRAF6 and NF-κB expression levels, as tumor promotors, were increased and correlated with tumor size, vascular and neural invasion, lymph node metastasis, differentiation, gross stage and clinical stage.
In the present study, a negative correlation between NLRX1 and TRAF6 was discovered using Spearman's rank correlation. Therefore, it was speculated that NLRX1 may also interact with TRAF6 to exert an antitumor effect in GC.
In conclusion, NLRX1 expression levels were discovered to be downregulated in GC tissues and the expression levels of NLRX1 were associated with the clinicopathological characteristics of GC. A negative correlation was identified between NLRX1 and TRAF6/NF-κB expression levels, while a positive correlation was observed between TRAF6 and NF-κB expression levels. Thus, NLRX1 may be a potential biomarker for the diagnosis of GC and warrants further investigation.
Acknowledgements
Not applicable.
Funding
The present study was funded by the Dalian Medical Science Research Project (grant no. 1711038), the National Natural Science Foundation of China (grant nos. 81701965 and 81872255), the Key Medical Talents Fund of Jiangsu Province (grant no. 2016KJQWZDRC-03) and the Natural Science Foundation of Liaoning Province (grant nos. 20180550116 and 2019-MS-069).
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Authors' contributions
ZF and JP designed the study. JP contributed to the collection and storage of the tissues. HW analyzed the pathology. ZF, JP, HW and YZ performed the remaining experiments. YZ revised the manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to participate
The present study was approved by the Ethics Committee of The Third People's Hospital of Dalian (approval no. 2017-KY-004). Written informed consent regarding information and tissue samples were acquired from the patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
- 1.Comprehensive molecular characterization of gastric adenocarcinoma. Nature. 2014;513:202–209. doi: 10.1038/nature13480. Cancer Genome Atlas Research Network. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Gong Z, Mu Y, Chen J, Chu H, Lian P, Wang C, Wang J, Jiang L. Expression and significance of cyclophilin J in primary gastric adenocarcinoma. Anticancer Res. 2017;37:4475–4481. doi: 10.21873/anticanres.11843. [DOI] [PubMed] [Google Scholar]
- 3.Wadhwa R, Song S, Lee JS, Yao Y, Wei Q, Ajani JA. Gastric cancer-molecular and clinical dimensions. Nat Rev Clin Oncol. 2013;10:643–655. doi: 10.1038/nrclinonc.2013.170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Siegel RL, Miller KD, Jemal A. Cancer statistics, 2016. CA Cancer J Clin. 2016;66:7–30. doi: 10.3322/caac.21332. [DOI] [PubMed] [Google Scholar]
- 5.McGuire S. World Cancer Report 2014 Geneva, Switzerland: World Health Organization, International Agency for Research on Cancer, WHO Press, 2015. Adv Nutr. 2016;7:418–419. doi: 10.3945/an.116.012211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Graziosi L, Marino E, Donini A. Minimally invasive surgery for advanced gastric cancer: Are we sure? Gastric Cancer. 2017;20:1013–1014. doi: 10.1007/s10120-017-0727-4. [DOI] [PubMed] [Google Scholar]
- 7.Peddanna N, Holt S, Verma RS. Genetics of gastric cancer. Anticancer Res. 1995;15:2055–2064. [PubMed] [Google Scholar]
- 8.Lv Y, Zhao Y, Wang X, Chen N, Mao F, Teng Y, Wang T, Peng L, Zhang J, Cheng P, et al. Increased intratumoral mast cells foster immune suppression and gastric cancer progression through TNF-α-PD-L1 pathway. J Immunother Cancer. 2019;7(54) doi: 10.1186/s40425-019-0530-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Xie Y, Li F, Li Z, Shi Z. miR-135a suppresses migration of gastric cancer cells by targeting TRAF5-mediated NF-κB activation. Onco Targets Ther. 2019;12:975–984. doi: 10.2147/OTT.S189976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Deptala A, Bedner E, Gorczyca W, Darzynkiewicz Z. Activation of nuclear factor kappa B (NF-kappaB) assayed by laser scanning cytometry (LSC) Cytometry. 1998;33:376–382. doi: 10.1002/(sici)1097-0320(19981101)33:3<376::aid-cyto13>3.0.co;2-q. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Han F, Zhang L, Qiu W, Yi X. TRAF6 promotes the invasion and metastasis and predicts a poor prognosis in gastric cancer. Pathol Res Pract. 2016;212:31–37. doi: 10.1016/j.prp.2015.11.005. [DOI] [PubMed] [Google Scholar]
- 12.Inoue J, Gohda J, Akiyama T. Characteristics and biological functions of TRAF6. Adv Exp Med Biol. 2007;597:72–79. doi: 10.1007/978-0-387-70630-6_6. [DOI] [PubMed] [Google Scholar]
- 13.Sun YS, Ye ZY, Qian ZY, Xu XD, Hu JF. Expression of TRAF6 and ubiquitin mRNA in skeletal muscle of gastric cancer patients. J Exp Clin Cancer Res. 2012;31(81) doi: 10.1186/1756-9966-31-81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Maeda S, Yoshida H, Ogura K, Mitsuno Y, Hirata Y, Yamaji Y, Akanuma M, Shiratori Y, Omata M. H. pylori activates NF-kappaB through a signaling pathway involving IkappaB kinases, NF-kappaB-inducing kinase, TRAF2, and TRAF6 in gastric cancer cells. Gastroenterology. 2000;119:97–108. doi: 10.1053/gast.2000.8540. [DOI] [PubMed] [Google Scholar]
- 15.Takeuchi O, Akira S. Innate immunity to virus infection. Immun Rev. 2009;227:75–86. doi: 10.1111/j.1600-065X.2008.00737.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Allen IC, Moore CB, Schneider M, Lei Y, Davis BK, Scull MA, Gris D, Roney KE, Zimmermann AG, Bowzard JB, et al. NLRX1 protein attenuates inflammatory responses to infection by interfering with the RIG-I-MAVS and TRAF6-NF-κB signaling pathways. Immunity. 2011;34:854–865. doi: 10.1016/j.immuni.2011.03.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Moore CB, Bergstralh DT, Duncan JA, Lei Y, Morrison TE, Zimmermann AG, Accavitti-Loper MA, Madden VJ, Sun L, Ye Z, et al. NLRX1 is a regulator of mitochondrial antiviral immunity. Nature. 2008;451:573–577. doi: 10.1038/nature06501. [DOI] [PubMed] [Google Scholar]
- 18.Coutermarsh-Ott S, Simmons A, Capria V, LeRoith T, Wilson JE, Heid B, Philipson CW, Qin Q, Hontecillas-Magarzo R, Bassaganya-Riera J, et al. NLRX1 suppresses tumorigenesis and attenuates histiocytic sarcoma through the negative regulation of NF-κB signaling. Oncotarget. 2016;7:33096–33110. doi: 10.18632/oncotarget.8861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hu B, Ding GY, Fu PY, Zhu XD, Ji Y, Shi GM, Shen YH, Cai JB, Yang Z, Zhou J, et al. NOD-like receptor X1 functions as a tumor suppressor by inhibiting epithelial-mesenchymal transition and inducing aging in hepatocellular carcinoma cells. J Hematol Oncol. 2018;11(28) doi: 10.1186/s13045-018-0573-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Fléjou JF. WHO Classification of digestive tumors: The fourth edition. Ann Pathol. 2011;31 (5 Suppl):S27–S31. doi: 10.1016/j.annpat.2011.08.001. (In French) [DOI] [PubMed] [Google Scholar]
- 21.Zheng M, Zang S, Xie L, Fang X, Zhang YU, Ma X, Liu J, Lin D, Huang A. Rheb phosphorylation is involved in p38-regulated/activated protein kinase-mediated tumor suppression in liver cancer. Oncol Lett. 2015;10:1655–1661. doi: 10.3892/ol.2015.3406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lyros O, Thomaidis T, Muller M, Sivanathan V, Grimminger P, Lang H, Gockel I, Hartmann JT, Moehler M. External Validation of the Proposed Kiel Staging System and Comparison with the Old (6th edition) and the Currently Used (7th edition) TNM Classification in Gastric Cancer. Oncol Res Treat. 2018;41:122–128. doi: 10.1159/000484320. [DOI] [PubMed] [Google Scholar]
- 23.Harino Y, Imura S, Kanemura H, Morine Y, Fujii M, Ikegami T, Uehara H, Shimada M. Role of tumor angiogenesis in gallbladder carcinoma: With special reference to thymidine phosphorylase. Int J Clin Oncol. 2008;13:452–457. doi: 10.1007/s10147-008-0778-y. [DOI] [PubMed] [Google Scholar]
- 24.Bertuccio P, Chatenoud L, Levi F, Praud D, Ferlay J, Negri E, Malvezzi M, La Vecchia C. Recent patterns in gastric cancer: A global overview. Int J Cancer. 2009;125:666–673. doi: 10.1002/ijc.24290. [DOI] [PubMed] [Google Scholar]
- 25.Wang FH, Shen L, Li J, Zhou ZW, Liang H, Zhang XT, Tang L, Xin Y, Jin J, Zhang YJ, et al. The Chinese society of clinical oncology (CSCO): Clinical guidelines for the diagnosis and treatment of gastric cancer. Cancer Commun (Lond) 2019;39(10) doi: 10.1186/s40880-019-0349-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Makino Y, Nishimura Y, Oshita S, Mizosoe T, Akihiro T. Storage in high-barrier pouches increases the sulforaphane concentration in broccoli florets. PLoS One. 2018;13(e0192342) doi: 10.1371/journal.pone.0192342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Chen W, Sun K, Zheng R, Zeng H, Zhang S, Xia C, Yang Z, Li H, Zou X, He J. Cancer incidence and mortality in China, 2014. Chin J Cancer Res. 2018;30:1–12. doi: 10.21147/j.issn.1000-9604.2018.01.01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ma D, Zhao Y, She J, Zhu Y, Zhao Y, Liu L, Zhang Y. NLRX1 alleviates lipopolysaccharide-induced apoptosis and inflammation in chondrocytes by suppressing the activation of NF-κB signaling. Int Immunopharmacol. 2019;71:7–13. doi: 10.1016/j.intimp.2019.03.001. [DOI] [PubMed] [Google Scholar]
- 29.Scantlebery AML, Uil M, Butter LM, Poelman R, Claessen N, Girardin SE, Florquin S, Roelofs JJTH, Leemans JC. NLRX1 does not play a role in diabetes nor the development of diabetic nephropathy induced by multiple low doses of streptozotocin. PLoS One. 2019;14(e0214437) doi: 10.1371/journal.pone.0214437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Soares F, Tattoli I, Rahman MA, Robertson SJ, Belcheva A, Liu D, Streutker C, Winer S, Winer DA, Martin A, et al. The mitochondrial protein NLRX1 controls the balance between extrinsic and intrinsic apoptosis. J Biol Chem. 2014;289:19317–19330. doi: 10.1074/jbc.M114.550111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Tattoli I, Killackey SA, Foerster EG, Molinaro R, Maisonneuve C, Rahman MA, Winer S, Winer DA, Streutker CJ, Philpott DJ, Girardin SE. NLRX1 acts as an epithelial-intrinsic tumor suppressor through the modulation of TNF-mediated proliferation. Cell Rep. 2016;14:2576–2586. doi: 10.1016/j.celrep.2016.02.065. [DOI] [PubMed] [Google Scholar]
- 32.Wang X, Yang C, Liao X, Han C, Yu T, Huang K, Yu L, Qin W, Zhu G, Su H, et al. NLRC and NLRX gene family mRNA expression and prognostic value in hepatocellular carcinoma. Cancer Med. 2017;6:2660–2672. doi: 10.1002/cam4.1202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Castaño-Rodríguez N, Kaakoush NO, Goh KL, Fock KM, Mitchell HM. The NOD-like receptor signalling pathway in Helicobacter pylori infection and related gastric cancer: A case-control study and gene expression analyses. PLoS One. 2014;9(e98899) doi: 10.1371/journal.pone.0098899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Singh K, Poteryakhina A, Zheltukhin A, Bhatelia K, Prajapati P, Sripada L, Tomar D, Singh R, Singh AK, Chumakov PM, Singh R. NLRX1 acts as tumor suppressor by regulating TNF-α induced apoptosis and metabolism in cancer cells. Biochim Biophys Acta. 2015;1853:1073–1086. doi: 10.1016/j.bbamcr.2015.01.016. [DOI] [PubMed] [Google Scholar]
- 35.Zhang M, Pan L, Xu D, Cao C, Shi R, Han S, Liu J, Li X, Li M. The NFκB signaling pathway serves an important regulatory role in Klebsiella pneumoniae liver abscesses. Exp Ther Med. 2018;15:5443–5449. doi: 10.3892/etm.2018.6096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ahmmed B, Khan MN, Nisar MA, Kampo S, Zheng Q, Li Y, Yan Q. Tunicamycin enhances the suppressive effects of cisplatin on lung cancer growth through PTX3 glycosylation via AKT/NF-κB signaling pathway. Int J Oncol. 2019;54:431–442. doi: 10.3892/ijo.2018.4650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Bishop RT, Marino S, de Ridder D, Allen RJ, Lefley DV, Sims AH, Wang N, Ottewell PD, Idris AI. Pharmacological inhibition of the IKKε/TBK-1 axis potentiates the anti-tumour and anti-metastatic effects of Docetaxel in mouse models of breast cancer. Cancer Lett. 2019;450:76–87. doi: 10.1016/j.canlet.2019.02.032. [DOI] [PubMed] [Google Scholar]
- 38.Marino S, Bishop RT, Carrasco G, Logan JG, Li B, Idris AI. Pharmacological inhibition of NFκB reduces prostate cancer related osteoclastogenesis in vitro and osteolysis ex vivo. Calcif Tissue Int. 2019;105:193–204. doi: 10.1007/s00223-019-00538-9. [DOI] [PubMed] [Google Scholar]
- 39.Fu J, Yu L, Luo J, Huo R, Zhu B. Paeonol induces the apoptosis of the SGC-7901 gastric cancer cell line by downregulating ERBB2 and inhibiting the NF-κB signaling pathway. Int J Mol Med. 2018;42:1473–1483. doi: 10.3892/ijmm.2018.3704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Novack DV. Role of NF-κB in the skeleton. Cell Res. 2011;21:169–182. doi: 10.1038/cr.2010.159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Jeong Y, Lim JW, Kim H. Lycopene inhibits reactive oxygen species-mediated NF-κB signaling and induces apoptosis in pancreatic cancer cells. Nutrients. 2019;11(762) doi: 10.3390/nu11040762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Li T, Li M, Xu C, Xu X, Ding J, Cheng L, Ou R. miR-146a regulates the function of Th17 cell differentiation to modulate cervical cancer cell growth and apoptosis through NF-κB signaling by targeting TRAF6. Oncol Rep. 2019;41:2897–2908. doi: 10.3892/or.2019.7046. [DOI] [PubMed] [Google Scholar]
- 43.Morgan JJ, McAvera RM, Crawford LJ. TRAF6 silencing attenuates multiple myeloma cell adhesion to bone marrow stromal cells. Int J Mol Sci. 2019;20(702) doi: 10.3390/ijms20030702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Xia X, Cui J, Wang HY, Zhu L, Matsueda S, Wang Q, Yang X, Hong J, Songyang Z, Chen ZJ, Wang RF. NLRX1 negatively regulates TLR-induced NF-κB signaling by targeting TRAF6 and IKK. Immunity. 2011;34:843–853. doi: 10.1016/j.immuni.2011.02.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Paik JH, Jang JY, Jeon YK, Kim WY, Kim TM, Heo DS, Kim CW. MicroRNA-146a downregulates NFκB activity via targeting TRAF6 and functions as a tumor suppressor having strong prognostic implications in NK/T cell lymphoma. Clin Cancer Res. 2011;17:4761–4771. doi: 10.1158/1078-0432.CCR-11-0494. [DOI] [PubMed] [Google Scholar]
- 46.Yang WL, Wang J, Chan CH, Lee SW, Campos AD, Lamothe B, Hur L, Grabiner BC, Lin X, Darnay BG, Lin HK. The E3 ligase TRAF6 regulates Akt ubiquitination and activation. Science. 2009;325:1134–1138. doi: 10.1126/science.1175065. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.