

Abstract
Microglia-associated neuroinflammation plays an important role in the pathophysiology of ischemic stroke. Microglial activation and polarization, and the inflammatory response mediated by these cells play important roles in the development, progression and outcome of brain injury after ischemic stroke. Currently, there is no effective strategy for treating ischemic stroke in clinical practice. Therefore, it is clinically important to study the role and regulation of microglia in stroke. In this review, we discuss the involvement of microglia in the neuroinflammatory process in ischemic stroke, with the aim of providing a better understanding of the relationship between ischemic stroke and microglia.
Key Words: brain, cells, central nervous system, inflammation, injury, macrophages, repair, review, stroke
Introduction
Stroke is a major life-threatening disease worldwide (Dal-Ré, 2011), with a high disability and mortality rate, which places a huge burden on the economy and society (Bai et al., 2017). Ischemic stroke is the most common type, accounting for 87% of all strokes (Benjamin et al., 2018). The pathophysiological mechanisms of ischemic stroke are complex, involving microglia-mediated neuroinflammation, oxidative stress, excitotoxicity, and destruction of the blood-brain barrier (BBB) (Dirnagl et al., 1999; Allen and Bayraktutan, 2009; George and Steinberg, 2015). Thrombolytic therapy is currently recognized as the most effective method for the prevention and treatment of ischemic stroke, but it has a very narrow time window (Henderson et al., 2018). Thus, for patients who cannot receive thrombolytic therapy, we need to identify new therapeutic targets.
Numerous recent studies have shown that the microglia-mediated inflammatory response is an important pathological mechanism in ischemic stroke (Benarroch, 2013; Guruswamy and ElAli, 2017). Microglial cells derived from macrophages are important immune cells that play a critical role in the repair and regeneration of the central nervous system (CNS) (Xiong et al., 2016). Microglial cells are innate immune cells of the central nervous system (CNS) and are the main mediators of neuroinflammation (Amor and Woodroofe, 2014; Skaper et al., 2014). They can be activated by various stimuli to secrete factors that exert proinflammatory or anti-inflammatory effects (Herder et al., 2015; Xia et al., 2015; Yu et al., 2015). A recent study showed that M1 phenotype microglia have a destructive effect on the brain, while M2 phenotype microglia have neuroprotective effects (Hu et al., 2015). Therefore, it is beneficial for neuroprotection to promote the M2 phenotype and inhibit the M1 phenotype in microglia in ischemic stroke. Microglia are among the first cells to respond to stroke and other brain injuries (Streit et al., 2004; Kawabori and Yenari, 2015). Ischemic stroke and other brain injuries can cause local inflammatory responses, including activation of microglia (Heindl et al., 2018). In this review, we discuss the role of microglia in neuroinflammation and their impact on ischemic stroke. We also clarify the mechanisms underlying the role of microglia in stroke pathology, with a highlight on microglia-associated neuroinflammation, which may be a potential drug target. This review should provide new prospects and directions for the treatment of ischemic stroke.
Search Strategy
The articles included in this review were retrieved by an electronic search of the PubMed database from inception to 2020 for literature describing the role of microglia-associated neuroinflammation in ischemic stroke. Searches were conducted using the phrase “ischemic stroke” combined with the following key words: microglia, neuroinflammation, polarization, M1 phenotype, M2 phenotype, upstream and therapeutic target. Articles were included if they were deemed to contribute to the understanding of the role of microglia-associated neuroinflammation in ischemic stroke.
Features of Microglial Cells
In the 1920s and 1930s, Pio del Rio-Hortega first investigated the characteristics of microglia and described their unique morphological phenotypes (Penfield, 1932). Microglia are the most abundant of the resident macrophage populations in the CNS (Ransohoff and Perry, 2009). Under physiological conditions, microglia are in a resting state, branching with small cell bodies and long protrusions (Michell-Robinson et al., 2015). However, resting microglia are not completely inert—although they do not display phagocytic functions, they can still move and pinocytose (Cherry et al., 2014; Prinz and Priller, 2014), and can secrete growth factors that nourish, support and protect the electrophysiological functions of neurons to maintain CNS homeostasis (Eyo and Dailey, 2013). In addition, microglia can also dynamically monitor synaptic function through synaptic contact. In the CNS, abnormal synaptic transmission or dysfunction can trigger microglia to respond. In addition to clearing dead cell debris, Wake et al. (2013) showed that glial cells also help maintain normal synaptic function, trimming or clearing damaged or redundant synapses. Stimulated by ischemia, infection and other factors, microglia can be activated within minutes, and then proliferate, migrate to the diseased site, shorten their cell body protrusions, and change from a branched to a round or ameboid morphology. At this time, activated microglial cells proliferate, phagocytose and clear damaged and dead neurons (Jia, 2012). In the early stage of inflammation, microglia appear to be of the proinflammatory type, which is the classic activated (M1) type, secreting a variety of inflammatory factors. In the later stages of acute inflammation, these M1 microglia transform into the anti-inflammatory (M2) type, which secrete anti-inflammatory cytokines that contribute to neuroprotection (Liu et al., 2019). Microglia act as the first line of defense in the brain, and play a critical role in maintaining brain homeostasis (Perry et al., 2010; Xia et al., 2015). After an ischemic event, along with a reduction in cerebral blood flow, microglia undergo morphological changes in preparation for the upcoming immune response (Masuda et al., 2011). In a word, microglia are associated with neuroinflammation, which plays an important role in ischemic stroke.
Role of Microglia Associated Neuroinflammation in Ischemic Stroke
Microglia are major immune system cells, and they play an important role in the repair and regeneration of the CNS (Hanisch and Kettenmann, 2007). In addition to their neuroprotective effects, microglia are also the main producers of proinflammatory cytokines, which can greatly inhibit brain repair and promote neurogenesis (Ekdahl et al., 2003). The inflammatory microenvironment has a great impact on microglial phenotypic changes, and can change gene expression patterns and biological functions in brain tissue (Xiong et al., 2016).
Neuroinflammation, the inflammatory reaction in the CNS, is a key pathological event after ischemic stroke, and can lead to secondary brain tissue injury and poor functional recovery. During ischemic injury, microglial cells are activated and release neuroinflammatory factors. In addition, the peripheral immune cell infiltration following BBB breakdown further amplifies the neuroinflammatory response, eventually leading to neuronal damage (Rajkovic et al., 2018).
In ischemic stroke, various proinflammatory factors, such as tumor necrosis factor-α (TNF-α), interleukin (IL)-1β, IL-6 and interferon-γ (IFN-γ), chemokines, nitric oxide, reactive oxygen species (ROS) and matrix metallopeptidase-9, activate microglia, further aggravating the neuroinflammatory reaction in the brain and exacerbating brain damage (Luo et al., 2016). After ischemic stroke, activated microglial cells exhibit distinct phenotypes, namely M1 or M2, according to the affected brain area, and the specific site and severity of injury, and respond differently, either exerting proinflammatory or anti-inflammatory effects (Skirving and Dan, 2001; Sieweke and Allen, 2013). The different microglial phenotypes represent the two extremes of their activation status. M1 phenotypic polarization causes the release of proinflammatory mediators that inhibit CNS recovery, while M2 phenotypic polarization leads to the release of anti-inflammatory cytokines that promote tissue regeneration and repair. M1 microglia mainly secrete proinflam-matory factors, such as IL-1, TNF-α, IL-6 and IFN-γ, to increase expression of inducible nitric oxide synthase, which has cytotoxic effects on neurons, resulting in neuronal loss (Lan et al., 2017). This leads to disruption of the BBB and the degradation of the extracellular matrix (Haley and Lawrence, 2017). Subsequently, peripheral blood leukocytes, plasma fibronectin, fibrinogen and other fibrinolytic factors enter the brain tissue (Dejonckheere et al., 2011), aggravating tissue damage (Anrather and Iadecola, 2016). Compared with M1 microglia, M2 microglia have a stronger ability to induce phagocytosis of dead neurons, which can curtail the secondary inflammatory response and promote tissue regeneration (Xia et al., 2015). The M2 phenotype can be divided into M2a, M2b and M2c subtypes, based on function and stimulatory factors (Hu et al., 2012; Chhor et al., 2013; Sudduth et al., 2013; Franco and Fernán-dez-Suárez, 2015; Latta et al., 2015). The M2a subtype is induced by IL-4 and IL-13 stimulation (Latta et al., 2015). The M2b subtype is an immunomodulatory phenotype that can be induced by lipopolysaccharide (LPS) and IL-1 receptor agonists (Chhor et al., 2013; Franco and Fernández-Suárez, 2015). The M2c subtype, called deactivated microglia, are stimulated by transforming growth factor-β, IL-10 and glucocorticoids (Franco and Fernández-Suárez, 2015). The M2 phenotype mainly secretes anti-inflammatory factors, such as IL-3, IL-4, IL-10, transforming growth factor-β and insulin-like growth factor-1, to inhibit inflammation (Xiong et al., 2016). M2 microglia also produce the antioxidative factors hemeoxygenase-1 and glutathione, which may inhibit the recognition of viable neurons, promoting viable neurons repair (Xia et al., 2015). It was found that activated M1 phenotypic microglia can be reactivated to the M2 phenotype under certain conditions. M1 polarized microglia can inhibit the actions of M2 polarized microglia to a degree. Therefore, microglia may produce proinflammatory or anti-inflammatory cytokines and chemokines (Lan et al., 2017), which have differential effects on neuroinflammation in ischemic stroke (Figure 1).
Figure 1.
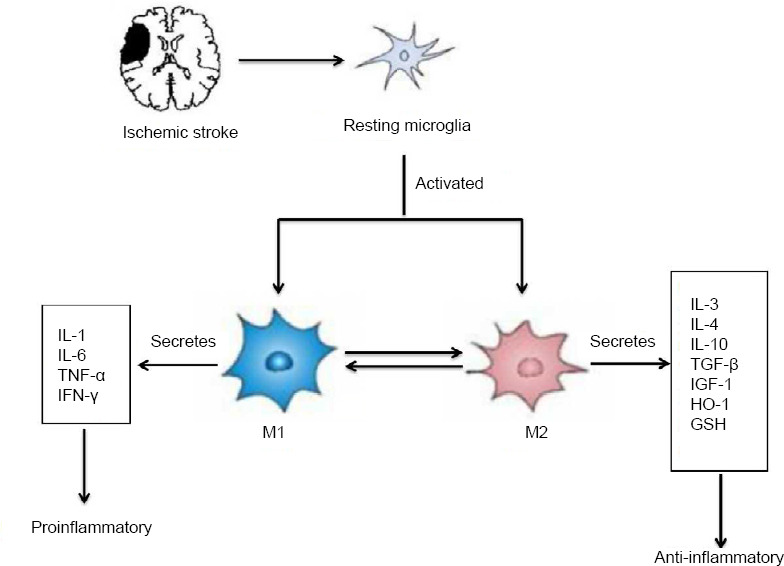
Phenotypic polarization of microglia.
Microglia become polarized towards the M1 or M2 phenotype at various stages after central nervous system injuries, and have distinct roles in the restoration of the neurovascular network. M1 phenotypic microglia mainly secrete proinflammatory factors such as IL-1, IL-6, TNF-α and IFN-γ, which have cytotoxic effects on neurons, resulting in neuronal loss. The M2 phenotype mainly secretes anti-inflammatory factors such as IL-3, IL-4, IL-10, TGF-β and IGF-1 to inhibit inflammation. M2 microglia also produce the anti-oxidative factors HO-1 and GSH, which may reduce neuronal loss and promote neuronal repair. GSH: Glutathione; HO-1: heme oxygenase-1; IFN-γ: interferon-γ; IGF-1: insulin-like growth factor-1; IL: interleukin; TGF-β: transforming growth factor-β; TNF-α: tumor necrosis factor-α.
Proinflammatory response of microglial cells in ischemic stroke
Neuroinflammation is considered a major contributor to cerebral ischemia-induced brain damage. Neuroinflammation is initiated in response to ischemic stroke, and is usually characterized by microglial activation and collateral brain damage, and is caused by a strong inflammatory reaction. Excessive activation of microglial cells results in the release of a large number of proinflammatory factors that aggravate cerebral ischemia-reperfusion injury (Ekdahl et al., 2003; Xiong et al., 2016). Therefore, inhibiting the release of proinflammatory factors from microglial cells could be a major strategy for alleviating ischemic stroke (Lu et al., 2019). The proinflammatory M1 microglia can kill pathogens and promote tissue repair by enhancing phagocytosis. However, excessive release of IL-6, IL-1β, IL-12, IL-23 and TNF-α and the production of other proinflammatory factors such as inducible nitric oxide synthase and ROS can cause in-flammation and aggravate brain damage (Xiong et al., 2016; Voet et al., 2018).
Studies have shown that the neuroinflammatory response mediated by microglia may be triggered by substances that stimulate M1 phenotypic microglia to produce proinflammatory factors (He et al., 2016, 2017). It is well known that LPS induces an inflammatory cytokine response in mice. LPS induces microglial activation through the mitogen-activated protein kinase pathway and increases the expression of proinflammatory cytokines (He et al., 2017; Ding et al., 2019). He et al. (2016) found that chemokine ligand 2 and chemokine ligand receptor 2 mediate the polarization of microglia into the M1 phenotype and promote the release of inflammatory factors during CNS inflammation. Indeed, inflammatory cytokines, such as IL-1β, IL-6 and TNF-α, are significantly increased in mice with ischemic stroke (Hénaut et al., 2019). Among these cytokines, TNF-α is mainly secreted by microglia, and is an important factor contributing to ischemic/hypoxic injury. Microglia can stimulate the activation of inflammatory cells, such as neutrophils, and promote inflammatory responses (Wang et al., 2013). Chen et al. (2019) further demonstrated that M1 phenotypic microglia can cause BBB destruction after ischemic stroke, mainly because these cells secrete TNF-α, thereby promoting the inflammatory response. Therefore, finding an effective clinical drug to block microglial-associated neuroinflammation may reduce BBB breakdown and alleviate tissue damage after ischemic stroke. Triggering receptor expressed on myeloid cells 1 (TREM1), an amplifier of the innate immune response, is a critical regulator of inflammation. It has been found that TREM1 is upregulated after cerebral ischemic injury and accelerates the neuroinflammatory response (Xu et al., 2019). Soluble Fas ligand (sFasL) plays an important role in M1 microglial migration and the inflammatory response in the mouse model of middle cerebral artery occlusion. sFasL can increase M1 microglia, exacerbating neuroinflammation (Meng et al., 2016). Therefore, neutralizing sFasL is a potential new treatment strategy to inhibit post-stroke inflammation and M1 phenotypic microglial polarization, which may improve stroke prognosis. Papaverine inhibits the production of nitric oxide and proinflammatory cytokines by microglia stimulated by LPS by regulating various inflammatory signals (Lee et al., 2019). While the mechanisms by which microglia promote neuroinflammation remain unclear, the accumulating evidence suggests that M1 phenotypic microglial polarization promotes an inflammatory response and is a risk factor for ischemic stroke. Therefore, inhibiting the polarization of microglia into the M1 phenotype may have therapeutic potential for the treatment of stroke.
Anti-inflammatory effect of microglial cells in ischemic stroke
Activated microglial cells have different roles in the CNS inflammatory environment. The M2 phenotype microglial cells that are activated after ischemic stroke promote brain tissue repair mainly by of the removal of cell debris, the reduction of local inflammation, and the release of a large number of trophic factors (Hanisch and Kettenmann, 2007; Thored et al., 2009; Kwon et al., 2013; Miron et al., 2013). M2 microglia have an anti-inflammatory role and secretes anti-inflammatory mediators and neurotrophic factors, such as IL-10, IL-13, transforming growth factor-β, brain-derived neurotrophic factor, glial neurotrophic factor, to reduce inflammatory response, protect neurons, and promote tissue repair. Perego et al. (2011) showed that selectively activated M2 phenotypic microglia secrete various anti-inflammatory factors such as IL-4, IL-10 and transforming growth factor-β, which are of great significance to promote brain repair and neuron regeneration in ischemic stroke. Adenosine 5'-monophosphate-activated protein kinase (AMPK) is the main molecular switch that activates the M2 phenotype (Mounier et al., 2013). Activation of the AMPK pathway by synthetic or natural compounds can reduce inflammatory damage and promote microglial M2 polarization (Lu et al., 2010; Zhou et al., 2014; Zhu et al., 2019). ROS produced by NADPH oxidase in microglia play an important role in neuronal injury after ischemic stroke. The voltage-gated proton channel Hv1 is selectively ex-pressed in microglia and is necessary for the production of ROS by NADPH oxidase in the brain. Compared with wild-type mice, mice lacking Hv1 exhibit reduced ROS production, and polarized microglia are transformed from the M1 to the M2 phenotype, which slows brain tissue damage (Tian et al., 2016). These observations suggest that microglial cells can reduce inflammation and tissue damage in the infarcted area.
Studies have demonstrated that inhibition of the suppressor of cytokine signaling 1 (SOCS1)/Janus kinase 2 (JAK2)/STAT3 signaling pathway promotes M2 phenotypic polarization, thereby reducing the neuroinflammatory response in rats with ischemic stroke (Qin et al., 2012; Tian et al., 2016; Wang et al., 2017b; Li et al., 2019a). The nuclear hormone receptor peroxisome proliferator-activated receptor γ is another transcription factor that regulates the differentiation of monocytes into M2 phenotypic microglia, and has anti-inflammatory effects, thereby reducing neuroinflammation (Bouhlel et al., 2007). The various subtypes of interferon regulatory factors play differing roles in microglial polarization. For example, interferon regulatory factor– 4 is a key transcription factor that controls M2 polarization (Satoh et al., 2010) and reduces inflammation and tissue damage in the infarcted area. Further studies on the functional characteristics of activated microglia are needed to determine how to polarize microglia toward the M2 phenotype and maintain them in an anti-inflammatory state in the brain. Finding a way to induce microglial cells to undergo M2 phenotypic polarization is therefore a potential new approach for the treatment of ischemic stroke (Figure 2).
Figure 2.
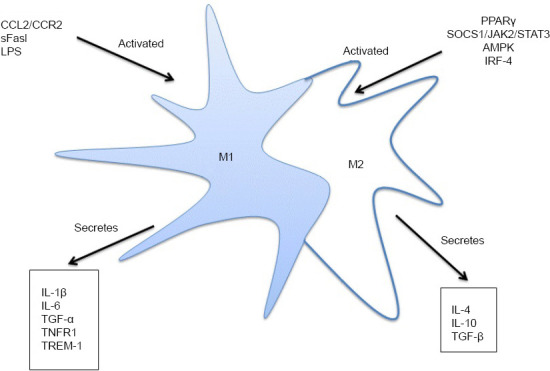
Activated microglial cells secrete inflammatory cytokines.
CCL2/CCR2, sFasL and LPS can activate M1 microglial cells, and activated M1 microglial cells can secrete inflammatory cytokines, such as IL-1β, IL-6 and TNF-α. PPARγ, SOCS1/JAK2/STAT3, AMPK and IRF-4 can activate M2 microglial cells, and activated M2 microglial cells can secrete inflammatory cytokines, such as IL-4, IL-10 and TGF-β. AMPK: Adenosine 5'-monophosphate-activated protein kinase; CCL2: chemokine ligand-2; CCR2: chemokine ligand receptor-2; IL: interleukin; IRF-4: interferon regulatory factor-4; LPS: lipopolysaccharide; PPARγ: peroxisome proliferator-activated receptor γ; sFasL: soluble Fas ligand; SOCS1/JAK2/STAT3: suppressor of cytokine signaling 1/Janus kinase 2/signal transducer and activator of transcription 3; TGF-β: transforming growth factor-β; TNFR1: tumor necrosis factor receptor 1; TNF-α: tumor necrosis factor-α; TREM-1: triggering receptor expressed on myeloid cells-1.
Upstream Regulators of Microglia-Associated Neuroinflammation in Ischemic Stroke
Microglial cells are highly plastic. Microglial polarization and microglial-associated neuroinflammation are dynamic processes. In the early stage of cerebral ischemia, inhibiting microglial polarization into the M1 phenotype and promoting their polarization into the M2 phenotype, to maintain the balance between anti-inflammatory and proinflammatory responses, is a potential new strategy for the treatment of ischemic stroke.
The signal transducer and activator of transcription (STAT) family of transcription factors play a key role in cytokine and immune regulation. For example, STAT6 is important in establishing the M2 phenotype following IL-13 and IL-4 stimulation (Kuroda et al., 2009). The STAT3 signaling pathway is thought to be an important pathway that regulates microglial phenotype and participates in the microglial-associated inflammatory response in ischemic stroke (Qin et al., 2012). Downregulation of the STAT3 signaling pathway can inhibit the death of neurons and promote microglial polarization into the M2 phenotype in ischemic stroke (Li et al., 2019a). sFasL activates microglia and promotes M1 polarization by phosphorylating the JAK2/STAT3/nuclear factor-κB (NF-κB) pathway in ischemic stroke, while AG490 (a JAK2 antagonist) not only inhibits STAT3/NF-κB phosphorylation, but sFasL-induced M1 phenotypic polarization and inflammation as well (Meng et al., 2016). Recent studies have shown that inhibiting the SOCS1/JAK2/STAT3 pathway can lower the loss of neural function and the apoptosis of neuronal cells, and may therefore serve as a new target for the clinical treatment of ischemic stroke. In addition, it has been reported that inhibition of the SOCS1/JAK2/STAT3 pathway reduces the neuroinflammatory response in the brain of rats with ischemic stroke (Wang et al., 2017b).
Exosomes are efficient tools for delivering gene-based drugs to the ischemic cortex. For example, exosomes loaded with miR-30d-5p can protect against ischemia and autophagy-mediated brain damage by promoting M2 microglial polarization. In the acute phase of stroke, the use of exosomes from adipose-derived stem cells overexpressing miR-30d-5p may reduce brain damage by inhibiting the inflammatory response (Jiang et al., 2018b). Therefore, exosomes loaded with mir-30d-5p have the potential to be a therapeutic strategy for ischemic stroke by promoting M2 microglial polarization.
In ischemic stroke, class A scavenger receptor (SR-A) and cannabinoid 2 receptor are involved in the regulation of microglial-associated neuroinflammation. After ischemic stroke, the SR-A receptor promotes microglial M1 phenotypic polarization by activating NF-κB. The number of M1 microglial cells therefore increases rapidly, and the inflammatory response and brain damage worsen. In SR-A knockout mice, compared with control mice, the number of M1 microglial cells in ischemic tissue is decreased and the number of M2 microglial cells is increased (Xu et al., 2012). IL-10 is an anti-inflammatory cytokine that inhibits inflammatory responses and antigen presentation. For example, compared with normal mice, mice with IL-10 deficiency have larger and more severe lesions, and worse prognosis after brain injury. M2 microglia in mice lacking IL-10 are significantly decreased, suggesting that IL-10 promotes M2 polarization and neuronal recovery, and reduces the inflammatory response (De Córdoba et al., 2015). Microglial-associated neuroinflammation is a complex process involving intricate molecular regulatory pathways. Therefore, identifying key targets upstream of microglial-associated neuroinflammation will likely contribute to more effective treatment of ischemic stroke.
Therapeutic Targeting of Microglia-Associated Neuroinflammation in Ischemic Stroke
Ischemic stroke is a complex process involving multiple factors, interactions and targets. Therefore, the treatment of ischemic stroke is also a complex process involving multiple links and targets. Intravenous thrombolysis and mechanical thrombectomy are effective strategies for the treatment of ischemic stroke (Badhiwala et al., 2016). However, because of the narrow treatment time window, few patients can receive intravenous thrombolysis and mechanical thrombectomy. However, we can selectively develop new drugs and identify new targets for the prevention and treatment of ischemic stroke based on the dual nature of microglial activation after stroke. For example, rosiglitazone is thought to improve ischemia-induced white matter damage by promoting microglial polarization into the anti-inflammatory M2 phenotype (Han et al., 2015). Therefore, promoting microglial M2 polarization may be an effective treatment strategy for ischemic stroke. Curcumin inhibits LPS and IFN-γ-induced M1 polarization, while promoting M2 polarization and inhibiting microglial-mediated proinflammatory responses (Liu et al., 2017). Therefore, curcumin can maintain the M1/M2 phenotypic balance and exert a protective effect in ischemic stroke. It was reported that in the rat middle cerebral artery occlusion model of stroke, α-lipoic acid regulates microglial M1/M2 polarization, reduces proinflammatory cytokine expression, increases anti-inflammatory cytokine levels and inhibits the NF-κB pathway, thereby exerting a neuroprotective effect (Wang et al., 2018). Furthermore, the histone deacetylase inhibitors valproic acid and sodium butyrate can inhibit the activation of microglia, reduce the number of microglia, and inhibit other inflammatory effectors in the ischemic brain (Kim et al., 2007). M1 microglia produce matrix metallopeptidase-9, and inhibiting this enzyme has therapeutic potential for ischemic stroke. Recent clinical trials have shown that the efficacy of minocycline in improving NIH stroke scale scores is negligible. However, it remains unclear how minocycline regulates microglial-associated neuroinflammation (Machado et al., 2006; Switzer et al., 2011; Veltkamp and Gill, 2016; Serhan et al., 2020). The clinical trial of Ma et al. (2017) showed that chronic metformin treatment suppresses the expression of M1 phenotype-associated genes (CD32, IL-1b, CD16) and enhances the expression of M2-associated genes (CD206, Arg1), providing new avenues for the treatment of ischemic stroke. In addition, one study reported that glycine inhibits M1 polarization. Glycine is a simple nonessential amino acid and is known to have neuroprotective functions. Glycine is an important component of many proteins and is also a major inhibitory neurotransmitter that binds to glycine receptors to inhibit postsynaptic neurons in ischemic stroke (Stephan and Friauf, 2014). In the pathophysiology of ischemic stroke, glycine inhibits phosphatase and tensin homologue deleted on chromosome ten (PTEN), which activates protein kinase B and inhibits NF-κB p65 and hypoxia-inducible factor-1α, thereby inhibiting ischemia-induced M1 microglial polarization and exerting anti-inflammatory effects, thereby indirectly reducing ischemia-induced neuronal death (Liu et al., 2019). Therefore, treatment with glycine can reduce infarct volume, improve neurologic function, and reduce neuronal and microglial death in ischemic stroke injury. These findings offer new therapeutic strategies and approaches for targeting microglia-associated neuroinflammation in ischemic stroke. Table 1 summarizes the use of microglia in the treatment of ischemic stroke.
Table 1.
Potential targets of microglia-associated neuroinflammation for the treatment of ischemic stroke
| Target | Polarization | Conclusion | Reference |
| IL-13 | M2 | Peripheral administration of IL-13 induces anti-inflammatory microglial responses and provides neuroprotection in ischemic stroke. | Kolosowska et al. (2019) |
| IL-33/ST2 | M2 | M2 phenotype microglial cells can prevent brain damage caused by acute ischemic stroke through the IL-33/ST2 axis. | Jiang et al. (2018a) |
| LncRNA-1810034E14Rik | No polar | LncRNA-1810034E14Rik plays an anti-inflammatory role in ischemic stroke and regulates p65 phosphorylation. | Zhang et al. (2019) |
| Nrf2 | No polar | The induction of Nrf2 by D3T is required for its suppressive effect on microglia activation. | Kuo et al. (2017) |
| TXA2R | No polar | TXA2R antagonist SQ29548 reduces ischemic stroke-induced microglia activation and enrichment. | Yan et al. (2016) |
| LncRNA-H19 | M1 | LncRNA-H19 promotes neuroinflammation by driving HDAC1-dependent M1 microglial polarization. | Wang et al. (2017a) |
| TLR4 | M1 | A 21-day of alcohol exposure during adolescence induces a phagocytic activated proinflammatory state of M1 microglia through TLR4 activation. | Li et al. (2019b) |
D3T: 3H-1,2-dithiole-3-thione; HDAC1: histone deacetylase 1; IL: interleukin; lncrna: long non-coding RNA; ST2: suppression of tumorigenicity 2; TLR4: Toll-like- receptor 4; TXA2R: thromboxane A2 receptor.
Conclusion
Inflammatory responses mediated by microglial activation play a key pathogenetic role in ischemic stroke. Microglial cells can rapidly undergo morphological changes based on subtle changes in the brain environment. M1 phenotype microglial cells are associated with an increased inflammatory response, which damages the BBB by increasing the release of proinflammatory factors, thereby aggravating damage to ischemic brain tissue. In contrast, M2 phenotype microglia promote neuronal repair and regeneration by secreting anti-inflammatory and neurotrophic factors. Therefore, modulating microglial polarization in the different stages of ischemic stroke will be a good strategy for the treatment of ischemic stroke by reducing neural inflammation. However, the classification and regulation of microglia after ischemic stroke are extremely complex, and more studies are needed to further elucidate their pathophysiological role. An important component of this needed research is a comprehensive and in-depth study of the mechanisms of microglia-associated neuroinflammation. The results of this research will likely help provide new approaches for the prevention and treatment of ischemic stroke.
Footnotes
Conflicts of interest: The authors declare no conflicts of interest.
Financial support: This work was supported by the National Natural Science Foundation of China, Nos. 31871169 (to YT), 81600040 (to APW); Key Project of Department of Education of Hunan Province, China, No. 18A243 (to APW); Innovation Guidance Project of Hunan Province, China, No. 2018SK51606 (to SXG); the Natural Science Foundation of Hunan Province of China, No. 2017JJ3279 (to APW). The funding sources had no role in study conception and design, data analysis or interpretation, paper writing or deciding to submit this paper for publication.
Copyright license agreement: The Copyright License Agreement has been signed by all authors before publication.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Funding: This work was supported by the National Natural Science Foundation of China, Nos. 31871169 (to YT), 81600040 (to APW); Key Project of Department of Education of Hunan Province, China, No. 18A243 (to APW); Innovation Guidance Project of Hunan Province, China, No. 2018SK51606 (to SXG); the Natural Science Foundation of Hunan Province of China, No. 2017JJ3279 (to APW).
C-Editor: Zhao M; S-Editors: Yu J, Li CH; L-Editors: Patel D, Yu J, Song CP; T-Editor: Jia Y
References
- 1.Allen CL, Bayraktutan U. Oxidative stress and its role in the pathogenesis of ischaemic stroke. Int J Stroke. 2009;4:461–470. doi: 10.1111/j.1747-4949.2009.00387.x. [DOI] [PubMed] [Google Scholar]
- 2.Amor S, Woodroofe MN. Innate and adaptive immune responses in neurodegeneration and repair. Immunology. 2014;141:287–291. doi: 10.1111/imm.12134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Anrather J, Iadecola C. Inflammation and stroke: an overview. Neurotherapeutics. 2016;13:661–670. doi: 10.1007/s13311-016-0483-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Badhiwala JH, Manoranjan B, Almenawer SA. Mechanical thrombectomy for ischemic stroke. J Am Coll Cardiol. 2016;67:2449–2450. doi: 10.1016/j.jacc.2016.02.068. [DOI] [PubMed] [Google Scholar]
- 5.Bai B, Yan Z, Hao Y, Zhang Z, Li G, Dekker J, Qiu C. A randomised controlled multimodal intervention trial in patients with ischaemic stroke in Shandong, China: design and rationale. Lancet. 2017;390:S13. [Google Scholar]
- 6.Benarroch EE. Microglia: Multiple roles in surveillance, circuit shaping, and response to injury. Neurology. 2013;81:1079–1088. doi: 10.1212/WNL.0b013e3182a4a577. [DOI] [PubMed] [Google Scholar]
- 7.Benjamin EJ, Virani SS, Callaway CW, Chamberlain AM, Chang AR, Cheng S, Chiuve SE, Cushman M, Delling FN, Deo R, de Ferranti SD, Ferguson JF, Fornage M, Gillespie C, Isasi CR, Jiménez MC, Jordan LC, Judd SE, Lackland D, Lichtman JH, et al. Heart disease and stroke statistics-2018 update: a report from the American Heart Association. Circulation. 2018;137:e67–e492. doi: 10.1161/CIR.0000000000000558. [DOI] [PubMed] [Google Scholar]
- 8.Bouhlel MA, Derudas B, Rigamonti E, Dièvart R, Brozek J, Haulon S, Zawadzki C, Jude B, Torpier G, Marx N, Staels B, Chinetti-Gbaguidi G. PPARgamma activation primes human monocytes into alternative M2 macrophages with anti-inflammatory properties. Cell Metab. 2007;6:137–143. doi: 10.1016/j.cmet.2007.06.010. [DOI] [PubMed] [Google Scholar]
- 9.Chen AQ, Fang Z, Chen XL, Yang S, Zhou YF, Mao L, Xia YP, Jin HJ, Li YN, You MF, Wang XX, Lei H, He QW, Hu B. Microglia-derived TNF-α mediates endothelial necroptosis aggravating blood brain-barrier disruption after ischemic stroke. Cell Death Dis. 2019;10:487. doi: 10.1038/s41419-019-1716-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cherry JD, Olschowka JA, O’Banion MK. Neuroinflammation and M2 microglia: the good, the bad, and the inflamed. J Neuroinflammation. 2014;11:98. doi: 10.1186/1742-2094-11-98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Chhor V, Le Charpentier T, Lebon S, Oré MV, Celador IL, Josserand J, Degos V, Jacotot E, Hagberg H, Sävman K, Mallard C, Gressens P, Fleiss B. Characterization of phenotype markers and neuronotoxic potential of polarised primary microglia in vitro. Brain Behav Immun. 2013;32:70–85. doi: 10.1016/j.bbi.2013.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Dal-Ré R. Worldwide clinical interventional studies on leading causes of death: a descriptive analysis. Ann Epidemiol. 2011;21:727–731. doi: 10.1016/j.annepidem.2011.03.010. [DOI] [PubMed] [Google Scholar]
- 13.De Córdoba JL, García Bach M, Isach N, Piles S. Percutaneous balloon compression for trigeminal neuralgia: imaging and technical aspects. Reg Anesth Pain Med. 2015;40:616–622. doi: 10.1097/AAP.0000000000000292. [DOI] [PubMed] [Google Scholar]
- 14.Dejonckheere E, Vandenbroucke RE, Libert C. Matrix metalloproteinases as drug targets in ischemia/reperfusion injury. Drug Discov Today. 2011;16:762–778. doi: 10.1016/j.drudis.2011.06.009. [DOI] [PubMed] [Google Scholar]
- 15.Ding YH, Song YD, Wu YX, He HQ, Yu TH, Hu YD, Zhang DP, Jiang HC, Yu KK, Li XZ, Sun L, Qian F. Isoalantolactone suppresses LPS-induced inflammation by inhibiting TRAF6 ubiquitination and alleviates acute lung injury. Acta Pharmacol Sin. 2019;40:64–74. doi: 10.1038/s41401-018-0061-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dirnagl U, Iadecola C, Moskowitz MA. Pathobiology of ischaemic stroke: an integrated view. Trends Neurosci. 1999;22:391–397. doi: 10.1016/s0166-2236(99)01401-0. [DOI] [PubMed] [Google Scholar]
- 17.Ekdahl CT, Claasen JH, Bonde S, Kokaia Z, Lindvall O. Inflammation is detrimental for neurogenesis in adult brain. Proc Natl Acad Sci U S A. 2003;100:13632–13637. doi: 10.1073/pnas.2234031100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Eyo UB, Dailey ME. Microglia: key elements in neural development, plasticity, and pathology. J Neuroimmune Pharmacol. 2013;8:494–509. doi: 10.1007/s11481-013-9434-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Franco R, Fernández-Suárez D. Alternatively activated microglia and macrophages in the central nervous system. Prog Neurobiol. 2015;131:65–86. doi: 10.1016/j.pneurobio.2015.05.003. [DOI] [PubMed] [Google Scholar]
- 20.George PM, Steinberg GK. Novel stroke therapeutics: unraveling stroke pathophysiology and its impact on clinical treatments. Neuron. 2015;87:297–309. doi: 10.1016/j.neuron.2015.05.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Guruswamy R, ElAli A. Complex roles of microglial cells in ischemic stroke pathobiology: new insights and future directions. Int J Mol Sci. 2017;18:496. doi: 10.3390/ijms18030496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Haley MJ, Lawrence CB. The blood-brain barrier after stroke: Structural studies and the role of transcytotic vesicles. J Cereb Blood Flow Metab. 2017;37:456–470. doi: 10.1177/0271678X16629976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Han L, Cai W, Mao L, Liu J, Li P, Leak RK, Xu Y, Hu X, Chen J. Rosiglitazone promotes white matter integrity and long-term functional recovery after focal cerebral ischemia. Stroke. 2015;46:2628–2636. doi: 10.1161/STROKEAHA.115.010091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hanisch UK, Kettenmann H. Microglia: active sensor and versatile effector cells in the normal and pathologic brain. Nat Neurosci. 2007;10:1387–1394. doi: 10.1038/nn1997. [DOI] [PubMed] [Google Scholar]
- 25.He HQ, Wu YX, Nie YJ, Wang J, Ge M, Qian F. LYRM03, an ubenimex derivative, attenuates LPS-induced acute lung injury in mice by suppressing the TLR4 signaling pathway. Acta Pharmacol Sin. 2017;38:342–350. doi: 10.1038/aps.2016.141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.He M, Dong H, Huang Y, Lu S, Zhang S, Qian Y, Jin W. Astrocyte-derived CCL2 is associated with M1 activation and recruitment of cultured microglial cells. Cell Physiol Biochem. 2016;38:859–870. doi: 10.1159/000443040. [DOI] [PubMed] [Google Scholar]
- 27.Heindl S, Gesierich B, Benakis C, Llovera G, Duering M, Liesz A. Automated morphological analysis of microglia after stroke. Front Cell Neurosci. 2018;12:106. doi: 10.3389/fncel.2018.00106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Hénaut L, Grissi M, Brazier F, Assem M, Poirot-Leclercq S, Lenglet G, Boudot C, Avondo C, Boullier A, Choukroun G, Massy ZA, Kamel S, Chillon JM. Cellular and molecular mechanisms associated with ischemic stroke severity in female mice with chronic kidney disease. Sci Rep. 2019;9:6432. doi: 10.1038/s41598-019-42933-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Henderson SJ, Weitz JI, Kim PY. Fibrinolysis: strategies to enhance the treatment of acute ischemic stroke. J Thromb Haemost. 2018;16:1932–1940. doi: 10.1111/jth.14215. [DOI] [PubMed] [Google Scholar]
- 30.Herder V, Iskandar CD, Kegler K, Hansmann F, Elmarabet SA, Khan MA, Kalkuhl A, Deschl U, Baumgärtner W, Ulrich R, Beineke A. Dynamic changes of microglia/macrophage m1 and m2 polarization in theiler’s murine encephalomyelitis. Brain Pathol. 2015;25:712–723. doi: 10.1111/bpa.12238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hu X, Leak RK, Shi Y, Suenaga J, Gao Y, Zheng P, Chen J. Microglial and macrophage polarization—new prospects for brain repair. Nat Rev Neurol. 2015;11:56–64. doi: 10.1038/nrneurol.2014.207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Hu X, Li P, Guo Y, Wang H, Leak RK, Chen S, Gao Y, Chen J. Microglia/macrophage polarization dynamics reveal novel mechanism of injury expansion after focal cerebral ischemia. Stroke. 2012;43:3063–3070. doi: 10.1161/STROKEAHA.112.659656. [DOI] [PubMed] [Google Scholar]
- 33.Jia Y. Microglia and their roles in response to injury of the central nervous system. Yixue Yanjiusheng Xuebao. 2012;25:418–421. [Google Scholar]
- 34.Jiang M, Liu X, Zhang D, Wang Y, Hu X, Xu F, Jin M, Cao F, Xu L. Celastrol treatment protects against acute ischemic stroke-induced brain injury by promoting an IL-33/ST2 axis-mediated microglia/macrophage M2 polarization. J Neuroinflammation. 2018a;15:78. doi: 10.1186/s12974-018-1124-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Jiang M, Wang H, Jin M, Yang X, Ji H, Jiang Y, Zhang H, Wu F, Wu G, Lai X, Cai L, Hu R, Xu L, Li L. Exosomes from miR-30d-5p-ADSCs reverse acute ischemic stroke-induced, autophagy-mediated brain injury by promoting M2 microglial/macrophage polarization. Cell Physiol Biochem. 2018b;47:864–878. doi: 10.1159/000490078. [DOI] [PubMed] [Google Scholar]
- 36.Kawabori M, Yenari MA. The role of the microglia in acute CNS injury. Metab Brain Dis. 2015;30:381–392. doi: 10.1007/s11011-014-9531-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kim HJ, Rowe M, Ren M, Hong JS, Chen PS, Chuang DM. Histone deacetylase inhibitors exhibit anti-inflammatory and neuroprotective effects in a rat permanent ischemic model of stroke: multiple mechanisms of action. J Pharmacol Exp Ther. 2007;321:892–901. doi: 10.1124/jpet.107.120188. [DOI] [PubMed] [Google Scholar]
- 38.Kolosowska N, Keuters MH, Wojciechowski S, Keksa-Goldsteine V, Laine M, Malm T, Goldsteins G, Koistinaho J, Dhungana H. Peripheral administration of IL-13 induces anti-inflammatory microglial/macrophage responses and provides neuroprotection in ischemic stroke. Neurotherapeutics. 2019;16:1304–1319. doi: 10.1007/s13311-019-00761-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kuo PC, Yu IC, Scofield BA, Brown DA, Curfman ET, Paraiso HC, Chang FL, Yen JH. 3H-1, 2-Dithiole-3-thione as a novel therapeutic agent for the treatment of ischemic stroke through Nrf2 defense pathway. Brain Behav Immun. 2017;62:180–192. doi: 10.1016/j.bbi.2017.01.018. [DOI] [PubMed] [Google Scholar]
- 40.Kuroda E, Ho V, Ruschmann J, Antignano F, Hamilton M, Rauh MJ, Antov A, Flavell RA, Sly LM, Krystal G. SHIP represses the generation of IL-3-induced M2 macrophages by inhibiting IL-4 production from basophils. J Immunol. 2009;183:3652–3660. doi: 10.4049/jimmunol.0900864. [DOI] [PubMed] [Google Scholar]
- 41.Kwon MJ, Kim J, Shin H, Jeong SR, Kang YM, Choi JY, Hwang DH, Kim BG. Contribution of macrophages to enhanced regenerative capacity of dorsal root ganglia sensory neurons by conditioning injury. J Neurosci. 2013;33:15095–15108. doi: 10.1523/JNEUROSCI.0278-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Lan X, Han X, Li Q, Yang QW, Wang J. Modulators of microglial activation and polarization after intracerebral haemorrhage. Nat Rev Neurol. 2017;13:420–433. doi: 10.1038/nrneurol.2017.69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Latta CH, Sudduth TL, Weekman EM, Brothers HM, Abner EL, Popa GJ, Mendenhall MD, Gonzalez-Oregon F, Braun K, Wilcock DM. Determining the role of IL-4 induced neuroinflammation in microglial activity and amyloid-β using BV2 microglial cells and APP/PS1 transgenic mice. J Neuroinflammation. 2015;12:41. doi: 10.1186/s12974-015-0243-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Lee YY, Park JS, Leem YH, Park JE, Kim DY, Choi YH, Park EM, Kang JL, Kim HS. The phosphodiesterase 10 inhibitor papaverine exerts anti-inflammatory and neuroprotective effects via the PKA signaling pathway in neuroinflammation and Parkinson’s disease mouse models. J Neuroinflammation. 2019;16:246. doi: 10.1186/s12974-019-1649-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Li F, Zhao H, Han Z, Wang R, Tao Z, Fan Z, Zhang S, Li G, Chen Z, Luo Y. Xuesaitong may protect against ischemic stroke by modulating microglial phenotypes and inhibiting neuronal cell apoptosis via the STAT3 signaling pathway. CNS Neurol Disord Drug Targets. 2019a;18:115–123. doi: 10.2174/1871527317666181114140340. [DOI] [PubMed] [Google Scholar]
- 46.Li Q, Liu D, Pan F, Ho CSH, Ho RCM. Ethanol exposure induces microglia activation and neuroinflammation through TLR4 activation and SENP6 modulation in the adolescent rat hippocampus. Neural Plast. 2019b;2019:1648736. doi: 10.1155/2019/1648736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Liu R, Liao XY, Pan MX, Tang JC, Chen SF, Zhang Y, Lu PX, Lu LJ, Zou YY, Qin XP, Bu LH, Wan Q. Glycine exhibits neuroprotective effects in ischemic stroke in rats through the inhibition of M1 microglial polarization via the NF-κB p65/Hif-1α signaling pathway. J Immunol. 2019;202:1704–1714. doi: 10.4049/jimmunol.1801166. [DOI] [PubMed] [Google Scholar]
- 48.Liu Z, Ran Y, Huang S, Wen S, Zhang W, Liu X, Ji Z, Geng X, Ji X, Du H, Leak RK, Hu X. Curcumin protects against ischemic stroke by titrating micro-glia/macrophage polarization. Front Aging Neurosci. 2017;9:233. doi: 10.3389/fnagi.2017.00233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Lu D, Shen L, Mai H, Zang J, Liu Y, Tsang CK, Li K, Xu A. HMG-CoA reductase inhibitors attenuate neuronal damage by suppressing oxygen glucose deprivation-induced activated microglial cells. Neural Plast. 2019;2019:7675496. doi: 10.1155/2019/7675496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Lu DY, Tang CH, Chen YH, Wei IH. Berberine suppresses neuroinflammatory responses through AMP-activated protein kinase activation in BV-2 microglia. J Cell Biochem. 2010;110:697–705. doi: 10.1002/jcb.22580. [DOI] [PubMed] [Google Scholar]
- 51.Luo XL, Liu SY, Wang LJ, Zhang QY, Xu P, Pan LL, Hu JF. A tetramethoxychalcone from Chloranthus henryi suppresses lipopolysaccharide-induced inflam-matory responses in BV2 microglia. Eur J Pharmacol. 2016;774:135–143. doi: 10.1016/j.ejphar.2016.02.013. [DOI] [PubMed] [Google Scholar]
- 52.Ma Y, Wang J, Wang Y, Yang GY. The biphasic function of microglia in ischemic stroke. Prog Neurobiol. 2017;157:247–272. doi: 10.1016/j.pneurobio.2016.01.005. [DOI] [PubMed] [Google Scholar]
- 53.Machado LS, Kozak A, Ergul A, Hess DC, Borlongan CV, Fagan SC. Delayed minocycline inhibits ischemia-activated matrix metalloproteinases 2 and 9 after experimental stroke. BMC Neurosci. 2006;7:56. doi: 10.1186/1471-2202-7-56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Masuda T, Croom D, Hida H, Kirov SA. Capillary blood flow around microglial somata determines dynamics of microglial processes in ischemic conditions. Glia. 2011;59:1744–1753. doi: 10.1002/glia.21220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Meng HL, Li XX, Chen YT, Yu LJ, Zhang H, Lao JM, Zhang X, Xu Y. Neuronal soluble fas ligand drives M1-microglia polarization after cerebral ischemia. CNS Neurosci Ther. 2016;22:771–781. doi: 10.1111/cns.12575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Michell-Robinson MA, Touil H, Healy LM, Owen DR, Durafourt BA, Bar-Or A, Antel JP, Moore CS. Roles of microglia in brain development, tissue maintenance and repair. Brain. 2015;138:1138–1159. doi: 10.1093/brain/awv066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Miron VE, Boyd A, Zhao JW, Yuen TJ, Ruckh JM, Shadrach JL, van Wijngaarden P, Wagers AJ, Williams A, Franklin RJM, Ffrench-Constant C. M2 microglia and macrophages drive oligodendrocyte differentiation during CNS remyelination. Nat Neurosci. 2013;16:1211–1218. doi: 10.1038/nn.3469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Mounier R, Théret M, Arnold L, Cuvellier S, Bultot L, Göransson O, Sanz N, Ferry A, Sakamoto K, Foretz M, Viollet B, Chazaud B. AMPKα1 regulates macrophage skewing at the time of resolution of inflammation during skeletal muscle regeneration. Cell Metab. 2013;18:251–264. doi: 10.1016/j.cmet.2013.06.017. [DOI] [PubMed] [Google Scholar]
- 59.Penfield W. Cytology & cellular pathology of the nervous system. New York: PB Hoeber, Inc; 1932. [Google Scholar]
- 60.Perego C, Fumagalli S, De Simoni MG. Temporal pattern of expression and colocalization of microglia/macrophage phenotype markers following brain ischemic injury in mice. J Neuroinflammation. 2011;8:174. doi: 10.1186/1742-2094-8-174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Perry VH, Nicoll JA, Holmes C. Microglia in neurodegenerative disease. Nat Rev Neurol. 2010;6:193–201. doi: 10.1038/nrneurol.2010.17. [DOI] [PubMed] [Google Scholar]
- 62.Prinz M, Priller J. Microglia and brain macrophages in the molecular age: from origin to neuropsychiatric disease. Nat Rev Neurosci. 2014;15:300–312. doi: 10.1038/nrn3722. [DOI] [PubMed] [Google Scholar]
- 63.Qin H, Yeh WI, De Sarno P, Holdbrooks AT, Liu Y, Muldowney MT, Reynolds SL, Yanagisawa LL, Fox TH, 3rd, Park K, Harrington LE, Raman C, Benveniste EN. Signal transducer and activator of transcription-3/suppressor of cytokine signaling-3 (STAT3/SOCS3) axis in myeloid cells regulates neuroin-flammation. Proc Natl Acad Sci U S A. 2012;109:5004–5009. doi: 10.1073/pnas.1117218109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Rajkovic O, Potjewyd G, Pinteaux E. Regenerative medicine therapies for targeting neuroinflammation after stroke. Front Neurol. 2018;9:734. doi: 10.3389/fneur.2018.00734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Ransohoff RM, Perry VH. Microglial physiology: unique stimuli, specialized responses. Annu Rev Immunol. 2009;27:119–145. doi: 10.1146/annurev.immunol.021908.132528. [DOI] [PubMed] [Google Scholar]
- 66.Satoh T, Takeuchi O, Vandenbon A, Yasuda K, Tanaka Y, Kumagai Y, Miyake T, Matsushita K, Okazaki T, Saitoh T, Honma K, Matsuyama T, Yui K, Tsujimura T, Standley DM, Nakanishi K, Nakai K, Akira S. The Jmjd3-Irf4 axis regulates M2 macrophage polarization and host responses against helminth infection. Nat Immunol. 2010;11:936–944. doi: 10.1038/ni.1920. [DOI] [PubMed] [Google Scholar]
- 67.Serhan A, Aerts JL, Boddeke E, Kooijman R. Neuroprotection by insulin-like growth factor-1 in rats with ischemic stroke is associated with microglial changes and a reduction in neuroinflammation. Neuroscience. 2020;426:101–114. doi: 10.1016/j.neuroscience.2019.11.035. [DOI] [PubMed] [Google Scholar]
- 68.Sieweke MH, Allen JE. Beyond stem cells: self-renewal of differentiated macrophages. Science. 2013;342:1242974. doi: 10.1126/science.1242974. [DOI] [PubMed] [Google Scholar]
- 69.Skaper SD, Facci L, Giusti P. Neuroinflammation, microglia and mast cells in the pathophysiology of neurocognitive disorders: a review. CNS Neurol Disord Drug Targets. 2014;13:1654–1666. doi: 10.2174/1871527313666141130224206. [DOI] [PubMed] [Google Scholar]
- 70.Skirving DJ, Dan NG. A 20-year review of percutaneous balloon compression of the trigeminal ganglion. J Neurosurg. 2001;94:913–917. doi: 10.3171/jns.2001.94.6.0913. [DOI] [PubMed] [Google Scholar]
- 71.Stephan J, Friauf E. Functional analysis of the inhibitory neurotransmitter transporters GlyT1, GAT-1, and GAT-3 in astrocytes of the lateral superior olive. Glia. 2014;62:1992–2003. doi: 10.1002/glia.22720. [DOI] [PubMed] [Google Scholar]
- 72.Streit WJ, Mrak RE, Griffin WS. Microglia and neuroinflammation: a pathological perspective. J Neuroinflammation. 2004;1:14. doi: 10.1186/1742-2094-1-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Sudduth TL, Schmitt FA, Nelson PT, Wilcock DM. Neuroinflammatory phenotype in early Alzheimer’s disease. Neurobiol Aging. 2013;34:1051–1059. doi: 10.1016/j.neurobiolaging.2012.09.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Switzer JA, Hess DC, Ergul A, Waller JL, Machado LS, Portik-Dobos V, Pettigrew LC, Clark WM, Fagan SC. Matrix metalloproteinase-9 in an exploratory trial of intravenous minocycline for acute ischemic stroke. Stroke. 2011;42:2633–2635. doi: 10.1161/STROKEAHA.111.618215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Thored P, Heldmann U, Gomes-Leal W, Gisler R, Darsalia V, Taneera J, Nygren JM, Jacobsen SE, Ekdahl CT, Kokaia Z, Lindvall O. Long-term accumulation of microglia with proneurogenic phenotype concomitant with persistent neurogenesis in adult subventricular zone after stroke. Glia. 2009;57:835–849. doi: 10.1002/glia.20810. [DOI] [PubMed] [Google Scholar]
- 76.Tian DS, Li CY, Qin C, Murugan M, Wu LJ, Liu JL. Deficiency in the voltage-gated proton channel Hv1 increases M2 polarization of microglia and attenuates brain damage from photothrombotic ischemic stroke. J Neurochem. 2016;139:96–105. doi: 10.1111/jnc.13751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Veltkamp R, Gill D. Clinical trials of immunomodulation in ischemic stroke. Neurotherapeutics. 2016;13:791–800. doi: 10.1007/s13311-016-0458-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Voet S, Mc Guire C, Hagemeyer N, Martens A, Schroeder A, Wieghofer P, Daems C, Staszewski O, Vande Walle L, Jordao MJC, Sze M, Vikkula HK, Demeestere D, Van Imschoot G, Scott CL, Hoste E, Gonçalves A, Guilliams M, Lippens S, Libert C, et al. A20 critically controls microglia activation and inhibits inflammasome-dependent neuroinflammation. Nat Commun. 2018;9:2036. doi: 10.1038/s41467-018-04376-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Wake H, Moorhouse AJ, Miyamoto A, Nabekura J. Microglia: actively surveying and shaping neuronal circuit structure and function. Trends Neurosci. 2013;36:209–217. doi: 10.1016/j.tins.2012.11.007. [DOI] [PubMed] [Google Scholar]
- 80.Wang J, Yang Z, Liu C, Zhao Y, Chen Y. Activated microglia provide a neuroprotective role by balancing glial cell-line derived neurotrophic factor and tumor necrosis factor-α secretion after subacute cerebral ischemia. Int J Mol Med. 2013;31:172–178. doi: 10.3892/ijmm.2012.1179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Wang J, Zhao H, Fan Z, Li G, Ma Q, Tao Z, Wang R, Feng J, Luo Y. Long noncoding RNA H19 promotes neuroinflammation in ischemic stroke by driving histone deacetylase 1-dependent M1 microglial polarization. Stroke. 2017a;48:2211–2221. doi: 10.1161/STROKEAHA.117.017387. [DOI] [PubMed] [Google Scholar]
- 82.Wang Q, Lv C, Sun Y, Han X, Wang S, Mao Z, Xin Y, Zhang B. The role of alpha-lipoic acid in the pathomechanism of acute ischemic stroke. Cell Physiol Biochem. 2018;48:42–53. doi: 10.1159/000491661. [DOI] [PubMed] [Google Scholar]
- 83.Wang XL, Qiao CM, Liu JO, Li CY. Inhibition of the SOCS1-JAK2-STAT3 signaling pathway confers neuroprotection in rats with ischemic stroke. Cell Physiol Biochem. 2017b;44:85–98. doi: 10.1159/000484585. [DOI] [PubMed] [Google Scholar]
- 84.Xia CY, Zhang S, Gao Y, Wang ZZ, Chen NH. Selective modulation of microglia polarization to M2 phenotype for stroke treatment. Int Immunopharmacol. 2015;25:377–382. doi: 10.1016/j.intimp.2015.02.019. [DOI] [PubMed] [Google Scholar]
- 85.Xiong XY, Liu L, Yang QW. Functions and mechanisms of microglia/macrophages in neuroinflammation and neurogenesis after stroke. Prog Neurobiol. 2016;142:23–44. doi: 10.1016/j.pneurobio.2016.05.001. [DOI] [PubMed] [Google Scholar]
- 86.Xu P, Zhang X, Liu Q, Xie Y, Shi X, Chen J, Li Y, Guo H, Sun R, Hong Y, Liu X, Xu G. Microglial TREM-1 receptor mediates neuroinflammatory injury via interaction with SYK in experimental ischemic stroke. Cell Death Dis. 2019;10:555. doi: 10.1038/s41419-019-1777-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Xu Y, Qian L, Zong G, Ma K, Zhu X, Zhang H, Li N, Yang Q, Bai H, Ben J, Li X, Xu Y, Chen Q. Class A scavenger receptor promotes cerebral ischemic injury by pivoting microglia/macrophage polarization. Neuroscience. 2012;218:35–48. doi: 10.1016/j.neuroscience.2012.05.036. [DOI] [PubMed] [Google Scholar]
- 88.Yan A, Zhang T, Yang X, Shao J, Fu N, Shen F, Fu Y, Xia W. Thromboxane A2 receptor antagonist SQ29548 reduces ischemic stroke-induced micro-glia/macrophages activation and enrichment, and ameliorates brain injury. Sci Rep. 2016;6:35885. doi: 10.1038/srep35885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Yu Z, Sun D, Feng J, Tan W, Fang X, Zhao M, Zhao X, Pu Y, Huang A, Xiang Z, Cao L, He C. MSX3 switches microglia polarization and protects from inflammation-induced demyelination. J Neurosci. 2015;35:6350–6365. doi: 10.1523/JNEUROSCI.2468-14.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Zhang X, Zhu XL, Ji BY, Cao X, Yu LJ, Zhang Y, Bao XY, Xu Y, Jin JL. LncRNA-1810034E14Rik reduces microglia activation in experimental ischemic stroke. J Neuroinflammation. 2019;16:75. doi: 10.1186/s12974-019-1464-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Zhou X, Cao Y, Ao G, Hu L, Liu H, Wu J, Wang X, Jin M, Zheng S, Zhen X, Alkayed NJ, Jia J, Cheng J. CaMKKβ-dependent activation of AMP-activated protein kinase is critical to suppressive effects of hydrogen sulfide on neuroinflammation. Antioxid Redox Signal. 2014;21:1741–1758. doi: 10.1089/ars.2013.5587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Zhu J, Cao D, Guo C, Liu M, Tao Y, Zhou J, Wang F, Zhao Y, Wei J, Zhang Y, Fang W, Li Y. Berberine facilitates angiogenesis against ischemic stroke through modulating microglial polarization via AMPK signaling. Cell Mol Neurobiol. 2019;39:751–768. doi: 10.1007/s10571-019-00675-7. [DOI] [PMC free article] [PubMed] [Google Scholar]