

Abstract
Prostate cancer (PCa) is one of the most common malignancies in Western countries. Studies have shown that androgen contributes to the progression of PCa, but how androgen promotes PCa remains largely unknown. Here, we demonstrated that androgen suppressed the expression of miR-760 depending on the interaction between androgen and androgen receptor (AR). miR-760 was downregulated in prostate cancer tissues compared with normal tissues. Functional experiments showed that miR-760 downregulation promoted the proliferation and growth of LNCaP and 22rv1 cells. In contrast, miR-760 ectopic expression inhibited the proliferation of LNCaP and 22rv1 cells. DNA synthesis was suppressed by miR-760. Mechanistically, miR-760 bound to the 3'UTR of interleukin 6 (IL6). A mutation in the binding site disrupted their interaction. In addition, silencing of IL6 suppressed the proliferation of LNCaP and 22rv1 cells. IL6 was upregulated in PCa tissues. Our study reveals that androgen downregulates miR-760 to promote the growth of PCa cells by regulating IL6.
Keywords: androgen, interleukin 6, miR-760, prostate cancer
INTRODUCTION
Prostate cancer (PCa) is one of the most common malignancies and the second leading cause of cancer-related death among men in developed countries.1,2 Decades of efforts have demonstrated that androgen plays an essential role in PCa development. PCa patients respond favorably to androgen deprivation therapy (ADT), but most fail to be cured and develop castration-resistant prostate cancer (CRPC).3,4 Recently, a gene expression profiling study determined that androgen receptor (AR) contributed to the development of CRPC.5 Nevertheless, knowledge of the mechanism is very limited. Understanding the critical molecular events may help develop effective treatment strategies for CRPC.
MicroRNAs are a group of approximately 24 nucleotide long noncoding RNA molecules that posttranscriptionally regulate the expression of different genes by binding to mRNAs.6,7 The connection between androgen-AR and microRNAs has been reported in PCa. Jia et al.8 found that AR stimulated miR-193a-3p to promote the migration of PCa cells. They also identified various upregulated or downregulated microRNAs in CRPC cells. miR-182-5p is another positive downstream effector of AR. Upregulation of miR-182-5p promotes cell proliferation, invasion, and migration by negatively regulating arrestin domain-containing protein 3 (ARRDC3).9 Recent studies have demonstrated that downregulation of miR-760 plays a pivotal role in the development and progression of different cancers, including glioma, lung, and colon cancer.10,11,12 However, the relationship between miR-760 and androgen-associated PCa remains unknown.
In this study, we aimed to investigate the significance of miR-760 in PCa. AR was knocked down or overexpressed in PCa cells, and the expression of miR-760 was evaluated after androgen stimulation. The results showed that androgen downregulated miR-760 depending on the expression of AR. miR-760 was downregulated in PCa tissues. In vitro, miR-760 suppressed the proliferation and growth of PCa cells. Furthermore, miR-760 was found to directly target IL6. Our study demonstrates that miR-760 functions as a tumor suppressive microRNA in PCa.
MATERIALS AND METHODS
Prostate cancer tissues
All prostate cancer and normal tissues were collected between March 2017 and September 2019 at Peking University Cancer Hospital (Beijing, China). The research was carried out according to the World Medical Association Declaration of Helsinki and was approved by the Clinical Research Ethics Committee of the hospital. Written informed consent was obtained from each patient. Adjacent normal tissues 2 cm in proximity to the cancer lesion were collected. All tissues were obtained before therapeutic intervention. The expression of miR-760 was detected using quantitative real-time polymerase chain reaction (qRT-PCR) assay.
TCGA database analysis of miR-760
The transcript of miR-760 in PCa patients was analyzed from The Cancer Genome Atlas (TCGA) website (http://cancergenome.nih.gov). The correlation between miR-760 and PCa survival was also analyzed.
Cell culture
The PCa cells LNCaP, 22rv1, DU145, and PC3 were obtained from the American Type Culture Collection (Manassas, VA, USA). All the cells were cultured in RPMI 1640 (Invitrogen, Carlsbad, CA, USA), which contained 10% fetal bovine serum (Gibco, Grand Island, NY, USA) and 1% penicillin and streptomycin solution (Corning Inc., Corning, NY, USA). The cell culture was maintained in a 37°C incubator with 5% CO2.
Oligonucleotide transfection
miR-760 inhibitor and control inhibitor, miR-760 mimics and control mimics, antagomir miR-760 and control antagomir, and agomir miR-760 and control agomir were synthesized by RiboBio (Guangzhou, China). Oligonucleotides were transfected using Lipofectamine 3000 reagent (Invitrogen) following the manufacturer's instructions.
AR overexpression assay
The coding sequence (CDS) of AR was synthesized and cloned into pcDNA3.1. pcDNA3.1 empty or pcDNA3.1-AR vector was transfected into DU145 or PC3 cells using Lipofectamine 3000. Forty-eight hours later, DU145 and PC3 cells overexpressing AR were treated with or without R1881.
RNA interference
siRNAs against AR and IL6 were purchased from GenePharma company (Shanghai, China). siCtrl, siAR, or siIL6 oligonucleotides were used to transfect the indicated cells with RNAiMAX reagent (Invitrogen), following the manufacturer's instructions. The target sequences of AR and IL6 were CCGAGGAGCTTTCCAGAAT and GCAGGACATGACAACTCAT, respectively.
RNA extraction and qRT-PCR
Total RNA was extracted from the indicated cells using TRIzol reagent (Invitrogen). For microRNA qRT-PCR, a High-Capacity RNA-to-cDNA kit was used for reverse transcription. A TaqMan probe (Roche, Basel, Switzerland) was used to detect the product of the quantitative polymerase chain reaction (qPCR). The level of miR-760 was measured with a master mix (Thermo Fisher Scientific, Waltham, MA, USA). U6 served as the internal control.
For mRNA qRT-PCR, equal amounts of total RNA were subjected to reverse transcription using Moloney Murine Leukemia Virus reverse transcriptase (M-MLV) reverse transcriptase (Promega, Madison, WI, USA). qRT-PCR was performed using SYBR master mix (Takara, Otsu, Japan) on a Bio-Rad IQ 5 machine (Hercules, CA, USA). The sequences of the primers were as follows: IL6 forward, 5'-ACTCACCTCTTCAGAACGAATTG-3' and reverse, 5'-CCATCTTTGGAAGGTTCAGGTTG-3'; glyceraldehyde-3-phosphate dehydrogenase (GAPDH) forward, 5'-TGACTTCAACAGCGACACCCA-3' and reverse, 5'-CACCCTGTTGCTGTAGCCAAA-3'. GAPDH served as the internal control.
Western blot assays
Proteins were extracted from LNCaP and 22rv1 cells using radioimmunoprecipitation assay (RIPA) buffer (Beyotime, Shanghai, China). The protein concentration was determined by a BCA kit (Pierce, Rockford, IL, USA). Fifty micrograms of the total protein was separated on sodium dodecyl sulfate-polyacrylamide gel electrophoresis gels and transferred to polyvinylidene fluoride membranes. The membranes were blocked with 5% nonfat milk at room temperature for 1 h, followed by incubation with primary antibodies at 4°C overnight. After washing with phosphate buffered saline with Tween (PBST), the membranes were incubated with secondary antibodies. Then, the protein abundance was detected by an ECL-Plus kit (Amersham Biosciences, Boston, MA, USA). Antibodies against IL6 were purchased from Abcam (Ab9324, Cambridge, United Kingdom). Anti-β-actin primary antibody was obtained from Santa Cruz Biotechnology (sc-47778, Santa Cruz, CA, USA). All secondary antibodies were from ABclonal (AS003 and AS014, Wuhan, China).
Cell proliferation analysis
The proliferation of PC cells was measured by Cell Counting Kit assay (YEASEN, Shanghai, China). In brief, LNCaP and 22rv1 cells were transfected with inhibitors (control or miR-760) or mimics (control or miR-760). A total of 2000 LNCaP or 22rv1 cells were seeded in 96-well plates. One day, 2, 3, and 4 days later, 20 μl of CCK buffer was added to each well, respectively, and the plates were maintained at 37°C for 2 h. The optical density (OD) value at 450 nm was measured on a microplate reader.
Colony formation assay
Equal numbers of LNCaP or 22rv1 cells, which were transfected with antagomir control or miR-760 (durable inhibitors) or transfected with agomir control or miR-760 (durable mimics), were seeded and cultured at 37°C. Seven to 10 days later, the plates were washed with PBS twice and fixed with methanol for half an hour. Subsequently, the colonies were stained with crystal violet solution. The images were acquired with a camera.
EdU staining
A BeyoClick™ EdU Cell Proliferation Kit with Alexa Fluor 555 (Beyotime) was used for EdU staining. LNCaP or 22rv1 cells (3 × 105) were seeded on coverslips in 6-well plates. Twenty-four hours later, the cells were stained with EdU following the manufacturer's protocols. Nuclei were stained with 4',6-diamdino-2-phenylindole (DAPI). Images were obtained under a fluorescence microscope (Olympus, Tokyo, Japan).
Luciferase reporter assay
The wild type (WT) or mutated (MU) 3' UTR sequence of IL6 was synthesized and cloned into a psi-CHECK vector (Promega). Lipofectamine 2000 (Invitrogen) was used to transfect the vectors into LNCaP cells according to the manufacturer's instructions. The luciferase activity was determined by the Dual-Luciferase Reporter Assay System (Promega). Renilla luciferase activity served as the internal control.
Statistical analyses
GraphPad Prism (GraphPad Software, San Diego, CA, USA) was used to analyze the data. Paired or unpaired Student's t-test was used to analyze the difference between two groups. Differences were considered statistically significant when P < 0.05.
RESULTS
Androgen inhibits the expression of miR-760 in LNCaP and 22rv1 cells
Androgen plays an important role in PCa growth and survival.13 To explore the role of miR-760 in AR-related PCa, the AR-positive PCa cells LNCaP and 22rv1 and AR-negative PCa cells DU145 and PC3 were stimulated with the androgen analogue R1881 and subjected to qRT-PCR analysis of miR-760. The results showed that R1881 suppressed the expression of miR-760 in LNCaP (P < 0.05), and 22rv1 (P < 0.01) cells but had no effect on miR-760 expression in DU145 and PC3 cells (Figure 1a and 1b). To confirm that R1881 regulates miR-760 depending on AR expression, we silenced AR in LNCaP and 22rv1 cells and overexpressed AR in DU145 and PC3 cells. The miR-760 transcript was analyzed after R1881 stimulation. In AR-overexpressing DU145 and PC3 cells, the expression of miR-760 was suppressed by R1881 (P < 0.05). In contrast, it remained unchanged after R1881 treatment of AR-silenced LNCaP (P > 0.05) and 22rv1 cells (P > 0.05; Figure 1c and 1d). These results suggest that androgen negatively regulates miR-760 expression depending on AR.
Figure 1.
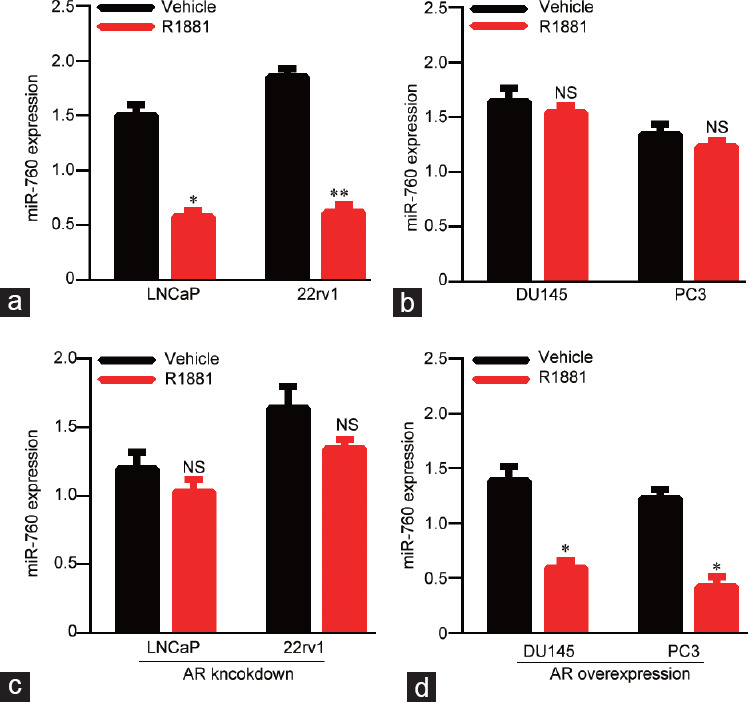
Androgen inhibits miR-760 expression depending on the androgen receptor. (a) qPCR analysis of miR-760 in LNCaP and 22rv1 cells that were treated with vehicle or R1881. (b) qPCR analysis of miR-760 in DU145 and PC3 cells that were treated with vehicle or R1881. (c) qPCR analysis of miR-760 in AR knockdown LNCaP and 22rv1 cells that were treated with vehicle or R1881. (d) qPCR analysis of miR-760 in AR-overexpressing DU145 and PC3 cells that were treated with vehicle or R1881. *P < 0.05, **P < 0.01. NS: no significance; qPCR: quantitative polymerase chain reaction; AR: androgen receptor.
The expression of miR-760 is reduced in PCa tissues
To investigate the clinical relevance of miR-760 in PCa, we collected 10 pairs of PCa tissues and their adjacent normal tissues. qRT-PCR was used to detect the levels of miR-760. The results showed that miR-760 expression was decreased in PCa tissues compared with adjacent tissues (P < 0.01; Figure 2a). Then, the miR-760 level was also measured in nonpaired PCa tissues. We found that miR-760 was lower in PCa tissues than in nonpaired normal tissues (P < 0.001; Figure 2b). To confirm our results, we analyzed the TCGA database and found that miR-760 transcript was significantly decreased in PCa tissues compared with normal tissues (P < 0.001; Figure 2c). The database analysis also showed that low miR-760 expression predicted relatively shorter survival in PCa patients (Figure 2d). Overall, miR-760 is downregulated in PCa tissues.
Figure 2.
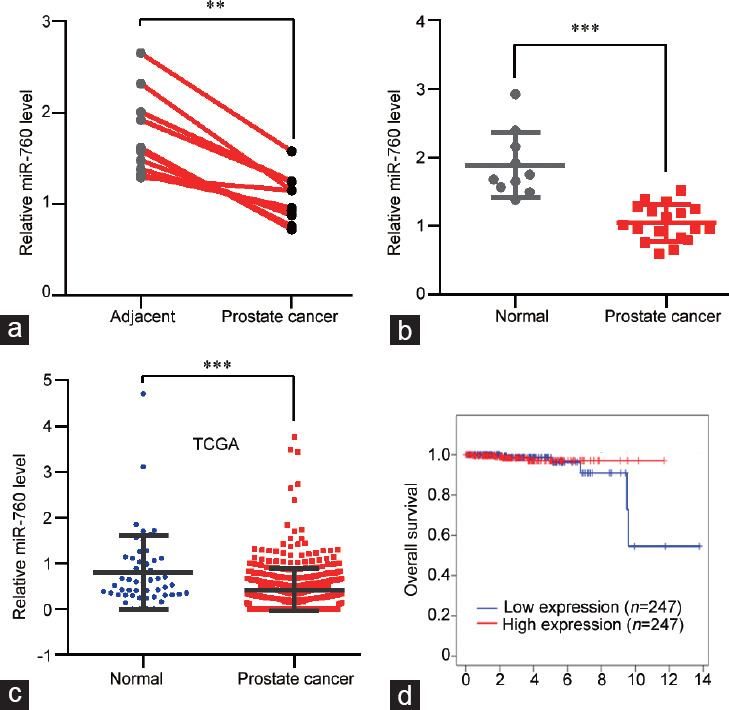
miR-760 levels are reduced in prostate cancer tissues. (a) qPCR analysis of miR-760 levels in prostate cancer and adjacent normal tissues (n = 10). (b) qPCR analysis of miR-760 levels in prostate cancer (n = 18) and nonpaired normal (n = 10) tissues. (c) miR-760 expression was analyzed from The Cancer Genome Atlas (TCGA) database data in prostate cancer (n = 499) and nonpaired normal (n = 52) tissues. (d) The overall survival of prostate cancer patients was analyzed from TCGA database in the miR-760 low (n = 247) and high (n = 247) expression groups (P > 0.05). **P < 0.01, ***P < 0.001. qPCR: quantitative polymerase chain reaction.
miR-760 inhibits the proliferation and growth of PCa cells
We next investigated the role of miR-760 in PCa cells by silencing and overexpressing miR-760 with miR-760 inhibitors and mimics. CCK results indicated that miR-760 inhibitors promoted the proliferation of LNCaP (P < 0.05) and 22rv1 (P < 0.01) cells, while the opposite results were observed in miR-760-overexpressing cells (P < 0.05; Figure 3a and 3b). Based on stable miR-760 knockdown and overexpression, we found that miR-760 had a similar effect on PCa cell colony formation (Figure 3c). DNA synthesis is a hallmark of cell proliferation. We then subjected PCa cells to EdU staining after miR-760 silencing or overexpression. The results showed that miR-760 knockdown enhanced EdU incorporation in LNCaP and 22rv1 cells. In contrast, the opposite results were found in miR-760-overexpressing PCa cells (Figure 3d). Overall, miR-760 is a tumor suppressor in PCa.
Figure 3.
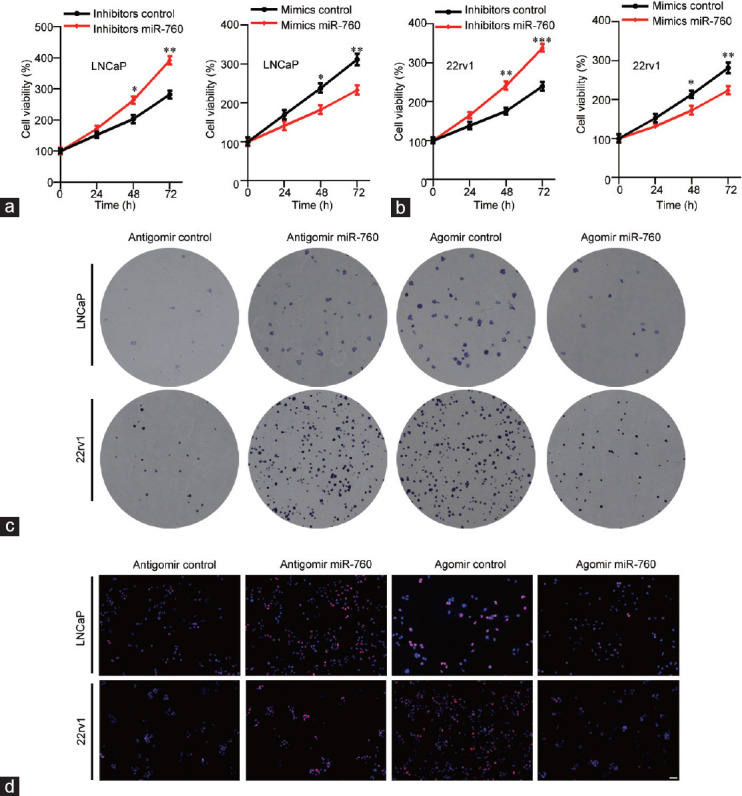
miR-760 suppresses the growth of prostate cancer cells. (a) LNCaP and (b) 22rv1cells transfected with control inhibitor or miR-760 inhibitor (left) or transfected with control mimics or miR-760 mimics (right) were subjected to Cell Counting Kit analysis of cell proliferation. LNCaP and 22rv1 cells transfected with control antagomir or miR-760 antagomir (left) or transfected with control agomir or miR-760 agomir (right) were subjected (c) to colony formation assays and (d) to EdU staining. Scale bars = 50 μm. *P < 0.05, **P < 0.01, ***P < 0.001.
miR-760 inhibits the expression of IL6 by binding to its 3'UTR
To study the molecular mechanisms of the tumor suppressive role of miR-760 in PCa, the online program TargetScan was used to predict the potential targets of miR-760. The diagram showed that miR-760 directly bound to the 3'UTR of IL6, which is a potential modulator of PCa (Figure 4a). We therefore used a luciferase reporter assay to verify the connection between miR-760 and IL6 by constructing reporters containing the WT or MU 3'UTR of IL6. The results showed that miR-760 reduced the luciferase activity of the WT but not the MU IL6 3'UTR reporter, suggesting that miR-760 directly binds to the 3'UTR of IL6 (P < 0.05; Figure 4b). qRT-PCR results indicated that miR-760 inhibited the mRNA expression of IL6 (P < 0.01; Figure 4c). Then, the role of IL6 was investigated in 22rv1 and LNCaP cells using siRNA. IL6 knockdown significantly suppressed the proliferation of 22rv1(P < 0.05) and LNCaP cells (P < 0.01; Figure 4d and 4e). Moreover, the mRNA level of IL6 was higher in PCa tissues than in paired and nonpaired normal tissues (P < 0.01 and P < 0.001, respectively; Figure 4f and 4g). Taken together, the results demonstrate that miR-760 suppresses IL6 to inhibit the viability of PCa cells.
Figure 4.
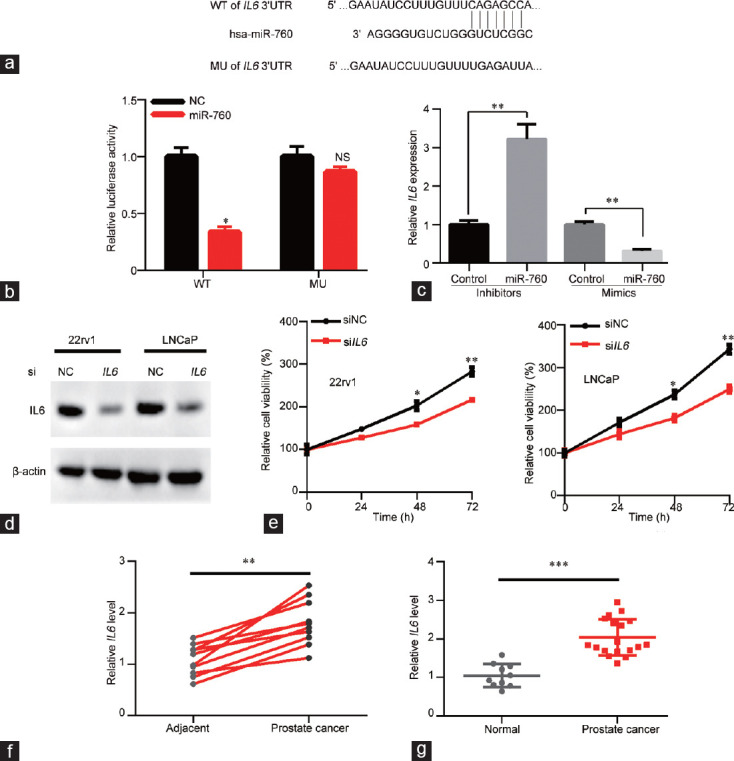
miR-760 directly targets IL6 by binding to its 3’UTR. (a) The putative binding site of miR-760 in the 3’UTR of IL6. The mutated site is also shown in the diagram. (b) Control or miR-760 mimics were cotransfected with psiCHECK reporters containing the wild-type or mutated 3’UTR of IL6. Luciferase activity was measured as indicated. (c) qPCR analysis of IL6 in LNCaP cells that were transfected with control inhibitor or miR-760 inhibitor or transfected with conrol mimics or miR-760 mimics. siNC or siIL6 LNCaP and 22rv1 cells were subjected to (d) western blot analysis of IL6 and (e) CCK analysis of cell proliferation. (f) qPCR analysis of IL6 levels in prostate cancer and adjacent normal tissues (n = 10). (g) qPCR analysis of IL6 levels in prostate cancer (n = 18) and nonpaired normal (n = 10) tissues. *P < 0.05, **P < 0.01, ***P < 0.001. NS: no significance. IL6: interleukin 6; CCK: Cell Counting Kit; NC: negative control; qPCR: quantitative polymerase chain reaction; WT: wide type; MU: mutant.
DISCUSSION
Androgen is a major risk factor for PCa through the regulation of AR signaling. The downstream effectors of AR remain to be clarified. In this study, we determined that miR-760 is a downstream target of AR and plays an important role in PCa. Androgen repressed the expression of miR-760 in AR-positive but not AR-negative PCa cells. TCGA database analysis and our experimental results showed that miR-760 was downregulated in PCa tissues. Functional studies demonstrated the tumor suppressive role of miR-760 in PCa. Mechanistically, miR-760 directly bound to the 3'UTR of IL6 to suppress its expression. We conclude that the androgen-AR complex contributes to PCa development by suppressing miR-760.
Dysregulation of miRNAs has been proven to contribute to the proliferation, growth, metastasis, and invasion of cancers by negatively regulating important downstream mRNAs. Increasing evidence demonstrates that miR-760 is dysregulated in a set of cancers. For example, miR-760 is upregulated by estrogen stimulation in breast cancer.14 Hu et al.15 found that miR-760 levels were reduced in doxorubicin (DOX)-resistant MCF-7 cells. Han et al.16 showed that miR-760 reduced breast cancer stemness by silencing Nanog homeobox (NANOG). Overexpression of miR-760 alleviates the resistance of breast cancer cells to anticancer drugs.15 Moreover, miR-760 was downregulated in colon cancer tissues, and it suppressed the growth of colon cancer by targeting basic leucine zipper transcription factor ATF-like 3 (BATF3)/ activator protein 1 (AP-1)/cyclinD1 signaling.17–19 However, another study demonstrated that miR-760 was an oncogenic microRNA in ovarian cancer through the downregulation of PH domain and leucine-rich repeat protein phosphatase 2 (PHLPP2).20 miR-760 also acts as an oncogene in hepatocellular carcinoma. The downregulation of miR-760 predicted a better survival of the patients.21 These studies indicate that miR-760 has different roles in cancer development depending on the cancer type. Nevertheless, the significance of miR-760 in PCa is unclear. In this study, we initially observed that miR-760 was downregulated in LNCaP and 22rv1 cells but not in PC3 and DU145 cells after R1881 stimulation. Because LNCaP and 22rv1 cells are AR-positive cells, while PC3 and DU145 cells are AR-negative cells, we knocked down AR in LNCaP and 22rv1 and overexpressed AR in PC3 and DU145 cells. The results were opposite to the present results, indicating that androgen suppressed the expression of miR-760 depending on AR signaling. Furthermore, we found that miR-760 was downregulated in PCa tissues compared with paired or nonpaired normal tissues according to our results and the TCGA database. The TCGA database analysis also showed that patients with low miR-760 expression exhibited poorer survival than those with high miR-760 expression. Importantly, functional experiments demonstrated that miR-760 suppressed the DNA synthesis, proliferation, and growth of PCa cells. These results reveal that androgen-AR-mediated repression of miR-760 enhances the viability of PCa cells.
IL6 is an important inflammatory factor. Enhanced IL6 production has been proven to promote the development of CRPC in a mouse model.22 Furthermore, IL6 signaling activation contributes to prostate cancer growth triggered by high-fat diet-stimulated inflammation.23 The melanoma differentiation associated gene-9 (mda-9)/insulin-like growth factor-1 receptor (IGF-1R)/signal transducer and activator of transcription 3 (STAT3) cascade induces prostate cancer invasion.24 In addition, the expression of IL6 is suppressed by let-7 microRNA in prostate cancer progression.25 IL6 also participates in AR-mediated gene expression in PCa,26 but the direct effect of IL6 on PCa cell growth is poorly understood. Here, we showed that the mRNA and protein expression of IL6 was repressed by miR-760. Luciferase reporter activity results showed that miR-760 potentially bound to the CAGAGCC sequence in the 3'UTR of IL6. Importantly, the miR-760-mediated suppression of IL6 was confirmed to have an anticancer role in PCa because knockdown of IL6 significantly suppressed the viability of PCa cells. Furthermore, IL6 was downregulated in PCa tissues compared with normal tissues. Therefore, miR-760 suppresses the proliferation and growth of PCa cells at least partly by targeting IL6.
CONCLUSION
We demonstrated for the first time that miR-760 functions as a tumor suppressive microRNA in PCa and is suppressed by androgen. Androgen represses the expression of miR-760 depending on AR. miR-760 inhibits the viability of PCa cells. The expression of IL6 was silenced by miR-760. Therefore, androgen-AR promotes PCa growth through upregulation of IL6 by negatively regulating miR-760.
AUTHOR CONTRIBUTIONS
SW and PD designed the study, wrote the manuscript draft and revised the manuscript. SW, YY, YDC, XXT, and PD performed the experiments and analyzed the data. All authors read and approved the final manuscript.
COMPETING INTERESTS
All authors declared no competing interests.
ACKNOWLEDGMENTS
The research was financially supported by the Capital Special Scientific Projects for Health Development (2018-2-1025).
REFERENCES
- 1.Siegel RL, Miller KD, Jemal A. Cancer Statistics, 2017. CA Cancer J Clin. 2017;67:7–30. doi: 10.3322/caac.21387. [DOI] [PubMed] [Google Scholar]
- 2.Litwin MS, Tan HJ. The Diagnosis and treatment of prostate cancer: a review. JAMA. 2017;317:2532–42. doi: 10.1001/jama.2017.7248. [DOI] [PubMed] [Google Scholar]
- 3.Harris WP, Mostaghel EA, Nelson PS, Montgomery B. Androgen deprivation therapy: progress in understanding mechanisms of resistance and optimizing androgen depletion. Nat Clin Pract Urol. 2009;6:76–85. doi: 10.1038/ncpuro1296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Chandrasekar T, Yang JC, Gao AC, Evans CP. Targeting molecular resistance in castration-resistant prostate cancer. BMC Med. 2015;13:206. doi: 10.1186/s12916-015-0457-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Watson PA, Arora VK, Sawyers CL. Emerging mechanisms of resistance to androgen receptor inhibitors in prostate cancer. Nat Rev Cancer. 2015;15:701–11. doi: 10.1038/nrc4016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Krol J, Loedige I, Filipowicz W. The widespread regulation of microRNA biogenesis, function and decay. Nat Rev Genet. 2010;11:597–610. doi: 10.1038/nrg2843. [DOI] [PubMed] [Google Scholar]
- 7.Voinnet O. Origin, biogenesis, and activity of plant microRNAs. Cell. 2009;136:669–87. doi: 10.1016/j.cell.2009.01.046. [DOI] [PubMed] [Google Scholar]
- 8.Jia L, Gui B, Zheng D, Decker KF, Tinay I, et al. Androgen receptor-regulated miRNA-193a-3p targets AJUBA to promote prostate cancer cell migration. Prostate. 2017;77:1000–11. doi: 10.1002/pros.23356. [DOI] [PubMed] [Google Scholar]
- 9.Yao J, Xu C, Fang Z, Li Y, Liu H, Wang Y, et al. Androgen receptor regulated microRNA miR-182-5p promotes prostate cancer progression by targeting the ARRDC3/ITGB4 pathway. Biochem Biophys Res Commun. 2016;474:213–9. doi: 10.1016/j.bbrc.2016.04.107. [DOI] [PubMed] [Google Scholar]
- 10.Yao J, Xu C, Fang Z, Li Y, Liu H, et al. Long non-coding RNA LOC730100 enhances proliferation and invasion of glioma cells through competitively sponging miR-760 from FOXA1 mRNA. Biochem Biophys Res Commun. 2019;512:558–63. doi: 10.1016/j.bbrc.2019.03.124. [DOI] [PubMed] [Google Scholar]
- 11.Zhang X, Wang L, Liu Y, Huang W, Cheng D. MiR-760 enhances TRAIL sensitivity in non-small cell lung cancer via targeting the protein FOXA1. Biomed Pharmacother. 2018;99:523–9. doi: 10.1016/j.biopha.2018.01.076. [DOI] [PubMed] [Google Scholar]
- 12.Zhu Y, Xing Y, Chi F, Sun W, Zhang Z, et al. Long noncoding RNA SNHG6 promotes the progression of colorectal cancer through sponging miR-760 and activation of FOXC1. Onco Targets Ther. 2018;11:5743–52. doi: 10.2147/OTT.S170246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Knudsen KE, Scher HI. Starving the addiction: new opportunities for durable suppression of AR signaling in prostate cancer. Clin Cancer Res. 2009;15:4792–8. doi: 10.1158/1078-0432.CCR-08-2660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Cicatiello L, Mutarelli M, Grober OM, Paris O, Ferraro L, et al. Estrogen receptor alpha controls a gene network in luminal-like breast cancer cells comprising multiple transcription factors and microRNAs. Am J Pathol. 2010;176:2113–30. doi: 10.2353/ajpath.2010.090837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hu SH, Wang CH, Huang ZJ, Liu F, Xu CW, et al. miR-760 mediates chemoresistance through inhibition of epithelial mesenchymal transition in breast cancer cells. Eur Rev Med Pharmacol Sci. 2016;20:5002–8. [PubMed] [Google Scholar]
- 16.Han ML, Wang F, Gu YT, Pei XH, Ge X, et al. MicroR-760 suppresses cancer stem cell subpopulation and breast cancer cell proliferation and metastasis: by down-regulating NANOG. Biomed Pharmacother. 2016;80:304–10. doi: 10.1016/j.biopha.2016.03.024. [DOI] [PubMed] [Google Scholar]
- 17.Li H, Zhang H, Lu G, Li Q, Gu J, et al. Mechanism analysis of colorectal cancer according to the microRNA expression profile. Oncol Lett. 2016;12:2329–36. doi: 10.3892/ol.2016.5027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Wang Q, Huang Z, Ni S, Xiao X, Xu Q, et al. Plasma miR-601 and miR-760 are novel biomarkers for the early detection of colorectal cancer. PLoS One. 2012;7:e44398. doi: 10.1371/journal.pone.0044398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Cao L, Liu Y, Wang D, Huang L, Li F, et al. MiR-760 suppresses human colorectal cancer growth by targeting BATF3/AP-1/cyclinD1 signaling. J Exp Clin Cancer Res. 2018;37:83. doi: 10.1186/s13046-018-0757-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Liao Y, Deng Y, Liu J, Ye Z, You Z, et al. MiR-760 overexpression promotes proliferation in ovarian cancer by downregulation of PHLPP2 expression. Gynecol Oncol. 2016;143:655–63. doi: 10.1016/j.ygyno.2016.09.010. [DOI] [PubMed] [Google Scholar]
- 21.Sun D, Lu J, Hu C, Zhang Q, Wang X, et al. Prognostic role of miR-760 in hepatocellular carcinoma. Oncol Lett. 2018;16:7239–44. doi: 10.3892/ol.2018.9546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Patel R, Fleming J, Mui E, Loveridge C, Repiscak P, et al. Sprouty2 loss-induced IL6 drives castration-resistant prostate cancer through scavenger receptor B1. EMBO Mol Med. 2018;10:e8347. doi: 10.15252/emmm.201708347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hayashi T, Fujita K, Nojima S, Hayashi Y, Nakano K, et al. High-fat diet-induced inflammation accelerates prostate cancer growth via IL6 signaling. Clin Cancer Res. 2018;24:4309–18. doi: 10.1158/1078-0432.CCR-18-0106. [DOI] [PubMed] [Google Scholar]
- 24.Das SK, Pradhan AK, Bhoopathi P, Talukdar S, Shen XN, et al. The MDA-9/Syntenin/IGF1R/STAT3 axis directs prostate cancer invasion. Cancer Res. 2018;78:2852–63. doi: 10.1158/0008-5472.CAN-17-2992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sung SY, Liao CH, Wu HP, Hsiao WC, Wu IH, et al. Loss of let-7 microRNA upregulates IL-6 in bone marrow-derived mesenchymal stem cells triggering a reactive stromal response to prostate cancer. PLoS One. 2013;8:e71637. doi: 10.1371/journal.pone.0071637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Chen T, Wang LH, Farrar WL. Interleukin 6 activates androgen receptor-mediated gene expression through a signal transducer and activator of transcription 3-dependent pathway in LNCaP prostate cancer cells. Cancer Res. 2000;60:2132–5. [PubMed] [Google Scholar]