

Abstract
Introduction
Recent reports suggest SARS-CoV-2, the virus causing COVID-19, may be transmittable from pregnant mother to placenta and fetus, albeit rarely. The efficacy of vertical transmission of SARS-CoV-2 critically depends on the availability of its receptor, ACE2, in the placenta. In the present study, we tested the hypothesis that placental ACE2 expression is oxygenation-dependent by studying the expression of ACE2 and associated cell entry regulators in the monochorionic twin anemia-polycythemia (TAPS) placenta, a model of discordant placental oxygenation.
Methods
We performed a retrospective comparative immunohistochemical, immunofluorescence and Western blot analysis of ACE2, TMPRSS2 and Cathepsin B expression in anemic and polycythemic territories of TAPS placentas (N = 14).
Results
ACE2 protein levels were significantly higher in the anemic twin territories than in the corresponding polycythemic territories, associated with upregulation of the key ACE2-related cell entry regulators, TMPRSS2 and Cathepsin B, immunolocalized to villous trophoblastic and stromal cells. Cellular colocalization of ACE2 and TMPRSS2, suggestive of functionality of this cell entry axis, was demonstrated by double immunofluorescence studies.
Discussion
Placental hypoxia is associated with upregulation of ACE2 expression, concomitant with increased expression of its key cell entry proteases. ACE2-regulated placental functions, both infection- and non-infection related, may be highly oxygenation-dependent.
Keywords: Hypoxia, Monochorionic, Coronavirus
1. Introduction
The rapid global spread of the 2019 novel coronavirus, SARS-CoV-2, and the resulting disease, COVID-19 [[1], [2], [3]], from its origins in Wuhan China has created an unrelenting public health threat, affecting more than 30 million persons worldwide and resulting in more than one million deaths as of September, 2020 [World Health Organization situation report] [Johns Hopkins COVID-19 Case Tracker, https://coronavirus.jhu.edu/]. Similar to other coronaviruses linked to epidemics, such as severe acute respiratory syndrome-related coronavirus (SARS-CoV) [4] and Middle East respiratory syndrome coronavirus (MERS-CoV) [5], SARS-CoV-2 primarily targets the respiratory tract, while other routes of transmission (indirect, fecal-oral, ocular, sexual) are less common and/or still under consideration [6].
At present data on vertical transmission of SARS-CoV-2 from mother to fetus during pregnancy are limited. Early studies including large case series of COVID-19-positive pregnant women from Wuhan, China, suggested that transmission of SARS-CoV-2 in utero or intrapartum is unlikely, as amniotic fluid, cord blood and breast milk samples from the mothers were negative for SARS-CoV-2 [7]. Other studies similarly failed to find evidence of vertical transmission in neonates or placentas [[8], [9], [10], [11], [12], [13]].
Conversely, the detection of anti-SARS-CoV-2 IgM antibodies in some neonates soon (<24 h) after birth to COVID-19-positive mothers [[14], [15], [16]] raised the strong possibility of intrauterine transmission of SARS-COV-2, since IgM antibodies cannot cross the placenta and typically appear only 3–7 days after infection due to their complex molecular structure [15,16]. However, the reliability of SARS-CoV-2 IgM assays as indicator of intrauterine transmission has been questioned [14] especially in the absence of additional reflex testing [17]. IgM assays in general are prone to technical errors, leading to false negative and false positive results [14,18]. Further, transfer of maternal IgM across the placental barrier may occur in inflammatory conditions, such as maternal hypoxia, viral cytotoxic effects, or ascending infections of the lower genital tract [17,19]. Finally, the observed sharp postnatal decline of SARS-CoV-2 IgM levels differs from the general behavior of these antibodies in other congenital infections, such as rubella or Zikavirus infection [14].Whereas the significance of SARS-CoV-2 IgM antibodies in neonates thus remains unclear, a growing number of suspected or confirmed intrauterine or intrapartum-acquired neonatal SARS-CoV-2 infections have been reported, suggesting intrauterine vertical transmission of SARS-CoV-2 from pregnant mother to placenta and/or fetus may be possible, albeit exceptional [[20], [21], [22], [23], [24], [25], [26], [27], [28]]. In these reports, the assumption of vertical transmission was generally based on SARS-CoV-2 nucleic acid amplification of nasopharyngeal swab samples obtained from neonates of COVID-19-positive mothers proximate to delivery [[20], [21], [22], [23], [24],26,27,29,30], or on documented evidence of placental or amniotic fluid SARS-CoV-2 infection [21,[24], [25], [26],29,[31], [32], [33]]. Other studies have described the presence of SARS-CoV-2 virions in villous syncytiotrophoblastic cells by electron microscopy [24,25,32,34], providing complimentary histopathologic evidence of intrauterine transmission of SARS-CoV-2. It deserves mention that immunohistochemical studies of SARS-Cov-2 infected placentas thus far have failed to show definitive localization of the virus beyond the trophoblast into the fetal compartment. Furthermore, some of the ultrastructural studies [32,34] appear to demonstrate confusion between virions and coated vesicles.
As in other organ systems, the capacity for transmission of SARS-CoV-2 across the maternal-fetal interface barrier critically depends on the permissibility of the placenta as barrier, and in particular: on the availability and functionality of its viral entry mechanisms in the syncytiotrophoblast layer. SARS-CoV-2 gains cellular entry by attachment of its spike protein (S) to the angiotensin-converting enzyme 2 (ACE2) receptor [[35], [36], [37]], followed by activation of the virus internalization process by host cell proteases, in particular the cell surface protease, TMPRSS2 (transmembrane protease, serine 2), and lysosomal cathepsin proteases [[37], [38], [39], [40], [41]]. Whereas the regulation of ACE2 expression in other organs has been well documented, the regulatory mechanisms underlying placental ACE2 expression are less well known, which is surprising in view of its critical role as member of the placental renin-angiotensin system (RAS).
In animal models, placental ACE2 expression has been determined to be modulated by maternal nutrient restriction [42], maternal glucocorticoid administration [43], and placental insufficiency [44]. Previous studies have also demonstrated, with seemingly conflicting results, that the placental RAS system and ACE2 expression are dysregulated by maternal hypoxia. In a mouse model of maternal hypoxia, Cuffe et al. [45] described decreased placental Ace2 mRNA and protein expression. In a similar murine model, Goyal et al. [46] reported decreased Ace2 mRNA levels, but increased protein levels, suggestive of post-transcriptional regulatory mechanisms involved with hypoxic stress. In a rat model of placental insufficiency, which is presumably characterized, at least in part, by placental hypoxia, Goyal et al. [44] demonstrated increased, rather than decreased, placental Ace2 mRNA expression.
In the current study, we utilized the twin anemia-polycythemia (TAPS) human placenta as model of intertwin differential placental oxygenation to study the effects of hypoxia on placental expression of ACE2 and its cell entry regulators, TMPRSS2 and cathepsin B, in vivo. Twin anemia-polycythemia sequence is a unique complication of monochorionic twinning, characterized by chronic and slow intertwin blood transfusion that results in large intertwin hemoglobin differences without associated (severe) twin oligohydramnios-polyhydramnios sequence [47]. We previously demonstrated that TAPS placentas display significant intertwin discordant tissue oxygenation, characterized by significant tissue hypoxia in the anemic territories compared with their polycythemic counterparts [48]. Importantly, selection of the monochorionic TAPS twin placenta as model system for this study allowed elimination of most maternal, pregnancy-related and genetic confounding factors, as twins are exposed to the same maternal and uterine milieu, and as monochorionic (ie monozygotic) twins (and their placentas) have a near-identical genetic makeup. The aim of this study was to test the hypothesis that TAPS placentas, characterized by striking intertwin hemoglobin and oxygenation discordance, show corresponding intertwin differences in placental expression of ACE2 and associated cellular entry regulators.
2. Materials and methods
2.1. Patient population
We performed a clinicoplacental analysis of a consecutive series of diamniotic-monochorionic twin placentas from pregnancies complicated by TAPS, examined at the Department of Pathology at Women and Infants Hospital of Rhode Island between 2010 and 2019. The diagnosis of twin anemia-polycythemia sequence was based on the following proposed criteria for postnatal diagnosis of TAPS: intertwin Hb difference > 8 g/dL and very small intertwin anastomoses [49,50]. Placentas with fetal demise and placentas from pregnancies complicated by twin-to-twin transfusion syndrome (TTTS, twin oligohydramnios-polyhydramnios sequence) were excluded from this study. The accompanying charts were reviewed for relevant maternal and fetal/neonatal information. Relevant neonatal information included birth weight, Apgar scores, gender, and first hematologic blood count values (hemoglobin and hematocrit, reticulocyte count, obtained immediately after delivery). Findings in the anemic twin territory were compared with those of the adjacent polycythemic twin territory. The study was approved by the Institutional Review Board.
2.2. Processing of the placenta
Immediately upon receipt in the Department of Pathology, placental parenchyma from four randomly selected quadrants of each twin territory was sampled for subsequent molecular (Western blot) analyses. To avoid inclusion of shared cotyledons, these samples were obtained from villous parenchyma lateral to the respective cord insertions, at a distance from the vascular equator. Areas of placental parenchyma with calcification, infarction, fibrin deposition or hemorrhage were avoided. Placental samples were placed in RNAlater (Amnion Inc., Austin, TX) and stored at −20 °C. Further examination of the placenta, including injection and categorization of the chorionic vasculature, was performed as previously described in detail [[51], [52], [53]]. For histological, immunohistochemical and immunofluorescence studies, tissues were obtained from at least 5 randomly selected areas per twin territory. These tissues were formalin-fixed and paraffin-embedded according to standard methods.
2.3. Immunohistochemical and immunofluorescence analysis
Immunohistochemical staining was performed using a Dako Autostainer (Dako, Carpenteria, CA), as previously described in detail [54]. For this peroxidase-based staining, we used rabbit monoclonal anti-ACE2 antibody (concentration: 1:100) and anti-TMPRSS2 antibodies (concentration: 1:1000) (both: Abcam, Cambridge, MA). Placental sections from corresponding twin territories were prepared and immunostained in one single session. Controls for specificity consisted of incubation with isotype IgG instead of the respective primary antibodies, which abolished all immunoreactivity.
Anatomic colocalization of ACE2 and its canonical protease, TMPRSS2, was assessed by combining anti-ACE2 with anti-TMPRSS2 immunofluorescence staining. For these double immunofluorescence studies, tissue sections were incubated sequentially with monoclonal rabbit anti-TMPRSS2 antibody, Alexa Fluor 594-conjugated anti-rabbit IgG (Jackson ImmunoResearch Laboratories, Inc., West Grove, PA), mouse monoclonal anti-ACE2 antibody (ThermoFisher Scientific (Invitrogen), Waltham, MA), biotinylated anti-mouse IgG (Vector Laboratories, Inc., Burlingame, CA) and Alexa Fluor 488-conjugated streptavidin (Jackson ImmunoResearch Laboratories, Inc.). Sections were covered with aqueous mounting medium containing 4′,6-diamidine-2′-phenylindole dihydrochloride (DAPI, Vector Laboratories, Inc.). Controls consisted of omission of one or both primary antibodies, which abolished the respective immunoreactivities. Confocal images were acquired with a Nikon Ti-E spinning disk confocal microscope (Nikon Inc., Melville, NY) using diode lasers 405, 488, and 561, and processed as previously described [48].
2.4. Western blot analysis
ACE2 and SARS-CoV-2 cellular entry-related protein levels were evaluated by Western blot analysis of placental lysates, according to methods described in detail elsewhere [55], using the following antibodies: anti-ACE2 rabbit monoclonal antibody (abcam), anti-TMPRSS2 rabbit monoclonal antibody (abcam), and anti-Cathepsin B (D1C7Y) rabbit monoclonal antibody (Cell Signaling Technology, Danvers, MA). A housekeeping gene, GAPDH (glyceraldehyde 3-phosphate dehydrogenase, Santa Cruz Biotechnology, Dallas, Texas) served as internal loading control. Band intensity was expressed as the integrated optical density (IOD) normalized to the IOD of GAPDH.
2.5. Data analysis
Values are expressed as mean ± standard deviation (SD) or standard error of mean (SEM), or median (range). The significance of differences between groups was determined by Student t-test, Mann-Whitney U test, ANOVA with post-hoc Scheffe test, Fisher's exact test, or Wilcoxon matched-pairs signed rank test, where applicable. Data were analyzed and graphically represented using GraphPad Prism 5 software (GraphPad Prism; GraphPad Software, Inc., San Diego, CA). Data depicted in modified (Tukey) box plots reflect group median, upper and lower quartiles (box), maximum and minimum values excluding outliers (whiskers), and outliers (more than 3/2 times upper quartile). The significance level was set at P < 0.05.
3. Results
3.1. General clinical and placental data
ACE2 and ACE2-related expression was studied in a consecutive cohort of 14 placentas from pregnancies complicated by spontaneous TAPS (i.e. not iatrogenic following laser treatment for twin-to-twin transfusion syndrome). Relevant clinical and placental data are summarized in Table 1 . The intertwin Hb difference was >8 g/dL in all cases, fulfilling the proposed hematologic criteria for TAPS, and ranged from 8.9 g/dL to 20.3 g/dL (median: 12.8 g/dL). Reticulocyte counts obtained immediately after birth (prior to transfusion) were only available in three cases (reticulocyte count ratios: 4.0, 4.5 and 5.2).
Table 1.
Patient and placental data of TAPS pregnancies.
| Polycythemic twin/placental side (14) | Anemic twin/placental side (14) | P | |
|---|---|---|---|
| Gestational age (wks) | 34 (26–37) | ||
| Birth weight (g) | 1715 ± 635 | 1456 ± 618 | <0.05 |
| Median Apgar 1 min | 7 (3–8) | 7 (1–9) | NS |
| Apgar <7 at 1 min | 4/14 | 6/14 | NS |
| Median Apgar 5 min | 8.5 (6–9) | 8 (6–9) | NS |
| Hgb (g/dL) | 23.4 ± 1.5 | 9.1 ± 2.7 | <0.01 |
| Hct (%) | 65.4 ± 4.9 | 26.9 ± 5.9 | <0.01 |
| Placenta weight (g) | 683 ± 129 | ||
| Placental share (%) | 47 (39–61) | 53 (45–61) | NS |
| Peripheral cord (M/V) | 6/14 | 11/14 | NS |
All placental weights were appropriate for gestational age. Anemic twins tended to have peripheral (marginal or velamentous) cord insertion more frequently than polycythemic twins (11/14 versus 6/14) but this trend was not significant. In agreement with the proposed placental criteria for TAPS diagnosis, all TAPS placentas displayed few, relatively small intertwin choriovascular anastomoses (Fig. 1 A). In 11/14 cases, the maternal surface showed marked color (redness) discordance with clear demarcation of anemic and polycythemic territories (Fig. 1B). Histologic examination of the TAPS placentas revealed varying degrees of congestion of the villous capillaries in the polycythemic twin share (Fig. 1C), and small, collapsed capillaries with increased numbers of circulating nucleated erythroid precursors (erythroblastosis) in the anemic twin share (Fig. 1D).
Fig. 1.

Representative gross and microscopic appearance of TAPS placenta.
A. Fetal surface following removal of the intertwin membrane and injection of the chorionic vasculature. Vascular injection highlights the near-complete separation of the choriovascular beds. A single minuscule artery-to-artery anastomosis is shown by arrow. Color code: Left twin: artery: red, vein: green; right twin: artery: red, vein: yellow (venous bed only partially filled due to vascular disruption).
B. Maternal surface demonstrating marked color (redness) discordance with sharp demarcation of polycythemic (left) and anemic (right) territories. C-D. Representative micrographs of plethoric and pale placental parenchyma, respectively. Insert: ×600 magnification of Fig. 1D showing circulating erythroid precursors (hematoxylin-eosin staining, original magnification ×200. Scale bar = 20 μm)
3.2. ACE2 and ACE2-associated protease expression in TAPS placentas
Placental protein levels of ACE2 were assayed by Western blot analysis of whole placental lysates. As shown in Fig. 2 A, immunoreactive ACE2 protein levels, appropriately sized at 120 kDa [56], were consistently higher in anemic placental shares than in the corresponding polycythemic placental shares. As functionality of the ACE2 receptor depends on its colocalization and coexpression with TMPRSS2 and other ACE2-associated host cellular proteases [37,57], we assessed the expression of these critical cell entry regulators. As shown in Fig. 2A, appropriately sized immunoreactive bands for TMPRSS2 and Cathepsin B were readily detected in all polycythemic and anemic placental shares. In all twin sets studied, levels of these proteins were significantly higher in the anemic twin territories than in the corresponding polycythemic territories. Results of densitometric quantitation of band intensities, normalized to GAPDH, are shown in Fig. 2B.
Fig. 2.

ACE2 and ACE2-associated expression in TAPS placentas.
A Western blot analysis of ACE2, TMPRSS2 and CTSB expression in lysates of polycythemic (P) and anemic (A) territories of representative TAPS placentas. GAPDH served as loading control. B. Densitometric analysis of Western blot. IOD: integrated optical density; *: P < 0.05; **: P < 0.01; ***: P < 0.001 (Wilcoxon matched-pairs signed rank test). ACE2: angiotensin converting enzyme 2; TMPRSS2: transmembrane serine protease 2; CTSB: cathepsin B.
Concordant with the immunoblot data, immunohistochemical analysis demonstrated significantly stronger ACE2 and TMPRSS2 immunoreactivity in anemic than in polycythemic twin shares (Fig. 3 ). Both proteins were immunolocalized to villous syncytiotrophoblastic, cytotrophoblastic, endothelial and stromal cells (Fig. 3). Colocalization of ACE2 and its canonical associated cellular protease, TMPRSS2, was studied by double immunofluorescence studies in the anemic twin territories. Foci of apparent colocalization (yellow fluorescence) of ACE2 (green fluorescence) and TMPRSS2 (red fluorescence) were readily observed (Fig. 4 ).
Fig. 3.
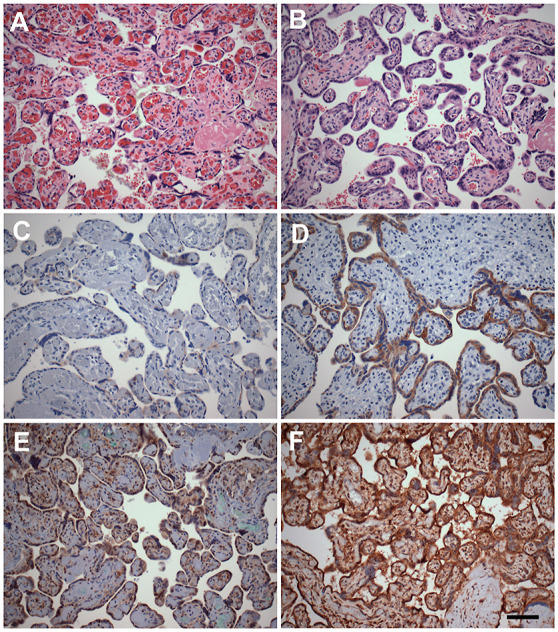
Representative immunohistochemical analysis of ACE2 and TMPRSS2 protein expression in TAPS placenta.
Representative immunohistochemical analysis of expression of ACE2 (C-D), and TMPRSS2 (E-F) in polycythemic (left) and corresponding anemic (right) territories of TAPS placentas. (A-B: hematoxylin-eosin stain; C–F: DAB-peroxidase system with hematoxylin counterstain, original magnification ×200. Scale bar = 20 μm).
Fig. 4.

Combined immunofluorescence analysis of placental colocalization of ACE and TMPRSS2.
Confocal fluorescence microscopy of the anemic territory of a TAPS placenta subjected to combined anti-ACE2 (green) and anti-TMPRSS2 (red) immunofluorescence, captured at the same settings. Yellow dots represent regions of colocalization of ACE2 and TMPRSS2, suggestive of membrane fusion. In selected fields, colocalization was confirmed quantitatively using Pearson's correlation coefficient analysis whereby a cutoff of r2 > 0.5 was used to indicate positive colocalization.
4. Discussion
With mounting infections, fatalities and economic losses caused by COVID-19, there is an urgent need to better understand the basic mechanisms underlying SARS-CoV-2 infection, and, in particular, its cell entry mechanisms. In line with the reported effects of hypoxia on ACE2 expression in a wide range of tissues and organ systems [[58], [59], [60], [61], [62]], we determined that ACE2 expression was significantly upregulated in the hypoxic anemic territories of TAPS placentas. As in previous studies [63,64], ACE2 was immunolocalized to villous trophoblastic and stromal cells.
Several recent studies have described robust ACE2 expression in placentas [38,65,66]. In these studies, placental ACE2 expression was associated with low or absent TMPRSS2 expression [17,38,66,67]. This absence or paucity of ACE2/TMPRSS2 co-expression was implicated in the relative insensitivity to transplacental SARS-CoV-2 infection [17,66], as co-expression of these proteins is crucial for cytoplasmic entry of SARS-CoV-2. In sharp contrast to these prior studies, we determined that ACE2 expression in hypoxic placental territories was increased in tandem with increased expression of TMPRSS2 and Cathepsin B, suggesting the placental ACE2-protease axis may have potential functionality, at least in hypoxic conditions.
The clinical implications of the observed relatively high placental expression of ACE2 and associated cellular proteases in hypoxic conditions remain unclear. Intuitively, upregulation of ACE2 expression might be interpreted as predisposing to increased cellular infectivity. By analogy, it has been suggested that the use of drugs such as ACE inhibitors, Ang II-receptor blockers (ARBs) or ibuprofen, may increase the risk of COVID-19 infection by upregulating the cellular expression of ACE2 [[68], [69], [70]]. However, there is at present no convincing clinical evidence linking ACE inhibitors and/or ARBs to COVID-19 severity and mortality [71,72]. Similarly, it remains to be determined whether placental hypoxia-related ACE2 upregulation predisposes to increased cellular infectivity of trophoblastic cells and increased vertical viral transmission. Increased understanding of the biology of ACE2 may contribute to further development of intervention strategies targeting viral cell entry mechanisms. Currently proposed approaches focused on ACE2 as a putative target for host immune surveillance and therapeutic strategies include competitive inhibition using recombinant ACE2 [73], an excess of soluble ACE2 [74] or decoy ACE2-expressing extracellular vesicles [75].
In addition to its function as cognate receptor for SARS-CoV-2 (and SARS-CoV), ACE2 has multiple other enzymatic and non-enzymatic functions. As ACE homologue and member of the RAS system, ACE2 has a wide range of substrates, including kinins, apelin, neurotensin, dynorphin, ghrelin, amyloid and angiotensins [71], and an equally wide range of functions. ACE2 is further involved in placentation, including trophoblast migration, vascular remodeling and vasodilation of the maternal vasculature [64,76]. Dysregulated ACE2 expression has been implicated in pregnancy complications, such as miscarriage, ectopic pregnancy and preeclampsia [63]. In the present study, the birth weight of the anemic TAPS twins was significantly lower than that of the corresponding polycythemic twins, as is characteristic of TAPS pregnancies in general [47]. Whether enhanced ACE2 expression may have contributed to the growth restriction of anemic TAPS twins is unclear.
In summary, this study demonstrates that TAPS placentas display intertwin discordance in expression of ACE2, the SARS-CoV-2 receptor, as well as its associated cellular proteases, TMPRSS2 and cathepsin-B. In view of the known effects of hypoxia on ACE2 expression in trophoblastic cells in vitro and in animal models, and our previous demonstration of decreased tissue oxygen levels in the anemic twin territory, we speculate that the discordance in ACE2/TMPSSR2 expression in TAPS placentas may be attributable, in large part, to differential tissue oxygenation. The concordant upregulation of ACE2 and TMPRSS2/Cathepsin B expression in the anemic twin shares suggests hypoxic conditions may enhance functionality of this axis, potentially predisposing to viral cell entry and vertical transmission. TAPS twin placentas provide an in vivo model for study of the role and regulation of ACE2 expression as adaptive stress response to intrauterine hypoxia, and ultimately may contribute to deeper insight into the role of ACE2 expression and its modulation in reproductive health, both in general and in the context of coronavirus infections.
Declaration of competing interest
The authors declare no conflict of interest.
References
- 1.Zhu N., Zhang D., Wang W., Li X., Yang B., Song J., Zhao X., Huang B., Shi W., Lu R., Niu P., Zhan F., Ma X., Wang D., Xu W., Wu G., Gao G.F., Tan W., China Novel Coronavirus I., Research T. A novel coronavirus from patients with pneumonia in China, 2019. N. Engl. J. Med. 2020;382(8):727–733. doi: 10.1056/NEJMoa2001017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Chen N., Zhou M., Dong X., Qu J., Gong F., Han Y., Qiu Y., Wang J., Liu Y., Wei Y., Xia J., Yu T., Zhang X., Zhang L. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: a descriptive study. Lancet. 2020;395(10223):507–513. doi: 10.1016/S0140-6736(20)30211-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Huang C., Wang Y., Li X., Ren L., Zhao J., Hu Y., Zhang L., Fan G., Xu J., Gu X., Cheng Z., Yu T., Xia J., Wei Y., Wu W., Xie X., Yin W., Li H., Liu M., Xiao Y., Gao H., Guo L., Xie J., Wang G., Jiang R., Gao Z., Jin Q., Wang J., Cao B. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet. 2020;395(10223):497–506. doi: 10.1016/S0140-6736(20)30183-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ksiazek T.G., Erdman D., Goldsmith C.S., Zaki S.R., Peret T., Emery S., Tong S., Urbani C., Comer J.A., Lim W., Rollin P.E., Dowell S.F., Ling A.E., Humphrey C.D., Shieh W.J., Guarner J., Paddock C.D., Rota P., Fields B., DeRisi J., Yang J.Y., Cox N., Hughes J.M., LeDuc J.W., Bellini W.J., Anderson L.J., Group S.W. A novel coronavirus associated with severe acute respiratory syndrome. N. Engl. J. Med. 2003;348(20):1953–1966. doi: 10.1056/NEJMoa030781. [DOI] [PubMed] [Google Scholar]
- 5.Zaki A.M., van Boheemen S., Bestebroer T.M., Osterhaus A.D., Fouchier R.A. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N. Engl. J. Med. 2012;367(19):1814–1820. doi: 10.1056/NEJMoa1211721. [DOI] [PubMed] [Google Scholar]
- 6.Patel K.P., Vunnam S.R., Patel P.A., Krill K.L., Korbitz P.M., Gallagher J.P., Suh J.E., Vunnam R.R. Transmission of SARS-CoV-2: an update of current literature. Eur. J. Clin. Microbiol. Infect. Dis. 2020;39(11):2005–2011. doi: 10.1007/s10096-020-03961-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Schwartz D.A. An analysis of 38 pregnant women with COVID-19, their newborn Infants, and maternal-fetal transmission of SARS-CoV-2: maternal coronavirus infections and pregnancy outcomes. Arch. Pathol. Lab Med. 2020;144(7):799–805. doi: 10.5858/arpa.2020-0901-SA. [DOI] [PubMed] [Google Scholar]
- 8.Karimi-Zarchi M., Neamatzadeh H., Dastgheib S.A., Abbasi H., Mirjalili S.R., Behforouz A., Ferdosian F., Bahrami R. Vertical transmission of coronavirus disease 19 (COVID-19) from infected pregnant mothers to neonates: a review. Fetal Pediatr. Pathol. 2020;39(3):246–250. doi: 10.1080/15513815.2020.1747120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Chen H., Guo J., Wang C., Luo F., Yu X., Zhang W., Li J., Zhao D., Xu D., Gong Q., Liao J., Yang H., Hou W., Zhang Y. Clinical characteristics and intrauterine vertical transmission potential of COVID-19 infection in nine pregnant women: a retrospective review of medical records. Lancet. 2020;395(10226):809–815. doi: 10.1016/S0140-6736(20)30360-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Yan J., Guo J., Fan C., Juan J., Yu X., Li J., Feng L., Li C., Chen H., Qiao Y., Lei D., Wang C., Xiong G., Xiao F., He W., Pang Q., Hu X., Wang S., Chen D., Zhang Y., Poon L.C., Yang H. Coronavirus disease 2019 in pregnant women: a report based on 116 cases. Am. J. Obstet. Gynecol. 2020;223(1):111 e1–111 e14. doi: 10.1016/j.ajog.2020.04.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Masmejan S., Pomar L., Favre G., Panchaud A., Giannoni E., Greub G., Baud D. 11th. Vol. 26. Clin. Microbiol. Infect.; 2020. Vertical transmission and materno-fetal outcomes in 13 patients with COVID-19; pp. 1585–1587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Grimminck K., Santegoets L.A.M., Siemens F.C., Fraaij P.L.A., Reiss I.K.M., Schoenmakers S. No evidence of vertical transmission of SARS-CoV-2 after induction of labour in an immune-suppressed SARS-CoV-2-positive patient. BMJ Case Rep. 2020;13(6):e235581. doi: 10.1136/bcr-2020-235581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Shanes E.D., Mithal L.B., Otero S., Azad H.A., Miller E.S., Goldstein J.A. Placental Pathology in COVID-19. Am. J. Clin. Pathol. 2020;154(1):23–32. doi: 10.1093/ajcp/aqaa089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kimberlin D.W., Stagno S. Can SARS-CoV-2 infection Be acquired in utero?: more definitive evidence is needed. J. Am. Med. Assoc. 2020;323(18):1788–1789. doi: 10.1001/jama.2020.4868. [DOI] [PubMed] [Google Scholar]
- 15.Dong L., Tian J., He S., Zhu C., Wang J., Liu C., Yang J. Possible vertical transmission of SARS-CoV-2 from an infected mother to her newborn. J. Am. Med. Assoc. 2020;323(18):1846–1848. doi: 10.1001/jama.2020.4621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zeng H., Xu C., Fan J., Tang Y., Deng Q., Zhang W., Long X. Antibodies in Infants born to mothers with COVID-19 pneumonia. J. Am. Med. Assoc. 2020;323(18):1848–1849. doi: 10.1001/jama.2020.4861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mahyuddin A.P., Kanneganti A., Wong J.J.J.L., Dimri P.S., Su L.L., Biswas A., Illanes S.E., Mattar C., Huang R.J., Choolani M. Mechanisms and Evidence of Vertical Transmission of Infections in Pregnancy Including SARS-CoV-2. Prenat Diagn. 2020;40(13):1655–1670. doi: 10.1002/pd.5765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Nielsen C.M., Hansen K., Andersen H.M., Gerstoft J., Vestergaard B.F. An enzyme labelled nuclear antigen immunoassay for detection of cytomegalovirus IgM antibodies in human serum: specific and non-specific reactions. J. Med. Virol. 1987;22(1):67–76. doi: 10.1002/jmv.1890220109. [DOI] [PubMed] [Google Scholar]
- 19.Ben-Hur H., Gurevich P., Elhayany A., Avinoach I., Schneider D.F., Zusman I. Transport of maternal immunoglobulins through the human placental barrier in normal pregnancy and during inflammation. Int. J. Mol. Med. 2005;16(3):401–407. [PubMed] [Google Scholar]
- 20.Zeng L., Xia S., Yuan W., Yan K., Xiao F., Shao J., Zhou W. Neonatal early-onset infection with SARS-CoV-2 in 33 neonates born to mothers with COVID-19 in wuhan, China. JAMA Pediatr. 2020;174(7):722–725. doi: 10.1001/jamapediatrics.2020.0878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zamaniyan M., Ebadi A., Aghajanpoor S., Rahmani Z., Haghshenas M., Azizi S. Preterm delivery, maternal death, and vertical transmission in a pregnant woman with COVID-19 infection. Prenat. Diagn. 2020;40(13):1759–1761. doi: 10.1002/pd.5713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Alzamora M.C., Paredes T., Caceres D., Webb C.M., Valdez L.M., La Rosa M. Severe COVID-19 during pregnancy and possible vertical transmission. Am. J. Perinatol. 2020;37(8):861–865. doi: 10.1055/s-0040-1710050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kirtsman M., Diambomba Y., Poutanen S.M., Malinowski A.K., Vlachodimitropoulou E., Parks W.T., Erdman L., Morris S.K., Shah P.S. Probable congenital SARS-CoV-2 infection in a neonate born to a woman with active SARS-CoV-2 infection. CMAJ (Can. Med. Assoc. J.) 2020;192(24):E647–E650. doi: 10.1503/cmaj.200821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Patane L., Morotti D., Giunta M.R., Sigismondi C., Piccoli M.G., Frigerio L., Mangili G., Arosio M., Cornolti G. Vertical transmission of COVID-19: SARS-CoV-2 RNA on the fetal side of the placenta in pregnancies with COVID-19 positive mothers and neonates at birth. Am. J. Obstet. Gynecol. MFM. 2020;2(3):100145. doi: 10.1016/j.ajogmf.2020.100145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sisman J., Jaleel M.A., Moreno W., Rajaram V., Collins R.R.J., Savani R.C., Rakheja D., Evans A.S. Intrauterine transmission of SARS-CoV-2 infection in a preterm infant. Pediatr. Infect. Dis. J. 2020;39(9):e265–e267. doi: 10.1097/INF.0000000000002815. [DOI] [PubMed] [Google Scholar]
- 26.Pulinx B., Kieffer D., Michiels I., Petermans S., Strybol D., Delvaux S., Baldewijns M., Raymaekers M., Cartuyvels R., Maurissen W. Vertical transmission of SARS-CoV-2 infection and preterm birth. Eur. J. Clin. Microbiol. Infect. Dis. 2020;39(12):2441–2445. doi: 10.1007/s10096-020-03964-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Demirjian A., Singh C., Tebruegge M., Herbert R., Draz N., Mirfenderesky M., Jones V., Hinstridge P., Seneviratne R., Myers R., Ellis J., Miah S., Meadows C.I., Kenny J., Alonso A., Handforth J., Perkins S., Butler M., Blackburn F., Douthwaite S., Pillay D., Chand M., Zambon M. Probable vertical transmission of SARS-CoV-2 infection. Pediatr. Infect. Dis. J. 2020;39(9):e257–e260. doi: 10.1097/INF.0000000000002821. [DOI] [PubMed] [Google Scholar]
- 28.Sharps M.C., Hayes D.J.L., Lee S., Zou Z., Brady C.A., Almoghrabi Y., Kerby A., Tamber K.K., Jones C.J., Adams Waldorf K.M., Heazell A.E.P. A structured review of placental morphology and histopathological lesions associated with SARS-CoV-2 infection. Placenta. 2020;101:13–29. doi: 10.1016/j.placenta.2020.08.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Vivanti A.J., Vauloup-Fellous C., Prevot S., Zupan V., Suffee C., Do Cao J., Benachi A., De Luca D. Transplacental transmission of SARS-CoV-2 infection. Nat. Commun. 2020;11(1):3572. doi: 10.1038/s41467-020-17436-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wang S., Guo L., Chen L., Liu W., Cao Y., Zhang J., Feng L. A Case Report of Neonatal 2019 Coronavirus Disease in China. Clin. Infect. Dis. 2020;71(15):853–857. doi: 10.1093/cid/ciaa225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Penfield C.A., Brubaker S.G., Limaye M.A., Lighter J., Ratner A.J., Thomas K.M., Meyer J., Roman A.S. Detection of SARS-COV-2 in placental and fetal membrane samples. Am. J. Obstet. Gynecol. MFM. 2020;2(3):100133. doi: 10.1016/j.ajogmf.2020.100133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Hosier H., Farhadian S.F., Morotti R.A., Deshmukh U., Lu-Culligan A., Campbell K.H., Yasumoto Y., Vogels C.B., Casanovas-Massana A., Vijayakumar P., Geng B., Odio C.D., Fournier J., Brito A.F., Fauver J.R., Liu F., Alpert T., Tal R., Szigeti-Buck K., Perincheri S., Larsen C.P., Gariepy A.M., Aguilar G., Fardelmann K.L., Harigopal M., Taylor H.S., Pettker C.M., Wyllie A.L., Dela Cruz C.S., Ring A.M., Grubaugh N.D., Ko A.I., Horvath T.L., Iwasaki A., Reddy U.M., Lipkind H.S. SARS-CoV-2 infection of the placenta. J. Clin. Invest. 2020;130(9):4947–4953. doi: 10.1172/JCI139569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ferraiolo A., Barra F., Kratochwila C., Paudice M., Vellone V.G., Godano E., Varesano S., Noberasco G., Ferrero S., Arioni C. Report of positive placental swabs for SARS-CoV-2 in an asymptomatic pregnant woman with COVID-19. Medicina (Kaunas) 2020;56(6):306. doi: 10.3390/medicina56060306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Algarroba G.N., Rekawek P., Vahanian S.A., Khullar P., Palaia T., Peltier M.R., Chavez M.R., Vintzileos A.M. Visualization of SARS-CoV-2 virus invading the human placenta using electron microscopy. Am. J. Obstet. Gynecol. 2020;223(2):275–278. doi: 10.1016/j.ajog.2020.05.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Li W., Moore M.J., Vasilieva N., Sui J., Wong S.K., Berne M.A., Somasundaran M., Sullivan J.L., Luzuriaga K., Greenough T.C., Choe H., Farzan M. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature. 2003;426(6965):450–454. doi: 10.1038/nature02145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Liu Z., Xiao X., Wei X., Li J., Yang J., Tan H., Zhu J., Zhang Q., Wu J., Liu L. Composition and divergence of coronavirus spike proteins and host ACE2 receptors predict potential intermediate hosts of SARS-CoV-2. J. Med. Virol. 2020;92(6):595–601. doi: 10.1002/jmv.25726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hoffmann M., Kleine-Weber H., Schroeder S., Kruger N., Herrler T., Erichsen S., Schiergens T.S., Herrler G., Wu N.H., Nitsche A., Muller M.A., Drosten C., Pohlmann S. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell. 2020;181(2):271–280 e8. doi: 10.1016/j.cell.2020.02.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sungnak W., Huang N., Becavin C., Berg M., Queen R., Litvinukova M., Talavera-Lopez C., Maatz H., Reichart D., Sampaziotis F., Worlock K.B., Yoshida M., Barnes J.L., Network H.C.A.L.B. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat. Med. 2020;26(5):681–687. doi: 10.1038/s41591-020-0868-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Stopsack K.H., Mucci L.A., Antonarakis E.S., Nelson P.S., Kantoff P.W. TMPRSS2 and COVID-19: serendipity or opportunity for intervention? Canc. Discov. 2020;10(6):779–782. doi: 10.1158/2159-8290.CD-20-0451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Lu R., Zhao X., Li J., Niu P., Yang B., Wu H., Wang W., Song H., Huang B., Zhu N., Bi Y., Ma X., Zhan F., Wang L., Hu T., Zhou H., Hu Z., Zhou W., Zhao L., Chen J., Meng Y., Wang J., Lin Y., Yuan J., Xie Z., Ma J., Liu W.J., Wang D., Xu W., Holmes E.C., Gao G.F., Wu G., Chen W., Shi W., Tan W. Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding. Lancet. 2020;395(10224):565–574. doi: 10.1016/S0140-6736(20)30251-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Walls A.C., Park Y.J., Tortorici M.A., Wall A., McGuire A.T., Veesler D. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell. 2020;181(2):281–292 e6. doi: 10.1016/j.cell.2020.02.058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Gao H., Yallampalli U., Yallampalli C. Maternal protein restriction reduces expression of angiotensin I-converting enzyme 2 in rat placental labyrinth zone in late pregnancy. Biol. Reprod. 2012;86(2):31. doi: 10.1095/biolreprod.111.094607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ghadhanfar E., Alsalem A., Al-Kandari S., Naser J., Babiker F., Al-Bader M. The role of ACE2, angiotensin-(1-7) and Mas1 receptor axis in glucocorticoid-induced intrauterine growth restriction. Reprod. Biol. Endocrinol. 2017;15(1):97. doi: 10.1186/s12958-017-0316-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Goyal R., Yellon S.M., Longo L.D., Mata-Greenwood E. Placental gene expression in a rat 'model' of placental insufficiency. Placenta. 2010;31(7):568–575. doi: 10.1016/j.placenta.2010.05.004. [DOI] [PubMed] [Google Scholar]
- 45.Cuffe J.S., Walton S.L., Steane S.E., Singh R.R., Simmons D.G., Moritz K.M. The effects of gestational age and maternal hypoxia on the placental renin angiotensin system in the mouse. Placenta. 2014;35(11):953–961. doi: 10.1016/j.placenta.2014.09.004. [DOI] [PubMed] [Google Scholar]
- 46.Goyal R., Lister R., Leitzke A., Goyal D., Gheorghe C.P., Longo L.D. Antenatal maternal hypoxic stress: adaptations of the placental renin-angiotensin system in the mouse. Placenta. 2011;32(2):134–139. doi: 10.1016/j.placenta.2010.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Tollenaar L.S., Slaghekke F., Middeldorp J.M., Klumper F.J., Haak M.C., Oepkes D., Lopriore E. Twin anemia polycythemia sequence: current views on pathogenesis, diagnostic criteria, perinatal management, and outcome, twin research and human genetics. Off. J. Int. Soc.Twin Stud. 2016;19(3):222–233. doi: 10.1017/thg.2016.18. [DOI] [PubMed] [Google Scholar]
- 48.Mao Q., Chu S., Shapiro S., Yao H., De Paepe M.E. Discordant placental oxygenation and autophagy in twin anemia-polycythemia sequence (TAPS) Placenta. 2020;90:9–17. doi: 10.1016/j.placenta.2019.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Lopriore E., Slaghekke F., Oepkes D., Middeldorp J.M., Vandenbussche F.P., Walther F.J. Hematological characteristics in neonates with twin anemia-polycythemia sequence (TAPS) Prenat. Diagn. 2010;30(3):251–255. doi: 10.1002/pd.2453. [DOI] [PubMed] [Google Scholar]
- 50.Couck I., Lewi L. The placenta in twin-to-twin transfusion syndrome and twin anemia polycythemia sequence, twin research and human genetics. Off. J. Int. Soc.Twin Stud. 2016;19(3):184–190. doi: 10.1017/thg.2016.29. [DOI] [PubMed] [Google Scholar]
- 51.De Paepe M.E., Burke S., Luks F.I., Pinar H., Singer D.B. Demonstration of placental vascular anatomy in monochorionic twin gestations. Pediatr. Dev. Pathol. 2002;5(1):37–44. doi: 10.1007/s10024-001-0089-z. [DOI] [PubMed] [Google Scholar]
- 52.Paepe M.E. Examination of the twin placenta. Semin. Perinatol. 2015;39(1):27–35. doi: 10.1053/j.semperi.2014.10.005. [DOI] [PubMed] [Google Scholar]
- 53.De Paepe M.E., Gundogan F., Mao Q., Chu S., Shapiro S. Redness discordance in monochorionic twin placentas: correlation with clinical and placental findings. Placenta. 2017;60:54–60. doi: 10.1016/j.placenta.2017.10.007. [DOI] [PubMed] [Google Scholar]
- 54.Ravishankar S., Bourjeily G., Lambert-Messerlian G., He M., De Paepe M.E., Gundogan F. Evidence of placental hypoxia in maternal sleep disordered breathing. Pediatr. Dev. Pathol. 2015;18(5):380–386. doi: 10.2350/15-06-1647-OA.1. [DOI] [PubMed] [Google Scholar]
- 55.Chu S., Mao Q., Shapiro S., De Paepe M.E. Placental endoglin levels in diamniotic-monochorionic twin gestations: correlation with clinical and placental characteristics. Placenta. 2013;34(3):261–268. doi: 10.1016/j.placenta.2012.12.012. [DOI] [PubMed] [Google Scholar]
- 56.Danilczyk U., Eriksson U., Crackower M.A., Penninger J.M. A story of two ACEs. J. Mol. Med. (Berl.) 2003;81(4):227–234. doi: 10.1007/s00109-003-0419-x. [DOI] [PubMed] [Google Scholar]
- 57.Matsuyama S., Nagata N., Shirato K., Kawase M., Takeda M., Taguchi F. Efficient activation of the severe acute respiratory syndrome coronavirus spike protein by the transmembrane protease TMPRSS2. J. Virol. 2010;84(24):12658–12664. doi: 10.1128/JVI.01542-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Burrell L.M., Risvanis J., Kubota E., Dean R.G., MacDonald P.S., Lu S., Tikellis C., Grant S.L., Lew R.A., Smith A.I., Cooper M.E., Johnston C.I. Myocardial infarction increases ACE2 expression in rat and humans. Eur. Heart J. 2005;26(4):369–375. doi: 10.1093/eurheartj/ehi114. discussion 322-4. [DOI] [PubMed] [Google Scholar]
- 59.Joshi S., Wollenzien H., Leclerc E., Jarajapu Y.P. Hypoxic regulation of angiotensin-converting enzyme 2 and Mas receptor in human CD34(+) cells. J. Cell. Physiol. 2019;234(11):20420–20431. doi: 10.1002/jcp.28643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Joshi S., Balasubramanian N., Vasam G., Jarajapu Y.P. Angiotensin converting enzyme versus angiotensin converting enzyme-2 selectivity of MLN-4760 and DX600 in human and murine bone marrow-derived cells. Eur. J. Pharmacol. 2016;774:25–33. doi: 10.1016/j.ejphar.2016.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Clarke N.E., Belyaev N.D., Lambert D.W., Turner A.J. Epigenetic regulation of angiotensin-converting enzyme 2 (ACE2) by SIRT1 under conditions of cell energy stress. Clin. Sci. (Lond.) 2014;126(7):507–516. doi: 10.1042/CS20130291. [DOI] [PubMed] [Google Scholar]
- 62.Delforce S.J., Wang Y., Van-Aalst M.E., Corbisier de Meaultsart C., Morris B.J., Broughton-Pipkin F., Roberts C.T., Lumbers E.R., Pringle K.G. Effect of oxygen on the expression of renin-angiotensin system components in a human trophoblast cell line. Placenta. 2016;37:1–6. doi: 10.1016/j.placenta.2015.11.011. [DOI] [PubMed] [Google Scholar]
- 63.Valdes G., Neves L.A., Anton L., Corthorn J., Chacon C., Germain A.M., Merrill D.C., Ferrario C.M., Sarao R., Penninger J., Brosnihan K.B. Distribution of angiotensin-(1-7) and ACE2 in human placentas of normal and pathological pregnancies. Placenta. 2006;27(2–3):200–207. doi: 10.1016/j.placenta.2005.02.015. [DOI] [PubMed] [Google Scholar]
- 64.Pringle K.G., Tadros M.A., Callister R.J., Lumbers E.R. The expression and localization of the human placental prorenin/renin-angiotensin system throughout pregnancy: roles in trophoblast invasion and angiogenesis? Placenta. 2011;32(12):956–962. doi: 10.1016/j.placenta.2011.09.020. [DOI] [PubMed] [Google Scholar]
- 65.Li M., Chen L., Zhang J., Xiong C., Li X. The SARS-CoV-2 receptor ACE2 expression of maternal-fetal interface and fetal organs by single-cell transcriptome study. PloS One. 2020;15(4) doi: 10.1371/journal.pone.0230295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Pique-Regi R., Romero R., Tarca A.L., Luca F., Xu Y., Alazizi A., Leng Y., Hsu C.D., Gomez-Lopez N. Does the human placenta express the canonical cell entry mediators for SARS-CoV-2? Elife. 2020;9:e58716. doi: 10.7554/eLife.58716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Uhlen M., Oksvold P., Fagerberg L., Lundberg E., Jonasson K., Forsberg M., Zwahlen M., Kampf C., Wester K., Hober S., Wernerus H., Bjorling L., Ponten F. Towards a knowledge-based human protein atlas. Nat. Biotechnol. 2010;28(12):1248–1250. doi: 10.1038/nbt1210-1248. [DOI] [PubMed] [Google Scholar]
- 68.Zheng Y.Y., Ma Y.T., Zhang J.Y., Xie X. COVID-19 and the cardiovascular system. Nat. Rev. Cardiol. 2020;17(5):259–260. doi: 10.1038/s41569-020-0360-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Esler M., Esler D. Can angiotensin receptor-blocking drugs perhaps be harmful in the COVID-19 pandemic? J. Hypertens. 2020;38(5):781–782. doi: 10.1097/HJH.0000000000002450. [DOI] [PubMed] [Google Scholar]
- 70.Fang L., Karakiulakis G., Roth M. Are patients with hypertension and diabetes mellitus at increased risk for COVID-19 infection?, the Lancet. Respir. Med. 2020;8(4):e21. doi: 10.1016/S2213-2600(20)30116-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Touyz R.M., Li H., Delles C. ACE2 the Janus-faced protein - from cardiovascular protection to severe acute respiratory syndrome-coronavirus and COVID-19. Clin. Sci. (Lond.) 2020;134(7):747–750. doi: 10.1042/CS20200363. [DOI] [PubMed] [Google Scholar]
- 72.Kuster G.M., Pfister O., Burkard T., Zhou Q., Twerenbold R., Haaf P., Widmer A.F., Osswald S. SARS-CoV2: should inhibitors of the renin-angiotensin system be withdrawn in patients with COVID-19? Eur. Heart J. 2020;41(19):1801–1803. doi: 10.1093/eurheartj/ehaa235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Monteil V., Kwon H., Prado P., Hagelkruys A., Wimmer R.A., Stahl M., Leopoldi A., Garreta E., Hurtado Del Pozo C., Prosper F., Romero J.P., Wirnsberger G., Zhang H., Slutsky A.S., Conder R., Montserrat N., Mirazimi A., Penninger J.M. Inhibition of SARS-CoV-2 infections in engineered human tissues using clinical-grade soluble human ACE2. Cell. 2020;181(4):905–913 e7. doi: 10.1016/j.cell.2020.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Batlle D., Wysocki J., Satchell K. Soluble angiotensin-converting enzyme 2: a potential approach for coronavirus infection therapy? Clin. Sci. (Lond.) 2020;134(5):543–545. doi: 10.1042/CS20200163. [DOI] [PubMed] [Google Scholar]
- 75.Inal J.M. Decoy ACE2-expressing extracellular vesicles that competitively bind SARS-CoV-2 as a possible COVID-19 therapy. Clin. Sci. (Lond.) 2020;134(12):1301–1304. doi: 10.1042/CS20200623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Valdes G., Corthorn J., Bharadwaj M.S., Joyner J., Schneider D., Brosnihan K.B. Utero-placental expression of angiotensin-(1-7) and ACE2 in the pregnant Guinea-pig. Reprod. Biol. Endocrinol. 2013;11:5. doi: 10.1186/1477-7827-11-5. [DOI] [PMC free article] [PubMed] [Google Scholar]