

Abstract
Evolutionarily conserved microRNAs (miRNAs) usually have high copy numbers in the genome. The redundant and specific roles of each member of a multimember miRNA gene family are poorly understood. Previous studies have shown that the miR156-SPL-miR172 axis constitutes a signaling cascade in regulating plant developmental transitions. Here, we report the feasibility and utility of CRISPR-Cas9 technology to investigate the functions of all 5 MIR172 family members in Arabidopsis. We show that an Arabidopsis plant devoid of miR172 is viable, although it displays pleiotropic morphological defects. MIR172 family members exhibit distinct expression pattern and exert functional specificity in regulating meristem size, trichome initiation, stem elongation, shoot branching, and floral competence. In particular, we find that the miR156-SPL-miR172 cascade is bifurcated into specific flowering responses by matching pairs of coexpressed SPL and MIR172 genes in different tissues. Our results thus highlight the spatiotemporal changes in gene expression that underlie evolutionary novelties of a miRNA gene family in nature. The expansion of MIR172 genes in the Arabidopsis genome provides molecular substrates for the integration of diverse floral inductive cues, which ensures that plants flower at the optimal time to maximize seed yields.
This study uses CRISPR-Cas9 technology to investigate the functions of all five miR172 genes in Arabidopsis, finding that miRNA172 family members exhibit distinct expression pattern and exert functional specificity in regulating meristem size, trichome initiation, stem elongation, shoot branching and floral competence.
Introduction
MicroRNAs (miRNAs), a class of small single-stranded noncoding RNAs that range in length from 20 to 22 nucleotides (nt), play important roles in regulating gene expression [1–5]. It has been proposed that miRNAs originate from inverted duplication of target gene fragments, and then undergo diversification through genome-wide duplication, tandem duplication, and segmental duplication, a similar process that drives the evolution of protein gene families in plants [6–9]. As such, the ancient (i.e., evolutionarily conserved) miRNAs such as miR156, miR159/319, miR160, miR165/6, miR171, and miR172 are present in high copy numbers in the genome of Arabidopsis thaliana [10].
Due to the small sizes of the genes, simultaneous inactivation of all miRNA family members by generation of multiple transfer-DNA (T-DNA) mutant lines has so far been achieved for only 2 relatively small families, MIR164 (i.e., MIR164A-C) and MIR159 (i.e., MIR159A-B) [11,12], in Arabidopsis. These 2 studies showed that, similar to protein-coding genes, the MIRNA genes in the same family are functionally redundant. For instance, the mir159ab double mutant displays pleiotropic morphological defects that include curled leaves, short stature, and shorter siliques, whereas the mir159a and mir159b single mutants are phenotypically normal [12].
Target mimicry technologies, such as target mimicry (MIM) and short tandem target mimicry (STTM), have provided effective tools to silence miRNA gene families with more than 3 members by blocking endogenous mature miRNA activity [13–17]. Although generally effective, MIM and STTM apparently have their limitations. For example, residual miRNA is detectable in transgenic MIM plants, suggesting that overexpression of a target mimic is not sufficient to silence the miRNA completely. In addition, because they target the mature miRNA, both MIM and STTM fail to distinguish the functional diversity of each MIRNA gene. Over the past decade, the development of genome editing technology based on the clustered regularly interspaced short palindromic repeats (CRISPR)-Cas9 system has greatly advanced our ability to manipulate specific genome sequences in plants [18–22]. In particular, this method can be used to efficiently construct loss-of-function (null allele) mutants for gene families with multiple members and small gene sizes [23].
In Arabidopsis, miR172 targets a group of transcription factor genes including APETALA2 (AP2), TARGET OF EAT1 (TOE1), TOE2, TOE3, SCHLAFMUTZE (SMZ), and SCHNARCHZAPFEN (SNZ) [24,25]. Previous studies have shown that miR172 and its targets play critical roles in plant developmental transitions [26–29]. miR172 acts downstream of miR156, a miRNA in which the level gradually decreases with time after seed germination [30,31]. Accordingly, miR172 shows a temporal expression pattern that is the opposite of that of miR156 [32]. The increasing level of miR172 promotes the appearance of adult traits including the formation of trichomes (leaf hairs) on the abaxial leaf surfaces [33]. In addition, high amounts of miR172 contribute to the acquisition of floral competence [34–36]. Overexpression of miR172 leads to early flowering [32,37,38], whereas increased levels of its targets, mRNAs from SMZ or SNZ, results in a late flowering phenotype [39]. Genome-wide identification of SMZ and AP2 targets reveals that miR172-targeted AP2-like transcription factors repress flowering through inactivation of the florigen gene FLOWERING LOCUS T (FT) in leaves and MADS-box genes such as APETALA1 (AP1) and SUPPRESSOR OF OVEREXPRESSION OF CO 1 (SOC1) at the shoot apices [39,40].
It has been shown that miR172 also plays a regulatory role in floral patterning in Arabidopsis. AP2 and miR172 show a complementary expression manner in developing floral organs: AP2 is predominantly expressed in the outer floral whorls, while miR172 accumulates to high levels in the centers of the floral primordia [41,42]. The expression of AP2 and miR172 partially overlaps at the boundary between the perianth and the reproductive organs in the third whorl. Reducing miR172 activity by overexpressing MIM172 in the third whorl converts stamens into petals [42].
miR172 belongs to one of the miRNA gene families that are ubiquitous and generally highly expressed across terrestrial plant species [43–46]. Careful sequence analysis has shown that miR172 is not present in either the bryophyte Physcomitrella patens or the lycophyte Selaginella moellendorffii [47], suggesting that the origin of miR172 may play a critical function in the evolution of the vascular plants. However, it is currently unknown whether miR172 is absolutely required for viability in higher plants such as Arabidopsis and rice. In addition, the functional redundancy and specificities among MIR172 genes remain to be clarified. Here, we report the functional investigation of the MIR172 gene family in Arabidopsis using CRISPR-Cas9 technology. We find that the 5 MIR172 gene family members exert functional specificities in regulating diverse aspects of plant developmental processes. The detailed analysis of the role of miR172 in the floral transition suggests that the specificities of MIR172 genes can be explained by their distinct expression patterns and different responsiveness to plant age, ambient temperature, and photoperiod.
Results
Generation of MIR172 knockout mutants by CRISPR-Cas9 technology
In the Arabidopsis genome, the miR172 gene family consists of 5 members, MIR172A to MIR172E (Fig 1A) [25,48]. Because of the small sizes of these genes (<200 base pairs) and their noncoding property, loss-of-function mutants of MIR172 family members in the same genetic background are rarely obtained by canonical mutagenesis methods such as ethyl methanesulfonate treatment, fast neutron irradiation, or T-DNA tagging. To circumvent this problem, we generated mir172 mutants in the Columbia-0 (Col-0) accession of A. thaliana using CRISPR-Cas9 technology with the egg cell-specific promoter [49]. To generate null alleles, we sought to create mutants with large fragment deletions by designing 2 single guide RNAs (sgRNAs) within or flanking the stem-loop region (Fig 1B). Using PCR genotyping, we successfully identified plants carrying mutations in every MIR172 family member in the T1 generation (Fig 1B; S1 Fig). For the MIR172A and MIR172C loci, the entire stem-loop regions were deleted; for the MIR172B and MIR172D loci, the deletions occurred in the loops of the stem-loop regions; for the MIR172E locus, the sequence corresponding to the miRNA* (passenger strand) was deleted.
Fig 1. Generation of MIR172 mutants by CRISPR-Cas9 technology.

(A) Location of the MIR172 genes on Arabidopsis chromosomes. The short and long arms of chromosomes 2, 3, and 5 are shown. (B) The genomic structure of the MIR172 gene loci in WT and the mir172 mutants. Orange, stem-loop of MIR172 gene; green, miRNA*; blue, miRNA. (C) Validation of the mir172 mutants. The WT or mutated versions of MIR172 genes were cloned and overexpressed in WT. Please note that overexpression of the WT version of MIR172 led to early flowering time phenotype. Scale bars represent 1 cm. (D) Flowering time measurement. The number of rosette leaves was counted. Plants were grown at 22°C in long days. The statistically significant differences are determined by ordinary one-way ANOVA (p < 0.05). See also S4 Table. The data underlying this figure are included in S1 Data. CRISPR, clustered regularly interspaced short palindromic repeats; miRNA, microRNA; sgRNA, single guide RNA; WT, wild type.
To confirm that every mutation gave a null allele, the mutated MIR172 genes were cloned and overexpressed individually in wild-type (WT) plants using the constitutive CaMV 35S promoter. Overexpression of the WT MIR172A led to an early flowering phenotype under long days [32,41,50], whereas the overexpression of the mutated versions of the MIR172A, MIR172C, and MIR172E genes did not alter flowering time (Fig 1C and 1D; S1 Table). In agreement with the nature of mutations (i.e., deletions in the loops of the stem-loop regions), the transgenic plants with high levels of mir172b and mir172d flowered slightly earlier than WT (Fig 1C and 1D; S1 Table). Therefore, we conclude that the mir172a, mir172c, and mir172e mutant genes are completely null. In contrast, residual miR172 may exist in the mir172b and mir172d mutants. The mutants were then backcrossed to WT to remove the transgene and off-target mutations. In total, 24 mir172 multiple-mutant lines in different combinations were generated by crossing and PCR-based genotyping (S2 Table).
We examined mature miR172 levels by quantitative real-time PCR (qRT-PCR). As shown in S2A Fig, miR172 accumulates at low levels in the seedling stage and gradually increases with development under long days. The highest expression level was observed in inflorescences (S2A Fig). miR172 was barely detected in the mir172 quintuple mutant (S2B Fig). The comparison of miR172 abundance between WT and the mir172 mutants showed that MIR172A and MIR172B contribute to most of the mature miR172 pool in 12-day-old plants in long days (S2B Fig). In contrast, mutation in MIR172C led to a slight decrease in miR172. The mir172d and mir172e mutants accumulated the same level of miR172 as the WT. Consistent with the notion that miR172 represses its targets mainly through translational inhibition [41], we did not find significant change in abundance of miR172 targets in the mir172 mutants (S2C to S2L Fig). Thus, these results confirm that miR172 expression is nearly abolished in the mir172 quintuple mutants.
Phenotypic analyses of mir172 multiple-mutant lines
As described earlier, although the target mimicry approach can be effective, miR172 is still detectable in the transgenic MIM172 plants [14]. To understand whether miR172 is absolutely required for plant growth and development, we performed phenotypic analyses of the mir172 quintuple mutants. The mir172 quintuple mutants are viable when grown at 22°C in the growth chamber under both long-day and short-day conditions. The quintuple mutants could also survive and produce a large number of seeds when grown outdoors (S3 Fig). This result indicates that, despite its ancient origin, miR172 is not essential for the completion of the seed-to-seed life cycle in Arabidopsis.
Compared to the WT, the mir172 quintuple mutants displayed pleiotropic phenotypes. During the vegetative phase, the development of abaxial trichomes was delayed in the mir172 quintuple mutants under long days (Fig 2A; S4 Fig), which is consistent with previous observations [33]. The contributions of each gene to abaxial trichome initiation were not equal: The mir172ab double mutant exhibited the same early trichome phenotype as the mir172 quintuple mutant (Fig 2A and 2B), whereas mutations in the other MIR172 genes did not affect the timing of trichome initiation. Thus, these results suggest that MIR172A and MIR172B play dominant roles in the timing of trichome initiation (Fig 2I).
Fig 2. Phenotypic analyses of mir172 mutants.

(A) Abaxial trichomes phenotype. The abaxial surface of the 7th leaf was shown. Scale bars represent 200 μm. (B) Quantification of the abaxial trichomes in different genotypes. The 1st leaf with abaxial trichomes was scored. Error bars represent SEM (n = 18–28). (C) Shoot branch phenotype. Scale bars represent 1.0 cm. (D) Quantification of shoot branch numbers in different genotypes. Error bars represent SEM (n = 6–37). The statistically significant differences are determined by ordinary one-way ANOVA (p < 0.05). (E) Internode length phenotype. Scale bars represent 1.0 cm. (F) Quantification of internode length. Error bars represent SEM (n = 27–153). The statistically significant differences are determined by ordinary one-way ANOVA (p < 0.05). (G) Inflorescence SAM size. Orange line, diameter of inflorescence SAM. Scale bars represent 20 μm. (H) Quantification of the size of inflorescence SAM. Error bars represent SEM (n = 10–29). The statistically significant differences are determined by ordinary one-way ANOVA (p < 0.05). (I) Sankey diagram showing the contribution of each MIR172 gene to diverse developmental processes. The individual MIR172 gene was shown in different color. Line thickness stands for the contribution of each gene. The data underlying this figure are included in S1 Data. LD, long days; SAM, shoot apical meristem; SD, short days; WT, wild type.
The mir172 quintuple mutants exhibited striking phenotypes after flowering. The number of branches on the primary bolt was markedly increased (Fig 2C and 2D), accompanied by shortened internodes (Fig 2E and 2F). Interestingly, we found that the meristem size in the quintuple mutant was increased (Fig 2G to 2I). This phenotype is in a good agreement with a recent report that a MADS-box transcription factor FRUITFULL (FUL) regulates the timing of shoot apical meristem (SAM) termination through miR172-targeted AP2-like genes [51]. The comparisons among multiple-mutant lines further revealed that MIR172D plays a key role in regulating SAM size, whereas all of the MIR172 genes except MIR172E cooperatively control internode elongation (Fig 2I). Branch number is mainly regulated by MIR172A and MIR172D. Therefore, our results indicate that the 5 MIR172 gene family members are functionally redundant but exert individual specificities in regulating diverse aspects of plant development (Fig 2I).
It has been proposed that miR172 regulates floral patterning by modulating AP2 expression at the third whorl [35,42,52–54]. Indeed, we found that the first few flowers on the primary bolt showed homeotic transformation (S5A Fig). However, this defect disappeared quickly with the development of the inflorescence (S5B Fig), implying that miR172 may play a minor role in floral patterning during the late reproductive stages under growth chamber conditions.
Flowering time analyses of mir172 mutants under long-day conditions
A previous study showed that down-regulation of miR172 activity by overexpression of a target mimic (35S::MIM172) leads to a late flowering phenotype [14]. However, it remains unclear whether all the MIR172 family members are involved in flowering time regulation and, if so, whether the different members contribute differently to this process. To answer these questions, we scored the flowering times of the single and higher-order mir172 mutant plants grown under both long and short days. As shown in Fig 3B and S1 Table, the mir172a mutant, but not other mir172 single mutants, exhibited a late flowering phenotype in long days. Consistently, the mir172bcde quadruple mutant flowered at the same time as WT (Fig 3A and 3B; S1 Table). Moreover, the double or triple mutant combinations that included mir172a flowered significantly later than WT, whereas the combinations without the mir172a mutation flowered normally, like WT plants (Fig 3A and 3B; S1 Table). MIR172E appears to play only a minor role in regulating flowering time because the mir172abcd quadruple mutant had the same flowering time as the mir172 quintuple mutant. Both the mir172abcd and mir172abde quadruple mutants flowered later than mir172acde, suggesting that MIR172B makes a modest contribution to the floral transition. Taken together, we conclude that the contribution of each MIR172 gene family member to flowering time under long days is: MIR172A>MIR172B>MIR172D>MIR172C>MIR172E (Fig 2I).
Fig 3. Flowering time of WT and the mir172 mutants in long days.
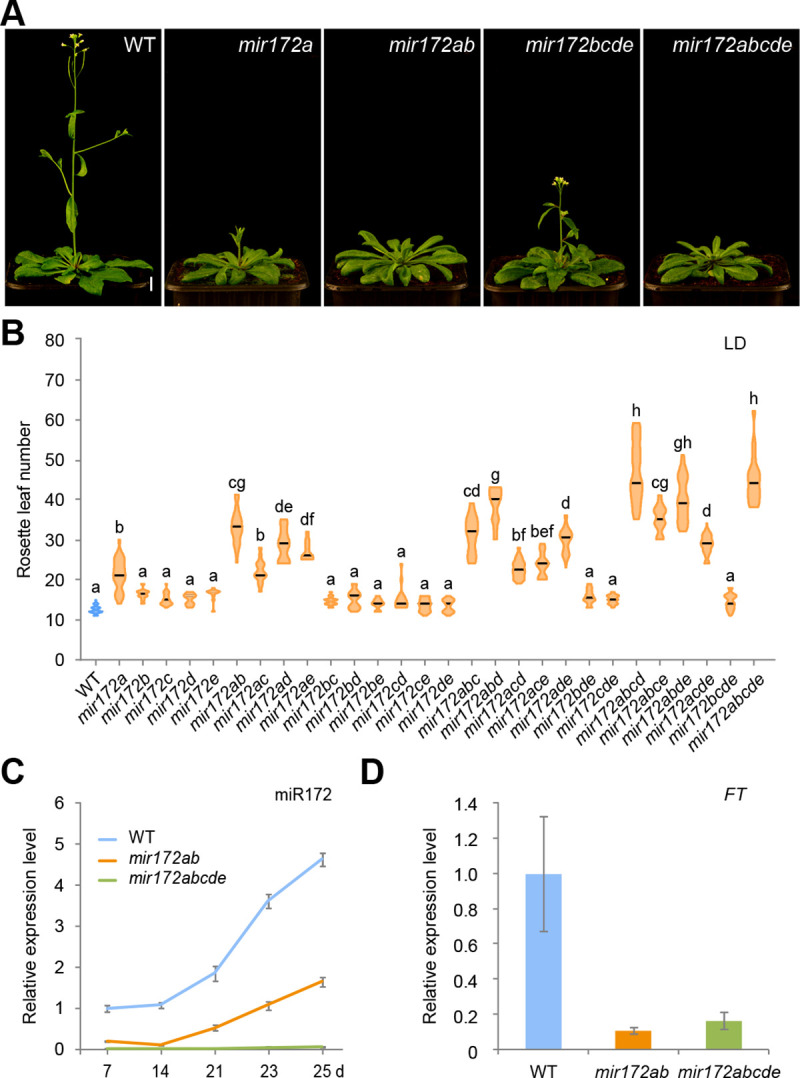
(A) Flowering time phenotype of the mir172 mutants. Plants were grown at 22°C in long days. One representative plant is shown. (B) Quantification of flowering times of the mir172 mutants. Lines show mean (n = 7–36). The statistically significant differences are determined by ordinary one-way ANOVA (p < 0.05). See also S1 and S4 Tables. (C) Expression of miR172 in WT, mir172ab, and mir172abcde mutants. Plants were grown at 22°C in long days. The shoot apices and developing leaves were used for qRT-PCR analysis. Expression was normalized to TUB. Two technical replicates for each biological replicate (n = 2) were performed. Error bars represent SD. (D) Expression of FT in WT and the mir172 mutants. The leaves of 15-day-old plants grown at 22°C in long days were harvested at ZT16 and used for qRT-PCR analyses. Expression was normalized to TUB. Two technical replicates for each biological replicate (n = 3) were performed. Error bars represent SD. The data underlying this figure are included in S1 Data. FT, FLOWERING LOCUS T; LD, long days; qRT-PCR, quantitative real-time PCR; TUB, β-TUBULIN-2; WT, wild type; ZT16, Zeitgeber time 16.
It has been shown that the level of miR172 increases with age, and this promotes the gain of reproductive competence [33]. qRT-PCR assays showed that the gradual increase in miR172 levels was largely compromised in the mir172ab double mutant (Fig 3C). Induction of FT by the photoperiod pathway plays a critical role in flowering in long days [55]. The mir172ab double mutant plants had lower levels of FT than did WT plants in long days (Fig 3D), indicating that these 2 MIR172 members alleviate the repression of FT expression by miR172-targeted AP2s in leaves.
Expression pattern of MIR172 genes in plants grown under long days
We generated the fluorescent protein [green fluorescent protein (GFP) or Venus]-based reporters for the 5 MIR172 gene family members. To ensure that the promoter regions covered all of the regulatory sequences, we surveyed our transposase-accessible chromatin sequencing (ATAC-seq) datasets, which reveal the chromatin accessibility at a given gene locus [56]. The promoter sequences that contained all of the accessible regions in the intergenic region were cloned and placed upstream of the coding regions of GFP or Venus (S6 Fig). For each construct, we examined over 20 individual T1-generation lines that gave consistent and reliable expression patterns. We chose 1 representative T2 line for the subsequent analyses. We did not detect the GFP fluorescent signals for MIR172E under long days (S7 Fig), suggesting that MIR172E is weakly expressed and is not required for development when the plants are grown under LD conditions.
We next focused on the expression of the MIR172 reporter genes in the leaf vascular tissues or the SAM, where the floral induction occurs. In long days, MIR172B and MIR172C were expressed at low levels in seedlings and became detectable in the vascular tissues when the plants entered the adult phase and eventually flowered (Fig 4B and 4C). MIR172A is actively transcribed in both vascular tissue and the SAM, and its promoter strength increased as development progressed (Fig 4A). MIR172D exhibited a different expression pattern in which the Venus fluorescence was constitutively observed at the margins of Arabidopsis leaves (Fig 4D). A high level of expression of MIR172D was evident in the SAM of 16-day-old plants. The temporal expression pattern of these reporters was verified by qRT-PCR and quantification of GFP or Venus densities (Fig 4E to 4I; S8 Fig). Overall, these analyses are consistent with the abovementioned genetic analysis data where MIR172A and MIR172B play predominant roles in flowering in long days. The activation of these 2 genes in the leaf vascular tissues primes the activation of FT by the photoperiod pathway.
Fig 4. Expression pattern of MIR172 genes in long days.

(A–D) Analyses of MIR172 reporters. Plants were grown at 22°C in long days. The leaves (upper panels) and shoot apices (lower panels) were examined. Please note that the plants start to flower 3 weeks (approximately 21 days) after seed germination. Over 20 T1 independent lines for each reporter were examined, and the results of 1 representative T2 line are shown. Dash line marks the SAM. Scale bars represent 100 μm. (E–I) Expression of MIR172 genes (E–H) and AP1 (I) in WT. AP1 was monitored as an indication for floral transition. Plants were grown at 22°C in long days and harvested at different time points. Expression was normalized to TUB. Two technical replicates for each biological replicate (n = 2) were performed. Error bars represent SD. (J) Expression of the MIR172B reporter in WT and the spl9 mutant. One representative leaf of the same developmental age is shown. Scale bars represent 100 μm. The same confocal settings were used for scanning for each reporter line (A–D and I). The data underlying this figure are included in S1 Data. AP1, APETALA1; SAM, shoot apical meristem; TUB, β-TUBULIN-2; WT, wild type.
Genetic analyses have placed miR172 downstream of miR156-targeted SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) genes [57–60]. The gradual decrease in miR156 levels with increasing plant age leads to the up-regulation of SPL genes, which subsequently activates miR172 [33,61]. In situ hybridization assays have shown that SPL9 is expressed in leaf anlagen and the vascular tissues [30,62]. Compared to WT, the promoter activities of MIR172B and MIR172C were moderately reduced in the leaves of spl9 mutant plants (Fig 4J; S9B and S9D Fig), suggesting that SPL9 contributes to the increased level of miR172 in the leaves of adult plants through these 2 MIR172 gene family members. The transcriptional activity of MIR172A was not altered in the spl9 mutant, suggesting that one or more other miR156-targeted SPLs may regulate its expression (S9A Fig). Indeed, our recent work has shown that the SPL2, SPL10, and SPL11 genes are also highly expressed in leaf vascular tissues [63].
MIR172A and MIR172D play dominant roles in determining flowering time under short days
In short days, we found that MIR172D played a critical role in flowering because the mir172d single mutant exhibited an obvious late flowering phenotype (Fig 5A and 5B; S1 Table). This phenotype was further enhanced by mutation of MIR172A. The mir172ad double mutant plants flowered nearly as late as the mir172 quintuple mutant plants (Fig 5B; S1 Table). In contrast, the flowering times of the other double mutants was comparable to that of WT. It is well documented that the activation of a subset of MADS-box genes and LFAFY in the SAM evokes flowering in short days [55,64]. In situ hybridization showed that miR172 abundance is markedly reduced in the shoot apices of the mir172ad double mutant (Fig 5C). Expression analyses further revealed that the up-regulation of MADS-box genes such as FUL and SOC1 is delayed in the shoot apices of mir172ad mutant (Fig 5D). Therefore, we conclude that the relative contribution of each MIR172 gene family member to flowering time in short days is: MIR172D>MIR172A>MIR172B = MIR172C>MIR172E (Fig 2I).
Fig 5. Flowering time of WT and the mir172 mutants in short days.
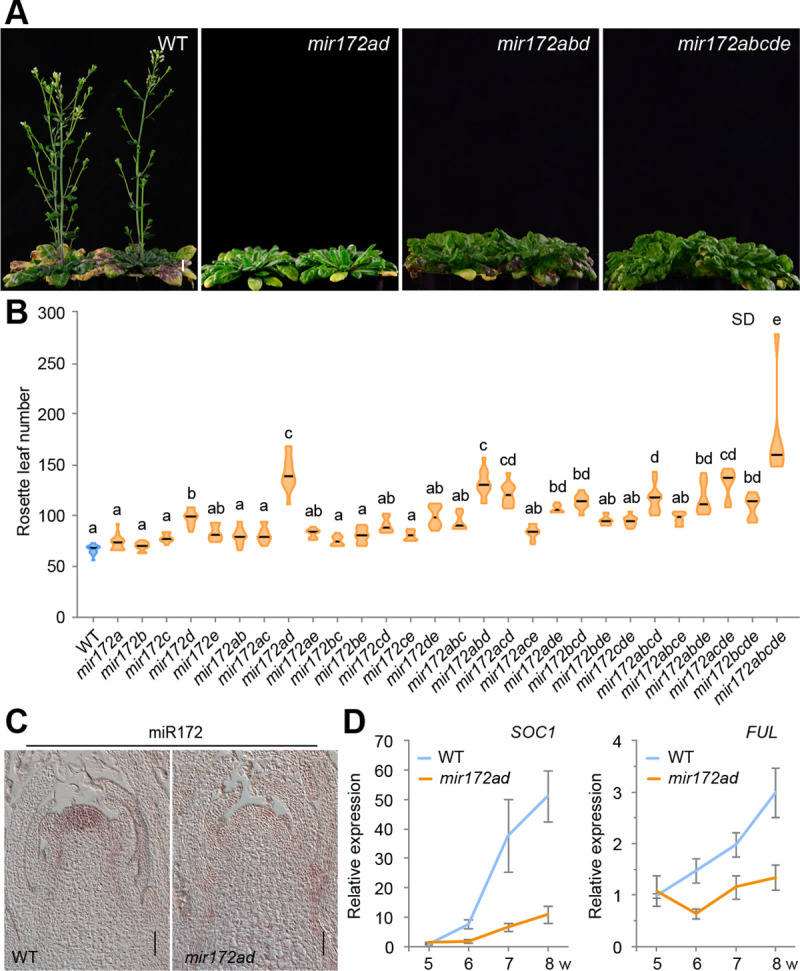
(A) Flowering time phenotype of the mir172 mutants. Plants were grown at 22°C in short days (SD). Four representative plants are shown. (B) Quantification of flowering times of the mir172 mutants. Lines show mean (n = 6–16). The statistically significant differences are determined by ordinary one-way ANOVA (p < 0.05). See also S1 and S4 Tables. (C) Expression of miR172 in the shoot apices of WT and the mir172ad mutant. Plants were grown at 22°C in short days. Scale bars represent 50 μm. (D) Expression of SOC1 and FUL in WT and the mir172ad mutants. Plants were grown at 22°C in short days, and the shoot apices were harvested at different time points as indicated. Expression was normalized to TUB. Two technical replicates for each biological replicate (n = 2) were performed. Error bars represent SD. The data underlying this figure are included in S1 Data. FUL, FRUITFULL; SOC1, SUPPRESSOR OF OVEREXPRESSION OF CO 1; TUB, β-TUBULIN-2; WT, wild type.
We next analyzed the MIR172 reporter activities in plants grown in short days. Expression of MIR172B and MIR172C was barely detectable in the vegetative phase (Fig 6B and 6C; S10 Fig). Consistent with the role of MIR172A and MIR172D in flowering under short days, both reporters were active in the SAM, with the promoter strength increasing as development progressed (Fig 6A and 6D; S10 Fig). Among the miR156-targeted SPL genes, SPL15 is predominantly expressed in the SAM and coordinates the basal floral promotion pathways required for flowering under noninductive short-day conditions [61]. Consistent with this notion, the expression of MIR172D was greatly reduced in the SAM of the spl15 mutant (Fig 6F; S9C and S9D Fig). On the contrary, MIR172A expression was largely unaffected by the mutation in SPL15 (S9A and S9D Fig).
Fig 6. Expression pattern of MIR172 genes in short days.

(A–E) Analyses of MIR172 reporters. Plants were grown at 22°C in short days. The shoot apices were examined. Please note that the plants start to flower after 10 weeks. Over 20 T1 independent lines for each reporter were examined, and the results of 1 representative T2 line are shown. Dash line marks the SAM. Scale bars represent 100 μm. (F) Expression of MIR172D reporter in WT and the spl15 mutant. Dash line marks the SAM. Scale bars represent 100 μm. The same confocal settings were used for scanning for each reporter line. SAM, shoot apical meristem; WT, wild type.
In summary, the flowering times and expression analyses described above suggest that MIR172A, MIR172B, and MIR172D are the primary regulators of flowering time in the MIR172 gene family. The spatially localized SPL-MIR172 pairs promote the acquisition of floral competence in different tissues under different growth conditions (Fig 8A and 8B). In long days, miR172 functions in both the SAM and vascular tissues, with MIR172A, MIR172B, and MIR172D playing dominant roles. In the vasculature of leaves, MIR172B is induced by SPL9, whereas MIR172A is activated probably by other miR156-targeted SPLs. The up-regulation of MIR172A and MIR172B relieves the repression of FT by miR172-targeted genes that encode AP2-like transcription factors. Consequently, FT is activated in leaves by the photoperiod pathway, and the mobile protein is transported to the SAM where it induces the floral transition. The SPL15-MIR172D pair in the SAM plays a less important role in flowering under long-day conditions because the photoperiod pathway is dominant in Arabidopsis, and the repressive role of miR172-targted AP2s on floral transition in the shoot apex can be eventually bypassed by FT.
Fig 8. Distinct expression pattern and different responsiveness of MIR172 genes to plant age, ambient temperature, and photoperiod.

(A) Schematic of the expression pattern of MIR172 genes. Individual MIR172 gene is shown in different color. Two SPL/MIR172 pairs (SPL9-MIR172B/C and SPL15-MIR172D) contribute to the acquisition of floral competence in leaf vascular tissue and shoot apex, respectively. (B) Mode of MIR172 genes in regulating flowering time in LD and SD. Different combinations of MIR172 genes are shown in different colors. In LD, the products of FT move from leaves to shoot apex (black lines). FT, FLOWERING LOCUS T; FUL, FRUITFULL; LD, long days; SD, short days; SOC1, SUPPRESSOR OF OVEREXPRESSION OF CO 1.
In short days, miR172 activity is crucially important for floral transition at the SAM, with MIR172A and MIR172D being the most important (Fig 8A and 8B). The gradual increase in SPL15 levels promotes the transcription of MIR172D in the SAM. As a result, the accumulation of mature miR172 leads to down-regulation of miR172-targeted AP2-like genes, which eventually facilitates the activation of floral promoting MADS-box genes. How MIR172A is progressively activated in the SAM in short days is currently unknown. Possible activators include SPL2 and SPL10 which are also highly abundant in the SAM (S11 Fig). It should be noted that the contribution of MIR172B to floral transition in short days may be underscored because the mutant allele is not completely null (Fig 1C and 1D).
The role of MIR172 genes in response to ambient temperature
Previous studies have shown that overexpression of miR172 leads to an early flowering phenotype at both 16 and 23°C [65,66]. Moreover, miR172 levels are higher at 23°C than at 16°C, probably due to an enhancement in the processing of the primary miR172 transcripts mediated by the Arabidopsis RNA-binding protein FLOWERING CONTROL LOCUS A (FCA) [66]. However, due to the extremely early flowering phenotype of the miR172-overexpression line, it remains unclear whether miR172 is indeed required for thermosensory flowering. We found that the flowering of both WT and mir172 quintuple mutant plants was accelerated at 28°C (Fig 7A), indicating that elevated ambient temperature promotes flowering independent of miR172. In contrast, although the flowering of the WT plants was delayed in 16°C, the mir172 quintuple mutants started to bolt with nearly the same number of rosette leaves at 16 and 22°C (Fig 7B and 7C). Thus, this result indicates that low ambient temperature regulates flowering largely through miR172.
Fig 7. Expression of MIR172 genes in response to environment cues.

(A) Comparison of the flowering time of WT and the mir172abcde mutants grown at 22°C and 28°C in long days. Rosette number ratio was calculated by the mean value at 22°C divided by that at 28°C. (B) Comparison of the flowering time of WT and the mir172 mutants grown at 16°C and 22°C in long days. Rosette number ratio was determined by the mean value at 16°C divided by that at 22°C. (C) Quantification of flowering time phenotype of the mir172 mutants at 16°C in long days. Lines show mean (n = 18–24). The statistically significant differences are determined by ordinary one-way ANOVA (p < 0.05). See also S1 and S4 Tables. (D and E) Expression of miR172 (D) and MIR172 genes (E) at 16°C and 22°C in long days. Expression was normalized to TUB. Two technical replicates for each biological replicate (n = 3) were performed. Error bars represent SD. (F) Analyses of MIR172C and MIR172D reporters in the shoot apices. Plants were grown at 16°C in long days. (G) Expression of MIR172A, MIR172B, and MIR172D reporters in response to photoperiod. The plants were grown in short days (SD) for 3 weeks and shifted to long days (shift + 3 d or 5 d). The same confocal settings were used for scanning for each reporter line (F and G). Dash lines mark the SAM. See also S13 Fig. The data underlying this figure are included in S1 Data. LD, long days; SAM, shoot apical meristem; SD, short days; TUB, β-TUBULIN-2; WT, wild type.
To understand which MIR172 genes are responsible for the temperature response, we compared the flowering times of the single and higher-order mir172 mutants grown at different temperatures. As shown in Fig 7B, all single mir172 mutants showed moderate reduction in thermosensory flowering response with MIR172A likely playing a major role within the gene family (Fig 7B and 7C). The double-mutant plants harboring the mutation in MIR172A (mir172ab, mir172ac, and mir172ad) flowered at nearly the same time when grown at 16°C compared to 22°C (Fig 7B and 7C).
We next explored the expression of individual MIR172 genes in response to low ambient temperature. Consistent with previous results [65,66], qRT-PCR assays revealed that the abundance of miR172 was elevated in plants grown at 22°C, as compared to 16°C (Fig 7D). Among the reporters examined, we found that MIR172C and MIR172D responded to the temperature change (Fig 7E; S12 Fig). The expression of MIR172C was greatly increased in the vasculature of leaves and the inflorescence stem, whereas the promoter activity of MIR172D was enhanced with the increase in temperature, mainly in the vascular tissues and leaf epidermis (Fig 7F). Altogether, these results indicate that, in addition to modulating miR172 abundance at the posttranscriptional level as previously proposed [66], low ambient temperature may control flowering by regulating the expression of MIR172C and MIR172D at the transcriptional level (Fig 8A).
The expression of MIR172 genes in response to photoperiod
Finally, we explored whether photoperiod, another exogenous floral inductive cue, can affect the transcription of MIR172 genes. To this end, we grew the transgenic plants expressing the MIR172 reporters in short days for 3 weeks and then shifted them to long days. As shown in Fig 7G and S13 Fig, we did not detect strong induction of all the reporters 1 day after the plants were shifted to long-day conditions. At day 3, the transcriptional activities of MIR172A and MIR172D were strongly induced in the shoot apex, whereas increases in MIR172A and MIR172B expression were observed in the leaf vascular tissues (Fig 7G; S13 Fig). The expression of MIR172C did not respond to long days (S13 Fig). We did not find ectopic expression of Venus or GFP, suggesting that long-day conditions only regulate the amplitude of MIR172 expression. Because the expression of floral-promoting genes such as FUL and SOC1 are rapidly induced in the SAM 1 day after the shift [30], the increased levels of MIR172A and MIR172D in the SAM may be caused by a cell fate transition rather than by a direct photoperiod response. Therefore, photoperiod modulates miR172 abundance at least at two different levels: transcriptional activation of MIR172A and MIR172B in the vasculature of leaves (Fig 8A) and promoting the processing of miR172 precursors through the plant circadian oscillation regulator GIGANTEA (GI) [67].
Discussion
Evolutionarily conserved miRNAs are usually encoded by multiple gene families. The comprehensive characterization of the function of each of the gene family members is difficult, largely due to a lack of null knockout mutants. With the development of CRISPR-Cas9 technology, it is now possible to generate the corresponding mutant alleles and investigate their biological functions. More importantly, the generation of higher-order mutants carrying different combinations of mutations in each miRNA gene family member allows us to explore the function and contribution of each member. In this study, we have demonstrated that the Arabidopsis MIR172 gene family members are not only redundant but also confer functional specification (Fig 2I). Our reporter analyses further reveal that this specificity can be attributed to their distinct expression pattern, in addition to variations in miRNA processing efficiency. For example, consistent with its being highly expressed in the SAM (Fig 4D), MIR172D is the only family member that regulates SAM size. The promote activity of MIR172D could also be detected in other tissues such as lateral shoots and vascular bundles in the stem (Fig 4D), where it participates in the control of shoot branching and internode elongation. Similar result has been reported for the Arabidopsis MIR164 family, where MIR164C plays a specific role in regulation of petal number [68].
How MIR172 family members are differentially regulated by SPLs in Arabidopsis is poorly understood. First, although the SPL mRNA level is extremely low, miR172 is still detectable in the juvenile phase. This result suggests that some unknown transcription factors contribute to the basal level of miR172 at this developmental stage. It is unlikely that this putative transcription factor activates miR172 expression through MIR172E because our genetic and reporter analyses showed that MIR172E is largely not functional under normal growth conditions. Second, it is also unclear why MIR172A, MIR172B, and MIR172C are highly expressed in the leaf vascular tissues. It is possible that this tissue specificity is generated by transcriptional activators, including SPLs (SPL9, SPL10, and SPL11) and GI, which are predominantly expressed in the vascular tissues [63,67]. Third, we have shown that the SPL15-MIR172D pair regulates floral transition in the SAM (Fig 8). Interestingly, the predominant expression of SPL15 in the SAM is conserved within species in the Brassicaceae [69]. Therefore, identification and analysis of the cis-elements that confer this tissue specificity is an important future research direction. It also remains unknown why SPL9 or SPL10 are not able to activate MIR172D in leaves. One possibility is that the MIR172D locus is transcriptionally competent only in the SAM. In this scenario, SPL15 would activate MIR172D with the help of a cofactor which is exclusively expressed in the meristem. A promising candidate is FD, which is expressed only in the SAM and induces flowering through its interaction with FT [70,71]. Moreover, in support of this hypothesis, the ATAC-seq datasets have revealed that the MIR172D locus is largely inaccessible in the 1st and 7th leaves (S6 Fig).
Previous reports have shown that the miR172 level varies in response to the ambient temperature [65,66,72]. Interestingly, the data presented in this manuscript show that, in addition to being regulated posttranscriptionally by FCA, environmental temperature can influence miR172 levels by modulating at least 2 MIR172 genes (MIR172C and MIR172D) at the transcriptional level [66]. The molecular link between temperature sensing and MIR172 transcription is unknown at present. Genetic analyses have demonstrated that 2 MADS-box floral repressors SHORT VEGETATIVE PHASE (SVP) and FLOWERING LOCUS M (FLM) play critical roles in flowering time at 16°C [73–76]. While FLM is regulated by temperature-dependent alternative splicing [77–79], SVP is subjected to temperature-dependent destabilization [80]. The FLM-SVP dimer represses flowering through FT and SOC1 [80]. Therefore, it is plausible to assume that the FLM-SVP complex may bind to the promoter of MIR172C and MIR172D and suppress their expression at 16°C. Genome-wide mapping of the binding sites of the FLM-SVP complex and identification of the temperature responsive cis-elements in the MIR172C and MIR172D promoters will be necessary to test this hypothesis in the future.
Our findings have useful implications with respect to the regulation of flowering time in crops. Genome-wide analyses have found that, as in Arabidopsis, the MIR172 genes are present as multigene families in the rice, maize, and wheat genomes [81–86]. The functions of the individual MIR172 genes in flowering and floral patterning is largely unexplored. The Hake lab has shown that tasselseed4 (ts4) is encoded by a MIR172 gene in maize [87]. The mutation in ts4 permits carpel development in the tassel while increasing meristem branching. Thus, this result implies that MIR172 genes may also undergo functional specification in some crop species. Undoubtedly, careful comparisons between wild species and cultivars will shed light on whether the natural variations in SPL-MIR172 pairs underlie the evolution of plasticity in flowering time that has occurred during crop domestication.
In conclusion, our results reveal that the expansion of the MIR172 gene family in the Arabidopsis genome provides molecular substrates for the integration of diverse floral inductive cues including age, photoperiod, and temperature. The matching pairs of coexpressed SPL and MIR172 genes ensure the precise acquisition of floral competence under different conditions in order to maximize seed yields.
Methods
Plant materials and growth conditions
A. thaliana (ecotype Col-0) plants were grown at 22°C (day)/19°C (night) in long days (16 hours light/8 hours dark) or at 21°C (day)/18°C (night) short days (8 hours light/16 hours dark). For growth under natural conditions, plants were sowed on the soil and grown in the pots (5 × 5 cm) in Shanghai (SIPPE, Fenglin campus) from December 2018 to May 2019. The average temperature was 10°C (day)/5°C (night). The day length varied from 12 hours light/12 hours dark (December 2018) to 14 hours light/10 hours dark (May 2019). For temperature assays, the plants were grown in the chamber (Percival) at 16°C or 28°C in long days. For transgenic A. thaliana plants, the binary constructs were delivered into Agrobacterium tumefaciens strain GV3101 (pMP90) by freeze-thaw method. Transgenic plants were generated by floral dipping method [88] and screened with 0.05% glufosinate (Basta) on soil, 40 μg/ml hygromycin or 50 μg/mL kanamycin on half-strength Murashige and Skoog (MS) media.
Constructs
The oligonucleotide primers for all the constructs are given in S3 Table. The SnapGene map for each construct is available upon request. To generate MIR172 reporters, the promoter regions of MIR172 genes were PCR amplified and cloned in front of GFP-N7 or 3xVenus-N7 coding region in the binary vector VV00 (S6 Fig).
Generation of mir172 mutants by CRISPR-Cas9
Gene-targeting vectors were constructed based on the pHEE2A-TRI (rbcS-E9t) system provided by Dr. Qi-Jun Chen [49]. The sgRNA sequences for each MIR172 gene were designed using tools at CRISPR-P 2.0 (http://crispr.hzau.edu.cn/CRISPR2/) according to website instructions [89]. The oligonucleotide primers for sgRNAs are given in S3 Table. The mutants were identified by PCR. The mutants were backcrossed to WT to remove transgene and potential off-target mutations. The high-order mutants were generated by crossing and PCR-based genotyping.
Phenotypic analyses
For flowering time measurement, the seeds of different genotypes were sowed on soil. The plants were grown under long-day or short-day conditions. Because the leaf initiation rate of the mir172 mutants was comparable to that of WT, the flowering time was directly measured by counting the total number of rosette leaves when plants started bolting (10 cm in height). The branches on the primary bolt and the average internode length were scored after the inflorescence SAMs were fully terminated. The floral patterning phenotype were examined using Olympus BX63 equipped with DP73 digital camera.
Statistical analyses
We used ordinary one-way ANOVA to perform statistical analyses. Brown–Forsythe test and Bartlett test were performed whenever multiple samples were compared. Statistical significance was determined at p < 0.05 unless otherwise indicated. Statistical test results are given in S4 Table. The original data were given in S1 and S2 Data and deposited in https://figshare.com/articles/dataset/202011_Lian_PLOS_B_Original_data_xlsx/13293722.
Microscopy
The inflorescence SAM samples were processed and scanned using JSM-6360LV Scanning Electron Microscope (SEM, JEOL). The SEM images were analyzed with Image J software as described [90,91].
Phenotypic analyses and imaging of floral organs and meristems were examined using Olympus BX63 equipped with DP73 digital camera as described [90,91]. For RNA in situ imaging, slides were mounted with water as described and observed under Olympus BX63 equipped with DP73 digital camera and differential interference contrast module. To compare the MIR172 reporter activities between WT and the spl mutants, the MIR172 reporter line was crossed to the spl9 or spl15 mutant. The homozygous mutants were identified in F2 and used for analyses.
For confocal imaging, the flowers and inflorescences were dissected on 2% agar plate under a stereo microscope. For the vegetative samples collected from seedlings, the tissues were collected and immediately placed in the vials with ice-cold phosphate buffered saline (PBS) containing 2.5% paraformaldehyde (PFA (pH 7.0)). The samples were infiltrated for 30 minutes by vacuum and stored at 4°C overnight. Tissues were then washed with sucrose gradient PBS-PFA solution, embedded with 6% low melting agarose, and sliced with a Lecia Sliding Microtome 1200S at the thickness of 50 μm. For the vegetative samples collected from adult plants, the shoot apices were dissected by free-hand sectioning under a stereo microscope. Selected sections were stained in 10 μM FM4-64 solution (Sigma, Merck, China) or mounted directly with water without staining.
Prepared specimen was observed and scanned with Olympus FV3000 or Leica SP8 confocal microscope. Proper filter sets and lasers were selected for fluorescence signal scanning. All specimens were scanned with 10× objective (HCX PL APO CS 10X/0.40 DRY on Leica SP8 or UPLXAPO 10X/0.40 on Olympus FV3000). For GFP, excitation light wave length was 488 nm; emission, 500 to 550 nm. For Venus, excitation light wave length was 415 nm (on Leica SP8) or 488 nm (on Olympus FV3000); emission, 520 to 550 nm. For chlorophyll and FM4-64, excitation light wave length was 415 nm (on Leica SP8) or 488 nm (on Leica SP8 or Olympus FV3000), emission, 650 to 750 nm. The same settings for visualizing GFP or Venus were used for each sample. The settings for visualizing plant cell walls by FM4-64 was modified for presentation purposes. The interpretation of the results was not affected, and the original images are available upon request.
Quantification of reporter intensity (S8 Fig; S10B Fig) was performed using Image J software. Briefly, the intensities of GFP or Venus in the selected area (vascular tissues or the SAM) were measured. For each MIR172 reporter line, the images taken from 3 to 6 individual plants were measured.
Expression analyses
Total RNA was extracted from seedlings, leaves, roots (harvested from 7-day-old seedlings), or shoot apices with Trizol reagent (ThermoFisher, Cat No./ID: 15596018). A total of 1 μg of total RNA was DNase I-treated (1 unit/mL; ThermoFisher, Cat No./ID: EN0521) and used for cDNA synthesis with oligo (dT) primer and/or miR172-RT-Primer A primer using RevertAid RT Reverse Transcription Kit (ThermoFisher, Cat No./ID: K1691) (S14A Fig). The average expression levels and standard errors were calculated from 2−ΔΔCt values. Two or three biological replicates were performed. For each biological replicate, 2 technical replicates were performed. The qRT-PCR primers for TUB have been described [30]. The oligonucleotide primers for all the genes are given in S3 Table. qRT-PCR on mature miR172 was performed as described [92,93]. The difference among miR172 isoforms resides in the last nucleotide (S14A Fig). The miR172-RT-Primer A primer used in this study did not distinguish these isoforms (S14B Fig).
RNA in situ hybridization
Shoot apices from short-day grown plants of different ages were dissected and fixed in formalin:acetic acid:ethanol (1:1:18). Paraffin-embedded materials were sectioned to 8 μm thickness. RNA in situ hybridization was performed as described [62,90]. For SPL2 and SPL10 probes, the cDNA fragments of SPL2 and SPL10 were PCR amplified and cloned into pBluescript SK, respectively. In vitro transcription was performed with T3 or T7 RNA polymerase (ThermoFisher, Cat No./ID: EP0101/EP0111), in which linearized vectors were used as templates. For miR172 probe, Locked Nucleic Acid (LNA) oligonucleotide was end labeled with the DIG oligonucleotide 3′-end labeling kit (Roche, Cat No./ID: 03 353 575 910).
Accession numbers
Sequence data from this article can be found in the Arabidopsis Genome Initiative or GenBank/EMBL databases under the following accession numbers: MIR172A (At2g28056), MIR172B (At5g04275), MIR172C (At3g11435), MIR172D (At3g55512), MIR172E (At5g59505), SPL2 (At5g43270), SPL9 (At2g42200), SPL10 (At1g27370), and SPL15 (At3g57920).
Supporting information
The genomic sequence of WT and mutated MIR172 loci. Orange, stem-loop of MIR172; green, miRNA*; blue, miRNA. CRISPR, clustered regularly interspaced short palindromic repeats; miRNA, microRNA; WT, wild type.
(TIF)
(A) Expression of miR172 in different tissues in WT. Seven-day-old seedlings, the 6th and 8th leaves, roots, and inflorescence were used. Two technical replicates for each biological replicate (n = 2) were performed. Error bars represent SD. (B) Expression of miR172 in the mir172 mutants. Twelve-day-old plants in long days were used for qRT-PCR analyses. Two technical replicates for each biological replicate (n = 2) were performed. Error bars represent SD. (C to G) Expression of miR172 targets in the mir172 mutants. Twelve-day-old plants in long days were used for qRT-PCR analyses. Two technical replicates were performed. Error bars represent SD. (H to L) Time course analysis of abundance of miR172 targets. Seven-day, 14-day, and 21/25-day-old plants in long days were used for qRT-PCR analyses. Two technical replicates for each biological replicate (n = 2) were performed. Error bars represent SD. For all the qRT-PCR assays, the expression level was normalized to that of TUB. The data underlying this figure are included in S2 Data. qRT-PCR, quantitative real-time PCR; TUB, β-TUBULIN-2; WT, wild type.
(TIF)
The photos were taken after plants flowered. Scale bar represents 1 cm.
(TIF)
WT and the mir172 mutants were grown at 22°C in long days. The abaxial surfaces of the leaves are shown. Scale bar represents 200 μm.
(TIF)
(A) The flowers of WT and the mir172 mutants. The flowers with abnormal phenotypes are shown. Scale bars represent 500 μm. (B) Quantification of abnormal floral phenotype in different genotypes. The floral phenotype of 5 to 21 plants for each genotype was examined. The data underlying this figure are included in S2 Data.
(TIF)
The ATAC-seq tracks for the MIR172 genes are shown. The datasets are derived from 4 plant tissues including the 1st leaf, the 7th leaf, embryo, and seedlings [56]. The orange box and line indicate stem loop region and the sequences used for generation of MIR172 reporter, respectively. For MIR172E reporter, the 1.3 kilobase pair (kb) downstream sequence was also included. The ATAC-seq datasets are deposited in Beijing Institute of Genomics Data Center (http://bigd.big.ac.cn) with the accession number (BioProject PRJCA002620 and BioProject PRJCA003872). The data underlying this figure are included in S2 Data.
(TIF)
The leaves (A) and shoot apices (B) are shown. Plants were grown at 22°C in long days. The same pinhole size was used for scanning. Dash line marks the SAM. Scale bars represent 100 μm.
(TIF)
Plants were grown at 22°C in long days. See also Fig 4. The data underlying this figure are included in S2 Data.
(TIF)
(A to C) Expression of MIR172 reporter genes in the spl9 and spl15 mutants. Plants were grown at 22°C in long days. Please note that the promoter activity of MIR172C was attenuated in the spl9 mutant but not in the spl15 mutant (B). In contrast, the transcription of MIR172A was largely unaffected by the mutation in SPL9 or SPL15 (A). The promoter activity of MIR172D was decreased in the spl15 mutant (C). The same confocal settings were used for scanning for each reporter line. Dash line marks the SAM. Scale bars represent 100 μm. (D) Expression of MIR172 genes in 18-day-old spl9 and spl15 mutants grown in LD. We harvested plants with the cotyledons and the first 5 rosette leaves manually removed. We could not get faithful data for MIR172C because its transcript level was very low. The expression level in WT is set to 1.0. Expression was normalized to TUB. Two technical replicates for each biological replicate (n = 2) were performed. Error bars represent SD. The data underlying this figure are included in S2 Data. LD, long days; SAM, shoot apical meristem; TUB, β-TUBULIN-2; WT, wild type.
(TIF)
(A) qRT-PCR analyses of MIR172 genes. Due to low expression levels of MIR172C and MIR172D in short days, their expression were not faithfully quantified. Two technical replicates for each biological replicate (n = 2) were performed. Error bars represent SD. See also Fig 7. (B) Quantification of MIR172 reporter genes. See Fig 6. The data underlying this figure are included in S2 Data.
(TIF)
Scale bars represent 50 μm.
(TIF)
Expression of MIR172A (A) and MIR172B (B) in leaf and shoot apices. Plants were grown at 16°C or 22°C in long days. The same confocal settings were used for scanning for each reporter line. Dash line marks the SAM. Scale bars represent 100 μm.
(TIF)
(A to D) The expression of MIR172 reporter genes before and after shift. Plants were grown in short days for 3 weeks (SD 3w) and shifted to long days (shift + n days). (E) The expression of MIR172 reporter genes after 4 weeks in short days (SD 4w). Please note that all the plants were still in the vegetative phase and the expression pattern of each reporter was the same as that of 3-week-old plants. The same confocal settings were used for scanning for each reporter line. Dash line marks the SAM. Scale bars represent 100 μm.
(TIF)
(A) The comparison of miR172 isoforms. The difference among miR172 isoforms resides in the last nucleotide (red). Two miR172-RT primers are designed: The miR172-RT-Primer A is fully complementary to miR172a, miR172b, and miR172e, whereas the miR172-RT-Prime C is fully complementary to miR172c and miR172d. These RT primers bind to the 3′ portion of miR172 molecules (underlined), initiating reverse transcription. (B) The expression of miR172 in WT and the mir172 mutants. We harvested 14-day-old plants grown in long days and set up 2 qRT-PCR experiments using miR172-RT-Primer A (left) or miR172-RT-Primer C (right) as the RT primer, respectively. Since MIR172D is highly expressed in the SAM, we could not detect its contribution to the mature miR172 pool. As such, the abundance of miR172 is markedly decreased in the mir172abe mutants. Both RT primers gave rise to similar results. However, the miR172-RT-Primer A gave higher reverse transcription efficiency than miR172-RT-Primer C. Notably, we could still detect miR172 in the mir172cd mutant using miR172-RT-Primer A as the RT primer, indicating that this primer does not discriminate miR172 isoforms. Expression was normalized to TUB. Two technical replicates for each biological replicate (n = 2) were performed. Error bars represent SD. The data underlying this figure are included in S2 Data. qRT-PCR, quantitative real-time PCR; SAM, shoot apical meristem; TUB, β-TUBULIN-2; WT, wild type.
(TIF)
(XLSX)
(XLSX)
(XLSX)
(XLSX)
From Figs 1D, 2B, 2D, 2F, 2H, 3B–3D, 4E–4J, 5B, 5D, 7A–7E.
(XLSX)
From Figs S2A–S2G, 5, 6, 8, 9D, 10A, 10B, 14B.
(XLSX)
Acknowledgments
We thank Ji-Qin Li, Zhi-Ping Zhang, and Xiao-Yan Gao (SIPPE, CAS) for skillful technical assistance; George Coupland (Max Planck Institute for Plant Breeding Research, Germany) and members in J.-W. Wang lab for discussion and comments on the manuscript.
Abbreviations
- AP1
APETALA1
- AP2
APETALA2
- ATAC-seq
transposase-accessible chromatin sequencing
- Col-0
Columbia-0
- CRISPR
clustered regularly interspaced short palindromic repeats
- FCA
FLOWERING CONTROL LOCUS A
- FLM
FLOWERING LOCUS M
- FT
FLOWERING LOCUS T
- FUL
FRUITFULL
- GFP
green fluorescent protein
- GI
GIGANTEA
- MIM
target mimicry
- miRNA
microRNA
- nt
nucleotide
- qRT-PCR
quantitative real-time PCR
- SAM
shoot apical meristem
- sgRNA
single guide RNA
- SMZ
SCHLAFMUTZE
- SNZ
SCHNARCHZAPFEN
- SOC1
SUPPRESSOR OF OVEREXPRESSION OF CO 1
- SPL
SQUAMOSA PROMOTER BINDING PROTEIN-LIKE
- STTM
short tandem target mimicry
- SVP
SHORT VEGETATIVE PHASE
- T-DNA
transfer-DNA
- TOE1
TARGET OF EAT1
- ts4
tasselseed4
- TUB
β-TUBULIN-2
- WT
wild type
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by the grants from National Natural Science Foundation of China (31788103; 31525004; 31721001; 31401026) and Strategic Priority Research Program of the Chinese Academy of Sciences (XDB27030101). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Rogers K, Chen X. Biogenesis, Turnover, and Mode of Action of Plant MicroRNAs. Plant Cell. 2013;25(7):2383–99. 10.1105/tpc.113.113159 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Wu G. Plant microRNAs and development. Journal of genetics and genomics = Yi chuan xue bao. 2013;40(5):217–30. Epub 2013/05/28. 10.1016/j.jgg.2013.04.002 . [DOI] [PubMed] [Google Scholar]
- 3.Xie M, Zhang S, Yu B. microRNA biogenesis, degradation and activity in plants. Cell Mol Life Sci. 2015;72(1):87–99. Epub 2014/09/12. 10.1007/s00018-014-1728-7 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Li S, Castillo-Gonzalez C, Yu B, Zhang X. The functions of plant small RNAs in development and in stress responses. Plant J. 2017;90(4):654–70. Epub 2016/12/13. 10.1111/tpj.13444 . [DOI] [PubMed] [Google Scholar]
- 5.Voinnet O. Origin, biogenesis, and activity of plant microRNAs. Cell. 2009;136(4):669–87. Epub 2009/02/26. 10.1016/j.cell.2009.01.046 . [DOI] [PubMed] [Google Scholar]
- 6.Allen E, Xie Z, Gustafson AM, Sung GH, Spatafora JW, Carrington JC. Evolution of microRNA genes by inverted duplication of target gene sequences in Arabidopsis thaliana. Nat Genet. 2004;36(12):1282–90. 10.1038/ng1478 . [DOI] [PubMed] [Google Scholar]
- 7.Baldrich P, Beric A, Meyers BC. Despacito: the slow evolutionary changes in plant microRNAs. Curr Opin Plant Biol. 2018;42:16–22. Epub 2018/02/16. 10.1016/j.pbi.2018.01.007 . [DOI] [PubMed] [Google Scholar]
- 8.Maher C, Stein L, Ware D. Evolution of Arabidopsis microRNA families through duplication events. Genome Res. 2006;16(4):510–9. Epub 2006/03/08. 10.1101/gr.4680506 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cui J, You C, Chen X. The evolution of microRNAs in plants. Curr Opin Plant Biol. 2016;35:61–7. 10.1016/j.pbi.2016.11.006 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fahlgren N, Howell MD, Kasschau KD, Chapman EJ, Sullivan CM, Cumbie JS, et al. High-throughput sequencing of Arabidopsis microRNAs: evidence for frequent birth and death of MIRNA genes. PLoS ONE. 2007;2(2):e219 Epub 2007/02/15. 10.1371/journal.pone.0000219 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sieber P, Wellmer F, Gheyselinck J, Riechmann JL, Meyerowitz EM. Redundancy and specialization among plant microRNAs: role of the MIR164 family in developmental robustness. Development. 2007;134(6):1051–60. 10.1242/dev.02817 . [DOI] [PubMed] [Google Scholar]
- 12.Allen RS, Li J, Stahle MI, Dubroue A, Gubler F, Millar AA. Genetic analysis reveals functional redundancy and the major target genes of the Arabidopsis miR159 family. Proc Natl Acad Sci U S A. 2007;104(41):16371–6. 10.1073/pnas.0707653104 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Franco-Zorrilla JM, Valli A, Todesco M, Mateos I, Puga MI, Rubio-Somoza I, et al. Target mimicry provides a new mechanism for regulation of microRNA activity. Nat Genet. 2007;39(8):1033–7. 10.1038/ng2079 . [DOI] [PubMed] [Google Scholar]
- 14.Todesco M, Rubio-Somoza I, Paz-Ares J, Weigel D. A collection of target mimics for comprehensive analysis of microRNA function in Arabidopsis thaliana. PLoS Genet. 2010;6(7):e1001031 10.1371/journal.pgen.1001031 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Zhang H, Zhang J, Yan J, Gou F, Mao Y, Tang G, et al. Short tandem target mimic rice lines uncover functions of miRNAs in regulating important agronomic traits. Proc Natl Acad Sci U S A. 2017;114(20):5277–82. Epub 2017/05/04. 10.1073/pnas.1703752114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Yan J, Gu Y, Jia X, Kang W, Pan S, Tang X, et al. Effective small RNA destruction by the expression of a short tandem target mimic in Arabidopsis. Plant Cell. 2012;24(2):415–27. 10.1105/tpc.111.094144 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Peng T, Qiao M, Liu H, Teotia S, Zhang Z, Zhao Y, et al. A Resource for Inactivation of MicroRNAs Using Short Tandem Target Mimic Technology in Model and Crop Plants. Mol Plant. 2018;11(11):1400–17. Epub 2018/09/24. 10.1016/j.molp.2018.09.003 . [DOI] [PubMed] [Google Scholar]
- 18.Yin K, Gao C, Qiu JL. Progress and prospects in plant genome editing. Nat Plants. 2017;3:17107 10.1038/nplants.2017.107 . [DOI] [PubMed] [Google Scholar]
- 19.Ran Y, Liang Z, Gao C. Current and future editing reagent delivery systems for plant genome editing. Sci China Life Sci. 2017;60(5):490–505. 10.1007/s11427-017-9022-1 . [DOI] [PubMed] [Google Scholar]
- 20.Manghwar H, Lindsey K, Zhang X, Jin S. CRISPR/Cas System: Recent Advances and Future Prospects for Genome Editing. Trends Plant Sci. 2019;24(12):1102–25. Epub 2019/11/16. 10.1016/j.tplants.2019.09.006 . [DOI] [PubMed] [Google Scholar]
- 21.Chen K, Wang Y, Zhang R, Zhang H, Gao C. CRISPR/Cas Genome Editing and Precision Plant Breeding in Agriculture. Annu Rev Plant Biol. 2019;70:667–97. Epub 2019/03/06. 10.1146/annurev-arplant-050718-100049 . [DOI] [PubMed] [Google Scholar]
- 22.Hua K, Zhang J, Botella JR, Ma C, Kong F, Liu B, et al. Perspectives on the Application of Genome-Editing Technologies in Crop Breeding. Mol Plant. 2019;12(8):1047–59. Epub 2019/07/02. 10.1016/j.molp.2019.06.009 . [DOI] [PubMed] [Google Scholar]
- 23.Yamaguchi YL, Ishida T, Yoshimura M, Imamura Y, Shimaoka C, Sawa S. A collection of mutants for CLE-peptide-encoding genes in Arabidopsis generated by CRISPR/Cas9 mediated gene targeting. Plant Cell Physiol. 2017;58(11):1848–56. 10.1093/pcp/pcx139 . [DOI] [PubMed] [Google Scholar]
- 24.Park W, Li J, Song R, Messing J, Chen X. CARPEL FACTORY, a Dicer homolog, and HEN1, a novel protein, act in microRNA metabolism in Arabidopsis thaliana. Curr Biol. 2002;12(17):1484–95. 10.1016/s0960-9822(02)01017-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Rhoades MW, Reinhart BJ, Lim LP, Burge CB, Bartel B, Bartel DP. Prediction of plant microRNA targets. Cell. 2002;110(4):513–20. 10.1016/s0092-8674(02)00863-2 . [DOI] [PubMed] [Google Scholar]
- 26.Spanudakis E, Jackson S. The role of microRNAs in the control of flowering time. J Exp Bot. 2014;65(2):365–80. 10.1093/jxb/ert453 . [DOI] [PubMed] [Google Scholar]
- 27.Teotia S, Tang G. To bloom or not to bloom: role of microRNAs in plant flowering. Mol Plant. 2015;8(3):359–77. Epub 2015/03/05. 10.1016/j.molp.2014.12.018 . [DOI] [PubMed] [Google Scholar]
- 28.Wang JW. Regulation of flowering time by the miR156-mediated age pathway. J Exp Bot. 2014;65(17):4723–30. 10.1093/jxb/eru246 . [DOI] [PubMed] [Google Scholar]
- 29.Zhou CM, Wang JW. Regulation of flowering time by microRNAs. Journal of genetics and genomics = Yi chuan xue bao. 2013;40(5):211–5. 10.1016/j.jgg.2012.12.003 . [DOI] [PubMed] [Google Scholar]
- 30.Wang JW, Czech B, Weigel D. miR156-regulated SPL transcription factors define an endogenous flowering pathway in Arabidopsis thaliana. Cell. 2009;138(4):738–49. 10.1016/j.cell.2009.06.014 . [DOI] [PubMed] [Google Scholar]
- 31.Wu G, Poethig RS. Temporal regulation of shoot development in Arabidopsis thaliana by miR156 and its target SPL3. Development. 2006;133(18):3539–47. 10.1242/dev.02521 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Aukerman MJ, Sakai H. Regulation of flowering time and floral organ identity by a MicroRNA and its APETALA2-like target genes. Plant Cell. 2003;15(11):2730–41. 10.1105/tpc.016238 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wu G, Park MY, Conway SR, Wang JW, Weigel D, Poethig RS. The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis. Cell. 2009;138(4):750–9. 10.1016/j.cell.2009.06.031 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Huijser P, Schmid M. The control of developmental phase transitions in plants. Development. 2011;138(19):4117–29. 10.1242/dev.063511 . [DOI] [PubMed] [Google Scholar]
- 35.Zhu QH, Helliwell CA. Regulation of flowering time and floral patterning by miR172. J Exp Bot. 2011;62(2):487–95. 10.1093/jxb/erq295 . [DOI] [PubMed] [Google Scholar]
- 36.Yu S, Galvao VC, Zhang YC, Horrer D, Zhang TQ, Hao YH, et al. Gibberellin regulates the Arabidopsis floral transition through miR156-targeted SQUAMOSA PROMOTER BINDING-LIKE transcription factors. Plant Cell. 2012;24(8):3320–32. 10.1105/tpc.112.101014 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Jung JH, Seo PJ, Kang SK, Park CM. miR172 signals are incorporated into the miR156 signaling pathway at the SPL3/4/5 genes in Arabidopsis developmental transitions. Plant Mol Biol. 2011;76(1–2):35–45. 10.1007/s11103-011-9759-z [DOI] [PubMed] [Google Scholar]
- 38.Huo H, Wei S, Bradford KJ. DELAY OF GERMINATION1 (DOG1) regulates both seed dormancy and flowering time through microRNA pathways. Proc Natl Acad Sci U S A. 2016. 10.1073/pnas.1600558113 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Mathieu J, Yant LJ, Murdter F, Kuttner F, Schmid M. Repression of flowering by the miR172 target SMZ. PLoS Biol. 2009;7(7):e1000148 10.1371/journal.pbio.1000148 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Yant L, Mathieu J, Dinh TT, Ott F, Lanz C, Wollmann H, et al. Orchestration of the floral transition and floral development in Arabidopsis by the bifunctional transcription factor APETALA2. Plant Cell. 2010;22(7):2156–70. 10.1105/tpc.110.075606 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Chen X. A microRNA as a translational repressor of APETALA2 in Arabidopsis flower development. Science. 2004;303(5666):2022–5. 10.1126/science.1088060 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Wollmann H, Mica E, Todesco M, Long JA, Weigel D. On reconciling the interactions between APETALA2, miR172 and AGAMOUS with the ABC model of flower development. Development. 2010;137(21):3633–42. 10.1242/dev.036673 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Chavez Montes RA, de Fatima Rosas-Cardenas F, De Paoli E, Accerbi M, Rymarquis LA, Mahalingam G, et al. Sample sequencing of vascular plants demonstrates widespread conservation and divergence of microRNAs. Nat Commun. 2014;5:3722 Epub 2014/04/25. 10.1038/ncomms4722 . [DOI] [PubMed] [Google Scholar]
- 44.Cuperus JT, Fahlgren N, Carrington JC. Evolution and functional diversification of MIRNA genes. Plant Cell. 2011;23(2):431–42. Epub 2011/02/15. 10.1105/tpc.110.082784 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Axtell MJ, Bowman JL. Evolution of plant microRNAs and their targets. Trends Plant Sci. 2008;13(7):343–9. 10.1016/j.tplants.2008.03.009 . [DOI] [PubMed] [Google Scholar]
- 46.Jones-Rhoades MW. Conservation and divergence in plant microRNAs. Plant Mol Biol. 2012;80(1):3–16. Epub 2011/10/15. 10.1007/s11103-011-9829-2 . [DOI] [PubMed] [Google Scholar]
- 47.You C, Cui J, Wang H, Qi X, Kuo LY, Ma H, et al. Conservation and divergence of small RNA pathways and microRNAs in land plants. Genome Biol. 2017;18(1):158 Epub 2017/08/25. 10.1186/s13059-017-1291-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Reinhart BJ, Weinstein EG, Rhoades MW, Bartel B, Bartel DP. MicroRNAs in plants. Genes Dev. 2002;16(13):1616–26. 10.1101/gad.1004402 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wang ZP, Xing HL, Dong L, Zhang HY, Han CY, Wang XC, et al. Egg cell-specific promoter-controlled CRISPR/Cas9 efficiently generates homozygous mutants for multiple target genes in Arabidopsis in a single generation. Genome Biol. 2015;16:144 10.1186/s13059-015-0715-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Schwab R, Palatnik JF, Riester M, Schommer C, Schmid M, Weigel D. Specific effects of microRNAs on the plant transcriptome. Dev Cell. 2005;8(4):517–27. 10.1016/j.devcel.2005.01.018 . [DOI] [PubMed] [Google Scholar]
- 51.Balanza V, Martinez-Fernandez I, Sato S, Yanofsky MF, Kaufmann K, Angenent GC, et al. Genetic control of meristem arrest and life span in Arabidopsis by a FRUITFULL-APETALA2 pathway. Nat Commun. 2018;9(1):565 10.1038/s41467-018-03067-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Jung JH, Lee S, Yun J, Lee M, Park CM. The miR172 target TOE3 represses AGAMOUS expression during Arabidopsis floral patterning. Plant Sci. 2014;215–216C:29–38. 10.1016/j.plantsci.2014.09.010 . [DOI] [PubMed] [Google Scholar]
- 53.Ji L, Liu X, Yan J, Wang W, Yumul RE, Kim YJ, et al. ARGONAUTE10 and ARGONAUTE1 regulate the termination of floral stem cells through two microRNAs in Arabidopsis. PLoS Genet. 2011;7(3):e1001358 10.1371/journal.pgen.1001358 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Zhao L, Kim Y, Dinh TT, Chen X. miR172 regulates stem cell fate and defines the inner boundary of APETALA3 and PISTILLATA expression domain in Arabidopsis floral meristems. Plant J. 2007;51(5):840–9. 10.1111/j.1365-313X.2007.03181.x . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Andres F, Coupland G. The genetic basis of flowering responses to seasonal cues. Nat Rev Genet. 2012;13(9):627–39. 10.1038/nrg3291 . [DOI] [PubMed] [Google Scholar]
- 56.Wang FX, Shang GD, Wu LY, Xu ZG, Zhao XY, Wang JW. Chromatin Accessibility Dynamics and a Hierarchical Transcriptional Regulatory Network Structure for Plant Somatic Embryogenesis. Dev Cell. 2020;54: 10.1016/j.devcel.2020.07.003 [DOI] [PubMed] [Google Scholar]
- 57.Yu S, Lian H, Wang JW. Plant developmental transitions: the role of microRNAs and sugars. Curr Opin Plant Biol. 2015;27:1–7. 10.1016/j.pbi.2015.05.009 . [DOI] [PubMed] [Google Scholar]
- 58.Poethig RS. Vegetative phase change and shoot maturation in plants. Curr Top Dev Biol. 2013;105:125–52. 10.1016/B978-0-12-396968-2.00005-1 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Poethig RS. The past, present, and future of vegetative phase change. Plant Physiol. 2010;154(2):541–4. 10.1104/pp.110.161620 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Poethig RS. Small RNAs and developmental timing in plants. Curr Opin Genet Dev. 2009;19(4):374–8. 10.1016/j.gde.2009.06.001 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Hyun Y, Richter R, Vincent C, Martinez-Gallegos R, Porri A, Coupland G. Multi-layered Regulation of SPL15 and Cooperation with SOC1 Integrate Endogenous Flowering Pathways at the Arabidopsis Shoot Meristem. Dev Cell. 2016;37(3):254–66. 10.1016/j.devcel.2016.04.001 . [DOI] [PubMed] [Google Scholar]
- 62.Wang JW, Schwab R, Czech B, Mica E, Weigel D. Dual effects of miR156-targeted SPL genes and CYP78A5/KLUH on plastochron length and organ size in Arabidopsis thaliana. Plant Cell. 2008;20(5):1231–43. 10.1105/tpc.108.058180 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ye BB, Shang GD, Pan Y, Xu ZG, Zhou CM, Mao YB, et al. AP2/ERF Transcription Factors Integrate Age and Wound Signals for Root Regeneration. Plant Cell. 2020;32(1):226–41. Epub 2019/10/28. 10.1105/tpc.19.00378 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Hyun Y, Richter R, Coupland G. Competence to Flower: Age-Controlled Sensitivity to Environmental Cues. Plant Physiol. 2017;173(1):36–46. 10.1104/pp.16.01523 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Lee H, Yoo SJ, Lee JH, Kim W, Yoo SK, Fitzgerald H, et al. Genetic framework for flowering-time regulation by ambient temperature-responsive miRNAs in Arabidopsis. Nucleic Acids Res. 2010;38(9):3081–93. 10.1093/nar/gkp1240 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Jung JH, Seo PJ, Ahn JH, Park CM. Arabidopsis RNA-binding Protein FCA Regulates MicroRNA172 Processing in Thermosensory Flowering. J Biol Chem. 2012;287(19):16007–16. 10.1074/jbc.M111.337485 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Jung JH, Seo YH, Seo PJ, Reyes JL, Yun J, Chua NH, et al. The GIGANTEA-regulated microRNA172 mediates photoperiodic flowering independent of CONSTANS in Arabidopsis. Plant Cell. 2007;19(9):2736–48. 10.1105/tpc.107.054528 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Baker CC, Sieber P, Wellmer F, Meyerowitz EM. The early extra petals1 mutant uncovers a role for microRNA miR164c in regulating petal number in Arabidopsis. Curr Biol. 2005;15(4):303–15. 10.1016/j.cub.2005.02.017 . [DOI] [PubMed] [Google Scholar]
- 69.Hyun Y, Vincent C, Tilmes V, Bergonzi S, Kiefer C, Richter R, et al. A regulatory circuit conferring varied flowering response to cold in annual and perennial plants. Science. 2019;363(6425):409–12. Epub 2019/01/27. 10.1126/science.aau8197 . [DOI] [PubMed] [Google Scholar]
- 70.Abe M, Kobayashi Y, Yamamoto S, Daimon Y, Yamaguchi A, Ikeda Y, et al. FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science. 2005;309(5737):1052–6. 10.1126/science.1115983 . [DOI] [PubMed] [Google Scholar]
- 71.Wigge PA, Kim MC, Jaeger KE, Busch W, Schmid M, Lohmann JU, et al. Integration of spatial and temporal information during floral induction in Arabidopsis. Science. 2005;309(5737):1056–9. 10.1126/science.1114358 [DOI] [PubMed] [Google Scholar]
- 72.Cho HJ, Kim JJ, Lee JH, Kim W, Jung JH, Park CM, et al. SHORT VEGETATIVE PHASE (SVP) protein negatively regulates miR172 transcription via direct binding to the pri-miR172a promoter in Arabidopsis. FEBS Lett. 2012;586(16):2332–7. 10.1016/j.febslet.2012.05.035 . [DOI] [PubMed] [Google Scholar]
- 73.Susila H, Nasim Z, Ahn JH. Ambient Temperature-Responsive Mechanisms Coordinate Regulation of Flowering Time. Int J Mol Sci. 2018;19(10). Epub 2018/10/20. 10.3390/ijms19103196 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Capovilla G, Schmid M, Pose D. Control of flowering by ambient temperature. J Exp Bot. 2014. 10.1093/jxb/eru416 . [DOI] [PubMed] [Google Scholar]
- 75.Wigge PA. Ambient temperature signalling in plants. Curr Opin Plant Biol. 2013;16(5):661–6. Epub 2013/09/12. 10.1016/j.pbi.2013.08.004 . [DOI] [PubMed] [Google Scholar]
- 76.Song YH, Ito S, Imaizumi T. Flowering time regulation: photoperiod- and temperature-sensing in leaves. Trends Plant Sci. 2013;18(10):575–83. 10.1016/j.tplants.2013.05.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Capovilla G, Symeonidi E, Wu R, Schmid M. Contribution of major FLM isoforms to temperature-dependent flowering in Arabidopsis thaliana. J Exp Bot. 2017. 10.1093/jxb/erx328 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Sureshkumar S, Dent C, Seleznev A, Tasset C, Balasubramanian S. Nonsense-mediated mRNA decay modulates FLM-dependent thermosensory flowering response in Arabidopsis. Nat Plants. 2016;2(5):16055 10.1038/nplants.2016.55 [DOI] [PubMed] [Google Scholar]
- 79.Lutz U, Nussbaumer T, Spannagl M, Diener J, Mayer KF, Schwechheimer C. Natural haplotypes of FLM non-coding sequences fine-tune flowering time in ambient spring temperatures in Arabidopsis. Elife. 2017;6 10.7554/eLife.22114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Lee JH, Ryu HS, Chung KS, Pose D, Kim S, Schmid M, et al. Regulation of temperature-responsive flowering by MADS-box transcription factor repressors. Science. 2013;342(6158):628–32. 10.1126/science.1241097 . [DOI] [PubMed] [Google Scholar]
- 81.Lee DY, Lee J, Moon S, Park SY, An G. The rice heterochronic gene SUPERNUMERARY BRACT regulates the transition from spikelet meristem to floral meristem. Plant J. 2007;49(1):64–78. Epub 2006/12/06. 10.1111/j.1365-313X.2006.02941.x . [DOI] [PubMed] [Google Scholar]
- 82.Zhu QH, Upadhyaya NM, Gubler F, Helliwell CA. Over-expression of miR172 causes loss of spikelet determinacy and floral organ abnormalities in rice (Oryza sativa). BMC Plant Biol. 2009;9:149 10.1186/1471-2229-9-149 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Chuck G, Meeley R, Hake S. Floral meristem initiation and meristem cell fate are regulated by the maize AP2 genes ids1 and sid1. Development. 2008;135(18):3013–9. Epub 2008/08/15. 10.1242/dev.024273 . [DOI] [PubMed] [Google Scholar]
- 84.Yin ZJ, Shen FF. Identification and characterization of conserved microRNAs and their target genes in wheat (Triticum aestivum). Genetics and molecular research: GMR. 2010;9(2):1186–96. Epub 2010/07/01. 10.4238/vol9-2gmr805 . [DOI] [PubMed] [Google Scholar]
- 85.Debernardi JM, Lin H, Chuck G, Faris JD, Dubcovsky J. microRNA172 plays a crucial role in wheat spike morphogenesis and grain threshability. Development. 2017;144(11):1966–75. 10.1242/dev.146399 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Debernardi JM, Greenwood JR, Jean Finnegan E, Jernstedt J, Dubcovsky J. APETALA 2-like genes AP2L2 and Q specify lemma identity and axillary floral meristem development in wheat. Plant J. 2020;101(1):171–87. Epub 2019/09/09. 10.1111/tpj.14528 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Chuck G, Meeley R, Irish E, Sakai H, Hake S. The maize tasselseed4 microRNA controls sex determination and meristem cell fate by targeting Tasselseed6/indeterminate spikelet1. Nat Genet. 2007;39(12):1517–21. Epub 2007/11/21. 10.1038/ng.2007.20 . [DOI] [PubMed] [Google Scholar]
- 88.Clough SJ, Bent AF. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998;16(6):735–43. 10.1046/j.1365-313x.1998.00343.x . [DOI] [PubMed] [Google Scholar]
- 89.Liu H, Ding Y, Zhou Y, Jin W, Xie K, Chen LL. CRISPR-P 2.0: An Improved CRISPR-Cas9 Tool for Genome Editing in Plants. Mol Plant. 2017;10(3):530–2. 10.1016/j.molp.2017.01.003 . [DOI] [PubMed] [Google Scholar]
- 90.Lian H, Li X, Liu Z, He Y. HYL1 is required for establishment of stamen architecture with four microsporangia in Arabidopsis. J Exp Bot. 2013;64(11):3397–410. 10.1093/jxb/ert178 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Li X, Lian H, Zhao Q, He Y. MicroRNA166 Monitors SPOROCYTELESS/NOZZLE for Building of the Anther Internal Boundary. Plant Physiol. 2019;181(1):208–20. Epub 2019/06/30. 10.1104/pp.19.00336 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Yu S, Cao L, Zhou CM, Zhang TQ, Lian H, Sun Y, et al. Sugar is an endogenous cue for juvenile-to-adult phase transition in plants. Elife. 2013;2:e00269 10.7554/eLife.00269 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Varkonyi-Gasic E, Wu R, Wood M, Walton EF, Hellens RP. Protocol: a highly sensitive RT-PCR method for detection and quantification of microRNAs. Plant Methods. 2007;3:12 10.1186/1746-4811-3-12 [DOI] [PMC free article] [PubMed] [Google Scholar]