

Abstract
Sepsis is a severe disease characterized by an uncontrolled systemic inflammation and consequent organ dysfunction generated in response to an infection. Extracellular ATP acting through the P2X7 receptor induces the maturation and release of pro-inflammatory cytokines (i.e., IL-1β) and the production of reactive nitrogen and oxygen species that lead to oxidative tissue damage. Here, we investigated the role of the P2X7 receptor in inflammation, oxidative stress, and liver injury in sepsis. Sepsis was induced by cecal ligation and puncture (CLP) in wild-type (WT) and P2X7 knockout (P2X7−/−) mice. The oxidative stress in the liver of septic mice was assessed by 2′,7′-dichlorofluorescein oxidation reaction (DCF), thiobarbituric acid-reactive substances (TBARS), and nitrite levels dosage. The status of the endogenous defense system was evaluated through catalase (CAT) and superoxide dismutase (SOD) activities. The inflammation was assessed histologically and by determining the expression of inflammatory cytokines and chemokines by RT-qPCR. We observed an increase in the reactive species and lipid peroxidation in the liver of septic WT mice, but not in the liver from P2X7−/− animals. We found an imbalance SOD/CAT ratio, also only WT septic animals. The number of inflammatory cells and the gene expression of IL-1 β, IL-6, TNF-α, IL-10, CXCL1, and CXCL2 were higher in the liver of WT septic mice in comparison to P2X7−/− septic animals. In summary, our results suggest that the P2X7 receptor might be a therapeutic target to limit oxidative stress damage and liver injury during sepsis.
Keywords: Sepsis, Purinergic signaling, ATP, Liver, Oxidative stress
Introduction
Sepsis is a life-threatening organ dysfunction characterized by an uncontrolled and excessive inflammatory response to an infection [1, 2]. Although considerable progress has been made in medical assistance over the last decades, sepsis remains a healthcare challenge worldwide [3, 4]. The inflammatory response and organ injury in sepsis initiate when pathogen-associated molecular patterns (PAMPs) bind to pattern recognition receptors (PRR), such as toll-like receptors (TLR), on the surface of immune cells [5, 6]. PRRs trigger activation of inflammatory signaling pathways and the production of inflammatory mediators, including cytokines, chemokines, and oxygen and nitrogen reactive species [7–9]. When the inflammation becomes systemic and excessive, the overproduction of inflammatory mediators affects the functionality of several vital organs, including the liver [10–12].
The liver is the largest gland in the human body. This organ mediates several essential metabolic processes and modulates immunological responses [12–14]. The strategic position of the liver in the bloodstream permanently exposes the organ to several antigens and microbial products with a potential immunostimulatory capacity [14, 15]. Therefore, the liver plays a central role in the development of systemic inflammation in the initial course of sepsis by sensing danger signals; recognizing pathogens; producing cytokines, chemokines, and acute-phase proteins; and regulating metabolism. Therefore, liver protection is critical for host survival in sepsis [12, 16–18].
Extracellular adenosine triphosphate (eATP) plasma levels increase during sepsis [19]. This nucleotide is an important danger signal that regulates the recruitment and function of inflammatory cells via purinergic receptors [20, 21]. The P2 purinergic family is composed of G protein-coupled P2Y and ionotropic P2X receptors [22]. Among the P2 receptor, the P2X7 receptor has been the most associated with the activation of oxidative and inflammatory mechanisms in several inflammatory and infectious diseases [23–28]. The P2X7 receptor is a critical player in the activation of the NLRP3 inflammasome, acting as a second signal for caspase-1 activation and subsequent release of IL1-β [29, 30]. Besides, this receptor mediates the production of ROS and NO in inflammatory cells [31, 32]. In sepsis, P2X7 genetic deletion improves survival and reduces circulating inflammatory cytokines and systemic inflammation [23, 33]. Moreover, P2X7 receptor pharmacological inhibition reduces sepsis-induced intestinal barrier disruption [27] and brain inflammation and oxidative damage [24, 34]. P2X7 receptor induces inflammation and oxidative damage in a model of carbon tetrachloride-induced steatohepatitis in obese mice by NADPH oxidase-dependent mechanisms [35]. Therefore, here we sought to investigate the role of ATP-gated P2X7 receptor in sepsis-induced inflammation, oxidative damage, and liver injury in sepsis.
Materials and methods
Animals
We used adult male (8–10 weeks old; 25–30 g weight) wild-type (WT) and P2X7 receptor-deficient (P2X7−/−) C57BL/6 mice originally obtained from Jackson Laboratories (Bar Harbor, ME, USA). The animals had free access to food and water and were maintained on a 12:12-h light-dark cycle at a temperature of 22 ± 1 °C. Experiments were performed with age- and weight-matched animals, according to the guidelines of the Brazilian College of Animal Experimentation (COBEA). The Commission for the Ethical Use of Research Animals (CEUA) from the Federal University of Rio de Janeiro (UFRJ) approved all experiments (protocol number: IBCCF138).
Sepsis induction by cecal ligation and puncture
Sepsis was induced by cecal ligation and puncture (CLP), a procedure that generates acute polymicrobial peritonitis and, subsequently, a systemic inflammation, as previously described [36, 37]. Mice were anesthetized by the injection of ketamine (80 mg/kg) and xylazine (5 mg/kg) intraperitoneally, and a 1-cm incision was made on the abdomen. The cecum was exposed and ligated below the ileocecal junction. A double puncture was made using a 21-gauge needle, a small amount of stool was gently squeezed out to induce peritonitis, and the peritoneal cavity and abdominal wall were closed by suture. Sham-operated animals (controls) underwent an identical laparotomy but without ligation and perforation. Animals received 1 mL of 0.9% isotonic NaCl sterile solution subcutaneously to compensate for the third spacing that occurred during the surgery. The liver samples were collected 24 h after the procedure.
Alanine aminotransferase and aspartate aminotransferase dosage
For the determination of ALT and AST, kinetic kits commercially available (Bioclin®, Belo Horizonte, MG, Brazil; cat#K049-6 and K048-6, respectively) were used, and the experiments were carried out according to the manufacturer’s instructions. Briefly, 24 h after sepsis induction, the animals were euthanized in a chamber with an atmosphere rich in CO2, and the blood was collected through cardiac puncture using a 1-mL syringe/27-G needle with sodium citrate (5%). Plasma was obtained through centrifugation (2000×g for 10 min) and used for enzymatic analysis. The enzyme reaction was analyzed on a SpectraMax2 spectrophotometer (Thermo Fisher Scientific, Waltham, Massachusetts). The results are expressed in units per liter (U/L).
Oxidative profile
Tissue preparation for oxidative analysis
The liver tissue was homogenized in 10 volumes (1:10 w/v) of 20 mM sodium phosphate buffer (pH 7.4) containing 140 mM KCl and centrifuged at 800×g for 10 min at 4 °C. The supernatant was used immediately for the determination of oxidative parameters.
2′,7′-Dichlorofluorescein (H2DCF) oxidation assay
The production of reactive species was measured according to the methodology based on the oxidation of the 2′,7′-dichlorofluorescein (H2DCF) proposed by LeBel et al. [38]. The samples were incubated in the dark in a 2′,7′-dichlorofluorescein diacetate solution (H2DCF-DA) for 30 min at 37 °C. H2DCF-DA is cleaved by cellular esterases, and the resulting H2DCF is eventually oxidized by the reactive species present in the samples. This last reaction produces a fluorescent product: dichlorofluorescein (DCF), which was measured at 488-nm excitation and 525-nm emission. Results are presented in nmol DCF per milligram protein [38].
Dosage of thiobarbituric acid-reactive substances
As previously as described by Ohkawa et al. [39], tissue supernatant (200 μL) was mixed with 50 μL 8.1% SDS; 375 μL of 20% acetic acid in aqueous solution (v/v) pH 3.5; and 375 μL of 0.8% thiobarbituric acid and mixed in the vortex. The reaction is produced in a boiling water bath for 1 h. The mixture was cooled in water for 5 min and centrifuged 750×g for 10 min. TBARS levels were determined by spectrophotometry at 535 nm. A calibration curve with the standard 1,1,3,3-tetrametoxypropane was used. TBARS levels are expressed as TBARS nanomole per milligram protein.
Determination of total sulfhydryl content
The oxidation of thiols in samples leads to the formation of disulfide bonds. Sulfhydryl content is inversely proportional to the oxidative damage in proteins. In this method, 5,5′-dithiobis (2-nitrobenzoic) acid (DTNB) undergoes a chemical modification in the presence of sulfhydryl groups, generating 5-thio-2-nitrobenzoate (TNB). We mixed 50 μL of homogenate with 980 μL of PBS buffer pH 7.4 containing 1 mM EDTA. The reaction was started by the addition of 30 μL of 10 mM DTNB and incubated for 30 min at room temperature in a dark room. The absorbance of this yellow product (TNB) was measured spectrophotometrically at 412 nm. The results were presented as TNB per milligram protein [40].
Nitric oxide production
Nitrite, which is a metabolite of nitric oxide (NO), was evaluated using Griess reagent (1:1 mixture of 1% sulfanilamide in 5% phosphoric acid and 0.1% N-(1-Naphthyl)ethylenediamine dihydrochloride in water. The samples were incubated 10 min at room temperature in a Griess reagent. Nitrite was used as the standard, and absorbance was measured at 543 nm. The results were expressed as micromole of nitrite per milligram of protein [41].
Catalase assay
CAT assay is based on the consumption of H2O2, which is measured by a spectrophotometer at 240 nm in a reaction medium containing 20 mM H2O2, 0.1% Triton X-100, 10 mM phosphate buffer solution pH 7.0, and an amount of protein from 0.1 to 0.3 mg per mL of reaction medium [42]. A unit of CAT is defined as 1 mmol of H2O2 consumed per min, and the specific activity was represented as units of CAT/mg of protein.
Superoxide dismutase assay
SOD activity is based on the ability of the pyrogallol compound to self-oxidize in the presence of superoxide, which is also a substrate for the SOD enzyme. The inhibition of the auto-oxidation of this compound occurs in the presence of SOD, whose activity was measured indirectly by spectrophotometry at 420 nm [43]. A calibration curve was made with purified SOD as a standard to calculate SOD activity in the samples. The results were presented as units of SOD per milligram of protein.
Protein quantification
The determination of the total amount of proteins was made using a spectrophotometric method using the Pierce BCA Kit (Thermo Fisher Scientific) and the SpectraMax2 plate reader (Thermo Fisher Scientific) under the manufacturer’s instructions. This method combines the reduction of copper (II) to copper (I) cations by proteins in an alkaline medium (the biuret method) with the selective and sensitive colorimetric detection of copper (I) cations. The purple reaction product exhibits an absorbance at 562 nm, whose increase is practically linear with the increase in protein concentration. Protein concentrations were reported on a calibration curve using bovine serum albumin (BSA) absorbance values as standard. The expression of the total protein concentration in the samples is shown in microgram per milliliter.
RNA isolation and real-time quantitative PCR
For the extraction of total RNA from the liver tissue, the TRIzol® reagent (Thermo Fisher Scientific, Somerset, NJ, USA) was used according to the manufacturer’s instructions. RNA samples were quantified, and the purity was assessed in a NanoDrop BioChrom apparatus (Harvard Bioscience, Holliston, Massachusetts, USA). The synthesis of cDNA was performed with 1 μg of total RNA using the High Capacity Reverse Transcription Kit with RNase Inhibitor (Thermo Fisher Scientific, Somerset, NJ, USA) according to the manufacturer’s instructions in a Master Cycler Gradient thermocycler (Eppendorf, Hamburg, Germany).
The real-time quantitative PCR reactions (RT-qPCR) were performed using the Master Mix SYBR Green PCR (Applied Biosystems, Foster City, California, USA) in a StepOnePlus real-time PCR system (Applied Biosystems, Foster City, California, USA). The reactions were performed in a final volume of 10 μL, using 2 μL of diluted cDNA (1:10) and 300 nM of each of the primer forward and reverse. The relative gene expression was calculated using the comparative cycle threshold (Ct) method (ΔΔCt). The β-actin gene (Actb) was used as the endogenous control. The sham WT group was chosen as the control group. The primer sequences are provided in Table 1. The results are presented as relative expression of the gene of interest/Actb.
Table 1.
PCR primer sequences
| Gene | Sequence (5′–3′) | |
|---|---|---|
| P2rX7 | Sense | AAT CGG TGT GTT TCC TTT GG |
| P2rX7 | Antisense | CCG GGT GAC TTT GTT TGT CT |
| Nox1 | Sense | TAA ATT GCC CCT CCA TTT CC |
| Nox1 | Antisense | CCC TGC TGC TCG AAT ATG AA |
| Nox2 | Sense | CCC TTT GGT ACA GCC AGT GAA GAT |
| Nox2 | Antisense | CAA TCC CGG CTC CCA CTA ACA TCA |
| Nos2 | Sense | ACA TCG ACC CGT CCA CAG TAT |
| Nos2 | Antisense | CAG AGG GGT AGG CTT GTC TC |
| Il1β | Sense | TTC AGG CAG GCA GTA TCA CTC |
| Il1β | Antisense | CCA CGG GAA AGA CAC AGG TAG |
| Il6 | Sense | CTG CAA GAG ACT TCC ATC CAG |
| Il6 | Antisense | AGT GGT ATA GAC AGG TCT GTT GG |
| Tnf | Sense | GGT CCC CAA AGG GAT GAG AAG TTC |
| Tnf | Antisense | CCA CTT GGT GGT TTG CTA CGA CG |
| Il10 | Sense | GCT GGA CAA CAT ACT GCT AAC C |
| Il10 | Antisense | ATT TCC GAT AAG GCT TGG CAA |
| Cxcl1 | Sense | ACT GCA CCC AAA CCG AAG TC |
| Cxcl1 | Antisense | TGG GGA CAC CTT TTA GCA TCT T |
| Cxcl2 | Sense | CCA ACC ACC AGG CTA CAG G |
| Cxcl2 | Antisense | GCG TCA CAC TCA AGC TCT G |
| Actb | Sense | TAT GCC AAC ACA GTG CTG TCT GG |
| Actb | Antisense | ACT CCT GCT TGC TGA TCC ACA T |
Histological analysis
The liver was harvested 24 h after induced sepsis and fixed with 4% paraformaldehyde, embedded in paraffin, and 5-μm-thick slices were stained with hematoxylin-eosin. A pathologist, who was blinded to the experimental groups, analyzed the histopathology and performed the semiquantitative analyses.
A histological scoring system was applied to evaluate the degree of liver injury associated with sepsis after observation of all liver sections of each animal. The histological parameters steatosis, spotty (focal) necrosis, and ballooning degeneration were considered. The hepatic injury scoring measures features of liver injury induced by sepsis and is represented as the unweighted sum of scores of steatosis (0–3), hepatocellular ballooning or degeneration (0–2), and spotty (focal) necrosis (0–2) with the final score ranging between 0 and 7. This analysis was adapted from the histological scoring system for the nonalcoholic fatty liver disease [44].
The degree of steatosis (steatosis identified as homogeneous lipid droplet in the hepatocyte cytoplasm present as microvesicular or macrovesicular) was graded and scored as follows: 0, none; 1, ≤ 30% hepatocytes containing fat; 2, 30%–70% hepatocytes containing fat; 3, ≥ 70% hepatocytes containing fat. Ballooning degeneration was graded and scored as follows: 0, normal hepatocytes with cuboidal shape and pink eosinophilic cytoplasm; 1, presence of clusters of hepatocytes with a rounded shape and pale, reticulated cytoplasm and the same size of normal ones; 2, clusters of rounded, enlarged hepatocytes. The grade of spotty necrosis considered was as follows: 0, lack of focal necrosis; 1, one focus viewed with a 10× objective lens; 2, more than two foci viewed with a 10× objective lens.
Lobular inflammation was evaluated in histological sections of the liver stained with HE and represented the presence of polynucleated cells (neutrophils) or mononucleate cells (macrophages/monocytes or lymphocytes) in the sinusoidal lumen. Fifteen photomicrographs were obtained from each animal using a light microscope Eclipse E800 (Nikon, Japan) assisted with a digital camera Evolution VR Cooled Color 13 bits (Media Cybernetics, Bethesda, USA). The software Q-Capture 2.95.0, version 2.0.5 (Silicon Graphics Inc., EUA), was utilized to obtain high-resolution images (2048 X1536 pixels buffer) using the 20× objective lens. Results were expressed as the total number of inflammatory cells/histological field.
Statistical analysis
The Kolmogorov-Smirnov statistical test was used to verify whether the data obtained in this work has a normal distribution. The results were analyzed through a two-way analysis of variance (two-way ANOVA) followed by Tukey’s multiple tests, considering p < 0.05 as a statistical difference. The GraphPad Prism Version 5 program (GraphPad Software Incorporation) was used for data analysis.
Results
P2X7 receptor expression increases in the liver of septic mice
ATP-P2X7 receptor signaling seems to be relevant for the excessive inflammation in sepsis. Initially, we found that P2X7 receptor expression significantly increases in the liver of WT septic animals 24 h after sepsis induction (Fig. 1; p < 0.05), suggesting the participation of this receptor in hepatic dysfunction in sepsis.
Fig. 1.
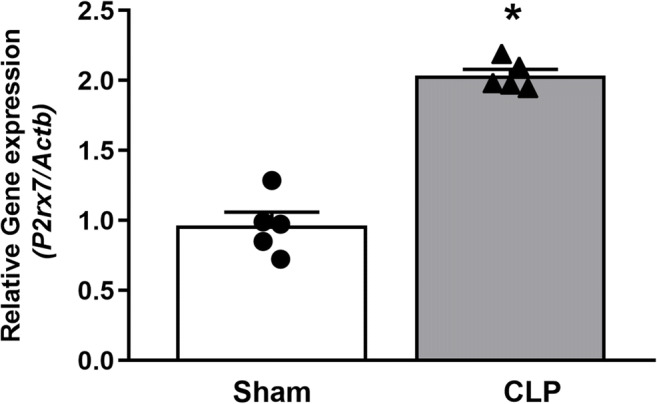
P2X7 receptor gene expression in the liver of septic mice. P2X7 gene expression was analyzed by RT-qPCR in the liver from septic mice 24 h after surgery. Data are expressed as mean ± S.E.M. (n = 5 for both sham and CLP groups). Data were analyzed by t test. Statistically significant differences between sham and CLP groups are represented by asterisks (*p < 0.05)
P2X7 receptor genetic deletion attenuates liver injury in septic mice
Hepatocyte lesions were determined by quantifying the activity of liver enzymes (ALT and AST) in the mice serum 24 h after sepsis induction. The presence of these enzymes in the circulation provides evidence of the extent and magnitude of the hepatic lesion. An increase in AST and ALT was observed in the serum from both WT and P2X7−/− septic animals when compared to sham groups (p < 0.05) (Fig. 2 a and b, respectively). However, this increase in both enzymes was significantly lower in P2X7−/− septic animals in comparison to WT septic mice (p < 0.05) (Fig. 2). Thus, the absence of the P2X7 receptor seems to be important to protect liver integrity during the initial 24 h of sepsis.
Fig. 2.
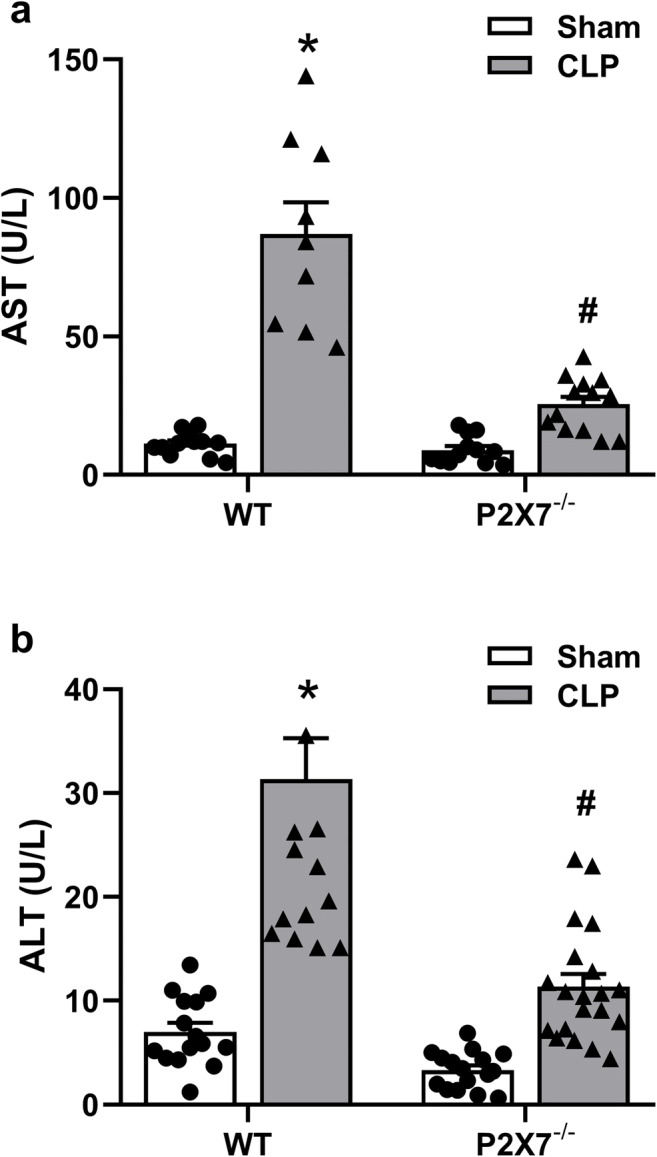
AST and ALT in the serum of WT and P2X7−/− animals 24 h after sepsis induction. Serum AST in WT (left (a)) and P2X7−/− (right (a)) mice. Serum ALT in WT (left (b)) P2X7−/− (right (b)) mice. The asterisk represents statistical difference (*p < 0.05) when compared to sham groups with their respective CLP group, while the number sign represents statistical difference (#p < 0.05) when compared to CLP groups (WT CLP vs. P2X7−/− CLP). Data were analyzed by two-way ANOVA and they are expressed as mean + S.E.M. (n ≥ 10 animals per group)
P2X7 receptor genetic deletion reduces sepsis-induced oxidative stress in mice liver
We observed a significant increase (p < 0.05) in the production of reactive species in septic WT animals when compared to the WT sham group, confirming the well-reported oxidative stress-induced by sepsis (Fig. 3a, left). P2X7-deficient septic mice did not show an increase in DCF oxidation when compared to their respective sham group (p > 0.05). Therefore, the genetic deletion of P2X7 blocked the increase in DCF oxidation in the liver homogenates from septic animals compared to the septic WT group (p < 0.05) (Fig. 3a, right). In addition to the increase in free radical production, we observed an increase (p < 0.05) in lipid peroxidation and nitrite content in WT septic animals in comparison with the WT sham group (Fig. 3 b and d, left). No increases (p > 0.05) were observed in P2X7−/− septic mice when compared to the P2X7−/− sham group (Fig. 3 b and d, right). No differences were detected in the sulfhydryl content (Fig. 3c; p > 0.05). Our results confirm the oxidative stress induced by sepsis in the liver and show that P2X7-deficient animals were protected, suggesting that the P2X7 receptor may be necessary for the production of reactive species in the liver of septic animals 24 h after surgery.
Fig. 3.

The genetic deletion of the P2X7 receptor blocks the production of free radicals in the liver of septic mice. a The production of reactive species as measured by the oxidation of 2′,7′-dichlorofluorescein (DCF), b lipid peroxidation measured by substances reactive to thiobarbituric acid (TBARS), c total sulfhydryl content, and d nitrite levels in the liver of WT and P2X7−/− mice 24 after sepsis induction. The asterisk represents statistical difference (*p < 0.05) when compared to sham groups with their respective CLP group, while the number sign represents statistical difference (#p < 0.05) when compared to CLP groups (WT CLP vs. P2X7 CLP−/−). Data were analyzed by two-way ANOVA and they are expressed as mean ± S.E.M. (n ≥ 4 animals per group)
P2X7 receptor contributes to an imbalance in the activities of antioxidant enzymes (SOD/CAT) in the liver septic animals
SOD and CAT activities were analyzed in the liver of septic mice. A decrease in CAT activity was observed (Fig. 4b, left) in septic WT animals (p < 0.05), but no statistical differences were found in SOD activity in CLP WT animals when compared with their respective sham group (Fig. 4a; p > 0.05). When we evaluated the P2X7−/− animals, we did not verify significant differences in SOD or CAT activities when compared with their sham group (Fig. 4 a and b; p > 0.05). These data show an imbalance in SOD/CAT activity in the liver of septic WT animals (p < 0.05), which can result in an accumulation of H2O2 (Fig. 4c, left). This imbalance between the ratio of SOD/CAT activities was not observed in the liver P2X7−/− septic animals (Fig. 4c, right).
Fig. 4.
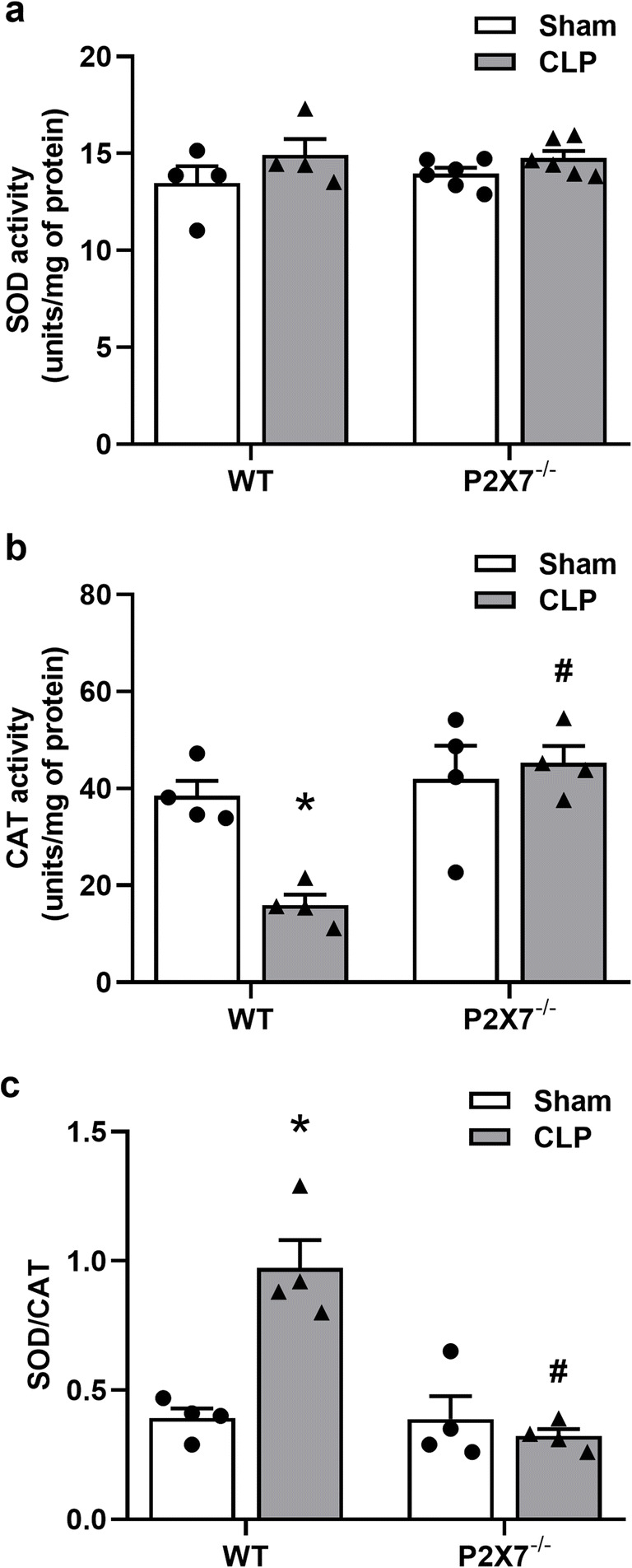
P2X7 receptor induces an imbalance in SOD/CAT activities in the liver from septic animals. a SOD activity was measured by auto-oxidation of the pyrogallol compound and b CAT activity was assessed by the disappearance of H2O2 liver homogenates from WT and P2X7−/− septic mice. c The ratio between SOD/CAT activities. The asterisk represents statistical difference (*p < 0.05) when compared to sham groups with their respective CLP group, while the number sign represents statistical difference (#p < 0.05) when compared to CLP groups (WT CLP vs. P2X7 CLP −/−). Data were analyzed by two-way ANOVA and they are expressed as mean + S.E.M. (n ≥ 4 animals per group)
Gene expression of iNOS and NADPH oxidases increase in the liver of septic animals
Biomarkers of redox status were assessed in the liver of septic animals 24 h after sepsis. Besides, we investigated the role of the P2X7 receptor in modulating the expression of enzymes involved in oxidative metabolism, such as NADPH oxidases and iNOS in liver tissue. Our results show an increase (p < 0.05) in the relative expression of Nox1, Nox2, and Nos2 in WT septic animals (Fig. 5a–c, left). The P2X7 receptor deletion prevented these increases since no differences in Nox1, Nox2 (p > 0.05), and a partial increase in Nos2 expression was observed in P2X7−/− septic mice in comparison to the P2X7−/− sham group (Fig. 5a–c right). Taken together, these results indicate that the P2X7 receptor contributes to the activation of NADPH oxidases and iNOS in liver tissue during sepsis.
Fig. 5.
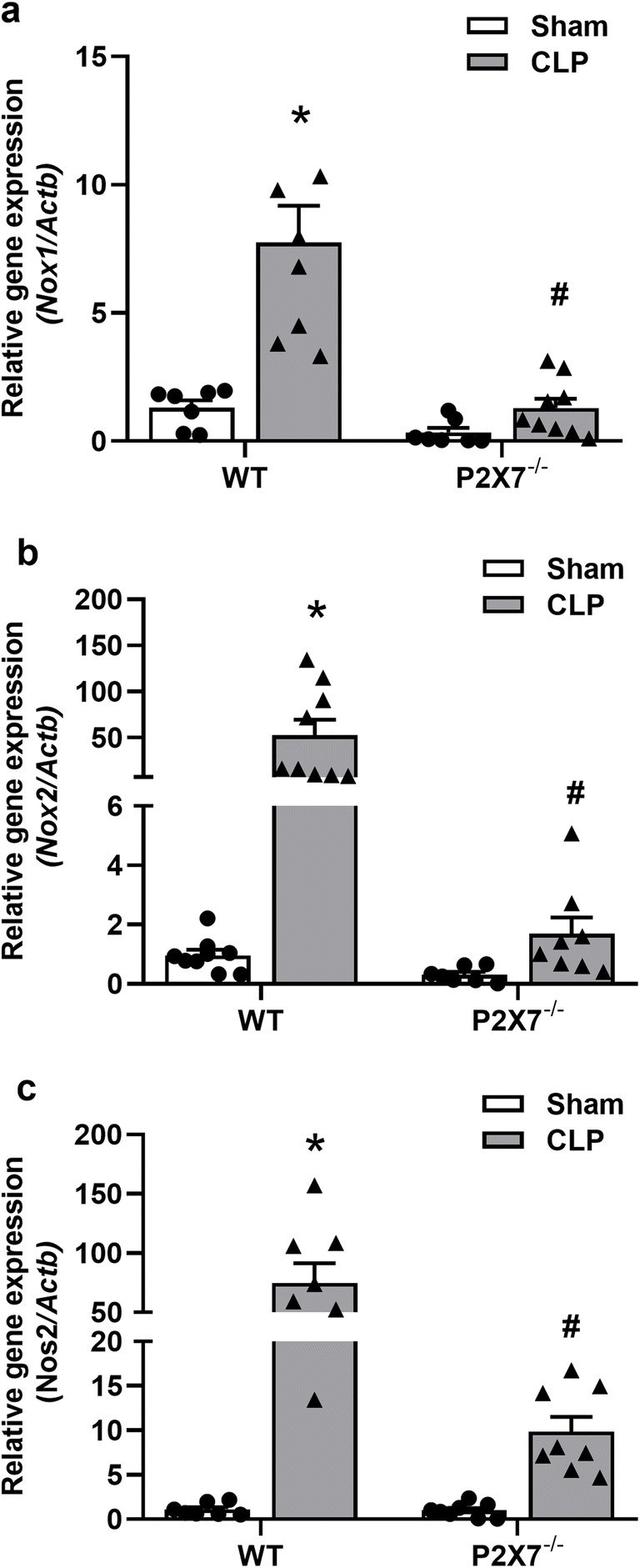
Gene expression of iNOS and NADPH oxidases increases in the liver of WT but not in P2X7-deficient septic mice. The relative expression of the a Nox1, b Nox2, and c Nos2 genes was analyzed by RT-qPCR in the liver of septic mice 24 h after surgery. The asterisk represents statistical difference (*p < 0.05) when compared to sham groups with their respective CLP group, while the number sign represents statistical difference (#p < 0.05) when compared to CLP groups (WT CLP vs. P2X7 CLP−/−). Data were analyzed by two-way ANOVA and they are expressed as mean ± S.E.M. (n ≥ 7 animals per group)
P2X7 receptor functionality is crucial to upregulate cytokine and chemokine expression in the liver of septic animals
The liver is one of the foremost vital organs severely affected during sepsis, and the macrophages Kupffer cells (KC) residing in the liver produce pro-inflammatory cytokines that contribute to the liver damage in sepsis. Thus, we investigated the contribution of the P2X7 receptor to IL-1β, IL-6, TNF-α, and IL-10 expression in the liver of septic animals. As depicted in Fig. 6, both IL-1β, IL-6, TNF-α, and IL-10 increase in the liver of WT septic animals when compared to the sham group (p < 0.05) (Fig. 6 a, b, c, and d respectively, left). In the absence of the P2X7 receptor, no significant increases in the expression of these cytokines were observed in the organ 24 h after CLP induction compared to the sham group (Fig. 6 a, b, c, and d, right) (p > 0.05).
Fig. 6.

Gene expression of inflammatory cytokines and chemokines in the liver of septic mice. The relative gene expression of the genes of a Il1β, b Il6, c Tnf, d Il10, e Cxcl1, and f CxCl2 was analyzed by RT-qPCR in the liver of septic mice 24 h after surgery. The asterisk represents statistical difference (*p < 0.05) when comparing the sham WT groups with their respective CLP group, while the number sign represents statistical difference (#p < 0.05) when comparing the CLP P2X7−/− groups with CLP WT. Data were analyzed by two-way ANOVA and they are expressed as mean ± S.E.M. of 3 independent experiments (n ≥ 6 animals per group)
Chemokines, such as CXCL1 and CXCL2, are small molecular weight proteins that are important for acute liver damage, and hepatocytes, KC, HCS, and LSEC produce them in response to injury. The expression of CXCL1 and CXCL2 by these cells generates a chemoattractive gradient that promotes the infiltration of monocytes, macrophages, NK cells, and neutrophils from the circulation to the inflammation site. To determine whether the P2X7 receptor is involved in mechanisms that upregulate the expression of these pro-inflammatory chemokines, we also evaluated the relative gene expression of the Cxcl1 and Cxcl2 genes in the liver of septic mice 24 h after surgery. We observed an increase (p < 0.05) in the relative expression of Cxcl1 in both septic WT and P2X7−/− animals (Fig. 6e), while Cxcl2 increases (p < 0.05) only in WT septic mice (Fig. 6f, left). However, Cxcl1 expression was higher (p < 0.05) in WT than P2X7−/− septic animals (Fig. 6e). These results suggest that the P2X7 receptor is important to the production of cytokines and chemokines in the liver during the acute phase of sepsis.
P2X7 receptor deletion reduces the number of inflammatory cells in liver tissue from septic mice
Liver histology demonstrated significant liver injury 24 h after sepsis induction in both WT and P2X7−/− mice when compared to their respective sham groups (Fig. 7a–g). Our histological analysis also shows an increased number of inflammatory cells in the liver parenchyma 24 h after sepsis induction in both septic WT and P2X7−/− animals (p < 0.05) (Fig. 7a–d). The number of inflammatory cells was higher in WT septic mice in comparison to P2X7−/− septic animals (p < 0.05) (Fig. 7e). Therefore, the P2X7 receptor seems to be involved in the recruitment of cells to liver parenchyma, promoting inflammation and tissue damage.
Fig. 7.

P2X7 genetic deletion reduces the number of inflammatory cells in the liver from septic mice. Representative photomicrographs of hepatic parenchyma of WT and P2X7−/− mice before (a and b, respectively) and 24 after surgery (c and f WT; and d and g P2X7−/−). c Liver parenchyma from CLP WT mice showing numerous inflammatory cells inside the sinusoids (arrows). d Less number of inflammatory cells in liver parenchyma of CLP P2X7−/− mice. e Quantitative analysis of the number of inflammatory cells per field in the liver parenchyma. Representative images of hepatocyte alterations in WT (f) and P2X7−/− (g) septic mice. Blue arrows indicate steatosis in hepatocytes, and blue stars indicate swelling/ballooning. h Liver injury score. The asterisk represents statistical difference (*p < 0.05) when compared to sham groups with their respective CLP group, while the number sign represents statistical difference (#p < 0.05) when compared to CLP groups (WT CLP vs. P2X7−/− CLP). Data were analyzed by two-way ANOVA and they are expressed as mean ± S.E.M. (n ≥ 5 animals per group). Twenty fields were analyzed by histological section for WT and P2X7−/− animals. Scale bar = 100 μm
Steatosis, ballooning degeneration, focal necrosis, and inflammation constituted the morphological patterns of liver injury in sepsis. Therefore, an index of liver injury (Fig. 7h) and a histological score of liver inflammation (Fig. 7e). Histological analysis of septic animals demonstrated that hepatic steatosis—characterized by the presence of cytoplasm vacuolization—was observed in both WT and P2X7−/− CLP mice, albeit slightly pronounced in WT animals (Fig. 7f–h). Indeed, P2X7−/− mice have an abnormal liver structure in which steatosis and cytoplasm vacuolization is already present independent of sepsis induction (Fig. 7 b and h), as reported by Arguin et al. [45].
Discussion
Sepsis is initiated by an infection that generates a systemic inflammatory response disrupting the immune system and triggering a cascade of events that can progress to multiple organ failure and host death [1]. Sepsis is the leading cause of death in non-cardiac intensive care units worldwide and represents a significant public health concern [4, 46]. P2X7 receptor deletion attenuates the systemic inflammatory responses and improves outcomes in a murine model of sepsis [33]. This receptor aggravates inflammation and tissue damage in different organs during sepsis, including the lungs, intestine, and brain [24, 27, 33, 34]. Here, we show that the P2X7 receptor deletion attenuates inflammation and oxidative damage in the liver from septic mice.
The P2X7 receptor mediates liver oxidative stress, inflammatory mechanisms [23, 47], and fibrogenesis [48]. This receptor triggers Kupffer cell inflammatory cellular responses and hepatocyte damage in liver diseases [20, 23, 49]. Indeed, we found that P2X7 receptor expression increases in the liver of septic mice. Moreover, AST and ALT enzymes released by hepatic cell disruption are reduced in the serum of septic P2X7-deficient mice, providing evidence for the relevance of this receptor in sepsis-associated liver injury. Accordingly, P2X7-deficient mice showed a significant reduction in serum ALT in experimental models of acetaminophen-induced acute liver injury [49] and nonalcoholic steatohepatitis [47]. Nevertheless, our histological analysis showed that P2X7-deficient mice have basal hepatic steatosis, and because of that, no differences were observed between WT and P2X7−/− in the hepatic liver score. These results are in accordance with a previous study that showed a disturbed glucose metabolism with consequent higher glycemia, dyslipidemia, increased susceptibility to glucose intolerance, and insulin resistance, with consequent hepatic steatosis in P2X7-deficient mice [45]. Therefore, the reduction in serum hepatic enzymes and oxidative stress in P2X7−/− septic mice suggest that these alterations are secondary to sepsis, while hepatocyte injury score in these animals depends on the altered glucose metabolism with basal hepatic steatosis, as detected in sham mice.
Oxidative damage contributes to metabolic and inflammatory liver diseases [50]. Oxidative stress represents an imbalance between oxidizing and antioxidant agents. Cellular proteins, membrane lipids, and DNA are cellular structures that are primarily affected by ROS and RNS [51]. The oxidative imbalance results in structural and functional abnormalities in the liver. One of the main characteristics of sepsis pathophysiology is the activation of tissue-resident immune cells (i.e., Kupffer cells) and the infiltration of phagocytic cells in liver parenchyma (i.e., PMN and monocytes). These cells respond to an infection producing superoxide, hydrogen peroxide, and nitric oxide inducing tissue damage [50, 52, 53]. Our results show an increase in the production of reactive species in the liver of septic WT animals, as already reported in sepsis [52, 54].
Interestingly, the P2X7 genetic deletion blocked this effect 24 h after surgery, indicating a lower production of reactive species in these animals. In addition to the increase in free radical production, we observed an increase in lipid peroxidation and nitrite content in septic WT animals, but not in P2X7-deficient septic animals. These results are in accordance with previous reports showing P2X7-dependent mechanisms for the generation of ROS and RNS by murine macrophages [28, 31, 32, 55] and in the brain of septic mice [24].
In an experimental model of nonalcoholic steatohepatitis, the P2X7 receptor triggers oxidative stress, inflammation, and fibrosis [47, 56]. P2X7 receptor-selective antagonist Brilliant Blue G also prevents acetaminophen hepatotoxicity by reducing tissue oxidative stress and inflammation [57]. Indeed, Chatterjee et al. [35] showed that the P2X7 receptor induces the production of reactive species through NADPH oxidase-dependent mechanisms in a model of carbon tetrachloride-induced steatohepatitis. In this study, the P2X7 receptor increases the expression of the p47 phox subunit that binds to membrane subunit gp91 phox producing reactive species. We found an increased expression of Nox1 and Nox2 in the liver of WT but not in the liver of P2X7−/− mice. Therefore, the P2X7 receptor seems to be involved in the activation of oxidative mechanisms in the liver, contributing to sepsis pathogenesis.
The antioxidant enzymes SOD and CAT have a protective role in several diseases [51, 58]. Our results evaluating the activity of endogenous antioxidant enzymes in the hepatic tissue showed a decrease in CAT activity in WT septic animals but not in SOD activity. The ratio between the enzymatic activities of SOD/CAT in septic WT animals indicates an accumulation of H2O2. This imbalance was not observed in P2X7−/− septic animals, providing evidence that the functionality of this receptor also alters antioxidant mechanisms. These data are relevant since the antioxidant capacity in patients with sepsis correlates with the severity of the disease and outcomes [59].
Activated KCs have the ability to produce NO through the induction of iNOS expression [60]. The NO produced reacts with ROS to form a highly reactive and toxic compound, the ONOO−. In our settings, nitrite levels and the Nos2 expression are significantly attenuated in P2X7−/− septic animals when compared with WT septic mice. Several factors modulate the iNOS gene expression pathway. The cytokines TNF-α and IL-1β can induce the expression of NOS2 in the liver [60]. We also found a significantly diminished expression of IL-1β, IL-6, TNF-α, IL-10, CXCL, and CXCL2 in the liver of P2X7−/− septic mice. We and others have shown that genetic deletion or pharmacological inhibition of the P2X7 receptor decreases the production of inflammatory cytokines (IL-1β, IL-6, and IL-10), NO production, and the recruitment of neutrophils into the peritoneal cavity after sepsis induction [23, 27, 33]. In this line of evidence, our histological analysis showed a reduced number of inflammatory cells in the liver P2X7−/− septic mice, suggesting that the activity of this receptor is relevant for recruiting inflammatory cells to the liver during sepsis progression. These results provide evidence for a critical role of the P2X7 receptor in the development of liver inflammation and tissue damage in sepsis. Such findings are also in accordance with studies in humans that identified P2X7 receptor single-nucleotide polymorphism of gain-of-function related with an increased sepsis severity [61].
Conclusion
The present study shows that P2X7 receptor functionality contributes to sepsis-associated liver oxidative damage and inflammation favoring the progression of the disease. Therefore, these findings suggest possible therapeutic approaches based on P2X7 genotyping and administration of P2X7 receptor blockers to limit oxidative damage, inflammation, and liver injury during the acute phase of sepsis.
Funding
This work was supported by funds from the Conselho Nacional de Desenvolvimento Cientifico e Tecnológico do Brasil – CNPq (306839/2019-9 to RCS), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), and Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro - FAPERJ (E-26/202.701/2019 to LEBS; 26/010.101036/2018 and E-26/202.774/2018 to RCS).
Compliance with ethical standards
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
Experiments were performed with age- and weight-matched animals, according to the guidelines of the Brazilian College of Animal Experimentation (COBEA). The Commission for the Ethical Use of Research Animals (CEUA) from the Federal University of Rio de Janeiro (UFRJ) approved all experiments (protocol number: IBCCF138).
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Robson Coutinho-Silva and Luiz Eduardo Baggio Savio are joint senior authors.
Contributor Information
Robson Coutinho-Silva, Email: rcsilva@biof.ufrj.br.
Luiz Eduardo Baggio Savio, Email: savio@biof.ufrj.br.
References
- 1.Singer M, Deutschman CS, Seymour C, et al. The third international consensus definitions for sepsis and septic shock (sepsis-3) JAMA - J Am Med Assoc. 2016;315:801–810. doi: 10.1001/jama.2016.0287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Angus DC, Van Der Poll T. Severe sepsis and septic shock. N Engl J Med. 2013;369:840–851. doi: 10.1056/NEJMra1208623. [DOI] [PubMed] [Google Scholar]
- 3.Angus DC, Linde-Zwirble WT, Lidicker J, Clermont G, Carcillo J, Pinsky MR. Epidemiology of severe sepsis in the United States: analysis of incidence, outcome, and associated costs of care. Crit Care Med. 2001;29:1303–1310. doi: 10.1097/00003246-200107000-00002. [DOI] [PubMed] [Google Scholar]
- 4.Rudd KE, Johnson SC, Agesa KM, Shackelford KA, Tsoi D, Kievlan DR, Colombara DV, Ikuta KS, Kissoon N, Finfer S, Fleischmann-Struzek C, Machado FR, Reinhart KK, Rowan K, Seymour CW, Watson RS, West TE, Marinho F, Hay SI, Lozano R, Lopez AD, Angus DC, Murray CJL, Naghavi M. Global, regional, and national sepsis incidence and mortality, 1990–2017: analysis for the Global Burden of Disease Study. Lancet. 2020;395:200–211. doi: 10.1016/S0140-6736(19)32989-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.MEDZHITOV R, JANEWAY CJ. Innate immunity. N Engl J Med. 2000;83:338–344. doi: 10.7326/0003-4819-83-3-445_2. [DOI] [PubMed] [Google Scholar]
- 6.Iwasaki A, Medzhitov R. Control of adaptive immunity by the innate immune system. Nat Immunol. 2015;16:343–353. doi: 10.1038/ni.3123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Pandey S, Kawai T, Akira S. Microbial sensing by toll-like receptors and intracellular nucleic acid sensors. Cold Spring Harb Perspect Biol. 2015;7:1–18. doi: 10.1101/cshperspect.a016246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Takeuchi O, Akira S. Pattern recognition receptors and inflammation. Cell. 2010;140:805–820. doi: 10.1016/j.cell.2010.01.022. [DOI] [PubMed] [Google Scholar]
- 9.Aziz M, Jacob A, Yang W-L, Matsuda A, Wang P. Current trends in inflammatory and immunomodulatory mediators in sepsis. J Leukoc Biol. 2013;93:329–342. doi: 10.1189/jlb.0912437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Nesseler N, Launey Y, Aninat C, Morel F, Mallédant Y, Seguin P. Clinical review: the liver in sepsis. Crit Care. 2012;16:1–8. doi: 10.1186/cc11381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Possamai LA, Thursz MR, Wendon JA, Antoniades CG. Modulation of monocyte/macrophage function: a therapeutic strategy in the treatment of acute liver failure. J Hepatol. 2014;61:439–445. doi: 10.1016/j.jhep.2014.03.031. [DOI] [PubMed] [Google Scholar]
- 12.Yan J, Li S, Li S. The role of the liver in sepsis. Int Rev Immunol. 2014;33:498–510. doi: 10.3109/08830185.2014.889129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zhou Z, Xu MJ, Gao B. Hepatocytes: a key cell type for innate immunity. Cell Mol Immunol. 2016;13:301–315. doi: 10.1038/cmi.2015.97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Strnad P, Tacke F, Koch A, Trautwein C. Liver — guardian , modifier and target of sepsis. Nat Rev Gastroenterol Hepatol. 2017;14:55–66. doi: 10.1038/nrgastro.2016.168. [DOI] [PubMed] [Google Scholar]
- 15.Bogdanos DP, Gao B, Gershwin ME. Liver Immunology. Compr Physiol. 2013;3:567–598. doi: 10.1038/jid.2014.371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kramer L, Jordan B, Druml W, Bauer P, Metnitz PG, Austrian Epidemiologic Study on Intensive Care, ASDI Study Group Incidence and prognosis of early hepatic dysfunction in critically ill patients - a prospective multicenter study. Crit Care Med. 2007;35:1099–1104. doi: 10.1097/01.CCM.0000259462.97164.A0. [DOI] [PubMed] [Google Scholar]
- 17.Canabal JM, Kramer DJ. Management of sepsis in patients with liver failure. Curr Opin Crit Care. 2008;14:189–197. doi: 10.1097/MCC.0b013e3282f6a435. [DOI] [PubMed] [Google Scholar]
- 18.Protzer U, Maini MK, Knolle PA. Living in the liver: hepatic infections. Nat Rev Immunol. 2012;12:201–213. doi: 10.1038/nri3169. [DOI] [PubMed] [Google Scholar]
- 19.Sumi Y, Woehrle T, Chen Y, et al. Plasma ATP is required for neutrophil activation in a mouse sepsis model. Shock. 2014;42:142–147. doi: 10.1038/jid.2014.371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Vaughn BP, Robson SC, Burnstock G. Pathological roles of purinergic signaling in the liver. J Hepatol. 2012;57:916–920. doi: 10.1016/j.jhep.2012.06.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Burnstock G, Vaughn B, Robson SC. Purinergic signalling in the liver in health and disease. Purinergic Signal. 2013;10:51–70. doi: 10.1007/s11302-013-9398-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ralevic V, Burnstock G. Receptors for purines and pyrimidines. Pharmacol Rev. 1998;50:413–492. doi: 10.1007/978-3-642-28863-0_5. [DOI] [PubMed] [Google Scholar]
- 23.Savio LEB, de Andrade MP, Figliuolo VR, et al. CD39 limits P2X7 receptor inflammatory signaling and attenuates sepsis-induced liver injury. J Hepatol. 2017;67:716–726. doi: 10.1016/j.jhep.2017.05.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Savio LEB, Andrade MGJ, de Andrade MP, et al. P2X7 receptor signaling contributes to sepsis-associated brain dysfunction. Mol Neurobiol. 2017;54:6459–6470. doi: 10.1007/s12035-016-0168-9. [DOI] [PubMed] [Google Scholar]
- 25.Savio LEB, Coutinho-Silva R. Immunomodulatory effects of P2X7 receptor in intracellular parasite infections. Curr Opin Pharmacol. 2019;47:53–58. doi: 10.1016/j.coph.2019.02.005. [DOI] [PubMed] [Google Scholar]
- 26.Savio LEB, Mello P d A, da Silva CG, Coutinho-Silva R. The P2X7 receptor in inflammatory diseases: angel or demon? Front Pharmacol. 2018;9:1–31. doi: 10.3389/fphar.2018.00052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wu X, Ren J, Chen G, Wu L, Song X, Li G, Deng Y, Wang G, Gu G, Li J. Systemic blockade of P2X7 receptor protects against sepsis-induced intestinal barrier disruption. Sci Rep. 2017;7:1–13. doi: 10.1038/s41598-017-04231-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Moreira-Souza ACA, Almeida-da-Silva CLC, Rangel TP, Rocha GC, Bellio M, Zamboni DS, Vommaro RC, Coutinho-Silva R (2017) The P2X7 receptor mediates Toxoplasma gondii control in macrophages through canonical NLRP3 inflammasome activation and reactive oxygen species production. Front Immunol 8. 10.3389/fimmu.2017.01257 [DOI] [PMC free article] [PubMed]
- 29.Ferrari D, Pizzirani C, Adinolfi E, Lemoli RM, Curti A, Idzko M, Panther E, di Virgilio F. The P2X 7 receptor: a key player in IL-1 processing and release. J Immunol. 2006;176:3877–3883. doi: 10.4049/jimmunol.176.7.3877. [DOI] [PubMed] [Google Scholar]
- 30.Qu Y, Franchi L, Nunez G, Dubyak GR. Nonclassical IL-1β secretion stimulated by P2X7 receptors is dependent on inflammasome activation and correlated with exosome release in murine macrophages. J Immunol. 2007;179:1913–1925. doi: 10.4049/jimmunol.179.3.1913. [DOI] [PubMed] [Google Scholar]
- 31.Cruz CM, Rinna A, Forman HJ, Ventura ALM, Persechini PM, Ojcius DM. ATP activates a reactive oxygen species-dependent oxidative stress response and secretion of proinflammatory cytokines in macrophages. J Biol Chem. 2007;282:2871–2879. doi: 10.1074/jbc.M608083200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Hewinson J, MacKenzie AB. P2X7 receptor-mediated reactive oxygen and nitrogen species formation: from receptor to generators. Biochem Soc Trans. 2007;35:1168–1170. doi: 10.1042/BST0351168. [DOI] [PubMed] [Google Scholar]
- 33.Santana PT, Benjamim CF, Martinez CG, Kurtenbach E, Takiya CM, Coutinho-Silva R. The P2X7 receptor contributes to the development of the exacerbated inflammatory response associated with Sepsis. J Innate Immun. 2015;7:417–427. doi: 10.1159/000371388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Wang H, Hong LJ, Huang JY, Jiang Q, Tao RR, Tan C, Lu NN, Wang CK, Ahmed MM, Lu YM, Liu ZR, Shi WX, Lai EY, Wilcox CS, Han F. P2RX 7 sensitizes Mac-1/ICAM-1-dependent leukocyte-endothelial adhesion and promotes neurovascular injury during septic encephalopathy. Cell Res. 2015;25:674–690. doi: 10.1038/cr.2015.61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Chatterjee S, Rana R, Corbett J, Kadiiska MB, Goldstein J, Mason RP. P2X7 receptor-NADPH oxidase axis mediates protein radical formation and Kupffer cell activation in carbon tetrachloride-mediated steatohepatitis in obese mice. Free Radic Biol Med. 2012;52:1666–1679. doi: 10.1016/j.freeradbiomed.2012.02.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wichterman KA, Baue AE, Chaudry IH. Sepsis and septic shock -a review of laboratory and a proposal models. J Surg Res. 1980;29:189–201. doi: 10.1016/0022-4804(80)90037-2. [DOI] [PubMed] [Google Scholar]
- 37.Rittirsch D, Huber-lang MS, Flierl MA, Ward PA. 2009 Martinez-Cuantificación de apoptosis y senescencia celular. Nat Protoc. 2009;4:31–36. doi: 10.1038/nprot.2008.214.Immunodesign. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Lebel CP, Ischiropoulos H, Bondys SC. Evaluation of the probe 2′,7′-dichiorofluorescin as an indicator of reactive oxygen species formation and oxidative stress. Chem Res Toxicol. 1992;5:227–231. doi: 10.1021/tx00026a012. [DOI] [PubMed] [Google Scholar]
- 39.Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem. 1979;95:351–358. doi: 10.1016/0003-2697(79)90738-3. [DOI] [PubMed] [Google Scholar]
- 40.Aksenov MY, Markesbery WR. Changes in thiol content and expression of glutathione redox system genes in the hippocampus and cerebellum in Alzheimer ’ s disease. Neurosci Lett. 2001;302:141–145. doi: 10.1016/S0304-3940(01)01636-6. [DOI] [PubMed] [Google Scholar]
- 41.Green LC, Wagner DA, Glogowski J, Skipper PL, Wishnok JS, Tannenbaum SR. Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Anal Biochem. 1982;126:131–138. doi: 10.1016/0003-2697(82)90118-X. [DOI] [PubMed] [Google Scholar]
- 42.Aebi H. Oxygen radicals in biological systems. Methods Enzymol. 1984;105:121–126. doi: 10.1016/S0076-6879(84)05016-3. [DOI] [PubMed] [Google Scholar]
- 43.Marklund SL. Product of extracellular-superoxide dismutase catalysis. FEBS Lett. 1985;184:237–239. doi: 10.1016/0014-5793(85)80613-X. [DOI] [PubMed] [Google Scholar]
- 44.Kleiner DE, Brunt EM, Van Natta M, et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology. 2005;41:1313–1321. doi: 10.1002/hep.20701. [DOI] [PubMed] [Google Scholar]
- 45.Arguin G, Bourzac JF, Placet M, Molle CM, Paquette M, Beaudoin JF, Rousseau JA, Lecomte R, Plourde M, Gendron FP. The loss of P2X7 receptor expression leads to increase intestinal glucose transit and hepatic steatosis. Sci Rep. 2017;7:1–16. doi: 10.1038/s41598-017-13300-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Rhee C, Dantes R, Epstein L, et al. Incidence and trends of sepsis in US hospitals using clinical vs claims data, 2009-2014. JAMA. 2017;318:1241–1249. doi: 10.1001/jama.2017.13836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Das S, Seth RK, Kumar A, Kadiiska MB, Michelotti G, Diehl AM, Chatterjee S. Purinergic receptor X7 is a key modulator of metabolic oxidative stress-mediated autophagy and inflammation in experimental nonalcoholic steatohepatitis. Am J Physiol Liver Physiol. 2013;305:G950–G963. doi: 10.1152/ajpgi.00235.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Huang C, Yu W, Cui H, et al. P2X7 blockade attenuates mouse liver fibrosis. Mol Med Rep. 2014;9:57–62. doi: 10.3892/mmr.2013.1807. [DOI] [PubMed] [Google Scholar]
- 49.Hoque R, Sohail MA, Salhanick S et al (2012) P2X7 receptor-mediated purinergic signaling promotes liver injury in acetaminophen hepatotoxicity in mice. Am J Physiol - Gastrointest Liver Physiol 302. 10.1152/ajpgi.00352.2011 [DOI] [PMC free article] [PubMed]
- 50.Cichoz-Lach H, Michalak A. Oxidative stress as a crucial factor in liver diseases. World J Gastroenterol. 2014;20:8082–8091. doi: 10.3748/wjg.v20.i25.8082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Valko M, Leibfritz D, Moncol J, Cronin MTD, Mazur M, Telser J. Free radicals and antioxidants in normal physiological functions and human disease. Int J Biochem Cell Biol. 2007;39:44–84. doi: 10.1016/j.biocel.2006.07.001. [DOI] [PubMed] [Google Scholar]
- 52.Gujral JS, Hinson JA, Farhood A, Jaeschke H (2004) NADPH oxidase-derived oxidant stress is critical for neutrophil cytotoxicity during endotoxemia. Am J Physiol Gastrointest Liver Physiol 287(1):G243–52. 10.1152/ajpgi.00287.2003 [DOI] [PubMed]
- 53.Sakaguchi S, Furusawa S. Oxidative stress and septic shock: metabolic aspects of oxygen-derived free radicals generated in the liver during endotoxemia. FEMS Immunol Med Microbiol. 2006;47:167–177. doi: 10.1111/j.1574-695X.2006.00072.x. [DOI] [PubMed] [Google Scholar]
- 54.Kantrow SP, Taylor DE, Carraway MS, Piantadosi CA (1997) Oxidative metabolism in rat hepatocytes and mitochondria during sepsis. Arch Biochem Biophys 345(2):278–88. 10.1006/abbi.1997.0264 [DOI] [PubMed]
- 55.Pfeiffer ZA, Guerra AN, Hill LM, Gavala ML, Prabhu U, Aga M, Hall DJ, Bertics PJ. Nucleotide receptor signaling in murine macrophages is linked to reactive oxygen species generation. Free Radic Biol Med. 2007;42:1506–1516. doi: 10.1016/j.freeradbiomed.2007.02.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Chatterjee S, Das S. P2X7 receptor as a key player in oxidative stress-driven cell fate in nonalcoholic steatohepatitis. Oxidative Med Cell Longev. 2015;2015:1–7. doi: 10.1155/2015/172493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Abdelaziz HA, Shaker ME, Hamed MF, Gameil NM. Repression of acetaminophen-induced hepatotoxicity by a combination of celastrol and brilliant blue G. Toxicol Lett. 2017;275:6–18. doi: 10.1016/j.toxlet.2017.04.012. [DOI] [PubMed] [Google Scholar]
- 58.Halliwell B. Free radicals and antioxidants: updating a personal view. Nutr Rev. 2012;70:257–265. doi: 10.1111/j.1753-4887.2012.00476.x. [DOI] [PubMed] [Google Scholar]
- 59.Kumar S, Gupta E, Kaushik S, Kumar Srivastava V, Mehta S, Jyoti A. Evaluation of oxidative stress and antioxidant status: correlation with the severity of sepsis. Scand J Immunol. 2018;87:1–11. doi: 10.1111/sji.12653. [DOI] [PubMed] [Google Scholar]
- 60.Iwakiri Y, Kim MY. Nitric oxide in liver diseases. Trends Pharmacol Sci. 2015;36:524–536. doi: 10.1016/j.tips.2015.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Geistlinger J, Du W, Groll J, et al. P2RX7 genotype association in severe sepsis identified by a novel multi-individual array for rapid screening and replication of risk SNPs. Clin Chim Acta. 2012;413:39–47. doi: 10.1016/j.cca.2011.05.023. [DOI] [PubMed] [Google Scholar]