

Abstract
Microbes are ubiquitous in the natural environment of Caenorhabditis elegans. Bacteria serve as a food source for C. elegans but may also cause infection in the nematode host. The sensory nervous system of C. elegans detects diverse microbial molecules, ranging from metabolites produced by broad classes of bacteria to molecules synthesized by specific strains of bacteria. Innate recognition through chemosensation of bacterial metabolites or mechanosensation of bacteria can induce immediate behavioral responses. The ingestion of nutritive or pathogenic bacteria can modulate internal states that underlie long-lasting behavioral changes. Ingestion of nutritive bacteria leads to learned attraction and exploitation of the bacterial food source. Infection, which is accompanied by activation of innate immunity, stress responses, and host damage, leads to the development of aversive behavior. The integration of a multitude of microbial sensory cues in the environment is shaped by experience and context. Genetic, chemical, and neuronal studies of C. elegans behavior in the presence of bacteria have defined neural circuits and neuromodulatory systems that shape innate and learned behavioral responses to microbial cues. These studies have revealed the profound influence that host-microbe interactions have in governing the behavior of this simple animal host.
Microbiology
Caenorhabditis elegans lives in a microbe-rich environment that defines the ecology and has shaped the evolution of the organism (Schulenburg and Félix, 2017). The basic dichotomy for C. elegans in its interactions with this microbial community is that bacteria are an essential source of nutrition but may also be pathogenic and cause infection and death. Ecological survey and sequence analysis of bacteria isolated from the natural environment from which C. elegans are isolated has revealed a community of Proteobacteria, Bacteroidetes, Firmicutes, and Actinobacteria (Samuel et al., 2016), with notable enrichment of alpha-Proteobacteria genera such as Ochrobactrum and Pseudomonas in close association with C. elegans, likely colonizing the intestine (Dirksen et al., 2016). These associated bacteria may benefit the C. elegans host as a food source and in other ways. At the same time, the characterization of C. elegans strains in the wild has uncovered a broad range of pathogenic microorganisms, including bacteria, fungi, and viruses, which can cause sickness and death (Schulenburg and Félix, 2017).
The experimental study of C. elegans has typically involved laboratory cultivation on monoaxenic lawns of Escherichia coli OP50 seeded on agar plates supplemented with cholesterol (Brenner, 1974). Genetic and metabolomic characterization of alternative bacterial food sources for C. elegans, such as Comamonas, Bacillus subtilis and mutants of E. coli, has defined conserved requirements for micronutrients (Qi et al., 2017; Watson et al., 2014), novel mechanisms of host co-option of bacterial siderophores for iron acquisition (Qi and Han, 2018), and metabolic determinants of bacteria that can influence complex phenotypes such as lifespan (Han et al., 2017; Qi and Han, 2018; Saiki et al., 2008; Virk et al., 2012). Semi-defined axenic media has been developed for growth and cultivation of C. elegans, but live bacteria support optimal growth and development (Lenaerts et al., 2008).
Diverse bacteria are pathogenic to C. elegans. The human opportunistic pathogen Pseudomonas aeruginosa, which resides in soil and water, was shown to infect an evolutionarily diverse range of hosts, including C. elegans (Rahme et al., 1995; Tan et al., 1999). Bacillus thurigiensis produces a crystal pore-forming toxin that is highly toxic to C. elegans upon ingestion (Marroquin et al., 2000) and has attracted interest as a potential biocontrol method for nematodes that are pathogenic to animals and plants. The coryneform bacterium Microbacterium nematophilum causes a distinct mode of infection and host response through adherence to the rectal and post-anal cuticle, likely reflecting a natural infection of C. elegans (Hodgkin et al., 2000). A broad range of bacterial species, including environmental isolates and human pathogens, exhibit increased virulence towards C. elegans, compared with the survival of C. elegans on E. coli OP50 (Couillault and Ewbank, 2002). Experimental conditions and host status play an important role when considering the pathogenicity of a bacterial strain. As an illustration of this point, even E. coli OP50 can be considered as pathogenic in the presence of richer growth media or when colonizing aging or feeding-defective animals (Garigan et al., 2002; Garsin et al., 2001; Herndon et al., 2002; Kumar et al., 2019). Pathogenicity can also be altered by co-ingestion of non-pathogenic bacteria (Montalvo-Katz et al., 2013; Samuel et al., 2016). For example, factors secreted by Enterococcus faecium are protective against Salmonella pathogenesis (Rangan et al., 2016). The range of bacteria that can be pathogenic to C. elegans in the laboratory setting has been expanded by molecular engineering, for example by having E. coli strains carry plasmids expressing specific toxins of B. thurigiensis (Wei et al., 2003) or P. aeruginosa (McEwan et al., 2012), or even RNAi clones targeting essential C. elegans genes (Kamath et al., 2003).
Innate recognition
The innate recognition of bacteria by C. elegans is evident from observations of behavioral phenotypes of C. elegans in the presence of bacteria and its metabolites. Behaviors such as feeding (Avery and Horvitz, 1990), defecation (Thomas, 1990), egg-laying (Trent et al., 1983), and locomotion (Sawin et al., 2000) are affected by the presence of bacteria. Insights into the molecular cues sensed by C. elegans have come from the characterization of chemotaxis behaviors of C. elegans, which demonstrate that C. elegans propagated in the laboratory are attracted to a broad range of volatile organic molecules that are produced by bacterial metabolism and may serve as food cues (Bargmann and Horvitz, 1991; Bargmann et al., 1993; Ward, 1973). The genome of C. elegans encodes an expanded family of over 1000 chemoreceptor genes (Bargmann, 1998). Genetic analysis of chemotactic responses to diacetyl, which is produced by many bacteria, identified ODR-10 as a chemoreceptor for diacetyl (Sengupta et al., 1996). The chemical characterization of natural bacterial isolates that may serve as food for C. elegans in the wild has further revealed a number of volatile organic compounds that attract C. elegans (Worthy et al., 2018a). Bacterial food cues modulate dauer entry and exit (Golden and Riddle, 1984), and fatty acids derived from bacteria cause dauer larvae, which do not ingest bacteria, to exit dauer diapause (Kaul et al., 2014). In addition to chemosensation, C. elegans also detect the presence of bacteria through mechanosensation. Four classes of ciliated dopaminergic neurons are required for an innate slowing response that C. elegans display upon encountering a bacterial food source (Sawin et al., 2000). These neurons express mechanically-sensitive ion channels, display neuronal responses to mechanical forces, and also drive slowing in response to microbeads that are similar in size to bacteria (Kang et al., 2010; Sawin et al., 2000).
Gradients of molecular oxygen and carbon dioxide are generated by bacterial metabolism, and C. elegans exhibits robust detection and behavioral responses to these gases (Bretscher et al., 2008, 2011; Chang et al., 2006; Cheung et al., 2005; Gray et al., 2004; Hallem and Sternberg, 2008), which can drive the behavior of C. elegans in a microbial environment. C. elegans detects molecular oxygen with cytosolic guanylyl cyclase receptors expressed in the AQR, PQR, URX, and BAG neurons (Cheung et al., 2005; Gray et al., 2004; Zimmer et al., 2009) and exhibits a peaked preference for oxygen levels around ~8% in the absence of bacterial food (Gray et al., 2004). Multiple sensory neurons including BAG and AFD are responsive to carbon dioxide levels (Bretscher et al., 2008, 2011; Hallem and Sternberg, 2008), which may also serve as a cue that modulates behavioral responses to bacterial density. Ambient laboratory oxygen levels (~21%) would be expected to drive C. elegans into a bacterial lawn, where oxygen levels can be substantially lower (Gray et al., 2004; Reddy et al., 2011). However, the wild-type laboratory strain, N2, exhibits aerotaxis behavior that is altered in the presence of bacteria, such that the peaked preference for ~ 8% O2 levels is lost, with an increased attraction to higher levels of molecular oxygen. The altered aerotaxis behavior of N2 C. elegans in the presence of bacteria is caused by allele differences in the npr-1 gene, encoding a neuropeptide receptor (de Bono and Bargmann, 1998; Chang et al., 2006; Cheung et al., 2005; Gray et al., 2004), and the glb-5 gene, which encodes a neural globin (McGrath et al., 2009; Persson et al., 2009). The N2 strain carries a laboratory-acquired neomorphic 215V allele of npr-1, whereas natural isolates of C. elegans carry an ancestral 215F allele with reduced NPR-1 activity (de Bono and Bargmann, 1998; McGrath et al., 2009). Bacteria induce NPR-1-dependent differences in not only aerotaxis behavior, but a number of different behavioral phenotypes including differences in CO2 avoidance (Bretscher et al., 2008; Hallem and Sternberg, 2008), aggregation in feeding behavior, and roaming versus dwelling locomotion on bacterial lawns (Cheung et al., 2005; Gray et al., 2004). Altered behavioral responses to gradients of molecular oxygen also underlie the NPR-1-dependent avoidance of P. aeruginosa lawns (Reddy et al., 2009). Of note, behaviors dependent on NPR-1 in the N2 strain can be attenuated in the presence of mucoid Gram-negative bacterial strains, which have an altered surface due to the overproduction of an exopolysaccharide coat (Reddy et al., 2011). The pervasive influence that bacteria can have on the behavior of C. elegans is underscored by the modulation of behavior by NPR-1-dependent signaling circuitry (Macosko et al., 2009).
In addition to the innate responsiveness to molecules and mechanical cues that are produced by many bacteria, C. elegans also detects molecules that enable discrimination among specific bacterial species. Food choice assays and microdroplet-based assays have enabled the sensitive monitoring of head-turning behaviors to bacterial odors, which demonstrated that C. elegans can innately distinguish between odors emanating from E. coli OP50 and P. aeruginosa PA14 (Ha et al., 2010). C. elegans also exhibit chemotactic responses to autoinducer molecules produced by P. aeruginosa and Vibrio cholerae that mediate quorum sensing (Beale et al., 2006; Werner et al., 2014). Innate recognition of pathogenic Serratia marcescens has been shown to include the detection of volatile cues (Glater et al., 2014; Worthy et al., 2018b), as well as Serrawettin W2, a surfactant-like lipodepsipentapeptide, which acts as a chemical repellent of C. elegans (Pradel et al., 2007).
In vivo calcium imaging methods that directly measure activation of individual sensory neurons in response to bacterial cues has corroborated observations from behavioral assays and enabled the dissection of neuronal circuitry mediating innate recognition and preference. The activation of AWB and AWC chemosensory neurons, which mediate repulsive and attractive responses to volatile chemicals, was demonstrated and defined components of a circuit that mediates innate preference for P. aeruginosa PA14-conditioned media over E. coli OP50-conditioned media (Ha et al., 2010). Activation of the ASH neurons, in response to surfactant-like dodecanoic acid that is secreted by Streptomyces, was observed to be dependent on the SRB-6 olfactory receptor, defining a molecular mechanism for innate recognition of a repellant that induces immediate avoidance behavior (Tran et al., 2017). Comprehensive analyses of sensory responses in C. elegans have revealed at least 10 classes of sensory neurons that respond to nutritive E. coli supernatants (Zaslaver et al., 2015). The ASJ neurons were shown to be activated by both E. coli supernatants (Zaslaver et al., 2015), as well as nitric oxide (Hao et al., 2018) and phenazine-1-carboxamide (Meisel et al., 2014), which are produced by P. aeruginosa PA14.
Neuronal activation is accompanied by the rapid induction of gene transcription (Yap and Greenberg, 2018), and ASJ activation is accompanied by the induction of daf-7/TGF-beta transcription in the ASJ neurons within six minutes of exposure to P. aeruginosa and specifically its secondary metabolites, phenazine-1-carboxamide and the siderophore pyochelin (Meisel et al., 2014). DAF-7 is necessary for C. elegans avoidance of P. aeruginosa, and increased DAF-7 activity in the ASJ neurons induced by innate recognition of P. aeruginosa metabolites alters aerotaxis behavior to promote avoidance behavior (Meisel et al., 2014). Genetic analysis has defined the involvement of distinct cyclic-GMP-dependent signaling pathways in the ASJ neurons that couple the recognition of P. aeruginosa metabolites to the selective transcription of daf-7 (Park et al., 2020). The modulation of avoidance behavior induced by infection (discussed further below) by DAF-7 in response to innate recognition in the ASJ neurons of P. aeruginosa metabolites contrasts with the immediate attractive or repulsive responses that are induced by activation of the ASH neurons (Tran et al., 2017).
The characterization of C. elegans sensory responses to microbe-derived molecules in its environment suggest these host animals recognize molecules produced by broad classes of bacteria as well as molecules that are highly specific to bacterial strains. Many molecules are attractive to C. elegans, as might be anticipated for cues of bacterial food, but the diverse repertoire of molecules that can elicit sensory responses also suggests the ability to recognize and respond to specific strains of bacteria, depending on the physiological context.
Bacterial food and internal state
The ingestion of nutritive bacteria can influence the subsequent behavior of C. elegans. Associative learning paradigms that involve pairing either the presence or absence of nutritive, non-pathogenic E. coli bacteria with environmental conditions such as temperature result in a learned preference for the conditions associated with the fed, not starved, state (Hedgecock and Russell, 1975; Mori and Ohshima, 1995; Torayama et al., 2007). In addition, C. elegans that are fed non-pathogenic bacteria with a wide range of nutritive qualities — “good food” versus “bad food” that differ in their ease of ingestion and metabolic factors — can also exhibit a learned change in preference that is calibrated for better versus poorer food (Shtonda and Avery, 2006). These learned changes in behavior can last from minutes to days, depending on the conditioning protocol, thus reflecting a stable change in preference.
The foraging behaviors of C. elegans are also strongly influenced by past and present bacterial feeding conditions. While feeding on E. coli OP50, C. elegans alternate between active “roaming” states and inactive “dwelling” states in which they either explore or exploit their food source, respectively (Fujiwara et al., 2002). The proportion of time that C. elegans spends in roaming versus dwelling states is controlled by satiety levels, chemosensory inputs, and internal sensing of bacterial food ingestion (Ben Arous et al., 2009; Fujiwara et al., 2002; Shtonda and Avery, 2006). Animals that have been deprived of bacterial food exhibit an “enhanced slowing response” upon encountering a bacterial lawn, suggesting an important role for satiety state in these modes of food exploration (Sawin et al., 2000). The acute ingestion of bacterial food also plays a central role in food exploration: animals that are exposed to a lawn bacteria rendered largely inedible as a result of pharmacological treatment, spend almost all of their time in the roaming state (Ben Arous et al., 2009).
Serotonin signaling has been shown to have a key role in mediating the effects of the ingestion of nutritive bacteria on behavior. Serotonin biosynthesis and the serotonergic NSM neurons were shown to be required for the enhanced slowing response (Sawin et al., 2000) and for maintenance of the dwelling state (Flavell et al., 2013). NSM neurons extend a sensory dendrite to the surface of the pharyngeal lumen and are acutely activated upon bacterial food ingestion (Rhoades et al., 2019). Feeding-dependent NSM activation requires the acid-sensing ion channels DEL-3 and DEL-7 that localize to the NSM sensory dendrite and appear to mediate detection of a heat-stable bacterial component, connecting microbial recognition in the C. elegans alimentary canal to the modification of feeding behaviors.
While serotonin appears to promote states of slow locomotion, the neuropeptide pigment dispersing factor (PDF) is required for sustained roaming states (Flavell et al., 2013) and for mate search behaviors (Barrios et al., 2012), in which male C. elegans leave a bacterial food source to search for a mating partner (Lipton et al., 2004). PDF signaling appears to be antagonized by serotonin release (Flavell et al., 2013) and, in males, it promotes expression of daf-7 in ASJ neurons (Hilbert and Kim, 2018), which promotes mate searching behavior (Hilbert and Kim, 2017). daf-7 expression in ASJ is also positively regulated by satiety, thus allowing it to serve as a signal that integrates multiple internal cues to promote exploration. Increased motion through bacterial lawns during roaming states also activates dopamine signaling via the dopaminergic PDE neurons that appear to integrate the presence of food with the animal’s own motion (Cermak et al., 2020). Activation of dopaminergic neurons during roaming elevates egg-laying rates, allowing animals disperse their eggs across bacterial food sources.
Slower metabolic changes arising from bacterial food ingestion also influence foraging. Rictor/TORC2 signaling in intestinal cells promotes daf-7 expression in ASI neurons and elevates roaming behavior in a PDF-dependent manner (O’Donnell et al., 2018). The ETS-5 transcription factor functions in ASG and BAG sensory neurons to limit intestinal fat storage and promote PDF-dependent roaming (Juozaityte et al., 2017). Interestingly, the effects of ets-5 on roaming can be reversed by altering intestinal fat storage. In addition to ASG and BAG, the URX and ASI sensory neurons also impact fat storage (Palamiuc et al., 2017; Witham et al., 2016). URX has also been shown to detect the mobilization of peripheral fat stores, suggesting bi-directional communication (Witham et al., 2016). Genetic analysis of the sterol response element binding protein pathway for fat metabolism has also suggested a critical role for fat metabolism in the regulation of food-induced quiescence behaviors in C. elegans (Hyun et al., 2016). Reduced food intake can also alter the production of other metabolites that act on the nervous system. For example, 2 h of fasting reduces levels of kynurenic acid, which alters NMDA signaling and downstream serotonin signaling to impact feeding (Lemieux et al., 2015). Our current understanding of how specific species of bacteria might alter metabolic state to impact behavior remains more limited. However, a recent study showed that Providencia bacteria in the C. elegans gut produce the neurotransmitter tyramine, which is converted to octopamine by the C. elegans host to alter aversive sensory responses (O’Donnell et al., 2020), which were previously shown to be regulated by feeding state (Chao et al., 2004). A large number of neuroactive metabolites are produced by non-pathogenic bacteria, suggesting that other bacterial species-specific signals produced in the gut may similarly influence behavior.
The chemosensory and mechanosensory detection of bacteria can also influence C. elegans foraging states. When animals are removed from an E. coli food source, they exhibit a “local search” state where they display a high frequency of high-angle turns for ~15 minutes, before switching to a “global search” state where they dramatically reduce turning (Gray et al., 2005; Hills et al., 2004; Wakabayashi et al., 2004). The frequency of turning during the local search state depends on the density of the E. coli food lawn from which animals were removed (López-Cruz et al., 2019). In this case, chemosensory and mechanosensory neurons are required for bacterial food detection, as depletion of glutamate from both populations of sensory neurons abolishes local search (López-Cruz et al., 2019). Together, these studies of internal states reveal that the C. elegans nervous system surveys the past and present levels of non-pathogenic bacteria through multiple sensory modalities in order to change behavior over long time scales.
Infection and internal state
Infection following the ingestion of P. aeruginosa PA14, which is not only a good food source for C. elegans, but also highly pathogenic (Tan et al., 1999), causes an aversive learned response that is distinct from the effect of feeding on nutritive non-pathogenic bacteria. The initial innate preference of C. elegans for P. aeruginosa PA14 over E. coli OP50 was found to be reversed after feeding on P. aeruginosa PA14, with a subsequent preference for E. coli OP50 and aversion to P. aeruginosa PA14 (Ha et al., 2010; Zhang et al., 2005).
P. aeruginosa can kill C. elegans through multiple modes of toxicity that depend on bacterial strain and experimental conditions, including rapid toxicity from the secretion of diffusible toxins over the course of minutes (Darby et al., 1999; Kirienko et al., 2013; Mahajan-Miklos et al., 1999), or the development of an intestinal infection associated with intralumenal, extracellular bacterial proliferation and distention of the intestinal lumen with effacement of epithelial cells (Irazoqui et al., 2010; Tan et al., 1999) over the course of several hours (Tan et al., 1999). Infection of C. elegans by P. aeruginosa induces the activation of host innate immunity (Kim and Ewbank, 2018). The host response integrates innate immunity with cellular stress response pathways, such as the endoplasmic reticulum Unfolded Protein Response (Richardson et al., 2010) and mitochondrial stress pathways (Pellegrino et al., 2014), as well as responses to exogenous, toxin-mediated effects on mRNA translation (Dunbar et al., 2012; McEwan et al., 2012). The widespread induction of stress-activated signaling pathways, immune and stress-responsive genes, and morphological changes to the host reflect the disruption of normal physiology and homeostasis caused by infection with pathogenic bacteria. The broad activation of cellular stress responses with infection has itself been proposed to be a mechanism by which immune defense is activated (Liu et al., 2014; Pukkila-Worley, 2016; Reddy et al., 2016). Whereas specific microbial cues for the activation of innate immunity in C. elegans remain elusive, evidence points to a key role for host damage resulting from infection in activating innate immunity. The PMK-1 p38 mitogen-activated protein kinase pathway is activated by both bacterial infection (Fletcher et al., 2019; Kim et al., 2002; Troemel et al., 2006) and pore-forming toxin activity (Huffman et al., 2004). In the epidermal response to fungal infection, host damage may be signaled by the endogenously produced metabolite 4-hydroxyphenyllactic acid, which acts through GPCR signaling to activate PMK-1 and antifungal immunity (Zugasti et al., 2014).
The evolutionarily ancient role for host damage in the activation of innate immunity parallels its apparent role in the development of aversive behavior. Consistent with the kinetics of infection and modified choice behaviors, C. elegans exhibits avoidance of a lawn of pathogenic bacteria following bacterial infection (Melo and Ruvkun, 2012; Pradel et al., 2007; Pujol et al., 2001; Reddy et al., 2009; Schulenburg and Müller, 2004). Whereas morphological changes such as intestinal distention accompany the development of pathogen infection (Tan et al., 1999), such changes are only correlative, and host damage in the absence of such changes has been shown to be sufficient for C. elegans lawn avoidance behavior. C. elegans avoids bacteria such as Bacillus thurigiensis (Schulenburg and Müller, 2004), which produces a pore-forming toxin that rapidly kills C. elegans over a time scale of minutes without associated intestinal proliferation of bacteria (Marroquin et al., 2000). The development of a lawn avoidance response is also observed in the presence of Microbacterium nematophilum, which causes a distinct mode of infection resulting in the induction of a rectal swelling response and sickness (McMullan et al., 2012; Yook and Hodgkin, 2007). Moreover, E. coli strain HT115, which is used for feeding RNAi-based experiments and is a nutritious food source of C. elegans can be engineered to be toxic to C. elegans through the expression of dsRNA targeting genes that are essential for viability of C. elegans (Kamath et al., 2003). Aversion to the E. coli HT115 lawn develops over the time course that RNAi exerts toxic effects on the host, and even the addition of abiotic toxins to the lawn can also induce C. elegans to leave a bacterial lawn (Melo and Ruvkun, 2012). Notably, the subsequent lawn aversive behavior is not specific for E. coli HT115 only but also observed in response to other E. coli and even other bacterial species.
In addition to this generalized aversive response, exposure to pathogenic P. aeruginosa PA14 also causes a learned change in bacterial preference, where the preference for P. aeruginosa PA14 over E. coli OP50 is reversed. This change can be elicited by at least 4 h of P. aeruginosa PA14 exposure in adulthood or 12 h of exposure during the L1 larval stage (Jin et al., 2016; Zhang et al., 2005). In addition, 24 h of PA14 exposure beginning at the L4 larval stage can also reduce PA14 preference in progeny for up to four generations later (Moore et al., 2019). The molecular and circuit mechanisms by which exposure to P. aeruginosa elicits a change in C. elegans preference have been carefully examined. Infection with P. aeruginosa induces the increased transcription of tph-1 from the ADF neurons, and tph-1 mutants are defective for learned aversive choice behavior following P. aeruginosa PA14 infection (Zhang et al., 2005). tph-1 mutants exhibit increased susceptibility to killing by P. aeruginosa compared to wild-type, a difference that is abrogated when animals are constrained such that they cannot avoid the P. aeruginosa lawn (Shivers et al., 2009).
Circuit-level studies have localized the site of learning within sensorimotor circuits that underlies the learned change in bacterial preference. AWB and AWC olfactory neurons detect P. aeruginosa PA14 and E. coli OP50 odors, but their sensory responses to these cues are not altered after P. aeruginosa PA14 exposure (Ha et al., 2010). In addition, a subset of neurons in the downstream circuitry, such as AIY, AIZ, and AIB neurons, are required for navigation towards food sources in naïve animals, but not required for learned changes. In contrast, RIA and SMD neurons are not required for naïve food choice, but are required for the learned change in food preference after P. aeruginosa PA14 exposure (Jin et al., 2016; Zhang et al., 2005). These results suggest that RIA and SMD neurons are likely modulated during P. aeruginosa PA14 exposure. This modulation appears to require serotonergic signaling from ADF (Ha et al., 2010) and ins-6 and ins-7 insulin-like peptides whose expression also changes after learning (Chen et al., 2013). Thus, P. aeruginosa PA14 elicits changes in neuroendocrine signaling that impact specific nodes in the sensorimotor circuit to alter food preference. Interestingly, neuroendocrine signals from the gut are also critical for food aversion, as the dynamic expression of an intestinal insulin, ins-11, modulates aversive responses to P. aeruginosa (Lee and Mylonakis, 2017). In addition, changes in neuroendocrine signaling that modulate learning may be accompanied by changes in neuroendocrine signaling that alter innate behaviors, as is observed in the modulation of P. aeruginosa PA14 avoidance behavior by dynamic daf-7 expression (Meisel et al., 2014).
The observations that C. elegans exhibits a preference for one bacterial species over another, and that this preference can be changed based on experience, suggest that C. elegans can discriminate among bacterial species, which may be enabled in part by the diversity of microbial ligands that it can recognize. At the same time, the observations that C. elegans will leave a lawn of bacteria following infection and damage, and that the subsequent aversive response may not be specific for the bacteria causing the damage, suggest that a behavioral state characterized by a more general aversion to bacteria may also develop. This distinction underscores differences in assays for aversive behavior following infection. Lawn-leaving behavior may be influenced by changes in internal states that confer both specific and non-specific responses to bacteria. It is also possible that non-specific lawn aversion determinants, such as differential oxygen or carbon dioxide levels, may act differentially on the attraction or repulsion of C. elegans to particular bacteria to also influence the choice of C. elegans between two different bacterial strains.
Summary
The behavioral responses of C. elegans animals to microbial cues in their environment rely on the innate recognition of a vast and diverse set of bacterial sensory cues. Behavioral responses to these molecular and mechanosensory cues are influenced by experience and context, endowing C. elegans with a great deal of flexibility in how it responds and adapts to its microbial environment. Changes in satiety, tissue damage resulting from pathogenic infection, and recognition of specific bacterial metabolites can alter how the neural circuits in this animal process subsequent microbial cues. Nutritive bacteria can elicit a range of adaptive behavioral changes include learned attraction and stable switches to exploitative foraging behaviors. Pathogenic bacteria can elicit generalized responses, like bacterial aversion, as well as highly specific changes, like learned avoidance of harmful food sources. Thus, evolutionary ancient mechanisms to sense bacteria, as well as host damage arising from pathogenic bacteria, triggers not only cell-autonomous innate immune responses, but also organism-wide behavioral responses that are controlled by a nervous system that can flexibly respond to microbial sensory cues. These studies of host-microbe interactions in C. elegans may ultimately inform our understanding of how microbes impact nervous system function in more complex animals (Li and Liberles, 2015; Yang and Chiu, 2017).
Figure 1. C. elegans behavioral responses to bacteria unfold over multiple time scales.
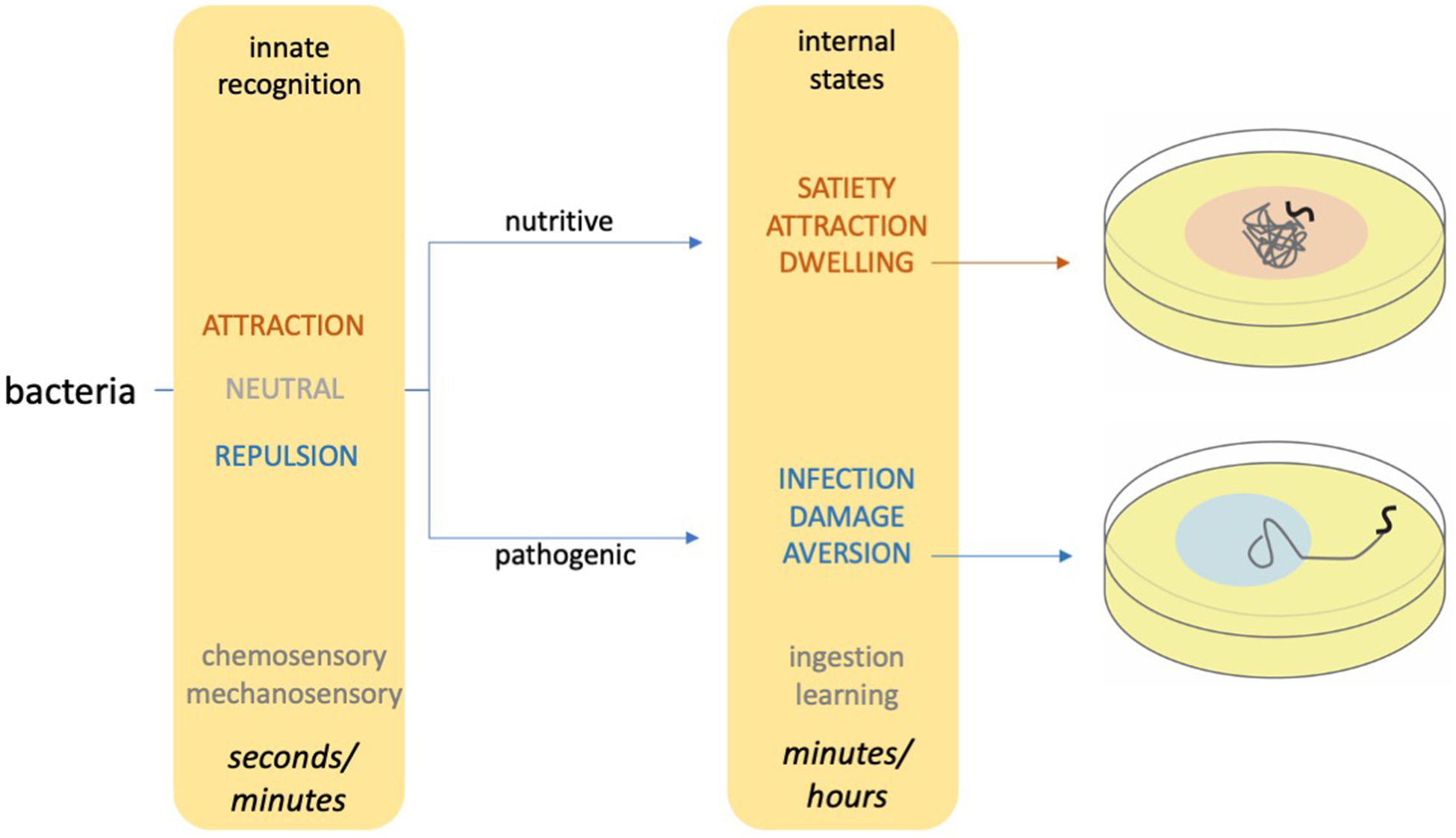
C. elegans consume diverse bacteria that differ in their nutritive qualities and their pathogenicity. Innate recognition of bacteria allows animals to generate rapid behavioral responses to bacterial odors and textures (left). After bacteria are ingested, animals undergo internal state changes that underlie long-lasting behavioral changes (middle). These long-lasting changes include alterations in their foraging strategies (right) and learned changes in bacterial preference.
Acknowledgements
We are grateful to the editors for the opportunity to write in honor of the memories of Sydney Brenner and John Sulston, who had central roles in the development of Caenorhabditis elegans as a simple organism for the molecular genetic analysis of behavior. We thank members of our research groups over the years who have joined us in studying host-microbe interactions of C. elegans. D.H.K. acknowledges funding from NIH GM084477. S.W.F acknowledges funding from NIH NS104892.
References
- Avery L, and Horvitz HR (1990). Effects of starvation and neuroactive drugs on feeding in Caenorhabditis elegans. J. Exp. Zool 253, 263–270. [DOI] [PubMed] [Google Scholar]
- Bargmann CI (1998). Neurobiology of the Caenorhabditis elegans genome. Science 282, 2028–2033. [DOI] [PubMed] [Google Scholar]
- Bargmann CI, and Horvitz HR (1991). Chemosensory neurons with overlapping functions direct chemotaxis to multiple chemicals in C. elegans. Neuron 7, 729–742. [DOI] [PubMed] [Google Scholar]
- Bargmann CI, Hartwieg E, and Horvitz HR (1993). Odorant-selective genes and neurons mediate olfaction in C. elegans. Cell 74, 515–527. [DOI] [PubMed] [Google Scholar]
- Barrios A, Ghosh R, Fang C, Emmons SW, and Barr MM (2012). PDF-1 neuropeptide signaling modulates a neural circuit for mate-searching behavior in C. elegans. Nat. Neurosci 15, 1675–1682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beale E, Li G, Tan M-W, and Rumbaugh KP (2006). Caenorhabditis elegans senses bacterial autoinducers. Appl. Environ. Microbiol 72, 5135–5137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ben Arous J, Laffont S, and Chatenay D (2009). Molecular and sensory basis of a food related two-state behavior in C. elegans. PloS One 4, e7584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Bono M, and Bargmann CI (1998). Natural variation in a neuropeptide Y receptor homolog modifies social behavior and food response in C. elegans. Cell 94, 679–689. [DOI] [PubMed] [Google Scholar]
- Brenner S (1974). The genetics of Caenorhabditis elegans. Genetics 77, 71–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bretscher AJ, Busch KE, and de Bono M (2008). A carbon dioxide avoidance behavior is integrated with responses to ambient oxygen and food in Caenorhabditis elegans. Proc. Natl. Acad. Sci. U. S. A 105, 8044–8049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bretscher AJ, Kodama-Namba E, Busch KE, Murphy RJ, Soltesz Z, Laurent P, and de Bono M (2011). Temperature, oxygen, and salt-sensing neurons in C. elegans are carbon dioxide sensors that control avoidance behavior. Neuron 69, 1099–1113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cermak N, Yu SK, Clark R, Huang Y-C, Baskoylu SN, and Flavell SW (2020). Whole-organism behavioral profiling reveals a role for dopamine in state-dependent motor program coupling in C. elegans. ELife 9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang AJ, Chronis N, Karow DS, Marletta MA, and Bargmann CI (2006). A distributed chemosensory circuit for oxygen preference in C. elegans. PLoS Biol. 4, e274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chao MY, Komatsu H, Fukuto HS, Dionne HM, and Hart AC (2004). Feeding status and serotonin rapidly and reversibly modulate a Caenorhabditis elegans chemosensory circuit. Proc. Natl. Acad. Sci. U. S. A 101, 15512–15517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Z, Hendricks M, Cornils A, Maier W, Alcedo J, and Zhang Y (2013). Two insulin-like peptides antagonistically regulate aversive olfactory learning in C. elegans. Neuron 77, 572–585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheung BHH, Cohen M, Rogers C, Albayram O, and de Bono M (2005). Experience-dependent modulation of C. elegans behavior by ambient oxygen. Curr. Biol. CB 15, 905–917. [DOI] [PubMed] [Google Scholar]
- Couillault C, and Ewbank JJ (2002). Diverse bacteria are pathogens of Caenorhabditis elegans. Infect. Immun 70, 4705–4707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darby C, Cosma CL, Thomas JH, and Manoil C (1999). Lethal paralysis of Caenorhabditis elegans by Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. U. S. A 96, 15202–15207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dirksen P, Marsh SA, Braker I, Heitland N, Wagner S, Nakad R, Mader S, Petersen C, Kowallik V, Rosenstiel P, et al. (2016). The native microbiome of the nematode Caenorhabditis elegans: gateway to a new host-microbiome model. BMC Biol. 14, 38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunbar TL, Yan Z, Balla KM, Smelkinson MG, and Troemel ER (2012). C. elegans detects pathogen-induced translational inhibition to activate immune signaling. Cell Host Microbe 11, 375–386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flavell SW, Pokala N, Macosko EZ, Albrecht DR, Larsch J, and Bargmann CI (2013). Serotonin and the neuropeptide PDF initiate and extend opposing behavioral states in C. elegans. Cell 154, 1023–1035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fletcher M, Tillman EJ, Butty VL, Levine SS, and Kim DH (2019). Global transcriptional regulation of innate immunity by ATF-7 in C. elegans. PLoS Genet. 15, e1007830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujiwara M, Sengupta P, and McIntire SL (2002). Regulation of body size and behavioral state of C. elegans by sensory perception and the EGL-4 cGMP-dependent protein kinase. Neuron 36, 1091–1102. [DOI] [PubMed] [Google Scholar]
- Garigan D, Hsu A-L, Fraser AG, Kamath RS, Ahringer J, and Kenyon C (2002). Genetic analysis of tissue aging in Caenorhabditis elegans: a role for heat-shock factor and bacterial proliferation. Genetics 161, 1101–1112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garsin DA, Sifri CD, Mylonakis E, Qin X, Singh KV, Murray BE, Calderwood SB, and Ausubel FM (2001). A simple model host for identifying Gram-positive virulence factors. Proc. Natl. Acad. Sci. U. S. A 98, 10892–10897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glater EE, Rockman MV, and Bargmann CI (2014). Multigenic natural variation underlies Caenorhabditis elegans olfactory preference for the bacterial pathogen Serratia marcescens. G3 Bethesda Md 4, 265–276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golden JW, and Riddle DL (1984). The Caenorhabditis elegans dauer larva: developmental effects of pheromone, food, and temperature. Dev. Biol 102, 368–378. [DOI] [PubMed] [Google Scholar]
- Gray JM, Karow DS, Lu H, Chang AJ, Chang JS, Ellis RE, Marletta MA, and Bargmann CI (2004). Oxygen sensation and social feeding mediated by a C. elegans guanylate cyclase homologue. Nature 430, 317–322. [DOI] [PubMed] [Google Scholar]
- Gray JM, Hill JJ, and Bargmann CI (2005). A circuit for navigation in Caenorhabditis elegans. Proc. Natl. Acad. Sci. U. S. A 102, 3184–3191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ha H, Hendricks M, Shen Y, Gabel CV, Fang-Yen C, Qin Y, Colón-Ramos D, Shen K, Samuel ADT, and Zhang Y (2010). Functional organization of a neural network for aversive olfactory learning in Caenorhabditis elegans. Neuron 68, 1173–1186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hallem EA, and Sternberg PW (2008). Acute carbon dioxide avoidance in Caenorhabditis elegans. Proc. Natl. Acad. Sci. U. S. A 105, 8038–8043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han S, Schroeder EA, Silva-García CG, Hebestreit K, Mair WB, and Brunet A (2017). Mono-unsaturated fatty acids link H3K4me3 modifiers to C. elegans lifespan. Nature 544, 185–190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hao Y, Yang W, Ren J, Hall Q, Zhang Y, and Kaplan JM (2018). Thioredoxin shapes the C. elegans sensory response to Pseudomonas produced nitric oxide. ELife 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hedgecock EM, and Russell RL (1975). Normal and mutant thermotaxis in the nematode Caenorhabditis elegans. Proc. Natl. Acad. Sci. U. S. A 72, 4061–4065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herndon LA, Schmeissner PJ, Dudaronek JM, Brown PA, Listner KM, Sakano Y, Paupard MC, Hall DH, and Driscoll M (2002). Stochastic and genetic factors influence tissue-specific decline in ageing C. elegans. Nature 419, 808–814. [DOI] [PubMed] [Google Scholar]
- Hilbert ZA, and Kim DH (2017). Sexually dimorphic control of gene expression in sensory neurons regulates decision-making behavior in C. elegans. ELife 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hilbert ZA, and Kim DH (2018). PDF-1 neuropeptide signaling regulates sexually dimorphic gene expression in shared sensory neurons of C. elegans. ELife 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hills T, Brockie PJ, and Maricq AV (2004). Dopamine and glutamate control area-restricted search behavior in Caenorhabditis elegans. J. Neurosci. Off. J. Soc. Neurosci 24, 1217–1225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hodgkin J, Kuwabara PE, and Corneliussen B (2000). A novel bacterial pathogen, Microbacterium nematophilum, induces morphological change in the nematode C. elegans. Curr. Biol. CB 10, 1615–1618. [DOI] [PubMed] [Google Scholar]
- Huffman DL, Abrami L, Sasik R, Corbeil J, van der Goot FG, and Aroian RV (2004). Mitogen-activated protein kinase pathways defend against bacterial pore-forming toxins. Proc. Natl. Acad. Sci. U. S. A 101, 10995–11000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hyun M, Davis K, Lee I, Kim J, Dumur C, and You Y-J (2016). Fat Metabolism Regulates Satiety Behavior in C. elegans. Sci. Rep 6, 24841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Irazoqui JE, Troemel ER, Feinbaum RL, Luhachack LG, Cezairliyan BO, and Ausubel FM (2010). Distinct pathogenesis and host responses during infection of C. elegans by P. aeruginosa and S. aureus. PLoS Pathog. 6, e1000982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jin X, Pokala N, and Bargmann CI (2016). Distinct Circuits for the Formation and Retrieval of an Imprinted Olfactory Memory. Cell 164, 632–643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Juozaityte V, Pladevall-Morera D, Podolska A, Nørgaard S, Neumann B, and Pocock R (2017). The ETS-5 transcription factor regulates activity states in Caenorhabditis elegans by controlling satiety. Proc. Natl. Acad. Sci. U. S. A 114, E1651–E1658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kamath RS, Fraser AG, Dong Y, Poulin G, Durbin R, Gotta M, Kanapin A, Le Bot N, Moreno S, Sohrmann M, et al. (2003). Systematic functional analysis of the Caenorhabditis elegans genome using RNAi. Nature 421, 231–237. [DOI] [PubMed] [Google Scholar]
- Kang L, Gao J, Schafer WR, Xie Z, and Xu XZS (2010). C. elegans TRP family protein TRP-4 is a pore-forming subunit of a native mechanotransduction channel. Neuron 67, 381–391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim DH, and Ewbank JJ (2018). Signaling in the innate immune response. WormBook Online Rev. C Elegans Biol 2018, 1–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim DH, Feinbaum R, Alloing G, Emerson FE, Garsin DA, Inoue H, Tanaka-Hino M, Hisamoto N, Matsumoto K, Tan M-W, et al. (2002). A conserved p38 MAP kinase pathway in Caenorhabditis elegans innate immunity. Science 297, 623–626. [DOI] [PubMed] [Google Scholar]
- Kirienko NV, Kirienko DR, Larkins-Ford J, Wählby C, Ruvkun G, and Ausubel FM (2013). Pseudomonas aeruginosa disrupts Caenorhabditis elegans iron homeostasis, causing a hypoxic response and death. Cell Host Microbe 13, 406–416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar S, Egan BM, Kocsisova Z, Schneider DL, Murphy JT, Diwan A, and Kornfeld K (2019). Lifespan Extension in C. elegans Caused by Bacterial Colonization of the Intestine and Subsequent Activation of an Innate Immune Response. Dev. Cell 49, 100–117.e6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee K, and Mylonakis E (2017). An Intestine-Derived Neuropeptide Controls Avoidance Behavior in Caenorhabditis elegans. Cell Rep. 20, 2501–2512. [DOI] [PubMed] [Google Scholar]
- Lemieux GA, Cunningham KA, Lin L, Mayer F, Werb Z, and Ashrafi K (2015). Kynurenic acid is a nutritional cue that enables behavioral plasticity. Cell 160, 119–131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lenaerts I, Walker GA, Van Hoorebeke L, Gems D, and Vanfleteren JR (2008). Dietary restriction of Caenorhabditis elegans by axenic culture reflects nutritional requirement for constituents provided by metabolically active microbes. J. Gerontol. A. Biol. Sci. Med. Sci 63, 242–252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Q, and Liberles SD (2015). Aversion and attraction through olfaction. Curr. Biol. CB 25, R120–R129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lipton J, Kleemann G, Ghosh R, Lints R, and Emmons SW (2004). Mate searching in Caenorhabditis elegans: a genetic model for sex drive in a simple invertebrate. J. Neurosci. Off. J. Soc. Neurosci 24, 7427–7434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Y, Samuel BS, Breen PC, and Ruvkun G (2014). Caenorhabditis elegans pathways that surveil and defend mitochondria. Nature 508, 406–410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- López-Cruz A, Sordillo A, Pokala N, Liu Q, McGrath PT, and Bargmann CI (2019). Parallel Multimodal Circuits Control an Innate Foraging Behavior. Neuron 102, 407–419.e8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macosko EZ, Pokala N, Feinberg EH, Chalasani SH, Butcher RA, Clardy J, and Bargmann CI (2009). A hub-and-spoke circuit drives pheromone attraction and social behaviour in C. elegans. Nature 458, 1171–1175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mahajan-Miklos S, Tan MW, Rahme LG, and Ausubel FM (1999). Molecular mechanisms of bacterial virulence elucidated using a Pseudomonas aeruginosa-Caenorhabditis elegans pathogenesis model. Cell 96, 47–56. [DOI] [PubMed] [Google Scholar]
- Marroquin LD, Elyassnia D, Griffitts JS, Feitelson JS, and Aroian RV (2000). Bacillus thuringiensis (Bt) toxin susceptibility and isolation of resistance mutants in the nematode Caenorhabditis elegans. Genetics 155, 1693–1699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McEwan DL, Kirienko NV, and Ausubel FM (2012). Host translational inhibition by Pseudomonas aeruginosa Exotoxin A Triggers an immune response in Caenorhabditis elegans. Cell Host Microbe 11, 364–374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGrath PT, Rockman MV, Zimmer M, Jang H, Macosko EZ, Kruglyak L, and Bargmann CI (2009). Quantitative mapping of a digenic behavioral trait implicates globin variation in C. elegans sensory behaviors. Neuron 61, 692–699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McMullan R, Anderson A, and Nurrish S (2012). Behavioral and immune responses to infection require Gαq- RhoA signaling in C. elegans. PLoS Pathog. 8, e1002530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meisel JD, Panda O, Mahanti P, Schroeder FC, and Kim DH (2014). Chemosensation of bacterial secondary metabolites modulates neuroendocrine signaling and behavior of C. elegans. Cell 159, 267–280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melo JA, and Ruvkun G (2012). Inactivation of conserved C. elegans genes engages pathogen- and xenobiotic-associated defenses. Cell 149, 452–466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Montalvo-Katz S, Huang H, Appel MD, Berg M, and Shapira M (2013). Association with soil bacteria enhances p38-dependent infection resistance in Caenorhabditis elegans. Infect. Immun 81, 514–520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moore RS, Kaletsky R, and Murphy CT (2019). Piwi/PRG-1 Argonaute and TGF-β Mediate Transgenerational Learned Pathogenic Avoidance. Cell 177, 1827–1841.e12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mori I, and Ohshima Y (1995). Neural regulation of thermotaxis in Caenorhabditis elegans. Nature 376, 344–348. [DOI] [PubMed] [Google Scholar]
- O’Donnell MP, Chao P-H, Kammenga JE, and Sengupta P (2018). Rictor/TORC2 mediates gut-to-brain signaling in the regulation of phenotypic plasticity in C. elegans. PLoS Genet. 14, e1007213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Donnell MP, Fox BW, Chao PH, Schroeder FC, Sengupta P (2020) A neurotransmitter produced by gut bacteria modulates host sensory behaviour. Nature, doi: 10.1038/s414586-020-2395-5. Online ahead of print [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palamiuc L, Noble T, Witham E, Ratanpal H, Vaughan M, and Srinivasan S (2017). A tachykinin-like neuroendocrine signalling axis couples central serotonin action and nutrient sensing with peripheral lipid metabolism. Nat. Commun 8, 14237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park J, Meisel JD, Kim DH (2020) Immediate activation of chemosensory neuron gene expression by bacterial metabolites is selectively induced by distinct cyclic-GMP-dependent pathways in C. elegans. PLoS Genet., In Press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pellegrino MW, Nargund AM, Kirienko NV, Gillis R, Fiorese CJ, and Haynes CM (2014). Mitochondrial UPR-regulated innate immunity provides resistance to pathogen infection. Nature 516, 414–417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Persson A, Gross E, Laurent P, Busch KE, Bretes H, and de Bono M (2009). Natural variation in a neural globin tunes oxygen sensing in wild Caenorhabditis elegans. Nature 458, 1030–1033. [DOI] [PubMed] [Google Scholar]
- Pradel E, Zhang Y, Pujol N, Matsuyama T, Bargmann CI, and Ewbank JJ (2007). Detection and avoidance of a natural product from the pathogenic bacterium Serratia marcescens by Caenorhabditis elegans. Proc. Natl. Acad. Sci. U. S. A 104, 2295–2300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pujol N, Link EM, Liu LX, Kurz CL, Alloing G, Tan MW, Ray KP, Solari R, Johnson CD, and Ewbank JJ (2001). A reverse genetic analysis of components of the Toll signaling pathway in Caenorhabditis elegans. Curr. Biol. CB 11, 809–821. [DOI] [PubMed] [Google Scholar]
- Pukkila-Worley R (2016). Surveillance Immunity: An Emerging Paradigm of Innate Defense Activation in Caenorhabditis elegans. PLoS Pathog. 12, e1005795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qi B, and Han M (2018). Microbial Siderophore Enterobactin Promotes Mitochondrial Iron Uptake and Development of the Host via Interaction with ATP Synthase. Cell 175, 571–582.e11. [DOI] [PubMed] [Google Scholar]
- Qi B, Kniazeva M, and Han M (2017). A vitamin-B2-sensing mechanism that regulates gut protease activity to impact animal’s food behavior and growth. ELife 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rahme LG, Stevens EJ, Wolfort SF, Shao J, Tompkins RG, and Ausubel FM (1995). Common virulence factors for bacterial pathogenicity in plants and animals. Science 268, 1899–1902. [DOI] [PubMed] [Google Scholar]
- Rangan KJ, Pedicord VA, Wang Y-C, Kim B, Lu Y, Shaham S, Mucida D, and Hang HC (2016). A secreted bacterial peptidoglycan hydrolase enhances tolerance to enteric pathogens. Science 353, 1434–1437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reddy KC, Andersen EC, Kruglyak L, and Kim DH (2009). A polymorphism in npr-1 is a behavioral determinant of pathogen susceptibility in C. elegans. Science 323, 382–384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reddy KC, Hunter RC, Bhatla N, Newman DK, and Kim DH (2011). Caenorhabditis elegans NPR-1-mediated behaviors are suppressed in the presence of mucoid bacteria. Proc. Natl. Acad. Sci. U. S. A 108, 12887–12892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reddy KC, Dunbar TL, Nargund AM, Haynes CM, and Troemel ER (2016). The C. elegans CCAAT-Enhancer-Binding Protein Gamma Is Required for Surveillance Immunity. Cell Rep. 14, 1581–1589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rhoades JL, Nelson JC, Nwabudike I, Yu SK, McLachlan IG, Madan GK, Abebe E, Powers JR, Colón-Ramos DA, and Flavell SW (2019). ASICs Mediate Food Responses in an Enteric Serotonergic Neuron that Controls Foraging Behaviors. Cell 176, 85–97.e14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richardson CE, Kooistra T, and Kim DH (2010). An essential role for XBP-1 in host protection against immune activation in C. elegans. Nature 463, 1092–1095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saiki R, Lunceford AL, Bixler T, Dang P, Lee W, Furukawa S, Larsen PL, and Clarke CF (2008). Altered bacterial metabolism, not coenzyme Q content, is responsible for the lifespan extension in Caenorhabditis elegans fed an Escherichia coli diet lacking coenzyme Q. Aging Cell 7, 291–304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Samuel BS, Rowedder H, Braendle C, Félix M-A, and Ruvkun G (2016). Caenorhabditis elegans responses to bacteria from its natural habitats. Proc. Natl. Acad. Sci. U. S. A 113, E3941–3949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sawin ER, Ranganathan R, and Horvitz HR (2000). C. elegans locomotory rate is modulated by the environment through a dopaminergic pathway and by experience through a serotonergic pathway. Neuron 26, 619–631. [DOI] [PubMed] [Google Scholar]
- Schulenburg H, and Félix M-A (2017). The Natural Biotic Environment of Caenorhabditis elegans. Genetics 206, 55–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schulenburg H, and Müller S (2004). Natural variation in the response of Caenorhabditis elegans towards Bacillus thuringiensis. Parasitology 128, 433–443. [DOI] [PubMed] [Google Scholar]
- Sengupta P, Chou JH, and Bargmann CI (1996). odr-10 encodes a seven transmembrane domain olfactory receptor required for responses to the odorant diacetyl. Cell 84, 899–909. [DOI] [PubMed] [Google Scholar]
- Shivers RP, Kooistra T, Chu SW, Pagano DJ, and Kim DH (2009). Tissue-specific activities of an immune signaling module regulate physiological responses to pathogenic and nutritional bacteria in C. elegans. Cell Host Microbe 6, 321–330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shtonda BB, and Avery L (2006). Dietary choice behavior in Caenorhabditis elegans. J. Exp. Biol 209, 89–102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan MW, Mahajan-Miklos S, and Ausubel FM (1999). Killing of Caenorhabditis elegans by Pseudomonas aeruginosa used to model mammalian bacterial pathogenesis. Proc. Natl. Acad. Sci. U. S. A 96, 715–720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas JH (1990). Genetic analysis of defecation in Caenorhabditis elegans. Genetics 124, 855–872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torayama I, Ishihara T, and Katsura I (2007). Caenorhabditis elegans integrates the signals of butanone and food to enhance chemotaxis to butanone. J. Neurosci. Off. J. Soc. Neurosci 27, 741–750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tran A, Tang A, O’Loughlin CT, Balistreri A, Chang E, Coto Villa D, Li J, Varshney A, Jimenez V, Pyle J, et al. (2017). C. elegans avoids toxin-producing Streptomyces using a seven transmembrane domain chemosensory receptor. ELife 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trent C, Tsuing N, and Horvitz HR (1983). Egg-laying defective mutants of the nematode Caenorhabditis elegans. Genetics 104, 619–647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Troemel ER, Chu SW, Reinke V, Lee SS, Ausubel FM, and Kim DH (2006). p38 MAPK regulates expression of immune response genes and contributes to longevity in C. elegans. PLoS Genet. 2, e183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Virk B, Correia G, Dixon DP, Feyst I, Jia J, Oberleitner N, Briggs Z, Hodge E, Edwards R, Ward J, et al. (2012). Excessive folate synthesis limits lifespan in the C. elegans: E. coli aging model. BMC Biol. 10, 67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wakabayashi T, Kitagawa I, and Shingai R (2004). Neurons regulating the duration of forward locomotion in Caenorhabditis elegans. Neurosci. Res 50, 103–111. [DOI] [PubMed] [Google Scholar]
- Ward S (1973). Chemotaxis by the nematode Caenorhabditis elegans: identification of attractants and analysis of the response by use of mutants. Proc. Natl. Acad. Sci. U. S. A 70, 817–821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watson E, MacNeil LT, Ritter AD, Yilmaz LS, Rosebrock AP, Caudy AA, and Walhout AJM (2014). Interspecies systems biology uncovers metabolites affecting C. elegans gene expression and life history traits. Cell 156, 759–770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wei J-Z, Hale K, Carta L, Platzer E, Wong C, Fang S-C, and Aroian RV (2003). Bacillus thuringiensis crystal proteins that target nematodes. Proc. Natl. Acad. Sci. U. S. A 100, 2760–2765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Werner KM, Perez LJ, Ghosh R, Semmelhack MF, and Bassler BL (2014). Caenorhabditis elegans recognizes a bacterial quorum-sensing signal molecule through the AWCON neuron. J. Biol. Chem 289, 26566–26573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Witham E, Comunian C, Ratanpal H, Skora S, Zimmer M, and Srinivasan S (2016). C. elegans Body Cavity Neurons Are Homeostatic Sensors that Integrate Fluctuations in Oxygen Availability and Internal Nutrient Reserves. Cell Rep. 14, 1641–1654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Worthy SE, Haynes L, Chambers M, Bethune D, Kan E, Chung K, Ota R, Taylor CJ, and Glater EE (2018a). Identification of attractive odorants released by preferred bacterial food found in the natural habitats of C. elegans. PloS One 13, e0201158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Worthy SE, Rojas GL, Taylor CJ, and Glater EE (2018b). Identification of Odor Blend Used by Caenorhabditis elegans for Pathogen Recognition. Chem. Senses 43, 169–180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang NJ, and Chiu IM (2017). Bacterial Signaling to the Nervous System through Toxins and Metabolites. J. Mol. Biol 429, 587–605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yap E-L, and Greenberg ME (2018). Activity-Regulated Transcription: Bridging the Gap between Neural Activity and Behavior. Neuron 100, 330–348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yook K, and Hodgkin J (2007). Mos1 mutagenesis reveals a diversity of mechanisms affecting response of Caenorhabditis elegans to the bacterial pathogen Microbacterium nematophilum. Genetics 175, 681–697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaslaver A, Liani I, Shtangel O, Ginzburg S, Yee L, and Sternberg PW (2015). Hierarchical sparse coding in the sensory system of Caenorhabditis elegans. Proc. Natl. Acad. Sci. U. S. A 112, 1185–1189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y, Lu H, and Bargmann CI (2005). Pathogenic bacteria induce aversive olfactory learning in Caenorhabditis elegans. Nature 438, 179–184. [DOI] [PubMed] [Google Scholar]
- Zimmer M, Gray JM, Pokala N, Chang AJ, Karow DS, Marletta MA, Hudson ML, Morton DB, Chronis N, and Bargmann CI (2009). Neurons detect increases and decreases in oxygen levels using distinct guanylate cyclases. Neuron 61, 865–879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zugasti O, Bose N, Squiban B, Belougne J, Kurz CL, Schroeder FC, Pujol N, and Ewbank JJ (2014). Activation of a G protein-coupled receptor by its endogenous ligand triggers the innate immune response of Caenorhabditis elegans. Nat. Immunol 15, 833–838. [DOI] [PMC free article] [PubMed] [Google Scholar]