

Abstract
Coronavirus disease 2019 (COVID-19) caused by SARS-CoV-2 has caused a global pandemics. To facilitate the detection of SARS-CoV-2 infection, various RT-LAMP assays using 19 sets of primers had been developed, but never been compared. We performed comparative evaluation of the 19 sets of primers using 4 RNA standards and 29 clinical samples from COVID-19 patients. Six of 15 sets of primers were firstly identified to have faster amplification when tested with four RNA standards, and were further subjected to parallel comparison with the remaining four primer sets using 29 clinical samples. Among these 10 primer sets, Set-4 had the highest positive detection rate of SARS-CoV-2 (82.8%), followed by Set-10, Set-11, and Set-13 and Set-17 (75.9%). Set-14 showed the fastest amplification speed (Tt value < 8.5 min), followed by Set-17 (Tt value < 12.5 min). Based on the overall detection performance, Set-4, Set-10, Set-11, Set-13, Set-14 and Set-17 that target Nsp3, S, S, E, N and N gene regions of SARS-CoV-2, respectively, were determined to be better than the other primer sets. Two RT-LAMP assays with the Set-4 primers in combination with any one of four other primer sets (Set-14, Set-10, Set-11, and Set-13) were recommended to be used in the COVID-19 surveillance.
Subject terms: Microbiology, Molecular biology
Introduction
Coronavirus disease 2019 (COVID-19), caused by the newly discovered coronavirus SARS-CoV-21, 2, is rapidly spreading throughout the world, posing a huge challenge to global public health security. As of 20 September, 2020, it has infected over 30.6 million people, and resulted in at least 950,000 deaths globally. In the absence of effective antiviral drugs or efficacious vaccines, early diagnosis of SARS-CoV-2 infection is essential for the containment of COVID-193,4, without which it is impossible to timely implement intervention and quarantine measures, and difficult to track contacts in order to limit virus spread.
Nucleic acid testing of various approaches are widely used as the primary tool for diagnosing COVID-193,4. Among them, reverse transcription quantitative PCR (RT-qPCR) methods have been set as the gold standard for laboratory confirmation of SARS-CoV-2 infection because of their proven track record as being the most robust technology in molecular diagnostics4–6. However, the RT-qPCR assay relies on sophisticated facilities with reliable supply of electricity and well-trained personnel in large general hospitals and health care facilities, or government labs (such as CDC), and it is relatively time-consuming (about 1.5–2 h). These limit its capacity in point-of-care settings. Moreover, visiting a clinical setting for testing increases the risk of spreading the virus. Therefore, an alternative, fast, simple, and sensitive point-of-care testing (POCT) is highly needed to facilitate the detection of SARS-CoV-2 infection in resource-limited settings3,7.
Loop-mediated isothermal amplification (LAMP) is a promising POCT method with high sensitivity, specificity, and rapidity, and it is easy-to-use8. To overcome the limitation of RT-qPCR assay, a number of RT-LAMP assays using at least 19 sets of different primers had been developed in the last few months for the detection of SARS-CoV-29–19. Although these assays had proven sensitive and effective for the detection of SARS-CoV-2, how do they compare with each other have not been evaluated. In this study, we compared all 19 sets of SARS-CoV-2-specific RT-LAMP primers using the mismatch-tolerant LAMP system that is faster and more sensitive than the conventional ones20,21, and screened the high-efficiency RT-LAMP assays for use in the detection of SARS-CoV-2.
Results
Strategy for the comparative evaluation
There were 19 sets of SARS-CoV-2 RT-LAMP primers available for the evaluation on 6 April, 20209–19. Among these primers, 2 sets were designed for binding to Nsp3 (non-structural proteins), 5 for RdRp (RNA-dependent RNA polymerase), 2 for E (envelope protein) and 6 for N (nucleocapsid protein) gene regions of SARS-CoV-2 (Fig. 1). These regions are highly conserved among SARS-CoV-2 and SARS-CoV, but distinct from five other human coronaviruses (MERS-CoV, OC43, 229E, NL63 and HKU1). Other 4 sets of primers were dispersed throughout the genome of SARS-CoV-2, and are located in the genomic regions of leader protein (Set-1), Nsp3 (Set-4), and S (spike protein) (Set-10 and Set-11) genes. Two to six sets of primers are adjacent to each other in the genomic location and 15 sets target to four genomic regions with lengths of 251–1954 bps. To minimize the consumption of clinical samples, and economize experimental efforts, we adopted a strategy that initiated by a preliminary evaluation of the primers binding to the four major genomic regions using in vitro-transcribed RNA standard, and followed by a further evaluation using clinical RNA samples (Fig. 1). Because four primer sets (i.e. S1, S4, S10 and S11) are dispersed throughout the genome (1–24,000 nt) and are not close to each other on the genome, it is difficult to obtain a long RNA template (about 24,000 nt length) covering the four primer sets by in vitro transcription (Fig. 1). Therefore, we directly moved the four primer sets to the next round of comparative experiments using clinical samples together with preliminarily selected primers.
Figure 1.

Genome location and evaluation strategy of 19 sets of SARS-CoV-2 RT-LAMP primers. The location of each primer set was detailed in Table 1.
Preliminary evaluation of primer sets
Using 3000 copies of in vitro-transcribed RNA standards of four gene segments (Targets 1–4) of SARS-CoV-2 (Fig. 1), we assessed the amplification performance of 15 sets of RT-LAMP primers. Except for Set-3 that failed in amplification, all other primer sets generated amplification curves with Time threshold (Tt) of 7.5–15.9 min and reached the plateau phase within 20 min (Fig. 2). In particular, six sets of the primers showed faster amplification with 10 min less Tt values than other primer sets (Fig. 2). The six sets of primers contain three (Set-14, Set-17 and Set-18) that bind to N gene and another three (Set-2, Set-5 and Set-13) that bind to Nsp, RdRp, and E genes, respectively. Faster amplification is often associated with higher detection sensitivity20. The six sets of primers were selected for further evaluation using clinical samples together with other four primer sets that bind to other genomic regions of the virus.
Figure 2.

Comparison of performance of 15 RT-LAMP assays using RNA standards. Two replicates were performed for each primer set. The replicates of the same primer set often generated completely overlapped amplification curves. A short bar was used to highlight the non-overlapping curves of the same primer. The curves of non-template control (NTC) are not shown. *The Tt values of the Set-8 were obtained by another repeated comparative experiments with Set-5and Set-9, both of which showed a consistent trend, but slightly lower Tt values than those shown here.
Comparative evaluation of ten primer sets using clinical samples
A total of 29 RNA samples extracted from clinical samples of COVID-19 patients were used at fourfold dilutions. Except one sample, all 29 RNA samples were detected as SARS-CoV-2 positive by at least one of the primer sets. Nine samples were detected as positive by all ten sets of primers and almost all reactions (except one with 49.5 min) had Tt values of less than 15.1 min, indicating high viral load. The primer Set-4 detected 24 positive samples, showing the highest positive detection rate (82.8%), followed by Set-10, Set-11, Set-13 and Set-17 that all detected 22 positive samples (75.9%) (Fig. 3A). Two primer sets, Set-1 and Set-18, had the lowest positive detection rates of 44.8% and 62.1%, respectively, and thus were excluded in the subsequent analyses. Comparison showed that the primer Set-14 had the lowest mean Tt value of less than 8.4 min, followed by Set-10, Set-11 and Set-13 that had mean Tt values of 11.1–11.5 min (Fig. 3A). These four fast-amplification sets of primers also had small standard deviations (SD) of 1.7–2.9, indicating that the RT-LAMP with these four primer sets were relatively more stable and faster than the other 15 primer sets. Compared with other primer sets, the Set-14 was the most efficient one that generated the fastest (the lowest Tt value) and the second fastest amplification in 14 and 7 samples, respectively, followed by Set-17 which was the fastest in 6 samples and second best in 9 samples, demonstrating these two primer sets had the best performance.
Figure 3.

Comparison of performance of 10 selected RT-LAMP primer sets using 41 clinical RNA samples. (A) Positive rates and Tt values of 10 selected RT-LAMP assays. The 41 clinical samples included 29 SARS-CoV-2 positive and 12 negative clinical samples that were previously determined by RT-qPCR assay28. The positive rate was calculated by dividing the number of positive sample by each primer set by total positive sample number of RT-qPCR assay (i.e. 29). (B) Paired comparison of Tt values of the primers Set-4, Set-10, Set-11, Set-13, Set-14 and Set-17. Because all RNA from clinical samples were fourfold diluted and some of them have very low viral load (high Ct values by RT-qPCR assay), some positive samples were not detected as positive by the RT-LAMP assay, which are defined as false-negative. We calculated the concordance rate by dividing the number of consistent results (true positive, true negative and false-negative) by any two primer sets by the total sample number (i.e. 41). SD standard deviation.
Because of their relatively high positive detection rates and lower Tt values, six primer sets including Set-4, Set-10, Set-11, Set-13, Set-14 and Set-17 were subjected to further pairwise comparison. The comparison showed that any two sets of these primers had high concordance performance (87.8–97.6%) for 41 clinical RNA samples (including 29 positive and 12 negative for SARS-CoV-2) (Fig. 3B). All the six primer sets had high amplification efficiency with mean Tt values of less than 12 min (Fig. 3B). Because of the highest positive detection rate, we further tested the sensitivity of the primer set-4. The results showed that it had the limit of the detection (LOD) of 3 copies per 25 µL reaction (Fig. 4), indicating a higher sensitivity than previously reported (Table 1)14.
Figure 4.
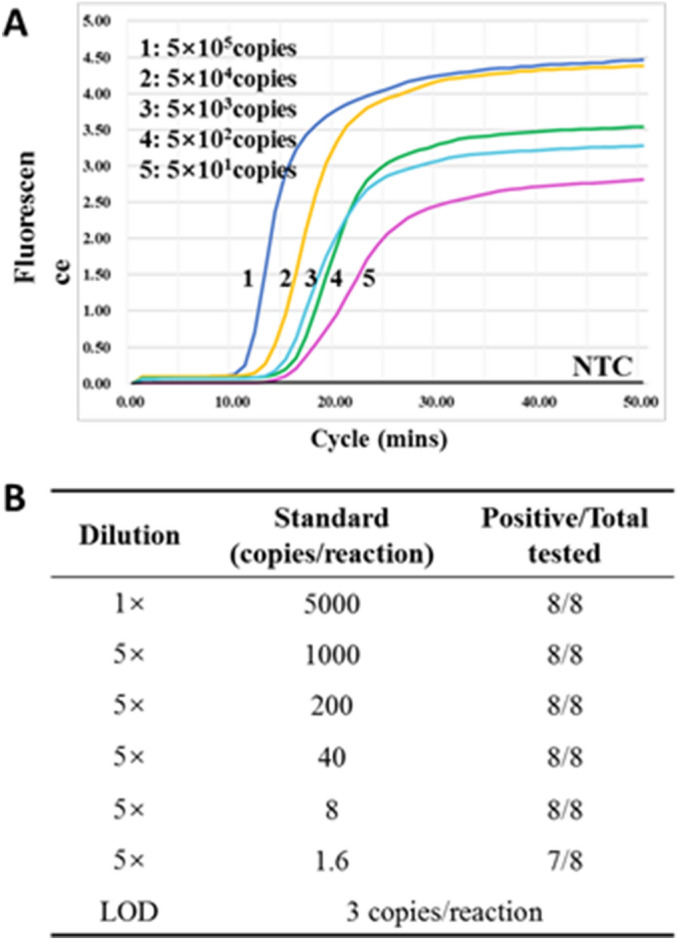
Sensitivity (A) and LOD (B) of the primer Set-4. NTC non-template control.
Table 1.
Information of 19 sets of RT-LAMP primers for the detection of SARS-CoV-2.
| Primer sets | Primer name | Primer sequence (5′-3′) | Target gene | Genomic location (nt) | Sensitivity (or LOD) | Refs |
|---|---|---|---|---|---|---|
| S1 | F3 | CTGCACCTCATGGTCATGTT | orf1ab | 498–711 | 1200 copies/25 μL reaction | 18 |
| B3 | AGCTCGTCGCCTAAGTCAA | |||||
| FIP | GAGGGACAAGGACACCAAGTGTATGGTTGAGCTGGTAGCAGA | |||||
| BIP | CCAGTGGCTTACCGCAAGGTTTTAGATCGGCGCCGTAAC | |||||
| LF | CCGTACTGAATGCCTTCGAGT | |||||
| LB | TTCGTAAGAACGGTAATAAAGGAGC | |||||
| S2 | F3 | TCCAGATGAGGATGAAGAAGA | orf1ab | 3043–3331 | 1.02 fg/25 μL reaction | 19 |
| B3 | AGTCTGAACAACTGGTGTAAG | |||||
| FIP | AGAGCAGCAGAAGTGGCACAGGTGATTGTGAAGAAGAAGAG | |||||
| BIP | TCAACCTGAAGAAGAGCAAGAACTGATTGTCCTCACTGCC | |||||
| LF | CTCATATTGAGTTGATGGCTCA | |||||
| LB | ACAAACTGTTGGTCAACAAGAC | |||||
| S3 | F3 | GGAATTTGGTGCCACTTC | orf1ab | 3145–3345 | 100 copies/15 μL reaction | 14 |
| B3 | CTATTCACTTCAATAGTCTGAACA | |||||
| FIP | CTTGTTGACCAACAGTTTGTTGACTTCAACCTGAAGAAGAGCAA | |||||
| BIP | CGGCAGTGAGGACAATCAGACACTGGTGTAAGTTCCATCTC | |||||
| LF | ATCATCATCTAACCAATCTTCTTC | |||||
| LB | TCAAACAATTGTTGAGGTTCAACC | |||||
| S4 | F3 | TGCAACTAATAAAGCCACG | orf1ab (Nsp3) | 6253–6446 | 100 copies/15 μL reaction | 14 |
| B3 | CGTCTTTCTGTATGGTAGGATT | |||||
| FIP | TCTGACTTCAGTACATCAAACGAATAAATACCTGGTGTATACGTTGTC | |||||
| BIP | GACGCGCAGGGAATGGATAATTCCACTACTTCTTCAGAGACT | |||||
| LF | TGTTTCAACTGGTTTTGTGCTCCA | |||||
| LB | TCTTGCCTGCGAAGATCTAAAAC | |||||
| S5 | F3 | TGCTTCAGTCAGCTGATG | orf1ab | 13,434–13,636 | 7 copies/10 µL reaction | 13 |
| B3 | TTAAATTGTCATCTTCGTCCTT | |||||
| FIP | TCAGTACTAGTGCCTGTGCCCACAATCGTTTTTAAACGGGT | |||||
| BIP | TCGTATACAGGGCTTTTGACATCTA TCTTGGAAGCGACAACAA | |||||
| LF | CTGCACTTACACCGCAA | |||||
| LB | GTAGCTGGTTTTGCTAAATTCC | |||||
| S6 | F3 | GGTATGATTTTGTAGAAAACCCA | orf1ab | 13,925–14,140 | 20 copies/25 µL reaction | 12 |
| B3 | CAACAGGAACTCCACTACC | |||||
| FIP | GGCATCACAGAATTGTACTGTTTTTGCGTATACGCCAACTTAGG | |||||
| BIP | AATGCTGGTATTGTTGGTGTACTGAGGTTTGTATGAAATCACCGAA | |||||
| LF | AACAAAGCTTGGCGTACACGTTCA | |||||
| S7 | F3 | GTTACGATGGTGGCTGTA | orf1ab | 14,885–15,081 | 5 copies/25 µL reaction | 16 |
| B3 | GGCATACTTAAGATTCATTTGAG | |||||
| FIP | AGCCTTACCCCATTTATTAAATGGAGCTAACCAAGTCATCGTCAA | |||||
| BIP | AATGAGTTATGAGGATCAAGATGCATTATAGTAGGGATGACATTACGT | |||||
| LF | AAACCAGCTGATTTGTCTAGGTTG | |||||
| S8 | F3 | AAACGTAATGTCATCCCTACT | orf1ab (RdRp) | 15,034–15,274 | 3 copies/25 μL reaction | 9 |
| B3 | GGTTTTCTACATCACTATAAACAGT | |||||
| FIP | ACAGATAGAGACACCAGCTACGCTCAAATGAATCTTAAGTATGCCA | |||||
| BIP | ATAGCCGCCACTAGAGGAGCCCAACCACCATAGAATTTGC | |||||
| LF | GTGCGAGCTCTATTCTTTGCACTA | |||||
| S9 | F3 | CCACTAGAGGAGCTACTGTA | orf1ab | 15,182–15,387 | 10 copies/20 µL reaction | 11 |
| B3 | TGACAAGCTACAACACGT | |||||
| FIP | AGGTGAGGGTTTTCTACATCACTATATTGGAACAAGCAAATTCTATGG | |||||
| BIP | ATGGGTTGGGATTATCCTAAATGTGTGCGAGCAAGAACAAGTG | |||||
| LF | CAGTTTTTAACATGTTGTGCCAACC | |||||
| LB | TAGAGCCATGCCTAACATGCT | |||||
| S10 | F3 | CTGACAAAGTTTTCAGATCCTCAG | S | 21,678–21,886 | NA | 14 |
| B3 | AGTACCAAAAATCCAGCCTCTT | |||||
| FIP | TCCCAGAGACATGTATAGCATGGAATCAACTCAGGACTTGTTCTTACC | |||||
| BIP | TGGTACTAAGAGGTTTGATAACCCTGTTAGACTTCTCAGTGGAAGCA | |||||
| LF | CCAAGTAACATTGGAAAAGAAA | |||||
| LB | GTCCTACCATTTAATGATGGTGTTT | |||||
| S11 | F3 | TCTATTGCCATACCCACAA | S | 23,693–23,937 | 200 copies/25 µL reaction | 12 |
| B3 | GGTGTTTTGTAAATTTGTTTGAC | |||||
| FIP | CATTCAGTTGAATCACCACAAATGTGTGTTACCACAGAAATTCTACC | |||||
| BIP | GTTGCAATATGGCAGTTTTTGTACATTGGGTGTTTTTGTCTTGTT | |||||
| LF | ACTGATGTCTTGGTCATAGACACT | |||||
| LB | TAAACCGTGCTTTAACTGGAATAGC | |||||
| S12 | F3 | CCGACGACGACTACTAGC | E | 26,191–26,424 | 20 copies/10 µL reaction | 15 |
| B3 | AGAGTAAACGTAAAAAGAAGGTT | |||||
| FIP | CTAGCCATCCTTACTGCGCTACTCACGTTAACAATATTGCA | |||||
| BIP | ACCTGTCTCTTCCGAAACGAATTTGTAAGCACAAGCTGATG | |||||
| LF | TCGATTGTGTGCGTACTGC | |||||
| LB | TGAGTACATAAGTTCGTAC | |||||
| S13 | F3 | AGCTGATGAGTACGAACTT | E | 26,226–26,441 | 5 copies /25 μL reaction | 16 |
| B3 | TTCAGATTTTTAACACGAGAGT | |||||
| FIP | ACCACGAAAGCAAGAAAAAGAAGTATTCGTTTCGGAAGAGACAG | |||||
| BIP | TTGCTAGTTACACTAGCCATCCTTAGGTTTTACAAGACTCACGT | |||||
| LB | CTGCGCTTCGATTGTGTGCGT | |||||
| S14 | F3 | CCAGAATGGAGAACGCAGTG | N | 28,354–28,569 | 1 copies/25 µL reaction | 17 |
| B3 | CCGTCACCACCACGAATT | |||||
| FIP | AGCGGTGAACCAAGACGCAGGGCGCGATCAAAACAACG | |||||
| BIP | AATTCCCTCGAGGACAAGGCGAGCTCTTCGGTAGTAGCCAA | |||||
| LF | TTATTGGGTAAACCTTGGGGC | |||||
| LB | TTCCAATTAACACCAATAGCAGTCC | |||||
| S15 | F3 | TGGCTACTACCGAAGAGCT | N | 28,525–28,741 | 120 copies/25 μL reaction | 18 |
| B3 | TGCAGCATTGTTAGCAGGAT | |||||
| FIP | TCTGGCCCAGTTCCTAGGTAGTCCAGACGAATTCGTGGTGG | |||||
| BIP | AGACGGCATCATATGGGTTGCACGGGTGCCAATGTGATCT | |||||
| LF | GGACTGAGATCTTTCATTTTACCGT | |||||
| LB | ACTGAGGGAGCCTTGAATACA | |||||
| S16 | F3 | AGATCACATTGGCACCCG | N | 28,702–28,914 | 5 copies/25 µL reaction | 16 |
| B3 | CCATTGCCAGCCATTCTAGC | |||||
| FIP | TGCTCCCTTCTGCGTAGAAGCCAATGCTGCAATCGTGCTAC | |||||
| BIP | GGCGGCAGTCAAGCCTCTTCCCTACTGCTGCCTGGAGTT | |||||
| LF | GCAATGTTGTTCCTTGAGGAAGTT | |||||
| LB | CGTAGTCGCAACAGTTAAGAAATTC | |||||
| S17 | F3 | GCCAAAAGGCTTCTACGCA | N | 28,774–28,971 | NA | 14 |
| B3 | TTGCTCTCAAGCTGGTTCAA | |||||
| FIP | TCCCCTACTGCTGCCTGGAGGCAGTCAAGCCTCTTCTCG | |||||
| BIP | TCTCCTGCTAGAATGGCTGGCATCTGTCAAGCAGCAGCAAAG | |||||
| LF | TGTTGCGACTACGTGATGAGGA | |||||
| LB | ATGGCGGTGATGCTGCTCT | |||||
| S18 | F3 | GCCAAAAGGCTTCTACGCA | N | 28,774–28,971 | 20 copies /25 µL reaction (118.6 copies/25 µL reaction) | 10 |
| B3 | TTGCTCTCAAGCTGGTTCAA | |||||
| FIP | TCCCCTACTGCTGCCTGGAGCAGTCAAGCCTCTTCTCGTT | |||||
| BIP | TCTCCTGCTAGAATGGCTGGCATCTGTCAAGCAGCAGCAAAG | |||||
| LB | TGGCGGTGATGCTGCTCTT | |||||
| S19 | F3 | AACACAAGCTTTCGGCAG | N | 29,083–29,311 | 20 copies/10 µL reaction | 15 |
| B3 | GAAATTTGGATCTTTGTCATCC | |||||
| FIP | CGCATTGGCATGGAAGTCACTTTGATGGCACCTGTGTAG | |||||
| BIP | TGCGGCCAATGTTTGTAATCAGCCAAGGAAATTTTGGGGAC | |||||
| LF | TTCCTTGTCTGATTAGTTC | |||||
| LB | ACCTTCGGGAACGTGGTT |
F3/B3: outer primers; FIP/BIP: forward and backward internal primers; LF/LB: forward and backward loop primers.
Specficity evaluation of six optimal primer sets based on sequence alignment
The specificity of these primer sets had been reported in previous studies9–19. In previous specificity experiments, common human respiratory pathogens were used, and none amplification curve or very weak amplification signals were observed. The pathogens used in the specificity experiment of the selected primer sets in the previous papers are listed in supplementary Table S2. To further examine the specificity of six recommended primer sets (Set-4, Set-10, Set-11, Set-13, Set-14 and Set-17) to other human coronaviruses, we performed sequence alignment analyses. SARS-CoV-2 shares 79.5% genomic homology with SARS-CoV1,2, indicating a relatively high sequence identity; but it is largely distinct from MERS-CoV and other four human coronaviruses (Supplementary Fig. S1). In particular, several primers of Set-4, Set-10 and Set-17 correspond to gaps or insertions of the genomes of MERS-CoV and other four common human coronaviruses OC43, 229E, NL63 and HKU1. These results implied that these six sets of primers were unable to bind to the genomes of MERS-CoV and four common human coronaviruses, therefore more specific for SARS-CoV-2. However, because of high sequence identity and the use of mismatch-tolerant RT-LAMP system that allows the presence of few mismatched bases between primers and templates, the SARS-CoV-2 RT-LAMP assays may generate a cross-amplification of SARS-CoV. In addition, the six primer sets did not generate amplification for all 12 COVID-19 negative RNA samples within 50 min, indicating that there was not or less non-specific amplification.
Discussion
SARS-CoV-2 transmission mainly occurs in the early and progressive stages of COVID-19 disease during which the patients and virus carriers have higher viral load than that in recovery stage22–24, and are generally more infectious. To contain the spread of the virus, early diagnosis is essential3,4. It helps to trigger timely intervention (e.g. quarantine, lockdown, and contact tracing), and facilitates to optimize clinical management. It is clear that serological assays are not suitable for this purpose, because detectable antibodies always appear several days after infection. Therefore, viral RNA testing is the primary method for early diagnosis of COVID-19. Despite being the most robust diagnostic tests, RT-qPCR-based assays are more centralized in core facilities, and they are not amenable for large-scale monitoring for asymptomatic and pre-symptomatic virus carriers in point-of-care settings (e.g. community and home). Therefore, community- and/or home-based nucleic acid assays that allow individuals to test in the community, at home, or other point-of-care sites without having to visit hospitals are convenient tools for the detection of SARS-CoV-2 infection by the general public3,7.
RT-LAMP assays are such needed tools8,20,21. In fact, various LAMP assays have been developed that included at least 19 sets of primers targeting to different genomic regions of SARS-CoV-2, with reported high detection sensitivity ranging from 1 to 1200 copies per 25 µL reaction9–19. However, these primers are never formally evaluated with clinical samples. The sensitivity and performance of a RT-LAMP assay are mainly determined by the primers, because other components of the reaction system are optimized and stable. Therefore, assessing the optimal RT-LAMP primer sets for the detection of SARS-CoV-2 infection is important for the selection of the best assay format to use for large field screening of COVID-19 patients.
Recently, the reaction system of RT-LAMP was further optimized to have higher sensitivity and faster amplification speed, even allowing the presence of few mismatched bases between primer and templates in a mismatch-tolerant version20,21. The new optimized reaction system containing an additional 0.15 U of high-fidelity DNA polymerase is called as mismatch-tolerant LAMP. The inclusion of an additional amount of high-fidelity DNA polymerase makes it a higher applicability to highly variable viruses, and a 10–15 min faster reaction speed than the conventional LAMP method. Using this new version, we assessed 19 sets of SARS-CoV-2 RT-LAMP primers. Six sets of primers showing faster amplification speed were firstly selected from 15 sets of primers using 4 RNA standards, and then tested with other 4 primer sets using 41 clinical samples. Eight sets of primers showed either comparable or better performance than the other 2 sets of primers (Set-1 and Set-18) as determined by positive detection rate. Of the 8 sets of primers, six were further selected based on high positive detection rate and/or overall faster amplification speed (with mean Tt value less than 13 min). The six primer sets are Set-4, Set-10, Set-11, Set-13, Set-14 and Set-17 that correspond to Nsp3, S, S, E, N, and N genes of SARS-CoV-2, respectively.
Among selected assays, the N gene-based RT-LAMP assays (Set-14 and Set-17) had the fastest amplification speed, followed by Orf, S and E gene-based assay (Set-4, Set-10, Set-11 and Set-13). This result suggested that the N gene-based RT-LAMP assay was more sensitive in detecting SARS-CoV-2 than that based on other genes, consisting with results of RT-qPCR assays5. In this study, Set-4 had the highest positive detection rates than all other primer sets, and had a LOD of 3 copies per 25 µL reaction, obviously more sensitive than the previously reported sensitivity of more than 100 copies per 25 µL reaction (Table 1 and Fig. 4)12,14. The sensitivity of Set-4 was comparable with highly sensitive primer sets Set-13 and Set-14 (less than 3 copies per 25 µL reaction)16,17. In addition, the sensitivity of primer Set-11 was less than 50 copies per 25 µL reaction (data not shown), obviously low 200 copies/25 µL reaction reported in previous study12. These indicated the mismatch-tolerant method significantly improved the detection sensitivity of RT-LAMP20. In addition, two of our previously reported primers, Set-8 and Set-18, exhibited high sensitivities of 3–20 copies per 25 µL reaction and good performance in the detection of clinical samples under the mismatch-tolerant reaction condition9,10, but they did not show better performance than other nine primer sets in this study. A reason might be that the use of the mismatch-tolerant reaction system generally improved the amplification efficiency of the primers reported by other groups20.
The analyzed primer sets showed high specificity in that they did not amplify any SARS-CoV-2 negative clinical samples. Sequence alignment analyses further supported that the six sets of optimal primers had good specificity to SARS-CoV-2, albeit they might generate non-specific amplification for SARS-CoV due to a high degree of sequence identity. However, given the lethal nature of both SARS-CoV-2 and SARS-CoV25, a non-specific positive result for SARS-CoV might also be of clinical importance.
Two nucleic acid assays targeting to different genes are suggested to be used in the detection of SARS-CoV-2 to avoid potential false-negative results5. Based on comparable performances, any two of the six optimal primer sets (Set-4, Set-10, Set-11, Set-13, Set-14 and Set-17) were recommend to be used in the detection of SARS-CoV-2. However, simultaneous use of Set-10 and Set-11, or Set-14 and Set-17 should be avoided because the former two sets target to the same S gene and the latter two sets target to the same N gene. In addition, because of highest positive detection rate and high sensitivity, Set-4 was strongly encouraged to be preferentially selected for the diagnosis of COVID-19 patients. Apart from the six recommend primer sets, other primers such as Set-2 and Set-5 also had good performance, and can also be used in the monitoring of COVID-19 infections.
Another advantage of our version of the RT-LAMP assay is that the results are easily visualized with a pH-sensitive indicator dye (e.g. cresol red and neutral red)26. Moreover, a combination of a nucleic acid extraction-free protocol and a master RT-LAMP mix containing all reagents (enzymes, primers, magnesium, nucleotides, dye and additives), except the template, enables the development of a simple kit that can be used at home, or a community-based diagnosis center for the detection of COVID-19 infection3,27.
In summary, we evaluated and selected six optimal primer sets from 19 sets of SARS-CoV-2 RT-LAMP primers through a comparative evaluation with clinical RNA samples from COVID-19 patients. Two RT-LAMP assays with the Set-4 primers and any one of the other four primer sets (Set-10, Set-11, Set-13 and Set-14) were recommended to be used in the COVID-19 surveillance to facilitate the early finding of asymptomatic and pre-symptomatic virus carriers in clinical and point-of-care settings, and the monitoring of environmental samples in the field.
Materials and methods
Ethics statement
The study was approved by Nantong Third Hospital Ethics Committee (E2020002: 3 February 2020). All experiments were performed in accordance with relevant guidelines and regulations. Written informed consents were obtained from each of the involved patients.
Preparation of RNA standard
To prepare RNA standard, four SARS-CoV-2 genomic segments (2720–3620 nt, 13,403–15,502 nt, 25,901–26,700 nt and 28,274–29,533 nt in Wuhan-Hu-1, GenBank: MN908947.3) were amplified from previously confirmed positive RNA sample with T7-promoter-containing primers (Supplementary Table S1). RNA standard was generated by in vitro transcription21, and quantitated by Qubit 4.0 Fluorometer (Thermo Fisher Scientific, USA). Copy number of RNA standard was estimated using the formula: RNA copies/mL = [RNA concentration (g/μL)/(nt transcript length × 340)] × 6.022 × 1023.
RNA samples of COVID-19 patients
A total of 29 RNA samples were obtained from COVID-19 patients described in our previous studies9,10. In brief, the throat swabs of COVID-19 patients were put into virus transport medium that contains Hank’s buffer, BSA, HEPES and antibiotics, and 300 μL were used for RNA extraction using RNA extraction Kit (Liferiver Biotechnology Co., Ltd., Shanghai) and eluted in 90 μL nuclease-free water. After screening and confirmation tests, the remaining RNA samples were stored at − 80 °C. When used for RT-LAMP assays, the stored SARS-CoV-2 positive RNA samples as confirmed by Novel Coronavirus (2019-nCoV) Real Time RT-PCR kit (Liferiver Biotechnology Co., Ltd., Shanghai) were thawed and fourfold diluted28. In addition, 12 SARS-CoV-2 negative clinical RNA samples were used as controls.
RT-LAMP assay
To assess the performance of 19 sets of RT-LAMP primers in the detection of SARS-CoV-2, an optimized mismatch-tolerant RT-LAMP method that has higher sensitivity and faster amplification speed than the conventional ones was used. A 25 µL RT-LAMP reaction mixture was prepared with 1 × isothermal amplification buffer, 6 mM MgSO4, 1.4 mM dNTPs, 8 units of WarmStart Bst 2.0 DNA polymerase, 7.5 units of WarmStartR RT, 0.15 unit of Q5 High-Fidelity DNA Polymerase, 0.2 μM each of primers of F3 and B3, 1.6 μM each of primers of FIP and BIP, 0.4 μM each of loop primer LF and/or LB, and 0.4 mM SYTO 9 (Life technologies, Carlsbad, CA, United States). The enzymes were all purchased from New England Biolabs (Beverly, MA, United States). In general, 3 μL of RNA standard or samples were added into each RT-LAMP reaction. The reaction was run at 63 °C for 50 min with real-time monitoring by the LightCycler 96 real-time PCR System (Roche Diagnostics, Mannheim, Germany).
Limit of detection (LOD)
Tenfold serial dilutions of the RNA standard were used as the standards to determine the sensitivity of selected primer sets. To test the LOD of the primer Set-4, fivefold serial dilutions of the RNA standard, from 5000 to 2 copies per 1 μL, were used. Each dilution was tested in a set of 8 replicates. A 95% probability of obtaining a positive result was used to define the LOD.
Supplementary Information
Acknowledgements
We thank the grants from the National Science and Technology Major Project of China (2019YFC1200603, 2017ZX10103009-002 and 2018ZX10711001) for financial support.
Author contributions
C.Z. conceived and designed the study, wrote the manuscript, and supervised the project. Y.D., X.W. and Y.L. performed the experiments. S.L., R.L. and Z.W. collected and screened clinical samples. C.Z., X.W., Y.D., and J.Q. analyzed the data. C.Z., G.Y. and X.J. interpreted the results. X.J. contributed to critical revision of the manuscript.
Data availability
All data was available from the article and author.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
These authors contributed equally: Yajuan Dong and Xiuming Wu.
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-020-80314-0.
References
- 1.Zhu N, et al. A novel coronavirus from patients with pneumonia in China, 2019. N. Engl. J. Med. 2020;382:727–733. doi: 10.1056/NEJMoa2001017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Zhou P, et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature. 2020;579:270–273. doi: 10.1038/s41586-020-2012-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Udugama B, et al. Diagnosing COVID-19: the disease and tools for detection. ACS Nano. 2020;14:3822–3835. doi: 10.1021/acsnano.0c02624. [DOI] [PubMed] [Google Scholar]
- 4.Lo YMD, Chiu RWK. Racing towards the development of diagnostics for a novel coronavirus (2019-nCoV) Clin. Chem. 2020;66:503–504. doi: 10.1093/clinchem/hvaa038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Chu DKW, et al. Molecular diagnosis of a novel coronavirus (2019-nCoV) causing an outbreak of pneumonia. Clin. Chem. 2020;66:549–555. doi: 10.1093/clinchem/hvaa029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Li Y, et al. A mismatch-tolerant RT-quantitative PCR: application to broad-spectrum detection of respiratory syncytial virus. Biotechniques. 2019;66:225–230. doi: 10.2144/btn-2018-0184. [DOI] [PubMed] [Google Scholar]
- 7.Nguyen T, Duong Bang D, Wolff A. Novel coronavirus disease (COVID-19): paving the road for rapid detection and point-of-care diagnostics. Micromachines (Basel) 2019;11:2020. doi: 10.3390/mi11030306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Notomi T, et al. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000;28:E63. doi: 10.1093/nar/28.12.e63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lu R, et al. Development of a novel reverse transcription loop-mediated isothermal amplification method for rapid detection of SARS-CoV-2. Virol. Sin. 2020;35:344–347. doi: 10.1007/s12250-020-00218-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lu R, et al. A Novel reverse transcription loop-mediated isothermal amplification method for rapid detection of SARS-CoV-2. Int. J. Mol. Sci. 2020;21:2826. doi: 10.3390/ijms21082826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Yu L, et al. Rapid detection of COVID-19 coronavirus using a reverse transcriptional loop-mediated isothermal amplification (RT-LAMP) diagnostic platform. Clin. Chem. 2020;66:975–977. doi: 10.1093/clinchem/hvaa102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Yan C, et al. Rapid and visual detection of 2019 novel coronavirus (SARS-CoV-2) by a reverse transcription loop-mediated isothermal amplification assay. Clin. Microbiol. Infect. 2020;26:773–779. doi: 10.1016/j.cmi.2020.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mohamed, E.-T., Haim H., B. & Jinzhao, S. A Single and Two-Stage, Closed-Tube, Molecular Test for the 2019 Novel Coronavirus (COVID-19) at Home, Clinic, and Points of Entry. (2020).
- 14.Park GS, et al. Development of reverse transcription loop-mediated isothermal amplification assays targeting SARS-CoV-2. J. Mol. Diag. 2020;22:729–735. doi: 10.1016/j.jmoldx.2020.03.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Broughton JP, et al. Rapid detection of 2019 novel coronavirus SARS-CoV-2 using a CRISPR-based DETECTR lateral flow assay. medRxiv. 2020 doi: 10.1101/2020.03.06.20032334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Yang W, et al. Rapid detection of SARS-CoV-2 using reverse transcription RT-LAMP method. Microb Biotechnol. 2020;13:950–961. doi: 10.1101/2020.03.02.20030130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Jiang M, et al. Development and validation of a rapid single-step reverse transcriptase loop-mediated isothermal amplification (RT-LAMP) system potentially to be used for reliable and high-throughput screening of COVID-19. Front Cell Infect Microbiol. 2020;10:331. doi: 10.3389/fcimb.2020.00331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Zhang Y, et al. Rapid molecular detection of SARS-CoV-2 (COVID-19) virus RNA using colorimetric LAMP. medRxiv. 2020 doi: 10.1101/2020.02.26.20028373. [DOI] [Google Scholar]
- 19.Lamb LE, Bartolone SN, Ward E, Chancellor MB. Rapid detection of novel coronavirus (COVID-19) by reverse transcription-loop-mediated isothermal amplification. PLoS One. 2020;15:e0234682. doi: 10.1371/journal.pone.0234682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhou Y, et al. A mismatch-tolerant reverse transcription loop-mediated isothermal amplification method and its application on simultaneous detection of all four serotype of dengue viruses. Front. Microbiol. 2019;10:1056. doi: 10.3389/fmicb.2019.01056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Li Y, et al. A mismatch-tolerant RT-LAMP method for molecular diagnosis of highly variable viruses. Bio-protocol. 2019;9:e3415. doi: 10.21769/BioProtoc.3415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Wang W, et al. Detection of SARS-CoV-2 in different types of clinical specimens. JAMA. 2020;323:1843–1844. doi: 10.1001/jama.2020.3786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zou L, et al. SARS-CoV-2 viral load in upper respiratory specimens of infected patients. N. Engl. J. Med. 2020;382:1177–1179. doi: 10.1056/NEJMc2001737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Yu F, et al. Quantitative detection and viral load analysis of SARS-CoV-2 in infected patients. Clin. Infect. Dis. 2020;71:793–798. doi: 10.1093/cid/ciaa345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wu Z, McGoogan JM. Characteristics of and important lessons from the coronavirus disease 2019 (COVID-19) outbreak in china: summary of a report of 72314 cases from the Chinese center for disease control and prevention. JAMA. 2020;323:1239–1242. doi: 10.1001/jama.2020.2648. [DOI] [PubMed] [Google Scholar]
- 26.Tanner NA, Zhang Y, Evans TC., Jr Visual detection of isothermal nucleic acid amplification using pH-sensitive dyes. Biotechniques. 2015;58:59–68. doi: 10.2144/000114253. [DOI] [PubMed] [Google Scholar]
- 27.Nie K, et al. Evaluation of a direct reverse transcription loop-mediated isothermal amplification method without RNA extraction for the detection of human enterovirus 71 subgenotype C4 in nasopharyngeal swab specimens. PLoS ONE. 2012;7:e52486. doi: 10.1371/journal.pone.0052486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lu R, et al. Epidemiological and clinical characteristics of COVID-19 patients in Nantong, China. J. Infect. Dev. Ctries. 2020;14:440–446. doi: 10.3855/jidc.12678. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All data was available from the article and author.