

Abstract
Ultrasound is emerging as a promising tool for both characterizing and fabricating engineered biomaterials. Ultrasound-based technologies offer a diverse toolbox with outstanding capacity for optimization and customization within a variety of therapeutic contexts, including improved extracellular matrix-based materials for regenerative medicine applications. Non-invasive ultrasound fabrication tools include the use of thermal and mechanical effects of acoustic waves to modify the structure and function of extracellular matrix scaffolds both directly, and indirectly via biochemical and cellular mediators. Materials derived from components of native extracellular matrix are an essential component of engineered biomaterials designed to stimulate cell and tissue functions and repair or replace injured tissues. Thus, continued investigations into biological and acoustic mechanisms by which ultrasound can be used to manipulate extracellular matrix components within three-dimensional hydrogels hold much potential to enable the production of improved biomaterials for clinical and research applications.
Keywords: Ultrasound, extracellular matrix, collagen, acoustics, biofabrication
1. Introduction
Tissue engineering combines the methodologies of cell biology, chemistry, and engineering to produce materials that replace or facilitate the repair of diseased or injured tissue [1]. Tissue engineering techniques typically utilize progenitor cells, biological scaffolds, and bioactive molecules, either alone or in combination, to achieve desired tissue characteristics [1, 2]. While the potential of tissue engineering to produce laboratory-grown, whole-organ transplants has gained widespread attention [2], other applications of tissue engineering include the development of materials that facilitate endogenous tissue repair [3]. Tissue engineering also offers the opportunity to address fundamental biological and physiological questions that cannot be answered with conventional approaches through the development of artificial tissues or organs for research purposes, as typified by “lab-on-a-chip” technologies [4].
The extracellular matrix (ECM) is a complex network of fibrillar glycoproteins and associated molecules that serves the critical role of defining tissue structure while also providing key biochemical and mechanical cues [5]. Cell-mediated assembly of ECMs plays essential roles in wound healing in many tissues [6, 7], and in tissue assembly during embryonic development [5]. As such, it is no surprise that therapeutic biomaterials targeting ECM assembly, structure, and function have become essential components from which to develop biomaterials for regenerative medicine applications. The potential clinical impact of ECM-based biomaterials is far-reaching, with utility for the treatment of injuries in tissues with low regenerative potential such as peripheral nerves [8], as well as chronic wounds in which the normal progression of healing is dysregulated [9]. In spite of significant progress, limitations still persist in the ability to engineer biomaterials that sufficiently recapitulate critical features of cell-assembled ECMs produced by healthy tissue [2, 10]. Thus, a critical need remains for technologies that can close the gap between the complex, highly organized structures of native ECMs and engineered products designed to enhance tissue repair.
Ultrasound is a versatile biomedical tool that has already revolutionized multiple domains of modern health care with non-invasive approaches to both diagnosis and therapy [11, 12]. Applications of ultrasound for tissue engineering have expanded rapidly [13], and include important contributions to both the characterization [14] and fabrication of engineered tissues [10]. In this review, we present recent innovations in the application of acoustic methodologies to manipulate both materials constructed from native ECM proteins, and synthetic scaffolds designed to mimic structural and functional aspects of the ECM in tissue homeostasis and repair. These applications include the strategic use of ultrasound to alter ECM protein structure via thermal effects and/or mechanical forces, as well as the use of engineered, acoustically responsive elements to manipulate scaffold properties. Finally, we discuss the use of ultrasound to enhance cell-mediated ECM remodeling behaviors, thereby indirectly modifying engineered scaffold structure. Together, these emerging ultrasound-based methodologies offer innovative strategies to enable non-invasive manufacturing and in situ translation of therapeutic materials for regenerative medicine.
2. Tissue Injury and Regeneration
Tissue repair in response to injury is an integral part of the physiology of many adult tissues but may be impaired as the result of comorbid health conditions. As well, the inherent repair capacity of some tissues is low. In both cases, failure to repair damaged tissue often leads to lifelong disability or premature death. Thus, interventions that support restoration of tissue function are of vital clinical relevance. In this section, we use dermal wound healing as a representative example to illustrate functional roles and clinical impacts of the ECM in tissue repair. It is important to note that the assembly, composition and organization of ECM structures are tissue-specific, and vary during course of development, with adult homeostasis, and during tissue repair in response to injury [15]. However, given the central role that ECMs play in organizing tissue structure and controlling function [5], the principles described herein apply to a variety of different tissues. Indeed, similar approaches for incorporating ECM-derived, tissue-specific cues into engineering frameworks have recently been reviewed for the treatment of injuries in bone [16], cartilage [17], tendon [18], cornea [19], muscle [20], and peripheral nerves [8, 21], and have been discussed extensively for chronic wounds [9, 22].
2.1. Role of Cell-Mediated ECM Remodeling in Wound Healing
Wound healing in response to injury relies on coordinated processes to rapidly induce hemostasis, followed by construction of replacement tissue [6]. Throughout this process, the ECM plays a critical role as a structure- and function-defining scaffold that coordinates cell and tissue responses [9]. Immediately following cutaneous injury, a transient ECM consisting primarily of fibrin and fibronectin forms quickly via the clotting cascade to restore homeostasis and barrier function [23]. This “provisional matrix” also supports subsequent stages of wound healing, including recruitment of immune mediators as well as activated fibroblasts from the surrounding tissue [24]. Cells that migrate into the wound space contribute to ongoing ECM remodeling through coordinated processes of matrix degradation, synthesis, and contraction [6, 25]. Several key signaling mechanisms are involved in the translation of ECM remodeling into coordinated tissue responses. These include cell-derived tensile forces and subsequent matrix contraction, which exposes cryptic, bioactive epitopes in matrix proteins [26], provides topological cues such as fiber alignment [27], and increases tissue tensile strength [28]. Matrix remodeling also acts as a cue for subsequent steps of tissue regeneration, as cell-derived tensile forces and associated changes in the surrounding ECM are key factors in recruitment and function of numerous cell types, including fibroblasts [29, 30], macrophages [31], and vascular cells [32]. In addition, many ECM components contain binding sites for growth factors, serving to sequester or present growth factors to cells in appropriate conformations [33]. Matrix remodeling and cell signaling are also influenced by proteolytic activity, particularly via activation of matrix metalloproteases (MMPs) and associated regulatory proteins (TIMPs), which are key facilitators of ECM turnover and release of soluble signaling factors [34].
2.2. ECM Dysfunction in Chronic Wounds
The collective result of these coordinated remodeling processes is the generation of replacement tissue in which the primary ECM components are types I and III collagen [6]. Although healed skin never reaches the full integrity of uninjured tissue, successful wound healing produces a stratified structure that sufficiently replaces the physiological function and mechanical integrity of uninjured skin tissue [35]. In contrast, failure of the wound healing process is associated with a number of chronic conditions, including obesity, diabetes, and peripheral vascular disease, many of which disproportionately affect aging populations [36, 37]. As a result, chronic and non-healing wounds are a widespread public health burden affecting an estimated 20 million people worldwide [38]. While the underlying etiologies of chronic wounds are complex and multifactorial, dysfunctional ECM remodeling may contribute to a number of wound healing pathologies. Long-term (> 20 years) diabetes is associated with increased glycation of dermal collagen [39]. In vitro investigations into downstream effects of collagen glycation have identified changes in fibroblast remodeling behavior, including decreased fibronectin matrix deposition and collagen gel contraction [40], as well as decreased proliferative capacity in endothelial cells [41]. Abnormal quantities and distribution of numerous ECM components have also been observed in histological sections taken from diabetic and venous ulcers in human patients [42, 43].
As impaired matrix remodeling has been associated with wound healing defects, cell-assembled matrices are emerging as valuable tools to treat non-healing wounds. Cell-remodeled collagen scaffolds were among the first engineered biomaterials to receive FDA approval and are still in use in today’s clinical environments [2, 44]. Critically, investigations of donor cell persistence within such materials suggest that donor cells are not maintained past 4 weeks post-transplantation [45]. This suggests that mechanisms by which cell-embedded biomaterials facilitate healing are not dependent on the persistence of donor cells, but rather the ability of cell-remodeled matrices to support host cell infiltration and subsequent healing [3]. The broad utility of cell-derived ECM as a regenerative template is further exemplified by the use of decellularized xenogenic or allogenic ECMs for a diverse array of regenerative medicine applications [44, 46]. In these applications, tissues such as porcine small intestinal submucosa or cadaveric tissue are decellularized, sterilized, and lyophilized to prepare acellular scaffolds comprised of a heterogeneous mixture of ECM proteins [47], glycosaminoglycans [48], and growth factors [49]. Decellularized ECM scaffolds are either in clinical use or under investigation as therapeutics for a broad spectrum of regenerative medicine applications, including vascular, urinary, skin, and nerve reconstruction [8, 46].
In summary, the ECM performs key structural and signaling functions throughout the wound healing process. Cell-mediated ECM remodeling via mechanical contraction, proteolysis, and matrix deposition directs cell signaling through exposure of neoepitopes, growth factor release, and topographic cues. The unique functional contributions of each of these components varies among tissues, injuries, and disease states. Yet, the versatility of decellularized tissue as a regenerative biomaterial suggests that many of these mechanisms are common to a variety of wound healing and regeneration processes. As such, incorporating key structural and functional characteristics of cell-remodeled ECMs into the design of therapeutic biomaterials is a promising strategy for facilitating tissue repair.
3. Biological Effects of Ultrasound
Biomedical ultrasound has been employed for a variety of diagnostic and therapeutic applications. Ultrasound is defined as sound with frequencies above the upper limit of human hearing (20 kHz), with current FDA-approved ultrasound devices operating up to 20 MHz [11–13]. Higher frequency devices are also used for a variety of applications, including acoustic microscopy [12]. Diagnostic ultrasound has an unparalleled safety record in comparison with other imaging modalities, many of which rely on ionizing radiation or hazardous contrast agents to acquire images [50, 51]. Extensive work devoted to understanding the interaction of sound waves with tissue, and the potential for biological effects, has been essential to establishing safety guidelines for diagnostic applications of ultrasound in clinical settings [51].
A growing number of ultrasound applications employ the deliberate induction of ultrasound bioeffects for therapeutic benefit. These techniques include well-established therapies such as shock wave lithotripsy, as well as non-invasive surgery using high-intensity focused ultrasound (HIFU) fields [11]. A new and rapidly expanding application is the use of ultrasound for tissue engineering and regenerative medicine. Ultrasound-based tissue engineering technologies have already yielded a number of innovative approaches for using ultrasound to organize cells, proteins and microparticles within in vitro environments [13]. In the following section, a brief overview of the known biological effects of ultrasound is presented. Several representative examples then follow to illustrate the wide range of strategies that have been employed to harness ultrasound for the purposes of producing biomaterials with enhanced functionality.
3.1. Sound Propagation Through Tissue
Propagation of sound waves through a medium results in a decrease in acoustic amplitude due to acoustic attenuation. In tissue, sound attenuation occurs via both scattering of sound by heterogeneous tissue structures, as well as direct absorption of acoustic energy [52, 53]. The attenuation coefficient (α) describes the rate at which sound energy is lost in a material over a given propagation distance and is dependent upon the frequency of the sound field. Typical values for the acoustic attenuation coefficient in solid tissue at 1 MHz frequency range from approximately 1 dB/cm in some soft tissues to upwards of 10 dB/cm for highly attenuating tissues such as bone [54]. Absorption of sound by tissue arises from the absorption behavior of its biochemical components, as well as macromolecular interactions between structures such as cells and proteins [55, 56]. The absorption properties of many proteins in dilute solutions have been characterized, and generally increase as a function of protein concentration, acoustic frequency [57, 58], and the extent of chemical crosslinking and other intermolecular interactions [55, 56].
As the most abundant protein in the body, the acoustic properties of collagen are of particular relevance for understanding the acoustic properties of tissue. However, technical limitations arising from the high viscosity and self-assembly capacity of purified collagen solutions have made quantitative characterization of collagen at physiologically-relevant concentrations infeasible [58]. Characterization studies of dilute collagen suspensions in acidic solution have demonstrated that collagen exhibits relatively high absorption in comparison to globular proteins at comparable concentrations and acoustic frequencies [58, 59]. Further, the fibrillar structure of collagen is thought to be a contributor to the acoustic scattering behavior of solid tissues [52, 60]. Recent work has demonstrated that changes in collagen fiber structure, due to concentration and polymerization temperature, influence the amount of backscattered acoustic signal in quantitative ultrasound imaging applications [61]. These results are consistent with reports of increased acoustic attenuation as a result of chemical crosslinking [56], however detailed investigations of how the acoustic properties of collagen change as fibers are formed have not been published.
3.2. Acoustic Mechanisms of Ultrasound-Induced Bioeffects
Absorption of sound in tissues can result in several biological effects with clinical relevance. First, significant heating may arise from the absorption of acoustic energy, and the extent of heating is dependent upon the acoustic exposure intensity, acoustic frequency, as well as the absorption properties of the propagation medium [53]. In therapeutic applications, HIFU fields have been used as a non-invasive surgical technique, in which acoustic beam forming methods selectively heat a small focal area without damage to the intervening tissue [11]. HIFU treatments have been FDA-approved for ablation of uterine leiomyomas, bone metastases, and prostate cancer, with many additional applications currently in different stages of clinical trials [62].
Sound propagation through a medium is also associated with a radiation pressure in the direction of acoustic propagation [63]. In solid materials, this pressure results in the generation of an acoustic radiation force and is of particular interest to ultrasound elastography applications, such as acoustic radiation force impulse (ARFI) imaging [64]. In fluids, the same radiation pressure results in fluid flow in the direction of acoustic propagation, referred to as acoustic streaming [65]. In experimental and simulation studies of streaming within cylindrical wells, such as those found in typical tissue culture systems, this includes the generation of cylindrical vortices around the axis of acoustic propagation [66]. Ultrasound-driven fluid streaming has been used to enhance fluid mixing in a variety of applications, including in vitro biological culture systems [67, 68].
In addition to radiation forces associated with travelling wave fields, interaction of multiple waves can result in the generation of ultrasound standing wave fields (USWF), which are characterized by stable regions of zero pressure (nodes) and pressure maxima (antinodes) that result from interference patterns between interacting fields [69]. Particles suspended within a standing wave field are subject to radiation forces, causing particles to cluster at either nodal or antinodal locations [70]. Radiation forces associated with USWFs are the primary mechanism for a number of applications in which ultrasound has been used to distribute particles, including cells and microbubbles into pre-defined patterns [13]. The use of ultrasound to non-invasively pattern cells is of growing interest to tissue engineering, as spatial cues such as relative position, spacing, and density of cells serve as important determinants of cellular behavior [71–73]. USWF exposures have been used to pattern a variety of cell types, including fibroblasts, endothelial cells, Schwann cells, and myocytes to produce enhanced collagen gel contraction [74], vascular network formation [75–77], and cellular alignment [78–81], respectively.
In addition to absorption-dependent mechanisms of ultrasound bioeffects, pressure oscillations associated with acoustic waves can cause expansion and compression of gas bubbles within both fluids and tissues [82, 83]. Bubble oscillations of small amplitude around their equilibrium size are known as stable (or non-inertial) cavitation [84, 85]. Stabilized microbubbles are widely used to enhance contrast during ultrasound imaging, particularly in cardiovascular applications [86]. In contrast, ultrasound-induced expansion of a gas bubble to several times its initial radius can cause inertial collapse of the microbubble. This effect, known as inertial or transient cavitation, produces extremely high pressures, temperatures, and fluid velocities with the potential to damage biological structures [83, 87]. Numerous applications of inertial cavitation are also under investigation for therapeutic applications, including drug delivery, clot lysis, and gene transfection through sonoporation techniques [88].
4. Acoustic Manipulation of ECM-Based Scaffolds
The ability of ultrasound to exert non-invasive, spatially- and temporally-localized effects within biological systems makes it an attractive tool for tissue engineering [13]. In particular, native protein components of the ECM are inherently sensitive to both temperature and mechanical forces [89–91], both of which can be induced non-invasively by ultrasound exposures of sufficient intensity [13, 92]. The versatility of ultrasound-mediated effects on ECM-based scaffolds has been furthered by the incorporation of thermal- or cavitation-responsive elements into ECM-based scaffolds to provide acoustic sensitivity. Finally, within the context of cell-embedded biomaterials, ultrasound may also indirectly influence ECM structure and function by stimulating cell behaviors involved in ECM remodeling. Together, these ultrasound-based technologies hold great potential for optimizing and customizing ECM-based biomaterials for regenerative medicine by non-invasively and site-specifically tuning their mechanical, chemical, and biological properties (Figure 1). In the remainder of this review, we discuss these approaches, with representative examples highlighted in Table 1.
Figure 1.
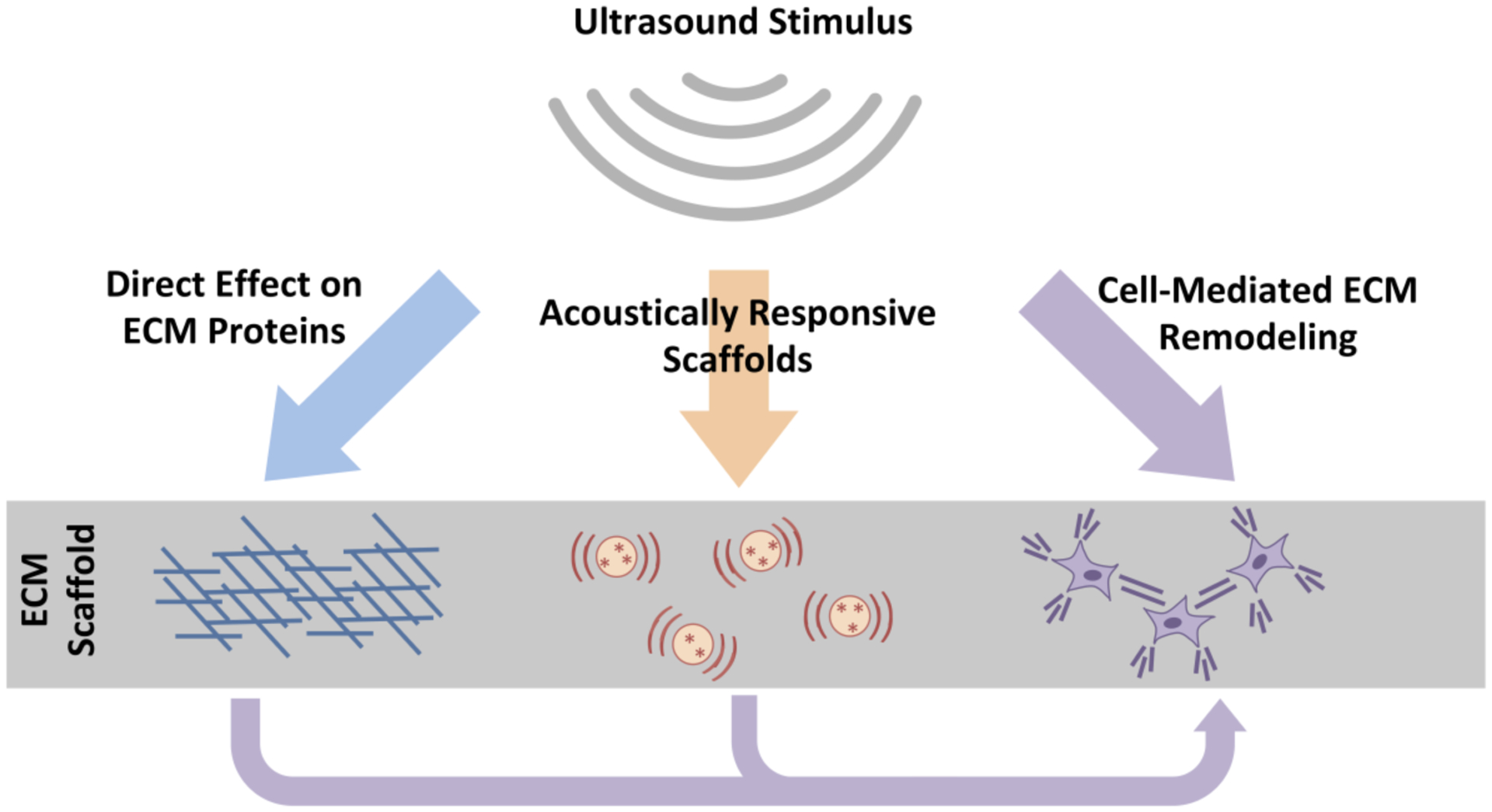
Mechanisms of Ultrasound-ECM Interactions. Ultrasound exposure parameters can be tuned to affect ECM proteins directly via heating, radiation force, or microbubble cavitation. Scaffolds can also be engineered with acoustically responsive elements for drug and protein delivery with spatial and temporal resolution. Additionally, mechanical forces associated with ultrasound may induce cell remodeling activity directly, or indirectly via changes in ECM protein composition or conformation.
Table 1.
Examples of acoustic modification of ECM scaffold structure and function. Abbreviations: CW, continuous wave; USWF, ultrasound standing wave field; ISPPA, spatial peak, pulse average intensity; ISPTA, spatial peak, temporal average intensity; PFC, perfluorocarbon; rtPA, recombinant tissue plasminogen activator.
| ECM/US Interaction | Acoustic Conditions | Acoustic Mechanism | Scaffold Composition | Biological Response | Reference |
|---|---|---|---|---|---|
| Direct effect on ECM proteins | 8 MHz, CW ISPPA ≤ 10 W/cm2 | Thermal and non-thermal | Collagen and fibronectin | Fiber alignment, fibronectin fibril formation; Fibroblast self-assembly | [93] |
| Direct effect on ECM proteins | 1 MHz, CW 1 – 8 W/cm2 | Cavitation | Fibrin and rtPA | Decreased fiber density, enhanced proteolysis | [94, 95] |
| Direct effect on ECM proteins | Benchtop sonicator | Cavitation | Decellularized tendon | Decreased fiber density and increased pore size; Enhanced cellular infiltration | [96] |
| Acoustically responsive scaffold | 20kHz, pulsed | Cavitation | Ca2+-spiked liposomes in transglutaminase-fibrinogen solution | Transglutaminase-triggered fibrinogen polymerization | [97] |
| Acoustically responsive scaffold | 3.5 MHz, pulsed MPa +(−) = 12.9 [6.0] | Cavitation | PFC-growth factor emulsions embedded in fibrin | Increased pore size, increased stiffness; Growth factor release | [98, 99] |
| Acoustically responsive scaffold | 2.5 MHz, CW ISPTA = 658–750 W/cm2 | Thermal | Cell-embedded fibrin (+/− hydroxyapatite) | Growth factor expression | [100, 101] |
| Cellular ECM Remodeling | 2–2.2 MHz, USWF 0.12 MPa | USWF | Collagen | Collagen contraction; Myoblast alignment along tensional axis | [80] |
| Cellular ECM Remodeling | 1 MHz, USWF 0.2MPa | USWF | Cell-embedded collagen | Enhanced gel contraction; Fiber alignment along vascular sprouts | [74, 75] |
| Direct effect on ECM proteins; Cellular ECM Remodeling | 8 MHz, CW ISPPA ≤ 10 W/cm2 | Non-thermal | Collagen | Altered pore density and radial fiber alignment; Fibroblast migration and ECM contraction | [102–104] |
4.1. Direct Effects of Ultrasound on ECM Proteins
Early investigations into effects of ultrasound on ECM structure and function demonstrated that exposing fibrin clots to ultrasound (0.2 – 4 MHz, 1–8 W/cm2) in the presence of proteolytic enzymes (i.e., tissue plasminogen activator, tPA) accelerated the rate of fibrin degradation [94, 105]. Subsequent mechanistic investigations demonstrated that ultrasound did not fragment fibrin scaffolds directly [94]. Rather, in the presence of ultrasound, fibrin fibers were separated into strands of reduced diameter and local density [95]. While the overall fiber structure of the fibrin gels returned to its pre-exposure state once ultrasound was removed, the transient reduction in fiber density was sufficient to accelerate proteolytic degradation [94, 95]. The primary acoustic mechanism by which ultrasound influenced fibrin fiber structure was thought to be cavitation, although heating and fluid streaming mechanisms may have also contributed to clot degradation [94, 106]. This work was among the first to demonstrate that ultrasound can be used to manipulate the structure of ECM proteins. Related techniques are under active investigation as therapeutic strategies to accelerate thrombolysis in animal models of ischemic stroke [107, 108].
Type I collagen is the most abundant protein in the human body, contributing to the ECMs of a variety of tissues, including skin, tendon, cornea, and bone [109] Collagen’s high abundance low antigenicity, and versatility have made it a promising starting material for the production of tissue-engineered scaffold structures in a variety of applications [110]. Under appropriate conditions in vitro, solubilized collagen can spontaneously self-assemble into 3D hydrogels, providing a valuable platform for regenerative medicine applications [44, 111]. Self-assembled collagen fibers mimic many of the features of native collagen ECM structures, including fiber diameter and periodicity [112]. However, 3D hydrogels assembled from purified collagen differ from native ECM collagen in several critical ways, including an absence of tissue-specific fiber structures and co-assembly with other ECM components [113, 114]. For this reason, numerous techniques for manipulating the structure and function of collagen within in vitro environments have been explored. Collagen polymerization parameters, including concentration, pH, and temperature, have been widely exploited to manipulate scaffold characteristics such as fiber diameter [91], pore size [115], and gel stiffness [116], and have been reviewed previously [110]. Collagen fiber characteristics directly influence cell behaviors important to wound healing, including cell spreading and adhesion, as well as migration and collagen fiber contraction [29, 115, 117, 118]. Several techniques have been developed to manipulate the hierarchical, macromolecular organization of collagen fibers within 3D hydrogels. These include the use of fluid flow [119], mechanical tension [120], electrospinning [121], and magnetic fields [122, 123] to produce aligned fibers within 3D collagen matrices. Other approaches have used PDMS or other non-adhesive molds to produce micron-scale microchannels within collagen gel structures [124].
The use of ultrasound to directly and non-invasively manipulate the structure and function of collagen hydrogels was first described by Garvin et al. [102]. This work demonstrated that ultrasound exposure (1- or 8.3-MHz, with acoustic intensities up to 30 W/cm2) during collagen polymerization could produce local changes in collagen fiber microstructure, including increased collagen fiber density and reduced fiber diameter [102]. Measurements of ultrasound-induced heating within the center of polymerizing gels demonstrated temperature increases of up to 10 °C above unexposed sham samples [102]. Ultrasound-induced changes in collagen fiber structure could be mimicked using a non-acoustic heat source, suggesting a thermal acoustic mechanism [102]. This conclusion is consistent with previous reports that elevated temperature during collagen polymerization is associated with thinner, more densely packed collagen fiber structures [91, 115]. One advantage of ultrasound is that local heating can be produced non-invasively and site specifically, thereby providing avenues for fabricating collagen hydrogels with spatial control of collagen fiber structure. In addition to thermal effects of ultrasound, acoustic exposure during collagen polymerization can produce collagen fiber alignment through non-thermal mechanisms [103]. This effect was only observed when ultrasound was present during the active phase of collagen fibril self-assembly [104]. The pattern of ultrasound-mediated collagen fiber alignment was consistent with simulation patterns of fluid flow within ultrasound-exposed cylindrical vessels [66, 68], and thus resembles other systems in which fluid streaming is induced by non-acoustic mechanisms, such as flow through microchannels [119].
Other groups have used laboratory benchtop sonicators to partially disrupt ECM structures. These devices operate at comparatively low frequencies (15–30 kHz), and high power (≥ 150 W total power) to induce acoustic cavitation within a fluid volume, and have been used to facilitate surface cleaning, chemical catalysis, and cell lysis [125, 126]. Examples in which benchtop sonicators have been used for regenerative medicine applications include sonication of detergent-decellularized porcine tendon, which produced fiber separation within the collagenous ECM structure [96]. Sonicated tendon scaffolds supported enhanced cellular infiltration, but did not support long-term viability of cells embedded within the center of the construct [96]. Likewise, Maller and colleagues utilized a model of sonicated type I collagen in combination with other ECM components to investigate effects on cellular morphology and integrin activation in mammary tumor cells [127]. In this study, volumes of soluble collagen were sonicated until the detection of fibers by second harmonic generation (SHG) imaging was eliminated, indicating unfolding or fragmentation of the collagen triple helical structure [127, 128]. Both examples demonstrate the feasibility of influencing collagen structure and function with ultrasound-induced bioeffects. However, typical benchtop sonicators do not offer precise control over acoustic parameters and vary among manufacturers, thereby limiting both optimization and reproducibility.
Finally, many ECM proteins contain regions that undergo conformational changes in response to temperature or mechanical force, thereby exposing new epitopes for cell and/or protein engagement [26, 89, 129]. A key example of this behavior is fibronectin, a large molecular weight glycoprotein whose incorporation into the fibrillar ECM requires application of cell-derived mechanical forces and exposure of cryptic, self-interacting epitopes [26, 130]. Interactions of fibronectin with other ECM components, including collagen, are also influenced by conformational changes that arise as a result of temperature [89] or the application of mechanical force [131]. Recent work describing the polymerization of collagen-fibronectin composite gels suggests that ultrasound exposure can trigger fibronectin fibril assembly at hydrogel surfaces [93]. Effects were observed only in the presence of ultrasound under permissive temperature conditions, suggesting that a combination of thermal and non-thermal effects of ultrasound are involved in triggering fibril formation within engineered hydrogels [93]. Collagen-fibronectin binding is one of many examples in which interactions among ECM components co-regulate downstream functions [132–134]. As investigations into effects of ultrasound expand to other composite ECM materials [101], additional examples of how ultrasound can be used to influence interactions among ECM components are likely to emerge.
4.2. Acoustically-Responsive Engineered Scaffolds
The development of novel techniques to enhance the susceptibility of biological systems to acoustic effects has expanded the versatility of ultrasound across both diagnostic and therapeutic domains. In particular, advances in microbubble chemistry have enabled the use of acoustically-responsive droplets as vehicles for the delivery of drugs, including recombinant proteins [135], small molecules [136], and genetic material [137]. In these systems, which vary among applications in their composition and preparation, a therapeutic payload is suspended in a cavitation-sensitive material, often gas or perfluorocarbons, and encapsulated within a stabilizing shell [138]. The development and potential applications of acoustically-responsive droplets has been reviewed recently [138, 139]. Here, we discuss specific examples of relevance to ECM-based scaffolds and within engineered environments.
A key advantage of acoustically-triggered release is its capacity to simultaneously enable spatial and temporal control over drug release [138]. Nele et al. recently reported using ultrasound to trigger calcium release from acoustically responsive liposomes to initiate fibrin hydrogel polymerization [97]. In this system, ultrasound-induced calcium release was used to activate transglutaminase activity, which in turn triggered catalysis of soluble fibrinogen to initiate hydrogel crosslinking [97]. The kinetics of calcium release and enzymatic activity, as well as the elastic modulus of the polymerized fibrin hydrogel, were sensitive to acoustic exposure duration [97]. Thus, ultrasound-induced calcium release simultaneously enabled hydrogel assembly and tuning of hydrogel mechanical properties. Many components of ECM signaling networks are sensitive to calcium, including matrix metalloprotease activation [140], integrin-mediated adhesion [141], and interactions between ECM proteins [142]. As such, techniques that take advantage of ultrasound-triggered calcium release to initiate enzymatic activity or cell signaling cascades in temporally- and spatially-defined patterns within ECM-based scaffolds are likely to expand.
Native ECM also serve as reservoirs for localized growth factor release [33]. The use of acoustically responsive droplets to mimic controlled release of growth factors has been demonstrated in fibrin gels. With this technique, growth factors are encapsulated into acoustically responsive droplets (<10 μm) which are then embedded in fibrin-based hydrogels. Exposing fibrin-embedded, basic fibroblast growth factor (bFGF)-loaded droplets to ultrasound triggered growth factor release into the surrounding media, and treatment of endothelial cells with this conditioned media enhanced cell viability [98] and vascular sprout formation [143]. Additionally, ultrasound-induced cavitation increased the porosity and stiffness of these fibrin scaffolds [98]. When similar constructs were loaded with a fluorescent tracer and implanted subcutaneously in mice, acoustic exposure supported accelerated payload release [99]. The kinetics of release were sensitive to acoustic pressure, with the composition [144] and concentration [145] of perfluorocarbons used in manufacture of acoustically responsive droplets having a significant influence on the threshold pressure at which cavitation and drug release were triggered. This technology has also been proposed as a method for sequential release of paired growth factors [146].
An alternative method to achieve ultrasound-triggered growth factor release has been to genetically engineer cells with heat-responsive gene elements. This technique exploits a well-characterized transcriptional response to heat shock [147], in which cells are transfected with plasmids containing a target gene sequence under the combined control of the HSP70B promoter and a rapamycin-dependent gene switch [148]. Thus, when engineered cells are cultured in the presence of rapamycin and exposed to high-intensity focused ultrasound (HIFU), localized heating induces expression of target genes with spatial and temporal specificity [100]. This approach has been used to induce expression of both BMP-2 [100] and VEGF [148], key growth factors for therapeutic bone fracture healing and angiogenesis, respectively [149]. Further tuning of growth factor expression has been achieved by addition of hydroxyapatite to cell-embedded fibrin gels to enhance acoustic absorption [101]. The translation potential of this approach has been demonstrated in vivo using ultrasound-induced luciferase expression in murine models [101]. Critically, the acoustic mechanisms used to induce growth factor release with heat-shock induction systems, namely absorption-dependent heating [100], are distinct from the cavitation-dependent release of growth factors from acoustically-responsive lipid droplets [98]. Thus, the secondary effects on surrounding native ECM components will likely be distinct, enabling an additional level of control in engineering custom scaffolds for specific applications.
4.3. Indirect Effects of Ultrasound on Cell-Mediated ECM Remodeling
A critical consideration in the fabrication of tissue engineered scaffolds is an understanding of the bi-directional relationship between the ECM and its resident cells. Within this framework, known as dynamic reciprocity, structural changes in the ECM induce changes in cell behaviors, which in turn remodel the ECM [150]. Key cellular programs associated with ECM remodeling include enhanced actin-myosin contractility and cell migration, as well as upregulation of ECM gene expression and matrix protein deposition [7]. The strategic use of intrinsic ECM-remodeling capabilities of cells has emerged as an essential tool for biomaterial fabrication. Examples include the use of geometric patterning and cell-derived tensile forces to achieve fiber alignment [27, 151, 152], and the use of tissue-specific cell types to assemble matrices with appropriate molecular composition [153, 154]. In combination, these techniques have been used to reconstruct complex biomaterials mimicking numerous features of tissue-specific ECMs, with particular utility in loadbearing tissues such as tendon [155], intervertebral discs [156] and menisci of the knee [157, 158].
Contraction of fibroblast-embedded collagen hydrogels and associated changes in cell migration and differentiation is a long-standing in vitro model for investigations of ECM remodeling in a wound-like environment [25]. Studies of fibroblast-embedded collagen hydrogels exposed to ultrasound standing wave fields during collagen polymerization showed that ultrasound exposure enhanced cell-mediated gel contraction compared to sham-exposed controls [74]. Subsequent studies demonstrated that exposing soluble collagen to ultrasound during hydrogel polymerization produces collagen fibrils that are more readily remodeled by cells than sham-exposed fibrils [102, 103]. Similar collagen remodeling was observed when primary cells derived from diabetic murine dermal explants were seeded on acoustically modified collagen hydrogels [103]. In these studies, cell-mediated collagen fibril remodeling and contraction were likely facilitated by ultrasound-induced changes in collagen structure [102, 103]. Given the role of ECM remodeling in the coordinated healing response of skin to injury [25, 159], these results raise the possibility that ultrasound-based techniques may improve current fabrication technologies for regenerative biomaterials by enhancing the healing capacity of acellular, collagen-based wound dressings.
The use of ultrasound standing wave fields as a technology to pattern cells in vitro has also emerged as a versatile technique for engineering cellular biomaterials, with applications to vascular [75–77], muscular [80, 81], and neuronal [78, 79] tissue engineering. In these systems, secondary radiation forces associated with ultrasound standing waves are used to pattern cells into planar bands or columns within solutions of soluble ECM proteins, often collagen [75] or fibrinogen [79]. Following hydrogel polymerization, cells are retained in the ultrasound-established pattern after the acoustic source is removed [160]. The majority of these studies have focused on cellular responses to acoustic patterning, including cellular morphogenesis [76] and differentiation [79]. Several pieces of evidence suggest that enhanced remodeling of the initial ECM template also contributes to cellular responses. First, vascular sprouting and collagen fiber alignment along the direction of vessel growth have been observed in acoustically-patterned systems containing vascular endothelial cells [75], consistent with observations of sprouting angiogenesis from microvessel explants [161]. Likewise, enhanced contraction of acoustically-patterned constructs has been reported in several studies [74, 80]. Critically, the direction of cell alignment can be directed to develop either parallel or perpendicular to the acoustic exposure axis by altering the orientation in which the engineered tissue is anchored [80]. Taken together, these findings suggest that the expression of tissue-mimicking cellular morphologies may arise from an enhanced capability of resident cells to remodeling the ultrasound-exposed scaffold.
5. Emerging Technologies
As technologies to manipulate biological systems advance, the uses of ultrasound in the fabrication of ECM-based scaffolds is also likely to expand. Many ultrasound properties are highly attractive for regenerative medicine applications, particularly the ability to be used non-invasively and with high temporal and spatial control. In this regard, ultrasound is one of several emerging modalities by which biomaterials, as well as cells and other components of the tissue-engineered environment, can be manipulated non-invasively [10]. Given the complexity of native tissues, producing engineered tissue substitutes with sufficient fidelity to restore healthy function in patients with severe disease and injury will likely require the synergy of multiple strategies and technologies. The use of tissue-specific cells to further remodel acoustically-modified ECM templates is one such example of this approach that has already generated promising preliminary success [75, 80, 103]. Here, we discuss additional examples in which ultrasound-based methodologies may extend the potential of other experimental strategies.
One technology of interest is the development of acoustic tweezing cytometry (ATC) to apply mechanical forces to cells or proteins. Pioneering work in this field demonstrated the feasibility of targeting microbubbles to the surface of cells via attachment of an integrin-binding peptide ligand to the microbubble shell [162, 163]. Exposure of microbubble-targeted cells to ultrasound induced microbubble displacement was followed by sustained generation of cellular traction forces [162, 163]. Cytoskeletal contraction required Rho- and Rho-associated kinase (ROCK)-mediated signaling [163], and was not observed when microbubbles were targeted to a non-integrin receptor [162]. Assembly of fibronectin fibrils, a key event during tissue repair [3, 23] is likewise dependent upon integrin ligation and Rho-mediated cytoskeletal contractility [130]. Thus, these studies suggest the possibility that ATC-stimulated cellular contractility may offer yet another technique to manipulate cellular ECM deposition and/or force-sensitive conformational changes to ECM proteins.
Another area of expanding potential is the use of acoustic techniques to assemble scaffold materials into increasingly complex patterns with high precision. The use of acoustic holograms, in which sound passes through a 3D-printed phase plate to generate an acoustic pressure field in a user-defined pattern, has emerged recently as a significant advance in the complexity of structures that can be achieved using ultrasound-mediated particle patterning [164]. This approach has been used to pattern cells [165] and PEG-DMA scaffolds [166] into complex geometries. The use of ultrasound to pattern PEG-based scaffolds is of particular interest to the engineering of biocompatible scaffolds, as PEG hydrogels can be functionalized with appropriate ligands to engineer specific ECM compositions and conformations [167]. Such an approach has the potential to enable the incorporation of a diverse array of bioactive signals [168], including adhesive [169, 170] and matricryptic [171] ligands, as well as enzymatically-sensitive [172] and growth factor-simulating [170, 172] sequences. Other technologies use ultrasound beam steering and vortex beams to move and steer particles in 3D through fluid materials [173]. Together, these technologies may be used in coordination to create engineered matrices of high spatial and biochemical complexity.
6. Conclusions
In summary, ultrasound is a versatile tool for manipulating biological systems and continues to find novel applications in the fabrication of biomaterials for regenerative medicine applications. Ultrasound has the capacity to induce several distinct effects within biological systems, including heating, fluid streaming, and microbubble cavitation. Numerous ultrasound-based techniques that have the capacity to manipulate biomaterials have emerged in recent years. These include the use of ultrasound to modify the conformation and organization of fibrillar ECM proteins either directly or indirectly, as well as innovative strategies to use ultrasound to enhance cellular activities within engineered scaffolds. The convergence of these developments with an evolving understanding of the role of ECM during tissue repair represents a significant opportunity to harness ultrasound as a non-invasive methodology for fabricating ECM-based scaffolds with enhanced complexity and regenerative capacity.
Funding
This research was supported by National Institutes of Health (NIH) Grants R21 GM131281 and R01 AG058746. E.G.N received support from the John R. Murlin Memorial Fund through the Department of Pharmacology and Physiology at the University of Rochester.
Footnotes
Competing Interests
The authors have declared that no competing interests exist.
References
- 1.Langer R, Vacanti JP. Tissue Eng. Science 1993; 260: 920–926. [DOI] [PubMed] [Google Scholar]
- 2.Place ES, Evans ND, Stevens MM. Complexity in biomaterials for Tissue Eng. Nat Mater 2009; 8: 457–470. [DOI] [PubMed] [Google Scholar]
- 3.Clark RA, Ghosh K, Tonnesen MG. Tissue Eng for cutaneous wounds. J Invest Dermatol. 2007; 127: 1018–1029. [DOI] [PubMed] [Google Scholar]
- 4.Borenstein JT, Vunjak-Novakovic G. Engineering tissue with BioMEMS. IEEE Pulse. 2011; 2: 28–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Rozario T, DeSimone DW. The extracellular matrix in development and morphogenesis: A dynamic view. Dev Biol. 2010; 341: 126–140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Singer AJ, Clark RA. Cutaneous wound healing. New Engl J Med. 1999; 341: 738–746. [DOI] [PubMed] [Google Scholar]
- 7.Lu P, Takai K, Weaver VM, Werb Z. Extracellular matrix degradation and remodeling in development and disease. Cold Spring Harb Perspect Biol. 2011; 3: a005058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gu X, Ding F, Williams DF. Neural Tissue Eng options for peripheral nerve regeneration. Biomaterials. 2014; 35: 6143–6156. [DOI] [PubMed] [Google Scholar]
- 9.Demidova-Rice TN, Hamblin MR, Herman IM. Acute and impaired wound healing: Pathophysiology and current methods for drug delivery, part 1: Normal and chronic wounds: Biology, causes, and approaches to care. Adv Skin Wound Care. 2012; 25: 304–314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Armstrong JP, Stevens MM. Using remote fields for complex Tissue Eng. Trends Biotechnol. 2020; 38: 254–263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Miller DL, Smith NB, Bailey MR, Czarnota GJ, Hynynen K, Makin IR, et al. Overview of therapeutic ultrasound applications and safety considerations. J Ultras Med 2012; 31: 623–634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Shung KK. High frequency ultrasound imaging. J Med Ultrasound. 2009; 17: 25–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Dalecki D, Hocking DC. Ultrasound technologies for biomaterials fabrication and imaging. Ann Biomed Eng. 2015; 43: 747–761. [DOI] [PubMed] [Google Scholar]
- 14.Deng CX, Hong X, Stegemann JP. Ultrasound imaging techniques for spatiotemporal characterizaiton of composition, microstructure, and mechanical properties in Tissue Eng. Tissue Eng Part B Rev. 2016; 22: 311–321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Daley WP, Peters SB, Larsen M. Extracellular matrix dynamics in development and regenerative medicine. J Cell Sci. 2008; 121: 255–264. [DOI] [PubMed] [Google Scholar]
- 16.Sundelacruz S, Kaplan DL. Stem cell- and scaffold-based Tissue Eng approaches to osteochondral regenerative medicine. Semin Cell Dev Biol. 2009; 20: 646–655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Setayeshmehr M, Esfandiari E, Rafieninia M, Hashemibeni B, Taheri-Kafrani A, Samadikuchaksaraei A, et al. Hybrid and composite scaffolds based on extracellular matrices for cartilage Tissue Eng. Tissue Eng Part B Rev. 2019; 25: 202–224. [DOI] [PubMed] [Google Scholar]
- 18.Voleti PB, Buckley MR, Soslowsky LJ. Tendon healing: Repair and regeneration. Annu Rev Biomed Eng. 2012; 14: 47–71. [DOI] [PubMed] [Google Scholar]
- 19.Ruberti JW, Zieske JD. Prelude to corneal Tissue Eng - gaining control of collagen organization. Prog Retin Eye Res. 2008; 27: 549–577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Fuoco C, Petrilli LL, Cannata S, Gargioli C. Matrix scaffolding for stem cell guidance toward skeletal muscle Tissue Eng. J Orthop Surg Res. 2016; 11: 86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Mukhatyar V, Karumbaiah L, Yeh J, Bellamokonda R. Tissue Eng strategies designed to facilitate the endogenous regenerative potential or peripheral nerves. Adv Mater. 2009; 21: 4670–4679. [Google Scholar]
- 22.Urciuolo F, Casale C, Imparato G, Netti PA. Bioengineered skin substitutes: The role of extracellular matrix and vascularization in the healing of deep wounds. J Clin Med. 2019; 8: 2083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Clark RA, Lanigan JM, DellaPelle P, Manseau E, Dvorak HF, Colvin RB. Fibronectin and fibrin provide a provisional matrix for epidermal cell migration during wound reepithelializaiton. J Invest Dermatol. 1982; 79: 264–269. [DOI] [PubMed] [Google Scholar]
- 24.Grinnell FR. Wound repair, keratinocyte activation and integrin modulation. J Cell Sci. 1992; 101: 1–5. [DOI] [PubMed] [Google Scholar]
- 25.Grinnell F, Petroll WM. Cell motility and mechanics in three-dimensional collagen matrices. Annu Rev Cell Dev Biol. 2010; 26: 335–361. [DOI] [PubMed] [Google Scholar]
- 26.Zhong C, Chrzanowska-Wodnicka M, Brown J, Shaub A, Belkin AM, Burridge K. Rho-mediated contractility exposes a cryptic site in fibronecitn and induces fibronectin matrix assembly. J Cell Biol. 1998; 141: 539–551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Provenzano PP, Inman DR, Eliceiri KW, Trier SM, Keely PJ. Contact guidance mediated three-dimensional cell migration is regulated by Rho/ROCK-dependent matrix reorganization. Biophys J. 2008; 95: 5374–5384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Gildner CD, Lerner AL, Hocking DC. Fibronectin matrix polymerization increases tensile strength of model tissue. Am J Physiol Heart Circ Physiol. 2004; 287: H46–H53. [DOI] [PubMed] [Google Scholar]
- 29.Miron-Mendoza M, Seemann J, Grinnell F. Collagen fibril flow and tissue translocation coupled to fibroblast migration in 3D collagen matrices. Mol Biol Cell. 2008; 19: 2051–2058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Sakar MS, Eyckmans J, Pieters R, Eberli D, Nelson BJ, Chen CS. Cellular forces and matrix assembly coordinate fibrous tissue repair. Nat Commun. 2016; 7: 11036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Pakshir P, Alizadehgiashi M, Wong B, Coelho NM, Chen X, Gong Z, et al. Dynamic fibroblast contractions attract remote macrophages in fibrillar collagen matrix. Nat Commun. 2019; 10: 1850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Korff T, Augustin HG. Tensional forces in fibrillar extracellular matrices control directional capilary sprouting. J Cell Sci. 1999; 112: 3249–3258. [DOI] [PubMed] [Google Scholar]
- 33.Schultz GS, Wysocki A. Interactions between extracellular matrix and growth factors in wound healing. Wound Repair Regen. 2009; 17: 153–162. [DOI] [PubMed] [Google Scholar]
- 34.Caley MP, Martins VL, O Toole EA. Metalloproteinases and wound healing. Adv Wound Care. 2015; 4: 225–234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Levenson SM, Geever EF, Crowley LV, Oates III JF, Berard CW, Rosen H. Healing of rat skin wounds. Ann Surg. 1965; 161: 293–308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Escandon J, Vivas AC, Tang J, Rowland KJ, Kirsner RS. High mortality in patients with chronic wounds. Wound Repair Regen. 2011; 19: 526–528. [DOI] [PubMed] [Google Scholar]
- 37.Sen CK. Human wounds and its burden: An updated compendium of estimates. Adv Wound Care. 2019; 8: 39–48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sen CK, Gordillo GM, Roy S, Kirsner R, Lambert L, Hunt TK, et al. Human skin wounds: A major and snowballing threat to public health and the economy. Wound Repair Regen. 2009; 17: 763–771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Vishwanath V, Frank KE, Elmets CA, Dauchot PJ, Monnier VM. Glycation of skin collagen in type I diabetes mellitus. Correlation with long-term complications. Diabetes. 1986; 35: 916921. [DOI] [PubMed] [Google Scholar]
- 40.Liao H, Zakhaleva J, Chen W. Cells and tissue interactions with glycated collagen and their relevance to delayed diabetic wound healing. Biomaterials. 2009; 30: 1689–1696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kuo PC, Kao CH, Chen JK. Glycated type 1 collagen induces endothelial dysfunction in culture. In Vitro Cell Dev Biol Anim. 2007; 43: 338–343. [DOI] [PubMed] [Google Scholar]
- 42.Loots MA, Lamme EN, Zeegelaar J, Mekkes JR, Bos JD, Middelkoop E. Differences in cellular infiltrate and extracellular matrix of chronic diabetic and venous ulcers versus acute wounds. J Invest Dermatol. 1997; 111: 850–857. [DOI] [PubMed] [Google Scholar]
- 43.Herrick SE, Sloan P, McGurk M, Freak L, McCollum CN, Ferguson MW. Sequential changes in histologic pattern and extracellular matrix deposition during the healing of chronic venous ulcers. Am J Pathol. 1992; 141: 1085–1096. [PMC free article] [PubMed] [Google Scholar]
- 44.Abou Neel EA, Bozec L, Knowles JC, Syed O, Mudera V, Day R, et al. Collagen - emerging collagen based therapies hit the patient. Adv Drug Deliver Rev 2013; 65: 429–456. [DOI] [PubMed] [Google Scholar]
- 45.Phillips TJ, Manzoor J, Rojas A, Isaacs C, Carson P, Sabolinski M, et al. The longevity of a bilayered skin substitute after application to venous ulcers. Arch Dermatol. 2002; 138: 1079–1081. [DOI] [PubMed] [Google Scholar]
- 46.Badylak SF. Xenogenic extracellular matrix as a scaffold for tissue reconstruction. Transpl Immunol. 2004; 12: 367–377. [DOI] [PubMed] [Google Scholar]
- 47.Lindberg K, Badylak SF. Porcine small intestinal submucosa (SIS): A bioscaffold supporting in vitro primary human epidermal cell differentiation and synthesis of basement membrane proteins. Burns. 2001; 27: 254–266. [DOI] [PubMed] [Google Scholar]
- 48.Hodde JP, Badylak SF, Brightman AO, Voytik-Harbin SL. Glycosaminoglycan content of small intesitinal submucosa: A bioscaffold for tissue replacement. Tissue Eng. 1996; 2: 209–217. [DOI] [PubMed] [Google Scholar]
- 49.Voytik-Harbin SL, Brightman AO, Kraine MR, Waisner B, Badylak SF. Identification of extractable growth factors from small intestinal submucosa. J Cell Biochem. 1997; 67: 478–491. [PubMed] [Google Scholar]
- 50.Arthurs OJ, Bjorkum AA. Safety in pediatric imaging: An update. Acta Radiol. 2013; 54: 983990. [DOI] [PubMed] [Google Scholar]
- 51.Zeqiri B Exposure criteria for medical diagnostic ultrasound: II. Criteria based on all known mechanisms:(NCRP Report No. 140) National Council on Radiation Protection and Measurements (NCRP), 2002. Ultrasound Med Biol. 2003; 29: 1809. [Google Scholar]
- 52.Insana MF, Wagner RF, Brown DG, Hall TJ. Describing small-scale structure in random media using pulse-echo ultrasound. J Acoust Soc Am. 1990; 87: 179–192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Fry WJ, Fry RB. Determination of absolute sound levels and acoustic absorption coefficients by thermocouple probes-Theory. J Acoust Soc Am. 1954; 26: 294–310. [Google Scholar]
- 54.Bamber JC. Ultrasonic properties of tissues Ultrasound in medicine. Philadelphia: Institute of Physics Publishing; 1998. p. 57–88. [Google Scholar]
- 55.Pauly H, Schwan HP. Mechanism of absorption of ultrasound in liver tissue. J Acoust Soc Am. 1971; 50: 692–699. [DOI] [PubMed] [Google Scholar]
- 56.Kremkau FW, Carstensen EL, Aldridge WG. Macromolecular interaction in the absorption of ultrasound in fixed erythrocytes. J Acoust Soc Am. 1973; 53: 1448–1451. [DOI] [PubMed] [Google Scholar]
- 57.Carstensen EL, Schwan HP. Acoustic properties of hemoglobin solutions. J Acoust Soc Am. 1959; 31: 305–311. [Google Scholar]
- 58.Goss SA, Dunn F. Ultrasonic propagation properties of collagen. Phys Med Biol. 1980; 25: 827–837. [DOI] [PubMed] [Google Scholar]
- 59.Goss SA, Frizzell LA, Dunn F, Dines KA. Dependence of the ultrasonic properties of biological tissue on constituent proteins. J Acoust Soc Am. 1980; 67: 1041–1044. [Google Scholar]
- 60.Fields S, Dunn F. Correlation of echographic visualizability of tissue with biological composition and physiological state J Acoust Soc Am. 1973; 54: 809–812. [DOI] [PubMed] [Google Scholar]
- 61.Mercado KP, Helguera M, Hocking DC, Dalecki D. Noninvasive quantitative imaging of collagen microstructure in three-dimensional hydrogels using high-frequency ultrasound. Tissue Eng Part C Methods. 2015; 21: 671–682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Elhelf IS, Albahar H, Shah U, Oto A, Cressman E, Almekkawy M. High intensity focused ultrasound: The fundamentals, clinical applications, and research trends. Diagn Interv Imag. 2018; 99: 349–359. [DOI] [PubMed] [Google Scholar]
- 63.Rooney JA, Nyborg WL. Acoustic radiation pressure in a traveling plane wave. Am J Phys 1972; 40: 1825–1830. [Google Scholar]
- 64.Parker KJ, Doyley MM, Rubens DJ. Imaging the elastic properties of tissue: The 20 year perspective. Phys Med Biol. 2011; 56: R1–R29. [DOI] [PubMed] [Google Scholar]
- 65.Starritt HC, Duck FA, Humphrey VF. An experimental investigation of streaming in pulsed diagnostic ultrasound fields. Ultrasound Med Biol. 1989; 15: 363–373. [DOI] [PubMed] [Google Scholar]
- 66.Green A, Marshall JS, Ma D, Wu J. Acoustic streaming and thermal instability of flow generated by ultrasound in a cylindrical container. Phys Fluids. 2016; 28: 104105. [Google Scholar]
- 67.Ma D, Green AM, Willsey GG, Marshall JS, Wargo MJ, Wu J. Effects of acoustic streaming from moderate-intensity pulsed ultrasound for enhancing biofilm mitigation effectiveness of drug-loaded liposomes. J Acoust Soc Am. 2015; 138: 1043–1051. [DOI] [PubMed] [Google Scholar]
- 68.Marshall JS, Wu J. Acoustic streaming, fluid mixing, and particle transport by a Gaussian ultrasound beam in a cylindrical container. Phys Fluids. 2015; 27: 103601. [Google Scholar]
- 69.Halliday D, Resnick R, Walker J. Waves - I. Fundamentals of Physics 9th ed. Hoboken: John Wiley & Sons, Inc.; 2011. p. 413–444. [Google Scholar]
- 70.Gould RK, Coakley WT. The effects of acoustic radiation forces on small particles in suspension Finite-amplitude wave effects in fluids: Proceedings of the 1973 symposium. Guildford: IPC Science and Technology Press Ltd; 1974. p. 252–257. [Google Scholar]
- 71.Comeau ES. Ultrasound standing wave field technology for cell patterning and microvessel network formation in vitro and in situ. Rochester: University of Rochester; 2017. [Google Scholar]
- 72.Ino K, Okochi M, Honda H. Application of magnetic force-based cell patterning for controling cell-cell interactions in angiogenesis. Biotechnol Bioeng. 2009; 102: 882–890. [DOI] [PubMed] [Google Scholar]
- 73.Glen CM, McDevitte TC, Kemp ML. Dynamic intracellular transport modulates the spatial patterning of differentiation during early neural committment. Nat Commun. 2018; 9: 4111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Garvin KA, Hocking DC, Dalecki D. Controlling the spatial organization of cells and extracellular matrix proteins in engineered tissues using ultrasound standing wave fields. Ultrasound Med Biol. 2010; 36: 1919–1932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Garvin KA, Dalecki D, Hocking DC. Vascularization of three-dimensional collagen hydrogels using ultrasound standing wave fields. Ultrasound Med Biol. 2011; 37: 1853–1864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Comeau ES, Hocking DC, Dalecki D. Ultrasound patterning technologies for studying vascular morphogenesis in 3D. J Cell Sci. 2017; 130: 232–242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Kang B, Shin J, Park HJ, Rhyou C, Kang D, Lee SJ, et al. High-resolution acoustophoretic 3D cell patterning to construct functional collateral cylindroids for ischemia therapy. Nat Commun. 2018; 9: 5402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Gesellchen F, Bernassau AL, Dejardin T, Cumming DR, Riehle M. Cell patterning with a heptagon acoustic tweezer - application in neurite guidance. Lab Chip. 2014; 14: 2266–2275. [DOI] [PubMed] [Google Scholar]
- 79.Bouyer C, Chen P, Güven S, Demirtas TT, Nieland TJ, Padilla F, et al. A bio-acoustic levitational (BAL) assembly method for engineering of multilayered, 3D brain-like constructs using human embryonic stem cell derived neuro-progenitors. Adv Mater. 2016; 28: 161–167. [DOI] [PubMed] [Google Scholar]
- 80.Armstrong JP, Puetzer JL, Serio A, Guex AG, Kapnisi M, Breant A, et al. Engineering anisotropic muscle tissue using acoustic cell patterning. Adv Mater. 2018; 30: 1802649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Serpooshan V, Chen P, Wu H, Lee S, Sharma A, Hu DA, et al. Bioacoustic-enabled patterning of human iPSC-derived cardiomyocytes into 3D cardiac tissue. Biomaterials. 2017; 131: 47–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Apfel RE, Holland CK. Gauging the likelihood of cavitation from short-pulse, low-duty cycle diagnostic ultrasound. Ultrasound Med Biol. 1991; 17: 179–185. [DOI] [PubMed] [Google Scholar]
- 83.Flynn HG. Generation of transient cavities in liquids by microsecond pulses of ultrasound. J Acoust Soc Am. 1982; 72: 1926–1932. [Google Scholar]
- 84.Bader KB, Holland CK. Gauging the likelihood of stable cavitation from ultrasound contrast agents. Phys Med Biol. 2013; 58: 127–144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Flynn HG. Physics of acoustic cavitation in liquids Physical Acoustics. Cambridge: Academic Press; 1964. p. 58–172. [Google Scholar]
- 86.Dalecki D Mechanical bioeffects of ultrasound. Annu Rev Biomed Eng. 2004; 6: 229–248. [DOI] [PubMed] [Google Scholar]
- 87.Carstensen EL, Flynn HG. The potential for transient cavitation with microsecond pulses of ultrasound. Ultrasound Med Biol. 1982; 8: L720–L724. [DOI] [PubMed] [Google Scholar]
- 88.Unger EC, Matsunaga TO, McCreery T, Schumann P, Sweitzer R, Quigley R. Therapeutic applications of microbubbles. Eur J Radiol. 2002; 42: 160–168. [DOI] [PubMed] [Google Scholar]
- 89.Ingham KC, Brew SA, Isaacs BS. Interaction of fibronectin and its gelatin binding domains with fluorescent-labeled chains of type I collagen. J Biol Chem. 1988; 263: 4624–4628. [PubMed] [Google Scholar]
- 90.Geiger B, Bershadsky A, Pankov R, Yamada KM. Extracellular matrix-cytoskeleton crosstalk. Nat Rev Mol Cell Bio. 2001; 2: 703–805. [DOI] [PubMed] [Google Scholar]
- 91.Raub CB, Suresh V, Krasieva T, Lyubovitsky J, Mih JD, Putnam AJ, et al. Noninvasive assessment of collagen gel microstructure and mechanics using multiphoton microscopy. Biophys J. 2007; 92: 2212–2222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.American Institute of Ultrasound in Medicine. Statement on mammalian biological effects of ultrasound in vivo. American Institute for Ultrasound in Medicine; 2015. [Google Scholar]
- 93.Norris EG, Dalecki D, Hocking DC. Acoustic fabrication of collagen-fibronectin composite gels accelerates microtissue formation. Appl Sci. 2020; 10: 2907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Blinc A, Francis CW, Trudnowski JL, Carstensen EL. Characterization of ultrasound-potentiated fibrinolysis in vitro. Blood. 1993; 81: 2636–2643. [PubMed] [Google Scholar]
- 95.Braaten JV, Goss RA, Francis CW. Ultrasound reversibly disaggregates fibrin fibers. Thromb Haemostasis. 1997; 78: 1063–1068. [PubMed] [Google Scholar]
- 96.Ingram JH, Korossis S, Howling G, Fisher J, Ingham E. The use of ultrasonication to aid recellularization of acellular natural tissue scaffolds for use in anterior cruciate ligament reconstruction. Tissue Eng. 2007; 13: 1561–1572. [DOI] [PubMed] [Google Scholar]
- 97.Nele V, Schutt CE, Wojciechowski JP, Kit-Anan W, Doutch JJ, Armstrong JP, et al. Ultrasoundtriggered enzymatic gelation. Adv Mater. 2020; 32: 1905914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Fabiilli ML, Wilson CG, Padilla F, Martín-Saavedra FM, Fowlkes JB, Franceschi RT. Acoustic droplet hydrogel composites for spatial and temporal control of growth factor delivery and scaffold stiffness. Acta Biomater. 2013; 9: 7399–7409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Moncion A, Arlotta KJ, O’Neill EG, Lin M, Mohr LA, Franceschi RT, et al. In in vitro and in vivo assessment of controlled release and degradation of acoustically-responsive scaffolds. Acta Biomater. 2016; 46: 221–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Wilson CG, Martín-Saavedra FM, Padilla F, Fabiilli ML, Zhang M, Baez AM, et al. Patterning expression of regenerative growth factors using high intensity focused ultrasound. Tissue Eng Part C Methods. 2014; 20: 769–779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Moncion A, Harmon JS, Li Y, Natla S, Farrell EC, Kripfgans OD, et al. Spatiotemporally-controlled transgene expression in hydroxyapatite-fibrin composite scaffolds using high intensity focused ultrasound. Biomaterials. 2019; 194: 14–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Garvin KA, VanderBurgh J, Hocking DC, Dalecki D. Controlling collagen fiber microstructure in three-dimensional hydrogels using ultrasound. J Acoust Soc Am. 2013; 134: 1491–1502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Norris EG, Dalecki D, Hocking DC. Acoustic modification of collagen hydrogels facilitates cellular remodeling. Materi Today Bio. 2019; 3: 100018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Norris EG, Majeski J, Wayson SE, Coleman H, Choe R, Dalecki D, et al. Non-invasive acoustic fabrication methods to enhance collagen hydrogel bioactivity. Mater Res Express. 2019; 6: 125410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Lauer CG, Burge R, Tang DB, Bass BG, Gomez ER, Alving BM. Effect of ultrasound on tissue-type plasminogen activator-induced thrombolysis. Circulation. 1992; 86: 1257–1262. [DOI] [PubMed] [Google Scholar]
- 106.Bader KB, Gruber MJ, Holland CK. Shaken and stirred: Mechanisms of ultrasound-enhanced thrombolysis. Ultrasound Med Biol. 2015; 41: 187–196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Huang S, Shekhar H, Holland CK. Comparative lytic efficacy of rt-PA and ultrasound in porcine versus human clots. PLoS ONE. 2017; 12: e0177786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Laing ST, Moody MR, Kim H, Smulevitz B, Huang SL, Holland CK, et al. Thrombolytic efficacy of tissue plasminogen activator-loaded echnogenic liposomes in a rabbit thrombus model. Thromb Res. 2012; 130: 629–635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Kadler KE, Baldock C, Bella J, Boot-Handford RP. Collagens at a glance. J Cell Sci. 2007; 120: 1955–1958. [DOI] [PubMed] [Google Scholar]
- 110.Antoine EE, Vlachos PP, Rylander MN. Review of collagen I hydrogels for bioengineered tissue microenvironments: Characterization of mechanics, structure, and transport. Tissue Eng Part B Rev. 2014; 20: 683–696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Wood GC. The formation of fibrils from collagen solutions. 2. A mechanism of collagen-fibril formation. Biochem J. 1960; 75: 598–605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Williams BR, Gelman RA, Poppke DC, Piez KA. Collagen fibril formation. Optimal in vitro conditions and preliminary kinetic results. J Biol Chem. 1978; 253: 6578–6585. [PubMed] [Google Scholar]
- 113.Cen L, Liu W, Cui L, Zhang W, Cao Y. Collagen tissue engineering: Development of novel biomaterials and applications. Pediatr Res. 2008; 63: 492–496. [DOI] [PubMed] [Google Scholar]
- 114.Schleifenbaum S, Prietzel T, Aust G, Boldt A, Fritsch S, Keil I, et al. Acellularization-induced changes in tensile properties are organ specific - An in-vitro mechanical and structural analysis of porcine soft tissues. PLoS ONE. 2016; 11: e0151223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Wolf K, Te Lindert M, Krause M, Alexander S, Te Riet J, Willis AL, et al. Physical limits of cell migration: Control by ECM space and nuclear deformation and tuning by proteolysis and traction force. J Cell Biol. 2013; 201: 1069–1084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Roeder BA, Kokini K, Sturgis JE, Robinson JP, Voytik-Harbin SL. Tensile mechanical properties of three-dimensional type I collagen extracellular matrices with varied microstructure. J of Biomech Eng. 2002; 124: 214–222. [DOI] [PubMed] [Google Scholar]
- 117.Miron-Mendoza M, Seemann J, Grinnell F. The differential regulation of cell motile activity through matrix stiffness and porosity in three dimensional collagen matrices. Biomaterials. 2010; 31: 6425–6435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Zaman MH, Trapani LM, Sieminski AL, MacKellar D, Gong H, Kamm RD, et al. Migration of tumor cells in 3D matrices is governed by matrix stiffness along with cell-matrix adhesion and proteolysis. P Natl Acad Sci USA. 2006; 103: 10889–10894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Lanfer B, Seib FP, Freudenberg U, Stamov D, Bley T, Bornhauser M, et al. The growth and differentiation of mesenchymal stem and progenitor cells cultured on aligned collagen matrices. Biomaterials. 2009; 30: 5950–5958. [DOI] [PubMed] [Google Scholar]
- 120.Riching KM, Cox BL, Salick MR, Pehlke C, Riching AS, Ponik SM, et al. 3D collagen alignment limits protrusions to enhance breast cancer cell persistence. Biophys J. 2015; 107: 2546–2558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Matthews JA, Wnek GE, Simpson DG, Bowlin GL. Electrospining of collagen nanofibers. Biomacromolecules. 2002; 3: 232–238. [DOI] [PubMed] [Google Scholar]
- 122.Torbet J, Ronziere MC. Magnetic alingment of collagen during self-assembly. Biochem J. 1984; 219: 1057–1059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Eguchi Y, Ohtori S, Sekino M, Ueno S. Effectiveness of magnetically aligned collagen for neural regeneration in vitro and in vivo. Bioelectromagnetics. 2015; 36: 233–243. [DOI] [PubMed] [Google Scholar]
- 124.Carey SP, Rahman A, Kraning-Rush CM, Romero B, Somasegar S, Torre OM, et al. Comparative mechanisms of cancer cell migration through 3D matrix and physiological microtracks. Am J Physiol Cell Physiol. 2015; 308: C436–C447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Suslick KS. Sonochemistry. Science. 1990; 247: 1439–1445. [DOI] [PubMed] [Google Scholar]
- 126.Grabski AC. Advances in preparation of biological extracts for protein purification Methods in Enzymology. 463 2nd ed. San Diego: Academic Press; 2009. p. 285–303. [DOI] [PubMed] [Google Scholar]
- 127.Maller O, Hansen KC, Lyons TR, Acerbi I, Weaver VM, Prekeris R, et al. Collagen architechture in pregnancy-induced protection from breast cancer. J Cell Sci. 2013; 126: 4108–4110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Williams RM, Zipfel WR, Webb WW. Interpreting second-harmonic generation images of collagen I fibrils. Biophys J. 2005; 88: 1377–1386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Zitnay JL, Li Y, Qin Z, San BH, Depalle B, Reese SP, et al. Molecular level detection and localization of mechanical damage in collagen enabled by collagen hybridizing peptides. Nat Commun. 2017; 8: 14913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Singh P, Carraher C, Schwarzbauer JE. Assembly of fibronectin extracellular matrix. Annu Rev Cell Dev Biol. 2010; 26: 397–419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Kubow KE, Vukmirovic R, Zhe L, Klotzsch E, Smith ML, Gourdon D, et al. Mechanical forces regulate the interactions of fibronectin and collagen I in extracellular matrix. Nat Commun. 2015; 6: 8026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Dzamba BJ, Wu H, Jaenisch R, Peters DM. Fibronectin binding site in type I collagen regulates fibronectin fibril formation. J Cell Biol. 1993; 121: 1165–1172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Sottile J, Shi F, Rublyevska I, Chiang HY, Lust J, Chandler J. Fibronectin-dependent collagen I deposition modulates the cell response to fibronectin. Am J Physiol Cell Physiol. 2007; 293: C1934–C1946. [DOI] [PubMed] [Google Scholar]
- 134.Gildner CD, Roy DC, Farrar CS, Hocking DC. Opposing effects of collagen I and vitronectin on fibronectin fibril structure and function. Matrix Biol. 2014; 34: 33–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Yuan H, Hu H, Sun J, Shi M, Yu H, Li C, et al. Ultrasound microbubble delivery targetting intraplaque neovascularization inhibits atherosclerotic plaque in an APOE-deficient mouse model. In Vivo. 2018; 32: 1025–1032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Zhu X, Guo J, He C, Geng H, Yu G, Li J, et al. Ultrasound triggered image-guided drug delivery to inhibit vascular reconstruction via paclitaxel-loaded microbubbles. Sci Rep. 2016; 6: 21683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Cao WJ, Rosenblat JD, Roth NC, Kuliszewski MA, Matkar PN, Rudenko D, et al. Therapeutic angiogenesis by ultrasound-mediated microRNA-126–3p delivery. Arterioscler Thromb Vasc Biol. 2015; 35: 2401–2411. [DOI] [PubMed] [Google Scholar]
- 138.Lea-Bank H, O’Reill MA, Hynynen K. Ultrasound-responsve droplets for therapy: A review. J Control Release. 2019; 293: 144–154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Kooiman K, Roovers S, Langeveld SA, Kleven RT, Dewitte H O’Reilly MA et al. Ultrasound responsive cavitation nuclei for therapy and drug delivery. Ultrasound Med Biol. 2020; 46: 1296–1325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Munshi HG, Wu YI, Ariztia EV, Stack MS. Calcium regulation of matrix metalloproteinase-mediated migration in oral squamous cell carcinoma cells. J Biol Chem. 2002; 277: 41480–41488. [DOI] [PubMed] [Google Scholar]
- 141.Zhang K, Chen J. The regulation of integrin function by divalent cations. Cell Adh Migr. 2012; 6: 20–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Tran H, VanDusen WJ, Argraves WS. The self-association and fibronectin-bining sites of fibulin-1 map to calcium-binding epidermal growth factor-like domain. J Biol Chem. 1997; 272: 22600–22606. [DOI] [PubMed] [Google Scholar]
- 143.Dong X, Lu X, Kingston K, Brewer E, Juliar BA, Kripfgans OD, et al. Controlled delivery of basic fibroblast growth factor (bFGF) using acoustic droplet vaporization stimulates endothelial network formation. Acta Biomater. 2019; 97: 409–419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Lu X, Dong X, Natla S, Kripfgans OD, Fowlkes JB, Wang X, et al. Parametric study of acoustic droplet vaporization thresholds and payload release from acoustically-responsive scaffolds. Ultrasound Med Biol. 2019; 45: 2471–2484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Aliabouzar M, Lu X, Kripfgans OD, Fowlkes JB, Fabiilli ML. Acoustic droplet vaporization in acoustically responsive scaffolds: Effects of frequency of excitation, volume fraction and threshold determination method. Ultrasound Med Biol. 2019; 45: 3246–3260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Moncion A, Lin M, Kripfgans OD, Franceschi RT, Putnam AJ, Fabiilli ML. Sequential payload release from acoustically-responsive scaffolds using focused ultrasound. Ultrasound Med Biol. 2018; 44: 2323–2335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Walther W, Stein U. Heat-responsive gene expression for gene therapy. Adv Drug Deliver Rev 2009; 61: 641–649. [DOI] [PubMed] [Google Scholar]
- 148.Martín-Saavedra FM, Wilson CG, Voellmy R, Vilaboa N, Franceschi R. Spatiotemporal control of vascular endothelial growth factor expression using a heat-shock activated rapamycin-dependent gene switch. Hum Gene Ther Methods. 2013; 24: 160–170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Ren X, Zhao M, Lash B, Martino MM, Julier Z. Growth factor engineering strategies for regenerative medicine applications. Front Bioeng Biotechnol. 2019; 7: 496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Bissell M, Hall HG, Parry G. How does the extracellular matrix direct gene expression? J Theor Biol. 1982; 99: 31–68. [DOI] [PubMed] [Google Scholar]
- 151.Schell JY, Wilks BT, Patel M, Franck C, Chalivendra V, Cao X, et al. Harnessing cellular-derived forces in self-assembled microtissues to control the synthesis and alignment of ECM. Biomaterials. 2016; 77: 120–129. [DOI] [PubMed] [Google Scholar]
- 152.Wilks BT, Evans EB, Nakhla MN, Morgan JR. Directing fibroblast self-assembly to fabricate highly-aligned, collagen-rich matrices. Acta Biomater. 2018; 81: 70–79. [DOI] [PubMed] [Google Scholar]
- 153.Chernousov MA, Stahl RC, Carey DJ. Schwann cells use a novel collagen-dependent mechanism for fibronectin fibril assembly. J Cell Sci. 1998; 111: 2763–2777. [DOI] [PubMed] [Google Scholar]
- 154.Quaroni A, Isselbacher KJ, Ruoslahti E. Fibronectin synthesis by epithelial crypt cells of rat small intestine. P Natl Acad Sci USA. 1978; 75: 5548–5552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Herchenhan A, Ulhlenbrock F, Eliasson P, Weis M, Eyre D, Kadler KE, et al. Lysyl oxidase activity is required for ordered collagen fibrillogenesis by tendon cells. J Biol Chem. 2015; 290: 16440–16450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Bowles RD, Williams RM, Zipfel WR, Bonassar LJ. Self-assembly of aligned tissue-engineered annulus fibrosus and intervertebral disk composite via collagen gel contraction. Tissue Eng Part A. 2010; 16: 1339–1348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Puetzer JL, Koo E, Bonassar LJ. Induction of fiber alignment and mechanical anisotropy in tissue engineered menisci with mechanical anchoring. J Biomech. 2015; 48: 1436–1443. [DOI] [PubMed] [Google Scholar]
- 158.Puetzer JL, Bonassar LJ. Physiologically distributed loading patterns drive the formation of zonally organized collagen structures in tissue engineered meniscus. Tissue Eng Part A. 2016; 22: 907–915. [DOI] [PubMed] [Google Scholar]
- 159.Falanga V Wound healing and its impairment in the diabetic foot. Lancet. 2005; 366: 1736–1743. [DOI] [PubMed] [Google Scholar]
- 160.Gherardini L, Cousins CM, Hawkes JJ, Spengler J, Radel S, Lawler H, et al. A new immobilization method to arrange particles in a gel matrix by ultrasound standing waves. Ultrasound Med Biol. 2005; 31: 261–272. [DOI] [PubMed] [Google Scholar]
- 161.Kirkpatrick ND, Andreou S, Hoying JB, Utzinger U. Live imaging of collagen remodeling during angiogenesis. Am J Physiol Heart Circ Physiol. 2007; 292: H3198–H3206. [DOI] [PubMed] [Google Scholar]
- 162.Chen D, Sun Y, Gudur MS, Hsiao YS, Wu Z, Fu J, et al. Two-bubble acoustic tweezing cytometry for biomechanical probing and stimulation of cells. Biophys J. 2015; 108: 32–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Fan Z, Sun Y, Chen D, Tay D, Chen W, Deng CX, et al. Acoustic tweezing cytometry for live-cell subcellular modulation of intracellular cytoskeleton contractility. Sci Rep. 2013; 3: 2176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Melde K, Mark AG, Qui T, Fisher P. Holograms for acoustics. Nature. 2016; 537: 518–22. [DOI] [PubMed] [Google Scholar]
- 165.Ma Z, Holle AW, Melde K, Qiu T, Poeppel K, Kadiri VM, et al. Acoustic holographic cell patterning in a biocompatible hydrogel. Adv Mater. 2020; 32: 1904181. [DOI] [PubMed] [Google Scholar]
- 166.Melde K, Choi E, Wu Z, Palagi S, Qiu T, Fischer P. Acoustic fabrication via the assembly and fusion of particles. Adv Mater. 2018; 30: 1704507. [DOI] [PubMed] [Google Scholar]
- 167.Vats K, Benoit DS. Dynamic manipulation of hydrogels to control cell behavior: A review. Tissue Eng Part B Rev. 2013; 19: 455–469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Zhu J Bioactive modification of poly (ethylene glycol) hydrogels for Tissue Eng. Biomaterials. 2010; 31: 4639–4656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Benoit DS, Anseth KS. The effect on osteoblast function of colocalized RGD and PHSRN epitopes on PEG surfaces. Biomaterials. 2005; 26: 5209–5220. [DOI] [PubMed] [Google Scholar]
- 170.He X, Ma J, Jabbari E. Effect of grafting RGD and BMP-2 protein-derived peptides to a hydrogel substrate on osteogenic differentiation of marrow stromal cells. Langmuir. 2008; 24: 12508–12516. [DOI] [PubMed] [Google Scholar]
- 171.Roy DC, Mooney NA, Raeman CH, Dalecki D, Hocking DC. Fibronectin matrix mimetics promote full-thickness wound repair in diabetic mice. Tissue Eng Part A. 2013; 19: 2517–2526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Seliktar D, Zisch AH, Lutolf MP, Wrana JL, Hubbell JA. MMP-2 sensitive, VEGF-bearing bioactive hydrogels for promotion of vascular healing. J Biomed Mater Res A. 2004; 68: 704–716. [DOI] [PubMed] [Google Scholar]
- 173.Ghanem MA, Maxwell AD, Sapozhnikov OA, Khokhlova VA, Bailey MR. Quantification of acoustic radiation forces on solid objects in fluids. Phys Rev Appl. 2019; 12: 044076. [DOI] [PMC free article] [PubMed] [Google Scholar]