

Abstract
Fatty acid-binding protein 4 (FABP4) is secreted from adipose tissue and acts as an adipokine, and an elevated circulating FABP4 level is associated with metabolic disorders and atherosclerosis. However, little is known about the causal link between circulating FABP4 level and mortality in a general population. We investigated the relationship between FABP4 concentration and mortality including cardiovascular death during a 12-year period in subjects of the Tanno-Sobetsu Study, a population-based cohort (n = 721, male/female: 302/419). FABP4 concentration at baseline was significantly higher in female subjects than in male subjects. All-cause death occurred in 123 (male/female: 74/49) subjects, and 34 (male/female: 20/14) and 42 (male/female: 26/16) subjects died of cardiovascular events and cancer, respectively. When divided into 3 groups according to tertiles of FABP4 level at baseline by sex (T1–T3), Kaplan–Meier survival curves showed that there were significant differences in rates of all-cause death and cardiovascular death, but not cancer death, among the groups. Multivariable Cox proportional hazard model analysis with a restricted cubic spline showed that hazard ratio (HR) for cardiovascular death, but not that for all-cause death, significantly increased with a higher FABP4 level at baseline after adjustment of age and sex. The risk of cardiovascular death after adjustment of age, sex, body mass index and levels of brain natriuretic peptide and high-sensitivity C-reactive protein in the 3rd tertile (T3) group (HR: 4.96, 95% confidence interval: 1.20–22.3) was significantly higher than that in the 1st tertile (T1) group as the reference. In conclusion, elevated circulating FABP4 concentration predicts cardiovascular death in a general population.
Subject terms: Cardiovascular diseases, Metabolic disorders
Introduction
Fatty acid-binding proteins (FABPs), about 14–15-kDa cytosolic proteins, can bind long-chain fatty acids1 and have been proposed to facilitate the transport of lipids to specific organelles in the cell1. Among FABPs, fatty acid-binding protein 4 (FABP4), also known as adipocyte P2 (aP2) or adipocyte FABP (A-FABP), is expressed in adipose tissue including adipocytes and macrophages1,2. Previous studies using animal models showed that FABP4 contributes to the development of metabolic disorders and cardiovascular disease in communication with metabolic and inflammatory pathways in adipocytes and macrophages3–5. We previously showed in experimental models that inhibition of FABP4 by a small molecule might be a novel therapeutic strategy against insulin resistance, type 2 diabetes mellitus and atherosclerosis6. In a human study, subjects with a genetic variation of the FABP4 locus (T-87C) were shown to have a decrease in FABP4 expression in adipose tissue and beneficial effects on cardiovascular and metabolic health7.
FABP4 has no typical secretory signal peptides in the sequence of FABP41,2, but secretion of FABP4 from adipocytes via a non-classical pathway has recently been confirmed8,9. Circulating FABP4 has been reported to act as an adipokine, an adipocyte-derived bioactive molecule, for the development of insulin resistance8 and atherosclerosis10 in experimental models. Furthermore, it has been reported that neutralization of secreted FABP4 with an antibody to FABP4 can be a novel strategy for treatment of insulin resistance, type 2 diabetes mellitus, atherosclerosis and vascular injury8,11,12.
Cross-sectional studies have shown that an elevated circulating FABP4 level is associated with obesity, insulin resistance, type 2 diabetes mellitus, hypertension, dyslipidemia, dysregulation of purine metabolism, atherosclerosis and disturbance of the heart, liver and kidney13–19. Longitudinal studies have also shown that FABP4 concentration can be a predictor for the development of metabolic syndrome20, type 2 diabetes mellitus21 and atherosclerosis22. Furthermore, FABP4 level has been reported to be associated with long-term cardiovascular events in patients with coronary heart disease23, type 2 diabetes mellitus24 and hemodialysis25. However, little is known about the causal link between circulating FABP4 level and mortality including cardiovascular death and cancer death as hard endpoints in a general population. In the present study, we investigated the association of FABP4 level with mortality during a 12-year period in subjects of a general population.
Results
Characteristics of the study subjects
Characteristics of the 721 recruited subjects (male/female: 302/419) are shown in supplementary Table S1. Mean age, body mass index (BMI) and waist circumference of the recruited subjects were 64 ± 14 years, 24.0 ± 3.9 and 85.3 ± 10.2 cm, respectively. FABP4 concentration was significantly lower in male subjects (median [interquartile range]: 16.9 [12.5–22.9] ng/mL) than in female subjects (23.4 [16.7–30.4] ng/mL).
Basal characteristics of male and female subjects divided by subgroups according to tertiles of FABP4 level at baseline are shown in Tables 1 and 2, respectively. There were significant differences in parameters, including age, BMI, waist circumference, blood pressure, blood urea nitrogen, creatinine, estimated glomerular filtration rate (eGFR), uric acid, triglycerides, insulin, homeostasis model assessment of insulin resistance (HOMA-R), hemoglobin A1c, brain natriuretic peptide (BNP) and high-sensitivity C-reactive protein (hsCRP), in both male and female subjects. In male subjects, there were significant differences in the use of a calcium channel blocker and thiazolidinedione among the groups (supplementary Table S2). In female subjects, there were significant differences in the use of an angiotensin II receptor blocker, calcium channel blocker, β blocker, diuretic, statin and eicosapentaenoic acid among the groups (Supplementary Table S3).
Table 1.
Basal characteristics of the male subjects with tertiles of FABP4 level at baseline (n = 302).
| Tertiles of FABP4 level | T1 | T2 | T3 | P |
|---|---|---|---|---|
| (5.3–14.2 ng/mL) | (14.2–21.0 ng/mL) | (21.2–50.6 ng/mL) | ||
| n = 100 | n = 101 | n = 101 | ||
| Age (years) | 61 ± 14 | 65 ± 13* | 69 ± 13* | < 0.001 |
| Body mass index | 23.2 ± 2.9 | 24.4 ± 2.6 | 25.5 ± 5.3* | < 0.001 |
| Waist circumference (cm) | 80.9 ± 8.6 | 85.7 ± 7.0* | 89.3 ± 10.0* | < 0.001 |
| Systolic blood pressure (mmHg) | 134 ± 23 | 143 ± 21* | 142 ± 20* | 0.006 |
| Diastolic blood pressure (mmHg) | 78 ± 11 | 82 ± 11* | 79 ± 12 | 0.028 |
| Pulse rate (beats/min) | 68 ± 9 | 66 ± 9 | 66 ± 10 | 0.616 |
| Smoking habit | 33 (33.0) | 25 (24.8) | 31 (30.7) | 0.416 |
| Alcohol drinking habit | 32 (32.0) | 35 (34.7) | 44 (43.6) | 0.204 |
| Comorbidity | ||||
| Hypertension | 42 (42.0) | 62 (61.4)* | 64 (63.4)* | 0.004 |
| Diabetes mellitus | 11 (11.0) | 11 (10.9) | 25 (24.8)* | 0.008 |
| Dyslipidemia | 41 (41.0) | 48 (47.5) | 55 (54.5) | 0.161 |
| Biochemical data | ||||
| AST (IU/L) | 24 (20–29) | 26 (21–34) | 25 (21–31) | 0.128 |
| ALT (IU/L) | 21 (16–28) | 25 (19–32) | 22 (15–34) | 0.139 |
| γGTP (IU/L) | 25 (19–42) | 30 (19–46) | 32 (22–55) | 0.073 |
| Blood urea nitrogen (mg/dL) | 16 ± 4 | 17 ± 4 | 18 ± 6* | 0.007 |
| Creatinine (mg/dL) | 0.7 ± 0.1 | 0.8 ± 0.1* | 0.9 ± 0.3* | < 0.001 |
| eGFR (ml/min/1.73 m2) | 89.1 ± 15.4 | 79.6 ± 15.5* | 74.0 ± 21.3* | < 0.001 |
| Uric acid (mg/dL) | 5.6 ± 1.1 | 5.9 ± 1.1 | 6.4 ± 1.3* | < 0.001 |
| Total cholesterol (mg/dL) | 197 ± 36 | 192 ± 34 | 201 ± 36 | 0.249 |
| LDL cholesterol (mg/dL) | 122 ± 32 | 117 ± 33 | 122 ± 34 | 0.462 |
| HDL cholesterol (mg/dL) | 58 ± 14 | 54 ± 12 | 54 ± 16 | 0.060 |
| Triglycerides (mg/dL) | 81 (66–108) | 100 (75–143) | 108 (80–146)* | 0.014 |
| Fasting glucose (mg/dL) | 101 ± 25 | 103 ± 21 | 108 ± 25 | 0.092 |
| Insulin (µU/ml) | 3.8 (2.7–5.4) | 4.8 (3.4–7.5) | 6.1 (4.0–10.3)* | < 0.001 |
| HOMA-R | 0.94 (0.64–1.45) | 1.22 (0.82–1.83) | 1.52 (0.92–2.95)* | < 0.001 |
| Hemoglobin A1c (%) | 5.2 ± 0.5 | 5.2 ± 0.6 | 5.4 ± 0.7* | 0.012 |
| BNP (pg/mL) | 15 (8–23) | 16 (10–36) | 20 (10–38)* | 0.038 |
| hsCRP (mg/dL) | 0.04 (0.02–0.07) | 0.06 (0.03–0.12) | 0.07 (0.04–0.12)* | 0.025 |
Variables are expressed as number (%), means ± SD or medians (interquartile ranges).
AST aspartate transaminase, ALT alanine transaminase, BNP brain natriuretic peptide, eGFR estimated glomerular filtration rate, FABP4 fatty acid-binding protein 4, γGTP γ-glutamyl transpeptidase, HDL high-density lipoprotein, HOMA-R homeostasis model assessment of insulin resistance, hsCRP high-sensitivity C-reactive protein, LDL low-density lipoprotein.
*P < 0.05 versus T1.
Table 2.
Basal characteristics of the female subjects with tertiles of FABP4 level at baseline (n = 419).
| Tertiles of FABP4 level | T1 | T2 | T3 | P |
|---|---|---|---|---|
| (6.4–18.4 ng/mL) | (18.4–27.3 ng/mL) | (27.4–85.5 ng/mL) | ||
| n = 139 | n = 140 | n = 140 | ||
| Age (years) | 56 ± 15 | 65 ± 12* | 70 ± 10* | < 0.001 |
| Body mass index | 21.3 ± 3.1 | 23.8 ± 3.0* | 25.9 ± 4.2* | < 0.001 |
| Waist circumference (cm) | 78.2 ± 9.3 | 85.7 ± 9.4* | 92.0 ± 9.4* | < 0.001 |
| Systolic blood pressure (mmHg) | 126 ± 25 | 136 ± 21* | 147 ± 27* | < 0.001 |
| Diastolic blood pressure (mmHg) | 74 ± 13 | 77 ± 12 | 80 ± 13* | 0.001 |
| Pulse rate (beats/min) | 69 ± 12 | 72 ± 12 | 71 ± 9 | 0.321 |
| Smoking habit | 22 (15.8) | 10 (7.1)* | 9 (6.4)* | 0.013 |
| Alcohol drinking habit | 85 (61.2) | 104 (74.3)* | 109 (77.9)* | 0.005 |
| Comorbidity | ||||
| Hypertension | 50 (36.0) | 79 (56.4)* | 94 (67.1)* | < 0.001 |
| Diabetes mellitus | 5 (3.6) | 11 (7.9) | 10 (7.1) | 0.288 |
| Dyslipidemia | 37 (26.6) | 79 (56.4)* | 76 (54.3)* | < 0.001 |
| Biochemical data | ||||
| AST (IU/L) | 21 (18–24) | 23 (19–27)* | 25 (21–30)* | < 0.001 |
| ALT (IU/L) | 15 (13–19) | 19 (14–24)* | 20 (15–26)* | < 0.001 |
| γGTP (IU/L) | 16 (13–22) | 18 (14–27) | 20 (14–28) | 0.103 |
| Blood urea nitrogen (mg/dL) | 13 ± 4 | 15 ± 4* | 16 ± 4* | < 0.001 |
| Creatinine (mg/dL) | 0.5 ± 0.1 | 0.6 ± 0.1 | 0.7 ± 0.6* | 0.003 |
| eGFR (ml/min/1.73 m2) | 91.5 ± 19.5 | 83.6 ± 15.7* | 74.1 ± 19.4* | < 0.001 |
| Uric acid (mg/dL) | 4.3 ± 0.9 | 4.7 ± 0.9* | 5.1 ± 1.2* | < 0.001 |
| Total cholesterol (mg/dL) | 199 ± 31 | 212 ± 31* | 212 ± 35* | 0.001 |
| LDL cholesterol (mg/dL) | 116 ± 28 | 127 ± 32* | 129 ± 32* | 0.001 |
| HDL cholesterol (mg/dL) | 67 ± 15 | 63 ± 17* | 61 ± 16* | 0.002 |
| Triglycerides (mg/dL) | 71 (56–100) | 95 (67–122)* | 94 (74–136)* | < 0.001 |
| Fasting glucose (mg/dL) | 92 ± 11 | 98 ± 19* | 100 ± 22* | 0.001 |
| Insulin (µU/ml) | 4.1 (3.0–5.5) | 4.9 (3.7–7.0)* | 5.9 (4.1–8.7)* | < 0.001 |
| HOMA-R | 0.90 (0.67–1.22) | 1.16 (0.80–1.78)* | 1.37 (0.94–2.29)* | < 0.001 |
| Hemoglobin A1c (%) | 4.9 ± 0.4 | 5.2 ± 0.6* | 5.3 ± 0.7* | < 0.001 |
| BNP (pg/mL) | 17 (11–23) | 19 (11–37)* | 23 (14–40)* | 0.003 |
| hsCRP (mg/dL) | 0.02 (0.01–0.05) | 0.04 (0.02–0.08) | 0.05 (0.03–0.10)* | < 0.001 |
Variables are expressed as number (%), means ± SD or medians (interquartile ranges).
AST aspartate transaminase, ALT alanine transaminase, BNP brain natriuretic peptide, eGFR estimated glomerular filtration rate, FABP4 fatty acid-binding protein 4, γGTP γ-glutamyl transpeptidase, HDL high-density lipoprotein, HOMA-R homeostasis model assessment of insulin resistance, hsCRP high-sensitivity C-reactive protein, LDL low-density lipoprotein.
*P < 0.05 versus T1.
Mortality and FABP4 level at baseline
The mean follow-up period was 10.4 years (range 0.1–12.2 years), and follow-up summation was 7512 (male/female: 3045/4467) person-years. The number of censored subjects other than death was 88, and the follow-up rate was 87.8%. During a 12-year period, all-cause death occurred in 123 (male/female: 74/49) subjects, and 34 (male/female: 20/14) and 42 (male/female: 26/16) subjects died of cardiovascular events and cancer, respectively. Causes of cardiovascular death included acute myocardial infarction (n = 3), sudden cardiac death (n = 2), heart failure (n = 12), stroke (n = 5), cardiovascular hemorrhage (n = 3) and other cardiovascular causes (n = 9). Organs of cancer in cases of cancer death included the lung (n = 13), stomach (n = 5), colon (n = 4), pancreas (n = 4) and others (n = 16). Cumulative incidences of all-cause death, cardiovascular death and cancer death were 17.1% (male/female: 24.5%/11.7%), 4.7% (male/female: 6.6%/3.3%) and 5.8% (male/female: 8.6%/3.8%), respectively.
When divided into 3 groups according to tertiles of FABP4 level at baseline (T1–T3), Kaplan–Meier survival curves showed that there were significant differences in rates of all-cause mortality among the groups in all of the subjects (log-rank test: P < 0.001) (Fig. 1A) as well as in male subjects (Fig. 1B) and female subjects (Fig. 1C) when divided by sex. Similarly, there were significant differences in rates of cardiovascular death (Fig. 1D–F), but not cancer death (supplementary Figure S1A–C), among the groups.
Figure 1.

Survival curves of subgroups divided by tertiles of FABP4 level at baseline. (A–C) Kaplan–Meier survival curves for all-cause death in the three groups (T1–T3) according to tertiles of fatty acid-binding protein 4 (FABP4) concentration at baseline in all of the subjects (A) and in male (B) and female (C) subjects. (D–F) Kaplan–Meier survival curves for cardiovascular death in the three groups (T1–T3) according to tertiles of FABP4 concentration at baseline in all of the subjects (D) and in male (E) and female (F) subjects. Dashed line (T1), dotted line (T2) and solid line (T3). Survival rate was compared by the log-rank test.
Prediction of mortality by FABP4 level at baseline
Receiver operating characteristic (ROC) analyses for predicting all-cause death showed that the cut-off points of FABP4 concentration at baseline in male and female subjects were 18.6 ng/mL (sensitivity: 64.9%, specificity: 63.2%, area under the curve [AUC]: 0.63) and 25.3 ng/mL (sensitivity: 65.3%, specificity: 60.3%, AUC: 0.64), respectively (supplementary Figure S2A, B). ROC analyses for predicting cardiovascular death showed that the cut-off points of FABP4 concentration at baseline in male and female subjects were 19.1 ng/mL (sensitivity: 80.0%, specificity: 61.7%, AUC: 0.72) and 25.3 ng/mL (sensitivity: 85.7%, specificity: 58.8%, AUC: 0.73), respectively (supplementary Figure S2C, D).
Impact of FABP4 level at baseline on mortality during the follow-up period
Multivariable Cox proportional hazard model analysis with a restricted cubic spline showed that hazard ratio (HR) for all-cause death after adjustment of age and sex tended to increase with a higher FABP4 level at baseline in all of the subjects (Fig. 2A), but the 95% confidence interval (CI) included the null value (HR = 1) calculated by a minimum value of FABP4 (5.3 ng/mL). The HR for cardiovascular death after adjustment of age and sex increased with a higher FABP4 level at baseline in all of the subjects (Fig. 2B), and the 95% CI did not include the null value (HR = 1) in most of the range of FABP4 levels.
Figure 2.
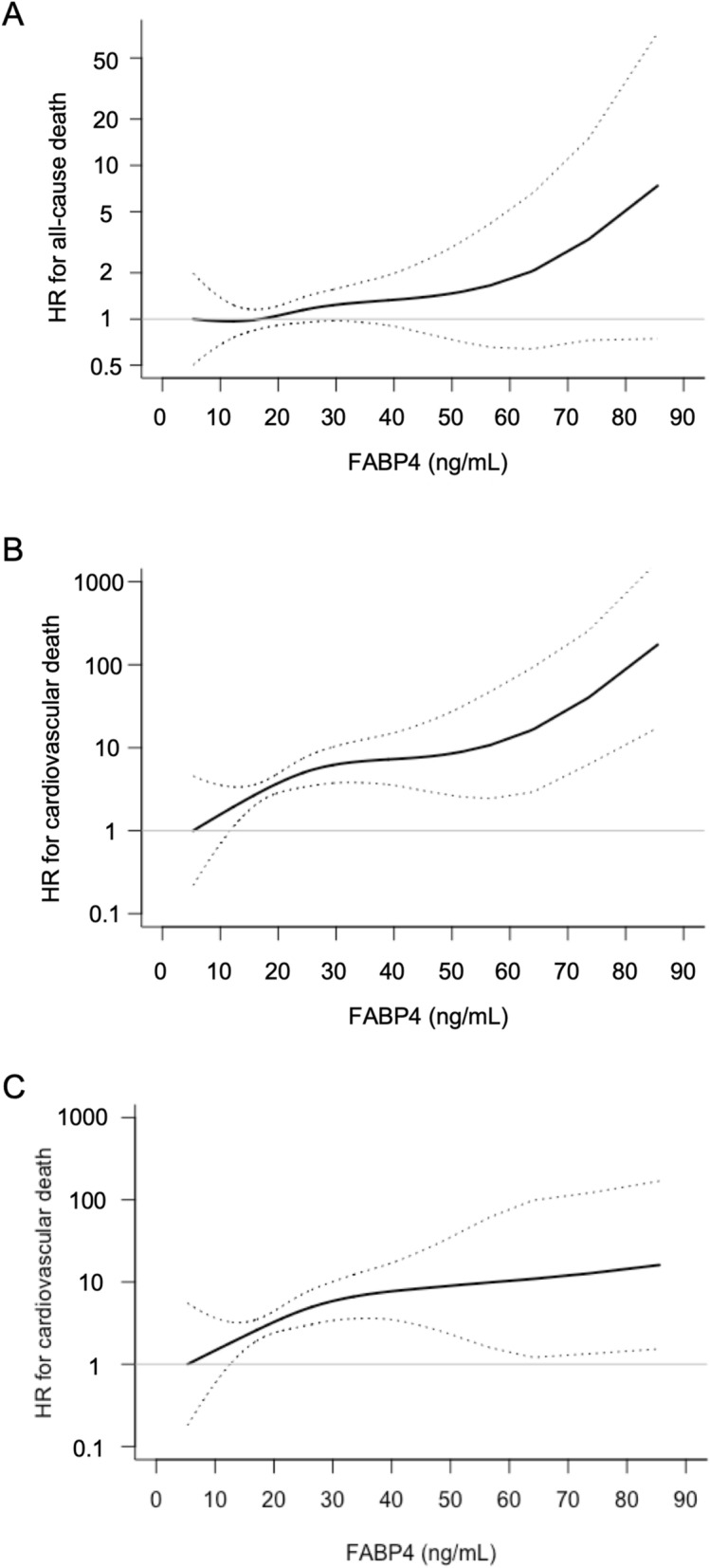
Hazard risk of FABP4 level for all-cause death and cardiovascular death. (A,B) Multivariable Cox proportional hazard analyses with a restricted cubic spline for all-cause death (A) and cardiovascular death (B) by fatty acid-binding protein 4 (FABP4) concentration at baseline after adjustment of age and sex during a 12-year follow-up period in all of the subjects. (C) Multivariable Cox proportional hazard analysis with a restricted cubic spline for cardiovascular death by FABP4 concentration at baseline after adjustment of age, sex, body mass index, brain natriuretic peptide and high-sensitivity C-reactive protein as the best-fit model during a 12-year follow-up period in all of the subjects. Solid line: hazard ratio (HR); dashed line: 95% confidence interval. The reference value of FABP4 was 5.3 ng/mL as a minimum value.
Multivariable Cox proportional hazard model analysis after adjustment of age and sex showed that HR for cardiovascular death in the 3rd tertile (T3) group of FABP4 level was significantly higher than that in the 1st tertile (T1) group as the reference (Model 1, Akaike’s Information Criterion [AIC]: 384) (Table 3). There was no significant interaction between sex and FABP4 tertiles for cardiovascular death (P = 0.951). After adjustment of age, sex, BMI and BNP, the risk of cardiovascular death in the T3 group was significantly higher than that in the T1 group, and there was no significant interaction between sex and FABP4 tertiles (Model 2, AIC: 366). When hsCRP was additionally incorporated into Model 2, the risk of cardiovascular death in the T3 group (HR: 4.96, 95% CI 1.20–22.3) was significantly higher than that in the T1 group (Model 3, AIC: 323). When hsCRP, smoking habit and diagnosis of hypertension, diabetes mellitus and dyslipidemia at baseline were additionally incorporated into Model 2, the risk of cardiovascular death in the T3 group was significantly higher than that in the T1 group (Model 4, AIC: 329).
Table 3.
Cox proportional hazard analyses for cardiovascular death in tertiles of FABP4 (n = 721).
| Model 1 | Model 2 | Model 3 | Model 4 | |||||
|---|---|---|---|---|---|---|---|---|
| HR (95% CI) | P | HR (95% CI) | P | HR (95% CI) | P | HR (95% CI) | P | |
| FABP4 | ||||||||
| T1 | Reference | – | Reference | – | Reference | – | Reference | – |
| T2 | 2.71 (0.59–12.5) | 0.201 | 2.76 (0.59–12.9) | 0.196 | 2.15 (0.44–10.5) | 0.344 | 2.07 (0.42–10.2) | 0.372 |
| T3 | 4.98 (1.15–21.6) | 0.032 | 5.38 (1.21–23.8) | 0.028 | 4.96 (1.10–22.3) | 0.037 | 4.83 (1.06–21.9) | 0.041 |
| Age (per 1 year) | 1.08 (1.03–1.14) | < 0.001 | 1.09 (1.05–1.13) | < 0.001 | 1.09 (1.04–1.13) | < 0.001 | 1.08 (1.03–1.13) | 0.001 |
| Sex (male) | 1.95 (0.68–5.62) | 0.214 | 1.98 (0.69–5.71) | 0.205 | 2.10 (0.71–6.27) | 0.180 | 2.29 (0.76–6.89) | 0.141 |
| Interaction* | 0.951 | 0.955 | 0.940 | 0.962 | ||||
| Body mass index | – | – | 0.91 (0.86–0.97) | 0.002 | 0.90 (0.85–0.97) | 0.001 | 0.91 (0.86–0.98) | 0.005 |
| BNP (per 1 pg/mL) | – | – | 1.01 (1.00–1.01) | < 0.001 | 1.01 (1.00–1.01) | < 0.001 | 1.01 (1.00–1.01) | < 0.001 |
| hsCRP (per 1 mg/dL) | – | – | – | – | 1.25 (0.01–49.4) | 0.915 | 1.07 (0.01–41.7) | 0.975 |
| Smoking habit | – | – | – | – | – | – | 0.85 (0.27–2.65) | 0.778 |
| Hypertension | – | – | – | – | – | – | 1.89 (0.61–5.81) | 0.267 |
| Diabetes mellitus | – | – | – | – | – | – | 0.44 (0.10–1.94) | 0.281 |
| Dyslipidemia | – | – | – | – | – | – | 0.90 (0.42–1.96) | 0.800 |
| (AIC = 384) | (AIC = 366) | (AIC = 323) | (AIC = 329) | |||||
*Interaction: Sex—FABP4 tertiles.
AIC Akaike's information criterion, BNP brain natriuretic peptide, CI confidence interval, FABP4 fatty acid-binding protein 4, HR hazard ratio, hsCRP high-sensitivity C-reactive protein.
The HR for cardiovascular death after adjustment of age, sex, BMI, BNP and hsCRP as the best-fit model using AIC increased with a higher FABP4 level at baseline in all of the subjects (Fig. 2C), and the 95% CI did not include the null value (HR = 1) in most of the range of FABP4 levels.
Discussion
The present study showed the impact of elevated circulating FABP4 concentration on the incidence of cardiovascular mortality in a general population. FABP4 has been shown to play a significant role in the development of insulin resistance, type 2 diabetes mellitus and atherosclerosis through its action at the interface of metabolic and inflammatory pathways in adipocytes and macrophages3–5. Furthermore, previous studies using in vitro and in vivo experiments showed that secreted FABP4 acts as an adipokine and directly affects various types of cells including hepatocytes, macrophages, cardiomyocytes, vascular endothelial cells and vascular smooth muscle cells8,10,12,26,27, though potential receptors for FABP4 have still not been identified2. Therefore, FABP4 may contribute to classical risk factors including insulin resistance and/or metabolic syndrome and might be a kind of “master regulatory factor” of metabolic risk factors. Moreover, FABP4 per se may directly contribute to atherosclerosis and/or cardiovascular damage, independent of classical risk factors. The combination of these possibilities such as a key master regulatory factor and an independently direct action of FABP4 may be related to cardiovascular death.
It has been reported that FABP4 concentration can predict not only the development of metabolic syndrome20, type 2 diabetes mellitus21 and atherosclerosis22 but also long-term cardiovascular events in patients with coronary heart disease23, type 2 diabetes mellitus24 and hemodialysis25. Obesity is a risk factor of several metabolic and cardiovascular diseases28, and FABP4 concentration has been reported to be strongly associated with BMI13,14. It has also been reported that FABP4 level reflects myocardial lipid storage as a putative effector of cardiac damage29. Therefore, BMI as a marker of obesity and BNP as an indicator of cardiac damage were incorporated as confounders in Cox proportional hazard model analyses in the present study (Table 3). A community-based cohort study showed that FABP4 concentration was associated with cardiovascular events including cardiovascular death during a 12-year follow-up period, though the predictive value was not significant when hsCRP was added in the adjustment for traditional risk factors30. In the present study, we demonstrated that elevated circulating FABP4 concentration predicts cardiovascular death, but not all-cause death or cancer death, as a hard endpoint after adjustment of confounding factors including hsCRP in a general population (Table 3, Model 3 and Model 4). These observations support the notion that measurement of FABP4 concentration is useful for risk stratification and for guiding treatment in observational or interventional studies.
Secretion of FABP4 from adipocytes via a non-classical pathway is associated with lipolysis8,9. Secretion of FABP4 from macrophages as well as adipocytes has also been confirmed10, but the main source of FABP4 in blood circulation is adipocytes5,8. Lipolysis is more active in visceral fat than in subcutaneous fat31, and visceral obesity has been reported to promote oxidative stress32. FABP4 normally prefers to bind essential polyunsaturated fatty acids, linoleic acid and α-linolenic acid, but the affinity of FABP4 would be changed to prefer binding a saturated fatty acid, palmitic acid, due to conformational structure change of FABP4 in a condition of visceral obesity-induced oxidative stress2,10. It has been reported that palmitic acid-bound, but not fatty acid-unbound or linoleic acid-bound, FABP4 can be a deteriorating adipokine and induce inflammatory responses in several target cells, including macrophages, smooth muscle cells, endothelial cells, adipocytes and other cells2,10. However, potential receptors of FABP4 have not been identified yet, and there have been some reports about internalization of FABP4 in the cell and interaction of cytokeratin 1 with FABP4 on the endothelial cell membrane2,12,33,34. Increased visceral obesity and consumption of palmitic acid-rich foods may lead to an increase in palmitic acid-bound FABP4, resulting in the development of cardiovascular events.
FABP4-deficient mice have a strong compensatory increase of FABP5 in adipose tissue3. Mice with combined deficiency of FABP4 and FABP5 exhibited protection against type 2 diabetes, fatty liver disease and atherosclerosis more than did FABP4- or FABP5-deficient mice35–37. There have been no reports about longevity in FABP4-deficient or FABP5-deficient mice, but it has recently been shown that the lifespan of FABP4/5 double knockout mice was not longer than that of wild-type mice despite the advantage of metabolic health in FABP4/5 double knockout mice38. It has been reported that FABP4/5 double knockout mice had reduced cardiac contraction due to disturbed utilization of fatty acids in the heart in a model of pressure overload-induced cardiac hypertrophy and failure by transverse aortic constriction39. Ageing-induced damage in several tissues with a high demand for fatty acid metabolism may overcome the advantage of metabolic health in FABP4/5 double knockout mice, suggesting that deficiency of either FABP4 or FABP5 is better than a combined deficiency of FABP4 and FABP5 in adipose tissue in terms of longevity.
Several drugs, including angiotensin II receptor blockers40,41, a statin42, omega-3 fatty acid ethyl esters43, and dipeptidyl peptidase-4 (DPP-4) inhibitors44,45, have been reported to decrease circulating FABP4 level. Randomized controlled trials using these drugs except DPP-4 inhibitors showed improvement of cardiovascular outcomes46–48, and DPP-4 inhibitors have been reported to reduce intima media thickness, a marker of atherosclerosis49,50. Reduction of FABP4 level by drugs would be a novel therapeutic strategy for prevention and reduction of cardiovascular mortality. Recent studies using experimental models demonstrated the possibility of a new strategy for treating metabolic diseases including diabetes mellitus, atherosclerosis and vascular injury by targeting serum FABP4 with a monoclonal antibody to FABP48,11,12. It is clearly necessary to prospectively evaluate whether a change in the FABP4 value by direct inhibition, neutralization and/or blockade of unidentified receptors indeed reflects conditions of metabolic syndrome and atherosclerosis and predicts long-term cardiovascular outcomes in the future.
The present study has several limitations. First, since the recruited subjects were only Japanese people, it is unclear whether the present findings can be generalized to other ethnicities. Second, recent studies have demonstrated modulation of FABP4 concentration by therapeutic drugs for hypertension, dyslipidemia and diabetes mellitus40–45,51,52, which might have affected cardiovascular events, though diagnosis of hypertension, dyslipidemia and diabetes mellitus was adjusted in Cox proportional hazard model analysis (Table 3, Model 4). Finally, FABP4 level was not investigated during the follow-up period. Investigation of the relationship between cardiovascular mortality and change in FABP4 level during a natural course and/or interventional studies would be needed in the future.
Conclusion
Elevated circulating FABP4 concentration predicts cardiovascular death in a general population. FABP4 concentration may be not only a marker of metabolic disorders but also a predictor of cardiovascular mortality in a general population as well as in patients with metabolic and cardiovascular diseases. A further understanding of the mechanism underlying the link between circulating FABP4 and cardiovascular events may enable the development of new therapeutic strategies for metabolic and cardiovascular diseases such as inhibition of FABP4, neutralization of FABP4 and blockade of the possible FABP4 receptor.
Methods
Study subjects
In the Tanno-Sobetsu Study, a study with a population-based cohort, a total of 721 Japanese subjects (male/female: 302/419, mean age: 64 ± 14 years) were recruited from residents of Sobetsu Town who received annual health examination in 2007. This study was performed with the approval of the Ethical Committee of Sapporo Medical University and conformed to the principles outlined in the Declaration of Helsinki. Written informed consent was received from all of the study subjects.
Follow-up and clinical outcome
All participants were followed up from 2007 to 2019, and their vital status, emigration status and cause of death were annually ascertained using residence registry data, death certificates, medical records in hospitals and/or questionnaires. Out-migrate individuals were defined as censored cases at the time of move-out day. The clinical endpoint was all-cause death including cardiovascular death and cancer death. Cardiovascular death included death resulting from acute myocardial infarction, sudden cardiac death and death due to heart failure, stroke, cardiovascular procedures, cardiovascular hemorrhage and other cardiovascular causes, as defined by the American Heart Association53.
Measurements
Medical check-ups were performed early in the morning after an overnight fast as previously described14. Blood pressure was measured using an automated sphygmomanometer (HEM-907, Omron Co., Kyoto, Japan). BMI was calculated as body weight (kg) divided by the square of body height (meter). Peripheral venous blood samples were analyzed immediately or stored at − 80 °C until biochemical analyses.
Measurements of biochemical parameters were performed as previously reported14. The concentration of FABP4 was measured using a commercially available enzyme-linked immunosorbent assay kit for FABP4 (Biovendor R&D, Modrice, Czech Republic). The accuracy, precision and reproducibility of the kit have been described previously13. BNP was measured using an assay kit (Shionogi & Co., Osaka, Japan). hsCRP was measured by a nephelometry method.
Low-density lipoprotein (LDL) cholesterol level was calculated by the Friedewald equation54. Hemoglobin A1c (HbA1c) was expressed in National Glycohemoglobin Standardization Program (NGSP) scale. HOMA-R as an index of insulin resistance was calculated by the following55: HOMA-R = insulin (μU/mL) × glucose (mg/dL)/405. eGFR was calculated by an equation for Japanese56: eGFR (ml/min/1.73 m2) = 194 × creatinine(−1.094) × age(−0.287) × 0.739 (if female).
A self-administered questionnaire survey was performed to obtain information on smoking habit, alcohol drinking habit, and use of drugs for diabetes mellitus, hypertension and dyslipidemia. Hypertension was diagnosed as systolic blood pressure ≥ 140 mmHg, diastolic blood pressure ≥ 90 mmHg or self-reported use of anti-hypertensive drugs. Diabetes mellitus was diagnosed in accordance with the guidelines of the American Diabetes Association57: fasting plasma glucose ≥ 126 mg/dL, HbA1c ≥ 6.5% or self-reported use of anti-diabetic drugs. Dyslipidemia was diagnosed as LDL cholesterol ≥ 140 mg/dL, high-density lipoprotein (HDL) cholesterol < 40 mg/dL, triglycerides ≥ 150 mg/dL or self-reported use of anti-dyslipidemic drugs.
Statistical analysis
Numeric variables are expressed as means ± standard deviation (SD) for normal distributions or medians (interquartile ranges) for skewed variables. The distribution of each parameter was tested for its normality using the Shapiro–Wilk W test. Intergroup differences in percentages of demographic parameters were examined by the chi-square test. Comparison between two groups was done with the Mann–Whitney's U test. One-way analysis of variance and Dunnett’s post hoc test were used for detecting significant differences in data between data in multiple groups. Survival rate was analyzed by the log-rank test of Kaplan–Meier curves. ROC analysis was performed to determine the inflection point at which the FABP4 level provided the most sensitive prediction of all-cause death and cardiovascular death in both male and female subjects, and the AUC of was calculated. The relationship between FABP4 concentration and hazard ratio (HR) for all-cause death, cardiovascular death or cancer death was analyzed by a multivariable Cox proportional hazard model with a restricted cubic spline after adjustment of age, sex and other confounders. In addition, the HRs and 95% CIs for cardiovascular death in three subgroups according to tertiles of FABP4 level at baseline (T1–T3) were calculated by adjustment of confounders including age, sex, BMI, smoking habit, levels of BNP and hsCRP, and diagnosis of hypertension, diabetes mellitus and dyslipidemia at baseline. Interaction of sex and tertiles of FABP4 level was also investigated. Among the candidate models, the best-fit model using AIC for each dependent variable was selected. A p value of < 0.05 was considered statistically significant. All data were analyzed by using JMP 14.3.0 for Macintosh (SAS Institute, Cary, NC) and R3.6.2.
Supplementary Information
Author contributions
M.F. conceived the project, performed data analyses and wrote the paper. N.S., M.K, H.A., M.T., N.M., H.O., S.S., N.U. performed data collection. M.T., M.K. and HO performed the statistical analyses. K.S. and T.M. supervised the analyses. All authors approved the final version of manuscript.
Funding
M.F. has been supported by grants from JSPS KAKENHI, SENSHIN Medical Research Foundation, Akiyama Life Science Foundation and Terumo Life Science Foundation.
Data availability
The datasets analyzed during the current study are available from the corresponding author on reasonable request.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
These authors contributed equally: Norie Saito and Masato Furuhashi.
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-021-83494-5.
References
- 1.Furuhashi M, Hotamisligil GS. Fatty acid-binding proteins: role in metabolic diseases and potential as drug targets. Nat. Rev. Drug Discov. 2008;7:489–503. doi: 10.1038/nrd2589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Furuhashi M. Fatty acid-binding protein 4 in cardiovascular and metabolic diseases. J. Atheroscler. Thromb. 2019;26:216–232. doi: 10.5551/jat.48710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hotamisligil GS, et al. Uncoupling of obesity from insulin resistance through a targeted mutation in aP2, the adipocyte fatty acid binding protein. Science. 1996;274:1377–1379. doi: 10.1126/science.274.5291.1377. [DOI] [PubMed] [Google Scholar]
- 4.Makowski L, et al. Lack of macrophage fatty-acid-binding protein aP2 protects mice deficient in apolipoprotein E against atherosclerosis. Nat. Med. 2001;7:699–705. doi: 10.1038/89076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Furuhashi M, et al. Adipocyte/macrophage fatty acid-binding proteins contribute to metabolic deterioration through actions in both macrophages and adipocytes in mice. J. Clin. Investig. 2008;118:2640–2650. doi: 10.1172/JCI34750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Furuhashi M, et al. Treatment of diabetes and atherosclerosis by inhibiting fatty-acid-binding protein aP2. Nature. 2007;447:959–965. doi: 10.1038/nature05844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Tuncman G, et al. A genetic variant at the fatty acid-binding protein aP2 locus reduces the risk for hypertriglyceridemia, type 2 diabetes, and cardiovascular disease. Proc. Natl. Acad. Sci. USA. 2006;103:6970–6975. doi: 10.1073/pnas.0602178103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Cao H, et al. Adipocyte lipid chaperone AP2 is a secreted adipokine regulating hepatic glucose production. Cell Metab. 2013;17:768–778. doi: 10.1016/j.cmet.2013.04.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Mita T, et al. FABP4 is secreted from adipocytes by adenyl cyclase-PKA- and guanylyl cyclase-PKG-dependent lipolytic mechanisms. Obesity (Silver Spring) 2015;23:359–367. doi: 10.1002/oby.20954. [DOI] [PubMed] [Google Scholar]
- 10.Furuhashi M, et al. Local production of fatty acid-binding protein 4 in epicardial/perivascular fat and macrophages is linked to coronary atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2016;36:825–834. doi: 10.1161/ATVBAHA.116.307225. [DOI] [PubMed] [Google Scholar]
- 11.Burak MF, et al. Development of a therapeutic monoclonal antibody that targets secreted fatty acid-binding protein aP2 to treat type 2 diabetes. Sci. Transl. Med. 2015;7:319ra205. doi: 10.1126/scitranslmed.aac6336. [DOI] [PubMed] [Google Scholar]
- 12.Fuseya T, et al. Ectopic fatty acid-binding protein 4 expression in the vascular endothelium is involved in neointima formation after vascular injury. J. Am. Heart Assoc. 2017 doi: 10.1161/JAHA.117.006377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Xu A, et al. Adipocyte fatty acid-binding protein is a plasma biomarker closely associated with obesity and metabolic syndrome. Clin. Chem. 2006;52:405–413. doi: 10.1373/clinchem.2005.062463. [DOI] [PubMed] [Google Scholar]
- 14.Ishimura S, et al. Circulating levels of fatty acid-binding protein family and metabolic phenotype in the general population. PLoS ONE. 2013;8:e81318. doi: 10.1371/journal.pone.0081318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ota H, et al. Elevation of fatty acid-binding protein 4 is predisposed by family history of hypertension and contributes to blood pressure elevation. Am. J. Hypertens. 2012;25:1124–1130. doi: 10.1038/ajh.2012.88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Furuhashi M, et al. Independent link between levels of proprotein convertase subtilisin/kexin type 9 and FABP4 in a general population without medication. Am. J. Cardiol. 2016;118:198–203. doi: 10.1016/j.amjcard.2016.04.037. [DOI] [PubMed] [Google Scholar]
- 17.Fuseya T, et al. Elevation of circulating fatty acid-binding protein 4 is independently associated with left ventricular diastolic dysfunction in a general population. Cardiovasc. Diabetol. 2014;13:126. doi: 10.1186/s12933-014-0126-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Yeung DC, et al. Serum adipocyte fatty acid-binding protein levels were independently associated with carotid atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2007;27:1796–1802. doi: 10.1161/ATVBAHA.107.146274. [DOI] [PubMed] [Google Scholar]
- 19.Furuhashi M, et al. Independent links between plasma xanthine oxidoreductase activity and levels of adipokines. J. Diabetes Investig. 2019;10:1059–1067. doi: 10.1111/jdi.12982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Xu A, et al. Circulating adipocyte-fatty acid binding protein levels predict the development of the metabolic syndrome: a 5-year prospective study. Circulation. 2007;115:1537–1543. doi: 10.1161/CIRCULATIONAHA.106.647503. [DOI] [PubMed] [Google Scholar]
- 21.Tso AWK, et al. Serum adipocyte fatty acid binding protein as a new biomarker predicting the development of type 2 diabetes: a 10-year prospective study in a Chinese cohort. Diabetes Care. 2007;30:2667–2672. doi: 10.2337/dc07-0413. [DOI] [PubMed] [Google Scholar]
- 22.Furuhashi M, et al. Circulating fatty acid-binding protein 4 concentration predicts the progression of carotid atherosclerosis in a general population without medication. Circ. J. 2018;82:1121–1129. doi: 10.1253/circj.CJ-17-1295. [DOI] [PubMed] [Google Scholar]
- 23.von Eynatten M, et al. Circulating adipocyte fatty acid-binding protein levels and cardiovascular morbidity and mortality in patients with coronary heart disease: a 10-year prospective study. Arterioscler. Thromb. Vasc. Biol. 2012;32:2327–2335. doi: 10.1161/ATVBAHA.112.248609. [DOI] [PubMed] [Google Scholar]
- 24.Liu G, et al. Plasma levels of fatty acid-binding protein 4, retinol-binding protein 4, high-molecular-weight adiponectin, and cardiovascular mortality among men with type 2 diabetes: a 22-year prospective study. Arterioscler. Thromb. Vasc. Biol. 2016;36:2259–2267. doi: 10.1161/ATVBAHA.116.308320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Furuhashi M, et al. Serum fatty acid-binding protein 4 is a predictor of cardiovascular events in end-stage renal disease. PLoS ONE. 2011;6:e27356. doi: 10.1371/journal.pone.0027356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yamamoto T, et al. Transcriptome and metabolome analyses in exogenous FABP4- and FABP5-treated adipose-derived stem cells. PLoS ONE. 2016;11:e0167825. doi: 10.1371/journal.pone.0167825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lamounier-Zepter V, et al. Adipocyte fatty acid-binding protein suppresses cardiomyocyte contraction: a new link between obesity and heart disease. Circ. Res. 2009;105:326–334. doi: 10.1161/CIRCRESAHA.109.200501. [DOI] [PubMed] [Google Scholar]
- 28.Gregor MF, Hotamisligil GS. Inflammatory mechanisms in obesity. Annu. Rev. Immunol. 2011;29:415–445. doi: 10.1146/annurev-immunol-031210-101322. [DOI] [PubMed] [Google Scholar]
- 29.Rodriguez-Calvo R, et al. Fatty acid binding protein 4 (FABP4) as a potential biomarker reflecting myocardial lipid storage in type 2 diabetes. Metabolism. 2019;96:12–21. doi: 10.1016/j.metabol.2019.04.007. [DOI] [PubMed] [Google Scholar]
- 30.Chow WS, et al. Elevated circulating adipocyte-fatty acid binding protein levels predict incident cardiovascular events in a community-based cohort: a 12-year prospective study. J. Am. Heart Assoc. 2013;2:e004176. doi: 10.1161/JAHA.112.004176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wajchenberg BL. Subcutaneous and visceral adipose tissue: their relation to the metabolic syndrome. Endocr. Rev. 2000;21:697–738. doi: 10.1210/edrv.21.6.0415. [DOI] [PubMed] [Google Scholar]
- 32.Furukawa S, et al. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004;114:1752–1761. doi: 10.1172/JCI21625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Saavedra P, et al. New insights into circulating FABP4: interaction with cytokeratin 1 on endothelial cell membranes. Biochim. Biophys. Acta. 1853;2966–2974:2015. doi: 10.1016/j.bbamcr.2015.09.002. [DOI] [PubMed] [Google Scholar]
- 34.Martinez-Micaelo N, et al. Extracellular FABP4 uptake by endothelial cells is dependent on cytokeratin 1 expression. Biochim. Biophys. Acta Mol. Cell Biol. Lipids. 1864;234–244:2019. doi: 10.1016/j.bbalip.2018.11.011. [DOI] [PubMed] [Google Scholar]
- 35.Maeda K, et al. Adipocyte/macrophage fatty acid binding proteins control integrated metabolic responses in obesity and diabetes. Cell Metab. 2005;1:107–119. doi: 10.1016/j.cmet.2004.12.008. [DOI] [PubMed] [Google Scholar]
- 36.Cao H, et al. Regulation of metabolic responses by adipocyte/macrophage fatty acid-binding proteins in leptin-deficient mice. Diabetes. 2006;55:1915–1922. doi: 10.2337/db05-1496. [DOI] [PubMed] [Google Scholar]
- 37.Boord JB, et al. Combined adipocyte-macrophage fatty acid-binding protein deficiency improves metabolism, atherosclerosis, and survival in apolipoprotein E-deficient mice. Circulation. 2004;110:1492–1498. doi: 10.1161/01.CIR.0000141735.13202.B6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Charles KN, et al. Uncoupling of metabolic health from longevity through genetic alteration of adipose tissue lipid-binding proteins. Cell Rep. 2017;21:393–402. doi: 10.1016/j.celrep.2017.09.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Umbarawan Y, et al. Glucose is preferentially utilized for biomass synthesis in pressure-overloaded hearts: evidence from fatty acid-binding protein-4 and -5 knockout mice. Cardiovasc. Res. 2018;114:1132–1144. doi: 10.1093/cvr/cvy063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Miyoshi T, et al. Olmesartan reduces arterial stiffness and serum adipocyte fatty acid-binding protein in hypertensive patients. Heart Vessels. 2011;26:408–413. doi: 10.1007/s00380-010-0060-x. [DOI] [PubMed] [Google Scholar]
- 41.Furuhashi M, et al. Angiotensin II receptor blockers decrease serum concentration of fatty acid-binding protein 4 in patients with hypertension. Hypertens Res. 2015;38:252–259. doi: 10.1038/hr.2015.2. [DOI] [PubMed] [Google Scholar]
- 42.Karpisek M, et al. Treatment with atorvastatin reduces serum adipocyte-fatty acid binding protein value in patients with hyperlipidaemia. Eur. J. Clin. Investig. 2007;37:637–642. doi: 10.1111/j.1365-2362.2007.01835.x. [DOI] [PubMed] [Google Scholar]
- 43.Furuhashi M, et al. Reduction of circulating FABP4 level by treatment with omega-3 fatty acid ethyl esters. Lipids Health Dis. 2016;15:5. doi: 10.1186/s12944-016-0177-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Furuhashi M, et al. Reduction of serum FABP4 level by sitagliptin, a DPP-4 inhibitor, in patients with type 2 diabetes mellitus. J. Lipid Res. 2015;56:2372–2380. doi: 10.1194/jlr.M059469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Furuhashi M, et al. Treatment with anagliptin, a DPP-4 inhibitor, decreases FABP4 concentration in patients with type 2 diabetes mellitus at a high risk for cardiovascular disease who are receiving statin therapy. Cardiovasc. Diabetol. 2020;19:89. doi: 10.1186/s12933-020-01061-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Messerli FH, Bangalore S. Angiotensin receptor blockers reduce cardiovascular events, including the risk of myocardial infarction. Circulation. 2017;135:2085–2087. doi: 10.1161/CIRCULATIONAHA.116.025950. [DOI] [PubMed] [Google Scholar]
- 47.Zhou Q, Liao JK. Statins and cardiovascular diseases: from cholesterol lowering to pleiotropy. Curr. Pharm. Des. 2009;15:467–478. doi: 10.2174/138161209787315684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Abdelhamid AS, et al. Omega-3 fatty acids for the primary and secondary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2020;3:CD003177. doi: 10.1002/14651858.CD003177.pub5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Mita T, et al. Alogliptin, a dipeptidyl peptidase 4 inhibitor, prevents the progression of carotid atherosclerosis in patients with type 2 diabetes: the study of preventive effects of alogliptin on diabetic atherosclerosis (SPEAD-A) Diabetes Care. 2016;39:139–148. doi: 10.2337/dc15-0781. [DOI] [PubMed] [Google Scholar]
- 50.Mita T, et al. Sitagliptin attenuates the progression of carotid intima-media thickening in insulin-treated patients with type 2 diabetes: the sitagliptin preventive study of intima-media thickness evaluation (SPIKE): a randomized controlled trial. Diabetes Care. 2016;39:455–464. doi: 10.2337/dc15-2145. [DOI] [PubMed] [Google Scholar]
- 51.Furuhashi M, et al. Possible increase in serum FABP4 level despite adiposity reduction by canagliflozin, an SGLT2 inhibitor. PLoS ONE. 2016;11:e0154482. doi: 10.1371/journal.pone.0154482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Cabre A, et al. Fatty acid binding protein 4 is increased in metabolic syndrome and with thiazolidinedione treatment in diabetic patients. Atherosclerosis. 2007;195:e150–158. doi: 10.1016/j.atherosclerosis.2007.04.045. [DOI] [PubMed] [Google Scholar]
- 53.Hicks KA, et al. 2017 Cardiovascular and stroke endpoint definitions for clinical trials. Circulation. 2018;137:961–972. doi: 10.1161/CIRCULATIONAHA.117.033502. [DOI] [PubMed] [Google Scholar]
- 54.Friedewald WT, Levy RI, Fredrickson DS. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972;18:499–502. doi: 10.1093/clinchem/18.6.499. [DOI] [PubMed] [Google Scholar]
- 55.Matthews DR, et al. Homeostasis model assessment: insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia. 1985;28:412–419. doi: 10.1007/BF00280883. [DOI] [PubMed] [Google Scholar]
- 56.Matsuo S, et al. Revised equations for estimated GFR from serum creatinine in Japan. Am J Kidney Dis. 2009;53:982–992. doi: 10.1053/j.ajkd.2008.12.034. [DOI] [PubMed] [Google Scholar]
- 57.American Diabetes Association 2. Classification and diagnosis of diabetes. Diabetes Care. 2017;40:S11–S24. doi: 10.2337/dc17-S005. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The datasets analyzed during the current study are available from the corresponding author on reasonable request.