

Abstract
The trabecular meshwork (TM) constitutes the main pathway for aqueous humor drainage and is exposed to complex intraocular pressure fluctuations. The mechanism of homeostasis in which TM senses changes in intraocular pressure and leads to normal levels of outflow resistance is not yet well understood. Previous reports have shown that Piezo1, a mechanically-activated cation channel, is expressed in TM and isolated TM cells. Therefore, we tested hypothesis that Piezo1 may function in response to membrane tension and stretch in TM. In human trabecular meshwork (hTM) cells, PIEZO1 was showed to be abundantly expressed, and Piezo1 agonist Yoda1 and mechanical stretch caused a Piezo1-dependent Ca2+ influx and release of arachidonic acid and PGE2. Treatment with Yoda1 or PGE2 significantly inhibited hTM cell contraction. These results suggest that mechanical stretch stimuli in TM activates Piezo1 and subsequently regulates TM cell contraction by triggering Ca2+ influx and release of arachidonic acid and PGE2. Thus, Piezo1 could acts as a regulator of intraocular pressure (IOP) within the conventional outflow pathway and could be a novel therapeutic strategy to modulate IOP in glaucoma patients.
Subject terms: Lipidomics, Cell biology, Molecular biology
Introduction
Intraocular pressure (IOP) is regulated by the production and outflow of aqueous humor and has been associated with increased risk of glaucoma development. The main route of aqueous humor drainage is the conventional outflow pathway, which involves the flow of aqueous humor to the Schlemm's canal through the juxtacanalicular trabecular meshwork (TM)1,2. Conventional outflow tissues are constantly subjected to fluctuations in IOP, and cells in these tissues sense alterations in IOP as mechanical stretch or strain2,3. Changes in pressure of 8–30 mmHg can cause length extension by 50% in cells in the outflow pathway4,5. In response to mechanical stretch or strain, TM cells activate intracellular signaling pathways leading to changes in gene expression, extracellular matrix (ECM) turnover, contractile properties2,6.
The non-selective cation channel TRPV4 and the two-pore domain potassium channel TREK1 are expressed in TM cells, sense mechanical stretch generated by fluctuations in IOP, and play an essential role in IOP adjustment7–10. Following membrane stretch, TM-resident TRPV4 channels initiate calcium (Ca2+) signals and cytoskeletal reorganization, and TRPV4 antagonists increased the outflow facility and lowered IOP in glaucomatous mouse eyes10. Recent studies showed that TREK-1 collaborates with TRPV4, regulating TM tensile homeostasis8. Although numerous studies highlighted the importance of mechanosensation in modulating TM outflow, the molecular links between mechanotransduction, Ca2+ homeostasis, and reorganization of the conventional outflow pathway remain unclear. Hence, the mechanosensory mechanisms mediating force-coupling and IOP regulation should be further explored.
Piezo1 and Piezo2 were first described in 2010 as components of a mechanically activated cation channel expressed in neurons and neuronal cell lines11. Piezo1 has been implicated in mechanical stretch response and extracellular Ca2+ influx regulation in the bladder urothelium12, red blood cell13, and neural stem cells14. Nevertheless, its role in TM remains poorly understood. We hypothesized that in TM, Piezo1 might respond to mechanical stress and regulate IOP by locally releasing lipid mediators. Lipid mediators are well known to have an important role in various physiological events through rapid production from the cell membrane and working transiently and locally in the restricted tissue. Notably, several arachidonic acid-derived metabolites have been implicated in mechanical signaling15,16. Among lipid mediators, prostaglandins such as PGF2α and PGE2, lysophospholipids, or cannabinoids are associated to the IOP regulation17–20. Thus, we investigated the involvement of Piezo1 on the mechanical stress and the release of lipid mediators in primary cultured human TM cells.
Results
Expression of Piezo and TRPV family members in primary hTM cells
First, we examined the expression of human Piezo and TRPV family members in primary hTM cells by qPCR. Piezo1 mRNA levels were profoundly higher than Piezo2 and TRPV family mRNA levels in hTM cells (Fig. 1).
Figure 1.

Expression levels of TRPV and Piezo family in human trabecular meshwork cells. mRNA levels of TRPV1-6 and Piezo1-2 in the human trabecular meshwork (hTM) cells were determined by qPCR. The number of mRNA copies of the target genes was calculated by normalizing to the amount of total RNA. Data are presented as means ± SD (n = Eight hTM cell populations from eight donors).
Knockdown of Piezo1 using siRNA in hTM cells
To investigate the involvement of Piezo1 in response to cell stretch stimuli, primary hTM cells were transfected with Piezo1 or control siRNA. Piezo1 siRNA #1 or #2 transfection reduced Piezo1 mRNA levels in hTM cells to 42.1 ± 6.44% or 31.8 ± 4.31%, respectively, compared to the control (Fig. 2A). As Piezo1 is a Ca2+ permeable channel11, we evaluated Ca2+ influx induced by the Piezo1 agonist Yoda1 in hTM cells loaded with the Ca2+ indicator Fluo-8. The Yoda1-induced maximum fluorescence intensity change Fmax/F0 was 3.76 ± 0.07 in cells transfected with control siRNA, but 2.45 ± 0.05 or 2.20 ± 0.12 in cells transfected with Piezo1 siRNA # 1 or # 2, respectively (Fig. 2B–D). We confirmed that primary hTM cells expressed functional Piezo1 and that Piezo1 siRNA treatment significantly suppressed Piezo1 at the mRNA and functional levels.
Figure 2.

Efficacy of Piezo1 knockdown in human trabecular meshwork cells. (A) Relative Piezo1 mRNA levels in the human trabecular meshwork treated with control or Piezo1 siRNAs 24 h after transfection, as determined by qPCR. Data are normalized to GAPDH mRNA level. Data are presented as means ± SE (n = 4 samples of cDNA synthesized from RNA extracted from cells in 4 wells). **p < 0.01, Dunnett’s test. (B) Representative images illustrating intracellular Ca2+ changes in human trabecular meshwork cells transfected with control or Piezo1 siRNAs and stimulated with Piezo1 agonist Yoda1. Scale bar, 100 μm. (C) Time course of Yoda1-induced Ca2+ changes in control or Piezo1 siRNA-transfected human trabecular meshwork cells. (D) Yoda1-evoked intracellular Ca2+ elevation in human trabecular meshwork cells transfected with control or Piezo1 siRNAs. Data are expressed as the means ± SE (n = 4 experiments; ≥ 20 cells/experiment). **p < 0.01, Dunnett’s test.
Mechanical stretch stimulation triggers calcium influx via Piezo1 activation in hTM cells
Mechanical stretch stimulation in hTM cells promoted Ca2+ influx10. To investigate the relevance of Piezo1 in Ca2+ influx following mechanical stretch stimulation, we stimulated hTM cells grown on a fibronectin-coated silicon chamber with a 30% uniaxial stretch. Intracellular Ca2+ elevation induced by mechanical stretch was significantly suppressed in PIEZO1-knockdown cells (Fmax/F0 = 1.81 ± 5.65) compared with control cells (Fmax/F0 = 2.02 ± 7.19, Fig. 3A,B). These findings suggest that mechanical stretch stimulation in primary hTM cells induces Ca2+ influx in a Piezo1-dependent manner.
Figure 3.
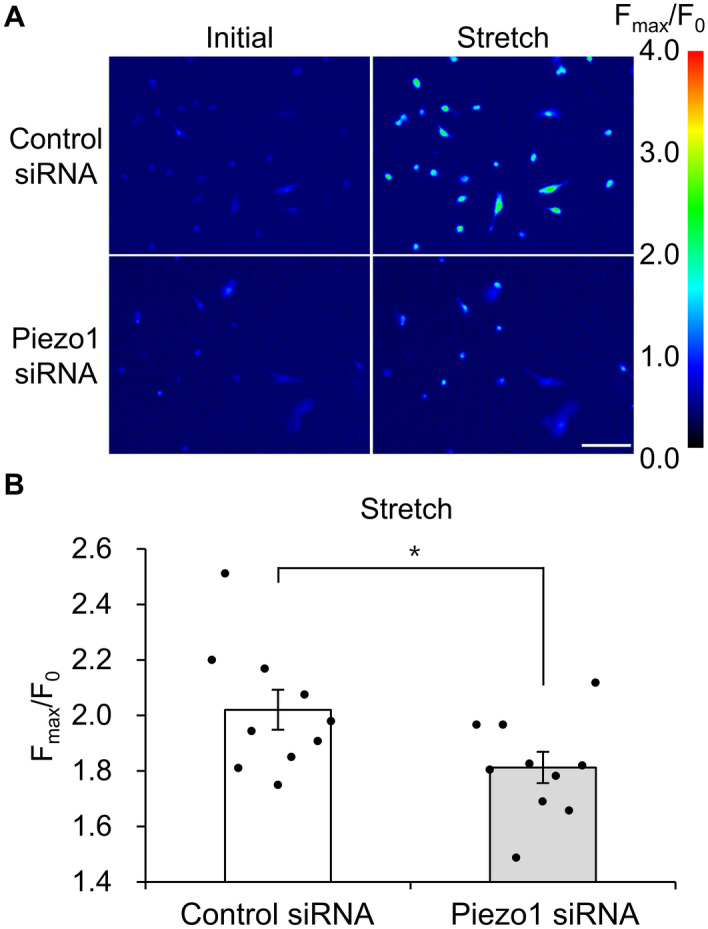
Intracellular Ca2+ changes upon stretch stimulation in human trabecular meshwork cells. (A) Images of intracellular Ca2+ changes upon stretch stimulation in human trabecular meshwork cells treated with control or Piezo1 siRNAs. Scale bar, 100 μm. (B) Intracellular Ca2+ levels following a single uniaxial stretch stimulation (1 way/s, 3.0 s pause) in human trabecular meshwork cells transfected with control or Piezo1 siRNA. Data are expressed as means ± SE (n = 5 experiments; ≥ 100 cells/experiment). *p < 0.05, two-tailed Student’s t-test.
Mechanical stretch stimulation in hTM cells promotes PGE2 release
We then performed lipid analysis to investigate which lipid mediators are produced by hTM cells following mechanical stretch stimulation. We found that mechanical stretch stimulation significantly increased the amounts of arachidonic acid and PGE2 released from hTM cells (Fig. 4A,B). The amount of arachidonic acid in the cell supernatant after 10, 30, 60 min in the Control (No stretch) group was 2533.62 ± 111.49 pg/ml, 2864 ± 295.56 pg/ml, and 3437 ± 212.68 pg/ml, and in the Stretch group was 3309.48 ± 84.64 pg/ml, 4716.96 ± 258.83 pg/ml, and 5593.08 ± 331.87 pg/ml. The amount of PGE2 in the cell supernatant after 10, 30, 60 min in the Control (No stretch) group was 32.96 ± 4.24 pg/ml, 29.13 ± 1.60 pg/ml, and 28.69 ± 2.89 pg/ml, and in the Stretch group was 196.70 ± 15.15 pg/ml, 176.80 ± 21.27 pg/ml, and 122.40 ± 3.85 pg/ml. PGD2 release was not significantly altered by stretch (Fig. 4C), and other lipid mediators such as PGF2α and TXB2 were not detected. Moreover, PIEZO1 silencing in hTM cells significantly abrogated the mechanical stretch stimulation-induced release of arachidonic acid and PGE2 (Fig. 5A,B). The amount of lipid mediators in the supernatant of Conrol siRNA-treated cells by stretch stimulation was 3382.93 ± 96.34 pg/ml of arachidonic acid and 49.19 ± 1.10 pg/ml of PGE2, while that of Piezo1 siRNA-treated cells was 2622.05 ± 117.42 pg/ml of arachidonic acid and 41.35 ± 2.56 pg/ml of PGE2. These results suggesting that Piezo1 plays an essential role in the release of arachidonic acid and PGE2 following stretch stimulation.
Figure 4.
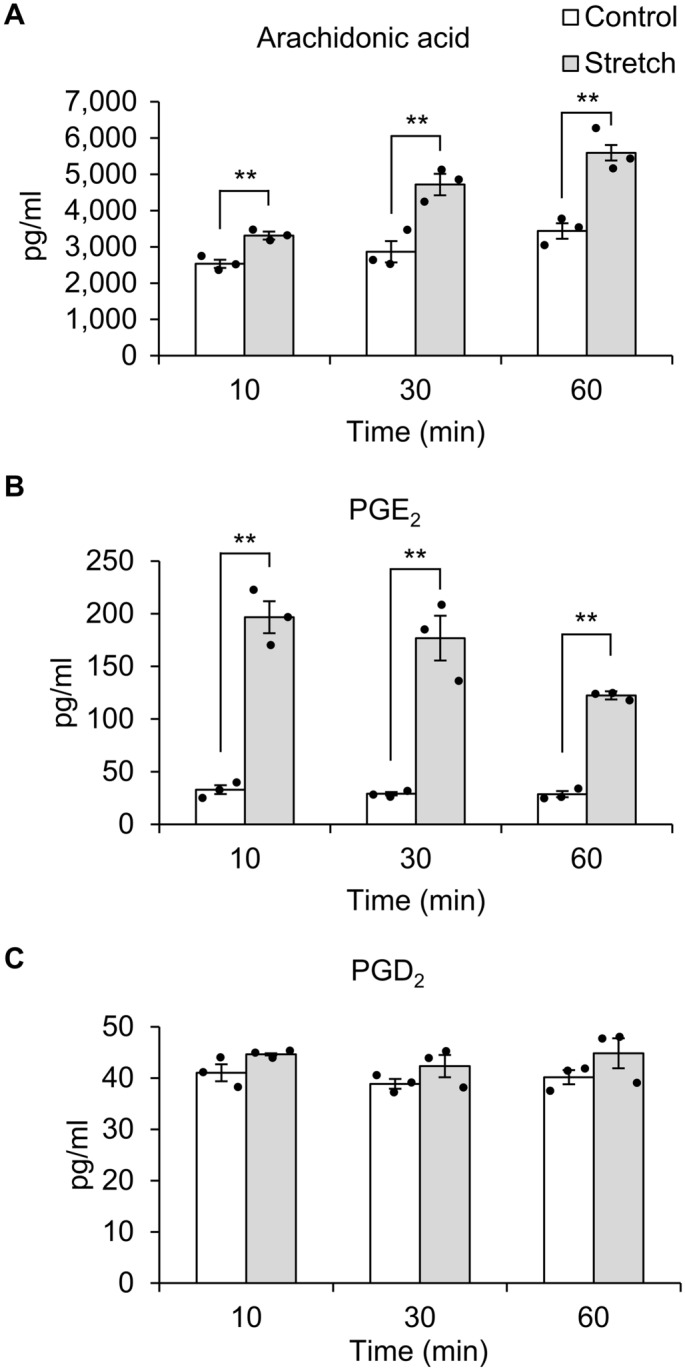
Mechanical stretch stimulation induces the release of arachidonic acid and PGE2 in human trabecular meshwork cells. Cell supernatants were collected 10, 30, and 60 min after a single uniaxial stretch stimulation and subjected to lipid analysis. Mechanical stretch stimulation in human trabecular meshwork cells triggered the release of arachidonic acid (A) and PGE2 (B), but not PGD2 (C). Data are expressed as means ± SE (n = 3 samples of lipid mediators extracted from cell supernatant in 3 chambers). **p < 0.01, two-tailed Student’s t-test.
Figure 5.
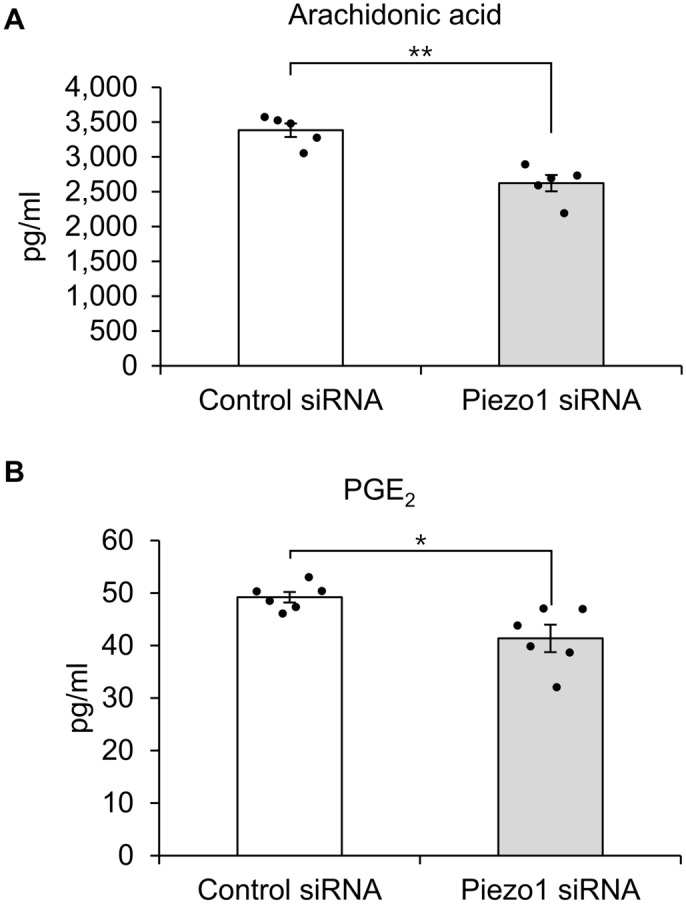
The impact of Piezo1 knockdown on the release of arachidonic acid and PGE2 following mechanical stretch stimulation in human trabecular meshwork cells. In human trabecular meshwork cells transfected with control or Piezo1 siRNAs, Cell supernatants were collected 10, 30, and 60 min after stretch stimulation. The levels of arachidonic acid (A), PGE2 (B) in the cell supernatant were measured. Piezo1 knockdown suppressed the release of arachidonic acid (A) and PGE2 (B). Data are expressed as mean ± SE (n = 3 samples of lipid mediators extracted from cell supernatant in 3 chambers). *p < 0.05. **p < 0.01, two-tailed Student’s t-test.
Yoda1 promotes PGE2 release in hTM cells
To assess the direct effect of Piezo1 activation, we measured lipid mediators in hTM cell supernatants after Yoda1 treatment. This analysis revealed that Yoda1 treatment significantly increased the levels of arachidonic acid and PGE2 released from hTM cells, but not PGD2 (Fig. 6). The amount of arachidonic acid in the cell supernatant after 10, 30, 60 min in the Control (DMSO) group was 2282.58 ± 3.20 pg/ml, 2539.86 ± 57.62 pg/ml, and 2620.76 ± 131.49 pg/ml, and in the Yoda1 group was 4500.13 ± 191.46 pg/ml, 5176.31 ± 89.15 pg/ml, and 7463.60 ± 188.21 pg/ml. The amount of PGE2 in the cell supernatant after 10, 30, 60 min in the Control (DMSO) group was 46.82 ± 2.27 pg/ml, 46.39 ± 3.43 pg/ml, and 35.28 ± 4.09 pg/ml, and in the Yoda1 group was 102.27 ± 1.12 pg/ml, 106.83 ± 4.52 pg/ml, and 100.25 ± 2.75 pg/ml. No other lipid mediators were detected. These data suggest that Piezo1 activation triggers the secretion of arachidonic acid and PGE2.
Figure 6.
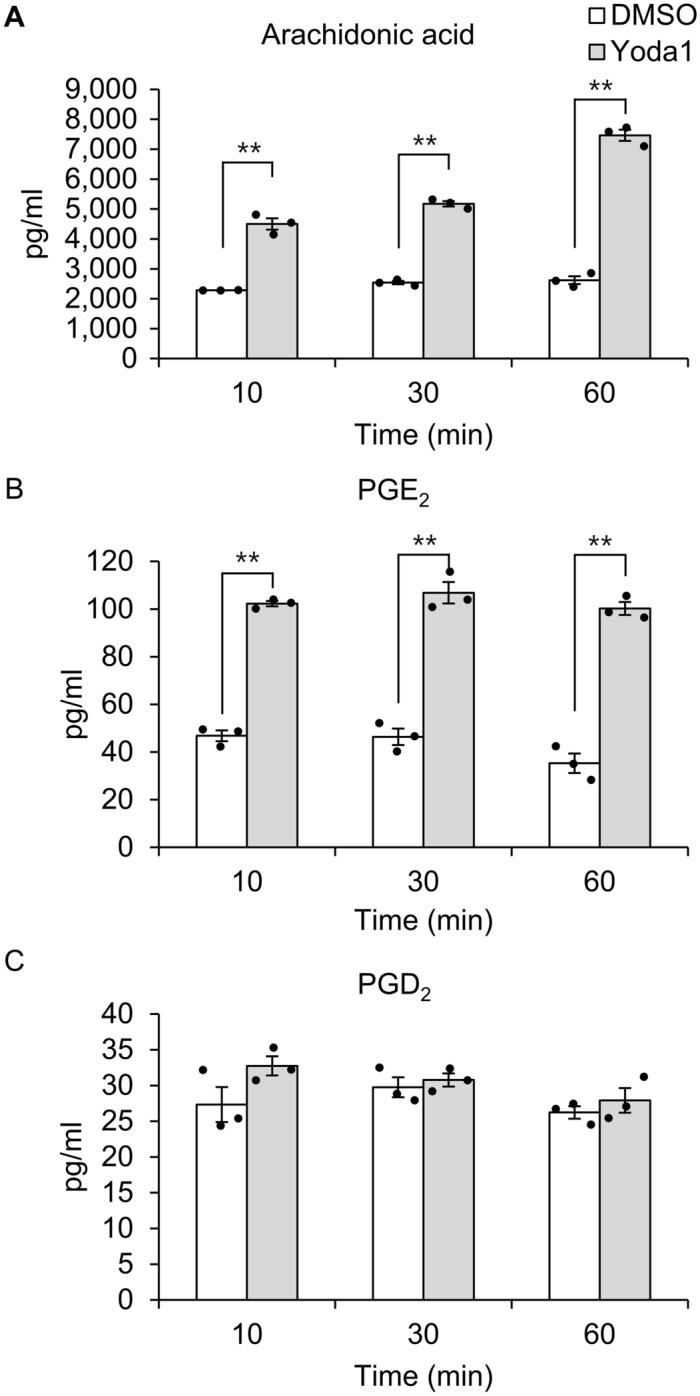
Direct activation of Piezo1 by Yoda1 induces the release of arachidonic acid and PGE2 in human trabecular meshwork cells. Human trabecular meshwork cells were incubated for the indicated time with or without Yoda1, and lipid mediators in the medium were analyzed. Yoda1 triggered the release of arachidonic acid (A) and PGE2 (B), but not PGD2 (C) in human trabecular meshwork cells. Data are expressed as the means ± SE (n = 3 samples of lipid mediators extracted from cell supernatant in 3 chambers). **p < 0.01, two-tailed Student’s t-test.
Effects of Yoda1 and PGE2 on collagen gel contraction
To investigate the effect of Yoda1 and the arachidonic acid metabolite PGE2 on hTM cells contraction, we performed an in vitro collagen gel contraction assay. This assay correlates that cells have contraction ability, which is supported by the fact that collagen alone does not cause contraction21,22. Compared to controls, Yoda1 significantly suppressed collagen gel contraction after 48 h and later, and PGE2 at 10 and 100 nM PGE2 after 24 h and later (Fig. 7A,B).
Figure 7.

Effect of Yoda1 and PGE2 on human trabecular meshwork cell-mediated collagen gel contraction. Changes in the collagen gel area in presence of Yoda1 (A) and PGE2 (B). Data are expressed as means ± SE (n = 4 gels separated from 4 wells). *p < 0.05. **p < 0.01, two-tailed Student’s t-test or Dunnett’s test.
Discussion
In this study, we demonstrated that Piezo1 was abundantly expressed in primary hTM cells, mediating mechanical stretch-induced Ca2+ influx and PGE2 release. Additionally, the Piezo1 agonist Yoda1 triggered Ca2+ influx and PGE2 release in hTM cells. Piezo1 activation in response to mechanical stimuli and subsequent PGE2 release suppressed hTM cell contraction. These results indicate that Piezo1 plays a crucial role in intracellular signaling induced by mechanical stretch and regulating TM contraction.
First, we analyzed the expression of TRPV1-6 and PIEZO1-2 in hTM cells and found that Piezo1 was most abundantly expressed at the mRNA level. Consistently, previous studies suggested that Piezo1 was one of 11 mechanotransduction channels identified in TM tissue and isolated TM cells23. Therefore, we hypothesized that Piezo1 might play a crucial role in TM physiology.
Next, we explored the role of Piezo1 signaling pathway in regulating TM contraction. As shown in Fig. 3, mechanical stretch-induced Ca2+ influx was inhibited when PIEZO1 was silenced. This finding is consistent with previous findings in urothelial cells12, red blood cell13, and neural stem cells14, suggesting that Piezo1 constitutes Ca2+ influx pathway in mechanical stretch of hTM cells. Arachidonic acid and its metabolites have been reported to play important roles in mechanical signaling15,16. Additionally, cyclic mechanical stress induced the release of arachidonic acid and its metabolic product PGE2 in porcine TM cells24. Therefore, we next investigated the profile of lipid mediators released after mechanical stretch and found a significant increase in the levels of arachidonic acid and PGE2 secreted from primary hTM cells following mechanical stretch (Fig. 4). We further found that the stretch-induced secretion of arachidonic acid and PGE2 was abrogated by PIEZO1 silencing (Fig. 5), and pharmacological activation of Piezo1 with Yoda1 triggered the release of arachidonic acid and PGE2 in hTM (Fig. 6), suggesting an essential role of Piezo1 in the mechanical stretch-induced secretion of these lipid mediators.
Moreover, we found that PGE2 produced by Piezo1 activation inhibited hTM contraction. The contraction of TM cells decreases the size of intercellular spaces and permeability, resulting in reduces aqueous humor outflow25. Agents that induce contraction of TM cells have been shown to increase IOP by reducing the outflow rate of aqueous humor, whereas relaxation of TM cells increases outflow and lowers IOP26–30. Consistent with these, EP2 agonists inhibited gel contraction29 and have an IOP lowering effect in vivo18,30,31. The response of TM cells by mechanical stimulation leading to the release of PGE2 may corroborate that recently introduced EP2 agonist, omidenepag, and previously reported EP2 agonists can significantly reduce IOP30,31. Our result suggests new homeostatic mechanism in the eye that elevated IOP stretches TM and subsequently TM cells respond to protect eyes through enhancing aqueous outflow via endogenously produced PGE2. The proposed mechanism underlying Stretch-mediated Piezo1 activation in human trabecular meshwork cells is diagrammatically summarized in Fig. 8.
Figure 8.
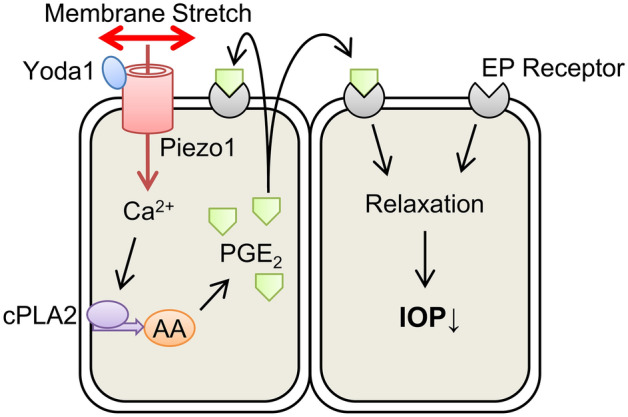
Proposed model of how Piezo1 activation contributes IOP regulation in human trabecular meshwork cells. Membrane stretch activates Piezo1 on hTM cells, causing Ca2+ influx. This increase in intracellular Ca2+ activates cytosolic phospholipase A2 (cPLA2) and releases arachidonic acid (AA) from membrane phospholipids. The liberated arachidonic acid is metabolized to produce PGE2, which causes cell relaxation via EP receptor, leading to a decrease in IOP.
To assess the effect of direct activation of Piezo1 in hTM cells, we used the Piezo1-selective activator Yoda1. Yoda1 was identified among approximately 3.25 million compounds as a synthetic small molecule that acts as an agonist of both human and mouse Piezo132. We investigated the effect of Yoda1 on Piezo1 activation in hTM cells and confirmed that it can be reliably activated at 10 μM (Fig. 2). Previous studies showed that high concentration (10–100 μM) of Yoda1 simultaneously caused substantial cytotoxicity and compromise of the HUVEC monolayer after 24 h33. In this study, we found that 10 μM Yoda1 reduced survival by approximately 10% after 24 h (See Supplementary Fig. S1 online). Although 10 μM Yoda1 significantly inhibited hTM cell contraction after 48–72 h in the gel contraction assay (Fig. 7A), we also showed the effect of PGE2 in gel contraction assay because of prolonged 10 μM Yoda1exposure (72 h) not only activates Piezo1 but may also have toxic effects. As shown in Fig. 7B, PGE2 significantly inhibited hTM collagen gel contraction in a dose-dependent manner, suggesting PGE2 production after Piezo1 activation contributes to IOP regulation. On the other hand, activation of EP2 (and EP1, EP4) receptors have been demonstrated to increase TM cell contractility34. However, this result is a response within 1 h of agonist exposure. PGE2 lowers IOP, and also temporarily increases IOP in the early stages after instillation18. The increase in TM cell contractility due to activation of EP2 receptor may reflect this transient increase in IOP and change at later time. EP2 receptor was also expressed in Schlemm's canal (SC) and its activation was shown to reduce SC cell stiffness34. Accordingly, PGE2 released from hTM cells may target SC in addition to TM.
While our findings highlighted the critical role of Piezo1 in IOP regulations upon TM stretch, there were several limitations in the present study. Piezo1 and TRPV4 were compared only for mRNA expression in this study. There are several reports comparing the role of Piezo1 and TRPV4 in response to stretch stimulation. In bladder urthelium, the sensitivity of Piezo1 to stretch stimuli was higher than that of TRPV412. In chondrocyte, Piezo1 was largely responsible for the stretch-activated current, whilst TRPV4 had no involvement in the specific mechanoelectrical transduction pathway35. However, there are no reports comparing the importance of Piezo1 and TRPV4 in TM. Thus, future studies are required to compare the sensitivity of Piezo1 and TRPV4 to stretch stimuli, as well as the relationship between Piezo1 and TRPV4 mRNA levels and function. We focused on lipid mediator and its involvement in the TM contraction, and didn’t further explore whether Piezo1 are involved in the stretch-induced cytoskeletal remodeling in hTM cells, which should be determined in the future study. In addition to lipid mediators, nitric oxide (NO) is a candidate that link intracellular Ca2+ elevation and regulation of TM outflow. NO is formed by the enzyme called nitric oxide synthase (NOS) and Three different isoforms have been identified including neuronal NOS or brain NOS (nNOS, bNOS or NOS1), endothelial NOS (eNOS or NOS3), and inducible NOS (iNOS or NOS2). nNOS1 and eNOS are constitutively expressed in cells and are activated by an increase in intracellular Ca2+36. iNOS is induced by a variety of stimuli, such as endotoxins and inflammatory cytokines, regardless of Ca2+ level. Three isoforms of NOS has been reported to be expressed in TM37. Several studies in vitro and in vivo have demonstrated IOP-lowering effects of NO, and treatment with the ROCK inhibitor Y-26732 increased the expression of eNOS and NO production more significantly38,39. Increased visual field damage results in decreased eNOS expression and Ca2+-dependent NOS activity in TM of POAG patients40. Thus, elevated intracellular Ca2+ in TM may increase eNOS expression and NO production, leading to decreased IOP. In this study, we applied a uniaxial stretch stimuli to Piezo1 knockdown cells, but by applying a multiaxial stretch stimuli to knockout cells, the exact contribution of Piezo1 in the cellular response to stretch stimuli could be obtained under more physiological conditions. Moreover, since our conclusions are based solely on evidence from in vitro experiments, we couldn’t determine the pathophysiological relevance of our in vitro observations in regulation of IOP or outflow facility. Further study using animal models or perfusion experiment will be needed to determine this issue.
In conclusion, this study clarified the role of new aqueous humor outflow regulation mediated by Piezo1 in TM. The present data has shown that Piezo1 regulates Ca2+ influx in response to mechanical stretching and that PGE2 is essential for Piezo1-mediated hTM cell contraction inhibition. Our study revealed a novel role for Piezo1 in hTM cells and proposed a new therapeutic strategy for the conventional outflow pathway.
Methods
Cell culture
Primary human trabecular meshwork (hTM) cells were isolated from human corneal scleral rims obtained from the Rocky Mountain Lions Eye Bank from 8 donors (both male and female, the mean age ± SD was 47.3 ± 25.9), as described previously41. All experiments were conducted according to the principles of the Declaration of Helsinki and approved by the ethics committee of the University of Tokyo. Written informed consent was obtained from donors. Briefly, corneal scleral rims maintained in Optisol were cut and divided into 6 to 8 sections. The trabecular meshwork strip was picked and isolated with forceps from sections. TM strips were cultured in a dish coated with collagen gel (Cellmatrix Type I-A, Nitta Gelatin Inc., Osaka, Japan). After the TM cells migrated from the tissue, the gel was digested with collagenase, and the collected TM cells were grown on dishes coated with 1 μg/mL fibronectin. hTM cells from passages 3 to 6 were used in subsequent experiments and maintained in Trabecular Meshwork Cell Medium (ScienCell Research Laboratories, Carlsbad, CA) ) containing 2% fetal bovine serum (ScienCell), 1% Trabecular Meshwork cell growth supplement (ScienCell) and 1% penicillin/streptomycin solution (ScienCell). The identity of the isolated cells was confirmed by the dexamethasone-induced upregulation of MYOC. After 7 days of exposure to vehicle (DMSO), 100 or 500 nM dexamethasone, dexamethasone enhanced MYOC expression (see Supplementary Fig. S2 online). Cells were transfected with PIEZO1-targeting siRNAs (SASI_Hs01_00208584, SASI_Hs01_00208585, Merck, Darmstadt, Germany) or MISSION siRNA Universal Negative Control #1 siRNAs (Merck). siRNA transfections were performed using MISSION siRNA Transfection Reagent (Merck) according to the manufacturer’s instructions.
Quantitative real-time polymerase chain reaction (qPCR) analysis
Total RNA was extracted from hTM cells at 24 h after siRNA transfection using ISOGEN (Nippon Gene, Tokyo, Japan). The concentration of Total RNA was measured by the absorbance at a wavelength of 260 nm using a spectrophotometer NanoDrop 2000 (Thermofisher, Waltham, MA). Complementary DNA (cDNA) was prepared using ReverTra Ace qPCR RT Master Mix with gDNA remover (Toyobo, Osaka, Japan). Quantitative real-time PCR was run on a Thermal Cycler Dice Realtime System (Takara Bio Inc., Shiga, Japan) using SYBR Premix Ex Taq™ II (Tli RNaseH Plus) (Takara Bio Inc.). Standard curves were used to determine mRNA transcript copy number in individual reactions. Primers specific for TRPV1-6 and PIEZO1-2 were designed by us or purchased from Takara Bio Inc., as detailed in Table 1.
Table 1.
Primers specific for TRPV1-6, PIEZO1-2, GAPDH, and MYOC.
| Gene Name | Primer | Sequences (5′–3′) | Product Size(bp) |
|---|---|---|---|
| TRPV1 | Forward | AAGTTCCTGCTGCAGAACTC | 130 |
| Reverse | TGCTCGTCACAAACTTCGTG | ||
| TRPV2 | Forward | AAACTGCTGCAGGCGAAATG | 95 |
| Reverse | GCAACAGCGGTGAAGATGAAC | ||
| TRPV3 | Forward | GTTAGCTACCCGCATTAAGCCTGA | 107 |
| Reverse | AGCAATTCTGGAATTCCCAGCTC | ||
| TRPV4 | Forward | TGCATGCGCCACCATTTTTG | 114 |
| Reverse | TATTGAGCACCGGCAAATCC | ||
| TRPV5 | Forward | GGAGACCTAATGCGTTTCTGCTG | 197 |
| Reverse | AAGGGCAAGTCCACGTCGTA | ||
| TRPV6 | Forward | AGCAGTGCCAATTGGGAAAG | 121 |
| Reverse | TGAGAACACGCAGTCAGATCTG | ||
| PIEZO1 | Forward | ATGCCAACGAGAAGCACATG | 128 |
| Reverse | ACGGATGTACTTGGGGAAGAG | ||
| PIEZO2 | Forward | ATTTCATTGTGCGGCCCAAC | 127 |
| Reverse | TTGTCACCTGCCATGATTCG | ||
| GAPDH | Forward | AATTCCATGGCACCGTCAAG | 104 |
| Reverse | ATCGCCCCACTTGATTTTGG | ||
| MYOC | Forward | TACACGGACATTGACTTGGC | 159 |
| Reverse | ATTGGCGACTGACTGCTTAC |
Calcium imaging and mechanical cell stretch stimulation
hTM cells or those 24 h after siRNA transfection were seeded on cover glass chambers (Iwaki, Shizuoka, Japan) or stretch chamber STB-CH-24 (STREX Inc., Osaka, Japan) coated with 1 μg/mL fibronectin. After washing with PBS, hTM were loaded with 0.1% BSA (Sigma) and 5 μM fluorescent Ca2+ indicator Fluo-8 AM (AAT Bioquest, Inc., Sunnyvale, CA) for 20 min. Subsequently, 10 μM Yoda1 (Cayman Chemical, Ann Arbor, MI)32 was applied directly to the cover glass chambers. STB-CH-24 was attached to a stretch device STB-150 (STB-150 in this study was modified to stretch up to 40% using STB-CH-24; STREX Inc.) on the stage of a fluorescence microscope and subjected to a single uniaxial stretch with a nominal 30% extension (1 way/s, 3.0 s pause) at room temperature. As evaluated from images of the chamber at 30% extension, the actual extension of the cell culture surface was about 20%. Fluorescent images were acquired under a fluorescence microscope (Keyence, Osaka, Japan). The fluorescence intensity of Fluo-8 was quantified using ImageJ software (https://imagej.nih.gov/ij/download.html). First, the captured images were stacked and the misalignment was corrected with the StackReg plugin. Next, the image with the highest fluorescence intensity from the cells was binarized by setting a threshold value by the triangle method, and the cells were selected with Analyze particles. Finally, the fluorescence intensity of the selected cells was measured on all images. The change ratio (FT/F0, Fmax/F0) was calculated using peak and basal values. FT, Fmax, and F0 represent the fluorescence intensity at that time, the maximum fluorescence intensity, and the fluorescence intensity before stimulation, respectively.
Lipid analysis
Lipid analysis was performed as previously described42. Cell supernatant was collected 10, 30, and 60 min after 10 μM Yoda1 treatment or stretch stimulation, and stored at − 80 ℃. Methanol was added to each sample at a ratio of 1:1, and 10 μL of internal standard was added. The internal standard contained 6-keto-PGF1α-d4, AEA-d4, arachidonic acid-d8, docosahexaenoic acid-d5, eicosapentaenoic acid-d5, OEA-d4, tetranor-PGEM-d6, TxB2-d4, PGF2α-d9, PGE2-d4, PGD2-d4, LTB4-d4, LTC4-d5, LTD4-d5, LTE4-d5, 5(S)-HETE-d8, 12-HETE-d8, 15(S)-HETE-d8, and PAF-C16-d4 (Cayman Chemical). After vortexing, samples were centrifuged at 15,000×g and 4 °C for 15 min to obtain phase separation. Supernatants were transferred into 15 mL glass tubes. The supernatant and 0.03% formic acid were mixed at a ratio of 1:4 with vortex. Samples were loaded onto a solid-phase extraction column (Oasis HLB, Waters Corporation, Milford, MA) pre-conditioned with methanol and 0.03% formic acid. The column was centrifuged at 800 rpm and 4 °C for 2 min, and washed with petroleum ether. Methanol containing 0.2% formic acid was added to the column, followed by centrifugation at 800 rpm and 4 °C for 2 min to elute lipids. After evaporation, eluents were reconstituted in methanol.
The samples were analyzed on a Nexera ultra-high performance liquid chromatograph connected to a triple quadrupole mass spectrometer LCMS-8060 (Shimazu, Kyoto, Japan). Chromatographic separation was performed using a Kinetex C8 column (2.6 μm, 2.1 × 150 mm, Phenomenex, Torrance, CA). The mass spectrometer was operated in selected reaction monitoring mode allowing for simultaneous detection of target lipid mediators. Quantification was performed by internal standard calibration method using chromatographic peak areas, as previously described42.
Gel contraction assay
Gel contraction assay was performed using the Collagen Gel Culturing Kit (Nitta Gelatin, Inc., Osaka, Japan) as per the manufacturer’s instructions. Collagen type I, 10 × MEM, and reconstitution buffer (pH 7.3) were mixed at a ratio of 7:1:1 at 4 °C. The final hTM cell concentration was adjusted to 1 × 106 cells/mL. The resultant mixture and cell suspension were mixed at a 9:1 ratio and 500 μL of the mixture was added to each well of 24-well plates and incubated at 37 °C to allow for collagen gel formation. After 60 min, gels were placed in 6-cm dishes containing 5 mL Trabecular Meshwork Cell Medium with DMSO, 10 μM Yoda1, or PGE2 (1, 10, or 100 nM). After 0, 12, 24, 48, and 72 h, gels were imaged with a gel imaging device, and the area was quantified with ImageJ software (https://imagej.nih.gov/ij/download.html).
Statistical analysis
Data were represented as mean ± standard deviation (SD) in Fig. 1, or as mean ± standard error (SE) in Figs. 2, 3, 4, 5, 6, 7 and Supplementary Figure with individual data plotted. Statistical analyses were performed using a two-tailed Student’s t-test or Dunnett’s test. A difference was considered statistically significant when p < 0.05.
Supplementary Information
Acknowledgements
The authors thank Yasuko Kuwabara, Junko Nihsida, and Chiyo Morita for their technical assistance. The English in this document has been checked by at least two professional editors, both native speakers of English. For a certificate, please see: http://www.textcheck.com/certificate/LAOOmC. This work was supported by JSPS KAKENHI Grant Number 18K16946.
Author contributions
T.U., Y.K., M.H. and M.A. designed and directed the project. T.U., S.S., R.Y., S.M.T. and M.A. designed experiments. T.U., S.S., R.Y., S.M.T. conducted the experiments, T.U. wrote the main manuscript text, and created the figures. All authors reviewed the manuscript.
Competing interests
Takatoshi Uchida and Shota Shimizu are employees of the Senju Pharmaceutical Co.,Ltd. The other authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-021-83713-z.
References
- 1.Lütjen-Drecoll E. Functional morphology of the trabecular meshwork in primate eyes. Prog. Retin. Eye Res. 1999;18:91–119. doi: 10.1016/S1350-9462(98)00011-1. [DOI] [PubMed] [Google Scholar]
- 2.Acott TS, et al. Intraocular pressure homeostasis: Maintaining balance in a high-pressure environment. J. Ocul. Pharmacol. Ther. 2014;30:94–101. doi: 10.1089/jop.2013.0185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Coleman DJ, Trokel S. Direct-recorded intraocular pressure variations in a human subject. Arch. Ophthalmol. 1969;82:637–640. doi: 10.1001/archopht.1969.00990020633011. [DOI] [PubMed] [Google Scholar]
- 4.Grierson I, Lee WR. Light microscopic quantitation of the endothelial vacuoles in Schlemm's canal. Am. J. Ophthalmol. 1977;84:234–246. doi: 10.1016/0002-9394(77)90857-1. [DOI] [PubMed] [Google Scholar]
- 5.Grierson I, Lee WR. The fine structure of the trabecular meshwork at graded levels of intraocular pressure. (1) Pressure effects within the near-physiological range (8–30 mmHg) Exp. Eye Res. 1975;20:505–521. doi: 10.1016/0014-4835(75)90218-3. [DOI] [PubMed] [Google Scholar]
- 6.Stamer WD, et al. Biomechanics of Schlemm's canal endothelium and intraocular pressure reduction. Prog. Retin. Eye Res. 2015;44:86–98. doi: 10.1016/j.preteyeres.2014.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Carreon TA, Castellanos A, Gasull X, Bhattacharya SK. Interaction of cochlin and mechanosensitive channel TREK-1 in trabecular meshwork cells influences the regulation of intraocular pressure. Sci. Rep. 2017;7:452. doi: 10.1038/s41598-017-00430-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Yarishkin O, et al. TREK-1 channels regulate pressure sensitivity and calcium signaling in trabecular meshwork cells. J. Gen. Physiol. 2018;150:1660–1675. doi: 10.1085/jgp.201812179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Luo N, et al. Primary cilia signaling mediates intraocular pressure sensation. Proc. Natl. Acad. Sci. USA. 2014;111:12871–12876. doi: 10.1073/pnas.1323292111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ryskamp DA, et al. TRPV4 regulates calcium homeostasis, cytoskeletal remodeling, conventional outflow and intraocular pressure in the mammalian eye. Sci. Rep. 2016;6:30583. doi: 10.1038/srep30583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Coste B, et al. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science. 2010;330:55–60. doi: 10.1126/science.1193270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Miyamoto T, et al. Functional role for Piezo1 in stretch-evoked Ca2+ influx and ATP release in urothelial cell cultures. J. Biol. Chem. 2014;289:16565–16575. doi: 10.1074/jbc.M113.528638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cahalan SM, et al. Piezo1 links mechanical forces to red blood cell volume. Elife. 2015;4:e07370. doi: 10.7554/eLife.07370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Pathak MM, et al. Stretch-activated ion channel Piezo1 directs lineage choice in human neural stem cells. Proc. Natl. Acad. Sci. USA. 2014;111:16148–16153. doi: 10.1073/pnas.1409802111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Burkholder TJ. Mechanotransduction in skeletal muscle. Front. Biosci. 2007;12:174–191. doi: 10.2741/2057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Munaron L. Shuffling the cards in signal transduction: Calcium, arachidonic acid and mechanosensitivity. World J. Biol. Chem. 2011;2:59–66. doi: 10.4331/wjbc.v2.i4.59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ota T, Aihara M, Narumiya S, Araie M. The effects of prostaglandin analogues on IOP in prostanoid FP-receptor-deficient mice. Investig. Ophthalmol. Vis. Sci. 2005;46:4159–4163. doi: 10.1167/iovs.05-0494. [DOI] [PubMed] [Google Scholar]
- 18.Saeki T, Ota T, Aihara M, Araie M. Effects of prostanoid EP agonists on mouse intraocular pressure. Investig. Ophthalmol. Vis. Sci. 2009;50:2201–2208. doi: 10.1167/iovs.08-2800. [DOI] [PubMed] [Google Scholar]
- 19.Honjo M, et al. Autotaxin-lysophosphatidic acid pathway in intraocular pressure regulation and glaucoma subtypes. Investig. Ophthalmol. Vis. Sci. 2018;59:693–701. doi: 10.1167/iovs.17-23218. [DOI] [PubMed] [Google Scholar]
- 20.Stamer WD, Read AT, Sumida GM, Ethier CR. Sphingosine-1-phosphate effects on the inner wall of Schlemm's canal and outflow facility in perfused human eyes. Exp Eye Res. 2009;89:980–988. doi: 10.1016/j.exer.2009.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Girão-Silva T, et al. Short-term mechanical stretch fails to differentiate human adipose-derived stem cells into cardiovascular cell phenotypes. Biomed. Eng. Online. 2014;13:54. doi: 10.1186/1475-925X-13-54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ying L, et al. The transient receptor potential vanilloid 4 channel modulates uterine tone during pregnancy. Sci. Transl. Med. 2015;7:319–204. doi: 10.1126/scitranslmed.aad0376. [DOI] [PubMed] [Google Scholar]
- 23.Tran VT, Ho PT, Cabrera L, Torres JE, Bhattacharya SK. Mechanotransduction channels of the trabecular meshwork. Curr. Eye Res. 2014;39:291–303. doi: 10.3109/02713683.2013.842593. [DOI] [PubMed] [Google Scholar]
- 24.Luna C, et al. Extracellular release of ATP mediated by cyclic mechanical stress leads to mobilization of AA in trabecular meshwork cells. Investig. Ophthalmol. Vis. Sci. 2009;50:5805–5810. doi: 10.1167/iovs.09-3796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Tamm ER. The trabecular meshwork outflow pathways: Structural and functional aspects. Exp. Eye Res. 2009;88:648–655. doi: 10.1016/j.exer.2009.02.007. [DOI] [PubMed] [Google Scholar]
- 26.Wiederholt M, et al. Regulation of outflow rate and resistance in the perfused anterior segment of the bovine eye. Exp. Eye Res. 1995;61:223–234. doi: 10.1016/S0014-4835(05)80042-9. [DOI] [PubMed] [Google Scholar]
- 27.Koga T. Rho-associated protein kinase inhibitor, Y-27632, induces alterations in adhesion, contraction and motility in cultured human trabecular meshwork cells. Exp. Eye Res. 2006;82:362–370. doi: 10.1016/j.exer.2005.07.006. [DOI] [PubMed] [Google Scholar]
- 28.Rao PV, Deng PF, Kumar J, Epstein DL. Modulation of aqueous humor outflow facility by the Rho kinase-specific inhibitor Y-27632. Investig. Ophthalmol. Vis. Sci. 2001;42:1029–1037. [PubMed] [Google Scholar]
- 29.Kalouche G, et al. Activation of prostaglandin FP and EP2 receptors differently modulates myofibroblast transition in a model of adult primary human trabecular meshwork cells. Investig. Ophthalmol. Vis. Sci. 2016;57:1816–1825. doi: 10.1167/iovs.15-17693. [DOI] [PubMed] [Google Scholar]
- 30.Kirihara T, et al. Pharmacologic characterization of omidenepag isopropyl, a novel selective EP2 receptor agonist, as an ocular hypotensive agent. Investig. Ophthalmol. Vis. Sci. 2018;59:145–153. doi: 10.1167/iovs.17-22745. [DOI] [PubMed] [Google Scholar]
- 31.Fuwa M, et al. Effects of a novel selective EP2 receptor agonist, omidenepag isopropyl, on aqueous humor dynamics in laser-induced ocular hypertensive monkeys. J. Ocul. Pharmacol. Ther. 2018;34:531–537. doi: 10.1089/jop.2017.0146. [DOI] [PubMed] [Google Scholar]
- 32.Syeda R, et al. Chemical activation of the mechanotransduction channel Piezo1. Elife. 2015;4:e07369. doi: 10.7554/eLife.07369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Davies JE, et al. Using Yoda-1 to mimic laminar flow in vitro: A tool to simplify drug testing. Biochem. Pharmacol. 2019;168:473–480. doi: 10.1016/j.bcp.2019.08.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Wang JW, Woodward DF, Stamer WD. Differential effects of prostaglandin E2-sensitive receptors on contractility of human ocular cells that regulate conventional outflow. Investig. Ophthalmol. Vis. Sci. 2013;54:4782–4790. doi: 10.1167/iovs.13-12363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Servin-Vences MR, Moroni M, Lewin GR, Poole K. Direct measurement of TRPV4 and PIEZO1 activity reveals multiple mechanotransduction pathways in chondrocytes. Elife. 2017;6:e21074. doi: 10.7554/eLife.21074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Förstermann U, Sessa WC. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012;33:829–837. doi: 10.1093/eurheartj/ehr304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wu RY, Ning MA. Expression of nitric oxide synthase and guanylate cyclase in the human ciliary body and trabecular meshwork. Chin. Med. J. 2012;125:129–133. [PubMed] [Google Scholar]
- 38.Schmetterer L, Polak K. Role of nitric oxide in the control of ocular blood flow. Prog. Retin. Eye Res. 2001;20:823–847. doi: 10.1016/S1350-9462(01)00014-3. [DOI] [PubMed] [Google Scholar]
- 39.Kim JW. Comparative study of the effects of trabecular meshwork outflow drugs on the permeability and nitric oxide production in trabecular meshwork cells. Korean J. Ophthalmol. 2017;31:452–459. doi: 10.3341/kjo.2017.0020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Fernández-Durango R, et al. Expression of nitrotyrosine and oxidative consequences in the trabecular meshwork of patients with primary open-angle glaucoma. Investig. Ophthalmol. Vis. Sci. 2008;49:2506–2511. doi: 10.1167/iovs.07-1363. [DOI] [PubMed] [Google Scholar]
- 41.Pattabiraman PP, Rao PV. Mechanistic basis of Rho GTPase-induced extracellular matrix synthesis in trabecular meshwork cells. Am. J. Physiol. Cell Physiol. 2010;298:C749–C763. doi: 10.1152/ajpcell.00317.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Yamada M, et al. A comprehensive quantification method for eicosanoids and related compounds by using liquid chromatography/mass spectrometry with high speed continuous ionization polarity switching. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2015;995–996:74–84. doi: 10.1016/j.jchromb.2015.05.015. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.