

Abstract
In hepatocellular carcinoma patients, due to the microenvironmental specificity of liver, the tumor microenvironment exhibits high immunosuppression and drug resistance, resulting in excessive or insufficient responses to immunotherapy. The dynamic interactions between tumor cells and immune modulators in the TME significantly impact the occurrence and development of tumors, efficacy, and drug resistance, which can create a much more positive response to immunotherapy. Moreover, with the wide application of single-cell sequencing technology in the TME, increasing evidence shows an interaction network among cells. Sequencing results suggest that specific tumor-associated macrophages are a hub node, connecting different cell populations in the cell interaction network, and can could regulate tumor generation and antitumor immunity. This review focused on therapeutic targets that could be targeted to remodel the tumor microenvironment and reprogram the tumor-associated macrophage phenotype in hepatocellular carcinoma patients, thereby improving immunotherapeutic efficacy.
Keywords: Tumor microenvironment, Hepatocellular carcinoma, Tumor-associated macrophages, Immunotherapy, Immune checkpoint, Antitumor immunity
Background
Hepatocellular carcinoma (HCC), accounting for more than 80% of primary liver cancer cases [1], is one of the most common malignant solid tumors and the fourth most frequent cause of cancer-related mortality worldwide [2]. Furthermore, the international trend for liver cancer incidence is still grim. The incidence of liver cancer continues to increase and is rising more rapidly than that of other types of cancer. From 2007 through 2016, the rate of increase was approximately 2 to 3% annually. According to cancer statistics, in 2020, 42,810 new cases of liver and intrahepatic bile duct cancers and 30,160 liver cancer-related deaths occurred in the United States [3]. The most recent liver cancer statistical data in China estimated 370,000 new cases (274,000 males and 96,000 females) of liver cancer in 2015. For liver cancer-related mortality, there were 326,000 new deaths (242,000 males and 84,000 females) in China [4]. As the predominant form of liver cancer, HCC tends to occur in patients with a history of cirrhosis, with these patients accounting for more than 70% of the HCC patients population [5]. Major risk factors that contribute to fibrosis and cirrhosis include chronic hepatitis B virus (HBV) infection, chronic hepatitis C virus (HCV) infection, obesity, diabetes, excess alcohol consumption, and metabolic diseases [6, 7]. In the setting of cirrhosis, genetic mutations, epigenetic deregulation, and abnormal molecular signaling pathway transduction are the most common causes of hepatocellular carcinogenesis [8].
With the deepening of tumor immunology research, studies have revealed that the tumor microenvironment (TME) is an intricate ecosystem around cancer cells that supports carcinogenesis from cancer initiation to metastasis and is consistently modulated by cellular metabolism, genetic alterations, dysfunctional oncogenic signaling, and epigenetic factors. The first discovery of the link between chronic inflammation and oncogenesis and the observation of leukocytes in neoplastic tissues by Rudolf Virchow indicated the need to develop a broader understanding of the TME in solid malignant tissue [9]. The skeleton of this complex tissue microenvironment is built by the extracellular matrix (ECM) and vascular networks, and the gaps are filled with different kinds of cells. In addition to tumor cells, many types of stromal cells, fibroblast, adaptive and innate immune cells, and noncellular components, such as cytokines, signaling proteins, and growth factors, are found in the TME [10]. Furthermore, researchers have verified that intratumoral heterogeneity is significantly related to different immune microenvironments [11], which dramatically influences precision medicine and will be elaborated later.
More specifically, the tumor immune microenvironment (TIME), the stage for the interactions between tumor cells and immune cells, plays a fundamental and indispensable role in HCC evolution and greatly influences immunotherapy outcomes. Within the TIME, many immune cells have been found to accumulate during tumor progression, such as myeloid-derived suppressor cells (MDSCs), regulatory T (Treg) cells, and tumor-associated macrophages (TAMs), which are responsible for building of immunosuppressive environment. In contrast, natural killer (NK) cells, cytotoxic CD8+ T cells, and CD4+ T cells with a proinflammatory T helper 1 phenotype work together to counteract protumor effects. Recently, according to the degree of immune infiltrate, Mikhail Binnewies et al. divided the TIME into three classes: infiltrated-excluded (I-E) TIMEs, infiltrated-inflamed (I-I) TIMEs, and tertiary lymphoid structure (TLS) TIMEs [12]. I-E TIMEs are filled with immune cells but lack cytotoxic lymphocytes (CTLs) in the tumor core, while I-I TIMEs are significantly infiltrated with CTLs expressing programmed cell death 1 (PD-1), and leukocytes and tumor cells within I-I TIMEs are characterized by the expression of immune-dampening PD-1 ligand (PD-L1). TLS-TIMEs are a subclass of I-I TIMEs and have been observed to include TLSs containing lymphoid aggregates; their cell composition is similar to that in the lymph nodes. These classifications of the immune contents within the TME are the leading information to recognize different the immunological compositions and immune statuses of tumors (i.e., activated or suppressed); for instance, tumors with an I-E TIME are known as immunologically “hot” tumors, which may produce positive responsiveness to immunotherapy and affect overall survival.
Generally, surgery (liver resection or transplantation) and radiofrequency ablation (RFA) are the standard curative therapies for HCC. In addition, transarterial chemoembolization (TACE) or transarterial radioembolization (TARE) are promising locoregional therapies that can reduce mortality and improve patient quality of life [13, 14]. For unresectable or advanced HCC, targeted molecular therapies, such as multikinase vascular endothelial growth factor (VEGF) inhibitors, are believed to be the future of HCC treatment. In the past decade, sorafenib has been the first-line agent most widely used for systemic chemotherapy [15]. As an FDA-approved tyrosine kinase inhibitor (TKI), sorafenib targets multiple receptors including platelet-derived growth factor (PDGFR) and fibroblast growth factor (FGFR1). It inhibits VEGF signaling, thereby downregulating the proliferation, migration, and angiogenesis of hepatocellular carcinoma cells and thus significantly prolonging HCC patient overall survival [16]. This year, a phase 3 trial enrolling patients with hepatocellular carcinoma showed that the immunotherapeutic combination of atezolizumab and bevacizumab produced a better outcome than sorafenib. With a 67.2% OS rate at 12 months and 6.8 months as the mPFS time, atezolizumab-bevacizumab was superior to sorafenib in terms of 12-month OS and PFS outcomes, revealing a promising therapeutic strategy for unresectable HCC patients [17, 18].
Given that the abovementioned curative therapies are only suitable for patients with a limited tumor burden, TIME-based approaches tailored to individual patients provide a prospective HCC treatment. This review focuses on the novel and innovative targets in the TME for immune checkpoint blockade (ICB) agents in HCC. In addition, since macrophage infiltration in the TME is a critical link in the development of liver cancer induced by steatosis, we discuss the potential of TAMs as targets for HCC immunotherapy. Furthermore, an inhibitor of colony-stimulating factor (CSF)-1 receptor targeting TAMs was shown to exert significant antitumor effects on the mouse glioblastoma and improve the survival rate [19]. We discuss emerging methods that could reprogram the tumor-associated macrophage phenotype and remodel the TME in HCC patients, thereby enhancing the antitumor immune response.
Rationales for using the TME as a therapeutic basis in HCC
The HCC tumor immune microenvironment is involved in the infiltration of various innate and adaptive immune cells and the diverse expression of genes that affect cancer immune surveillance and the response to immunotherapy. Different tumor subtypes show different patterns for the immune microenvironment, which is partially caused by intratumoral heterogeneity. HCC and intrahepatic cholangiocarcinoma (iCCA) are the two clinically and pathologically distinct categories of liver cancer [20], gene expression analysis, single-cell RNA sequencing, flow cytometric analysis while through histological analysis, such as multiplex immunohistochemistry, and histological analysis, such as multiplex immunohistochemistry, [21], have shown that the cellular composition of the TIME varies significantly among different tumor subtypes [11, 22, 23]. Therefore, with reference to the classification of histopathological subtypes according to the characteristics of the stromal features of HCC (e.g., lymphocyte-rich HCC, sclerosing HCC, and steatohepatitis HCC) [24, 25], a comprehensive evaluation of the immune microenvironment could serve as a classification standard and benefit precision medicine by stratifying the target population for immunotherapy.
Characteristics of the TIME in HCC
In the tumor immune microenvironment, the composition of immune cells (including most innate and adaptive immune cells) is roughly the same, but the degree of infiltration of different immune cells varies greatly, as shown in Fig. 1(a). For prognosis, after surgical HCC resection, a higher CD8+ T cell/Treg ratio is essential for effective antitumor immunity [26]. Moreover, ectopic lymphoid follicles, highly ordered structures formed by immune cells, rarely occur in HCC [27]. Through estimating immune cell infiltration, researchers have divided HCC into immune-class HCC and exhausted-class HCC [23, 28]. Immune-class HCC is characterized by high expression of T cell and B cell genes, interferon (IFN)-related genes and PD-1/PD-L1. Furthermore, in active-immune-class HCC, in addition to the elevated expression of the listed genes, the fibroblast-related gene signature is significantly lower than that in exhausted-class HCC [29]. Furthermore, histopathologically, according to T cell, B cell, and lymphoplasmacytic infiltration, immunity in the HCC microenvironment can be divided into three levels: immune-high, immune-mid, and immune-low. High co-infiltration of immune cells is the feature of the immune-high subtype associated with a relatively good prognosis, while the immune-mid and immune-low subtypes lack B cells, plasma cells and other immune cells. Although the immune-high subtype of HCC, which shows high expression of IFN-related genes and PD-1/PD-L1 molecules similar to immune-class HCC [23], represents active antitumor immunity, this subtype has been observed in poorly differentiated HCC with a poor prognosis [30, 31]. This finding suggests that it is reasonable to evaluate the TIME as one of the criteria for judging the prognosis of HCC.
Fig. 1.

Dynamic changes in cellular components in the HCC TME. a According to cell functions in the TME, the TME can be refined into the tumor immune microenvironment (TIME). Due to the specific features of the liver, the role of tumor-associated macrophages (TAMs) in the liver TIME is prominent. As shown, during the development of HCC, the expression of immunosuppressive checkpoint molecules (Tim-3, PD-1, and TLR) on the surface of TAMs affects the accumulation and antigen presentation of cell performing immune surveillance. b Blocking immune checkpoint molecule expression and reversing the phenotype of macrophages are two main approaches to regulate the tumor immune microenvironment discussed in this review. As shown above, increased infiltration of T cells and M2 macrophages can effectively inhibit the growth, metastasis, and invasion of tumor cells at the cellular level, which in turn achieves superior immunotherapeutic efficacy
In addition, as a classic multistage carcinogenesis model, the TIME of HCC also constantly changes during tumor progression. From low-grade dysplastic nodules (LGDNs) to high-grade dysplastic nodules (HGDNs), B cell infiltration and lymphoid follicle formation increase significantly but then decrease during the progression from early HCC to advanced HCC [32]. Moreover, from moderately differentiated lesions to poorly differentiated lesions, the number of Treg cells increases, while the infiltration of NK cells and macrophages and the CD8+ /CD4+ T cells ratio decrease [33].
Pro- and antitumor effect of immune cells in the TIME of HCC
As an immunomodulatory organ under disease-free conditions, the liver is responsible for processing multiple antigens derived from the gut and portal vein blood circulation via its intrinsic immune tolerogenicity that inhibits inappropriate inflammatory responses. However, once this complex immune tolerance system is disrupted due to chronic inflammatory liver disease, the functions of innate and adaptive cells become dysregulated, and cancer cells escape immune surveillance, thereby facilitating liver tumor development [34, 35]. According to the cell differentiation source and function, immune cells in the TIME of HCC can be grouped into three categories: antitumor lymphocytes (CTLs and NK cells), immunosuppressive lymphocytes (Tregs), and immunosuppressive myeloid cells (TAMs, DCs, and MDSCs). Compared to the peripheral blood, the liver has a higher ratio of CD8+ T cells to CD4+ T cells [36]. However, CD8+ T lymphocytes, the tumor-infiltrating lymphocyte (TIL) subset known to be the primary antitumor effector population in liver cancer, also play a conflicting role in facilitating a chronic proinflammatory microenvironment and spontaneous immune surveillance. CD8+ CTLs directly contact and lyse cells via secretion of perforin, granzyme A, and granzyme B; they also mediate signal transduction through cell-surface Fas-Fas ligand interactions and secretion of IFN-γ and TNF [37].
On the one hand, CD8+ CTLs have been found to promote HCC development by producing lymphotoxin-α and lymphotoxin-β, which are related to the construction of ectopic lymphoid structures (one of the poor prognostic factors of HCC), and tumor progression was shown to be slowed down when CD8+ T cells and lymphotoxin-β were depleted in an HCC model. In comparison, the number of IFN-γ-secreting CD8+ T cells, which are responsible for cytotoxic interactions in HCC, is positively correlated with improved OS [38]. Similarly, in a specific HCC model involving overexpression of the hepatocyte-specific proto-oncoprotein c-Myc with a methionine and choline-inadequate diet, depletion of CD4+ T cells accelerated the development of HCC [39], but after HCC resection, tumor-infiltrating CD4+ CTLs are known to be independent predictors of disease-free survival. Treg cells infiltration has a negative correlation with the prognosis of HCC. Therefore, we may expect further study on the opposing role of different T cell subsets in different stages of HCC pathogenesis.
Although the function of T lymphocytes in the progression of HCC and that of TIME-infiltrating T cells remain controversial, communication between DCs and T cells is irreplaceable for in effective antitumor immunity. DCs are the professional antigen-presenting cells (APCs) that activate effector CD4+ and CD8+ T cells. One DC subtype, CD103+ DCs, is the primary source of the chemokines CXCL9 and CXCL10, two necessary chemokines responsible for recruiting CD8+ T cells into the TME. When CD103+ DCs are lacking in the tumor, T cell migration into the tumor is impaired, damaging antitumor responses [40]. Additionally, the largest population of resident macrophages (Kupffer cells (KCs)) in the liver constitutes another major APC population. By expressing scavenger receptors, Toll-like receptors (TLRs), and other receptors, KCs identify antigens and internalize pathogens and apoptotic cells. However, KC-induced antigen-presentation can also upregulate the expression of PD-L1 and the release of immunosuppressive molecules, such as IL-10, TGFβ, and prostaglandin E2 (PGE2), which result in the induction of immunosuppressive Tregs and facilitates T cell suppression. TAMs are also present [41]. According to an analysis of HCC data in The Cancer Genome Atlas (TCGA), high TAM infiltration influences patient survival and correlates with poor outcomes in HCC [42]. To investigate the underlying mechanisms of TAM-mediated pro-HCC effects, a recent study using human HCC samples identified the TAM phenotype CD14+HLA-DR+PD-L1+Lag-3+ [43]. Importantly, further research shows that T cell immunoglobulin mucin-3 (Tim-3) on macrophages promotes the differentiation of primitive macrophages into M2 macrophages, thereby upregulating the secretion of anti-inflammatory cytokines and enhancing IL-6-induced tumor growth in patients with HCC. Underlying this, transforming growth factor-β (TGF-β) promotes the transcription of Tim-3 in TAMs [44]. Furthermore, Jindao et al. recently found that M2 macrophage-derived exosomes promoted HCC cell migratory activity by transferring functional CD11b/CD18 proteins from TAMs to HCC cells [45]. Interestingly, researchers also found that miR-92a-2-5p in macrophage-derived exosomes could decrease androgen receptor expression, which results in HCC cell invasion [46].
It is well accepted that there is a high density of immune cells in the liver, regardless of whether they serve as immunosurveillance effectors or exert immunosuppressive effects; they are all components of the TIME in HCC and act as immunological modulators by responding to various cell-surface ligands, producing a wide array of pro- and anti-inflammatory molecules, or directly killing cancer cells to affect the activity of tumor cells. Therefore, therapies that reactivate surveillance-capable immune cells or keep immunosuppressive cells of the innate and adaptive immune systems dormant in the TIME may produce promising effects to treating patients with HCC.
Immune checkpoint inhibition (ICI) for HCC treatment
It is well known that the tumor expression of immunoregulatory molecules may reflect the immune milieu, which is regulated by alterations in oncogenic cellular signals. Then, different expression levels of molecular subclasses and immune phenotypes lead to specific tumor immune microenvironments. Additionally, immune checkpoints are a group of signaling pathway molecules expressed by immune cells that regulate and control immune responses while maintaining self-tolerance, preventing the immune system from performing unregulated attacks on cells and preventing autoimmunity. Since the efficacy of immune responses is determined by the delicate balance between costimulatory and coinhibitory signals, immune checkpoints can be either stimulatory or inhibitory. For instance, stimulatory immune checkpoint molecules include CD27, CD28, CD40, CD122, and CD137. Inhibitory immune checkpoint molecules include A2AR, B7-H3, B7-H4, cytotoxic T-lymphocyte antigen 4 (CTLA-4), indoleamine 2,3-dioxygenase (IDO), lymphocyte activation gene 3 (LAG-3), PD-1, and Tim-3 [47–50]. However, immune checkpoints are usually “hijacked” by tumor cells to restrain the immune system’s ability to mount an effective antitumor response to protect the tumor from attack by the immune system. The expression of immune-checkpoint proteins, including CTLA-4, PD-1, PD-L1, and IDO-1, results in a protumor cytokine milieu and further moderates the antitumor response [51, 52].
Interestingly, Chew et al. found that compared with Tregs and T cells isolated from a non-TME location, Tregs and T cells within the TME expressed multiple markers responsible for T-cell exhaustion, including PD-1, Lag-3, and Tim-3 [43]. In addition, Brown et al. indicated that the high expression of IDO in patients with HCC is associated with tumor cell the resistance ICBs [53]. Therefore, the emerging goal of immunotherapy is to curb T cell depletion by tumor cells and increase the rate of active immune cell infiltration, thereby allowing detection and elimination of tumor cells and preventing further tumor development.
Targets for ICBs in the HCC tumor microenvironment
Through targeting of receptor or ligand interactions by molecular blockade, ICB / ICI has become the first generation of antibody-based immunological treatments that target the immunoregulatory dampening of T cell activation or the function of host responses to tumor-associated antigens. The ability to induce tumor regression in murine models by blocking the prototypical receptor CTLA-4 was discovered over 20 years ago [54]. Since anti-CTLA-4 (ipilimumab) was approved for melanoma treatment, which promotes T-cell activation by blocking the interaction of CTLA4 with CD80 and CD86 [55], the clinical successes achieved by blocking CTLA-4 and PD-1 have inspired further exploration of the potential of ICB therapies. In addition to the anti-CTLA-4 antibody ipilimumab in melanoma, clinical studies of the anti-PD-1 antibody nivolumab in advanced HCC patients have also proven this antibody to be efficient in phase I/II clinical trials with satisfactory safety and response data [56–58]. However, there is still a large population of patients who show an ineffective response after application of ICB therapy.
Retrospective analyses of HCC patients treated with ICB have indicated that various TIME subtypes associated with tumor progression exhibit different ICB responsiveness patterns [12]. In HCC tissue, as components of the TIME, CD8+ and CD4+ T cells expresses a higher surface levels of PD-1, Tim-3, and CTLA-4 than those in the peripheral blood or nontumor tissues that express the ligands for these receptors [59]. After further evaluation, researchers determined that the expression of PD-1, Tim-3, and LAG-3 is higher on the cell surface of tumor-associated antigen (TAA)-specific CD8+ TILs than those on that of other CD8+ TILs. Once antibodies block PD-L1, Tim-3, or LAG-3, T cell responses to tumor antigens in HCC could be restored, and combination treatment effects would also be enhanced. Although ICB studies of anti-PD-1/PD-L1 and anti-CTLA-4 antibodies have significant advantages in multiple tumor treatments, there has been an increasing incidence of resistance to these antibodies in recent years. In addition, accumulating co-expression of PD-1 and Tim-3 on CD8+ T cells has been observed in the TME [60, 61], and studies have also revealed that blockade of PD-1 simultaneously increases the expression of other checkpoints molecules on immune cells in the TME, including Tim-3 and LAG-3 [62, 63]. Therefore, anti-Tim-3 antibodies have become a new focus of curative HCC research and have shown antitumor efficacy in some preclinical studies. In the TME of HCC, Tim-3 is expressed at high levels on tumor cells, tumor-infiltrating T cells, Tregs, and TAMs, inducing TAM-immunosuppression and promoting HCC progression by dampening T cell function. Consistently, Li et al. reported that the relatively high infiltration of Tim-3+ T cells in HCC tissues leads to shortened patient survival [61].
In addition to the prognostic characteristics of Tim-3 in HCC, several anti-Tim-3 antibodies have been granted research patents. In monotherapy or combination therapy, the anti-Tim3 antibody RMT3–23 exhibits significant antitumor efficacy in several types of cancer, such as MCA-induced sarcoma [64], colon adenocarcinoma [64], lung cancer [60], ID8 ovarian cancer [65], melanoma [63], and murine glioma [66]. However, triggering antitumor immunity in tumors requires the co-existence of CD4+ T cells and IFN-γ-expressing CD8+ T cells when using anti-Tim-3 antibodies as a monotherapy [64]. Moreover, for the treatment of some cancer types, the efficacy of a monotherapy is significantly inferior to that of multi-immune checkpoint-targeted combination therapy. Co-blockade of Tim-3, PD-1, and CTLA-4 or combination with other therapeutic strategies, such as DNA methyltransferase inhibition or stereotactic radiosurgery, has shown synergistic antitumor effects in preclinic studies [67]. Although researchers have revealed that directly blocking the expression of Tim-3 and PD-1 can restore the antitumor function of TILs in HBV-HCC [68], there are almost no relevant experimental data showing that targeted blockade of Tim-3 can inhibit the growth of HCC and improve the survival rate of patients. Based on the antitumor effects of Tim-3 in other cancer types, Tim-3 could be a promising target for novel immunotherapeutic approaches in HCC, as shown in Fig. 1(b).
Potential regulatory strategy for ICI in HCC
Antibody application in phase III clinical trials assessing PD-1, and CTLA-4 ICB monotherapies has shown inspiring outcomes, but the production of monoclonal antibodies is difficult and costly; therefore, other forms of inhibitors or mechanistic regulation are alternative strategies.
MiRNAs, as dual regulators, are widely used in the study of tumorigenic mechanisms and therapeutic tools. Many reports have revealed that tumor-suppressing miRNAs are responsible for the antitumor immune response by directly regulating the transcription of immune checkpoint molecules, such as PD-1, PD-L1, and CTLA-4; targeting either the PD-1 or PD-L1 checkpoint protein; or regulating both transcription and related proteins simultaneously within the TME [69]. For PD-L1 checkpoint molecule expression, since the mechanism of PD-L1 overexpression that results in cancer immune evasion is the disruption of the PD-L1 3′-UTR in several types of cancer, multiple miRNAs bind to the 3′-UTR of PD-L1, resulting in translation inhibition at the posttranscriptional level. As an inhibitor of PD-L1 expression, miR-570 commonly exhibits downregulated expression during HCC oncogenesis, while miR-570 mimics can exert antiproliferative and antiangiogenic effects in vivo [70, 71]. In addition, in PD-L1 regulation, miRNAs can work either directly by targeting the 3′-UTR of PD-L1 as mentioned above or indirectly by regulating the upstream pathways of PD-L1, such as the PTEN/PI3K/Akt and JAK/STAT pathway [72–74]. Therefore, targeting the subset of immunomodulatory miRNAs, including miR-34, miR-126, miR-155, and the miR-25-93-106b cluster, and dysregulated miRNAs including miR-34, miR-570, miR-20b, miR-21b and miR-130b to regulate the expression of PD-L1 and control tumor immune evasion would be a promising approach to restore antitumor immunity and enhance the therapeutic response in HCC [71, 73, 75–77]. Since the liver contains the largest population of macrophages (also known as KCs) in the human body, TAMs, especially those with an M2 phenotype, acting as protumor macrophages within the TME of HCC have been “tamed” to facilitate tumor initiation, progression, and metastasis [78]. We summarize the recently proven regulatory approaches that can switch macrophage phenotypes from M2 to M1 in Table 1, including modulatory miRNA methods, immune checkpoint blockade, and other feasible approaches.
Table 1.
The verified regulating methods and targets that could repolarize TAMs to anti-HCC phenotypes
| Phenotypic Change | Regulating method | HCC model | Pathway | Effects |
|---|---|---|---|---|
| M2 to M1 | miR-99b transfection | Hepa1–6 cells injected mice |
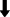 κB-Ras2 and/or mTOR; a positive feedback regulation loop of NF-κB κB-Ras2 and/or mTOR; a positive feedback regulation loop of NF-κB |
miR-99b amplifies M1 macrophage function, resulting in increased phagocytosis and antigen presentation, which impedes the growth of murine HCC [79] |
| M0 to M1; M2 to M1 | a nanoliposome-loaded C6-ceremide (LipC6) injection | C57BL/6 mice received injections of oncogenic hepatocytes |
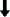 ROS signaling ROS signaling |
LipC6 enhances M1 cytokine production while inhibiting M2 cytokine production, thereby reversing immune suppression, and increasing CD8+ T cells activity [80] |
| M0 to M1; M2 to M1 | Listeria-based HCC vaccine, Lmdd-MPFG combined with PD-1 blockade | Hepa1–6/MPFG tumor-bearing mice |
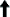 NF-κB pathway through the TLR2 and MyD88 pathway NF-κB pathway through the TLR2 and MyD88 pathway |
Lmdd-MPFG induces an increase in T cells number in the HCC TME and promotes the production of cytokines, such as IFN-γ [81]. |
| M1-type activated; M2-type decreased | dual anti-PD-1/VEGFR-2 therapy | HCA-1 in C3H mice and RIL-175 in C57Bl/6 mice. |
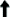 selectively upregulated pathways associated with myeloid cells and B cells selectively upregulated pathways associated with myeloid cells and B cells |
Combination PD-1and VEGFR-2 blockade therapy shifts the TAM ratios of F4/80+ CD80+ and/or CD86+ M1 TAMs to F4/80+CD206+ M2 TAMs, which also regulates the infiltration of other immune cells and reprograms the TME to an antitumor state in HCC [82]. |
| M0 to M1 | upregulate RIG-I expression | H22 liver cancer cells inoculated C57BL/6 mice |
 RIG-I/ MAVS/TRAF2/NF-κB pathway RIG-I/ MAVS/TRAF2/NF-κB pathway |
RIGI-induced M1 macrophages promoted apoptosis and death in HCC cells in vivo and in vitro [83]. |
| M0 to M1 | upregulated SIRT1 | HepG2 and RAW 264.7, HL-60 macrophages |
 NF-κB pathway NF-κB pathway |
SIRT1 overexpression enhances M1-like macrophage infiltration in HCC while inhibiting HCC cell growth, migration, and invasion [84]. |
| M2 to M1 | upregulate IL-37 expression | HepG2 and Huh-7 cells injected BALB/c nude mice and HCC-conditioned TAMs |
 IL-6/STAT3 signaling. IL-6/STAT3 signaling. |
IL-37 inhibits M2 polarization of TAMs and then suppresses HCC cell activities, including growth, migration, and invasion. In vivo, upregulated IL-37 expression in HCC-conditioned TAMs delays tumor growth [85]. |
| M2 to M1 | miR-98 mimics | HepG2 and SMMC7721 cells incubated with the culture medium of TAMs |
 targeting IL-10 targeting IL-10 |
miR-98 reverses M2 polarization in HCC and inhibits the TAM-mediated promotion of invasion, migration, and epithelial-mesenchymal transition in HCC [86, 87]. |
In addition, the novel inhibition approaches for the aforementioned Tim-3 checkpoint may provide new ideas for the regulation of other immune checkpoints. In the tumor immune microenvironment, cytokines are connected with immune cell activation and involved in the induction of Tim-3 expression in immune cells. It has been reported that the transcriptional induction of Tim-3 in T cells can be activated by IL-2, IL-7, IL-15, IL-21, and IL-27 [88, 89]. Increased serum IL-6 levels enhance PD- L1 expression in HCC monocytes and macrophages through JAK2/STAT1 and JAK2/STAT3/c- MYC signaling activation [90]. Specifically, Xie et al. analyzed immunohistochemistry and RNA-sequencing data for patients with HCC and demonstrated that high expression of OX40 regulated the activation of T cells, expansion of suppressive Tregs and upregulation of Tim-3 expression as well as CD8A, CD68, LAG3, and PD-1 expression [91], which provided a rationale for reversing multiple immunosuppressive pathways via therapeutic targeting of OX40 and blocking associated immune checkpoints in HCC patients. In addition to checkpoint molecule expression regulation, a new form of blockade now under preclinical scrutiny is also a therapeutic option. Similar to the recognition function of monoclonal antibodies, aptamers, single-stranded or peptide molecules, can bind to specific target molecules and display activity superior to that of currently used RMT3–23 monoclonal antibodies both in vivo and in vitro. Researchers have identified that a trimeric ligand of Tim-3 can efficiently eliminate the interaction of Tim-3 with Gal-9, thereby enhancing the proliferation of Tim-3+T cells and promoting their antitumor cytokine secretion with high affinity and specificity [92]. For Tim-3-expressing lymphocytes, Hervas-Stubbs et al. isolated a Tim-3 nonantigenic oligonucleotide aptamer (Tim-3 Apt) that binds to the extracellular domain of Tim-3 and increase the secretion of IFN-γ [93]. Moreover, MP7, an aptamer that blocks the immunoinhibitory PD-1, functionally restores IL-2 secretion in primary T cells by inhibiting the interaction between PD-1 and PD-L1 [94]. Since aptamers have potency equivalent to that of an antagonistic anti-checkpoint molecule antibody that provokes robust and durable antioncogenetic responses and inherent advantages, including a lack of immunogenicity, low cost, long shelf life, and antidote availability, the development of aptamers as novel ICBs for patients with HCC has bright research prospects.
Discussion
Tumor growth and metastasis are related not only to the intrinsic characteristics of tumor cells but also to the organ microenvironment where tumorigenesis is occurring, which is the dominant force promoting tumor growth [95]. The TME is recognized as a highly dynamic network during the occurrence, development, and prognosis of cancer or therapeutic interventions and one of the leading causes of tumor heterogeneity. The TME is composed of immune cells and stromal cells, the two major nontumor components that largely determine the effect and prognosis of cancer treatment [96]. Currently, to transform the TME into an immunologically “hot” status, novel methods that combine metabolic regulation and microbiome-based gene signatures that facilitate personalized immunotherapies are urgently needed to enhance antitumor immunity. Riera-Domingo et al. [97] emphasized hypoxic signatures that influence the immune response to immunotherapy. The hypoxic microenvironment has a great impact on the antitumor immune response. As a critical process that evolves in the TME, hypoxia affects epithelial-mesenchymal transition and angiogenesis and mainly changes the expression of immune checkpoint molecules, such as PD-L1, CD47, PD-1, and HLA-G, in liver cancer [98, 99]. With the help of HIF-1α, which binds the HRE in the promoter of the PD-L1 gene, the expression of PD-L1 is significantly increased in many types of tumor cells, including melanoma, lung cancer, breast cancer, and prostate cancer cells [100, 101]. In the hypoxic microenvironment of a steatotic-HCC mouse model, Jianxu et al. reported that upregulated HIF-2α expression led to lipid accumulation through PI3K-AKT-mTOR pathway activation, which reflected that the hypoxic conditions with HIF-2α upregulation could serve as a potential therapeutic target for liver cancer treatment [102]. Interestingly, there are always highly hypoxic conditions within the TME that promote immunosuppression and negatively affect antitumor immunity, yet one recent study found that CD8+ T cells activated under hypoxic condition exhibited increased cytotoxicity and enhanced antitumor immunity in vivo [103]. Therefore, approaches for regulating microenvironmental hypoxia in HCC should be given more attention in future studies, including direct modulation of HIF-mRNA, induction of HIF-1α degradation, hypoxia-activated drugs, small-molecule HIF inhibitors, and drugs targeting signaling pathways downstream of HIF [104, 105].
In addition to regulation of the microenvironment at the metabolic level, targeting the regulation of immune cells seems to be relatively feasible. As mentioned above, studies have consistently verified the potential of TAMs as a target for HCC immunotherapy. Therefore, the development of better tools, such as high-throughput single-cell approaches, could allow in-depth analysis of the phenotypic characteristics and functional features of diverse cell types within the TME and reveal their crosstalk at different stages of cancer progression and metastasis [10], thereby guiding clinical therapeutics for tumors and prognostic monitoring. Since the two major classes of tumor-associated macrophages have distinct opposing roles in the development of HCC, inducing the transition from protumor M2-like macrophage phenotype to a tumoricidal M1 phenotype is the main strategy to increase the antitumor effect of TAMs. Therefore, we also summarized the emerging potential regulatory methods that could achieve this transformation in Table 2. Furthermore, interfering with M2- like TAM survival and blocking M2 macrophage recruitment [113] are also two targeted regulatory mechanisms that cannot be ignored.
Table 2.
The potential targets and pathways that regulate HCC-TAMs phenotypic transition
| Target | Pathway | Cell line or HCC model | Pro-tumor Results |
|---|---|---|---|
| RIPK3 and FAO | ROS-caspase1-PPAR pathway | Murine H22 cells and C57BL/6 WT mice injected with diethylnitrosamine as HCC model | The RIPK3-FAO-ROS-caspase1-PPAR signaling axis is responsible for increased M2-TAM infiltration, which promotes HCC tumorigenesis [106]. |
| LINC00662 | WNT3A-Wnt/b-catenin signaling | HCCLM3, MHCC97H, Huh7, SK-HEP-1, and Hepa1–6 HCC cells | By upregulating WNT3A expression, LINC00662 activates the Wnt/b-catenin pathway and then induces M2 macrophage polarization, contributing to HCC tumorigenesis and invasion and repressing HCC cell apoptosis [107, 108]. |
| IL-25 | / | Human HCC cell lines MHCC97L and HepG2 cells; Murine HCC cell lines H22, and Hepa1–6; and BALB/c nude mice with the portal venous injection macrophages as HCC model. | IL-25 facilitates M2 TAM (CD206/CD68) infiltration, promotes secretion of the chemokine CXCL-10, and then induces HCC progression through the EMT pathway [109]. |
| CAF-induced endosialin | Interaction with CD68 and regulate GAS6 expression in CAF | Huh7 cell HFL-1 cells | In CAFs, via an interaction with CD68, endosialin promotes the expression of GSA6, which results in increased M2 macrophages recruitment and HCC progression [110]. |
| HCC-derived HMGB1 | ROS-TLR2-NOX2-autophagy axis | Mouse hepatoma cell line ML-14a cells and murine in a situ hepatoma model | Hepatoma-derived HMGB1 stimulates ROS via the TLR2/NOX2 axis, thereby inducing M2 macrophage polarization and subsequently supporting HCC growth [111]. |
| Nogo-B | Nogo-B-Yap/Taz pathway | Murine HCC cell lines Hepa1–6 cells and H22 cells, macrophage cell line RAW 264.7 cells | Elevated Nogo-B expression facilitates M2 TAM polarization and promotes HCC tumor growth in vivo by inducing Yap/Taz signaling [112]. |
Conclusion
In conclusion, as the liver is the body’s largest immune organ, liver cancer occurrence and development are already complicated, which results in the TME of liver cancer constantly changing. Nevertheless, modulation of the highly immunosuppressive and metabolically stressed TME is urgently needed to improve efficacy and identify protagonist cell types, such as TAMs, with high therapeutic relevance in the TME of both primary HCC and metastatic HCC. Furthermore, immune checkpoint inhibition has been studied in detail to explore the relevant mechanisms of PD-1/PD-L1 and CTLA-4, but specific useful ICB targets have not produced reflected a relatively ideal effect in HCC treatment. Additionally, recent works in the literature have recognized the role of Tim-3 as a potential target in HCC immunotherapy, which might both enhance T cell-induced antitumor immunity and revise macrophage-mediated immunosuppression in the HCC microenvironment, suggesting that future immune checkpoint investigations may focus more on Tim-3 for treatment of HCC or other solid tumors with similar pathogenesis. In addition to novel immune checkpoint discovery, emerging regulatory strategies have inspired HCC immunotherapy ideas. Immunotherapeutic aptamers, which bind specifically to the extracellular domain of immune checkpoints and block their interactions with receptors/ligands, significantly suppress growing tumor cells in vivo and represent an attractive alternative to monoclonal antibody therapeutics. Importantly, these antagonistic aptamer treatments have important advantages, including lower antigenicity, lower manufacturing price, and higher malleability. Additionally, at the molecular level of regulation, recent discoveries have shown that some miRNAs and hypoxia can negatively affect the antitumor immune response by modifying the expression of the main immune checkpoint molecules, setting up new therapeutic intervention opportunities to enhance the clinical benefit derived from ICBs. In the future, much attention should be paid to altering systemic metabolism or upstream modulatory sites in multiple facets of antitumor immunity within the HCC microenvironment, thereby significantly increasing the sensitivity of immune surveillance and enhancing treatment responses.
Acknowledgements
Not applicable.
Abbreviations
- HCC
Hepatocellular carcinoma
- HBV
Hepatitis B virus
- HCV
Hepatitis C virus
- TME
Tumor microenvironment
- ECM
Extracellular matrix
- TIME
Tumor immune microenvironment
- MDSCs
Myeloid-derived suppressor cells
- Treg
Regulatory T
- TAMs
Tumor-associated macrophages
- NKs
Natural killer cells
- I-E
Infiltrated-excluded
- I-I
Infiltrated-inflamed
- TLS
Tertiary lymphoid structure
- CTLs
Cytotoxic lymphocytes
- PD-1
Programmed cell death 1
- PD-L1
PD-1 ligand
- RFA
Radiofrequency ablation
- TACE
Trans arterial chemoembolization
- TARE
Trans arterial radioembolization
- VEGF
Vascular endothelial growth factor
- TKI
Tyrosine kinase inhibitor
- PDGFR
Platelet-derived growth factor
- FGFR
Fibroblast growth factor
- ICB
Immune checkpoint blockade
- CSF
Colony-stimulating factor
- iCCA
Intrahepatic cholangiocarcinoma
- IFN
Interferon
- LGDNs
Low-grade dysplastic nodules
- HGDNs
High-grade dysplastic nodules
- TIL
Tumor-infiltrating lymphocyte
- APCs
Antigen-presenting cells
- KCs
Kupffer cells
- TLRs
Toll-like receptors
- PGE2
Prostaglandin E2
- TCGA
The Cancer Genome Atlas
- Tim-3
Immunoglobulin mucin-3
- TGF-β
Transforming growth factor-β
- ICI
Immune checkpoint inhibition
- CTLA-4
Cytotoxic T-lymphocyte antigen 4
- IDO
Indoleamine 2,3-dioxygenase
- LAG-3
Lymphocyte activation gene 3
- TAA
Tumor-associated antigen
- Tim-3 Apt
Tim-3 nonantigenic oligonucleotide aptamer
Authors’ contributions
JZ performed the literature research, drafted the manuscript, and made the figure; WW revised the manuscript and directed the review to be more focused; QL gave the final approval for the article to be published. All authors read and approved the final manuscript.
Funding
The present study was supported by grants from the National Natural Science Foundation of China (grant NO. 82073214) and Outstanding disciplines leaders of Shanghai Municipal Commission of Health and Family Planning (No. 2018BR39) and Bethune Charitable Foundation.
Availability of data and materials
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Pinter M, Scheiner B, Peck-Radosavljevic M. Immunotherapy for advanced hepatocellular carcinoma: a focus on special subgroups. Gut. 2020. [DOI] [PMC free article] [PubMed]
- 2.Yang J, Hainaut P, Gores G, Amadou A, Plymoth A, Roberts L. A global view of hepatocellular carcinoma: trends, risk, prevention and management. Nat Rev Gastroenterol Hepatol. 2019;16(10):589–604. doi: 10.1038/s41575-019-0186-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Siegel R, Miller K, Jemal A. Cancer statistics, 2020. CA Cancer J Clin. 2020;70(1):7–30. doi: 10.3322/caac.21590. [DOI] [PubMed] [Google Scholar]
- 4.Lan A, Hongmei Z, Zheng R, Xi Z, Kexin S, Xiaonong Z, et al. Analysis of the prevalence of liver cancer in China in 2015. Chin J Oncol. 2019;41(10):721–727. [Google Scholar]
- 5.Forner A, Llovet J, Bruix J. Hepatocellular carcinoma. Lancet (London, England) 2012;379(9822):1245–1255. doi: 10.1016/S0140-6736(11)61347-0. [DOI] [PubMed] [Google Scholar]
- 6.Zhou W, Zhang Q, Qiao L. Pathogenesis of liver cirrhosis. World J Gastroenterol. 2014;20(23):7312–7324. doi: 10.3748/wjg.v20.i23.7312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Rao H, Wu E, Fu S, Yang M, Feng B, Lin A, et al. The higher prevalence of truncal obesity and diabetes in American than Chinese patients with chronic hepatitis C might contribute to more rapid progression to advanced liver disease. Aliment Pharmacol Ther. 2017;46(8):731–740. doi: 10.1111/apt.14273. [DOI] [PubMed] [Google Scholar]
- 8.Schulze K, Imbeaud S, Letouzé E, Alexandrov L, Calderaro J, Rebouissou S, et al. Exome sequencing of hepatocellular carcinomas identifies new mutational signatures and potential therapeutic targets. Nat Genet. 2015;47(5):505–511. doi: 10.1038/ng.3252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Balkwill F, Mantovani A. Inflammation and cancer: back to Virchow? Lancet (London, England) 2001;357(9255):539–545. doi: 10.1016/S0140-6736(00)04046-0. [DOI] [PubMed] [Google Scholar]
- 10.Duan Q, Zhang H, Zheng J, Zhang L. Turning cold into hot: firing up the tumor microenvironment. Trends Cancer. 2020;6(7):605–618. doi: 10.1016/j.trecan.2020.02.022. [DOI] [PubMed] [Google Scholar]
- 11.Kurebayashi Y, Ojima H, Tsujikawa H, Kubota N, Maehara J, Abe Y, et al. Landscape of immune microenvironment in hepatocellular carcinoma and its additional impact on histological and molecular classification. Hepatology (Baltimore, Md) 2018;68(3):1025–1041. doi: 10.1002/hep.29904. [DOI] [PubMed] [Google Scholar]
- 12.Binnewies M, Roberts E, Kersten K, Chan V, Fearon D, Merad M, et al. Understanding the tumor immune microenvironment (TIME) for effective therapy. Nat Med. 2018;24(5):541–550. doi: 10.1038/s41591-018-0014-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Couri T, Pillai A. Goals and targets for personalized therapy for HCC. Hepatol Int. 2019;13(2):125–137. doi: 10.1007/s12072-018-9919-1. [DOI] [PubMed] [Google Scholar]
- 14.Lee S, Kang T, Cha D, Song K, Lee M, Rhim H, et al. Radiofrequency ablation vs. surgery for perivascular hepatocellular carcinoma: propensity score analyses of long-term outcomes. J Hepatol. 2018;69(1):70–78. doi: 10.1016/j.jhep.2018.02.026. [DOI] [PubMed] [Google Scholar]
- 15.Cheng A, Kang Y, Chen Z, Tsao C, Qin S, Kim J, et al. Efficacy and safety of sorafenib in patients in the Asia-Pacific region with advanced hepatocellular carcinoma: a phase III randomised, double-blind, placebo-controlled trial. Lancet Oncol. 2009;10(1):25–34. doi: 10.1016/S1470-2045(08)70285-7. [DOI] [PubMed] [Google Scholar]
- 16.Llovet JM, Ricci S, Mazzaferro V, Hilgard P, Gane E, Blanc JF, et al. Sorafenib in advanced hepatocellular carcinoma. N Engl J Med. 2008;359(4):378–390. doi: 10.1056/NEJMoa0708857. [DOI] [PubMed] [Google Scholar]
- 17.Finn R, Qin S, Ikeda M, Galle P, Ducreux M, Kim T, et al. Atezolizumab plus Bevacizumab in Unresectable Hepatocellular Carcinoma. N Engl J Med. 2020;382(20):1894–1905. doi: 10.1056/NEJMoa1915745. [DOI] [PubMed] [Google Scholar]
- 18.Kelley R. Atezolizumab plus Bevacizumab - A Landmark in Liver Cancer. N Engl J Med. 2020;382(20):1953–1955. doi: 10.1056/NEJMe2004851. [DOI] [PubMed] [Google Scholar]
- 19.Han S, Zhen W, Guo T, Zou J, Li F. SETDB1 promotes glioblastoma growth via CSF-1-dependent macrophage recruitment by activating the AKT/mTOR signaling pathway. J Exp Clin Cancer Res. 2020;39(1):218. doi: 10.1186/s13046-020-01730-8. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 20.Calderaro J, Ziol M, Paradis V, Zucman-Rossi J. Molecular and histological correlations in liver cancer. J Hepatol. 2019;71(3):616–630. doi: 10.1016/j.jhep.2019.06.001. [DOI] [PubMed] [Google Scholar]
- 21.Zheng C, Zheng L, Yoo J, Guo H, Zhang Y, Guo X, et al. Landscape of Infiltrating T Cells in Liver Cancer Revealed by Single-Cell Sequencing. Cell. 2017;169(7):1342–1356.e16. [DOI] [PubMed]
- 22.Calderaro J, Couchy G, Imbeaud S, Amaddeo G, Letouzé E, Blanc J, et al. Histological subtypes of hepatocellular carcinoma are related to gene mutations and molecular tumour classification. J Hepatol. 2017;67(4):727–738. doi: 10.1016/j.jhep.2017.05.014. [DOI] [PubMed] [Google Scholar]
- 23.Sia D, Jiao Y, Martinez-Quetglas I, Kuchuk O, Villacorta-Martin C. Castro de Moura M, et al. identification of an immune-specific class of hepatocellular carcinoma, based on molecular features. Gastroenterology. 2017;153(3):812–826. doi: 10.1053/j.gastro.2017.06.007. [DOI] [PubMed] [Google Scholar]
- 24.Salomao M, Yu W, Brown R, Emond J, Lefkowitch J. Steatohepatitic hepatocellular carcinoma (SH-HCC): a distinctive histological variant of HCC in hepatitis C virus-related cirrhosis with associated NAFLD/NASH. Am J Surg Pathol. 2010;34(11):1630–1636. doi: 10.1097/PAS.0b013e3181f31caa. [DOI] [PubMed] [Google Scholar]
- 25.Chan A, Tong J, Pan Y, Chan S, Wong G, Wong V, et al. Lymphoepithelioma-like hepatocellular carcinoma: an uncommon variant of hepatocellular carcinoma with favorable outcome. Am J Surg Pathol. 2015;39(3):304–312. doi: 10.1097/PAS.0000000000000376. [DOI] [PubMed] [Google Scholar]
- 26.Nakagawa S, Umezaki N, Yamao T, Kaida T, Okabe H, Mima K, et al. Survival impact of lymphocyte infiltration into the tumor of hepatocellular carcinoma in hepatitis B virus-positive or non-B non-C patients who underwent curative resection. Hepatol Res. 2018;48(3):E126–EE32. doi: 10.1111/hepr.12936. [DOI] [PubMed] [Google Scholar]
- 27.Calderaro J, Petitprez F, Becht E, Laurent A, Hirsch T, Rousseau B, et al. Intra-tumoral tertiary lymphoid structures are associated with a low risk of early recurrence of hepatocellular carcinoma. J Hepatol. 2019;70(1):58–65. doi: 10.1016/j.jhep.2018.09.003. [DOI] [PubMed] [Google Scholar]
- 28.Llovet J, Montal R, Sia D, Finn R. Molecular therapies and precision medicine for hepatocellular carcinoma. Nat Rev Clin Oncol. 2018;15(10):599–616. doi: 10.1038/s41571-018-0073-4. [DOI] [PubMed] [Google Scholar]
- 29.Shimada S, Mogushi K, Akiyama Y, Furuyama T, Watanabe S, Ogura T, et al. Comprehensive molecular and immunological characterization of hepatocellular carcinoma. EBioMedicine. 2019;40:457–470. doi: 10.1016/j.ebiom.2018.12.058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Tsujikawa H, Masugi Y, Yamazaki K, Itano O, Kitagawa Y, Sakamoto M. Immunohistochemical molecular analysis indicates hepatocellular carcinoma subgroups that reflect tumor aggressiveness. Hum Pathol. 2016;50:24–33. doi: 10.1016/j.humpath.2015.10.014. [DOI] [PubMed] [Google Scholar]
- 31.Uenishi T, Kubo S, Yamamoto T, Shuto T, Ogawa M, Tanaka H, et al. Cytokeratin 19 expression in hepatocellular carcinoma predicts early postoperative recurrence. Cancer Sci. 2003;94(10):851–857. doi: 10.1111/j.1349-7006.2003.tb01366.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sakamoto M, Effendi K, Masugi Y. Molecular diagnosis of multistage hepatocarcinogenesis. Jpn J Clin Oncol. 2010;40(9):891–896. doi: 10.1093/jjco/hyq099. [DOI] [PubMed] [Google Scholar]
- 33.Meylan M, Petitprez F, Lacroix L, Di Tommaso L, Roncalli M, Bougoüin A, et al. Early hepatic lesions display immature tertiary lymphoid structures and show elevated expression of immune inhibitory and immunosuppressive molecules. Clin Cancer Res. 2020. [DOI] [PubMed]
- 34.Greten T, Duffy A, Korangy F. Hepatocellular carcinoma from an immunologic perspective. Clin Cancer Res. 2013;19(24):6678–6685. doi: 10.1158/1078-0432.CCR-13-1721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Robinson M, Harmon C, O'Farrelly C. Liver immunology and its role in inflammation and homeostasis. Cell Mol Immunol. 2016;13(3):267–276. doi: 10.1038/cmi.2016.3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Flecken T, Schmidt N, Hild S, Gostick E, Drognitz O, Zeiser R, et al. Immunodominance and functional alterations of tumor-associated antigen-specific CD8+ T-cell responses in hepatocellular carcinoma. Hepatology (Baltimore, Md) 2014;59(4):1415–1426. doi: 10.1002/hep.26731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Guidotti L, Inverso D, Sironi L, Di Lucia P, Fioravanti J, Ganzer L, et al. Immunosurveillance of the liver by intravascular effector CD8(+) T cells. Cell. 2015;161(3):486–500. doi: 10.1016/j.cell.2015.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Garnelo M, Tan A, Her Z, Yeong J, Lim C, Chen J, et al. Interaction between tumour-infiltrating B cells and T cells controls the progression of hepatocellular carcinoma. Gut. 2017;66(2):342–351. doi: 10.1136/gutjnl-2015-310814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ma C, Kesarwala A, Eggert T, Medina-Echeverz J, Kleiner D, Jin P, et al. NAFLD causes selective CD4(+) T lymphocyte loss and promotes hepatocarcinogenesis. Nature. 2016;531(7593):253–257. doi: 10.1038/nature16969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Spranger S, Dai D, Horton B, Gajewski T. Tumor-Residing Batf3 Dendritic Cells Are Required for Effector T Cell Trafficking and Adoptive T Cell Therapy. Cancer Cell. 2017;31(5):711–723.e4. [DOI] [PMC free article] [PubMed]
- 41.Doherty D. Immunity, tolerance and autoimmunity in the liver: a comprehensive review. J Autoimmun. 2016;66:60–75. doi: 10.1016/j.jaut.2015.08.020. [DOI] [PubMed] [Google Scholar]
- 42.Zhang Q, He Y, Luo N, Patel S, Han Y, Gao R, et al. Landscape and Dynamics of Single Immune Cells in Hepatocellular Carcinoma. Cell. 2019;179(4):829–845.e20. [DOI] [PubMed]
- 43.Chew V, Lai L, Pan L, Lim C, Li J, Ong R, et al. Delineation of an immunosuppressive gradient in hepatocellular carcinoma using high-dimensional proteomic and transcriptomic analyses. Proc Natl Acad Sci U S A. 2017;114(29):E5900–E59E9. doi: 10.1073/pnas.1706559114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Yan W, Liu X, Ma H, Zhang H, Song X, Gao L, et al. Tim-3 fosters HCC development by enhancing TGF-β-mediated alternative activation of macrophages. Gut. 2015;64(10):1593–1604. doi: 10.1136/gutjnl-2014-307671. [DOI] [PubMed] [Google Scholar]
- 45.J Wu, W Gao, Q Tang, Y Yu, W You, Z Wu, et al. M2 macrophage-derived exosomes facilitate hepatocarcinoma metastasis by transferring α β integrin to tumor cells. Hepatology (Baltimore, Md). 2020. [DOI] [PMC free article] [PubMed] [Retracted]
- 46.G Liu, X Ouyang, Y Sun, Y Xiao, B You, Y Gao, et al. The miR-92a-2-5p in exosomes from macrophages increases liver cancer cells invasion via altering the AR/PHLPP/p-AKT/β-catenin signaling. Cell death and differentiation. 2020. [DOI] [PMC free article] [PubMed]
- 47.Topalian S. Targeting immune checkpoints in Cancer therapy. JAMA. 2017;318(17):1647–1648. doi: 10.1001/jama.2017.14155. [DOI] [PubMed] [Google Scholar]
- 48.He X, Xu C. Immune checkpoint signaling and cancer immunotherapy. Cell Res. 2020;30(8):660–669. doi: 10.1038/s41422-020-0343-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Fallah-Mehrjardi K, Mirzaei H, Masoumi E, Jafarzadeh L, Rostamian H, Khakpoor-Koosheh M, et al. Pharmacological targeting of immune checkpoint A2aR improves function of anti-CD19 CAR T cells in vitro. Immunol Lett. 2020;223:44–52. doi: 10.1016/j.imlet.2020.04.005. [DOI] [PubMed] [Google Scholar]
- 50.Liu F, Liu Y, Chen Z. Tim-3 expression and its role in hepatocellular carcinoma. J Hematol Oncol. 2018;11(1):126. doi: 10.1186/s13045-018-0667-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Angelova M, Charoentong P, Hackl H, Fischer M, Snajder R, Krogsdam A, et al. Characterization of the immunophenotypes and antigenomes of colorectal cancers reveals distinct tumor escape mechanisms and novel targets for immunotherapy. Genome Biol. 2015;16:64. doi: 10.1186/s13059-015-0620-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Gatalica Z, Snyder C, Maney T, Ghazalpour A, Holterman D, Xiao N, et al. Programmed cell death 1 (PD-1) and its ligand (PD-L1) in common cancers and their correlation with molecular cancer type. Cancer #pidemiol, Biomark Prevent. 2014;23(12):2965–2970. doi: 10.1158/1055-9965.EPI-14-0654. [DOI] [PubMed] [Google Scholar]
- 53.Brown Z, Yu S, Heinrich B, Ma C, Fu Q, Sandhu M, et al. Indoleamine 2,3-dioxygenase provides adaptive resistance to immune checkpoint inhibitors in hepatocellular carcinoma. Cancer Immunol Immunother CII. 2018;67(8):1305–1315. doi: 10.1007/s00262-018-2190-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Leach D, Krummel M, Allison J. Enhancement of antitumor immunity by CTLA-4 blockade. Science (New York, NY). 1996;271(5256):1734–6. [DOI] [PubMed]
- 55.Melero I, Hervas-Stubbs S, Glennie M, Pardoll D, Chen L. Immunostimulatory monoclonal antibodies for cancer therapy. Nat Rev Cancer. 2007;7(2):95–106. doi: 10.1038/nrc2051. [DOI] [PubMed] [Google Scholar]
- 56.El-Khoueiry A, Sangro B, Yau T, Crocenzi T, Kudo M, Hsu C, et al. Nivolumab in patients with advanced hepatocellular carcinoma (CheckMate 040): an open-label, non-comparative, phase 1/2 dose escalation and expansion trial. Lancet (London, England, 2017;389(10088):2492–502. [DOI] [PMC free article] [PubMed]
- 57.Finkelmeier F, Waidmann O, Trojan J. Nivolumab for the treatment of hepatocellular carcinoma. Expert Rev Anticancer Ther. 2018;18(12):1169–1175. doi: 10.1080/14737140.2018.1535315. [DOI] [PubMed] [Google Scholar]
- 58.Finkelmeier F, Czauderna C, Perkhofer L, Ettrich T, Trojan J, Weinmann A, et al. Feasibility and safety of nivolumab in advanced hepatocellular carcinoma: real-life experience from three German centers. J Cancer Res Clin Oncol. 2019;145(1):253–259. doi: 10.1007/s00432-018-2780-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Zhou G, Sprengers D, Boor P, Doukas M, Schutz H, Mancham S, et al. Antibodies Against Immune Checkpoint Molecules Restore Functions of Tumor-Infiltrating T Cells in Hepatocellular Carcinomas. Gastroenterology. 2017;153(4):1107–1119.e10. [DOI] [PubMed]
- 60.Zhou Q, Munger M, Veenstra R, Weigel B, Hirashima M, Munn D, et al. Coexpression of Tim-3 and PD-1 identifies a CD8+ T-cell exhaustion phenotype in mice with disseminated acute myelogenous leukemia. Blood. 2011;117(17):4501–4510. doi: 10.1182/blood-2010-10-310425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Li J, Shayan G, Avery L, Jie H, Gildener-Leapman N, Schmitt N, et al. Tumor-infiltrating Tim-3 T cells proliferate avidly except when PD-1 is co-expressed: evidence for intracellular cross talk. Oncoimmunology. 2016;5(10):e1200778. doi: 10.1080/2162402X.2016.1200778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Sakuishi K, Apetoh L, Sullivan J, Blazar B, Kuchroo V, Anderson A. Targeting Tim-3 and PD-1 pathways to reverse T cell exhaustion and restore anti-tumor immunity. J Exp Med. 2010;207(10):2187–2194. doi: 10.1084/jem.20100643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Koyama S, Akbay E, Li Y, Herter-Sprie G, Buczkowski K, Richards W, et al. Adaptive resistance to therapeutic PD-1 blockade is associated with upregulation of alternative immune checkpoints. Nat Commun. 2016;7:10501. doi: 10.1038/ncomms10501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ngiow S, von Scheidt B, Akiba H, Yagita H, Teng M, Smyth M. Anti-TIM3 antibody promotes T cell IFN-γ-mediated antitumor immunity and suppresses established tumors. Cancer Res. 2011;71(10):3540–3551. doi: 10.1158/0008-5472.CAN-11-0096. [DOI] [PubMed] [Google Scholar]
- 65.Guo Z, Cheng D, Xia Z, Luan M, Wu L, Wang G, et al. Combined TIM-3 blockade and CD137 activation affords the long-term protection in a murine model of ovarian cancer. J Transl Med. 2013;11:215. doi: 10.1186/1479-5876-11-215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Kim J, Patel M, Mangraviti A, Kim E, Theodros D, Velarde E, et al. Combination therapy with anti-PD-1, anti-TIM-3, and focal radiation results in regression of murine Gliomas. Clin Cancer Res. 2017;23(1):124–136. doi: 10.1158/1078-0432.CCR-15-1535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Kang C, Dutta A, Chang L, Mahalingam J, Lin Y, Chiang J, et al. Apoptosis of tumor infiltrating effector TIM-3+CD8+ T cells in colon cancer. Sci Rep. 2015;5:15659. doi: 10.1038/srep15659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Liu F, Zeng G, Zhou S, He X, Sun N, Zhu X, et al. Blocking Tim-3 or/and PD-1 reverses dysfunction of tumor-infiltrating lymphocytes in HBV-related hepatocellular carcinoma. Bull Cancer. 2018;105(5):493–501. doi: 10.1016/j.bulcan.2018.01.018. [DOI] [PubMed] [Google Scholar]
- 69.Omar HA, E-Serafi AT, Hersi F, Arafa EA, Zaher DM, Madkour M, et al. Immunomodulatory MicroRNAs in cancer: targeting immune checkpoints and the tumor microenvironment. FEBS J. 2019;286(18):3540–3557. doi: 10.1111/febs.15000. [DOI] [PubMed] [Google Scholar]
- 70.Guo W, Tan W, Liu S, Huang X, Lin J, Liang R, et al. MiR-570 inhibited the cell proliferation and invasion through directly targeting B7-H1 in hepatocellular carcinoma. Tumour Biol. 2015;36(11):9049–9057. doi: 10.1007/s13277-015-3644-3. [DOI] [PubMed] [Google Scholar]
- 71.Lin Y, Liu S, Su L, Su Q, Lin J, Huang X, et al. miR-570 inhibits proliferation, angiogenesis, and immune escape of hepatocellular carcinoma. Cancer Biother Radiopharm. 2018;33(6):252–257. doi: 10.1089/cbr.2017.2389. [DOI] [PubMed] [Google Scholar]
- 72.Zhu J, Chen L, Zou L, Yang P, Wu R, Mao Y, et al. MiR-20b, −21, and -130b inhibit PTEN expression resulting in B7-H1 over-expression in advanced colorectal cancer. Hum Immunol. 2014;75(4):348–353. doi: 10.1016/j.humimm.2014.01.006. [DOI] [PubMed] [Google Scholar]
- 73.Xi J, Huang Q, Wang L, Ma X, Deng Q, Kumar M, et al. miR-21 depletion in macrophages promotes tumoricidal polarization and enhances PD-1 immunotherapy. Oncogene. 2018;37(23):3151–3165. doi: 10.1038/s41388-018-0178-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Li B, Lu Y, Yu L, Han X, Wang H, Mao J, et al. miR-221/222 promote cancer stem-like cell properties and tumor growth of breast cancer via targeting PTEN and sustained Akt/NF-κB/COX-2 activation. Chem-Biol Interact. 2017;277:33–42. doi: 10.1016/j.cbi.2017.08.014. [DOI] [PubMed] [Google Scholar]
- 75.Ma S, Liu M, Xu Z, Li Y, Guo H, Ge Y, et al. A double feedback loop mediated by microRNA-23a/27a/24-2 regulates M1 versus M2 macrophage polarization and thus regulates cancer progression. Oncotarget. 2016;7(12):13502–13519. doi: 10.18632/oncotarget.6284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Cortez M, Ivan C, Valdecanas D, Wang X, Peltier H, Ye Y, et al. PDL1 Regulation by p53 via miR-34. Journal of the National Cancer Institute. 2016;108(1). [DOI] [PMC free article] [PubMed]
- 77.Zonari E, Pucci F, Saini M, Mazzieri R, Politi L, Gentner B, et al. A role for miR-155 in enabling tumor-infiltrating innate immune cells to mount effective antitumor responses in mice. Blood. 2013;122(2):243–252. doi: 10.1182/blood-2012-08-449306. [DOI] [PubMed] [Google Scholar]
- 78.Komohara Y, Fujiwara Y, Ohnishi K, Takeya M. Tumor-associated macrophages: potential therapeutic targets for anti-cancer therapy. Adv Drug Deliv Rev. 2016;99:180–185. doi: 10.1016/j.addr.2015.11.009. [DOI] [PubMed] [Google Scholar]
- 79.Wang L, Hu Y, Zhao J, Huang F, Liang S, Dong L, et al. Targeted delivery of miR-99b reprograms tumor-associated macrophage phenotype leading to tumor regression. Journal for immunotherapy of cancer. 2020;8(2). [DOI] [PMC free article] [PubMed]
- 80.Li G, Liu D, Kimchi ET, Kaifi JT, Qi X, Manjunath Y, et al. Nanoliposome C6-Ceramide Increases the Anti-tumor Immune Response and Slows Growth of Liver Tumors in Mice. Gastroenterology. 2018;154(4):1024–1036.e9. [DOI] [PMC free article] [PubMed]
- 81.Xu G, Feng D, Yao Y, Li P, Sun H, Yang H, et al. Listeria-based hepatocellular carcinoma vaccine facilitates anti-PD-1 therapy by regulating macrophage polarization. Oncogene. 2020;39(7):1429–1444. doi: 10.1038/s41388-019-1072-3. [DOI] [PubMed] [Google Scholar]
- 82.Shigeta K, Datta M, Hato T, Kitahara S, Chen IX, Matsui A, et al. Dual programmed death Receptor-1 and vascular endothelial growth factor Receptor-2 blockade promotes vascular normalization and enhances antitumor immune responses in hepatocellular carcinoma. Hepatology. 2020;71(4):1247–1261. doi: 10.1002/hep.30889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Zhou B, Li C, Yang Y, Wang Z. RIG-I promotes cell death in hepatocellular carcinoma by inducing M1 polarization of Perineal macrophages through the RIG-I/MAVS/NF-κB pathway. OncoTargets Ther. 2020;13:8783–8794. doi: 10.2147/OTT.S258450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Zhou B, Yang Y, Li C. SIRT1 inhibits hepatocellular carcinoma metastasis by promoting M1 macrophage polarization via NF-κB pathway. OncoTargets Ther. 2019;12:2519–2529. doi: 10.2147/OTT.S195234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Zhang Z, Zhang J, He P, Han J, Sun C. Interleukin-37 suppresses hepatocellular carcinoma growth through inhibiting M2 polarization of tumor-associated macrophages. Mol Immunol. 2020;122:13–20. doi: 10.1016/j.molimm.2020.03.012. [DOI] [PubMed] [Google Scholar]
- 86.Li L, Sun P, Zhang C, Li Z, Cui K, Zhou W. MiR-98 modulates macrophage polarization and suppresses the effects of tumor-associated macrophages on promoting invasion and epithelial-mesenchymal transition of hepatocellular carcinoma. Cancer Cell Int. 2018;18:95. doi: 10.1186/s12935-018-0590-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Li L, Sun P, Zhang C, Li Z, Zhou W. MiR-98 suppresses the effects of tumor-associated macrophages on promoting migration and invasion of hepatocellular carcinoma cells by regulating IL-10. Biochimie. 2018;150:23–30. doi: 10.1016/j.biochi.2018.04.016. [DOI] [PubMed] [Google Scholar]
- 88.Das M, Zhu C, Kuchroo V. Tim-3 and its role in regulating anti-tumor immunity. Immunol Rev. 2017;276(1):97–111. doi: 10.1111/imr.12520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Mujib S, Jones R, Lo C, Aidarus N, Clayton K, Sakhdari A, et al. Antigen-independent induction of Tim-3 expression on human T cells by the common γ-chain cytokines IL-2, IL-7, IL-15, and IL-21 is associated with proliferation and is dependent on the phosphoinositide 3-kinase pathway. J Immunol (Baltimore, Md : 1950) 2012;188(8):3745–3756. doi: 10.4049/jimmunol.1102609. [DOI] [PubMed] [Google Scholar]
- 90.W Zhang, Y Liu, Z Yan, H Yang, W Sun, Y Yao, et al. IL-6 promotes PD-L1 expression in monocytes and macrophages by decreasing protein tyrosine phosphatase receptor type O expression in human hepatocellular carcinoma. Journal for immunotherapy of cancer. 2020;8(1). [DOI] [PMC free article] [PubMed]
- 91.Xie K, Xu L, Wu H, Liao H, Luo L, Liao M, et al. OX40 expression in hepatocellular carcinoma is associated with a distinct immune microenvironment, specific mutation signature, and poor prognosis. Oncoimmunology. 2018;7(4):e1404214. doi: 10.1080/2162402X.2017.1404214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Gefen T, Castro I, Muharemagic D, Puplampu-Dove Y, Patel S, Gilboa E. A TIM-3 oligonucleotide Aptamer enhances T cell functions and potentiates tumor immunity in mice. Mol Ther. 2017;25(10):2280–2288. doi: 10.1016/j.ymthe.2017.06.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Hervas-Stubbs S, Soldevilla M, Villanueva H, Mancheño U, Bendandi M, Pastor F. Identification of TIM3 2′-fluoro oligonucleotide aptamer by HT-SELEX for cancer immunotherapy. Oncotarget. 2016;7(4):4522–4530. doi: 10.18632/oncotarget.6608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Prodeus A, Abdul-Wahid A, Fischer N, Huang E, Cydzik M, Gariépy J. Targeting the PD-1/PD-L1 immune evasion Axis with DNA Aptamers as a novel therapeutic strategy for the treatment of disseminated cancers. Mol Ther Nucleic acids. 2015;4:e237. doi: 10.1038/mtna.2015.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Salmon H, Remark R, Gnjatic S, Merad M. Host tissue determinants of tumour immunity. Nat Rev Cancer. 2019;19(4):215–227. doi: 10.1038/s41568-019-0125-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Reina-Campos M, Moscat J, Diaz-Meco M. Metabolism shapes the tumor microenvironment. Curr Opin Cell Biol. 2017;48:47–53. doi: 10.1016/j.ceb.2017.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Riera-Domingo C, Audigé A, Granja S, Cheng W, Ho P, Baltazar F, et al. Immunity, hypoxia, and metabolism-the Ménage à Trois of Cancer: implications for immunotherapy. Physiol Rev. 2020;100(1):1–102. doi: 10.1152/physrev.00018.2019. [DOI] [PubMed] [Google Scholar]
- 98.MZ Norman, M Hasmim, A Lequeux, M Xiao, C Duhem, S Chouaib, et al. Improving Cancer Immunotherapy by Targeting the Hypoxic Tumor Microenvironment: New Opportunities and Challenges. Cells. 2019;8(9). [DOI] [PMC free article] [PubMed]
- 99.Zhang J, Zhang Q, Lou Y, Fu Q, Chen Q, Wei T, et al. Hypoxia-inducible factor-1α/interleukin-1β signaling enhances hepatoma epithelial-mesenchymal transition through macrophages in a hypoxic-inflammatory microenvironment. Hepatology (Baltimore, Md) 2018;67(5):1872–1889. doi: 10.1002/hep.29681. [DOI] [PubMed] [Google Scholar]
- 100.Noman M, Desantis G, Janji B, Hasmim M, Karray S, Dessen P, et al. PD-L1 is a novel direct target of HIF-1α, and its blockade under hypoxia enhanced MDSC-mediated T cell activation. J Exp Med. 2014;211(5):781–790. doi: 10.1084/jem.20131916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Barsoum I, Smallwood C, Siemens D, Graham C. A mechanism of hypoxia-mediated escape from adaptive immunity in cancer cells. Cancer Res. 2014;74(3):665–674. doi: 10.1158/0008-5472.CAN-13-0992. [DOI] [PubMed] [Google Scholar]
- 102.Chen J, Chen J, Huang J, Li Z, Gong Y, Zou B, et al. HIF-2α upregulation mediated by hypoxia promotes NAFLD-HCC progression by activating lipid synthesis via the PI3K-AKT-mTOR pathway. Aging. 2019;11(23):10839–10860. doi: 10.18632/aging.102488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Gropper Y, Feferman T, Shalit T, Salame T, Porat Z, Shakhar G. Culturing CTLs under hypoxic conditions enhances their cytolysis and improves their anti-tumor function. Cell Rep. 2017;20(11):2547–2555. doi: 10.1016/j.celrep.2017.08.071. [DOI] [PubMed] [Google Scholar]
- 104.Sormendi S, Wielockx B. Hypoxia Pathway Proteins As Central Mediators of Metabolism in the Tumor Cells and Their Microenvironment. Front Immunol. 2018;9:40. doi: 10.3389/fimmu.2018.00040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Chan MC, Holt-Martyn JP, Schofield CJ, Ratcliffe PJ. Pharmacological targeting of the HIF hydroxylases--a new field in medicine development. Mol Asp Med. 2016:54–75. [DOI] [PubMed]
- 106.Wu L, Zhang X, Zheng L, Zhao H, Yan G, Zhang Q, et al. RIPK3 orchestrates fatty acid metabolism in tumor-associated macrophages and Hepatocarcinogenesis. Cancer Immunol Res. 2020;8(5):710–721. doi: 10.1158/2326-6066.CIR-19-0261. [DOI] [PubMed] [Google Scholar]
- 107.Tian X, Wu Y, Yang Y, Wang J, Niu M, Gao S, et al. Long noncoding RNA LINC00662 promotes M2 macrophage polarization and hepatocellular carcinoma progression via activating Wnt/β-catenin signaling. Mol Oncol. 2020;14(2):462–483. doi: 10.1002/1878-0261.12606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Yang Y, Ye Y, Chen Y, Zhao J, Gao C, Han H, et al. Crosstalk between hepatic tumor cells and macrophages via Wnt/β-catenin signaling promotes M2-like macrophage polarization and reinforces tumor malignant behaviors. Cell Death Dis. 2018;9(8):793. doi: 10.1038/s41419-018-0818-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Li Q, Ma L, Shen S, Guo Y, Cao Q, Cai X, et al. Intestinal dysbacteriosis-induced IL-25 promotes development of HCC via alternative activation of macrophages in tumor microenvironment. J Exp Clin Cancer Res. 2019;38(1):303. doi: 10.1186/s13046-019-1271-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Yang F, Wei Y, Han D, Li Y, Shi S, Jiao D, et al. Interaction with CD68 and regulation of GAS6 expression by Endosialin in fibroblasts drives recruitment and polarization of macrophages in hepatocellular carcinoma. Cancer Res. 2020;80(18):3892–3905. doi: 10.1158/0008-5472.CAN-19-2691. [DOI] [PubMed] [Google Scholar]
- 111.Shiau DJ, Kuo WT, Davuluri GVN, Shieh CC, Tsai PJ, Chen CC, et al. Hepatocellular carcinoma-derived high mobility group box 1 triggers M2 macrophage polarization via a TLR2/NOX2/autophagy axis. Sci Rep. 2020;10(1):13582. doi: 10.1038/s41598-020-70137-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Zhao X, Wang X, You Y, Wen D, Feng Z, Zhou Y, et al. Nogo-B fosters HCC progression by enhancing yap/Taz-mediated tumor-associated macrophages M2 polarization. Exp Cell Res. 2020;391(1):111979. doi: 10.1016/j.yexcr.2020.111979. [DOI] [PubMed] [Google Scholar]
- 113.Cassetta L, Pollard J. Targeting macrophages: therapeutic approaches in cancer. Nat Rev Drug Discov. 2018;17(12):887–904. doi: 10.1038/nrd.2018.169. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.